Embed Size (px)
Citation preview

Page 1/30
Effect of thiram on rat kidney: inhibition of brushborder membrane and antioxidant enzymes,diminution of antioxidant capacity, enhanced DNAdamage and DNA-protein cross-linkingSamreen Salam
Aligarh Muslim UniversityAmin Arif
Aligarh Muslim University Faculty of Life SciencesAijaz Ahmed Khan
Aligarh Muslim University Faculty of MedicineRiaz Mahmood ( [email protected] )
Aligarh Muslim University, Aligarh, India https://orcid.org/0000-0001-6405-0013
Research Article
Keywords: thiram, kidney, Antioxidant enzymes, oxidative stress, DNA damage, metabolic enzymes
Posted Date: March 1st, 2021
DOI: https://doi.org/10.21203/rs.3.rs-205480/v1
License: This work is licensed under a Creative Commons Attribution 4.0 International License. Read Full License

Page 2/30
AbstractThiram is a dithiocarbamate pesticide that is widely used as a fungicide to protect crops and seeds,especially in China and India. Although thiram is considered relatively safe for humans but due to itspersistent nature it may become a health hazard for human beings and animal if long term exposuretakes place. The aim of the present work was to study the effects of oral administration of thiram onkidney of male rats given different doses of thiram (100, 250, 500, 750 mg/kg body weight) for 4consecutive days. This treatment signi�cantly reduced cellular glutathione and total sulfhydryl contentbut enhanced protein carbonyl and hydrogen peroxide levels. The plasma creatinine and BUN levels werealso elevated indicating nephrotoxicity. The activities of antioxidant enzymes catalase, superoxidedismutase, glutathione peroxidase, thioredoxin reductase and glutathione-S-transferase were signi�cantlydecreased. The antioxidant capacity was diminished resulting in less free radical quenching and metalreducing ability of kidney. Administration of thiram also led to inhibition of intestinal brush bordermembrane enzymes: alkaline phosphatase, leucine aminopeptidase, γ-glutamyl transferase and maltase.Activities of enzymes of glucose metabolism viz. glycolysis, citric acid cycle, pentose phosphate pathwayand gluconeogenesis were also inhibited. Histopathology of kidney tissue revealed tubular dilation,tubular cast, breaking of apical cytoplasm and intestinal hemorrhage. A signi�cant increase in DNAfragmentation, DNA strand breaks and DNA-protein cross-linking was also observed in thiram treated ratscompare to control group. These changes in kidney could be due to marked perturbation in antioxidantdefense system induced by free radicals generated upon exposure to thiram.
IntroductionDithiocarbamate fungicides form a large group of sulfur containing chemicals that have numerous usesin agriculture and medicine (Buac et al., 2012; Thind and Hollomon, 2017). They can be applied to thefoliage of plants for seed and fruits treatment and also used as bird and rodent repellents.Tetramethylthiuram disul�de, commonly known as thiram, is a dithiocarbamate fungicide that exhibits abroad spectrum of antifungal activity. It is widely used in agriculture due to its low cost, good e�cacy andrelatively low toxicity. It is also used as slimicide in water cooling systems, in sugar, pulp, and papermanufacturing and as vulcanization accelerator in the rubber industry (Alam et al., 2017; Cereser et al.,2001a). Thiram is also employed in the treatment of human scabies, as a sun screen and as a bactericideapplied directly to the skin or incorporated into soap. Thiram has metal chelating properties because ofwhich it is used as a scavenger in waste water treatment (Kanecoa et al., 2009). In agricultural �eld it isused to control colletotrichum lint on �ax, fungal diseases on sa�ower and cotton seed, as a repellent forrabbit, deer and black birds. Thiram is applied on seeds to control black root of sugar beet, apple scab,grey mould of strawberries, brown rot of stone fruit, against damping off of nursery seedlings and turfcrops (Sharma et al., 2003).
Despite several useful applications, the excessive use of thiram is harmful as it can enter into theenvironment by surface run-off, spraying on crops or in gardens, also via e�uents of waste watertreatment plants and can get incorporated into the food chain. People can be exposed to thiram by

Page 3/30
consuming food and liquid containing residues, by inhaling thiram contaminated air or by occupationalexposure. Thiram exposure is associated with several toxicological effects including endocrine disruption(Chen et al., 2018), hepatotoxicity (Saqib et al., 2005) neurotoxicity (Agrawal et al., 1997) and genotoxicity(Perocco et al., 1989). Thiram also interferes with the metabolism of xenobiotic compounds by inhibitingthe enzyme arylamine N-acetyltransferase-1 (Xu et al., 2017). Thiram and ziram induce neurotoxicity byincreasing the intracellular level of Ca2+ through non-selective cation channels (Han et al., 2003).Ingestion of seeds coated with thiram and imidacloprid decreases cellular immune response in red-lagged partridges (Alectoris rufa) (Lopez-Antia et al., 2015). Stoker et al. (1993) showed that thiramaffects oocyte fertilization by delaying ovulation. About one million birds suffer from depressed growth,soft egg shell and leg abnormalities from consuming thiram contaminated feed (Rath et al., 2005;Nageswara et al., 2017). At 100–500 ppm, thiram also inhibits the laying of eggs in hens, partridges andquails (Lorgue et al., 1996). Thiram is also a well know inducer of tibial dyschondroplasia, a commonmetabolic cartilage disease found in quickly growing poultry birds (Waqas et al., 2020). Since thiram canenter surface water by runoff and erosion its effects on the aquatic ecosystem have also been studied.Exposure to thiram at the early life stage affects the development of zebra �sh (Danio rerio) (Chen et al.,2018). It decreases growth rate and induces neurotoxicity in Daphnia magna, a small planktoniccrustacean (Belaid et al., 2019).
A strong correlation between pesticide toxicity and enhanced production of reactive oxygen species(ROS) and free radicals has been observed (Khan et al., 2005; Mansour and Mossa, 2010). Oxidativestress is characterized by an increase in the levels of ROS and free radicals that exceed the capacity ofcells to quench them. Thiram also induces oxidative stress by generating ROS and free radicals(Grosicka-Maciag et al., 2008; Cereser et al., 2001b; Kurpios-Piec et al., 2015). The formation of thesereactive species is the main cause for the pathogenesis of many diseases such as cancer,neurodegeneration, cardiovascular diseases and rheumatoid arthritis (Valko et al., 2007).
Due to the functional, biochemical and morphological heterogeneity of the kidney, several drugs andxenobiotics show site speci�c toxicity to this organ (Werner et al., 1995). Indeed, pesticides can causesevere renal damage including in�ammation, tubular cell toxicity, crystal nephropathy (Zager, 1997;Schetz et al., 2005) and nephrotoxicity in rat model (Behling et al., 2006). Blood chemistry showed thatthiram causes kidney damage in rats and beagle dogs (Maita et al., 1991; Lee et al., 1978). However,except for these two studies there are no reports investigating the nephrotoxic effect of thiram. In view ofthe paucity of reports on toxicity of thiram on kidney, we have examined the biochemical, histological andgenotoxic potential of this fungicide on rat kidney.
Material And MethodsChemicals
Thiram (purity, 97%) was purchased from Sigma Aldrich, USA. All other chemicals used were of analyticalgrade and obtained either from Sisco Research Laboratories (Mumbai, India), Himedia Laboratories

Page 4/30
(Mumbai, India) or Sigma-Aldrich (USA).
Experimental protocol
The animal experiments were done as per the guidelines provided by the Institutional Ethics Committee(IEC) of Aligarh Muslim University that monitors research involving animals (R. No. :714/GO/Re/S/02/CPCSEA). Adult male Sprague Dawley rats weighing 150-200 g were obtained fromNational Institute of Biologicals, Noida, India. The animals had unlimited access to standard pellet ratdiet and clean drinking water. After one week of acclimatization, 30 male rats were randomly divided into�ve groups of six animals each. Thiram was dissolved in corn oil and orally administered (by gavage) for4 days at doses of 100, 250, 500, 750 mg/kg body weight/day at an interval of 24 h. Untreated animals inthe control group received an equivalent volume of corn oil by gavage. There was no animal mortalitybecause these doses were below the LD50 value of thiram for rats (620 to over 1900 mg/kg body weight).All animals were sacri�ced 24 h after the administration of last dose of thiram, under anesthesia. Bothkidneys were carefully removed from each animal, decapsulated and used for the preparation ofhomogenates. Blood was removed from the heart using a syringe and transferred to heparinized tubes.
Renal function test
Plasma concentrations of creatinine and blood urea nitrogen (BUN) were used as markers of renalfunction. Blood was centrifuged at 1000 rpm for 10 min at 4ºC and supernatant (plasma) was saved. Theplasma was �rst deproteinized by adding 3% trichloroacetic acid in 1:3 ratio and centrifuged at 3000 rpmfor 5 min to pellet the precipitated proteins. The supernatant was used for BUN and creatinine estimationusing kits purchased from Arkray Healthcare, Mumbai, India. Creatinine was determined by modi�edJaffe’s method (Haugen, 1953) and BUN by diacetyl monoxime reagent (Rosenthal, 1955).
Preparation of kidney homogenates
Each kidney was divided into cortex and medulla using a sharp scalpel. The cortex and medullaryportions were homogenized in 2 mM Tris, 50 mM mannitol buffer, pH 7.5, to prepare a 10% (w/v)homogenate. Homogenates were further homogenized using an Ultra Turrex, Kunkel homogenizer bypassing �ve pulses of 30 sec each. Each homogenate was divided into aliquots and used immediately orquickly frozen at -80°C for further analyses.
Preparation of brush border membrane and assay of bound enzymes
The cortical homogenates prepared above were further processed to isolate brush border membrane(BBM) vesicles. To each homogenate 10 mM MgCl2 (�nal concentration) was added, samples left for 20min on ice with continuous stirring and then centrifuged at 2000 rpm for 15 min at 4ºC. The supernatantso obtained was centrifuged at 17,000 rpm (35000 x g) at 4°C for 30 min. The supernatant was discardedand pellet was suspended in 300 mM mannitol, 5 mM Tris-HCl buffer, pH 7.5, homogenized in a handheld homogenizer and again centrifuged at 17000 rpm for 30 min at 4°C. The supernatant was discarded

