Embed Size (px)
Citation preview

Cerebral Cortex August 2008;18:1758--1770
doi:10.1093/cercor/bhm199
Advance Access publication November 21, 2007
Cux-2 Controls the Proliferation ofNeuronal Intermediate Precursors of theCortical Subventricular Zone
Beatriz Cubelos1,2, Alvaro Sebastian-Serrano1, Seonhee Kim3,
Carmen Moreno-Ortiz1, Juan Miguel Redondo2, Christopher
A. Walsh3 and Marta Nieto1
1Centro Nacional de Biotecnologıa, CSIC, Darwin 3, Campus de
Cantoblanco, Madrid 28049, Spain, 2Fundacion Centro Nacional
de Investigaciones Cardiovasculares, Melchor Fenandez
Almagro 3, Madrid 28029, Spain and 3Division of Genetics,
Children’s Hospital Boston and Howard Hughes Medical
Institute, Beth Israel Deaconess Medical Center, Harvard
Medical School, Boston, MA 02115, USA
Whereas neurons of the lower layers (VI--V) of the cerebral cortexare first born from dividing precursors at the ventricular zone, upperlayer neurons (II--IV) subsequently arise from divisions of in-termediate neuronal precursors at the subventricular zone (SVZ).Little is known about mechanisms that control the proliferation ofSVZ neuronal precursors. We herein report that the restrictedexpression of the homeodomain transcription factor Cux-2 in theSVZ regulates the proliferation of intermediate neuronal precursorsand the number of upper layer neurons. In Cux-2-deficient mice(Cux-22/2), there is excessive number of upper layer neurons andselective expansion of SVZ neuronal precursors. Double-labelingexperiments demonstrate that Cux-22/2 upper layer precursorsreenter the cell cycle in a higher frequency than wild-typeprecursors. Overexpression studies indicate that Cux-2 controlscell cycle exit in a cell-autonomous manner. Analysis of Cux-12/2;Cux-22/2 double mutant revealed that Cux-2 controls SVZproliferation independently of Cux-1, demonstrating that this isa unique function of Cux-2, not redundant with Cux-1 activities. Ourresults point to Cux-2 as a key element in the control of theproliferation rates of the SVZ precursors and the number of uppercortical neurons, without altering the number of deep corticallayers.
Keywords: cerebral cortex, Cut, Cux-2, late-born neurons, progenitor,transcription factor
Introduction
The vertebrate cortex is organized into distinct layers, with the
neurons within each layer sharing similar functions, morphol-
ogy, and birthdates. An important advance in the understanding
of brain development is that we now know that the sequential
birth of the distinct cortical layers is achieved via the
generation of an intermediate neuronal precursor for the
upper layers. Whereas neurons of the lower layers arise first
from asymmetric divisions of the radial glial cells in the
ventricular zone (VZ) (Malatesta et al. 2000; Noctor et al.
2001), neurons of the upper cortical layers are born from
intermediate neuronal precursors dividing symmetrically in the
subventricular zone (SVZ) (Haubensak et al. 2004; Miyata et al.
2004; Nieto et al. 2004; Noctor et al. 2004; Zimmer et al. 2004;
Englund et al. 2005; Wu et al. 2005; Cappello et al. 2006). This
2-step mechanism of development has been proposed to
confer an evolutionary advantage by enabling both the
expansion of cortical layers and the appearance of new
neuronal types (Englund et al. 2005; Gotz and Huttner 2005;
Hill and Walsh 2005; Kriegstein et al. 2006).
Indeed, the evolution of the cerebral cortex extended the
cortical surface area and increased the number of neuronal
layers and neuronal circuits. Furthermore, the number of
distinguishable upper layers increases from mice to humans
(reviewed in Marin Padilla 2001) so that the circuit diagram
differs between primates and other mammals (Hill and Walsh
2005).
Remarkably, this increment in the number of superficial
layers of the cortex that occurs during evolution correlates
with an expansion of the SVZ during embryonic development.
This might be indeed attributable to an increase in the number
of intermediate neuronal precursors. Furthermore, the selec-
tive expansion of the upper layers in the outer part of the
cortical gyri and of their precursors at the SVZ has recently
been proposed to contribute to the formation of the gyri
typical of the cerebral cortex of carnivores and primates
(Kriegstein et al. 2006).
Any account of the evolution and development of the
cerebral cortex therefore requires an understanding of the
molecular basis regulating the proliferation of SVZ precursors
and differentiation of the upper layer neurons. Several
molecules have been reported to control precursor cell
proliferation (Takahashi et al. 1995; Chenn and Walsh 2002;
Nowakowski et al. 2002), and the sequential activation of
transcription factors governs the acquisition of specific laminar
fates (Englund et al. 2005; Guillemot et al. 2006). However,
little is known about factors involved in the selective pro-
liferation and differentiation of SVZ precursors and upper layer
neurons (Calegari et al. 2005; Guillemot et al. 2006; Glickstein
et al. 2007). Genes selectively expressed in the neuronal
precursor population of the SVZ during embryonic develop-
ment, such as SVET, Tbr-2, Nex/Math-2, and Cux-2 (Tarabykin
et al. 2001; Nieto et al. 2004; Zimmer et al. 2004; Englund et al.
2005; Wu et al. 2005), may participate in these processes.
Remarkably, 2 of these genes, Cux-2 and SVET, are also
expressed in the upper layer neurons.
Cux-2 encodes a vertebrate homolog of the Drosophila
transcription factor Cut (Quaggin et al. 1996). In the peripheral
nervous system, Cut determines the neuronal fate of external
sensory organ precursor cells (Bodmer et al. 1987; Blochlinger
et al. 1990) and the dendrite morphology of their neuronal
progeny (Grueber et al. 2003). Upper layer neurons express
Cux-2 and a second Cut homolog, Cux-1, which does not show
the same highly SVZ restricted expression during development
(Nieto et al. 2004). The function of mammalian Cux genes in
the nervous system is unknown. The overlapping expression
patterns of Cux-1 and Cux-2 suggest redundant functions for
these genes in neural cells and accordingly Cux-1-deficient
mice (Cux-1–/–) show no specific phenotype related to the
development of the nervous system (Luong et al. 2002).
However, the restricted expression pattern of Cux-2 in the
� The Author 2007. Published by Oxford University Press. All rights reserved.
For permissions, please e-mail: [email protected]

SVZ suggests specific functions for this transcription factor,
possibly in the generation of neurons of the upper cortical
layers. To address these issues, we generated a Cux-2-deficient
mouse strain by targeted gene deletion and investigated the
development of the cerebral cortex in these animals.
Our analysis of Cux-2–/– mice demonstrates that Cux-2
negatively regulates the proliferation of precursors of the
upper layer neurons in the SVZ, which reenter the cell cycle at
a higher frequency in the Cux-2–/– mice. Thus, our results
point to Cux-2 as a gene, the expression of which limits the
proliferation of SVZ intermediate precursor. In this context,
mechanisms modulating Cux-2 expression and functions
appear to be candidate for playing important roles in the
expansion of the cerebral cortex that occurs during evolution.
Experimental Procedures
Animals, Construction of a Mouse Cell Line ConditionallyTargeting Cux-2/Cutl2, and Generation of Cux-2–/– Mice
All animal procedures were approved by the Centro Nacional de
Biotecnologıa Animal Care and Use Committee, in compliance
with National and European Legislation. Cux-2 gene encodes 4
DNA-binding domains: 3 Cut repeats and 1 homeodomain. A
targeting construct was designed to conditionally eliminate
exons 22 and 23, which encode the third Cut repeat and part of
the homeodomain (exons 22 and 23) near the C terminus of the
Cux-2 protein, and to create a premature stop codon 3# after thedeletion. The targeting vector was constructed using a 1.9-kb
DNA fragment that contains regions located 1.4 kb 5# and 0.5 kb
3# of exon 21. The middle arm, containing exons 22 and 23, is
a 1.7-kb polymerase chain reaction (PCR) product. The long arm
is a 7.5-kb NdeI genomic fragment from a mouse lambda library.
