Embed Size (px)
Citation preview

Communication Vol. 268. No. 10, Issue of April 5, p 6835-6838. 1993 THE JOURNAL OF BIOLOGICAL CHEMISTRY
0 1993 hy The American Society for Biochemistry and More& Biology, Inc. Printed in U. S. A.
Demonstration of Protein Tyrosine Phosphatase Activity in the Second of Two Homologous Domains of CD45*
(Received for publication, November 3, 1992, and in revised form, December 17,1992)
Xuehai Tan, David R. Stover, and Kenneth A. WalshS From the Department of Biochemistry, University of Washington, Seattle, Washington 98195
It has been reported that alteration or deletion of critical residues within one of the two homologous protein tyrosine phosphatase (PTPase)-like domains of CD45 completely abolishes all activity, suggesting that only the more N-terminal domain is catalytically ac- tive. However, we now demonstrate, by two independ- ent techniques, that the second (C-terminal) domain is also a viable phosphatase. Limited proteolysis by en- doproteinase Lys-C or trypsin increased the phospha- tase activity toward reduced, carboxymethylated, and maleylated lysozyme approximately 8-fold. A 50-kDa fragment, isolated by ion exchange chromatography, was found to be responsible for this activity. N-termi- nal sequencing revealed that this fragment includes less than half of the first phosphatase domain and most, if not all, of the second. In a second experiment, 109 residues, including the presumed catalytic region, were removed from domain I by site-directed mutagen- esis. Expression of this construct in a mammalian cell line resulted in increased PTPase activity over non- transfected control cells. Isolation of the recombinant CD45 by immunoprecipitation and immunoaffinity chromatography revealed that it had phosphatase ac- tivity. Both of these experimental approaches demon- strate that the second conserved PTPase domain of CD45 is a functioning PTPase, but that external reg- ulation may be required to express its activity in the context of the native molecule.
The leukocyte-common antigens (L-CA, CD45) are a family of protein tyrosine phosphatases expressed exclusively by cells of hematopoietic origin as receptor-like transmembrane pro- teins (Thomas, 1989; Trowbridge, 1991). They have attracted great attention because of their abundance, distinctive tissue distribution, novel structural features, and role in the response of lymphocytes to antigen. In addition, since the transfor- mation of normal cells to neoplastic cells is often accompanied by changes in the state of phosphorylation of tyrosine residues in proteins (Hunter and Cooper, 1986; White and Kahn, 1986), the catalytic function of CD45 and its homologs in the
* This work was funded by National Institutes of Health Grant GM15731 (to K. A. W.) and by National Research Service Award GM07270 (to D. R. S.). The costs of publication of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked “aduertisernent” in accordance with 18 U.S.C. Section 1734 solely to indicate this fact.
$ To whom correspondence should be addressed.
dephosphorylation of tyrosine residues may be a critical factor in regulating cell growth.
PTPases’ include a large family of receptor-like transmem- brane proteins that generally contain two cytoplasmic do- mains each of which is homologous to soluble forms of PTPases (Charbonneau et al., 1988). Activity has been dem- onstrated in the more N-terminal of these domains (domain I) in several members of this family (Johnson et al., 1992; Wang and Pallen, 1991; Streuli et al., 1990; Itoh et al., 19921, after expression of domain I alone or after mutagenesis of the cysteine in the second domain that is essential in homologous single domain PTPases. Similar experiments with the second domain of these transmembrane PTPases have generally yielded an inactive protein. The second domain of human protein tyrosine phosphatase N was shown to be active against the nonpeptide substrate para-nitrophenyl phosphate; how- ever, it had no activity against one protein substrate and extremely low activity against another (Wang and Pallen, 1991). The observation that several transmembrane PTPases lack either the catalytic cysteine within the second domain (Guan and Dixon, 1990; Krueger and Saito, 1992) or the entire second domain (Krueger et al., 1990) has contributed to the hypothesis that the second domain of the transmem- brane PTPases plays a regulatory rather than a catalytic role.
