Embed Size (px)
Citation preview

Vol. 169, No. 4
Cyst Production and Brown Pigment Formation in Aging Cultures ofAzospirillum brasilense ATCC 29145t
LAKSHMI SADASIVAN* AND CARLOS A. NEYRA
Department of Biochemistry and Microbiology, New Jersey Agricultural Experiment Station, Cook College, RutgersUniversity, New Brunswick, New Jersey 08903
Received 2 October 1986/Accepted 22 December 1986
Encystation in Azospinillum brasilense ATCC 29145 was observed by using routine laboratory staining andphase-contrast and electron microscopy. Encystment occurred in liquid and in solid or semisolid mediacontaining fructose (8 mM) and KNO3 (0.5 mM). The encysted forms consisted of a central body filled withpoly-p-hydroxybutyric acid granules, an electron-transparent intinelike region, and a thick outer layer.Enlarged giant encysted forms with multiple central bodies were also observed during the germination of adesiccated brown colony. Morphogenetically different forms in an aging culture could be resolved by sucrosedensity gradient centrifugation. The dense encysted forms along with numerous granules in a fibrillar networkpelleted at 70% sucrose, while empty saclike envelopes along with vegetative cells and coccoid bodies peiletedat 55% sucrose. Different media induced various degrees of pigmentation in A. brasilense ATCC 29145 afteraging. The pigment possessed several of the properties reported for microbial melanins, including insolubilityin water and organic solvents, solubility in cold and hot alkali, and bleaching in hydrogen peroxide. The UVabsorption maxima of the alkali extract were at 280 and 310 nm. Electron micrographs of the brown pigmentshowed that it occurred as aggregated granules surrounding the encysting celis as well as being excreted intothe medium in an aging culture. It is concluded that A. brasilense ATCC 29145 produces compounds that forma brown pigment similar to melanin and are expressed under the influence of certain cultural conditionsconducive for encystment.
Azospirillum brasilense is known to fix nitrogen in culturemedia and in association with the roots of several grasses (3,23, 28, 33, 50). This microorganism has been used in fieldinoculation studies over the last decade with relative successby many workers (19, 20, 31, 42, 45), and crop yieldincreases as a result of inoculation are also well documented(3, 10, 40, 42, 48). Nonetheless, the response to inoculationby treated plants is known to be highly unpredictable (1, 50).It is generally assumed that bacteria added to therhizosphere will survive, multiply, and remain metabolicallyactive to support nitrogen fixation. Survival of the bacteria israrely monitored after inoculation in the field. These bacteriaare known to be highly pleomorphic and to change theirmetabolic activities swiftly in the face of changing environ-mental conditions (5, 15, 37). In the soil these may includethe availability of carbon and nitrogen sources, moisturecontent, and oxygen tension. Resistance to desiccation inthese bacteria is achieved by encystation, a phenomenon inwhich the vegetative cells undergo certain morphologicaland biochemical changes to form resistant resting cysts,which enables them to survive deleterious physical condi-tions, particularly desiccation (38, 39). Cyst formation inAzobacter spp. has been well documented by morphoge-netic, ultrastructural, and biochemical studies (17, 35, 43, 44,51, 52).
In Azospirillum spp., cysts are cells which have undergonemorphological changes as observed under phase-contrastand scanning electron microscopy and found to be resistantto desiccation from a few hours to 1 month (24, 32). We havepresented some morphological and ultrastructural details of
* Corresponding author.t New Jersey Agricultural Experiment Station publication no.
D-01204-02-86.
the encysting cells from a desiccated floc which was inducedin a liquid culture medium containing low levels of fructose(8 mM) as the carbon source and KNO3 (0.5 mM) as thenitrogen source (37). Under these conditions, the vegetativecells lost motility, assumed an enlarged spherical form, andaccumulated abundant poly-,-hydroxybutyric acid (PHB)granules and developed an outer undifferentiated layer (coat)of polysaccharides. We proposed that the phenomenon wasrelated to encystation. In an attempt to recognize the cul-tural conditions for complete encystment as defined forAzobacter spp., we grew Azospirillum brasilense ATCC29145 cells in the same medium containing fructose andnitrate but on solid agar. Growth under these culturalconditions produced brown-black pigmentation in a 1-weekperiod, which intensified as the culture aged. Pigmentationthus far reported in azospirilla has been primarily related tothe synthesis of carotenoids, which have been shown toprotect the nitrogenase enzyme of A. brasilense and alsoother strains from oxidative damages (15, 29).The objectives of this paper are to define the culture
conditions under which A. brasilense ATCC 29145 differen-tiates into cysts, to compare and present evidence that thePHB-rich encysted cells confer a survival advantage to A.brasilense ATCC 29145, and to report pigment (brownpigment) formation in azospirilla as a result of aging andpartially characterize the pigment.(A preliminary report of this work has been presented [L.
Sadasivan and C. A. Neyra, in W. Klingmuller, ed., Proc.Third Bayreuth Azospirillum Workshop, p. 230, 1985].)
MATERIALS AND METHODS
Bacterial strain and culture conditions. A. brasilense Sp7(ATCC 29145) was used for the encystation and pigment
1670
JOURNAL OF BACTERIOLOGY, Apr. 1987, p. 1670-16770021-9193/87/041670-08$02.00/0Copyright © 1987, American Society for Microbiology
on June 11, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

CYST PRODUCTION AND PIGMENT FORMATION IN A. BRASILENSE
formation studies. The bacterium was routinely subculturedbiweekly on nutrient agar (Difco Laboratories, Detroit,Mich.) slants and maintained at 30°C.Minimal salts medium (MSM) as described previously (37)
was used as the basic medium to induce encystation, withmodifications to incorporate different carbon sources. Mal-ate, fructose (at a concentration of 8 mM), and n-butanol (at0.3% concentration) were added individually as carbonsources. Three different conditions were used: liquid,solid, and semisolid media (0.175% agar [Difco]). In liquidand solid media, the nitrogen source was 0.5 mMKNO3.A log-phase culture from nutrient broth (NB; Difco) was
harvested by centrifugation at 5,000 rpm at 4°C and washedthree times in 100 mM KPO4 buffer (pH 6.8) to give an OD60of 0.8 when read in a spectrophotometer (model 240;Gilford). From this suspension, a loopful of bacteria wasstreaked on solid agar plates, and 0.1 ml was inoculated into100 ml of semisolid nitrogen-free basal (NFb) medium (37)and 10 ml into 100 ml of liquid medium. The solid plates andthe flasks with semisolid medium were kept stationary, whilethe liquid medium cultures were continuously shaken at 200rpm in a gyrotory shaker (New Brunswick Scientific Co.,Inc., New Brunswick, N.J.).