A total of 10 lg of targeting vector was linearized with NotI and
then transfected into J1-129 embryonic stem cells by electro-
poration. After selection in G418, surviving colonies were
expanded and analyzed by Southern blot to identify clones that
had undergone homologous recombination. Correctly, targeted
ES cell lines were microinjected into C57BL/6J blastocysts.
Resulting chimeric mice were crossed with C57BL6 mice to
obtain Cux-2+/loxP mice. Mice carrying the conditional allele
(Cux2loxP) were mated with mice expressing CRE recombi-
nase under the human beta-actin promoter (Tg (ACTB-
CRE)2Mrt deleter mice; Jackson Laboratories, Bar Harbor, ME)
on a Swiss Webster background, to obtain mice giving germ line
transmission of the floxed null allele. Heterozygous (het) mice
for this allele (hereafter Cux-2+/–) were crossed with wild-type
(WT) mice to segregate from the beta-actin transgene. Cux-2+/–animals were mated to obtain Cux-2 homozygous mutant mice
(Cux-2–/–). Previous studies with antibodies that recognize
different regions of the Cux-2 protein have found 2 isoforms of
Cux-2 protein in a human cell line. Both isoforms are recognized
by an antibody against the N-terminal part of the protein (anti-
Cux-2 antibody 356) encoded by the undeleted part of the
targeted Cux-2 gene. Anti-Cux-2 antibody 356 shows immuno-
reactivity with mouse Cux-2 (Gingras et al. 2005). Western blot
analysis of the expression of Cux-2 protein in Cux-2–/– mice
using antibody 356 reveals 2 bands in adult cerebral cortex
homogenates (Supplementary Fig. 1). No bands were detected
on brain homogenates from adult Cux-2–/– animals (Supple-
mentary Fig. 1). Immunohistochemistry (IHC) studies using
antibody 356 shows expression of Cux-2 protein in the WT
brains but not in Cux-2–/– animals. These experiments
demonstrate the complete absence of Cux-2 protein and lack
of Cux-2 truncated mutated forms of the protein in the Cux-2–/–
animals. Cux-1–/– mice have been described previously (Luong
et al. 2002) and were obtained from A. J. van Wijnen (University
of Massachusetts Medical School, MA, USA). Animals were
maintained on a C57BL6: Swiss Webster background. Morning
of the day of the appearance of the vaginal plug was defined as
embryonic day (E) 0.5.
Southern Blot and PCR
Genomic DNA was obtained from ES cell clones or tail biopsies
and digested with KpnI. Southern blot was performed using
ExpressHyb hybridization solution (BD Biosciences, Mountain
View, CA) according to the manufacturer’s protocol. A 1-kb
cDNA probe corresponding to exon 24 was used to screen for
positive ES cell clones and to genotype mice carrying the loxP
and null alleles. Mice were screened by Southern blot and the
PCR. Primers for the detection of the null floxed allele were 5#-AAGGGCGGTGATTACAGAGA-3# and 5#-GCCTGCTGTGGTA-GACAGGT-3#; primers for the WT allele were 5#-TCAGCA-CATGGTGTCTGGAT-3#and 5#-CCTACTTTCTGCCTGCTTG-3#.PCR was carried out over 35 cycles of 94 �C for 1 min, 60 �Cfor 1 min, and 72 �C for 1 min in 2.5 mM Mg2Cl and 5 mM
Betaine.
Antibodies, IHC, and Histology
Mice were perfused transcardially with 0.1 M phosphate-
buffered saline (PBS; pH 7.4) followed by cold 4% para-
formaldehyde in PBS. The perfused brains were removed and
postfixed in 4% paraformaldehyde at 4 �C overnight. Fixed
brains were cryoprotected in 30% sucrose in PBS and sectioned
on a cryostat to produce either 10--20 lM cryosections on
Superfrost Plus microscope slides (Fisher Scientific, Pittsburgh,
PA) or 50--100 lM floating cryosections. Sections were blocked
for 1 h at room temperature (r.t.) with 5% horse serum in PBS
containing 0.5% Triton-X 100 (blocking solution) and then
incubated for 1 h at r.t. or overnight at 4 �C with primary
antibodies diluted in blocking solution. Fluorescent-tagged
secondary antibodies (in PBS, 5% horse serum) were applied for
1 h at r.t., and sections were counterstained with Hoechst
33342 (Molecular Probes, Eugene, OR) and mounted in Aqua-
polymount mounting medium (Poly-Labo, Strasbourg, France).
Peroxidase/diaminobenzidine IHC staining was performed as
described (Cubelos et al. 2005). Briefly, after incubation with
primary antibody, sections were incubated with biotinylated
donkey anti-rabbit IgG (Sigma, St Louis, MO) for 1 h. Sections
were then washed 3 times in PBS, incubated with streptavidin-
biotinylated horseradish peroxidase complex, washed, and
incubated with 0.1 mg/ml H2O2 and 0.5 mg/mL diaminobenzi-
dine in PBS. Sections were mounted in glycerol--gelatin.
The following primary antibodies were used at the dilutions
indicated: rabbit polyclonal anti-Cux-1 (clone M222) (1:10) and
anti-Brn-1 (1:50) (Santa Cruz Biotechnologies Inc., St Cruz,
CA), rabbit polyclonal antiphosphohistone H3 (pH3) (1:500)
(Upstate, Spartanburg, SC), rabbit anticleaved caspase-3 (1:500)
(Cell Signaling Tech Inc., Danuers, MA), rabbit anti Ki67
(Novocastra, Newcastle on Tyne, UK), rabbit polyclonal anti-
Cux-2 (antibody 356, a gift from Dr Alex Nepveu of McGill
University Health Centre, Canada), rat anti-bromodeoxiuridine
(BrdU) (1:50) (BD Biosciences), and rabbit anti-Tbr-2 antibody
Cerebral Cortex August 2008, V 18 N 8 1759

(1:500) (Chemicon Inc., Hampshire, UK). Goat anti-rabbit and
goat anti-rabbit and anti-mouse secondary antibodies, conju-
gated to Alexa 488 and 594, respectively (Molecular Probes),
were applied at 1:500. Before staining for Cux-1, Cux-2, Ki67,
BrdU, clorodeoxiuridine (CldU), iododeoxiuridine (IdU), or
pH3, sections were boiled for 30 min in antigen retrieval
solution (Vector Laboratories, Burlingame, CA). This was
followed in the case of staining for BrdU, CldU, IdU, and pH3
by 30-min treatment with 2 N HCl.
Staining for CldU and IdU was performed as described (Aten
et al. 1994). Briefly, slides were incubated overnight at 4 �Cwith a mouse monoclonal anti-BrdU antibody (1:50; BD
Biosciences; cross-reactivity with IdU), then washed in Tris
buffer, pH 8.0, containing 0.5 M NaCl and 1% Tween 20 for
20 min at r.t. to eliminate background and possible cross-reactivity
with CldU, and incubated overnight at 4 �C with rat mono-
clonal anti-BrdU antibodies (1:250; Abcam, Cambridge, UK;
cross-reactivity with CldU). Secondary anti-mouse conjugated
to Alexa 495--conjugated and anti-rat Alexa 598--conjugated
antibodies (1:500; both from Molecular Probes) were applied
for 2 h at r.t. The presence of single positive neurons for IdU
and CldU in the stained cortical sections demonstrated the
specificity of the antibodies and the staining.
BrdU, CldU, and IdU Injections and Cell Counting
All BrdU, CldU, and IdU quantification analysis show results
obtained from the primary sensory cortex (primary somato-
sensory cortex barrel field [S1BF]. Interaural 2.34-2.22, Bregma
1.46-1.58 according to the mouse atlas of Paxinos and Franklin
1997). Similar results were observed in the primary motor
cortex and other regions of the sensory cortex (M1 and pri-
mary somatosensory cortex forelimb region [S1FL], according
to the mouse atlas of Paxinos and Franklin). BrdU, IdU, and
CldU were administered intraperitoneally at 5 mg/mL in PBS. A
single CldU injection was given 10 h after IdU injection at
E14.5. Anatomically matched sections were selected from each
mouse at each stage after BrdU injection (n = 3 Cux-2+/?control mice and n = 3 Cux-2–/– mice), and BrdU was detected
by IHC. The total cortical thickness was subdivided into 10 bins
of equal area, and the number of cells in which at least half of
the nucleus was BrdU positive was counted in each bin.