In the case of CD45, previous experiments have demon- strated that the external domain is not required for enzymatic activity, as the cytoplasmic segment expressed alone in bac- terial or baculovirus expression systems has intrinsic PTPase activity (Streuli et al., 1990; Ostergaard et al., 1989). The cytoplasmic segment features two tandem homologous do- mains (Fig. l ) , each of which resembles soluble forms of active PTPases (Charbonneau et al., 1988). Mutagenesis of murine CD45 has been used to dissect the critical regions required for activity (Johnson et al., 1992; Streuli et al., 1990). Their change of CYS’’~ (Cysa51 in human CD45) to Ser in domain I was reported to abolish all PTPase activity, indicating that domain I1 may be inactive in native CD45. However, domain 1 was not active when expressed independently, suggesting that domain I1 is required for the activity of domain I.
Although the second domain of CD45 is one of the least conserved PTPase domains, the degree of identity between it and other active domains (30-40%) remains quite high (Char- bonneau et d . , 1989). Its apparent lack of activity could have resulted from the use of inadequate expression systems, the lack of regulatory modifications, or the use of inappropriate substrates. In the present study, we tested whether the second domain is a viable phosphatase by applying limited proteolysis to generate a 50-kDa fragment, CD45(LysC50), that included domain I1 but little of domain I. This fragment was found to express activity similar to that of native CD45 toward 32P- labeled myelin basic protein (MBP), and much greater activity toward a second substrate, 32P-labeled reduced, carboxymeth- ylated and maleylated lysozyme (RCML), suggesting that a latent second domain was activated by proteolysis. To pre- clude the possibility that this activity was due to the residual
~~~~~
The abbreviations used are: PTPase, protein tyrosine phospha- tase; MBP, 32P-labeled myelin basic protein; RCML, 32P-labeled reduced, carboxymethylated, and maleylated lysozyme; MTX, meth- otrexate; PMSF, phenylmethanesulfony1 fluoride; 2-ME, 2-mercap- toethanol; BHK, baby hamster kidney; mAb, monoclonal antibody; MOPS, 4-morpholinepropanesulfonic acid.
6835

6836 Both PTPase-like Domains of CD45 Are Active CLl45(A825-933) CD45
GLYCOSYUTIONS DISULFIDE RICH
I I
FIG. 1. Schematic diagrams of the structure of CD46 and its truncated forms. The numbering system used is for unspliced human CD45 (Streuli et al., 1987). CD45(8825-933) was generated by excision of a BamHI fragment. A large C-terminal segment of domain 1 was excised, including a cysteine residue reported to be essential for domain I activity (Johnson et al., 1992; Streuli et al., 1990). The 50-kDa fragment CD45(LysC50) was generated by limited proteolysis with either trypsin or endoproteinase Lys-C. The peptide contains the C-terminal 106 amino acids of domain I, all of domain 11, and little if any of the C-terminal tail of CD45. The C terminus is estimated by N-terminal sequencing and SDS-gel mobility.
C-terminal segment of domain 1 in CD45(LysC50), we ex- amined a mutant form of CD45 lacking this segment, as expressed in a mammalian cell line, and found it to be an active PTPase.
EXPERIMENTAL PROCEDURES
Generation and Isolation of the Proteolytic Fragment-Approxi- mately 1.5 mg of CD45, isolated from Jurkat cells, was digested with 30 pg of endoproteinase Lys-C at 30 "C for 60 min in PTPase assay buffer (see below). The reaction was stopped by addition of 1 mM PMSF. Two ml of DE-52 Sepharose (Whatman) was then added and gently shaken for 10 min. The DE-52 slurry was poured into an Econo-Column (1 cm X 10 cm; Bio-Rad), and the resin was washed with 50 ml of 20 mM Tris, pH 7.2, containing 5 mM EDTA, and 0.1% 2-mercaptoethanol(2-ME). Proteolytic fragments were eluted with a gradient of NaCl to 400 mM in 50 ml of the same buffer. Two-ml fractions containing fragments of interest were identified by PTPase assays followed by SDS-polyacrylamide gel electrophoresis and West- ern analysis. These fractions were pooled, and the fragment was acetone-precipitated. An Applied Biosystems model 470A Sequencer was used to identify the N terminus of the major product obtained.