Page 5/30
and the white �uffy layer present in pellet, containing BBM, was suspended in 300 mM mannitol, 5 mMTris-HCl, pH 7.5 buffer (Khundmiri et al., 1997). The BBM preparations were used immediately or stored at-80ºC and used within 3-4 days. Protein concentration in homogenates and isolated BBM vesicles wasdetermined using Folin’s phenol reagent (Lowry et al., 1951).
Alkaline phosphatase (ALP), γ-glutamyl transferase (GGT) and leucine aminopeptidase (LAP) wereassayed by using p-nitrophenyl phosphate, γ-glutamyl p-nitroanilide and L-leucine p-nitroanilide assubstrates, respectively (Kempson et al., 1979; Kempson et al., 1985; Goldmann et al., 1976). Glucose-oxidase peroxidase method was employed to determine the enzymatic activity of maltase (Bergmeyer,1974).
Non-enzymatic oxidative stress markers
The concentration of reduced glutathione (GSH) was determined in protein-free homogenates using 5,5’-dithiobis-2-nitrobenzoic acid (DTNB), as described by Beutler et al. (1963). The total sulfhydryl groupcontent was quanti�ed from the concentration of the yellow thionitrobenzoate anion, which forms uponreaction of sulfhydryl/thiol groups with DTNB and absorbs maximally at 412 nm (Sedlak and Lindsay,1968). Protein oxidation was determined from carbonyl content as described by Levine et al. (1990). Inthis method, protein carbonyl groups react with 2,4 dinitrophenylhydrazine to form hydrazone adductwhich was quanti�ed from absorbance of solutions at 360 nm. Hydrogen peroxide (H2O2) concentration
was determined by the method of Gay and Gebicki (2000). H2O2 oxidizes Fe2+ to Fe3+ under acidic
conditions; Fe3+ reacts with xylenol orange to form a purple complex that absorbs at 560 nm. Allparameters were determined in kidney homogenates.
Antioxidant capacity
The antioxidant power (AO)/capacity of cortex and medullary homogenates was determined by usingFRAP (ferric reducing antioxidant power) and DPPH (2,2-diphenyl-1picrylhydrazyl) assays. In FRAP assay,0.1 ml homogenate was added to 1.5 ml FRAP reagent (300 mM sodium acetate, 10 mM 2,4,6-Tris(2-pyridyl)-s-triazine, 20 mM FeCl3, pH 3.6). The absorbance of samples was recorded after 5 min at 593 nm(Benzie and Strain, 1996). In DPPH assay, the AO power of homogenates was determined in terms ofpercent quenching of DPP• radical. Lower the quenching of DPP radical, lesser is the AO power. To 0.1 mlhomogenate were added 0.4 ml of 10 mM sodium phosphate buffer, pH 7.4, and 0.5 ml of 0.1 mM DPPHreagent. The samples were kept for half an hour in the dark, centrifuged at 12,000 x g and absorbance ofsupernatants was read at 517 nm (Mishra et al., 2012).
Antioxidant enzymes
The activity of catalase (CAT) was determined from its ability to convert H2O2 into H2O (Aebi, 1984) andCu, Zn-superoxide dismutase (SOD) from the inhibition of autoxidation of pyrogallol (Marklund andMarklund, 1974). Glutathione reductase (GR) was assayed from the reduction of oxidized glutathione(GSSG) to GSH with concomitant conversion of reduced nicotinamide adenine dinucleotide phosphate

Page 6/30
(NADPH) to its oxidized form (NADP+) as described by Carlberg and Mannervik (1985). Thioredoxinreductase (TR) was assayed from the reduction of DTNB to yellow thionitrobenzoate anion in presence ofNADPH; the absorbance of yellow color produced was read at 410 nm (Tamura and Stadtman, 1996).Glutathione peroxidase (GPx) was assayed by the method of Flohe and Gunzler (1984). In this methodNADPH is oxidized to NADP+, in presence of GSSG and GR, resulting in decrease in absorbance at 340nm. Glutathione-S-transferase (GST) is a phase II detoxifying enzyme which acts by conjugating harmfulxenobiotics to GSH. It was assayed using 1-chloro-2,4-dinitrobenzene and GSH as substrates. Theabsorbance of the conjugate formed was read at 340 nm. An extinction coe�cient of 9,600 M-1cm-1 wasused to calculate enzyme activity (Habig et al., 1974).
Metabolic enzymes
Hexokinase activity was determined from the decrease in concentration of free glucose as described byCrane and Sols (1953). Lactate dehydrogenase (LDH) was assayed from the oxidation of reducednicotinamide dinucleotide (NADH) to its oxidized form (NAD+) in the presence of sodium pyruvate; thedecrease in absorbance of solution was read at 340 nm (Khundmiri et al., 2004). Glucose 6-phosphatedehydrogenase (G6PDH) was assayed from the conversion of glucose 6-phosphate to 6-phosphogluconolactone, with concomitant reduction of NADP+ to NADPH (Shonk and Boxer, 1964). Theactivity of gluconeogenic enzymes, glucose 6 phosphatase (G6Pase) and fructose 1,6-bisphosphatase(FBPase), were determined from the release of inorganic phosphate from their substrates, glucose 6-phosphate and fructose 1,6-bisphosphate, respectively (Shull et al., 1956; Freedland and Harper, 1959).The amount of inorganic phosphate released was estimated by using Taussky and Shorr reagent(Taussky and Shorr, 1953). Malic enzyme was assayed from the oxidative decarboxylation of L-malate topyruvate. NADP+ serves as electron acceptor and is converted to NADPH resulting in increase inabsorbance of solution at 340 nm (Ochoa et al., 1948). Malate dehydrogenase was assayed by followingthe oxidation of NADH to NAD+ using oxaloacetate as substrate. The decrease in absorbance of solutionwas recorded at 340 nm (Mehler and Korenberg, 1948).
Acid phosphatase and total ATPase
Total ATPase activity was determined from the concentration of inorganic phosphate released uponhydrolysis of ATP (Bonting et al., 1961). Acid phosphatase acts on p-nitrophenyl phosphate to fromyellow p-nitrophenol that absorbs at 415 nm (Mohrenweiser and Novotny, 1982).
DNA fragmentation (DF) and DNA-protein crosslinks (DPCs)
DNA damage by thiram was assayed by diphenylamine and DNA-protein cross-linking (DPC) assays. Theconcentration of fragmented DNA was estimated using diphenylamine as described by Burton (1956).The homogenates were centrifuged and 0.5 M perchloric acid was added to both supernatant and pelletto precipitate the proteins. The samples were centrifuged and to 0.5 ml supernatant, 1 ml diphenylamine