All the cultures were incubated at 37°C. The culturesinoculated in NFb semisolid medium were incubated in twobatches, one under light and the other in dark to see anyeffect of light on pigment formation.
Phase-contrast and electron microscopy of encysted cells. A.brasilense cells grown in three conditions (solid, semisolid,and liquid) were desiccated at room temperature (28 ± 2°C)and observed for more than 6 months. The desiccatingcolonies from solid agar plates and pellicular growth fromsemisolid medium were periodically observed under a phase-contrast microscope either unstained or stained as describedby Vela and Wyss (52). After desiccation, they were scrapedand inoculated into a fresh NFb semisolid medium contain-ing fructose (8 mM) as the carbon source, incubated at 35°Cfor 24 to 48 h, and allowed to germinate. The newlydeveloping growth from the inoculum which was observed tocontain various encysted forms with single and multiplecental bodies was detached and photographed under a Nikonphase-contrast microscope.
Ultrastructural studies. The desiccated growth from allthree conditions was fixed in 1% glutaraldehyde-0.2 Mcacodylate buffer, pH 7.4, followed by 1% osmium tetroxidefixation for 10 h, and dehydrated in a series of graded ethylalcohol solutions after staining in uranyl acetate or ruthe-nium red by the procedures of Cole and Popkin (11) andCagle and Vela (9). The stained preparations were finallyembedded in Spurr plastic as described by Cole and Popkin(11). Ultrathin sections were cut on an ultramicrotome andobserved under a transmission electron microscope(Siemens model 1A Elmscope).
Separation of various encysted forms by sucrose densitygradients. Gradients of sucrose in water (55, 60, 65, and 70%,wt/wt) were prepared, and each concentration was layeredcarefully on top of the others with 2 ml of 70% sucrose at thebottom of a polypropylene tube. The desiccated pelliculargrowth from semisolid medium was removed, suspended in100 mM KPO4 buffer (pH 6.8), and layered on top of the 55%sucrose gradient. The tubes were then centrifuged at 20°Cfor 4 h at 40,000 rpm in an ultracentrifuge (Beckman modelL5-75B). The three bands resolved were individually with-drawn in a 1-ml hypodermic syringe from outside the poly-propylene tube and then observed under the transmission
electron microscope after negative staining with uranylacetate.Thermal resistance experiments. One milliliter of a suspen-
sion of cells from (i) 24-h-old NB-grown, highly motilevegetative cells lacking PHB granules, (ii) 24-h-old MSMliquid medium (supplemented with 8 mM malate and 5 mMNH4Cl)-grown cells containing mixed populations of highlymotile vibrioid and nonmotile round forms with PHB gran-ules, and (iii) 1-month-old MSM semisolid nitrogen-freemedium-grown cells having developed a brown pigment inthe pellicle were serially diluted in sterile distilled water.Dilutions of 105 and 106 of each of the above cultures wereimmersed in a series of water baths adjusted to temperaturesranging from 30 to 60°C. After 20 min, the tubes wereremoved and rapidly cooled before plating 100-pul samples ontryptic soy agar (TSA; Difco).
Desiccation experiments. One milliliter of a suspension ofthe three different types of cells described above was seriallydiluted in sterile distilled water. One hundred microlitersfrom dilutions of 104 and 105 were placed in sterile 1.5-mlEppendorf microfuge tubes, and the tubes were in turn keptopen in a sterile petri dish. The petri dishes with the tubeswere then placed in an incubator at 37°C. At selectedintervals, the dried cells from the microfuge tubes werewashed with 100 ptl of sterile distilled water with vigorousagitation to remove them quantitatively, and their viabilitywas determined by plating on TSA.
Pigment induction studies and characterization of brownpigment. Cells from a 24-h-old NB-grown culture werewashed twice in sterile saline, and then a loopful wasinoculated into various solid and semisolid agar media suchas nutrient agar and MSM containing malate or fructose asthe carbon source. The media were also supplemented withvarious amino acids, such as 0.1% tyrosine, 0.1% phenylal-anine, 0.1% cysteine, and 0.1% tyrosine plus cysteine, whichare known to be the precursors of brown pigment (melanin)formation.
Induction of pigment formation on TSA plates wasachieved as follows. One loopful of cells was inoculated into100 ml of NB medium in 250-ml flasks and incubated at 37°Cfor 1 to 4 weeks under constant shaking at 200 rpm on a
gyrotory shaker (New Brunswick Scientific). Every week,100-pu samples were withdrawn from the aging cultures andplated on TSA. The plates were then incubated at 30°C for 7days. Colonies forming brown pigments were counted andcompared with non-pigment-forming colonies.The solubility of the pigment formed in an NFb semisolid
medium was tested in water (cold and hot), alcohol, acetone,benzene, ether, 1 N HCI, and 1 N NaOH or KOH by themethod of Lingappa et al. (25).The absorption spectrum of the brown pigment soluble in
cold and hot alkali was done by the method of Ivins andHolmes (18) with a Beckman DU 40 recording spectropho-tometer on a range of wavelengths from 200 to 400 nm. Thespectrum obtained was compared with spectra obtainedfrom other brown pigments formed as a result of auto-oxidation of phenolic compounds (3,4-dihydroxyphenyl-alanine, catechol, and melanin).