Plasmid Constructs and Mutagenesis-Human CD45 cDNA (a generous gift of Dr. Deborah Cool, University of Washington) was cut with restriction enzymes at appropriate sites and subcloned into expression vector Zem229 (Zymogenetics Inc.). This vector contains a cloning site controlled by the mouse metallothionein-1 promoter. It also contains a murine dihydrofolate reductase gene, which allowed selection of transfected cells with methotrexate (MTX) (Mulvihill et al., 1988). Due to the large size of CD45 cDNA (>4 kilobase pairs), only fragments to be mutated were subcloned into pBluescript KS
bases upstream of the ATG initiation codon were deleted by phagemid (Stratagene) for mutagenesis. To optimize gene expression, about 70
site-directed mutagenesis (Kunkel, 1985, 1988). In addition, a trun- cated form of CD45 (CD45(6825-933)) was generated by BamHI digestion to remove a segment, coding residues between amino acids DPHL and MKKR, that includes 109 amino acids of the C-terminal sequence of domain I. Mutated fragments were re-ligated into expres- sion vectors. The mutations were identified by restriction enzyme analysis and DNA sequencing.
Cell Culture and DNA Transfection-BHK 570 cells (ATCC CRL 1632) were maintained in Dulbecco's modified Eagle's medium (JRH), supplemented with 10% (v/v) newborn bovine serum (GIBCO), and 50 pg/ml penicillin, 50 pg/ml streptomycin, 100 pg/ml neomycin. DNA transfection was accomplished by a modified calcium phosphate precipitation procedure (Graham and van der Eb, 1973). Twenty-five pg of DNA was used for each transfection. One p~ MTX was added
to the culture media 24 h post-transfection. Resistant colonies (at least 10) were randomly picked about 10 days later and continued in this medium. The cell lines were subjected to concentrations of MTX increasing to 100 pM to amplify the transfected genes. The cell lines that expressed CD45 or its mutant form were identified by Western blot analysis and PTPase activity assay.
Western Blot, Immunoprecipitation, and Affinity Purification- Cells grown to confluence were scraped, harvested by centrifugation at 800 x g for 5 min, and washed three times with phosphate-buffered saline. The cell pellet was homogenized in a Teflon Dounce homoge- nizer in buffer A (10 mM Tris-HC1, pH 7.5, 5 mM MgC12, 5 mM PMSF, 5 pg/ml leupeptin, 1 mM EDTA). The cell debris was removed by centrifugation at 5000 X g for 5 min. The supernatant containing the cell membrane fraction was then spun at 100,000 X g for 1 h. The precipitate was extracted with buffer B (50 mM Tris-HC1, pH 7.5, 100 mM NaC1, 1% Nonidet P-40, 10 mM EDTA, 0.1% 2-ME, 1 mM PMSF, 5 pg/ml leupeptin), and undissolved debris was removed by a 5-min centrifugation step.
The supernatant was subjected to Western blot analysis with an anti-CD45 monoclonal antibody (mAb 9.4) (a gift of Dr. John Hansen, Fred Hutchinson Cancer Research Center). Western transfers were carried out as described (Towbin et al., 1979; Johnson et al., 1984) and developed with a goat anti-mouse antibody conjugated to horse- radish peroxidase. Bands were visualized with Western blot developer solution (Promega).
Immunoprecipitations were carried out by incubating cell extracts with mAb 9.4 for 4-12 h at 4 "C and then precipitating the complex with 5 mg of protein A-Sepharose. The precipitates were washed twice in buffer A and twice in buffer A containing 500 mM NaCl before assaying for enzymatic activity.
