Embed Size (px)
Citation preview

2393Journal of Cell Science 108, 2393-2404 (1995)Printed in Great Britain © The Company of Biologists Limited 1995
Chromatin diminution is strictly correlated to somatic cell behavior in early
development of the nematode Parascaris univalens
M. R. Esteban, G. Giovinazzo and C. Goday*
Centro de Investigaciones Biológicas, Consejo Superior de Investigaciones Científicas, Velázquez 144, 28006 Madrid, Spain
*Author for correspondence
We have studied the relationship between the occurrenceof chromatin diminution and the developmental behaviorof somatic blastomeres in early development of thenematode Parascaris univalens. A cytological and immuno-cytochemical analysis of chromatin diminution wasperformed in P. univalens embryos exposed to ‘vegetaliz-ing’ (LiCl) and ‘animalizing’ (NaSCN) substances duringearly developmental stages. We have also analyzedchromatin diminution in embryos displaying only symmet-rical ‘somatic-like’ divisions due to a brief cytochalasin Btreatment at the pronuclear stage. The results show thatLiCl treatment induces chromatin diminution in P0-P4pregerminal blastomeres while NaSCN treatment preventsit. Pregerminal cells undergoing chromatin diminution in‘vegetalized’ embryos behaved like somatic cells withrespect to division and cleavage patterns. NaSCN
treatment results in undiminuted polynucleated embryosthat are not capable of cleavage. In cytochalasin B-pulsedembryos, chromatin diminution occurs in all blastomeres.From our results we conclude that chromatin diminutionand somatic cell behavior are not separable and constitutestrictly correlated events in Parascaris. Moreover, theresults indicate that the segregation of the cytoplasmicfactors involved in chromatin diminution in early Paras-caris development are microfilament-mediated. Here, wealso report the formation of a latter pregerminal cellprecursor (P5) not susceptible to LiCl-induced chromatindiminution.
Key words: nematode, Ascaridae, Parascaris univalens, chromatindiminution, vegetalization
SUMMARY
INTRODUCTION
A common feature in early development of nematodes is thata series of asymmetrical cleavages invariably occur during theseparation and establishment of germ versus somatic celllineages. This was first described by Boveri (1887) in hisclassical developmental studies on the parasitic nematodeParascaris. In Parascaris, in addition to differences in cellsize, cell cycle periods and spindle orientation, germlineprecursor cells and somatic line blastomeres greatly differ inboth DNA content and organization of chromatin. This is theresult of chromatin diminution, a process that consists of chro-mosome fragmentation and elimination of heterochromaticregions in all presomatic blastomeres at the initial stages ofembryogenesis (Fig. 1) (Boveri, 1887; Goday and Pimpinelli,1984). Since the discovery of chromatin diminution in Paras-caris, Boveri’s hypothesis on the significance of this phenom-enon with respect to germ vs soma differentiation has beengenerally accepted: in the nematodes Ascaridae, germline-limited chromatin is essential for the germinal quality of a blas-tomere while elimination of chromatin is necessary to ensurethe somatic fate of a blastomere. Recent evidence thatchromatin diminution in Ascaris suum and Parascarisunivalens involves the loss of single-copy germline-specificgenes constitutes the first molecular support to Boveri’s
hypothesis (Etter et al., 1991; Spicher et al., 1994). Moreover,it has been proposed that genome rearrangements due tochromatin diminution may provide a precise mechanism tocontrol gene expression and gene dosage in somatic cellsduring differentiation in these nematodes (Tobler et al., 1992;Goday and Pimpinelli, 1993). Boveri also concluded that thedifferential distribution of the egg cytoplasm by asymmetricalcleavages controls the early germ vs soma decision in Paras-caris (Boveri, 1910). A series of experiments by Boveri andother investigators have proven that specific, although yetunknown, cytoplasmic factors, most probably containingRNA, prevent chromatin diminution in pregerminal blas-tomeres (reviewed by Tobler et al., 1992). Alternative modelsfavouring the existence of chromatin diminution ‘activator’factors, differentially localized in the egg, have also beenreported (King and Beams, 1938; Seidl et al., 1988; reviewedby Tobler et al., 1992). However, the problem remainsunsolved since the cytoplasmic factors controlling chromatindiminution have not been yet identified.
Developmental studies in the free-living nematodeCaenorhabditis elegans where chromatin diminution has notbeen detected, have provided further information about the earlycell division patterns that characterize presomatic and pregermi-nal cells in nematodes. Early asymmetrical or unequal‘germline-like’ cleavages and symmetrical or equal ‘somatic-
2394 M. R. Esteban, G. Giovinazzo and C. Goday
like’ cleavages are known to be caused by differences in the ori-entation of the mitotic spindle which is directed by the centro-somes (Hyman and White, 1987; Hyman, 1989). Centrosomecell location, in turn, depends on the cytoskeleton microfilamentsystem. In this respect, treatment of C. elegans embryos with themicrofilament inhibitor cytochalasin B (CB), during a criticalshort period at the pronuclear stage, alters the typical earlycleavage pattern and produces symmetrical divisions (Stromeand Wood, 1983). Moreover, in C. elegans embryos, specificcytoplasmic factors called ‘P granules’ have been found to beconcentrated and distributed to pregerminal blastomeres fromthe very first asymmetrical division (Strome and Wood, 1982).Since P granule distribution is altered in CB-pulsed embryos,their location also depends on the microfilament system (Stromeand Wood, 1983). Despite their specific association to pregermi-nal blastomeres, it has not yet been proven that P granules con-stitute germinal determinants. In C. elegans early development,the strict correlation between unequal divisions, unequal germ-granule partitioning, and the generation of daughter blastomereswith different cell cycle timing, led to the conclusion that thethree processes are coupled (Hill and Strome, 1990). A class ofmaternal effect lethal mutants designated par (partitioning-defective) are known to affect the partitioning of cytoplasmicfactors controlling the cell cycle timing, cell division patternsand differentiation pathway of early C. elegans blastomeres(Kemphues et al., 1988; Kirby et al., 1990; Morton et al., 1992).In par mutants, asymmetric cleavages and P granule localizationare altered in a manner similar to that of embryos treated with
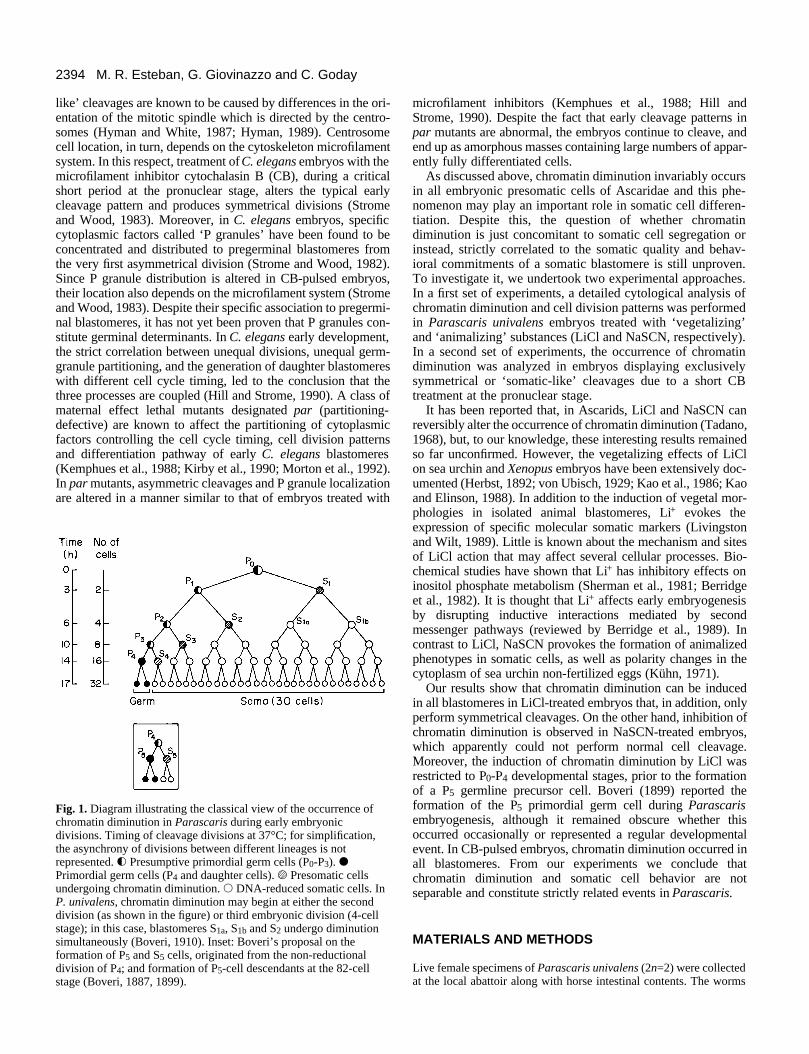
Fig. 1. Diagram illustrating the classical view of the occurrence ofchromatin diminution in Parascaris during early embryonicdivisions. Timing of cleavage divisions at 37°C; for simplification,the asynchrony of divisions between different lineages is notrepresented. f Presumptive primordial germ cells (P0-P3). dPrimordial germ cells (P4 and daughter cells). s Presomatic cellsundergoing chromatin diminution. s DNA-reduced somatic cells. InP. univalens, chromatin diminution may begin at either the seconddivision (as shown in the figure) or third embryonic division (4-cellstage); in this case, blastomeres S1a, S1b and S2 undergo diminutionsimultaneously (Boveri, 1910). Inset: Boveri’s proposal on theformation of P5 and S5 cells, originated from the non-reductionaldivision of P4; and formation of P5-cell descendants at the 82-cellstage (Boveri, 1887, 1899).
microfilament inhibitors (Kemphues et al., 1988; Hill andStrome, 1990). Despite the fact that early cleavage patterns inpar mutants are abnormal, the embryos continue to cleave, andend up as amorphous masses containing large numbers of appar-ently fully differentiated cells.