RESULTS
A. brasilense ATCC 29145 cells grown in semisolid nitro-gen-free malate medium were found to produce brownpigment in the aging pellicles. In nutrient agar plates, pig-mentation was rarely seen, but when the aging cultures fromNB were plated on TSA and incubated for 1 week or more,
1671VOL. 169, 1987
on June 11, 2018 by guesthttp://jb.asm
.org/D
ownloaded from
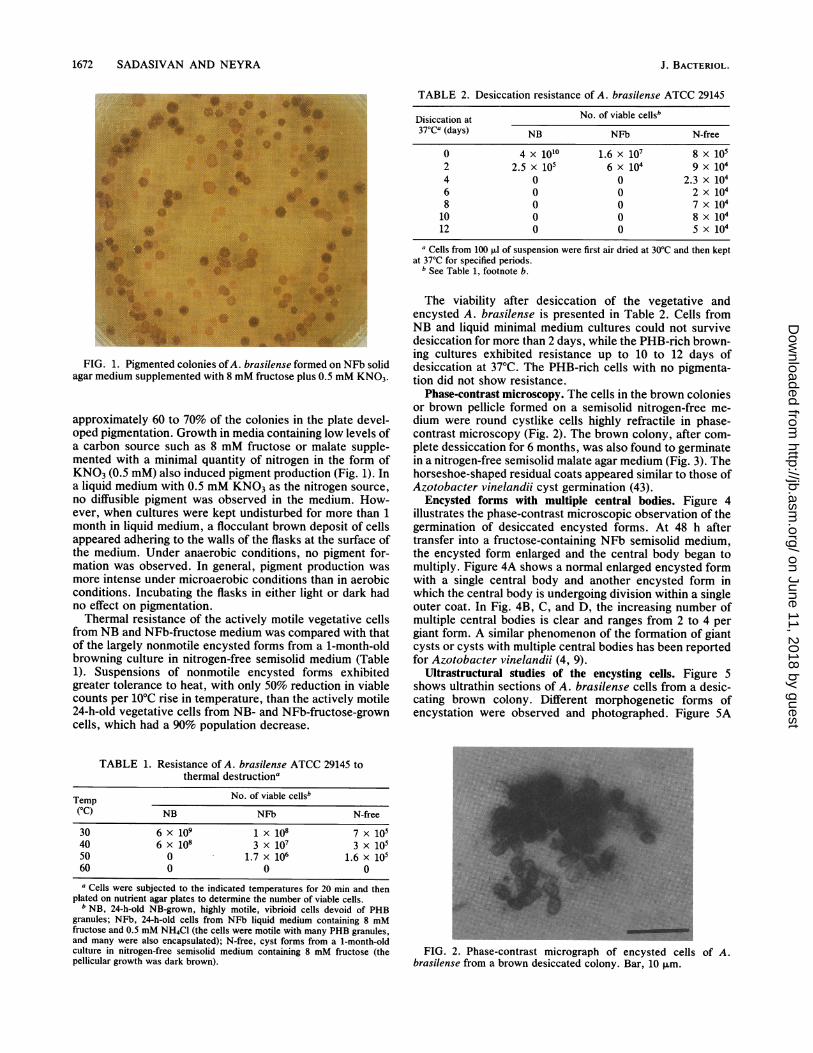
1672 SADASIVAN AND NEYRA
FIG. 1. Pigmented colonies ofA. brasilense formed on NFb solidagar medium supplemented with 8 mM fructose plus 0.5 mM KNO3.
approximately 60 to 70% of the colonies in the plate devel-oped pigmentation. Growth in media containing low levels ofa carbon source such as 8 mM fructose or malate supple-mented with a minimal quantity of nitrogen in the form ofKNO3 (0.5 mM) also induced pigment production (Fig. 1). Ina liquid medium with 0.5 mM KNO3 as the nitrogen source,no diffusible pigment was observed in the medium. How-ever, when cultures were kept undisturbed for more than 1month in liquid medium, a flocculant brown deposit of cellsappeared adhering to the walls of the flasks at the surface ofthe medium. Under anaerobic conditions, no pigment for-mation was observed. In general, pigment production wasmore intense under microaerobic conditions than in aerobicconditions. Incubating the flasks in either light or dark hadno effect on pigmentation.Thermal resistance of the actively motile vegetative cells
from NB and NFb-fructose medium was compared with thatof the largely nonmotile encysted forms from a 1-month-oldbrowning culture in nitrogen-free semisolid medium (Table1). Suspensions of nonmotile encysted forms exhibitedgreater tolerance to heat, with only 50% reduction in viablecounts per 10°C rise in temperature, than the actively motile24-h-old vegetative cells from NB- and NFb-fructose-growncells, which had a 90% population decrease.
TABLE 1. Resistance of A. brasilense ATCC 29145 tothermal destructiona
Temp No. of viable cellsb(OC) NB NFb N-free
30 6 x 109 1 x 108 7 x 1040 6 x 108 3 x 107 3 x 10550 0 1.7 x 106 1.6 x 10560 0 0 0a Cells were subjected to the indicated temperatures for 20 min and then
plated on nutrient agar plates to determine the number of viable cells.b NB, 24-h-old NB-grown, highly motile, vibrioid cells devoid of PHB
granules; NFb, 24-h-old cells from NFb liquid medium containing 8 mMfructose and 0.5 mM NH4Cl (the cells were motile with many PHB granules,and many were also encapsulated); N-free, cyst forms from a 1-month-oldculture in nitrogen-free semisolid medium containing 8 mM fructose (thepellicular growth was dark brown).
TABLE 2. Desiccation resistance of A. brasilense ATCC 29145
Disiccation at No. of viable cellsb37'Ca (days) NB NFb N-free
0 4 x 1010 1.6 x 107 8 x 1052 2.5 x 105 6 x 104 9 x 1044 0 0 2.3x 1046 0 0 2x1048 0 0 7x10410 0 0 8 x 10412 0 0 5 x 104
a Cells from 100 ,ul of suspension were first air dried at 30'C and then keptat 37°C for specified periods.
b See Table 1, footnote b.
The viability after desiccation of the vegetative andencysted A. brasilense is presented in Table 2. Cells fromNB and liquid minimal medium cultures could not survivedesiccation for more than 2 days, while the PHB-rich brown-ing cultures exhibited resistance up to 10 to 12 days ofdesiccation at 37°C. The PHB-rich cells with no pigmenta-tion did not show resistance.