Affinity purification of CD45(A825-933) was performed as previ- ously described for CD45 (Stover et aL, 1991) by loading membrane extract supernatant on a mAb 9.4 affinity column, then washing with buffer containing 50 mM imidazole, pH 7.2, 400 mM NaC1, 5 mM EDTA, 0.1% Nonidet P-40, plus protease inhibitors (as in buffer A). The protein was eluted with 30 mM triethanolamine, pH 11, 0.1% Nonidet P-40, 0.1% 2-ME. Collected fractions were neutralized im- mediately with 1 M MOPS buffer, pH 6.0, and desalted on a Sephadex G-25 column into buffer containing 20 mM imidazole, pH 7.2, 2 mM EDTA, 0.1% 2-ME, 0.1% Nonidet P-40. After concentration by ultrafiltration using an XM-100 membrane (Amicon), 10% glycerol was added and the solution stored at -70 "C. All procedures were carried out at 4 "C.
Assay of PTPase and Protein-Activity assay of CD45 was per- formed as described by Tonks et al. (1988) using reduced, carboxy- methylated, and maleylated lysozyme or myelin basic protein as 32P- phosphorylated substrates. The protein concentrations in cell mem- brane extracts were determined with a Bio-Rad D/C protein assay kit.
RESULTS AND DISCUSSION
Nearly all of the reported transmembrane PTPases include two cytoplasmic domains that share a high degree of similarity to single-domain, cytoplasmic PTPases (Fischer et al., 1991). Therefore, the suggestion that only one of these homologous domains in CD45 is a viable phosphatase is provocative. The experiments leading to this suggestion did not rule out the possibility that activity in domain 11 was missing because of improper folding of recombinant forms of CD45 (in recombi- nant prokaryotic cells), lack of regulatory signals, or the use of an inappropriate substrate. We sought to reevaluate whether the second domain is capable of catalysis when the recombinant protein was generated in a mammalian cell and when possible regions of negative regulation have been re- moved.
To circumvent the problem of improper folding of geneti- cally altered CD45 constructs expressed in foreign host cells, we isolated native CD45 from a human T cell line (Jurkat) and attempted to generate a fragment with an active second domain by limited proteolysis. Tonks et al. (1990) found that limited treatment with trypsin activated CD45 toward RCML but not toward MBP. We found that endoproteinase Lys-C at a 1:50 ratio (w:w) for 60 min at 30 "C activated CD45
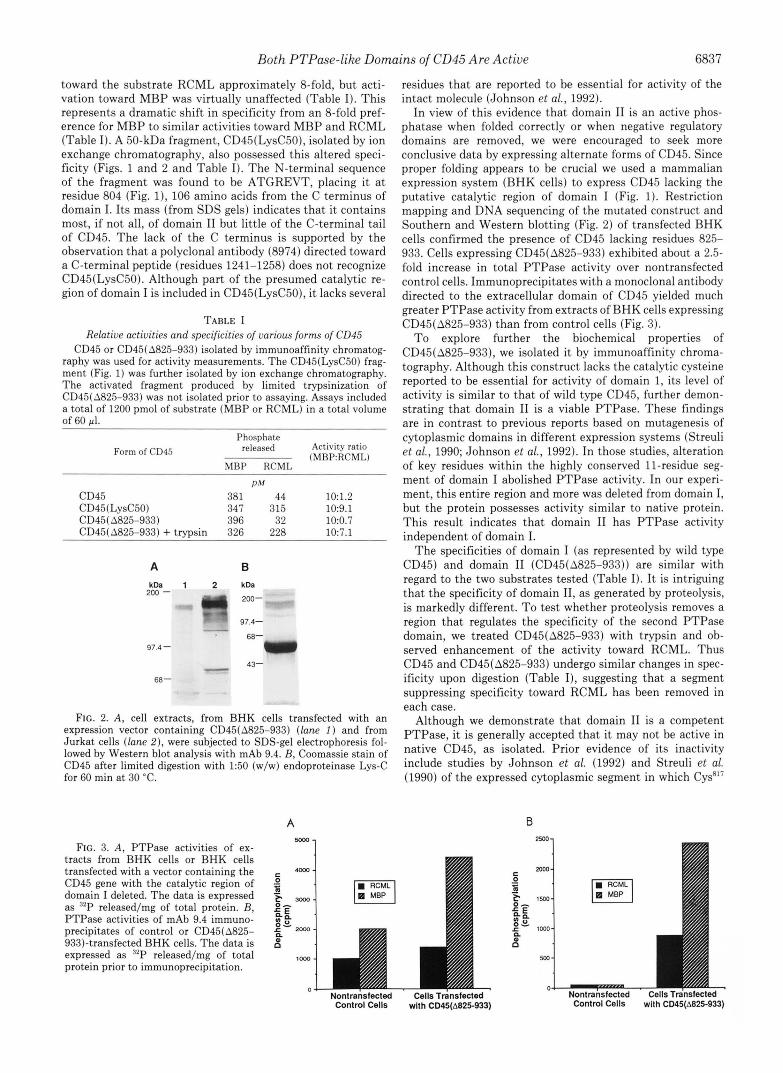
Both PTPase-like Domains of CD45 Are Active 6837