Page 7/30
reagent (1.5 gm diphenylamine, 100 ml acetic acid, 1.5 ml H2SO4 and 1.6 µg/ml acetaldehyde) wasadded. After 16-20 h at room temperature in the dark, absorbance of solutions was read at 600 nm.
DPCs are formed when cells are exposed to DNA damaging agents. The levels of DPCs were determinedusing K+/SDS assay, exactly as described by Zhitkovich and Costa (1992).
Comet assay
The genotoxic effect of thiram was analyzed by single cell gel electrophoresis using the method of Singhet al. (1988) but with slight modi�cations. Single cell suspension was prepared by mincing the tissue inRosewell Park Memorial Institute 1640 medium. A fraction of suspension was mixed with 1% low meltingpoint agarose (LMPA), poured over glass slides coated with 1% normal agarose and allowed to solidify.Another layer of LMPA (0.5%) was then spread over slides which were then dipped in chilled lysis buffer(100 mM EDTA, 2.5 m NaCl, 10 mM Tris-HCl, 1% Triton-X-100, pH 10.0) for 3 h in dark at 4ºC. The slideswere then transferred to electrophoresis buffer (1 mM EDTA, 300 mM NaOH, pH 13) for 30 min to allowDNA unwinding. Electrophoresis was carried out for 30 min at 0.7 V/cm and 300 mA at 4ºC. The slideswere then neutralized with 0.4 M Tris-HCl buffer, pH 7.4. The DNA was stained by adding a drop of 20µg/ml ethidium bromide on slides which were then washed with distilled water. The DNA was visualizedunder �uorescence microscope at 100 x and analyzed with Comet assay software from CaspLab.
Histopathology
Longitudinal sections of the kidney of 10 µm thickness were cut. The tissue was processed to prepareslides as described by Culling (2013) and stained with either eosin or hematoxylin. The slides were thenvisualized under microscope at 400 x magni�cation.
Statistical analysis
All data was expressed as mean ± standard error mean for six rats in each group and statisticalsigni�cance was reported using one way ANOVA. Student t-test was used for comparison between treatedand control groups. Differences were considered signi�cant at p < 0.05.
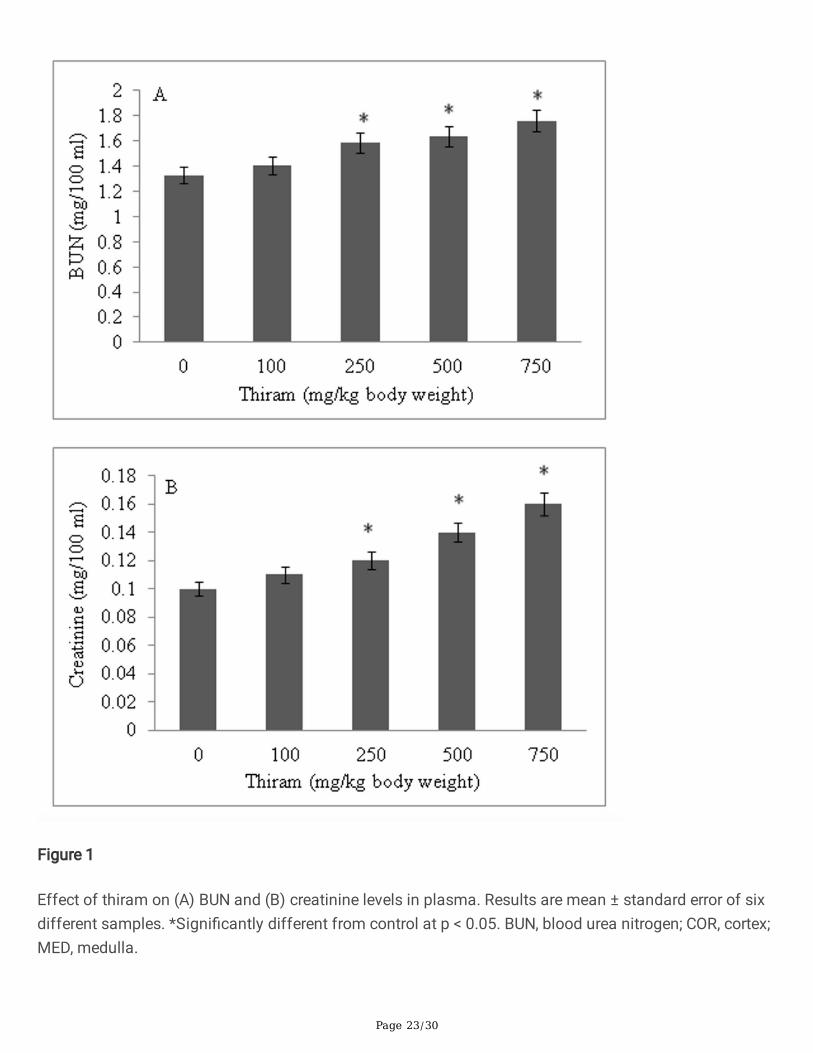
ResultsRenal function test
Both BUN and creatinine levels showed a thiram dose-dependent increase. The concentration of BUN inplasma increased to 1.6 fold in treated group (Fig. 1A) while creatinine increased by 1.32 fold, comparedto untreated rats (Fig. 1B).
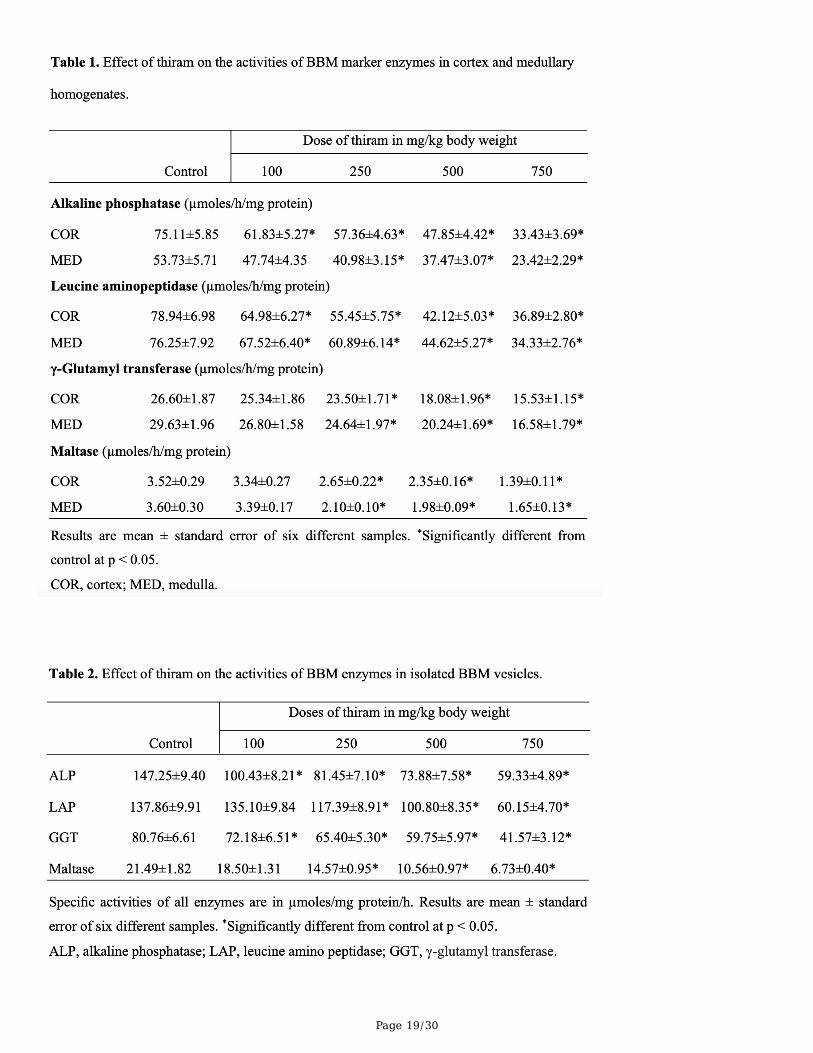
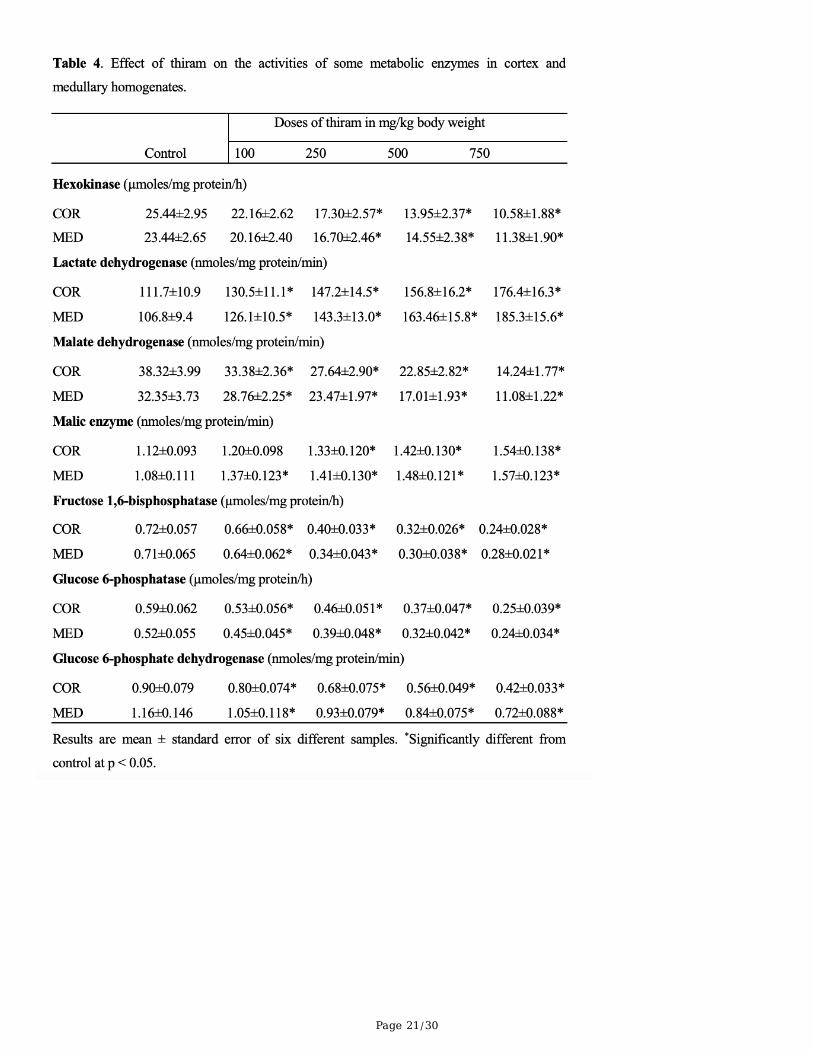
BBM enzymes