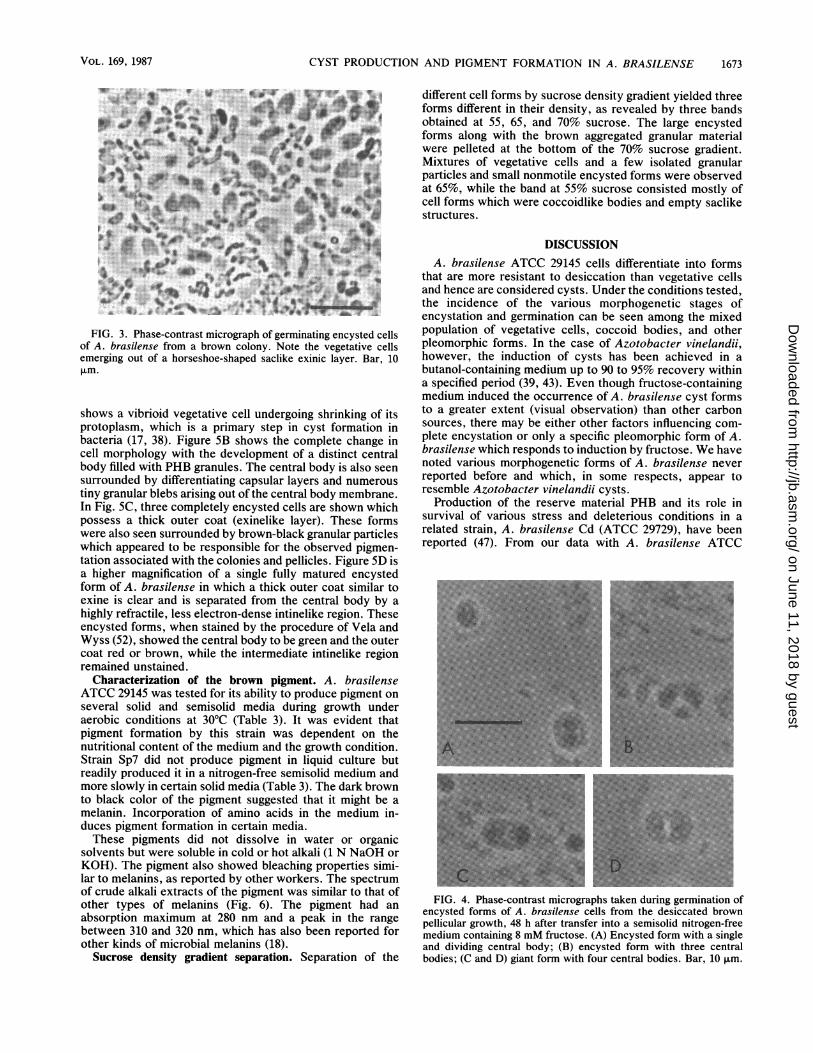
Phase-contrast microscopy. The cells in the brown coloniesor brown pellicle formed on a semisolid nitrogen-free me-dium were round cystlike cells highly refractile in phase-contrast microscopy (Fig. 2). The brown colony, after com-plete dessiccation for 6 months, was also found to germinatein a nitrogen-free semisolid malate agar medium (Fig. 3). Thehorseshoe-shaped residual coats appeared similar to those ofAzotobacter vinelandii cyst germination (43).
Encysted forms with multiple central bodies. Figure 4illustrates the phase-contrast microscopic observation of thegermination of desiccated encysted forms. At 48 h aftertransfer into a fructose-containing NFb semisolid medium,the encysted form enlarged and the central body began tomultiply. Figure 4A shows a normal enlarged encysted formwith a single central body and another encysted form inwhich the central body is undergoing division within a singleouter coat. In Fig. 4B, C, and D, the increasing number ofmultiple central bodies is clear and ranges from 2 to 4 pergiant form. A similar phenomenon of the formation of giantcysts or cysts with multiple central bodies has been reportedfor Azotobacter vinelandii (4, 9).
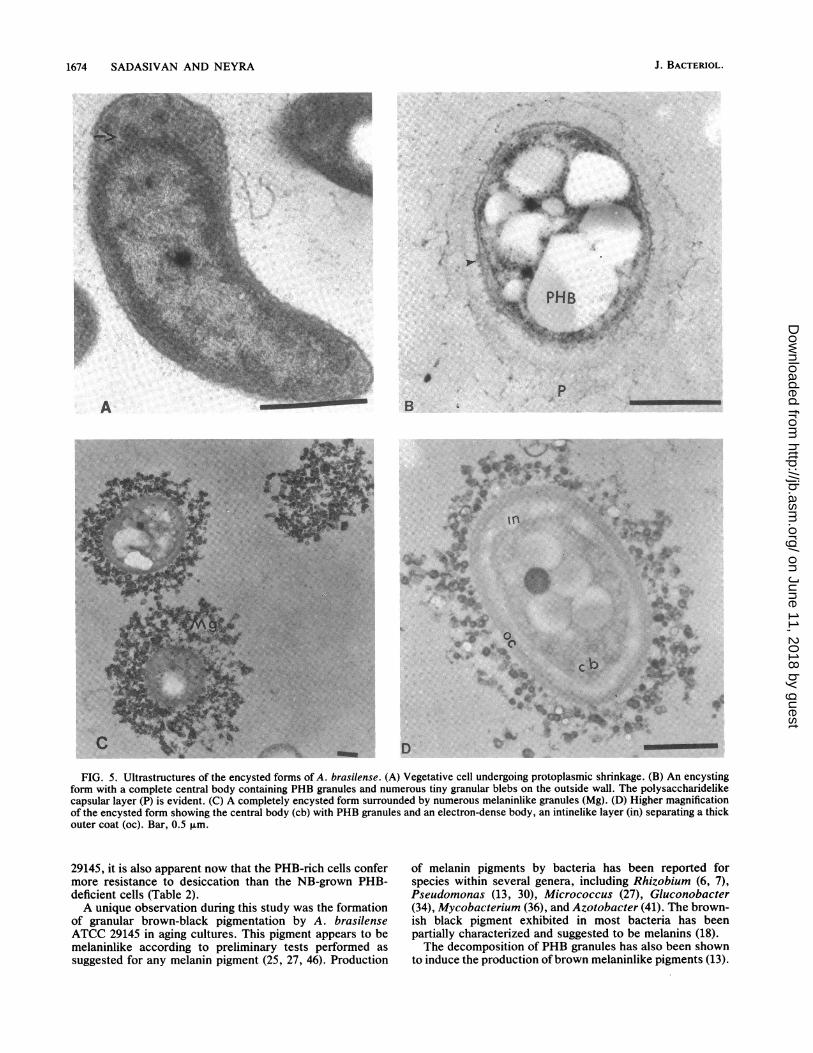
Ultrastructural studies of the encysting cells. Figure 5shows ultrathin sections of A. brasilense cells from a desic-cating brown colony. Different morphogenetic forms ofencystation were observed and photographed. Figure 5A
FIG. 2. Phase-contrast micrograph of encysted cells of A.brasilense from a brown desiccated colony. Bar, 10 ,um.