Identical results were obtained from independent analyses by
2 investigators.
Cortical Layer Thickness and Neuronal Density
For neuronal cell density and cortical thickness, serial sections
were cut from brains of control and Cux-2–/– animal (n = 3).
Then sections were Nissl stained, matched, and photographed.
Measurements and cell counts were performed on the primary
sensory cortex (S1BF, Interaural 2.34-2.22, Bregma 1.46-1.58
according to the mouse atlas of Paxinos and Franklin 1997).
Similar results of increased layer thickness were observed in
S1FL and M1 (Paxinos and Franklin 1997). Analysis was
performed blind by 2 investigators in independent analyses.
Cell density was calculated using the optical fractionator
method (Sterio 1984). Optical dissector was given by a
preexisting microscope grid (x, y, z = 61, 61, 10 lm). For cor-
tical thickness, 4--5 measurements were taken of each brain
region and mean values and standard deviations obtained. Statis-
tical significance was calculated using the 2-tailed Student’s
t-test.
Confocal Microscopy and Imaging
Confocal microscopy was performed with a Radiance 2100
(Bio-Rad, Hercules, CA) Laser Scanning System on a Zeiss
Axiovert 200 microscope. For fluorescence excitation, an argon
ion laser (488 nm), a Krypton--Neon laser (543 nm), and a red
diode (637 nm) were employed. The filter combinations used
for detection of Alexa 488 and Alexa 594 were a 560 DCLPXR
beam splitter and HQ 515/30 emission filter and a 650 DCLPXR
beam splitter with HQ 590/70, respectively. Sequential images
were taken with LaserSharp v5.0 software (Bio-Rad) and
analyzed using LaserPix v.4 image software (Bio-Rad).
Neurosphere Culture, Nucleofection, and Constructs
Dorsal telencephalons of E14.5 WT embryos were dissected
and cells dissociated as described (Nieto et al. 2004).
Dissociated single cells were cultured in DMEM-F12 supple-
mented with N2, fibroblast growth factor (10 ng/mL), and
epidermal growth factor (10 ng/mL) (Gibco, Invitrogen,
Carlsbad, CA). Neurosphere cultures were passaged every 3
days by mechanical dissociation. Dissociated cells were
cotransfected using the nucleofector technic (Amaxa Biosys-
tems, Gaithersburg, MD) with 2.5 lg each of DNA constructs
containing Cux-2 and green fluorescence protein (GFP) cDNAs
under the cytomegalovirus enhancer, chicken b-actin pro-
moter, and rabbit b-globin poly(A) signal (CAG) (cytomegalo-
virus [CMV] and beta-actin) promoter or the empty vector and
GFP construct. Neurosphere formation was monitored after
10 days in culture.
Results
Generation of Cux-2–/– Mice
To investigate the functions of Cux-2 in the development of
the nervous system, we generated mice carrying a null allele of
Cux-2 and analyzed the resulting brain phenotype. The
mutation was designed to remove exons 22--23 that encode
the third Cut repeat and the homeodomain near the C-terminal
end of the protein. Het Cux-2+/– mice were obtained by
breeding mice carrying a Cux-2 conditional allele (Cux-2+/loxP) with CRE deleter mice (see Experimental Procedure and
Fig. 1a--c). Homozygous Cux-2 mutant mice (Cux-2–/–) were
obtained by crossing het Cux-2+/– animals. Cux-2–/– mice
were born at the expected Mendelian ratios and showed
overall normal development and growth (not shown). To
confirm that the mutation of the Cux-2 C terminus effectively
removed expression of Cux-2 protein in mutant mice, postnatal
brain sections of WT and Cux-2–/– mice were stained with
a Cux-2 antiserum against the N-terminal part of the protein
that recognizes the 2 described isoforms of Cux-2 protein
but does not recognize Cux-1 (Gingras et al. 2005). This
demonstrated the complete absence of Cux-2 immunoreactiv-
ity in all brain regions of Cux-2–/– animals, including the upper
layer neurons and neurons of the ventrolateral and piriform
cortex (Fig. 1d, left panels). Western blot experiments
confirmed the complete absence of Cux-2 protein in Cux-2–/–
cortical homogenates (Supplementary Fig. 1). In contrast,
staining with a Cux-1-specific antiserum showed that the Cux-1
protein expression was indistinguishable between Cux-2–/–,
Cux-2+/–, and WT littermates (Fig. 1d and not shown). Thus,
loss of Cux-2 protein expression does not affect the expression
1760 Cux-2 Controls Upper Layer Precursor Proliferation d Cubelos et al.
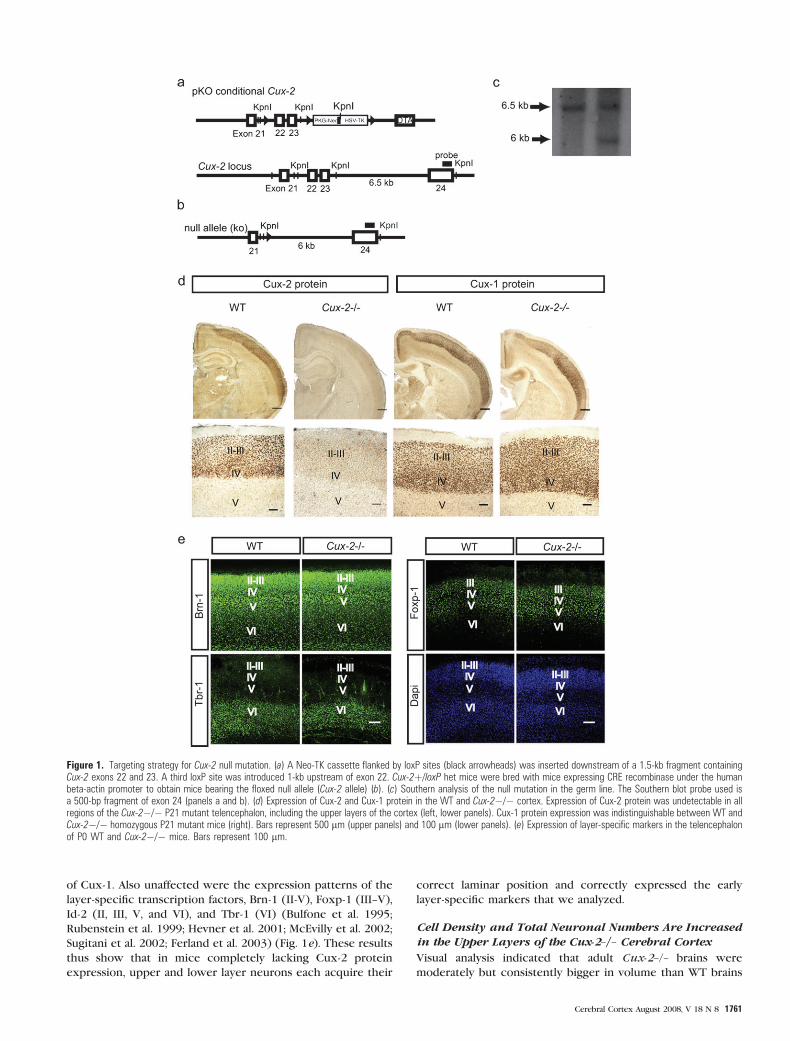
of Cux-1. Also unaffected were the expression patterns of the
layer-specific transcription factors, Brn-1 (II-V), Foxp-1 (III--V),
Id-2 (II, III, V, and VI), and Tbr-1 (VI) (Bulfone et al. 1995;
Rubenstein et al. 1999; Hevner et al. 2001; McEvilly et al. 2002;
Sugitani et al. 2002; Ferland et al. 2003) (Fig. 1e). These results
thus show that in mice completely lacking Cux-2 protein
expression, upper and lower layer neurons each acquire their
correct laminar position and correctly expressed the early
layer-specific markers that we analyzed.