As discussed above, chromatin diminution invariably occursin all embryonic presomatic cells of Ascaridae and this phe-nomenon may play an important role in somatic cell differen-tiation. Despite this, the question of whether chromatindiminution is just concomitant to somatic cell segregation orinstead, strictly correlated to the somatic quality and behav-ioral commitments of a somatic blastomere is still unproven.To investigate it, we undertook two experimental approaches.In a first set of experiments, a detailed cytological analysis ofchromatin diminution and cell division patterns was performedin Parascaris univalens embryos treated with ‘vegetalizing’and ‘animalizing’ substances (LiCl and NaSCN, respectively).In a second set of experiments, the occurrence of chromatindiminution was analyzed in embryos displaying exclusivelysymmetrical or ‘somatic-like’ cleavages due to a short CBtreatment at the pronuclear stage.
It has been reported that, in Ascarids, LiCl and NaSCN canreversibly alter the occurrence of chromatin diminution (Tadano,1968), but, to our knowledge, these interesting results remainedso far unconfirmed. However, the vegetalizing effects of LiClon sea urchin and Xenopus embryos have been extensively doc-umented (Herbst, 1892; von Ubisch, 1929; Kao et al., 1986; Kaoand Elinson, 1988). In addition to the induction of vegetal mor-phologies in isolated animal blastomeres, Li+ evokes theexpression of specific molecular somatic markers (Livingstonand Wilt, 1989). Little is known about the mechanism and sitesof LiCl action that may affect several cellular processes. Bio-chemical studies have shown that Li+ has inhibitory effects oninositol phosphate metabolism (Sherman et al., 1981; Berridgeet al., 1982). It is thought that Li+ affects early embryogenesisby disrupting inductive interactions mediated by secondmessenger pathways (reviewed by Berridge et al., 1989). Incontrast to LiCl, NaSCN provokes the formation of animalizedphenotypes in somatic cells, as well as polarity changes in thecytoplasm of sea urchin non-fertilized eggs (Kühn, 1971).
Our results show that chromatin diminution can be inducedin all blastomeres in LiCl-treated embryos that, in addition, onlyperform symmetrical cleavages. On the other hand, inhibition ofchromatin diminution is observed in NaSCN-treated embryos,which apparently could not perform normal cell cleavage.Moreover, the induction of chromatin diminution by LiCl wasrestricted to P0-P4 developmental stages, prior to the formationof a P5 germline precursor cell. Boveri (1899) reported theformation of the P5 primordial germ cell during Parascarisembryogenesis, although it remained obscure whether thisoccurred occasionally or represented a regular developmentalevent. In CB-pulsed embryos, chromatin diminution occurred inall blastomeres. From our experiments we conclude thatchromatin diminution and somatic cell behavior are notseparable and constitute strictly related events in Parascaris.
MATERIALS AND METHODS
Live female specimens of Parascaris univalens (2n=2) were collectedat the local abattoir along with horse intestinal contents. The worms

2395Chromatin diminution and somatic cells in Parascaris
were washed and maintained in a balanced salt solution (0.7% NaCl)at 37°C until dissected to remove gonads. Fertilized eggs wereextracted from the uteri and kept at 4°C. All eggs were dechorionatedand permeabilized as previously described (Goday et al., 1992),except that during treatment with 0.4 M KOH/17% NaOCl theembryos were held at room temperature to avoid growth. To obtaindechorionated embryos at different developmental stages, embryoswere incubated either in M9 buffer, pH 7 (Brenner, 1974) or 0.9%NaCl at 37°C and development was followed by phase contrast andNomarski optics. Since the efficiency of the dechorionation and per-meabilization method was not complete in all embryos, only thoseembryos lacking the impermeable eggshell layers were analyzed aftereach treatment as well as in control experiments. This was confirmedby observations with phase-contrast and Nomarski optics.
LiCl treatment Separate groups of embryos from the P0 cell stage (zygote) up to theP4 cell stage (16-cell embryo) were transferred in a 500 mM LiCl in0.9% NaCl solution. The embryo suspension was loaded into asyringe and passed through a 25-gauge needle. The embryos werethen transferred to Petri dishes and allowed to develop in 500 mMLiCl medium at 37°C for 5 hours. Part of these embryos were thenfixed as described below. The rest were washed six times in 0.9%NaCl at room temperature and allowed to develop further in 0.9%NaCl for 4, 18, 25 and 72 hours at 37°C. Embryos were then fixedand either Hoechst 33258 stained or processed for immunofluores-cence as described below. In control experiments, permeabilizedembryos from the P0 to P4 cell stages were transferred in 0.9% NaClsolution and processed as described.
To analyze the effects of LiCl on the descendants of cell P4, embryoswere allowed to develop in 0.9% NaCl to the 32-cell stage (17 hoursat 37°C) and first larval stage (72 hours at 37°C). They were then treatedwith a 500 mM LiCl in 0.9% NaCl solution as described above andincubated in the LiCl solution for 5, 8, 16, 24 and 40 hours at 37°Cbefore being fixed. For the analysis of untreated control embryos, per-meabilized fertilized eggs were allowed to develop in 0.9% NaClsolution at 37°C. Part of these embryos were collected and fixed atdifferent times of incubation (every 45 minutes) to examine them beforethe 32-cell stage. They were Hoechst 33258 stained to observe theoccurrence of chromatin diminution during early development. Whenchromatin diminution started at the 4-cell stage in three presomatic blas-tomeres simultaneously, it was considered to reflect two separate reduc-tional processes (S1 daughters and S2). The rest of the untreated controlembryos, from the 32-cell stage onwards, were allowed to developfurther in 0.9% NaCl for 4, 8, 18, 25 and 72 hours at 37°C. They werefixed and Hoechst 33258 stained as described below.
NaSCN treatment Embryos from zygote to 4-cell stages were transferred to a 250 mMNaSCN in 0.9% NaCl solution. The suspension was loaded into asyringe and passed through a 25-gauge needle. Embryos were thentransferred to Petri dishes and incubated in the NaSCN medium at37°C for 5 hours. Part of these embryos were then fixed as describedbelow and the rest washed six times in 0.9% NaCl at room tempera-ture and allowed to develop further in 0.9% NaCl at 37°C before beingfixed. In control experiments, permeabilized embryos from zygote to4-cell stages were transferred to 0.9% NaCl solution and processed asdescribed.
Cytochalasin B (CB) treatment Dechorionated fertilized eggs were transferred to Petri dishes,incubated in M9 buffer at 37°C and development was followed byNomarski optics. It was determined that the duration of the pronu-clear migration period in Parascaris is 2 hours. DihydrocytochalasinB (CB) (Sigma) was stored as a 1 mg/ml stock solution in DMSOat 4°C and diluted in M9 buffer to a 16 µM final concentration
immediately before use. In CB-pulsed embryos, dechorionated fer-tilized eggs were first incubated at 37°C in the presence of CBduring the 2 hour interval of pronuclear migration. Embryos werethen washed six times at room temperature with M9 buffer andincubated in M9 buffer during 4, 18, 25 and 72 hours at 37°C.Embryos were fixed and processed for immunofluorescence andHoechst 33258 staining as described below. In control experiments,permeabilized fertilized eggs were incubated in M9 buffer andprocessed as described.
Fixation and Hoechst 33258 stainingEmbryos were fixed with a mixture of methanol, acetic acid and chlo-roform (6:3:1) for 1 hour at room temperature or overnight at 4°C.Embryos were then washed twice with distilled water and stained ina 2.5 µg/ml Hoechst 33258 aqueous solution for 10 minutes. Thestaining solution was replaced by antifading solution consisting of 1mg/ml p-phenylendiamine (Sigma) in a 1:10 (v/v) mixture of PBS:glycerol (Johnson and de Nogueira Araujo, 1981). A drop of theembryo suspension was placed on a slide and the embryos wereallowed to stick. A coverslip was deposited on the top and gentlypressed before preparations were sealed with Rubber Cementadhesive.
Indirect immunofluorescenceFixationEmbryos were fixed in 100% methanol at −20°C for 10 minutes andthen permeabilized in acetone at −20°C for 1 minute. They werewashed in PBS for 5 minutes at room temperature three times, treatedwith 1% Triton X-100 in PBS for 10 minutes and subsequentlywashed in three changes of PBS for 5 minutes each. Before incuba-tion with antibodies, embryos were incubated with 1% BSA (bovineserum albumin; Sigma) in PBS at room temperature for 30 minutes.