J. BACTERIOL.
on June 11, 2018 by guesthttp://jb.asm
.org/D
ownloaded from
CYST PRODUCTION AND PIGMENT FORMATION IN A. BRASILENSE
.:U*tv^ Wt
i"-aN
4ir" vFIG. 3. Phase-contrast micrograph of germinating encysted cells
of A. brasilense from a brown colony. Note the vegetative cellsemerging out of a horseshoe-shaped saclike exinic layer. Bar, 10Lm.
shows a vibrioid vegetative cell undergoing shrinking of itsprotoplasm, which is a primary step in cyst formation inbacteria (17, 38). Figure SB shows the complete change incell morphology with the development of a distinct centralbody filled with PHB granules. The central body is also seensurrounded by differentiating capsular layers and numeroustiny granular blebs arising out of the central body membrane.In Fig. SC, three completely encysted cells are shown whichpossess a thick outer coat (exinelike layer). These formswere also seen surrounded by brown-black granular particleswhich appeared to be responsible for the observed pigmen-tation associated with the colonies and pellicles. Figure SD isa higher magnification of a single fully matured encystedform of A. brasilense in which a thick outer coat similar toexine is clear and is separated from the central body by ahighly refractile, less electron-dense intinelike region. Theseencysted forms, when stained by the procedure of Vela andWyss (52), showed the central body to be green and the outercoat red or brown, while the intermediate intinelike regionremained unstained.
Characterization of the brown pigment. A. brasilenseATCC 29145 was tested for its ability to produce pigment onseveral solid and semisolid media during growth underaerobic conditions at 30°C (Table 3). It was evident thatpigment formation by this strain was dependent on thenutritional content of the medium and the growth condition.Strain Sp7 did not produce pigment in liquid culture butreadily produced it in a nitrogen-free semisolid medium andmore slowly in certain solid media (Table 3). The dark brownto black color of the pigment suggested that it might be amelanin. Incorporation of amino acids in the medium in-duces pigment formation in certain media.These pigments did not dissolve in water or organic
solvents but were soluble in cold or hot alkali (1 N NaOH orKOH). The pigment also showed bleaching properties simi-lar to melanins, as reported by other workers. The spectrumof crude alkali extracts of the pigment was similar to that ofother types of melanins (Fig. 6). The pigment had anabsorption maximum at 280 nm and a peak in the rangebetween 310 and 320 nm, which has also been reported forother kinds of microbial melanins (18).
Sucrose density gradient separation. Separation of the
different cell forms by sucrose density gradient yielded threeforms different in their density, as revealed by three bandsobtained at 55, 65, and 70% sucrose. The large encystedforms along with the brown aggregated granular materialwere pelleted at the bottom of the 70% sucrose gradient.Mixtures of vegetative cells and a few isolated granularparticles and small nonmotile encysted forms were observedat 65%, while the band at 55% sucrose consisted mostly ofcell forms which were coccoidlike bodies and empty saclikestructures.
DISCUSSIONA. brasilense ATCC 29145 cells differentiate into forms
that are more resistant to desiccation than vegetative cellsand hence are considered cysts. Under the conditions tested,the incidence of the various morphogenetic stages ofencystation and germination can be seen among the mixedpopulation of vegetative cells, coccoid bodies, and otherpleomorphic forms. In the case of Azotobacter vinelandii,however, the induction of cysts has been achieved in abutanol-containing medium up to 90 to 95% recovery withina specified period (39, 43). Even though fructose-containingmedium induced the occurrence of A. brasilense cyst formsto a greater extent (visual observation) than other carbonsources, there may be either other factors influencing com-plete encystation or only a specific pleomorphic form of A.brasilense which responds to induction by fructose. We havenoted various morphogenetic forms of A. brasilense neverreported before and which, in some respects, appear toresemble Azotobacter vinelandii cysts.
Production of the reserve material PHB and its role insurvival of various stress and deleterious conditions in arelated strain, A. brasilense Cd (ATCC 29729), have beenreported (47). From our data with A. brasilense ATCC
I
FIG. 4. Phase-contrast micrographs taken during germination ofencysted forms of A. brasilense cells from the desiccated brownpellicular growth, 48 h after transfer into a semisolid nitrogen-freemedium containing 8 mM fructose. (A) Encysted form with a singleand dividing central body; (B) encysted form with three centralbodies; (C and D) giant form with four central bodies. Bar, 10 ,um.
VOL. 169, 1987 1673
on June 11, 2018 by guesthttp://jb.asm
.org/D
ownloaded from
1674 SADASIVAN AND NEYRA
p
4.
#'40-0 4L;9
FIG. 5. Ultrastructures of the encysted forms of A. brasilense. (A) Vegetative cell undergoing protoplasmic shrinkage. (B) An encystingform with a complete central body containing PHB granules and numerous tiny granular blebs on the outside wall. The polysaccharidelikecapsular layer (P) is evident. (C) A completely encysted form surrounded by numerous melaninlike granules (Mg). (D) Higher magnificationof the encysted form showing the central body (cb) with PHB granules and an electron-dense body, an intinelike layer (in) separating a thickouter coat (oc). Bar, 0.5 F±m.
29145, it is also apparent now that the PHB-rich cells confermore resistance to desiccation than the NB-grown PHB-deficient cells (Table 2).A unique observation during this study was the formation
of granular brown-black pigmentation by A. brasilenseATCC 29145 in aging cultures. This pigment appears to bemelaninlike according to preliminary tests performed as
suggested for any melanin pigment (25, 27, 46). Production
of melanin pigments by bacteria has been reported forspecies within several genera, including Rhizobium (6, 7),Pseudomonas (13, 30), Micrococcus (27), Gluconobacter(34), Mycobacterium (36), and Azotobacter (41). The brown-ish black pigment exhibited in most bacteria has beenpartially characterized and suggested to be melanins (18).The decomposition of PHB granules has also been shown
to induce the production of brown melaninlike pigments (13).
Dr #
I.