Cell Density and Total Neuronal Numbers Are Increasedin the Upper Layers of the Cux-2–/– Cerebral Cortex
Visual analysis indicated that adult Cux-2–/– brains were
moderately but consistently bigger in volume than WT brains
Figure 1. Targeting strategy for Cux-2 null mutation. (a) A Neo-TK cassette flanked by loxP sites (black arrowheads) was inserted downstream of a 1.5-kb fragment containingCux-2 exons 22 and 23. A third loxP site was introduced 1-kb upstream of exon 22. Cux-2þ/loxP het mice were bred with mice expressing CRE recombinase under the humanbeta-actin promoter to obtain mice bearing the floxed null allele (Cux-2 allele) (b). (c) Southern analysis of the null mutation in the germ line. The Southern blot probe used isa 500-bp fragment of exon 24 (panels a and b). (d) Expression of Cux-2 and Cux-1 protein in the WT and Cux-2�/� cortex. Expression of Cux-2 protein was undetectable in allregions of the Cux-2�/� P21 mutant telencephalon, including the upper layers of the cortex (left, lower panels). Cux-1 protein expression was indistinguishable between WT andCux-2�/� homozygous P21 mutant mice (right). Bars represent 500 lm (upper panels) and 100 lm (lower panels). (e) Expression of layer-specific markers in the telencephalonof P0 WT and Cux-2�/� mice. Bars represent 100 lm.
Cerebral Cortex August 2008, V 18 N 8 1761
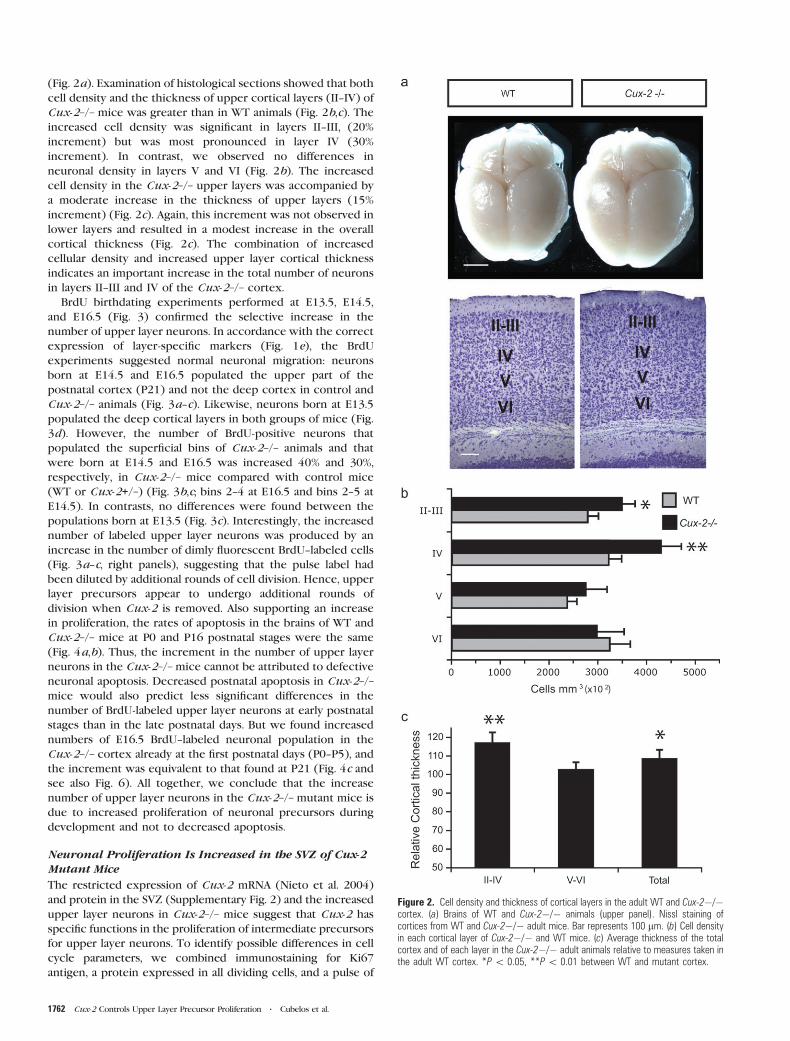
(Fig. 2a). Examination of histological sections showed that both
cell density and the thickness of upper cortical layers (II--IV) of
Cux-2–/– mice was greater than in WT animals (Fig. 2b,c). The
increased cell density was significant in layers II--III, (20%
increment) but was most pronounced in layer IV (30%
increment). In contrast, we observed no differences in
neuronal density in layers V and VI (Fig. 2b). The increased
cell density in the Cux-2–/– upper layers was accompanied by
a moderate increase in the thickness of upper layers (15%
increment) (Fig. 2c). Again, this increment was not observed in
lower layers and resulted in a modest increase in the overall
cortical thickness (Fig. 2c). The combination of increased
cellular density and increased upper layer cortical thickness
indicates an important increase in the total number of neurons
in layers II--III and IV of the Cux-2–/– cortex.
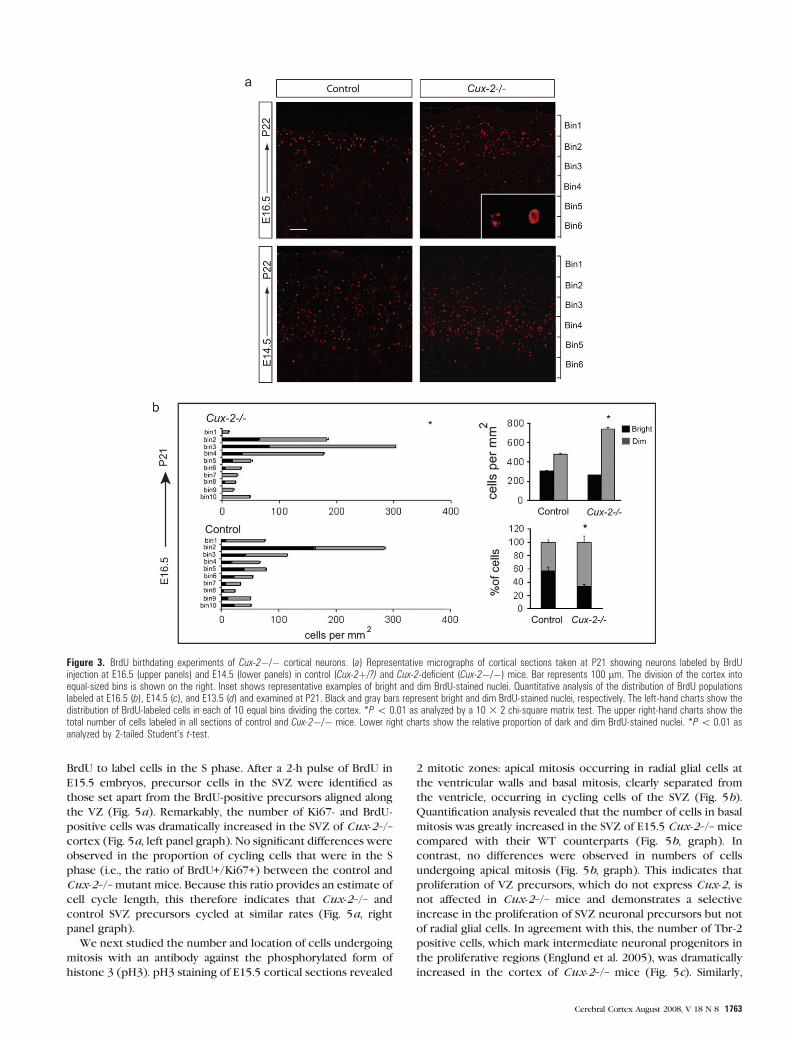
BrdU birthdating experiments performed at E13.5, E14.5,
and E16.5 (Fig. 3) confirmed the selective increase in the
number of upper layer neurons. In accordance with the correct
expression of layer-specific markers (Fig. 1e), the BrdU
experiments suggested normal neuronal migration: neurons
born at E14.5 and E16.5 populated the upper part of the
postnatal cortex (P21) and not the deep cortex in control and
Cux-2–/– animals (Fig. 3a--c). Likewise, neurons born at E13.5
populated the deep cortical layers in both groups of mice (Fig.