Page 8/30
The effect of thiram on the speci�c activities of BBM marker enzymes in isolated BBMV and cortical andmedullary homogenates is shown in Tables 1 and 2. Thiram exposure caused a signi�cant reduction inthe speci�c activities of all BBM enzymes in cortical and medullary homogenates. At the highest dose ofthiram (750 mg/kg body weight) the enzyme activities, relative to control, were: ALP, 44.50 and 43.59%;LAP, 46.73 and 45.02%; GGT, 58.38 and 55.15%; maltase, 39.48 and 45.83%. A similar pattern was alsoevident in the activities of these four BBM enzymes in isolated vesicles: ALP, 40.29%, LAP, 43.63%, GGT,51.47% and maltase, 31.31%.
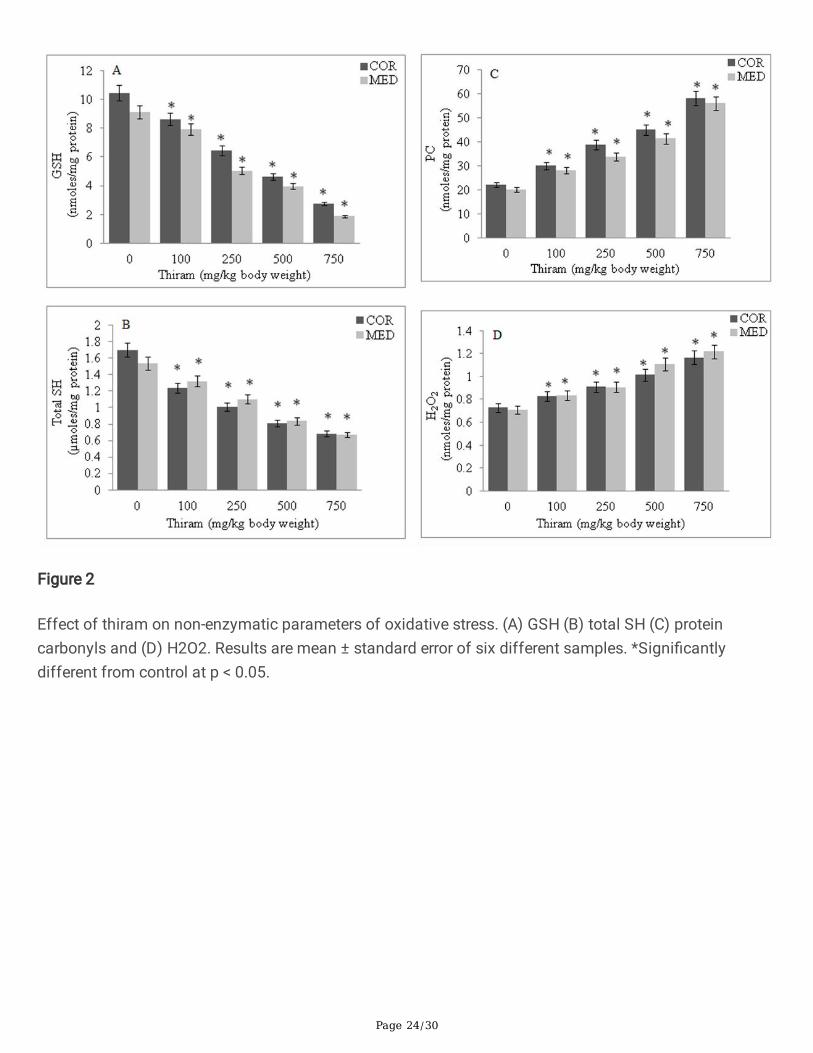
Non-enzymatic oxidative stress markers
The levels of both GSH (Fig. 2A) and total SH (Fig. 2B) showed a thiram dose-dependent decrease. Whilea signi�cant increase in carbonyl content (Fig. 2C) was seen along with an increase in H2O2 level (Fig.2D) in rats of thiram treated groups, compared to control group. The percent levels, relative to control, inthese parameters at the highest dose of thiram (750 mg/kg body weight), in cortical and medullaryhomogenates, were: GSH, 26.2 and 20.5; total SH, 50.0 and 55.6; protein carbonyls, 261.4 and 277.3;H2O2, 160.8 and 170.4.
Antioxidant capacity
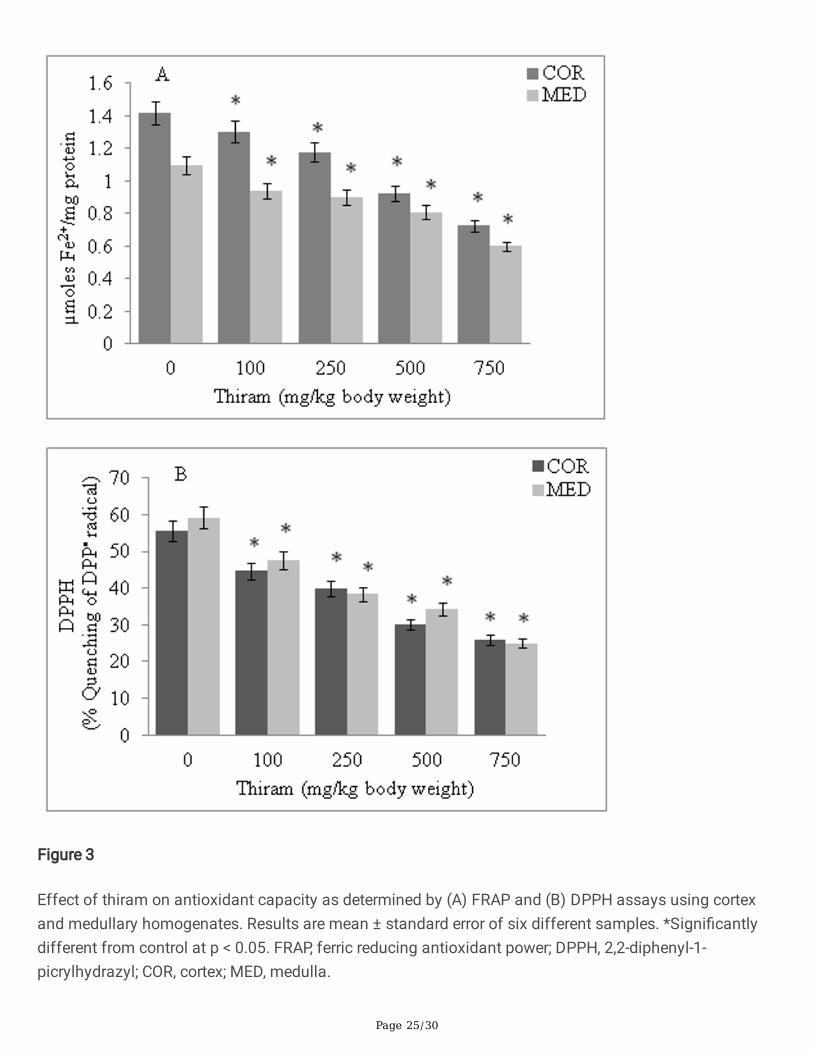
AO capacity of cortex and medulla decreased to 50.70% and 54.12%, respectively, as determined by theFRAP assay (Fig. 3A). In DPPH method, a reduction in DPPH quenching ability of cortical (46.25%) andmedullary (42.05%) homogenates was observed with increase concentration of thiram (Fig. 3B).
Antioxidant enzymes
SOD catalyzes the dismutation of superoxide anion to H2O2, which is then converted to oxygen and waterby CAT and GP. The speci�c activities of all three enzymes show a dose-dependent decline in cortical andmedullary homogenates of thiram treated rats. The SOD, CAT and GP activities, as percent control, were:34.78 and 23.13, 48.11 and 46.71, 56.89 and 58.97, respectively. The speci�c activities of secondary AOenzymes (GR, TR and GST) also decreased in a similar thiram dose-dependent manner. The percentactivities of these enzymes, relative to control, were: GR, 48.20 and 32.79; TR, 36.24 and 43.67 and GST,42.24 and 40.41 (Table 3).
Metabolic enzymes
The speci�c activities of hexokinase and G6PDH showed a thiram dose-dependent decrease in bothcortex and medullary homogenates: Hexokinase (41.58% in cortex and 48.54% in medulla) and G6PDH(46.66% in cortex and 62.06% in medulla). The activity of LDH increased and was 1.58 fold in corticaland 1.73 fold in medullary homogenates, relative to control. Similarly, the activities of gluconeogenicenzymes FBPase and G6Pase showed a thiram dose-dependent decrease. The activity of FBPasedecreased to 33.33% in cortex and 39.43% in medulla while G6Pase decreased to 42.37% in cortex and46.15% in medullary homogenates. The activity of malate dehydrogenase, also showed a gradual