toward the substrate RCML approximately 8-fold, but acti- vation toward MBP was virtually unaffected (Table I). This represents a dramatic shift in specificity from an 8-fold pref- erence for MBP to similar activities toward MBP and RCML (Table I). A 50-kDa fragment, CD45(LysC50), isolated by ion exchange chromatography, also possessed this altered speci- ficity (Figs. 1 and 2 and Table I). The N-terminal sequence of the fragment was found to be ATGREVT, placing it at residue 804 (Fig. I), 106 amino acids from the C terminus of domain I. Its mass (from SDS gels) indicates that it contains most, if not all, of domain I1 but little of the C-terminal tail of CD45. The lack of the C terminus is supported by the observation that a polyclonal antibody (8974) directed toward a C-terminal peptide (residues 1241-1258) does not recognize CD45(LysC50). Although part of the presumed catalytic re- gion of domain I is included in CD45(LysC50), it lacks several
TABLE I Relative activities and specificities of various forms of CD45
CD45 or CD45(6825-933) isolated by immunoaffinity chromatog- raphy was used for activity measurements. The CD45(LysC50) frag- ment (Fig. 1) was further isolated by ion exchange chromatography. The activated fragment produced by limited trypsinization of CD45(A825-933) was not isolated prior to assaying. Assays included a total of 1200 pmol of substrate (MBP or RCML) in a total volume of 60 ~ l .
PhosDhate Form of CD4.5 released Activity ratio
(MBPRCML) MBP RCML
PM CD45 381 44 CD45(LysC50)
101.2 347 315 10:9.1
10:0.7 CD45(A825-933) + trypsin 326 228 10:7.1 CD45(A825-933) 396 32
A B
200 - kDa 1 2 koa
97.4
~ .* 97.4
68
FIG. 2. A, cell extracts, from BHK cells transfected with an expression vector containing CD45(A825-933) (lane I ) and from Jurkat cells (lane Z), were subjected to SDS-gel electrophoresis fol- lowed by Western blot analysis with mAb 9.4. B, Coomassie stain of CD45 after limited digestion with 1:50 (w/w) endoproteinase Lys-C for 60 min at 30 "C.
FIG. 3. A, PTPase activities of ex- tracts from BHK cells or BHK cells transfected with a vector containing the CD45 gene with the catalytic region of domain I deleted. The data is expressed as 32P released/mg of total protein. B, PTPase activities of mAb 9.4 immuno- precipitates of control or CD45(A825- 933btransfected BHK cells. The data is expressed as "P released/mg of total protein prior to immunoprecipitation.
residues that are reported to be essential for activity of the intact molecule (Johnson et al., 1992).