3d). However, the number of BrdU-positive neurons that
populated the superficial bins of Cux-2–/– animals and that
were born at E14.5 and E16.5 was increased 40% and 30%,
respectively, in Cux-2–/– mice compared with control mice
(WT or Cux-2+/–) (Fig. 3b,c; bins 2--4 at E16.5 and bins 2--5 at
E14.5). In contrasts, no differences were found between the
populations born at E13.5 (Fig. 3c). Interestingly, the increased
number of labeled upper layer neurons was produced by an
increase in the number of dimly fluorescent BrdU--labeled cells
(Fig. 3a--c, right panels), suggesting that the pulse label had
been diluted by additional rounds of cell division. Hence, upper
layer precursors appear to undergo additional rounds of
division when Cux-2 is removed. Also supporting an increase
in proliferation, the rates of apoptosis in the brains of WT and
Cux-2–/– mice at P0 and P16 postnatal stages were the same
(Fig. 4a,b). Thus, the increment in the number of upper layer
neurons in the Cux-2–/– mice cannot be attributed to defective
neuronal apoptosis. Decreased postnatal apoptosis in Cux-2–/–
mice would also predict less significant differences in the
number of BrdU-labeled upper layer neurons at early postnatal
stages than in the late postnatal days. But we found increased
numbers of E16.5 BrdU--labeled neuronal population in the
Cux-2–/– cortex already at the first postnatal days (P0--P5), and
the increment was equivalent to that found at P21 (Fig. 4c and
see also Fig. 6). All together, we conclude that the increase
number of upper layer neurons in the Cux-2–/– mutant mice is
due to increased proliferation of neuronal precursors during
development and not to decreased apoptosis.
Neuronal Proliferation Is Increased in the SVZ of Cux-2Mutant Mice
The restricted expression of Cux-2 mRNA (Nieto et al. 2004)
and protein in the SVZ (Supplementary Fig. 2) and the increased
upper layer neurons in Cux-2–/– mice suggest that Cux-2 has
specific functions in the proliferation of intermediate precursors
for upper layer neurons. To identify possible differences in cell
cycle parameters, we combined immunostaining for Ki67
antigen, a protein expressed in all dividing cells, and a pulse of
Figure 2. Cell density and thickness of cortical layers in the adult WT and Cux-2�/�cortex. (a) Brains of WT and Cux-2�/� animals (upper panel). Nissl staining ofcortices from WT and Cux-2�/� adult mice. Bar represents 100 lm. (b) Cell densityin each cortical layer of Cux-2�/� and WT mice. (c) Average thickness of the totalcortex and of each layer in the Cux-2�/� adult animals relative to measures taken inthe adult WT cortex. *P\ 0.05, **P\ 0.01 between WT and mutant cortex.
1762 Cux-2 Controls Upper Layer Precursor Proliferation d Cubelos et al.
BrdU to label cells in the S phase. After a 2-h pulse of BrdU in
E15.5 embryos, precursor cells in the SVZ were identified as
those set apart from the BrdU-positive precursors aligned along
the VZ (Fig. 5a). Remarkably, the number of Ki67- and BrdU-
positive cells was dramatically increased in the SVZ of Cux-2–/–
cortex (Fig. 5a, left panel graph). No significant differences were
observed in the proportion of cycling cells that were in the S
phase (i.e., the ratio of BrdU+/Ki67+) between the control and
Cux-2–/– mutant mice. Because this ratio provides an estimate of
cell cycle length, this therefore indicates that Cux-2–/– and
control SVZ precursors cycled at similar rates (Fig. 5a, right
panel graph).
We next studied the number and location of cells undergoing
mitosis with an antibody against the phosphorylated form of
histone 3 (pH3). pH3 staining of E15.5 cortical sections revealed
2 mitotic zones: apical mitosis occurring in radial glial cells at
the ventricular walls and basal mitosis, clearly separated from
the ventricle, occurring in cycling cells of the SVZ (Fig. 5b).
Quantification analysis revealed that the number of cells in basal
mitosis was greatly increased in the SVZ of E15.5 Cux-2–/– mice
compared with their WT counterparts (Fig. 5b, graph). In
contrast, no differences were observed in numbers of cells
undergoing apical mitosis (Fig. 5b, graph). This indicates that
proliferation of VZ precursors, which do not express Cux-2, is
not affected in Cux-2–/– mice and demonstrates a selective
increase in the proliferation of SVZ neuronal precursors but not
of radial glial cells. In agreement with this, the number of Tbr-2
positive cells, which mark intermediate neuronal progenitors in
the proliferative regions (Englund et al. 2005), was dramatically
increased in the cortex of Cux-2–/– mice (Fig. 5c). Similarly,
Figure 3. BrdU birthdating experiments of Cux-2�/� cortical neurons. (a) Representative micrographs of cortical sections taken at P21 showing neurons labeled by BrdUinjection at E16.5 (upper panels) and E14.5 (lower panels) in control (Cux-2þ/?) and Cux-2-deficient (Cux-2�/�) mice. Bar represents 100 lm. The division of the cortex intoequal-sized bins is shown on the right. Inset shows representative examples of bright and dim BrdU-stained nuclei. Quantitative analysis of the distribution of BrdU populationslabeled at E16.5 (b), E14.5 (c), and E13.5 (d) and examined at P21. Black and gray bars represent bright and dim BrdU-stained nuclei, respectively. The left-hand charts show thedistribution of BrdU-labeled cells in each of 10 equal bins dividing the cortex. *P\ 0.01 as analyzed by a 10 3 2 chi-square matrix test. The upper right-hand charts show thetotal number of cells labeled in all sections of control and Cux-2�/� mice. Lower right charts show the relative proportion of dark and dim BrdU-stained nuclei. *P\ 0.01 asanalyzed by 2-tailed Student’s t-test.