Incubation with antibodiesPrimary and secondary antibodies were diluted in 3% BSA in PBSand used as follows: anti-tubulin YL1/2 (1:100) (Sera-lab) followedby FITC-conjugated anti-rat (1:100) (Sigma). The embryo suspensionwas incubated with primary antibody for 1 hour at room temperature,washed three times in 1% BSA in PBS for 7 minutes each, andincubated with secondary antibody for 45 minutes at room tempera-ture. Embryos were washed three times in 1% BSA in PBS and finallyin PBS for 10 minutes. PBS was replaced by the antifading solutioncontaining Hoechst 33258 at a 1 µg/ml final concentration (from a0.02 mg/ml stock solution in distilled water). A drop of the embryosuspension was placed on a slide, a coverslip was deposited on thetop and gently pressed before the preparations were sealed withRubber Cement adhesive.
Microscopy and photographyAll observations were made under a Nikon Labophot-2A equippedwith Nomarski differential interference contrast and epifluorescenceoptics. Photomicrographs were taken with Tri X and T max-100 films(Eastman Kodak Co., NY).
RESULTS
During the present study, chromatin diminution began in mostof the P. univalens embryos used as controls at the thirddivision (4-cell stage) in three blastomeres simultaneously (seeFig. 2E).
LiCl induces the occurrence of chromatindiminution in Parascaris pregerminal cellsThe analysis of Li+ effects on embryos at early successive
2396 M. R. Esteban, G. Giovinazzo and C. Goday
cleavages showed that unlike normal embryos (Fig. 2), in LiCl-treated embryos at the P0 stage, chromatin diminution occurredat the first division and that the first cleavage was symmetrical(Fig. 3A-D). When the embryos were treated immediately afterthe first division, chromatin diminution occurred simultane-ously in both blastomeres (Fig. 3E). In these embryos, incontrast to untreated ones (Fig. 2D), spindle orientation wasidentical for both blastomeres which, in addition, divided syn-chronously (Fig. 3E,F). In this respect, it is of interest to notethat in normal Parascaris embryos the pregerminal blastomereP1 initiates second cleavage before the presomatic S1 blas-tomere, even though it is the S1 cell which terminates thedivision first (Goday et al., 1992) (see Fig. 2C,D). Fig. 3Gshows an example of an embryo treated with LiCl at the 4-cellstage where all blastomeres underwent chromatin diminutionsimultaneously. In these embryos spindle orientation was alsoaltered with respect to controls (not shown). Similarly, inembryos treated during the 8 and 16-cell stages, pregerminalblastomeres underwent chromatin diminution (not shown). In
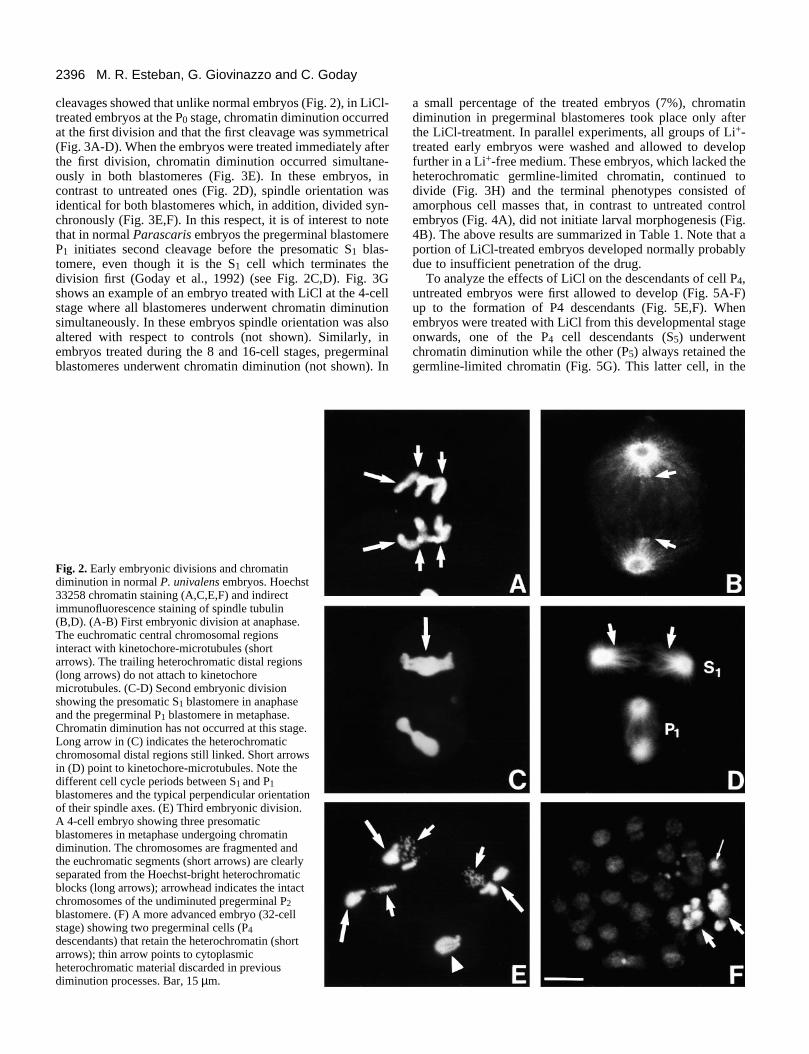
Fig. 2. Early embryonic divisions and chromatindiminution in normal P. univalens embryos. Hoechst33258 chromatin staining (A,C,E,F) and indirectimmunofluorescence staining of spindle tubulin(B,D). (A-B) First embryonic division at anaphase.The euchromatic central chromosomal regionsinteract with kinetochore-microtubules (shortarrows). The trailing heterochromatic distal regions(long arrows) do not attach to kinetochoremicrotubules. (C-D) Second embryonic divisionshowing the presomatic S1 blastomere in anaphaseand the pregerminal P1 blastomere in metaphase.Chromatin diminution has not occurred at this stage.Long arrow in (C) indicates the heterochromaticchromosomal distal regions still linked. Short arrowsin (D) point to kinetochore-microtubules. Note thedifferent cell cycle periods between S1 and P1blastomeres and the typical perpendicular orientationof their spindle axes. (E) Third embryonic division.A 4-cell embryo showing three presomaticblastomeres in metaphase undergoing chromatindiminution. The chromosomes are fragmented andthe euchromatic segments (short arrows) are clearlyseparated from the Hoechst-bright heterochromaticblocks (long arrows); arrowhead indicates the intactchromosomes of the undiminuted pregerminal P2blastomere. (F) A more advanced embryo (32-cellstage) showing two pregerminal cells (P4descendants) that retain the heterochromatin (shortarrows); thin arrow points to cytoplasmicheterochromatic material discarded in previousdiminution processes. Bar, 15 µm.
a small percentage of the treated embryos (7%), chromatindiminution in pregerminal blastomeres took place only afterthe LiCl-treatment. In parallel experiments, all groups of Li+-treated early embryos were washed and allowed to developfurther in a Li+-free medium. These embryos, which lacked theheterochromatic germline-limited chromatin, continued todivide (Fig. 3H) and the terminal phenotypes consisted ofamorphous cell masses that, in contrast to untreated controlembryos (Fig. 4A), did not initiate larval morphogenesis (Fig.4B). The above results are summarized in Table 1. Note that aportion of LiCl-treated embryos developed normally probablydue to insufficient penetration of the drug.