B
*I
;t4
D
J. BACTERIOL.
m_ --
t.
on June 11, 2018 by guesthttp://jb.asm
.org/D
ownloaded from
CYST PRODUCTION AND PIGMENT FORMATION IN A. BRASILENSE
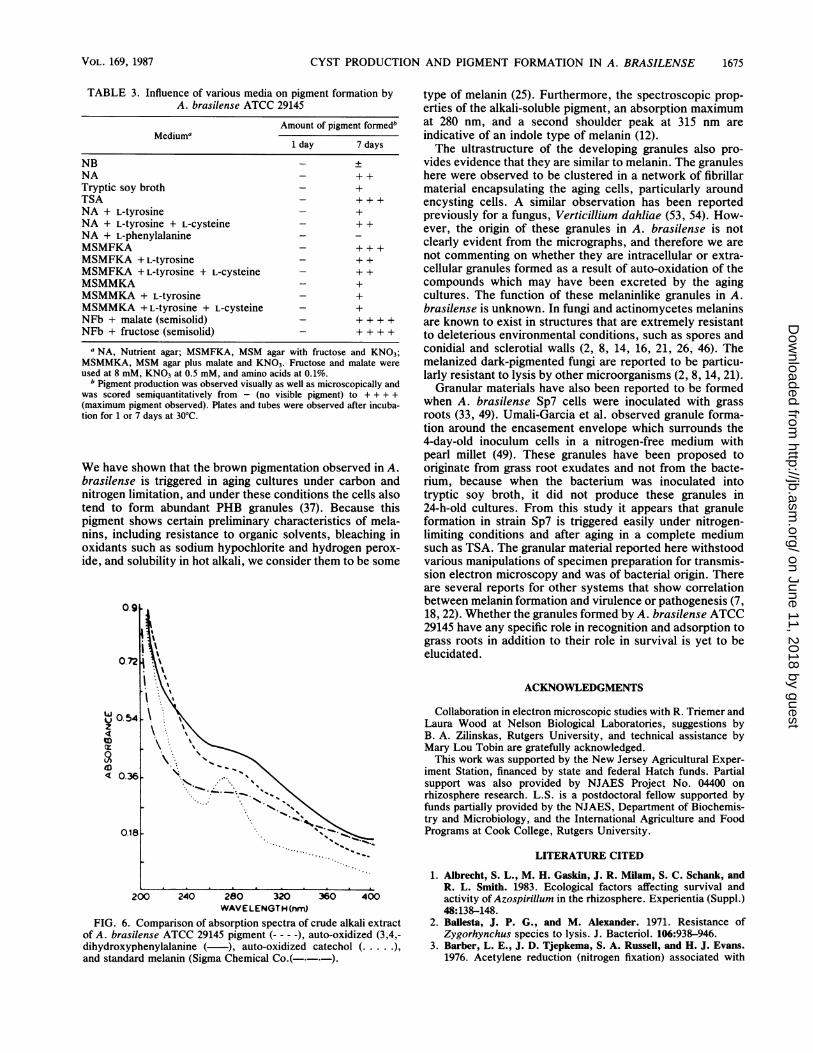
TABLE 3. Influence of various media on pigment formation byA. brasilense ATCC 29145
Amount of pigment formedbMediuma
1 day 7 days
NB - +NA - + +Tryptic soy broth - +TSA - + + +NA + L-tyrosine - +NA + L-tyrosine + L-cysteine - + +NA + L-phenylalanine -
MSMFKA - +++MSMFKA + L-tyrosine - + +MSMFKA + L-tyrosine + L-cysteine - + +MSMMKA - +MSMMKA + L-tyrosine - +MSMMKA +L-tyrosine + L-Cysteine - +NFb + malate (semisolid) - + + + +NFb + fructose (semisolid) - + + + +
a NA, Nutrient agar; MSMFKA, MSM agar with fructose and KNO3;MSMMKA, MSM agar plus malate and KNO3. Fructose and malate wereused at 8 mM, KNO3 at 0.5 mM, and amino acids at 0.1%.
b Pigment production was observed visually as well as microscopically andwas scored semiquantitatively from - (no visible pigment) to + + + +(maximum pigment observed). Plates and tubes were observed after incuba-tion for 1 or 7 days at 30°C.
We have shown that the brown pigmentation observed in A.brasilense is triggered in aging cultures under carbon andnitrogen limitation, and under these conditions the cells alsotend to form abundant PHB granules (37). Because thispigment shows certain preliminary characteristics of mela-nins, including resistance to organic solvents, bleaching inoxidants such as sodium hypochlorite and hydrogen perox-ide, and solubility in hot alkali, we consider them to be some
0.9
0.72
0.54z
4
ED
0.9
on
U 0 54
tD
ID
t;! \',\! \'sI 'X'M\ \
*\.*-. ''\
< 0.^ -'-X... s._vsO.tE ... .vs
0.181
200 240 280 320 360 400WAVELENGTH (nm)
FIG. 6. Comparison of absorption spectra of crude alkali extractof A. brasilense ATCC 29145 pigment (-- - -), auto-oxidized (3,4,-dihydroxyphenylalanine ( ), auto-oxidized catechol (..... ),and standard melanin (Sigma Chemical Co.(-.-.-).
type of melanin (25). Furthermore, the spectroscopic prop-erties of the alkali-soluble pigment, an absorption maximumat 280 nm, and a second shoulder peak at 315 nm areindicative of an indole type of melanin (12).The ultrastructure of the developing granules also pro-
vides evidence that they are similar to melanin. The granuleshere were observed to be clustered in a network of fibrillarmaterial encapsulating the aging cells, particularly aroundencysting cells. A similar observation has been reportedpreviously for a fungus, Verticillium dahliae (53, 54). How-ever, the origin of these granules in A. brasilense is notclearly evident from the micrographs, and therefore we arenot commenting on whether they are intracellular or extra-cellular granules formed as a result of auto-oxidation of thecompounds which may have been excreted by the agingcultures. The function of these melaninlike granules in A.brasilense is unknown. In fungi and actinomycetes melaninsare known to exist in structures that are extremely resistantto deleterious environmental conditions, such as spores andconidial and sclerotial walls (2, 8, 14, 16, 21, 26, 46). Themelanized dark-pigmented fungi are reported to be particu-larly resistant to lysis by other microorganisms (2, 8, 14, 21).
Granular materials have also been reported to be formedwhen A. brasilense Sp7 cells were inoculated with grassroots (33, 49). Umali-Garcia et al. observed granule forma-tion around the encasement envelope which surrounds the4-day-old inoculum cells in a nitrogen-free medium withpearl millet (49). These granules have been proposed tooriginate from grass root exudates and not from the bacte-rium, because when the bacterium was inoculated intotryptic soy broth, it did not produce these granules in24-h-old cultures. From this study it appears that granuleformation in strain Sp7 is triggered easily under nitrogen-limiting conditions and after aging in a complete mediumsuch as TSA. The granular material reported here withstoodvarious manipulations of specimen preparation for transmis-sion electron microscopy and was of bacterial origin. Thereare several reports for other systems that show correlationbetween melanin formation and virulence or pathogenesis (7,18, 22). Whether the granules formed by A. brasilense ATCC29145 have any specific role in recognition and adsorption tograss roots in addition to their role in survival is yet to beelucidated.