Cerebral Cortex August 2008, V 18 N 8 1763
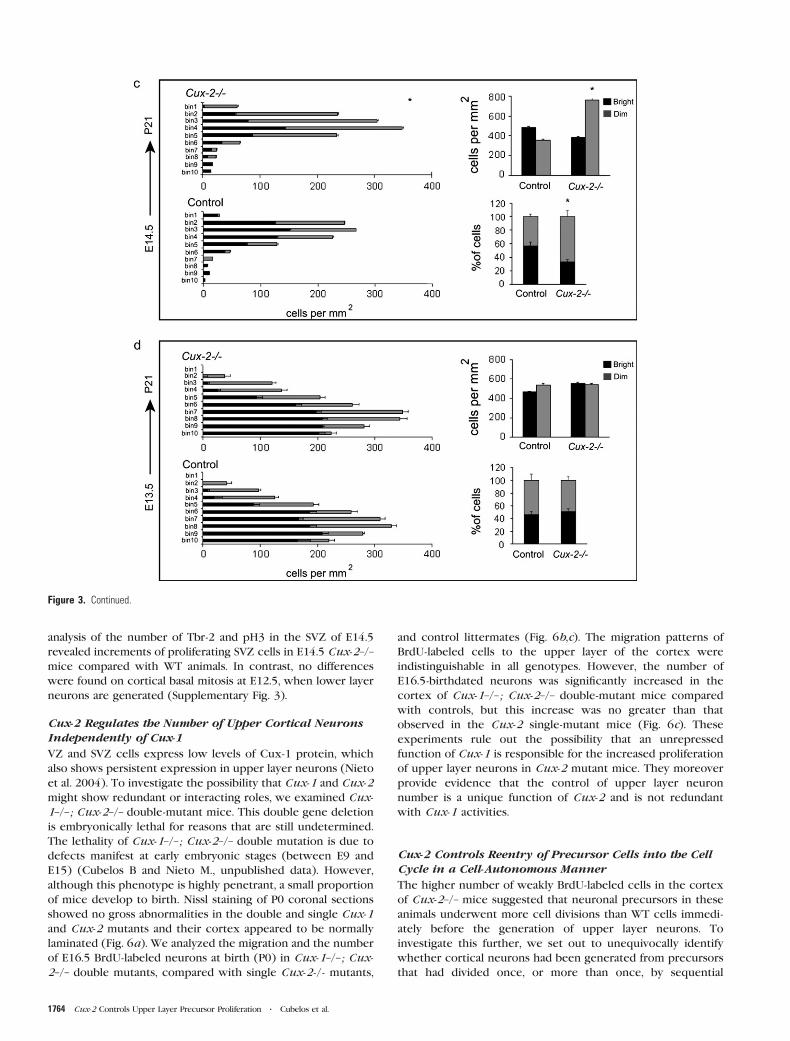
analysis of the number of Tbr-2 and pH3 in the SVZ of E14.5
revealed increments of proliferating SVZ cells in E14.5 Cux-2–/–
mice compared with WT animals. In contrast, no differences
were found on cortical basal mitosis at E12.5, when lower layer
neurons are generated (Supplementary Fig. 3).
Cux-2 Regulates the Number of Upper Cortical NeuronsIndependently of Cux-1
VZ and SVZ cells express low levels of Cux-1 protein, which
also shows persistent expression in upper layer neurons (Nieto
et al. 2004). To investigate the possibility that Cux-1 and Cux-2
might show redundant or interacting roles, we examined Cux-
1–/–; Cux-2–/– double-mutant mice. This double gene deletion
is embryonically lethal for reasons that are still undetermined.
The lethality of Cux-1–/–; Cux-2–/– double mutation is due to
defects manifest at early embryonic stages (between E9 and
E15) (Cubelos B and Nieto M., unpublished data). However,
although this phenotype is highly penetrant, a small proportion
of mice develop to birth. Nissl staining of P0 coronal sections
showed no gross abnormalities in the double and single Cux-1
and Cux-2 mutants and their cortex appeared to be normally
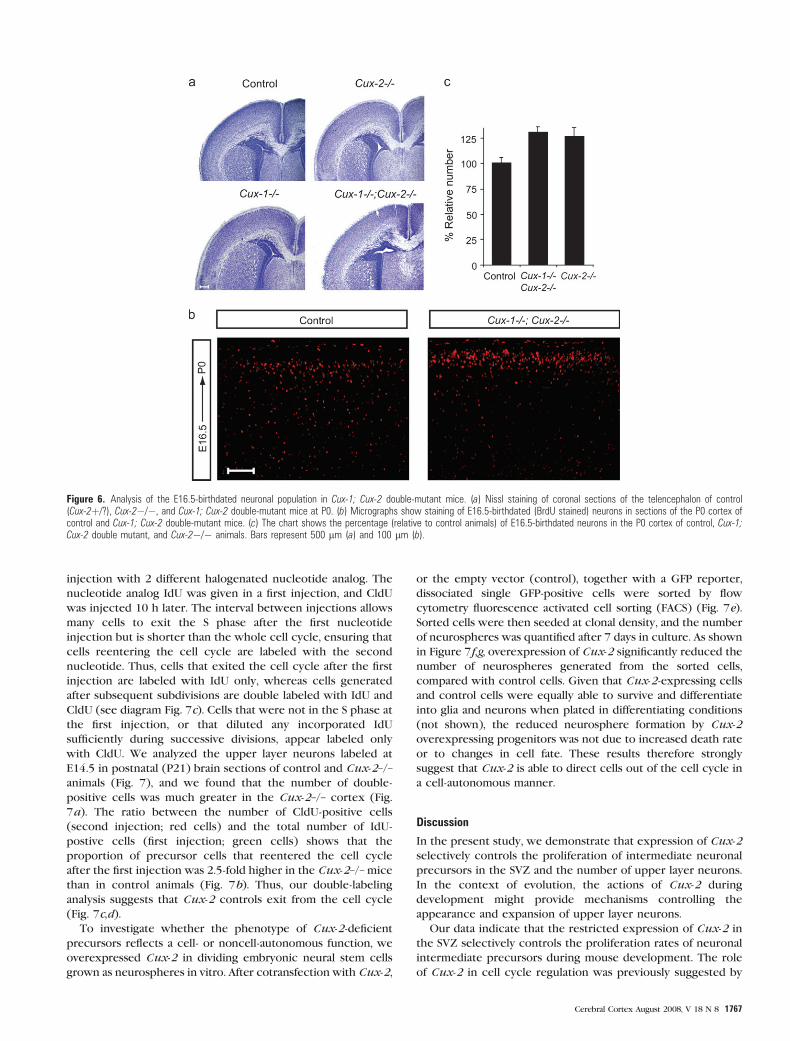
laminated (Fig. 6a). We analyzed the migration and the number
of E16.5 BrdU-labeled neurons at birth (P0) in Cux-1–/–; Cux-
2–/– double mutants, compared with single Cux-2-/- mutants,
and control littermates (Fig. 6b,c). The migration patterns of
BrdU-labeled cells to the upper layer of the cortex were
indistinguishable in all genotypes. However, the number of
E16.5-birthdated neurons was significantly increased in the
cortex of Cux-1–/–; Cux-2–/– double-mutant mice compared
with controls, but this increase was no greater than that
observed in the Cux-2 single-mutant mice (Fig. 6c). These
experiments rule out the possibility that an unrepressed
function of Cux-1 is responsible for the increased proliferation
of upper layer neurons in Cux-2 mutant mice. They moreover
provide evidence that the control of upper layer neuron
number is a unique function of Cux-2 and is not redundant
with Cux-1 activities.
Cux-2 Controls Reentry of Precursor Cells into the CellCycle in a Cell-Autonomous Manner
The higher number of weakly BrdU-labeled cells in the cortex
of Cux-2–/– mice suggested that neuronal precursors in these
animals underwent more cell divisions than WT cells immedi-
ately before the generation of upper layer neurons. To
investigate this further, we set out to unequivocally identify
whether cortical neurons had been generated from precursors
that had divided once, or more than once, by sequential
Figure 3. Continued.