Page 9/30
decrease. The percent decrease in its activity was 37.12 in cortex and 34.28 in medullary homogenate.Like LDH, the activity of malic enzyme also increased with increase concentration of thiram. A 1.37 foldincrease in the activity of malic enzyme was observed in cortex and a 1.45 fold increase in medullaryhomogenate (Table 4).
Acid phosphatase and total ATPase
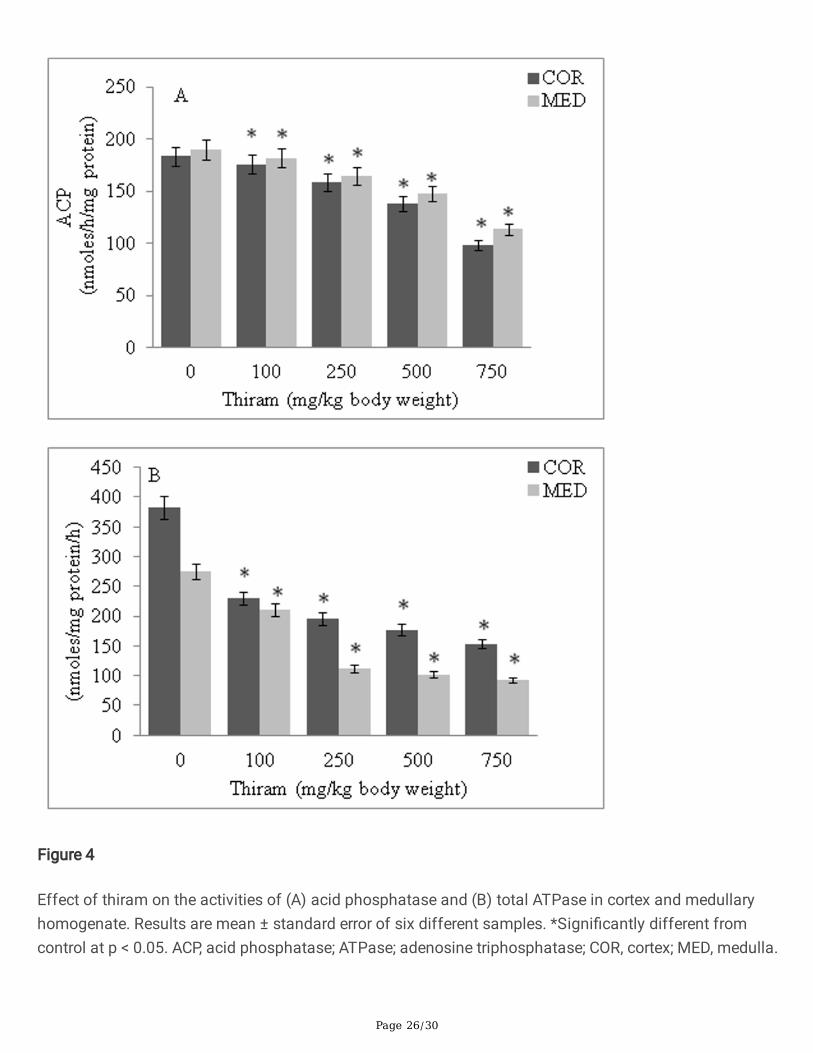
Both acid phosphatase and total ATPase showed a thiram dose-dependent decrease in activity. Theactivity of ACP declined to 53.40% in cortex and 59.65% in medullary homogenates (Fig 4A). TotalATPase was 40.20% in cortex and 33.74% in medullary homogenates, relative to control values (Fig 4B).
DNA damage
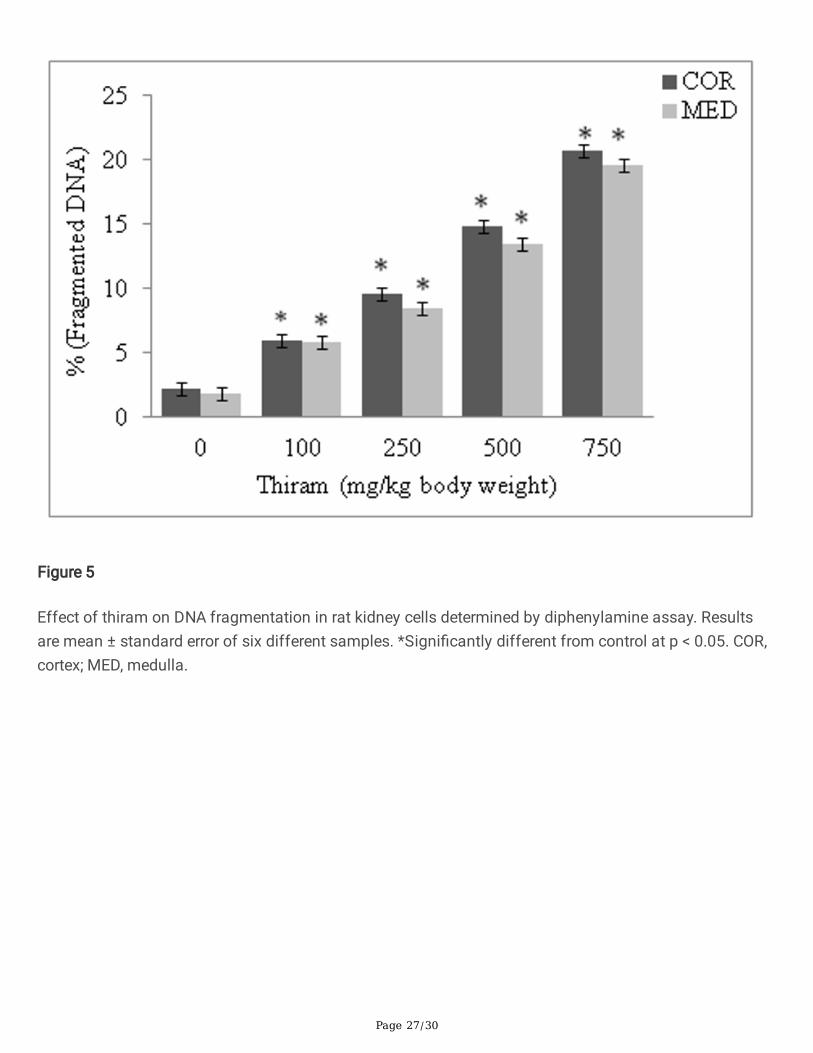
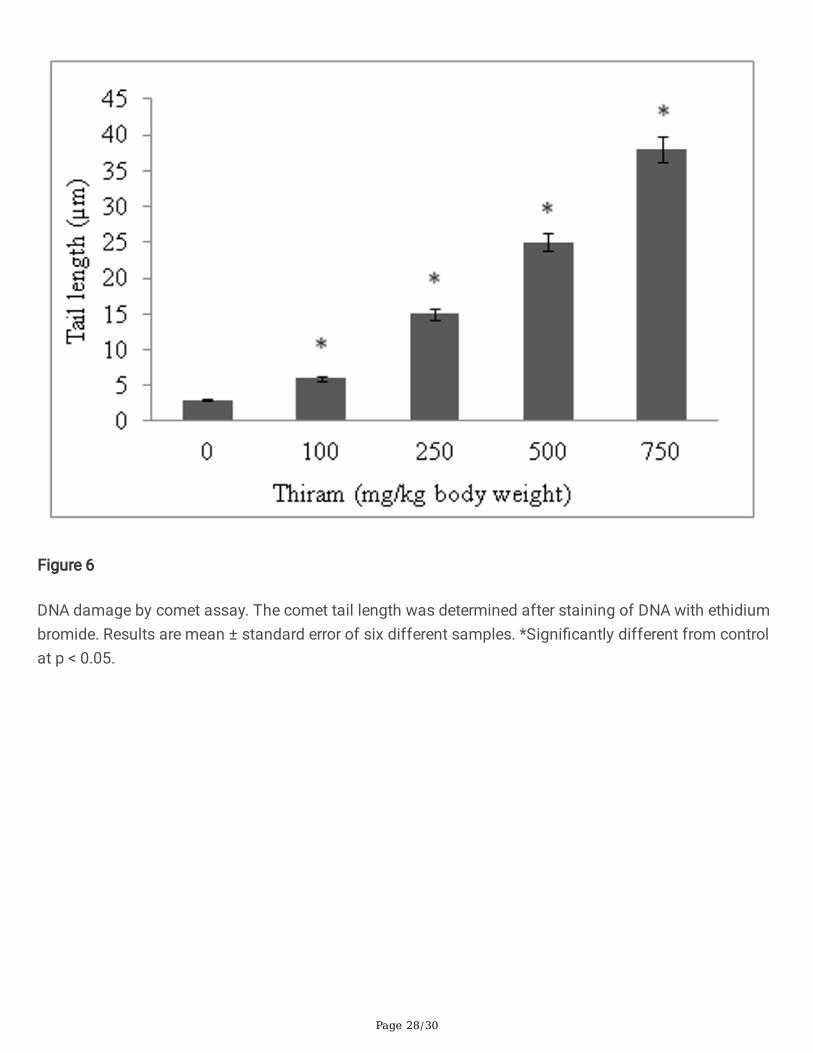
The genotoxic effect of thiram was determined by diphenylamine, DPC and comet assays. The levels offragmented DNA increase and was 9.7 fold in cortex and 10.8 fold in medullary homogenates, asmeasured by diphenylamine assay (Fig. 5). Similarly the levels of DPCs also increase in thiram treatedgroup compare to control group. The DPC increased by 2 fold in cortical and >2 fold in medullaryhomogenates (Table 5). An increase in tail length of DNA in comet assay was seen in a thiram dose-dependent manner. A 12.6 fold increase in comet tail length of DNA, relative to control, was seen at thehighest dose of thiram given to rats (Fig. 6).
Histopathology
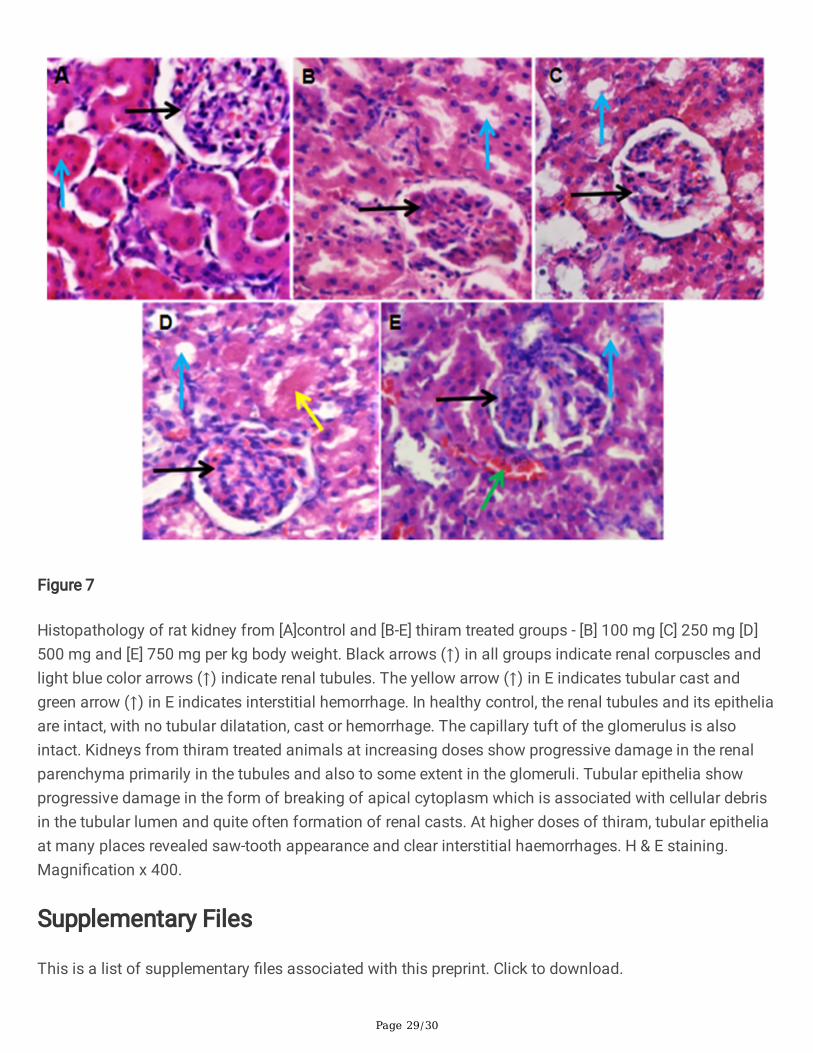
Histopathology of kidney tissues reveals normal structure in control group but a progressive damage intubular epithelia was observed at all doses of thiram. At 500 and 750 mg thiram/kg body weight, cellulardebris in tubular lumen, formation of tubular cast, saw tooth appearance and interstitial hemorrhage wereevident in cortex region of kidney (Fig 7).
DiscussionBUN and creatinine are metabolic waste products which are routinely excreted out of the body by kidney.A moderate increase in plasma creatinine and BUN levels was recorded in thiram treated rats. This couldbe due to decrease in glomerular �ltration that may have occurred upon reduced reabsorption by renaltubules because of degeneration of tubular epithelial cells. A similar trend in plasma BUN level wasobserved on oral administration of thiram in rats in a study conducted by Lee et al. (1978).
A signi�cant reduction in the activities of BBM enzymes; ALP, LAP, GGT and maltase was observed withincrease in dose of thiram in isolated BBMV and cortical homogenate. The decline in the speci�cactivities of these enzymes was more in isolated BBMV than in the homogenate. ALP and LAP aremetalloproteins whereas maltase and GGT have cysteine residues that are involved in binding ofsubstrate at the enzyme active site. The decrease in the activities of BBM marker enzymes could either bedue to metal chelating property of thiram or by formation of mixed disul�de with the critical cysteine