ACKNOWLEDGMENTS
Collaboration in electron microscopic studies with R. Triemer andLaura Wood at Nelson Biological Laboratories, suggestions byB. A. Zilinskas, Rutgers University, and technical assistance byMary Lou Tobin are gratefully acknowledged.
This work was supported by the New Jersey Agricultural Exper-iment Station, financed by state and federal Hatch funds. Partialsupport was also provided by NJAES Project No. 04400 onrhizosphere research. L.S. is a postdoctoral fellow supported byfunds partially provided by the NJAES, Department of Biochemis-try and Microbiology, and the International Agriculture and FoodPrograms at Cook College, Rutgers University.
LITERATURE CITED
1. Albrecht, S. L., M. H. Gaskin, J. R. Milam, S. C. Schank, andR. L. Smith. 1983. Ecological factors affecting survival andactivity of Azospirillum in the rhizosphere. Experientia (Suppl.)48:138-148.
2. BalHesta, J. P. G., and M. Alexander. 1971. Resistance ofZygorhynchus species to lysis. J. Bacteriol. 106:938-946.
3. Barber, L. E., J. D. Tjepkema, S. A. Russell, and H. J. Evans.1976. Acetylene reduction (nitrogen fixation) associated with
VOL. 169, 1987 1675
ri.
on June 11, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

1676 SADASIVAN AND NEYRA
corn inoculated with Spirillum. Appl. Environ. Microbiol.32:108-113.
4. Beamen, B. L., L. E. Jackson, and D. M. Shankel. 1968.Formation of multiple central bodies in giant cysts ofAzotobacter vinelandii. J. Bacteriol. 96:266-269.
5. Berg, R. M., M. E. Tyler, N. J. Novick, V. Vasil, and I. K. Vasil.1980. Biology of Azospirillum-sugarcane association: enhance-ment of nitrogenase activity. Appl. Environ. Microbiol.39:642-649.
6. Beringer, J. E., and P. E. Hirsch. 1984. Genetic adaptation tothe environment: the role of plasmids in microbial ecology, p.
63-70. In M. J. Klug and C. A. Reddy (ed.), Current perspec-
tives in microbial ecology. American Society for Microbiology,Washington, D.C.
7. Beynon, J. L., J. E. Beringer, and A. W. B. Johnston. 1980.Plasmids and host range in Rhizobium leguminosarum andRhizobium phaseoli. J. Gen. Microbiol. 120:421-429.
8. Bloomfield, B. J., and M. Alexander. 1967. Melanins and resist-ance of fungi to lysis. J. Bacteriol. 93:1276-1280.
9. Cagle, G. D., and G. R. Vela. 1971. Giant cysts and cysts withmultiple central bodies in Azotobacter vinelandii. J. Bacteriol.107:315-319.
10. Cohen, E., Y. Okon, J. Kigel, I. Nur, and Y. Henis. 1980.Increase in dry weight and total nitrogen content in Zea mays
and Setaria italica associated with nitrogen fixing Azospirillumspp. Plant Physiol. 66:746-749.
11. Cole, R. M., and T. J. Popkin. 1981. Electron microscopy, p.
34-51. In P. Gerhardt, R. G. E. Murray, R. N. Costilow, E. W.Nester, W. A. Hood, N. R. Kreig, and G. B. Phillips (ed.),Manual of methods for general bacteriology. American Societyfor Microbiology, Washington, D.C.
12. Das, K. C., M. B. Abramson, and R. Katzman. 1978. Neuronalpigments: spectroscopic characterization of human brain mela-nin. J. Neurochem. 30:601-605.
13. Delafield, I. P., M. Doudoroff, N. J. Palleroni, C. J. Lusty, andR. Contopoulos. 1965. Decomposition of poly-,-hydroxybut-yrate by Pseudomonas. J. Bacteriol. 90:1455-1466.
14. Ellis, D. H., and D. A. Griffiths. 1974. The location and analysisof melanins in the cell walls of some soil fungi. Can. J.Microbiol. 20:1379-1386.
15. Eskew, D. C., D. D. Focht, and I. P. Ting. 1977. Nitrogenfixation, denitrification, and pleomorphic growth in a highlypigmented Spirillum lipoferum. Appl. Environ. Microbiol. 34:582-585.
16. Griffith, D. A. 1970. The fine structure of developing microscle-rotia of Verticillium dahliae Kleb. Arch. Mikrobiol. 74:207-212.
17. Hitchinson, V. M., and H. L. Sadoff. 1970. Morphogenesis ofcysts in Azotobacter vinelandii. J. Bacteriol. 104:492-498.
18. Ivins, B. E., and R. K. Holmes. 1980. Isolation and character-ization of melanin-producing (mel) mutants of Vibrio cholerae.Infect. Immun. 27:721-729.
19. Kapulnik, Y., J. Kigel, Y. Okon, I. Nur, and Y. Henis. 1981.Effect of Azospirillum inoculation on some growth parametersand N-content of wheat, sorghum and panicum. Plant Soil61:65-70.
20. Kapulnik, Y., S. Sarig, I. Nur, Y. Okon, J. Kigel, and Y. Hennis.1981. -Yield increases in summer cereal crops in Israeli fieldsinoculated with Azospirillum. Exp. Agric. 17:179-187.
21. Kuo, M. J., and M. Alexander. 1967. Inhibition of the lysis offungi by melanins. J. Bacteriol. 94:624-629.
22. Kuon-drung, K. J., and J. C. Rhodes. 1986. Encapsulation andmelanin formation as indicators of virulence in Cryptococcusneoformans. Infect. Immun. 51:218-223.
23. Lakshmi, V., A. Satyanarayana Rao, K. ViJayalakshmi, M.
Lakshmi Kumari, K. V. B. R. Tilak, and N. S. Subba Rao. 1977.Establishment and survival of Spirillum lipoferum. Proc. IndianAcad. Sci. Sec. B 86:397-404.
24. Lamm, R. B., and C. A. Neyra. 1981. Characterization and cyst
production of azospirilla isolated from selected grasses growingin New Jersey and New York. Can. J. Microbiol. 27:1320-1325.