1764 Cux-2 Controls Upper Layer Precursor Proliferation d Cubelos et al.
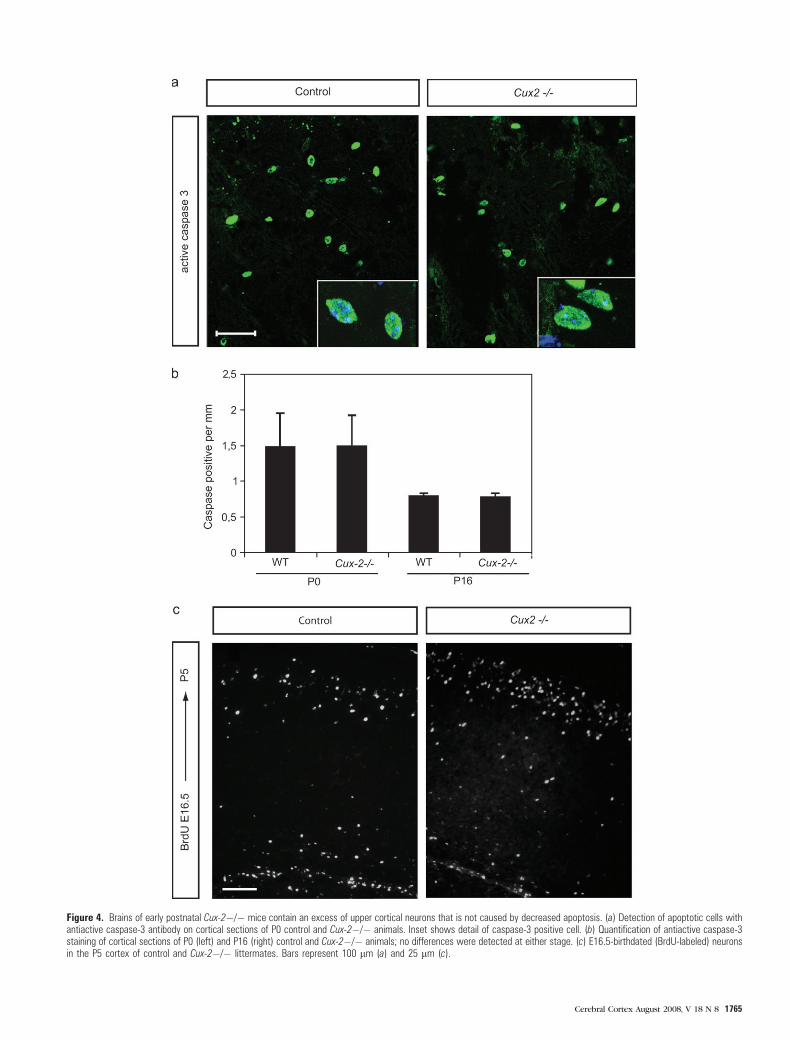
Figure 4. Brains of early postnatal Cux-2�/� mice contain an excess of upper cortical neurons that is not caused by decreased apoptosis. (a) Detection of apoptotic cells withantiactive caspase-3 antibody on cortical sections of P0 control and Cux-2�/� animals. Inset shows detail of caspase-3 positive cell. (b) Quantification of antiactive caspase-3staining of cortical sections of P0 (left) and P16 (right) control and Cux-2�/� animals; no differences were detected at either stage. (c) E16.5-birthdated (BrdU-labeled) neuronsin the P5 cortex of control and Cux-2�/� littermates. Bars represent 100 lm (a) and 25 lm (c).
Cerebral Cortex August 2008, V 18 N 8 1765
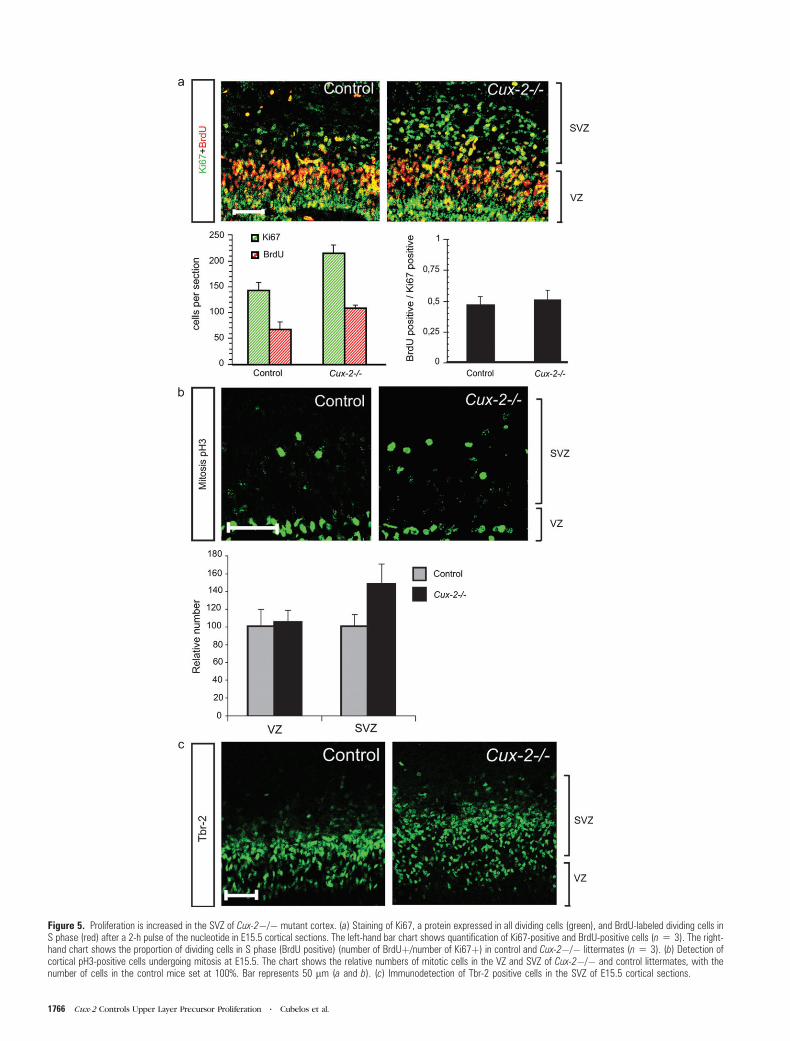
Figure 5. Proliferation is increased in the SVZ of Cux-2�/� mutant cortex. (a) Staining of Ki67, a protein expressed in all dividing cells (green), and BrdU-labeled dividing cells inS phase (red) after a 2-h pulse of the nucleotide in E15.5 cortical sections. The left-hand bar chart shows quantification of Ki67-positive and BrdU-positive cells (n 5 3). The right-hand chart shows the proportion of dividing cells in S phase (BrdU positive) (number of BrdUþ/number of Ki67þ) in control and Cux-2�/� littermates (n 5 3). (b) Detection ofcortical pH3-positive cells undergoing mitosis at E15.5. The chart shows the relative numbers of mitotic cells in the VZ and SVZ of Cux-2�/� and control littermates, with thenumber of cells in the control mice set at 100%. Bar represents 50 lm (a and b). (c) Immunodetection of Tbr-2 positive cells in the SVZ of E15.5 cortical sections.
1766 Cux-2 Controls Upper Layer Precursor Proliferation d Cubelos et al.
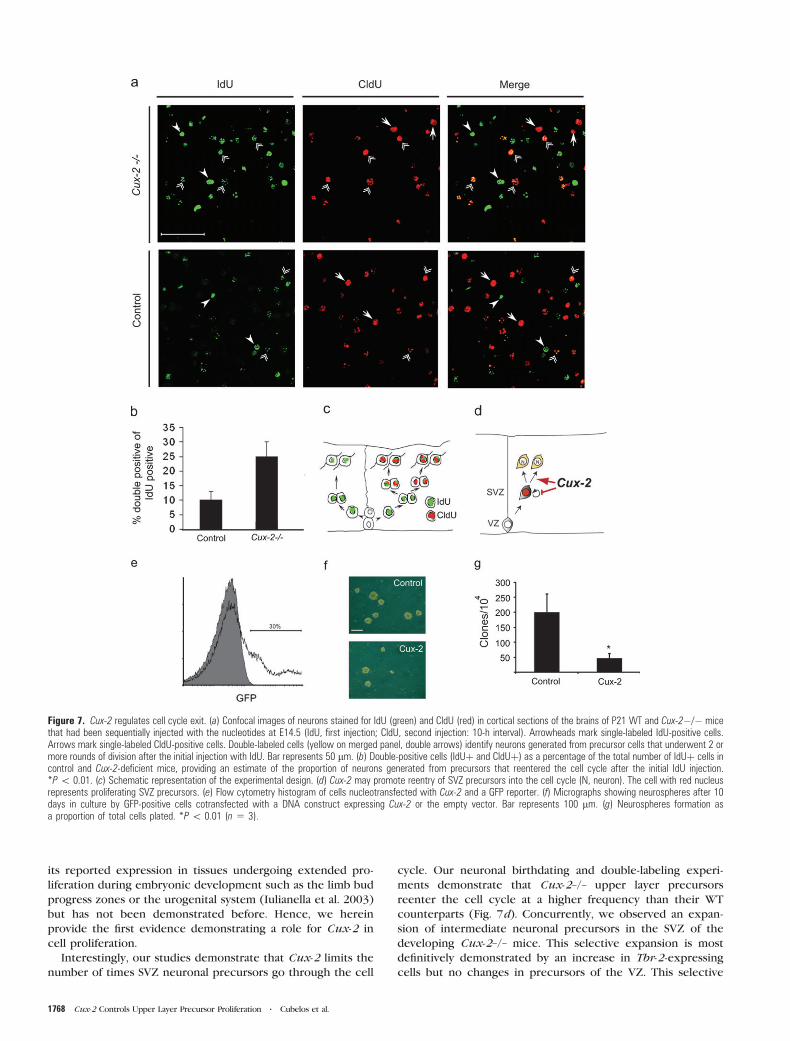
injection with 2 different halogenated nucleotide analog. The
nucleotide analog IdU was given in a first injection, and CldU
was injected 10 h later. The interval between injections allows
many cells to exit the S phase after the first nucleotide
injection but is shorter than the whole cell cycle, ensuring that
cells reentering the cell cycle are labeled with the second
nucleotide. Thus, cells that exited the cell cycle after the first
injection are labeled with IdU only, whereas cells generated
after subsequent subdivisions are double labeled with IdU and
CldU (see diagram Fig. 7c). Cells that were not in the S phase at
the first injection, or that diluted any incorporated IdU
sufficiently during successive divisions, appear labeled only
with CldU. We analyzed the upper layer neurons labeled at
E14.5 in postnatal (P21) brain sections of control and Cux-2–/–
animals (Fig. 7), and we found that the number of double-
positive cells was much greater in the Cux-2–/– cortex (Fig.