Page 10/30
residues, or their oxidation, present at the active site of enzyme (Marikovsky, 2002; Xu et al., 2017).Another reason could be loss of the enzymes into the lumen of the tubule upon membrane damage.
GSH, a tripeptide (glutamyl-cysteine-glycine) non-enzymatic antioxidant protects cells from free radicals.Within the cells GSH is found both in free form and bound to proteins. A gradual decrease in theconcentration of GSH and total SH was observed with increase in dose of thiram. This may be due tooxidation of thiol group of GSH by ROS generated by thiram, conversion to GSSG or formation ofdisul�de bridge with the thiram molecule. Another reason could be the reduced activity of enzyme GR thatregenerates GSH from GSSG (Cereser et al., 2001b). The increase in the levels of H2O2 could be due todecrease in the activities of AO enzymes CAT and GP that under normal conditions convert H2O2 into H2Oand O2. Carbonyl groups formed upon oxidation of side chain of amino acids are markers of proteinoxidation. A thiram dose-dependent increase in the protein oxidation was observed which could be due toincrease concentration of H2O2 that will lead to the formation of highly reactive hydroxyl radical (Parvezand Raisuddin, 2005).
Thiram exposure caused a signi�cant inhibition in the activities of primary (SOD, CAT and GP) as well assecondary (GR, TR and GST) AO enzymes. Thiram can chelate copper ion so inhibition in the enzymaticactivity of SOD could be due to sequestering of copper ion by thiram (Babo and Vasseur, 1992). CAT andGP convert H2O2 into H2O and O2 and the latter does so by conjugating it with GSH. The decrease in theactivity of enzyme GP could either be due to attack of ROS on selenium atom or carbomylation ofcysteine residue present at the active site. CAT can be inactivated by singlet oxygen generated fromexcess H2O2 (Kim et al., 2001). Elskens and Penninckx (1995) have reported that, in the presence ofNADPH, thiram acts as an inhibitor of GR. TR/thioredoxin system maintains thiol/disul�de redox balancein the cell and protects them from oxidative damage. A highly conserved sequence (Cys-Gly-Pro-Cys) ispresent at the active site and modi�cation of cysteine residues by thiram may be a reason for decreasedactivity of TR. GST is a major phase II detoxi�cation enzyme. It detoxi�es harmful xenobiotics that enterthe human body by conjugating them with GSH. Thiram, like other sufhydryl reagents (Smith andLitwack, 1989) may reduce the binding a�nity of substrate, 1-chloro 2,4-dinitrobenzene without alteringthe binding of glutathione and thus, decrease the activity of GST.
A dose-dependent decline in the non-enzymatic AO power of kidney cells also took place on exposure tothiram. Quenching of DPP• radical (DPPH assay) and reduction of ferric ion into ferrous ion (FRAP) byAOs present in cortex and medullary homogenate was used as a measure to quantify the AO power. Asigni�cant reduction in AO power was observed in both these assays. The reason may be diminishedenzymatic (AO enzymes) and non-enzymatic (GSH) AO defense system of the cell in thiram treatedanimals.
Hexokinase catalyzes the �rst step of glycolysis; it converts glucose to glucose 6-phosphate and commitsglucose to catabolism. The decreased activity of hexokinase, could be due to the interaction of thiramwith the free SH group present at the active site of this enzyme and essential for its catalytic function(Stromme, 1963). LDH converts pyruvate to lactate and regenerates NAD+. Increased activity of LDH

Page 11/30
could be attributed to its dissociation from membrane and its release into the cytosol (Rana andShivanaddappa, 2010). Devi et al. (2020) showed that binding of pesticides (rotenone and chlorpyrifos)induces alteration in the secondary structure of malate dehydrogenase and these conformationalchanges lead to the formation of cytotoxic conformers that generated oxidative stress. So, decrease inthe activity of malate dehydrogenase could be due to alteration in its structure occur due to binding ofthiram. Superoxide radical generated by reaction of H2O2 with Fe2+ can inhibit the enzymatic activity ofG6PDH (Bagnasco et al., 1991). Another reason could be decreased activity of hexokinase. Malic enzymedecarboxylates malate to pyruvate with concomitant reduction of NADP+ to NADPH. NADPH has twoimportant functions: it maintains the redox environment of cell and is involved in the synthesis of fattyacids. The increase in the activity of malic enzyme may be a defense mechanism of cell to protect itselffrom ROS by generating more NADPH (Corpas and Barroso, 2014). A signi�cant reduction in the activitiesof gluconeogenic enzymes, FBPase and G6Pase was seen with increase in dose of thiram. FBPaseconvert fructose 1,6-bisphosphate into fructose 6-phosphate while G6Pase dephosphorylate glucose 6-phosphate to glucose. The decrease in activity of G6Pase could be due to sequestering of metal ion bythiram (Nordlie and Johns, 1968) and that of FBPase due to increased generation of H2O2 (Halliwell andGutteridge, 1985).
ATPases are membrane bound enzymes that hydrolyze ATP to ADP and utilize the energy generatedduring this reaction to transport cations across the cell membrane. The decrease in the activity ofATPases may be due to the ability of dithiocarbamate to form complexes with metal activated enzymes(ATPases and oxidoreductase) (Owens, 1969) or changes in lipid composition and �uidity of plasmamembrane (Pentyalat and Chatty, 1993). Acid phosphatase performs diverse biological functionsincluding microbial killing, bone resorption, phosphate acquisition in plants and iron transport in animals.The activity of ACP widely depends on the reduced state of thiol group of cysteine residue present at itsactive site. Oxidation of this critical cysteine residue by ROS such as H2O2 may reduce the catalyticactivity of the enzyme (Gazo et al., 2015).
The genotoxic effect of thiram was studied using several methods. The level of fragmented DNAincreased in a thiram dose-dependent manner and these result are consistent with a previous studyconducted by Rath et al. (2005) in which they showed that thiram exposure induces DNA damage in tibialgrowth plate. A similar trend was observed in the other two assays used to study the genotoxic potentialof thiram. DPCs are formed when cells are exposed to xenobiotics like pesticides and are known to inhibitDNA metabolic processes such as replication and transcription. An error in DNA metabolic processesresults in the formation of sister chromatid exchange (SCE). Pienkowska and Zielenska (1990) showedthat thiram increases the frequency of SCE in human lymphocytes from this we can infer that thiram mayled to the formation of DPC. The comet assay is a sensitive method for detecting low levels of singlestrand breaks. A signi�cant increase in tail length of DNA was observed in a thiram dose-dependentmanner that corroborates the gentoxic effect of thiram (Emmanouil et al., 2008). Since previous studieshave shown the pro-oxidant nature of dithiocarbamate compound so we hypothesize that the thiram-induced DNA damage could be due to a compromised redox environment and greater generation of ROS

Page 12/30
that directly modify DNA bases and the deoxyribose sugar. The other reason could be the alkylation ofDNA, as seen in case of other dithiocarbamates (Gonzalez et al., 2003).
Histopathology of kidney tissues shows normal cellular architecture in control group but a progressivedamage in renal parenchyma was observed in thiram treated rats. At 100 and 250 mg/kg body weightdoses of thiram, degeneration of tubular epithelium was observed. At 500 mg/kg bw renal cast and at thehighest dose of thiram (750 mg/kg bw) interstitial hemorrhage and saw-tooth appearance were observedin the cortical part of kidney. The damage to tubule cells led to the formation of tubular cast in conditionssuch as necrosis while interstitial hemorrhages are associated with acute kidney injury.
ConclusionsThe �ndings of the present study suggested that acute exposure of male rats to thiram inducednephrotoxicity as evidenced by induction of oxidative stress, reduction in antioxidant power (enzymaticand non-enzymatic), metabolic dysfunction, biochemical and histological alterations and DNA damage.On the basis of our �ndings, we may propose that studies should be carried out to determine kidneyfunction in persons occupationally exposed to thiram.
DeclarationsFunding
The following schemes are gratefully acknowledged: UGC-SAP-DRS-III, DST-FIST-II and DST-PURSE forproviding �nancial aid to the Department of Biochemistry. SS is the recipient of Maulana Azad NationalFellowship sanctioned by UGC, New Delhi and AA is the recipient of Junior Research Fellowshipsanctioned by UGC, New Delhi.
Author Contributions
S.S performs experiments, analyzed data and wrote the manuscript. A.A performs experiment and carriedout statistical analysis. A.A.K performs Histopathology. R.M supervised the study, wrote and edited themanuscript as well as contributed to the interpretation of data. All authors revised, edited and approvedthe �nal version of the manuscript.
Data availability: All data generated or analyzed during this study are included in this article.
Compliance with ethical standards
Ethics approval: The animal experiments were done as per the guidelines provided by the InstitutionalEthics Committee (IEC) of Aligarh Muslim University that monitors research involving animals (R. No. :714/GO/Re/S/02/CPCSEA).
Consent to participate: Not applicable.