To analyze the effects of LiCl on the descendants of cell P4,untreated embryos were first allowed to develop (Fig. 5A-F)up to the formation of P4 descendants (Fig. 5E,F). Whenembryos were treated with LiCl from this developmental stageonwards, one of the P4 cell descendants (S5) underwentchromatin diminution while the other (P5) always retained thegermline-limited chromatin (Fig. 5G). This latter cell, in the
2397Chromatin diminution and somatic cells in Parascaris
continuous presence of LiCl, divided (Fig. 5H,I), giving rise totwo similar daughter primordial germ cells that could not beinduced either to undergo chromatin diminution or to dividefurther (Fig. 5J,K). These results are summarized in Table 2.Likewise, in LiCl-treated L1 larvae, no induction of chromatindiminution in the pregerminal cells was observed. In view ofthese unexpected results, we undertook a detailed cytologicalanalysis of untreated control P. univalens embryos. A popula-tion of embryos was allowed to develop in control conditionsand samples were fixed and Hoechst 33258 stained every 45minutes (see Materials and Methods). In this way, we verifiedcytologically that in nearly all control embryos (98.4%,
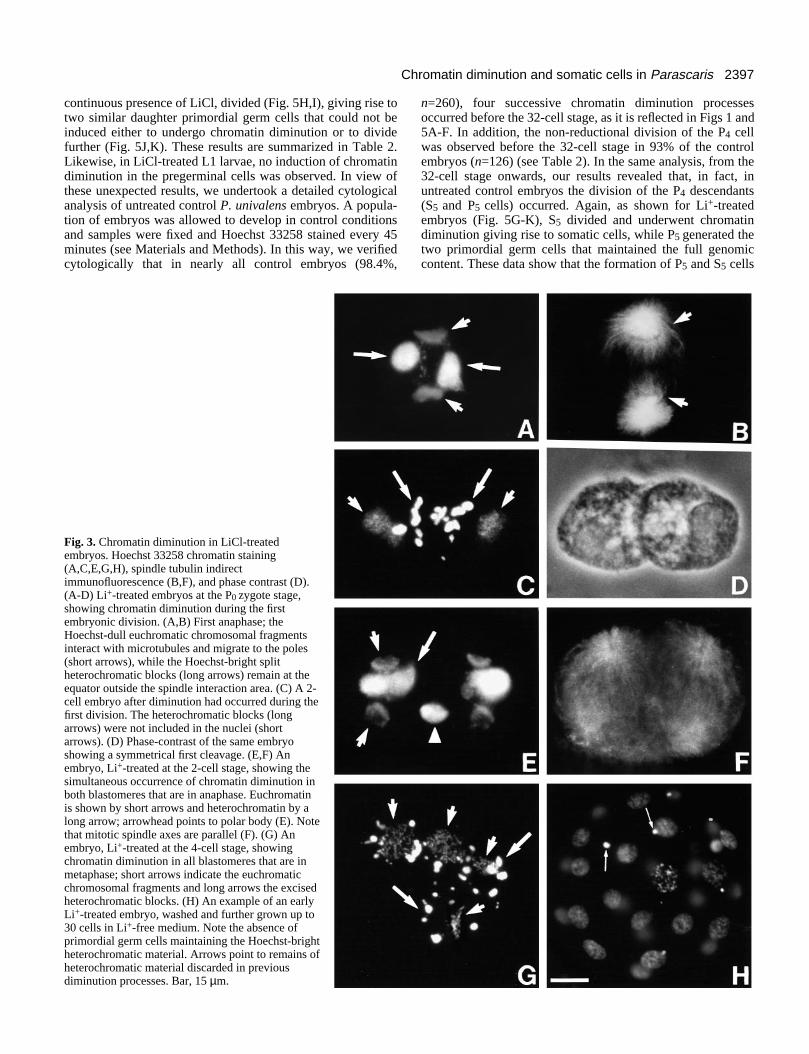
Fig. 3. Chromatin diminution in LiCl-treatedembryos. Hoechst 33258 chromatin staining(A,C,E,G,H), spindle tubulin indirectimmunofluorescence (B,F), and phase contrast (D).(A-D) Li+-treated embryos at the P0 zygote stage,showing chromatin diminution during the firstembryonic division. (A,B) First anaphase; theHoechst-dull euchromatic chromosomal fragmentsinteract with microtubules and migrate to the poles(short arrows), while the Hoechst-bright splitheterochromatic blocks (long arrows) remain at theequator outside the spindle interaction area. (C) A 2-cell embryo after diminution had occurred during thefirst division. The heterochromatic blocks (longarrows) were not included in the nuclei (shortarrows). (D) Phase-contrast of the same embryoshowing a symmetrical first cleavage. (E,F) Anembryo, Li+-treated at the 2-cell stage, showing thesimultaneous occurrence of chromatin diminution inboth blastomeres that are in anaphase. Euchromatinis shown by short arrows and heterochromatin by along arrow; arrowhead points to polar body (E). Notethat mitotic spindle axes are parallel (F). (G) Anembryo, Li+-treated at the 4-cell stage, showingchromatin diminution in all blastomeres that are inmetaphase; short arrows indicate the euchromaticchromosomal fragments and long arrows the excisedheterochromatic blocks. (H) An example of an earlyLi+-treated embryo, washed and further grown up to30 cells in Li+-free medium. Note the absence ofprimordial germ cells maintaining the Hoechst-brightheterochromatic material. Arrows point to remains ofheterochromatic material discarded in previousdiminution processes. Bar, 15 µm.
n=260), four successive chromatin diminution processesoccurred before the 32-cell stage, as it is reflected in Figs 1 and5A-F. In addition, the non-reductional division of the P4 cellwas observed before the 32-cell stage in 93% of the controlembryos (n=126) (see Table 2). In the same analysis, from the32-cell stage onwards, our results revealed that, in fact, inuntreated control embryos the division of the P4 descendants(S5 and P5 cells) occurred. Again, as shown for Li+-treatedembryos (Fig. 5G-K), S5 divided and underwent chromatindiminution giving rise to somatic cells, while P5 generated thetwo primordial germ cells that maintained the full genomiccontent. These data show that the formation of P5 and S5 cells

2398 M. R. Esteban, G. Giovinazzo and C. Goday
Fig. 4. (A) First larval stage of an untreated P.univalens embryo showing the two characteristicprimordial germ cells that maintain the Hoechst-brightheterochromatin (arrows). (B) A LiCl-treatedembryonic terminal phenotype consisting of anamorphous cell mass that does not initiate larvalmorphogenesis. Bars, 15 µm.
Table 1. Early developmental effects of LiCl treatment compared to untreated controls in P. univalens embryosInitiation of LiCl-treatment
Zygotes 2-cell embryos 4 to 16-cell embryos
Phenotypes Untreated Treated Untreated Treated Untreated Treated
Normal* 100/100 29/132 98/100 24/116 99/100 58/238
Abnormal chromatin diminution† 0/100 103/132 2/100 92/116 1/100 180/238+ abnormal division pattern‡
Terminal phenotypeLarval stages 50/50 9/50 49/50 11/50 98/100 33/100Amorphous cell masses§ 0/50 41/50 1/50 39/50 2/100 77/100
*Pregerminal blastomeres divide asymmetrically and only presomatic blastomeres undergo chromatin diminution beginning either at the 2-cell stage or at the4-cell stage.
†Chromatin diminution in all blastomeres of the embryo.‡Somatic-like symmetrical division, cell cycle timing and spindle orientation.§Arrest stage with several hundred cells without having initiated larval morphogenesis.
and their descendants takes place during early development ofthe Parascaris population we studied. As shown in Table 2, nosignificant differences have been observed between LiCl-treated and untreated control embryos. According to Boveri’sobservations (Boveri, 1899) (Fig. 1, inset), P5 division occursat the 82-cell developmental stage. From our observations,both in Li+-treated and untreated control embryos, the divisionof S5 occurs around the 50-cell stage and the division of P5immediately after the 80-cell stage is surpassed.
Fig. 5. Chromatin diminution in untreated normal P. univalensembryos up to the formation of P4 descendants (A-F) anddevelopment of P. univalens embryos treated with LiCl from the 32-cell stage onwards (G-K). (A-K) Hoechst 33258 chromatin staining.(A) Third embryonic division. A 4-cell embryo showing threepresomatic blastomeres undergoing chromatin diminution; S1a andS1b are in telophase and S2 is in metaphase. Arrows in S2 point to theHoechst-bright heterochromatic blocks clearly separated from theeuchromatic segments. The pregerminal blastomere P2 is inmetaphase and retains intact chromosomes. Arrowhead points topolar body. (B) An 8-cell embryo showing two cells (P2descendants) that retain the heterochromatin (thick arrows); thinarrows point to cytoplasmic heterochromatic material discarded inthe previous diminution processes. (C) A 12-cell embryo showing S3and P3 in metaphase. S3 undergoes chromatin diminution; arrowspoint to the separated heterochromatic material. P3 shows intactchromosomes. (D) A 24-cell embryo showing S4 in metaphaseundergoing chromatin diminution while P4 remains in interphase andmaintains the heterochromatin. Arrows point to the heterochromaticblocks separated from the euchromatic fragments of S4 cell. (E) A
NaSCN prevents chromatin diminutionWe have analyzed the effects of NaSCN during early embryo-genesis of P. univalens (see Materials and Methods). In 42%(n=319) of the treated embryos a normal pattern of develop-ment was observed and these embryos were indistinguishablefrom untreated control embryos (n=300); that is, pregerminalblastomeres divided asymmetrically, only presomatic blas-tomeres underwent chromatin diminution and developmentprogressed up to larval stages. However, the other 58% of the
29-cell embryo showing the non-reductional division of P4 cell;arrows point to cytoplasmic heterochromatic material discarded inthe previous diminution process. (F) A 38-cell embryo showing twointerphasic cells (thick arrows) retaining heterochomatin (P4descendants); thin arrow points to heterochromatic materialdiscarded in previous diminution processes. (G) A Hoechst-stainedembryo at the 50-cell stage showing the differential behavior of theP4 cell descendants (S5 and P5). S5 is in metaphase undergoingchromatin diminution while P5 remains in interphase. Arrows pointto the discarded heterochromatic material. (H) A Hoechst-stainedembryo that surpassed the 80-cell stage and shows the equal divisionof P5 that is in metaphase without chromatin diminution. The arrowpoints to heterochromatic material previously discarded. (I) AHoechst-stained embryo that surpassed the 100-cell stage showing P5in anaphase; the chromatids retain their heterochromatin. (J) AHoechst-stained embryo initiating larval morphogenesis showing thetwo interphasic primordial germ cells which originated from the P5division (arrows). (K) A larva immediately after hatching showingthe two last primordial germ cells generated by P5, that still remainin interphase (arrows). Bars, 15 µm.