25. Lingappa, Y., A. S. Sussman, and I. A. Bernstein. 1963. Effect of
light and media upon growth and melanin formation inAureobasidium pullans (De By) Am (Pullularia pullulans).Mycopathol. Mycol. Appl. 20:109-128.
26. Mencher, J. R., and A. M. Heim. 1962. Melanin biosynthesis byStreptomyces lavendulae. J. Gen. Microbiol. 28:665-670.
27. Mung, T. H., and L. M. Kondrat'eva. 1981. A new melaninsynthesizing species Micrococcus mellanogenerans. Mikrobio-logiya 50:122-127.
28. Neyra, C. A., and J. Dobereiner. 1977. Nitrogen fixation ingrasses. Adv. Agron. 29:1-38.
29. Nur, I., Y. Steinitz, Y. Okon, and Y. Henis. 1981. Carotenoidcomposition and function in nitrogen-fixing bacteria of the genusAzospirillum. J. Gen. Microbiol. 122:27-32.
30. Ogunnariuo, J., and J. M. Hamilton-Miller. 1975. Brown andred pigmented Pseudomonas aeroginosa. Differentiation be-tween melanin and pyrorubrin. J. Med. Microbiol. 8:199-203.
31. O'Hara, G. W., M. R. Davey, and J. A. Lucas. 1981. Effect ofinoculation of Zea mays with Azospirillum brasilense strainsunder temperate conditions. Can. J. Microbiol. 27:871-877.
32. Papen, M., and D. Werner. 1982. Organic acid utilization,succinate excretion, encystation and oscillating nitrogenaseactivity in Azospirillum brasilense under microaerobic condi-tions. Arch. Microbiol. 132:57-61.
33. Patriquin, D. G., J. Dobereiner, and D. K. Jain. 1983. Sites andprocesses of the association between diazotrophs and grasses.Can. J. Microbiol. 29:900-916.
34. Pavelenko, G. V., M. S. Loistsyanskaya, and N. I. Nemirons-kaya. 1981. Melanin pigment of Gluconobacter oxydans. Mikro-biologiya 50:718-722.
35. Pope, L. M., and 0. Wyss. 1970. Outer layers of Azotobactervinelandii cyst. J. Bacteriol. 102:234-239.
36. Prabhakaran, K., W. F. Kirchheimer, and E. B. Harris. 1968.Oxidation of phenolic compounds by Mycobacterium leprae:structure-activity relationships of substrates and comparisonwith other copper proteins and enzymes. Microbiology5:273-281.
37. Sadasivan, L., and C. A. Neyra. 1985. Flocculation in Azospiril-lum brasilense and Azospirillum lipoferum: exopolysaccharidesand cyst formation. J. Bacteriol. 163:716-723.
38. Sadoff, H. L. 1973. Comparative aspects of morphogenesis inthree prokaryotic genera. Annu. Rev. Microbiol. 27:133-153.
39. Sadoff, H. L. 1975. Encystment and germination in Azotobactervinelandii. Bacteriol. Rev. 39:516-539.
40. Schank, S. C., R. L. Smith, and G. C. Weiser. 1979. Response oftwo pearl millet grown in vitro after inoculation with Azospiril-lum brasilense. Proc. Soil Crop Sci. Soc. 39:112-115.
41. Sen, M., and P. Sen. 1965. Interspecific transformation inAzotobacter. J. Gen. Microbiol. 41:1-6.
42. Smith, R. L., J. M. Bouton, S. C. Schank, K. M. Quesenberry,M. E. Tyler, J. R. Milam, M. M. Gaskins, and R. C. Littell. 1976.Nitrogen fixation in grasses inoculated with Spirillum lipoferum.Science 193:1003-1005.
43. Socolofsky, M. D., and 0. Wyss. 1961. Cysts of Azotobacter. J.Bacteriol. 81:946-954.
44. Stevenson, L. H., and M. D. Socolofsky. 1972. Encystment ofAzotobacter vinelandii in liquid culture. Antonie van Leeu-wenhoek J. Microbiol. Serol. 38:605-610.
45. Subba Rao, N. S., K. V. B. R. Tilak, M. Lakshmi Kumari, andC. S. Singh. 1979. Azospirillum-a new bacterial fertilizer fortropical crops. Sci. Reporter 10:690-692.
46. Sussman, A. S., Y. Lingappa, and I. A. Bernstein. 1963. Effect oflight and media upon growth and melanin formation inCladosporium mansoni. Mycopathol. Mycol. Appl. 20:307-314.
47. Tal, S., and Y. Okon. 1985. Production of the reserve materialpoly-,-hydroxybutyrate and its function in Azospirillumbrasilense Cd. Can. J. Microbiol. 31:608-613.
48. Tien, T. M., M. H. Gaskin, and D. H. Hubbell. 1979. Plantgrowth substances produced by Azospirillum brasilense andtheir effect on the growth of pearl millet (Pennisetumamericanum L.). Appl. Environ. Microbiol. 37:219-226.
49. Umali-Garcia, M., D. H. Hubbell, M. H. Gaskins, and F. B.Dazzo. 1980. Association of Azospirillum with grass roots. Appl.
J. BACTERIOL.
on June 11, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

CYST PRODUCTION AND PIGMENT FORMATION IN A. BRASILENSE
Environ. Microbiol. 39:219-226.50. Van Berkuni, P., and B. B. Bohlool. 1980. Evaluation of nitrogen
fixation by bacteria in association with roots of tropical grasses.Microbiol. Rev. 44:491-517.
51. Vela, G. R. 1974. Survival of Azotobacter in dry soil. Appl.Microbiol. 28:77-79.
52. Vela, G. R., and 0. Wyss. 1963. Improved stain for visualizationof Azotobacter encystment. J. Bacteriol. 87:476-478.
53. Wheeler, M. H., W. J. Tolmsoff, A. A. Bell, and H. H.Mollenhauer. 1978. Ultrastructural and chemical distinction ofmelanins formed by Verticillium dahliae from (+)-scytalone,1,8-dihydroxyphenylalanine. Can. J. Microbiol. 24:289-297.
54. Wheeler, M. H., W. J. Tolmsoff, and S. Meola. 1976. Ultrastruc-ture of melanin formation in Verticillium dahliae with (+)-scytalone as a biosynthetic intermediate. Can. J. Microbiol.22:702-711.
VOL. 169, 1987 1677
on June 11, 2018 by guesthttp://jb.asm
.org/D
ownloaded from





