Page 13/30
Consent for publication: Not applicable.
Competing interest: The authors declare no con�ict of interest in this work.
ReferencesAebi H. 1984. Catalase in vitro. Methods Enzymol. 105, 121-126.
Agrawal RC, Shukla Y, Mehrotra NK. 1997. Assessment of mutagenic potential of thiram. Food ChemToxicol. 35, 523-525.
Alam JA, Bom L, Chbani A, Fajloun Z, Millet M. 2017. Analysis of dithiocarbamate fungicides invegetables matrices using HPLC-UV followed by atomic absorption spectrometry. J Chromatogr Sci.55(4), 429-435.
Babo S, Vasseur P. 1992. In vitro effects of thiram on liver antioxidant enzyme activities of rainbow trout(Oncorhynkus mykiss). Aquat Toxicol. 22, 61-68.
Bagnasco M, Camoirano A, De Flora S, Melodia F, Arillo A. 1991. Enhanced liver metabolism of mutagensand carcinogens in �sh living in polluted sea water. Mutat Res. 262, 129-137.
Behling EB, Sendao MC, Francescato HDC, Antunes LMG, Costa RS, MdeLP Bianchi. 2006. Comparativestudy of multiple dosage of quercetin against cisplatin-induced nephrotoxicity and oxidative stress in ratkidneys. Pharmacol Rep. 58(4), 526-532.
Belaid C, Sbartai I, Djebar MR. 2019. Populational effect of a dithiocarbamate (thiram) fungicide on afreshwater cladocerus Daphnia magna. Stud Univ Vasile Goldis Arad Ser Stiint Vietii. 29, 121-128.
Benzie IFF, Strain JJ. 1996. The ferric reducing ability of plasma (FRAP) as a measure of “antioxidantpower”: the FRAP assay. Anal Biochem. 239, 70-76.
Bergmeyer H. 1974. Enzymatic assay of maltase. In: Methods of enzymatic analysis, 2nd edn. AcademicPress, New York, pp 459-460.
Beutler E, Duron O, Kelly BM. 1963. Improved method for the determination of blood glutathione. J LabClin Med. 61, 882-888.
Bonting SL, Simon KA, Hawkins NM. 1961. Studies on sodium-potassium-activated adenosinetriphosphatase: I. quantitative distribution in several tissues of the cat. Arch Biochem Biophys. 95, 416-423.
Buac D, Schmitt S, Ventro G, Rani Kona F, Ping Dou Q. 2012. Dithiocarbamate-based coordinationcompounds as potent proteasome inhibitors in human cancer cells. Mini Rev Med Chem. 12(12), 1193-1201.

Page 14/30
Burton K. 1956. A study of the conditions and mechanism of the diphenylamine reaction for thecolorimetric estimation of deoxyribonucleic acid. Biochem J. 62(2), 315-323.
Carlberg I, Mannervik B. 1985. Glutathione reductase. Methods Enzymol. 113, 484-490.
Cereser C, Boget S, Parvaz P, Revol A. 2001a. An evaluation of thiram toxicity on cultured human skin�broblasts. Toxicology.162(2), 89-101.
Cereser C, Boget S, Parvaz P, Revol A. 2001b. Thiram induced cytotoxicity is accompanied by a rapid anddrastic oxidation of reduced glutathione with consecutive lipid peroxidation and cell death. Toxicology.163, 153-162.
Chen X, Fang M, Chernick M, Wang F, Yang J, Yu Y, Zheng N, Teraoka H, Nanba S, Hiraga T, Hinton DE,Dong W. 2018. The case for thyroid disruption in early life stage exposure to thiram in zebra�sh (Daniorerio). Gen Comp Endocr. 271, 73-81.
Corpas FJ, Barroso JB. 2014. NADPH-generating dehydrogenases: their role in the mechanism ofprotection against nitro-oxidative stress induced by adverse environmental conditions. Front Environ Sci.2(55), 1-5.
Crane RK, Sols A. 1953. The association of hexokinase with particulate fractions of brain and other tissuehomogenates. J Biol Chem. 203, 273-292.
Culling CFA. 2013. Handbook of histopathological and histochemical techniques: including museumtechniques. Butterworth-Heinemann.
Devi S, Karsauliya K, Srivastava T, Raj R, Kumar D, Priya S. 2020. Pesticides interactions inducealterations in secondary structure of malate dehydrogenase to cause destability and cytotoxicity.Chemosphere. 263, 128074.
Elskens MT, Penninckx MJ. 1995. In vitro inactivation of yeast gluthathione reductase bytetramethylthiuram disulphide. Eur J Biochem. 231, 667-672.
Emmanouil C, Kypriotakis S, Kungolos A, Machers K. 2008. Effect of thiram on MCPA acid on musell gillDNA. Discussion paper series. 14(24), 455-468.
Flohe L, Gunzler WA. 1984. Assays of glutathione peroxidase. Methods Enzymol. 105, 114-121.
Freedland RA, Harper AE. 1959. The study of metabolic pathways by means of metabolic adaptations. JBiol Chem. 234, 1350-1354.
Gay CA, Gebicki JM. 2000. A critical evaluation of the effect of sorbitol on the ferric-xylenol orangehydroperoxide assay. Anal Biochem. 284(2), 217-220.

Page 15/30
Gazo I, Koleova AS, Dietrich MA, Linhartova P, Shaliutina O, Cosson J. 2015. The effect of ractive oxygenspecies on motality parameters, DNA integrity, tyrosine phosphorylation and phosphatase activity ofCommon carp (Cyprinus carpio L.) spermatozoa. Mol Reprod. 82, 48-57.
Goldmann DR, Schlesinger H, Segal S. 1976. Isolation and characterization of the brush border fractionfrom newborn rat renal proximal tubule cells. Biochim Biophys Acta BBA-Biomembr. 419, 251-260.
Gonzalez M, Soloneski S, Reigosa MA, Larramendy ML. 2003. Effect of dithiocarbamate pesticide zineband its commercial formulation, azzuro IV. DNA damage and repair kinetics assessed by single cell gelelectrophoresis (SCGE) assay on chinese hamster ovary (CHO) cells. Mutat Res. 534, 145-154.
Grosicka-Maciag E, Kurpios D, Czeczot H, Szumilo M, Skrzycki M, Suchocki P, I Rahden-Staron I. 2008.Changes in antioxidant defense systems induced by thiram in V79 chinese hamster �broblasts. Toxicol InVitro. 22(1), 28-35.
Habig WH, Pabst MJ, Jakoby WB. 1974. The �rst enzymatic step in mercapturic acid formation. JBiol Chem. 249, 7130-7139.
Halliwell B, Gutteridge JMC. 1985. Free radicals in biology and medicine, Clarendon. Press, Oxford.
Han MS, Shin KJ, Kim YH, Kim SH, Lee T, Kim E, Ryu SH, Suh PG. 2003. Thiram and ziram stimulate non-selective cation channel and induce apoptosis in PC12 cells. Neurotoxicology. 24(3), 425-434.
Haugen HN. 1953. The determination of endogenous creatinine in plasma and urine. Scand J Clin LabInvest. 5(1), 48-57.
Kanecoa S, Lia N, Itoh KK, Katsumata H, Suzuki T, Ohta K. 2009. Titanium dioxide mediated solarphotocatalytic degradation of thiram in aqueous solution: Kinetics and mineralization. Chem Eng J.148(1), 50-56.
Kempson SA, Kim JK, Northrup TE, Knox FG, Dousa TP. 1979. Alkaline phosphatase in adaptation to lowdietary phosphate intake. Am J Physiol. 237, 465-473.
Khan PK, Sobti RC, Kataria L. 2005. Pesticide induced alteration in mice hepatooxidative status andprotective effect of black tea extract. Clin Chim Acta. 358, 131-138.
Khundmiri SJ, Asghar M, Khan F, Salim S, Yusu� AN. 2004. Effect of ischemia and reperfusion onenzymes of carbohydrate metabolism in rat kidney. J Nephrol. 17, 377-383.
Khundmiri SJ, Asghar M, Khan F, Salim S, Yusu� ANK. 1997. Effect of reversible and irreversible ischemiaon marker enzymes of BBM from renal cortical PT subpopulations. Am J Physiol. 273, F849-856.
Kim SY, kwon OJ, Park JW. 2001. Inactivation of catalase and superoxide dismutase by singlet oxygenderived from photoactivated dye. Biochemia. 83, 437-444.

Page 16/30
Kurpios-Piec D, Grosicka-Maciąg E, Wozniak K, Kowalewski C, Kiernozek E, Szumilo M, Rahden-Staron I.2015. Thiram activates NF-kappa B and enhances ICAM-1 expression in human microvascularendothelial HMEC-1 cells. Pestic Biochem Physiol. 118, 82-89.
Lee C, Russell JQ, Minor JL. 1978. Oral toxicity of ferric dimethyl‐dithiocarbamate (ferbam) andtetramethylthiuram disul�de (thiram) in rodents. J Toxicol Environ. 4(1), 93-106.
Levine RL, Garland D, Oliver CN, Amici A, Climent I, Lenz AG, Ahn BW, Shaltiel S, Stadtman ER. 1990.Determination of carbonyl content in oxidatively modi�ed proteins. Methods Enzymol. 186, 464-478.
Lopez-Antia A, Ortiz-Santaliestra M, Garcia-de Blas E, Camarero P, Mougeot F, Mateo R. 2015. Adverseeffects of thiram-treated seed ingestion on the reproductive performance and the offspring immunefunction of the red-legged partridge. Environ Toxicol Chem. 34(6), 1320-1329.
Lorgue G, Lechenet J, Riviere A, Chapman MJ. 1996. Clinical veterinary toxicology. Blackwell Science.
Lowry OH, Rosenbrough NJ, Farr AL, Randall RJ. 1951. Protein measurement with the Folin phenolreagent. J Biol Chem. 193, 265-275.
Maita K, Tsuda S, Shirasu Y. 1991. Chronic toxicity studies with thiram in wistar rats and beagle dogs.Fundam Appl Toxicol. 16(4), 667-686.
Mansour SA, Mossa AH. 2010. Oxidative damage, biochemical and histopathological alterations in ratsexposed to chlorpyrifos and the antioxidant role of zinc. Pestic Biochem Physiol. 96, 14-23.
Marikovsky M. 2002. Thiram inhibit angiogenesis and inhibits the development of experimental tumoursin mice. Br J Cancer. 86, 779-787.
Marklund S, Marklund G. 1974. Involvement of the superoxide anion radical in the autoxidation ofpyrogallol and a convenient assay for superoxide dismutase. Eur J Biochem. 47, 469-474.
Mehler AH, Kornberg A. 1948. The enzymatic mechanism of oxidation-reductions between malate orisocitrate and pyruvate. J Biol Chem. 174, 961-977.
Mishra K, Ojha H, Chaudhury NK. 2012. Estimation of antiradical properties of antioxidants using DPPHassay: a critical review and results. Food Chem. 130, 1036-1043.
Mohrenweiser HW, Novotny JE. 1982. ACP1GUA-1-a low-activity variant of human erythrocyte acidphosphatase: association with increased glutathione reductase activity. Am J Hum Genet. 34, 425-433.
Nageswara RNR, Srinivasa NB, Jagadish MN. 2017. Thiram a fungicide induced toxicity on glycogen andblood glucose level of freshwater �sh Cyprinus carpio (Hamilton). Int J Fish Aquat Stud. 5(6), 93-96.
Nordlie RC, Johns PT. 1968. The Inhibition of microsomal glucose 6-phosphatase by metal-bindingagents. Biochemistry. 7(4), 1473-1479.