2399Chromatin diminution and somatic cells in Parascaris
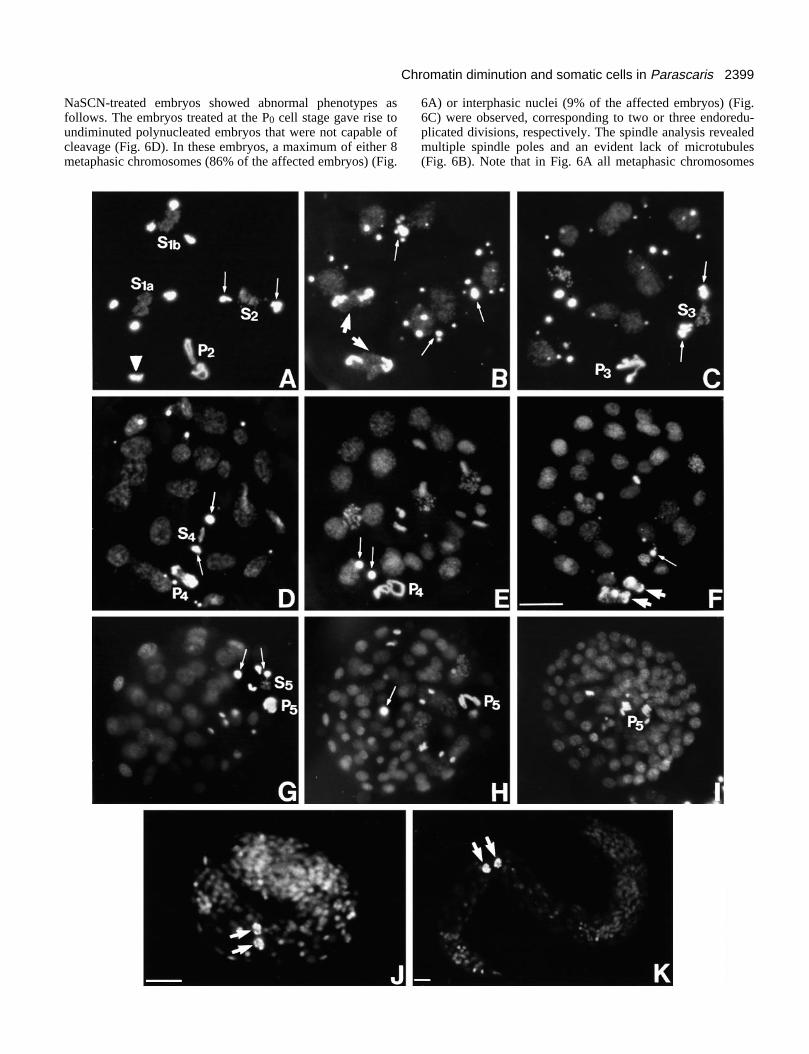
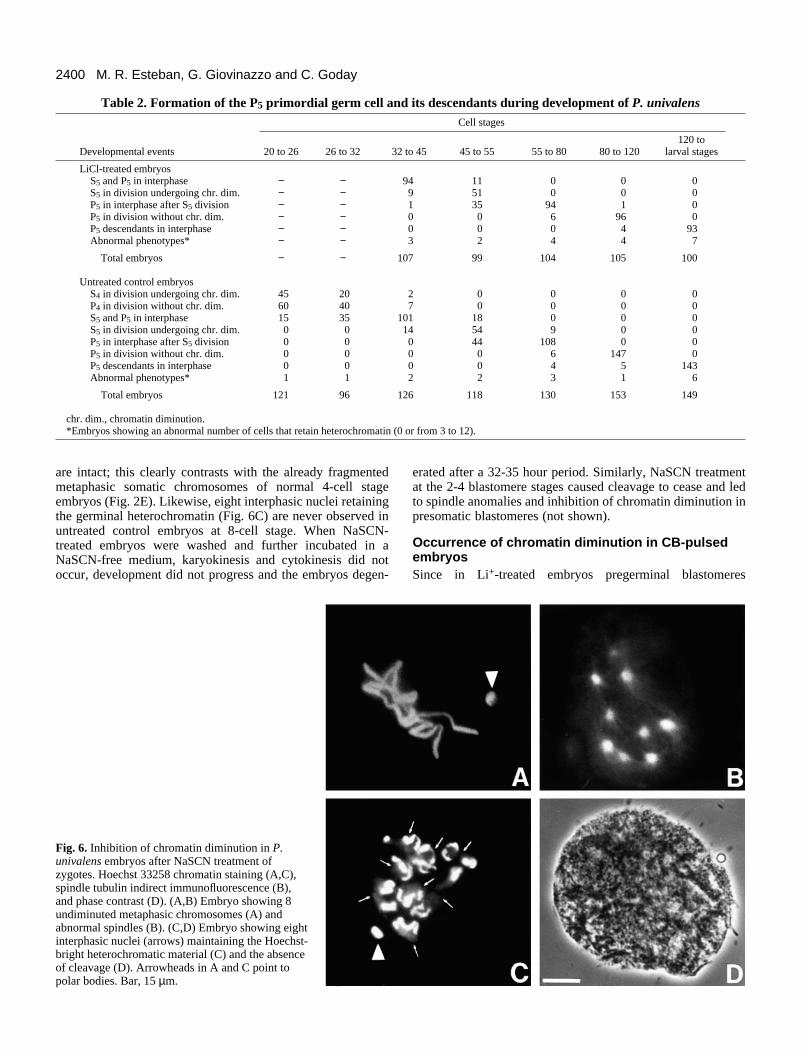
NaSCN-treated embryos showed abnormal phenotypes asfollows. The embryos treated at the P0 cell stage gave rise toundiminuted polynucleated embryos that were not capable ofcleavage (Fig. 6D). In these embryos, a maximum of either 8metaphasic chromosomes (86% of the affected embryos) (Fig.
6A) or interphasic nuclei (9% of the affected embryos) (Fig.6C) were observed, corresponding to two or three endoredu-plicated divisions, respectively. The spindle analysis revealedmultiple spindle poles and an evident lack of microtubules(Fig. 6B). Note that in Fig. 6A all metaphasic chromosomes
2400 M. R. Esteban, G. Giovinazzo and C. Goday
Table 2. Formation of the P5 primordial germ cell and its descendants during development of P. univalensCell stages
120 to Developmental events 20 to 26 26 to 32 32 to 45 45 to 55 55 to 80 80 to 120 larval stages
LiCl-treated embryosS5 and P5 in interphase − − 94 11 0 0 0S5 in division undergoing chr. dim. − − 9 51 0 0 0P5 in interphase after S5 division − − 1 35 94 1 0P5 in division without chr. dim. − − 0 0 6 96 0P5 descendants in interphase − − 0 0 0 4 93Abnormal phenotypes* − − 3 2 4 4 7
Total embryos − − 107 99 104 105 100
Untreated control embryosS4 in division undergoing chr. dim. 45 20 2 0 0 0 0P4 in division without chr. dim. 60 40 7 0 0 0 0S5 and P5 in interphase 15 35 101 18 0 0 0S5 in division undergoing chr. dim. 0 0 14 54 9 0 0P5 in interphase after S5 division 0 0 0 44 108 0 0P5 in division without chr. dim. 0 0 0 0 6 147 0P5 descendants in interphase 0 0 0 0 4 5 143Abnormal phenotypes* 1 1 2 2 3 1 6
Total embryos 121 96 126 118 130 153 149
chr. dim., chromatin diminution.*Embryos showing an abnormal number of cells that retain heterochromatin (0 or from 3 to 12).
are intact; this clearly contrasts with the already fragmentedmetaphasic somatic chromosomes of normal 4-cell stageembryos (Fig. 2E). Likewise, eight interphasic nuclei retainingthe germinal heterochromatin (Fig. 6C) are never observed inuntreated control embryos at 8-cell stage. When NaSCN-treated embryos were washed and further incubated in aNaSCN-free medium, karyokinesis and cytokinesis did notoccur, development did not progress and the embryos degen-
Fig. 6. Inhibition of chromatin diminution in P.univalens embryos after NaSCN treatment ofzygotes. Hoechst 33258 chromatin staining (A,C),spindle tubulin indirect immunofluorescence (B),and phase contrast (D). (A,B) Embryo showing 8undiminuted metaphasic chromosomes (A) andabnormal spindles (B). (C,D) Embryo showing eightinterphasic nuclei (arrows) maintaining the Hoechst-bright heterochromatic material (C) and the absenceof cleavage (D). Arrowheads in A and C point topolar bodies. Bar, 15 µm.
erated after a 32-35 hour period. Similarly, NaSCN treatmentat the 2-4 blastomere stages caused cleavage to cease and ledto spindle anomalies and inhibition of chromatin diminution inpresomatic blastomeres (not shown).