In view of this evidence that domain I1 is an active phos- phatase when folded correctly or when negative regulatory domains are removed, we were encouraged to seek more conclusive data by expressing alternate forms of CD45. Since proper folding appears to be crucial we used a mammalian expression system (BHK cells) to express CD45 lacking the putative catalytic region of domain I (Fig. 1). Restriction mapping and DNA sequencing of the mutated construct and Southern and Western blotting (Fig. 2) of transfected BHK cells confirmed the presence of CD45 lacking residues 825- 933. Cells expressing CD45(A825-933) exhibited about a 2.5- fold increase in total PTPase activity over nontransfected control cells. Immunoprecipitates with a monoclonal antibody directed to the extracellular domain of CD45 yielded much greater PTPase activity from extracts of BHK cells expressing CD45(A825-933) than from control cells (Fig. 3).
To explore further the biochemical properties of CD45(A825-933), we isolated i t by immunoaffinity chroma- tography. Although this construct lacks the catalytic cysteine reported to be essential for activity of domain 1, its level of activity is similar to that of wild type CD45, further demon- strating that domain I1 is a viable PTPase. These findings are in contrast to previous reports based on mutagenesis of cytoplasmic domains in different expression systems (Streuli et al., 1990; Johnson et al., 1992). In those studies, alteration of key residues within the highly conserved 11-residue seg- ment of domain I abolished PTPase activity. In our experi- ment, this entire region and more was deleted from domain I, but the protein possesses activity similar to native protein. This result indicates that domain I1 has PTPase activity independent of domain I.
The specificities of domain I (as represented by wild type CD45) and domain I1 (CD45(A825-933)) are similar with regard to the two substrates tested (Table I). It is intriguing that the specificity of domain 11, as generated by proteolysis, is markedly different. To test whether proteolysis removes a region that regulates the specificity of the second PTPase domain, we treated CD45(A825-933) with trypsin and ob- served enhancement of the activity toward RCML. Thus CD45 and CD45(A825-933) undergo similar changes in spec- ificity upon digestion (Table I), suggesting that a segment suppressing specificity toward RCML has been removed in each case.
Although we demonstrate that domain I1 is a competent PTPase, it is generally accepted that it may not be active in native CD45, as isolated. Prior evidence of its inactivity include studies by Johnson et al. (1992) and Streuli et al. (1990) of the expressed cytoplasmic segment in which CysR"
A B
-1
" . Nontransfected . Cells Transfected
Control Cells wlth CD45(A825-933)
2500-
m-
1500-
10W-
500-
0-
RCML
Control Cells wlth CD45(A825-933)

6838 Both PTPase-like Domains of CD45 Are Active
in domain I was changed to Ser; no activity was observed even though domain I1 was intact. However, analogous mu- tation of domain I1 inhibits approximately 50% of activity. When domain 11 was completely removed, leaving an intact domain I, no activity was observed, suggesting that domain I activity may require the presence of an intact domain 11. We have now demonstrated that removal of segments of domain I reveals an active domain 11, suggesting that domain I may suppress the activity of domain 11. These observations are consistent with the hypothesis that unidentified regulatory events (e.g. ligand binding or phosphorylation) switch on domain 11. Interdomain regulation is indicated by the depend- ence of domain I activity on the presence of an intact domain 11, the apparent activation of domain I1 upon disruption of domain I, and the observation that relatively harsh treatment with proteases yields a fragment that spans domain 11 and part of domain I rather than cleaving between the two do- mains as might have been expected.