Page 17/30
Ochoa S, Mehleb A, Kornberg A. 1948. Biosynthesis of dicarboxylic acids by carbon dioxide �xation.Isolation and properties of an enzyme from pigeon liver catalyzing the reversible oxidativedecarboxylation of malic acid. J Biol Chem. 174, 979-1000.
Owens RG. 1969. Chemical and physical interactions, In Fungicides, Torgoson DC (Ed.), Academic Press,New York, pp. 249-265.
Parvez S, Raisuddin S. 2005. Protein carbonyls: novel biomarkers of exposure to oxidative stress-inducingpesticides in freshwater �sh Channa punctata (Bloch). Environ Toxicol Pharmacol. 20, 112-117.
Pentyalat SN, Chetty CS. 1993. Comparative study on the changes in AChE and ATPase activities inneonate and adult rat brains under thiobencarb stress. J Appl Toxicol. 13(1), 39-42.
Perocco P, Santucci MA, Campani AG, Forti GC. 1989. Toxic and DNA damaging activities of thefungicides mancozeb and thiram (TMTD) on human lymphocytes in vitro. Teratog Carcinog Mutagen. 9,75-81.
Pienkowska M, Zielenska M. 1990. Genotoxic effects of thiram evaluated by sister-chromatid exchangesin human lymphocytes. Mutat Res. 245, 119-123.
Rana I, Shivanandappa T. 2010. Mechanism of potentiation of endosulfan cytotoxicity by thiram inehrlich ascites tumor cells. Toxicol In Vitro. 24, 40-44.
Rath NC, Richards MP, Huff WE, Huff GR, Balo J. 2005. Changes in tibial growth plates of chickens withthiram-induced dyschondroplasia. J Comp Path. 133, 41-52.
Rosenthal HL. 1955. Determination of urea in blood and urine with diacetyl monoxime. Anal Chem.27(12), 1980-1982.
Saqib TA, Naqvi SN, Siddiqui PA, Azmi MA. 2005. Detection of pesticide residues in muscles, liver and fatof 3 specise of labeo found in kalri and haleji lakes. J Environ Biol. 26(2), 433-438.
Schetz M, Dasta J, Goldstein S, Golper T. 2005. Drug-induced acute kidney injury. Curr Opin Crit Care.11(6), 555-565.
Sedlak J, Lindsay RH. 1968. Estimation of total, protein-bound and non-protein sulfhydryl groups in tissuewith Ellman's reagent. Anal Biochem. 25, 192-205.
Sharma VK, Aulakh JS, Malik AK. 2003. Thiram: degradation, applications and analytical methods. JEnviron Monit. 5, 717-723.
Shonk CC, Boxer GE. 1964. Enzyme patterns in human tissues I. Methods for the determination ofglycolytic enzymes. Cancer Res. 24, 709-721.

Page 18/30
Shull KH, Ashmore J, Mayer J. 1956. Hexokinase, glucose-6-phosphatase and phosphorylase levels inhereditarily obese-hyperglycemic mice. Arch Biochem Biophys. 62, 210-216.
Singh NP, McCoy MT, Tice RR, Schneider EL. 1988. A simple technique for quantitation of low levels ofDNA damage in individual cells. Exp Cell Res. 175(1), 184-191.
Smith GJ, Litwack G. 1989. Roles of ligandin and the glutathione S-transferase in binding steroidmetabolites, carcinogens and other compounds. Biochem Toxicol. 2, 1-47.
Stoker TE, Jerome M, Goldman JM, Cooper RL. 1993. The dithiocarbamate fungicide thiram disrupt thehormonal control of ovulation in the female rat. Reprod Toxicol. 7, 211-218.
Stromme JH. 1963. Inhibition of hexokinase by disul�ram and diethyldithiocarbamate. BiochemPharmacol. 12, 157-166.
Tamura T, Stadtman TC. 1996. A new selenoprotein from human lung adenocarcinoma cells: puri�cation,properties and thioredoxin reductase activity. Proc Natl Acad Sci USA. 93, 1006-1011.
Taussky HH, Shorr E. 1953. A microcolorimetric method for the determination of inorganic phosphorus. JBiol Chem 202, 675–685.
Thind TS, Hollomon DW. 2017. Thiocarbamate fungicides: reliable tools in resistance management andfuture outlook. Pest Manag Sci. 74(7), 1546-1551.
Valko M, Leibfritz D, Moncol J, Cronin MTD, Mazur M, Telser J. 2007. Free radicals and antioxidants innormal physiological functions and human disease. Int J Biochem Cell Biol. 39(1), 44-84.
Waqas M, Qamar H, Zhang J, Yao W, Li A, Wang Y, Iqbal M, Mehmood K, Jiang X, Li J. 2020. Puerarinenhance vascular proliferation and halt apoptosis in thiram-induced avian tibial dyschndroplasia byregulating HIF-1α, TIMP-3 and BCL-2 expressions. Ecotoxicol Environ Saf. 190, 110126.
Werner M, Costa MJ, Mitchell LG, Nayar R. 1995. Nephrotoxicity of xenobiotics. Clinica Chimica Acta.237(1-2), 107-154.
Xu X, Mathieu C, Berthelet J, Duval R, Bui LC, Busi F, Dupret JM, Rodrigues-Lima F. 2017. Humanarylamine N-acetyltransferase-1 is inhibited by the dithiocarbamte pesticide thiram. Mol Pharmacol.92(3), 358-365.
Zager RA. 1997. Pathogenetic mechanisms in nephrotoxic acute renal failure. Semin Nephrol. 17(1), 3-14.
Zhitkovich A, Costa M. 1992. A simple, sensitive assay to detect DNA-protein crosslinks in intact cells andin vivo. Carcinogenesis. 13(8), 1485-1489.
Tables
Page 19/30

Page 20/30
Page 21/30

Page 22/30
Figures
Page 23/30
Figure 1
Effect of thiram on (A) BUN and (B) creatinine levels in plasma. Results are mean ± standard error of sixdifferent samples. *Signi�cantly different from control at p < 0.05. BUN, blood urea nitrogen; COR, cortex;MED, medulla.
Page 24/30
Figure 2
Effect of thiram on non-enzymatic parameters of oxidative stress. (A) GSH (B) total SH (C) proteincarbonyls and (D) H2O2. Results are mean ± standard error of six different samples. *Signi�cantlydifferent from control at p < 0.05.
Page 25/30
Figure 3
Effect of thiram on antioxidant capacity as determined by (A) FRAP and (B) DPPH assays using cortexand medullary homogenates. Results are mean ± standard error of six different samples. *Signi�cantlydifferent from control at p < 0.05. FRAP, ferric reducing antioxidant power; DPPH, 2,2-diphenyl-1-picrylhydrazyl; COR, cortex; MED, medulla.
Page 26/30
Figure 4
Effect of thiram on the activities of (A) acid phosphatase and (B) total ATPase in cortex and medullaryhomogenate. Results are mean ± standard error of six different samples. *Signi�cantly different fromcontrol at p < 0.05. ACP, acid phosphatase; ATPase; adenosine triphosphatase; COR, cortex; MED, medulla.
Page 27/30
Figure 5
Effect of thiram on DNA fragmentation in rat kidney cells determined by diphenylamine assay. Resultsare mean ± standard error of six different samples. *Signi�cantly different from control at p < 0.05. COR,cortex; MED, medulla.
Page 28/30
Figure 6
DNA damage by comet assay. The comet tail length was determined after staining of DNA with ethidiumbromide. Results are mean ± standard error of six different samples. *Signi�cantly different from controlat p < 0.05.
Page 29/30
Figure 7
Histopathology of rat kidney from [A]control and [B-E] thiram treated groups - [B] 100 mg [C] 250 mg [D]500 mg and [E] 750 mg per kg body weight. Black arrows (↑) in all groups indicate renal corpuscles andlight blue color arrows (↑) indicate renal tubules. The yellow arrow (↑) in E indicates tubular cast andgreen arrow (↑) in E indicates interstitial hemorrhage. In healthy control, the renal tubules and its epitheliaare intact, with no tubular dilatation, cast or hemorrhage. The capillary tuft of the glomerulus is alsointact. Kidneys from thiram treated animals at increasing doses show progressive damage in the renalparenchyma primarily in the tubules and also to some extent in the glomeruli. Tubular epithelia showprogressive damage in the form of breaking of apical cytoplasm which is associated with cellular debrisin the tubular lumen and quite often formation of renal casts. At higher doses of thiram, tubular epitheliaat many places revealed saw-tooth appearance and clear interstitial haemorrhages. H & E staining.Magni�cation x 400.
Supplementary Files
This is a list of supplementary �les associated with this preprint. Click to download.

Page 30/30
Graphicalabstract.docx
Highlights.docx
![Jules Bordet La Lasertherapie o[/v ] µ · • Rapport eau libre/eau liée des tissus • ->diminution de l’œdème • ->perméabilité accrue des lymphatiques • Diminution des](https://img.pdfslide.us/doc/110x75/5b996b6809d3f2cb468bc717/jules-bordet-la-lasertherapie-ov-rapport-eau-libreeau-liee-des.jpg)




