Occurrence of chromatin diminution in CB-pulsedembryosSince in Li+-treated embryos pregerminal blastomeres
2401Chromatin diminution and somatic cells in Parascaris
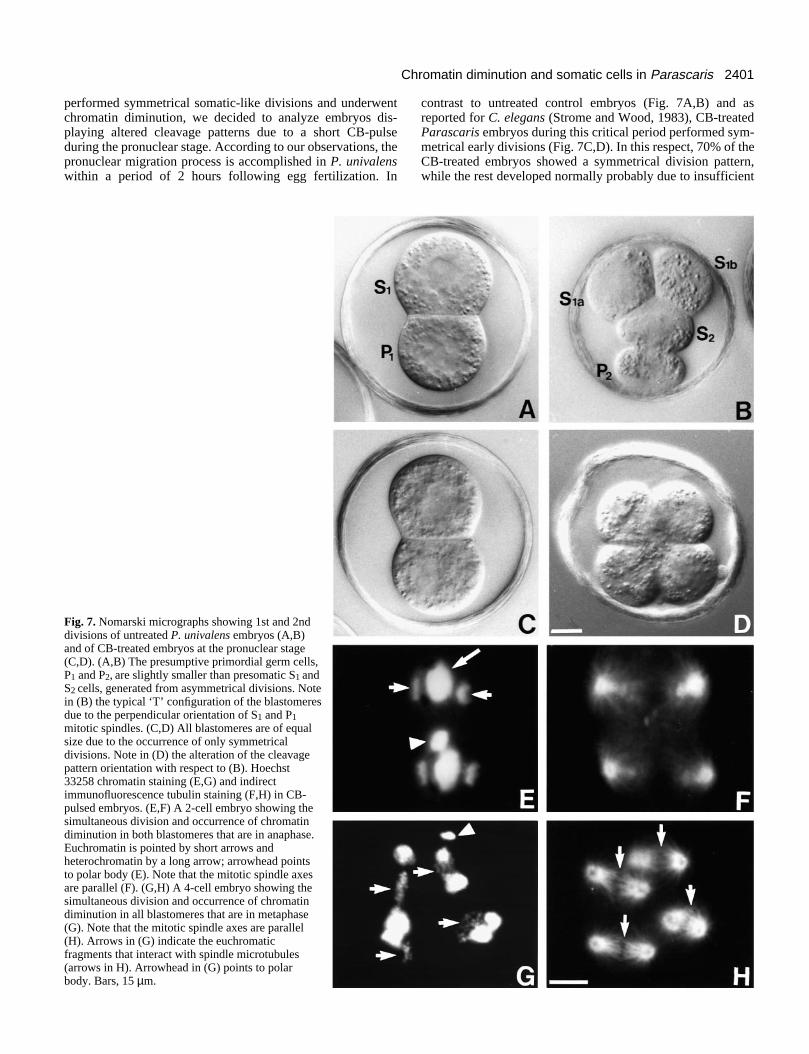
performed symmetrical somatic-like divisions and underwentchromatin diminution, we decided to analyze embryos dis-playing altered cleavage patterns due to a short CB-pulseduring the pronuclear stage. According to our observations, thepronuclear migration process is accomplished in P. univalenswithin a period of 2 hours following egg fertilization. In
Fig. 7. Nomarski micrographs showing 1st and 2nddivisions of untreated P. univalens embryos (A,B)and of CB-treated embryos at the pronuclear stage(C,D). (A,B) The presumptive primordial germ cells,P1 and P2, are slightly smaller than presomatic S1 andS2 cells, generated from asymmetrical divisions. Notein (B) the typical ‘T’ configuration of the blastomeresdue to the perpendicular orientation of S1 and P1mitotic spindles. (C,D) All blastomeres are of equalsize due to the occurrence of only symmetricaldivisions. Note in (D) the alteration of the cleavagepattern orientation with respect to (B). Hoechst33258 chromatin staining (E,G) and indirectimmunofluorescence tubulin staining (F,H) in CB-pulsed embryos. (E,F) A 2-cell embryo showing thesimultaneous division and occurrence of chromatindiminution in both blastomeres that are in anaphase.Euchromatin is pointed by short arrows andheterochromatin by a long arrow; arrowhead pointsto polar body (E). Note that the mitotic spindle axesare parallel (F). (G,H) A 4-cell embryo showing thesimultaneous division and occurrence of chromatindiminution in all blastomeres that are in metaphase(G). Note that the mitotic spindle axes are parallel(H). Arrows in (G) indicate the euchromaticfragments that interact with spindle microtubules(arrows in H). Arrowhead in (G) points to polarbody. Bars, 15 µm.
contrast to untreated control embryos (Fig. 7A,B) and asreported for C. elegans (Strome and Wood, 1983), CB-treatedParascaris embryos during this critical period performed sym-metrical early divisions (Fig. 7C,D). In this respect, 70% of theCB-treated embryos showed a symmetrical division pattern,while the rest developed normally probably due to insufficient
2402 M. R. Esteban, G. Giovinazzo and C. Goday
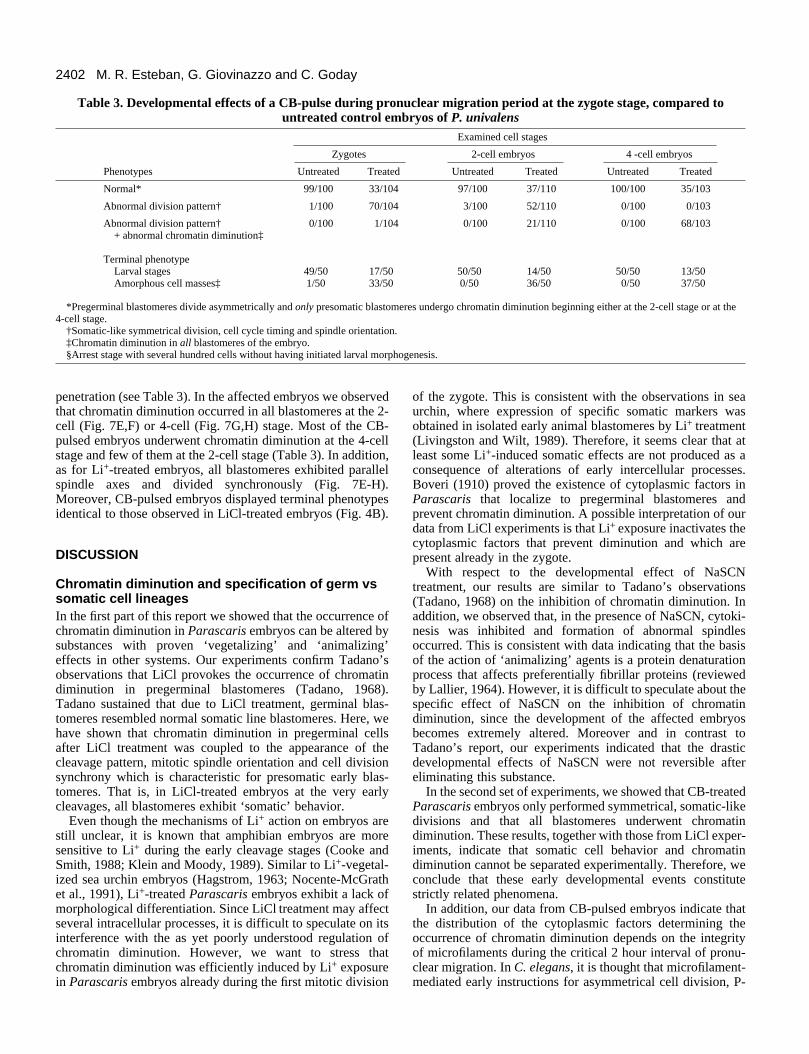
Table 3. Developmental effects of a CB-pulse during pronuclear migration period at the zygote stage, compared tountreated control embryos of P. univalens
Examined cell stages
Zygotes 2-cell embryos 4 -cell embryos
Phenotypes Untreated Treated Untreated Treated Untreated Treated
Normal* 99/100 33/104 97/100 37/110 100/100 35/103
Abnormal division pattern† 1/100 70/104 3/100 52/110 0/100 0/103
Abnormal division pattern† 0/100 1/104 0/100 21/110 0/100 68/103+ abnormal chromatin diminution‡
Terminal phenotypeLarval stages 49/50 17/50 50/50 14/50 50/50 13/50Amorphous cell masses‡ 1/50 33/50 0/50 36/50 0/50 37/50
*Pregerminal blastomeres divide asymmetrically and only presomatic blastomeres undergo chromatin diminution beginning either at the 2-cell stage or at the4-cell stage.
†Somatic-like symmetrical division, cell cycle timing and spindle orientation.‡Chromatin diminution in all blastomeres of the embryo.§Arrest stage with several hundred cells without having initiated larval morphogenesis.
penetration (see Table 3). In the affected embryos we observedthat chromatin diminution occurred in all blastomeres at the 2-cell (Fig. 7E,F) or 4-cell (Fig. 7G,H) stage. Most of the CB-pulsed embryos underwent chromatin diminution at the 4-cellstage and few of them at the 2-cell stage (Table 3). In addition,as for Li+-treated embryos, all blastomeres exhibited parallelspindle axes and divided synchronously (Fig. 7E-H).Moreover, CB-pulsed embryos displayed terminal phenotypesidentical to those observed in LiCl-treated embryos (Fig. 4B).