In summary, we have demonstrated that the C-terminal domain of a transmembrane PTPase, CD45, bears substantial levels of activity toward a peptide substrate. When CD45 lacking the catalytic region of the first domain was expressed in mammalian cells, it had a specificity and level of activity similar to wild type CD45. Limited proteolysis of either the wild type or the mutant form of CD45 altered its substrate specificity, suggesting that a region that influences the spec- ificity of the second PTPase domain had been removed. Because domain I is reported to be inactive in the absence of domain I1 and, conversely, domain I1 is reported to be inactive in the presence of domain I, we propose a model involving interdomain regulation where an unidentified regulatory switch turns on one domain while turning the other off.
Acknowkdgrnents-We are grateful to Santosh Kumar for protein sequence analysis, F. Lee for synthesis of oligonucleotides, Kurt Diltz for preparation of PTPase substrates, and Shaoming Huang for assistance in vector construction, cell culture, and transfection.
REFERENCES Walsh, K. A., and Fisher, E. H. (1988) Proc.
- """ meau, H., and Tonks, M. K. (1991) Science 2 5 3 , 401-
Gr'al;am F. L. and van der Eb A. J. (1973) Virology 52,456-466 Guan, K., and'Dixon, J. E. (1960) Science 249,553-556 Hunter, T., and Cooper, J. A. (1986) in The Enzymes (Bo er P D , and Krebs,
E. G., eds) Vol. 17, pp. 191-246, Academlc Press, New 4o;k ' '
Itoh, M., Streuli, M., Krueger, N. X., and Saito, H. (1992) J. Biol. Chem. 2 6 7 ,
Johnson, D. A., Gautsch, J. W., Sportsman, J. R., and Elder, J. H. (1984) Gene
Johnson, P., Ostergaard, H. L., Wasden, C. , and Trowbridge, I. S. (1992) J.
Krueger, N. X., and Saito, H. (1992) Proc. Natl. Acad. Sci. U. 5'. A. 8 9 , 7417-
Krueger, N. X Streuli, M., and Saito, H. (1990) EMBO J . 9 , 3241-3252 Kunkel, T. A. 11985) Proc. Natl. Acad. Sci. U. S. A. 82,488-492 Kunkel, T. A. (1988) in Nucleic Acids and Molecular Biology (Eckstein, F., and
Mulvihill, E., Berkner, K., Foster, D., Kumar, A., McKay, V., and Parker, G. Lilly, D. M. J., eds) Vol. 2, pp. 124-135, Springer-Verlag. Berlin
Ostergaard, H. L., Shackelford, D. A,, Hurley, T. R. Johnson, P., Hyman, R., (1988) European Patent Application 039944
Sefton, B. M., and Trowbridge, I. S. (1989) Procl Natl. Acad. SCL. U. S. A. 86,8959-8963
Stover, D. R., Charbonneau, H., Tonks, N. K., and Walsb, K. A. (1991) Proc. Natl. Acad. Sci. U. S. A. 8 8 , 7704-7707
Streuli, M., Hall, L. R., Saga, V., Schlossman, S. F., and Saito, H. (1987) J. Exp. Med. 166,1548-1566
Streuli, M., Krueger, N. X., Tsai, T., Tang, M., and Saito, H. (1990) EMBO J. 9,2399-2407
Thomas M. L. (1989) Annu. Rev. Immunol. 7,339-369 Tonks, N. K., Diltz, C. D., and Fisher, E. H. (1990) J . Biol. Chem. 266 , 10674-
12356-12363
Anal. Tech. 1, 3
Biol. Chem. 267,8035-8041
7421
1 nmn To%<;;, H. H., Staehelin, T., and Gordon, J. (1979) Proc. Natl. Acad. Sci.
Trowbridge, I. S. (1991) J. Biol Chem. 266,23517-23520 Wang Y., and Pallen, C. J. (1991) EMBO J. 10,3231-3237 White', M. F., and Kahn, C. R. (1986) in The Enzymes (Bo er, P. D., and Krebs,
U. S. A. 76,4350-4354
E. G., eds) Vol. 17, pp. 247-310, Academlc Press, New Bork





