DISCUSSION
Chromatin diminution and specification of germ vssomatic cell lineagesIn the first part of this report we showed that the occurrence ofchromatin diminution in Parascaris embryos can be altered bysubstances with proven ‘vegetalizing’ and ‘animalizing’effects in other systems. Our experiments confirm Tadano’sobservations that LiCl provokes the occurrence of chromatindiminution in pregerminal blastomeres (Tadano, 1968).Tadano sustained that due to LiCl treatment, germinal blas-tomeres resembled normal somatic line blastomeres. Here, wehave shown that chromatin diminution in pregerminal cellsafter LiCl treatment was coupled to the appearance of thecleavage pattern, mitotic spindle orientation and cell divisionsynchrony which is characteristic for presomatic early blas-tomeres. That is, in LiCl-treated embryos at the very earlycleavages, all blastomeres exhibit ‘somatic’ behavior.
Even though the mechanisms of Li+ action on embryos arestill unclear, it is known that amphibian embryos are moresensitive to Li+ during the early cleavage stages (Cooke andSmith, 1988; Klein and Moody, 1989). Similar to Li+-vegetal-ized sea urchin embryos (Hagstrom, 1963; Nocente-McGrathet al., 1991), Li+-treated Parascaris embryos exhibit a lack ofmorphological differentiation. Since LiCl treatment may affectseveral intracellular processes, it is difficult to speculate on itsinterference with the as yet poorly understood regulation ofchromatin diminution. However, we want to stress thatchromatin diminution was efficiently induced by Li+ exposurein Parascaris embryos already during the first mitotic division
of the zygote. This is consistent with the observations in seaurchin, where expression of specific somatic markers wasobtained in isolated early animal blastomeres by Li+ treatment(Livingston and Wilt, 1989). Therefore, it seems clear that atleast some Li+-induced somatic effects are not produced as aconsequence of alterations of early intercellular processes.Boveri (1910) proved the existence of cytoplasmic factors inParascaris that localize to pregerminal blastomeres andprevent chromatin diminution. A possible interpretation of ourdata from LiCl experiments is that Li+ exposure inactivates thecytoplasmic factors that prevent diminution and which arepresent already in the zygote.
With respect to the developmental effect of NaSCNtreatment, our results are similar to Tadano’s observations(Tadano, 1968) on the inhibition of chromatin diminution. Inaddition, we observed that, in the presence of NaSCN, cytoki-nesis was inhibited and formation of abnormal spindlesoccurred. This is consistent with data indicating that the basisof the action of ‘animalizing’ agents is a protein denaturationprocess that affects preferentially fibrillar proteins (reviewedby Lallier, 1964). However, it is difficult to speculate about thespecific effect of NaSCN on the inhibition of chromatindiminution, since the development of the affected embryosbecomes extremely altered. Moreover and in contrast toTadano’s report, our experiments indicated that the drasticdevelopmental effects of NaSCN were not reversible aftereliminating this substance.
In the second set of experiments, we showed that CB-treatedParascaris embryos only performed symmetrical, somatic-likedivisions and that all blastomeres underwent chromatindiminution. These results, together with those from LiCl exper-iments, indicate that somatic cell behavior and chromatindiminution cannot be separated experimentally. Therefore, weconclude that these early developmental events constitutestrictly related phenomena.
In addition, our data from CB-pulsed embryos indicate thatthe distribution of the cytoplasmic factors determining theoccurrence of chromatin diminution depends on the integrityof microfilaments during the critical 2 hour interval of pronu-clear migration. In C. elegans, it is thought that microfilament-mediated early instructions for asymmetrical cell division, P-

2403Chromatin diminution and somatic cells in Parascaris
granule segregation and duration of cell cycling are segregatedas a unit (Hill and Strome, 1990). Similarly, the factors con-trolling chromatin diminution in Parascaris seem to be parti-tioned in the same group of developmental instructions, alsoresponsible for early pattern formation. In C. elegans, parmutants and CB-treated embryos show similar defects withrespect to cleavage patterns and P granule distribution(Kemphues et al., 1988; Hill and Strome, 1990). This led tothe suggestion that some par genes may encode for actomyosincytoskeleton constituents (Strome, 1993). In both LiCl- andCB-treated Parascaris embryos we have found alterations inthe early division patterns similar to C. elegans CB-treatedembryos and par mutants. In addition, Parascaris terminalphenotypes resemble those of par mutants; that is, they consistof amorphous masses containing large numbers of cells.
At present, antigens similar to P granules found in C.elegans have not been identified in Parascaris pregerminalcells. However, preliminary observations from our laboratoryusing anti-myosin II antibodies on normal Parascaris embryosrevealed a differential distribution of myosin between preso-matic and pregerminal early blastomeres. In addition, in LiClembryos undergoing chromatin diminution in all blastomeres,no differences in myosin distribution between the cells wereobserved.
It is becoming evident that mRNA localization constitutesan effective way to establish the asymmetric distribution ofearly cytoplasmic determinants. Recently, in C. elegans aputative germ-line RNA helicase has been identified and con-sidered to represent a possible P granule component (Roussellet al., 1994). Studies in Drosophila and Xenopus embryossupport the idea that the specific localization of mRNAs con-stitutes a mechanism to establish gradients of proteins thatdetermine cell fate in early development (reviewed by Wilhelmand Vale, 1993). Microtubules and motor proteins are candi-dates for mRNA transport in different systems (Yisraeli et al.,1990; Pokrywka and Stephenson, 1991; Ainger et al., 1993).In addition, increasing evidence suggests that actin and myosinparticipate in the translocation of some RNAs (Sundell andSinger, 1991). Furthermore, cytoskeletal proteins, in particularactin, are thought to be responsible for the anchoring oflocalized mRNAs (Yisraeli et al., 1990). From our results it isalso conceivable that microfilament-mediated mRNA transportand/or anchoring may determine the localization of the cyto-plasmic factors involved in chromatin diminution during earlydevelopment in Parascaris.
On the formation of a P5 primordial germ cell and itsdescendantsBoveri (1899) described the formation of the P5 primordialgerm cell during Parascaris development and hypothesizedthat this additional division may occur at least occasionally incertain embryos (Boveri, 1910). However, the classical studiesby him and other authors supported the suggestion that P4actually represented the primordial germ cell (zur Strassen,1896; Müller, 1903; Boveri, 1910). Therefore, the questionconcerning the formation of a P5 primordial germ cellremained unsolved. Our cytological analysis of untreatedcontrol P. univalens embryos from the 32-cell stage onwards(see Table 2), confirms the formation of a P5 primordial germcell and its descendants (Fig. 1, inset) and strongly suggeststhat it constitutes a regular event of P. univalens early devel-
opment, at least in the population we examined. As shown, S5divides before P5 and undergoes chromatin diminution, whileP5 divides equally and without chromatin diminution and at alater developmental stage. Thus, the P4 cell represents a pre-sumptive primordial germ cell and only P5 descendants retainthe germline-limited heterochromatin and constitute the twolast primordial germ cells.
Our data concerning the formation of P0 to P4 cells are inaccordance with the generally assumed schematic representa-tion of the cell lineage and segregation of germ line andsomatic cells in Parascaris shown in Fig. 1 (for revision seeTobler, 1986). However, other studied Parascaris populationsexhibited a different timing of cleavage (zur Strassen, 1896;Boveri, 1899; Müller, 1903). In a more recent paper, thedivision of P4 is reported at the 160-cell stage (Seidl et al.,1988). The discrepancy among the published data may reflectheterochronical variations among different populations of P.univalens. In fact, there are also variations concerning thebeginning of the chromatin diminution process (second or thirddivision) depending on the population studied.
The formation of a P5 primordial germ cell has also beenreported in the free-living nematode Anguillula aceti (Pai,1928). According to Pai, the S5 blastomere is responsible forthe formation of gonadal somatic tissues while P5 representsthe first and definitively determinated germinal cell. However,in most nematodes, including C. elegans, the P4 cell is con-sidered to be the primordial germ cell and the somatic gonadis derived from S2 (Sulston et al., 1983).
We think it is of interest to point out that in P. univalenschromatin diminution could be Li+-induced in pregerminal P0-P4 cells but not in P5 and its descendants. This differentialresponse strongly suggests that, in contrast to early P0-P4pregerminal blastomeres, the germ quality and fate of P5 isstrictly determined and cannot be reversed or altered at thisspecific developmental stage. Our results may also reflect theexistence of different mechanisms that control germline deter-mination between the very early embryonic divisions and con-secutive cleavages during Parascaris development.
We thank Dr P. Ripoll, Dr L. Sánchez and Dr S. Pimpinelli forhelpful suggestions and comments on the manuscript and A. Hurtadofor the artwork. We acknowledge the support of DGICYT (grantPB90-0073).
REFERENCES
Ainger, K., Avossa, F., Morgan, F., Hill, S. J., Barry, E., Barbarese, E. andCarson, J. H. (1993). Transport and localization of exogenous myelin basicprotein mRNA microinjected into oligodendrocytes. J. Cell Biol. 123, 431-441.
Berridge, M. J., Downes, C. P. and Hanley, M. R. (1982). Lithium amplifiesagonist-dependent phosphatidylinositol responses in brain and salivaryglands. Biochem. J. 206, 587-595.
Berridge, M. J., Downes, C. P. and Hanley, M. R. (1989). Neural anddevelopmental actions of lithium: a unifying hypothesis. Cell 59, 411-419.
Boveri, T. (1887). Über Differenzierung der Zellkerne während der Furchungdes Eies von Ascaris megalocephala. Anat. Anz. 2, 688-693.
Boveri, T. (1899). Die Entwicklung von Ascaris megalocephala mitbesonderer Rücksicht auf die Kernverhältnisse. In Festschrift für C. vonKupffer, pp. 383-430. Fischer, Jena.
Boveri, T. (1910). Die Potenzen der Ascaris-Blastomeren bei abgeänderterFurchung. Zugleich ein Beitrag zur Frage qualitativ-ungleicher Chromosomen-Teilung. In Festschr für R. Hertwig III, pp. 131-214. Fischer, Jena.

2404 M. R. Esteban, G. Giovinazzo and C. Goday
Brenner, S. (1974). The genetics of Caenorhabditis elegans. Genetics 77, 71-94.
Cooke, J. and Smith, E. J. (1988). The restrictive effect of early exposure tolithium upon body pattern in Xenopus development, studied by quantitativeanatomy and immunofluorescence. Development 102, 85-99.
Etter, A., Aboutanos, M., Tobler, H. and Müller, F. (1991). Eliminatedchromatin of Ascaris contains a gene that encodes a putative ribosomalprotein. Proc. Nat. Acad. Sci. USA 88, 1593-1596.
Goday, C. and Pimpinelli, S. (1984). Chromosome organization andheterochromatin elimination in Parascaris. Science 224, 411-413.
Goday, C., Gonzalez-García, J. M., Esteban, M. R., Giovinazzo, G. andPimpinelli, S. (1992). Kinetochores and chromatin diminution in earlyembryos of Parascaris univalens. J. Cell Biol. 118, 23-32.
Goday, C. and Pimpinelli, S. (1993). The occurrence, role and evolution ofchromatin diminution in nematodes. Parasitol. Today 9, 319-322.
Hagstrom, B. E. (1963). The effect of lithium and o-iodobenzoic acid on theearly development of the sea urchin egg. Biol. Bull. 124, 55-64.
Herbst, C. (1892). Experimentelle Untersuchungen über den Einfluss derveränderten chemischen Zusammensetzung des umgebenden Mediums aufdie Entwicklung der Tiere, I Teil. Versuche an Seeigeleiern. Z. Wiss. Zool.55, 446-518.
Hill, D. P. and Strome, S. (1990). Brief cytochalasin-induced disruption ofmicrofilaments during a critical interval in 1-cell C. elegans embryos altersthe partitioning of developmental instructions to the 2-cell embryo.Development 108, 159-172.
Hyman, A. A. and White, J. G. (1987). Determination of cell division axes inthe early embryogenesis of C. elegans. J. Cell Biol. 105, 2123-2135.
Hyman, A. A. (1989). Centrosome movement in the early divisions ofCaenorhabditis elegans: A cortical site determining centrosome position. J.Cell Biol. 109, 1185-1193.
Johnson, G. D. and de Nogueira Araujo, G. M. (1981). A simple method ofreducig the fading immunofluorescence during microscopy. J. Immunol.Meth. 43, 349-350.
Kao, K. R., Masui, Y. and Elinson, R. P. (1986). Lithium-inducedrespecification of pattern in Xenopus laevis embryos. Nature 322, 371-373.
Kao, K. R. and Elinson, R. P. (1988). The entire mesodermal mantle behavesas Spemann’s organizer in dorsoanterior enhanced Xenopus laevis embryos.Dev. Biol. 127, 64-77.
Kemphues, K. J., Priess, J. R., Morton, D. G. and Cheng, N. (1988).Identification of genes required for cytoplasmic localization in early C.elegans embryos. Cell 52, 311-320.
King, R. L. and Beams, H. W. (1938). An experimental study of chromatindiminution in Ascaris. J. Exp. Zool. 77, 425-443.
Kirby, C., Kusch, M. and Kemphues, K. (1990). Mutations in the par genesof Caenorhabditis elegans affect cytoplasmic reorganization during the firstcell cycle. Dev. Biol. 142, 203-215.
Klein, S. L. and Moody, S. A. (1989). Lithium changes the ectodermal fate ofindividual frog blastomeres because it causes ectopic neural plate formation.Development 106, 599-610.
Kühn, A. (1971). Lectures on Developmental Physiology, pp. 166-199.Springer-Verlag, New York.
Lallier, R. (1964). Biochemical aspects of animalization and vegetalization inthe sea urchin embryo. Advan. Morphogen. 3, 147-196.
Livingston, B. T. and Wilt, F. H. (1989). Lithium evokes expression of vegetalspecific molecules in the animal blastomeres of sea urchin embryos. Proc.Nat. Acad. Sci. USA 86, 3669-3673.
Morton, D. G., Roos, J. M. and Kemphues, K. J. (1992). par-4 gene requiredfor cytoplasmic localization and determination of specific cell types inCaenorhabditis elegans embryogenesis. Genetics 130, 771-790.
Müller, H. (1903). Beiträge zur Embryonalentwicklung der Ascarismegalocephala. Zoologica 41, 30.
Nocente-McGrath, C., McIsaac, R. and Ernst, S. G. (1991). Altered cell fatein LiCl-treated sea urchin embryos. Dev. Biol. 147, 445-450.
Pai, S. (1928). Die Phasen des Lebenscyclus der Anguillula aceti. Ehrbg.Zietschr. wiss. Zool. 131, 293-344.
Pokrywka, N. J. and Stephenson, E. C. (1991). Microtubules mediate thelocalization of bicoid RNA during Drosophila oogenesis. Development 113,55-66.
Roussell, D., Gruidl, M. and Bennett, K. (1994). Germ-line determination inCaenorhabditis and Ascaris: Will a helicase begin to unravel the mystery?Parasitol. Today 10, 110-113.
Seidl, C., Bauer, M. and Moritz, K. B. (1988). Chromatin diminution andearly cleavage in Parascaris univalens (Nematoda). Roux’s Arch. Dev. Biol.197, 307-320.
Sherman, W. R., Leavitt, A. L., Honchar, M. P., Hallcher, L. M. andPhillips, B. E. (1981). Evidence that lithium alters phosphoinositidemetabolism: Chronic administration elevates primary D-myo-inositol-1-phosphate in cerebral cortex of the rat. J. Neurochem. 36, 1947-1951.
Spicher, A., Etter, A., Bernard, V. Tobler, H. and Müller, F. (1994).Extremely stable transcripts may compensate for the elimination of the genefert-1 from all Ascaris lumbricoides somatic cells. Dev. Biol. 164, 72-86.
Strome, S. and Wood, W. (1982). Immunofluorescence visualization of germ-line-specific cytoplasmic granules in embryos, larvae, and adults ofCaenorhabditis elegans. Proc. Nat. Acad. Sci. USA 79, 1558-1562.
Strome, S. and Wood, W. B. (1983). Generation of asymmetry andsegregation of germ-line granules in early C.elegans embryos. Cell 35, 15-25.
Strome, S. (1993). Determination of cleavage planes. Cell 72, 3-6.Sulston, J. Schierenberg, E., White, J. and Thomson, N. (1983). The
embryonic cell lineages of the nematode Caenorhabditis elegans. Dev. Biol.100, 67-119.
Sundell, C. L. and Singer, R. H. (1991). Requirement of microfilaments insorting of actin messenger RNA. Science 253, 1275-1277.
Tadano, M. (1968). Nemathelminthes. In Invertebrate Embryology (ed. M.Kumé and K. Dan), pp. 159-191. Nolit, Belgrad.
Tobler, H., Etter, A. and Müller, F. (1992). Chromatin diminution innematode development. Trends Genet. 8, 427-432.
von Ubisch, L. (1929). Über die Determination der larvalen Organe under derImaginalanage bei Seeigeln. Wilhelm Roux Arch. Entw. mech. Org. 117, 81-122.
Wilhelm, J. E. and Vale, R. D. (1993). RNA on the move: The mRNAlocalization pathway. J. Cell Biol. 123, 269-274.
Yisraeli, J. K., Sokol, S. and Melton, D. A. (1990). A two-step model for thelocalization of maternal mRNA in Xenopus oocytes: involvement ofmicrotubules and microfilaments in the translocation and anchoring of Vg1nRNA. Development 108, 289-298.
zur Strassen, O. (1896). Embryonalentwicklung der Ascaris megalocephala.Arch. Entwicklungsmech. 3, 27-105, 133-190.
(Received 29 July 1994 - Accepted 7 March 1995)
![Long Noncoding RNAs, Chromatin, and Developmentdownloads.hindawi.com/journals/tswj/2010/180798.pdf · active chromatin modifications and a more open chromatin conformation[26,39,40,41,42]](https://img.pdfslide.us/doc/110x75/5f8885d811957319d07a36bf/long-noncoding-rnas-chromatin-and-active-chromatin-modifications-and-a-more-open.jpg)




