7a). The ratio between the number of CldU-positive cells
(second injection; red cells) and the total number of IdU-
postive cells (first injection; green cells) shows that the
proportion of precursor cells that reentered the cell cycle
after the first injection was 2.5-fold higher in the Cux-2–/– mice
than in control animals (Fig. 7b). Thus, our double-labeling
analysis suggests that Cux-2 controls exit from the cell cycle
(Fig. 7c,d).
To investigate whether the phenotype of Cux-2-deficient
precursors reflects a cell- or noncell-autonomous function, we
overexpressed Cux-2 in dividing embryonic neural stem cells
grown as neurospheres in vitro. After cotransfection with Cux-2,
or the empty vector (control), together with a GFP reporter,
dissociated single GFP-positive cells were sorted by flow
cytometry fluorescence activated cell sorting (FACS) (Fig. 7e).
Sorted cells were then seeded at clonal density, and the number
of neurospheres was quantified after 7 days in culture. As shown
in Figure 7f,g, overexpression of Cux-2 significantly reduced the
number of neurospheres generated from the sorted cells,
compared with control cells. Given that Cux-2-expressing cells
and control cells were equally able to survive and differentiate
into glia and neurons when plated in differentiating conditions
(not shown), the reduced neurosphere formation by Cux-2
overexpressing progenitors was not due to increased death rate
or to changes in cell fate. These results therefore strongly
suggest that Cux-2 is able to direct cells out of the cell cycle in
a cell-autonomous manner.
Discussion
In the present study, we demonstrate that expression of Cux-2
selectively controls the proliferation of intermediate neuronal
precursors in the SVZ and the number of upper layer neurons.
In the context of evolution, the actions of Cux-2 during
development might provide mechanisms controlling the
appearance and expansion of upper layer neurons.
Our data indicate that the restricted expression of Cux-2 in
the SVZ selectively controls the proliferation rates of neuronal
intermediate precursors during mouse development. The role
of Cux-2 in cell cycle regulation was previously suggested by
Figure 6. Analysis of the E16.5-birthdated neuronal population in Cux-1; Cux-2 double-mutant mice. (a) Nissl staining of coronal sections of the telencephalon of control(Cux-2þ/?), Cux-2�/�, and Cux-1; Cux-2 double-mutant mice at P0. (b) Micrographs show staining of E16.5-birthdated (BrdU stained) neurons in sections of the P0 cortex ofcontrol and Cux-1; Cux-2 double-mutant mice. (c) The chart shows the percentage (relative to control animals) of E16.5-birthdated neurons in the P0 cortex of control, Cux-1;Cux-2 double mutant, and Cux-2�/� animals. Bars represent 500 lm (a) and 100 lm (b).
Cerebral Cortex August 2008, V 18 N 8 1767
its reported expression in tissues undergoing extended pro-
liferation during embryonic development such as the limb bud
progress zones or the urogenital system (Iulianella et al. 2003)
but has not been demonstrated before. Hence, we herein
provide the first evidence demonstrating a role for Cux-2 in
cell proliferation.
Interestingly, our studies demonstrate that Cux-2 limits the
number of times SVZ neuronal precursors go through the cell
cycle. Our neuronal birthdating and double-labeling experi-
ments demonstrate that Cux-2–/– upper layer precursors
reenter the cell cycle at a higher frequency than their WT
counterparts (Fig. 7d). Concurrently, we observed an expan-
sion of intermediate neuronal precursors in the SVZ of the
developing Cux-2–/– mice. This selective expansion is most
definitively demonstrated by an increase in Tbr-2-expressing
cells but no changes in precursors of the VZ. This selective
Figure 7. Cux-2 regulates cell cycle exit. (a) Confocal images of neurons stained for IdU (green) and CldU (red) in cortical sections of the brains of P21 WT and Cux-2�/� micethat had been sequentially injected with the nucleotides at E14.5 (IdU, first injection; CldU, second injection: 10-h interval). Arrowheads mark single-labeled IdU-positive cells.Arrows mark single-labeled CldU-positive cells. Double-labeled cells (yellow on merged panel, double arrows) identify neurons generated from precursor cells that underwent 2 ormore rounds of division after the initial injection with IdU. Bar represents 50 lm. (b) Double-positive cells (IdUþ and CldUþ) as a percentage of the total number of IdUþ cells incontrol and Cux-2-deficient mice, providing an estimate of the proportion of neurons generated from precursors that reentered the cell cycle after the initial IdU injection.*P\ 0.01. (c) Schematic representation of the experimental design. (d) Cux-2 may promote reentry of SVZ precursors into the cell cycle (N, neuron). The cell with red nucleusrepresents proliferating SVZ precursors. (e) Flow cytometry histogram of cells nucleotransfected with Cux-2 and a GFP reporter. (f) Micrographs showing neurospheres after 10days in culture by GFP-positive cells cotransfected with a DNA construct expressing Cux-2 or the empty vector. Bar represents 100 lm. (g) Neurospheres formation asa proportion of total cells plated. *P\ 0.01 (n 5 3).
1768 Cux-2 Controls Upper Layer Precursor Proliferation d Cubelos et al.

effect of Cux-2 in SVZ cells supports the notion that the
functions of Cux-2 are cell autonomous. Cell-autonomous
mechanisms are also strongly supported by our experiments
showing that overexpression of Cux-2 decreases proliferation
in neural progenitor cells in vitro.
The increased numbers in the number of SVZ precursors in
Cux-2–/– mice are quite significant (about 40% increment).
This excessive proliferation temporally correlates with in-
creased production of late-born neurons that migrate to the
upper layers in normal patterns (30--40% in BrdU experiments).
This causes parallel increments in cell density of layers II--IV
(20--30% increment), as well as a moderate increase in upper
layer thickness (15% increment).
Somewhat surprisingly, Cux-2 does not appear to be
functionally redundant with Cux-1 in the SVZ precursor. The
nonredundancy of Cux-2 in intermediate neuronal precursors
is suggested by its restricted expression in the SVZ, which is
not shown by Cux-1, because it is expressed in VZ precursors
as well. Our analysis of Cux-1–/–; Cux-2–/– double-mutant mice
confirms this notion and demonstrates that the proliferation of
upper layer neuronal precursor does not depend on Cux-1
activity. The 2 Cux genes thus do not cooperate to control
these aspects of neuronal development.
Our findings are of particular interest in the search for
a mechanism to explain how the appearance and selective
expansion of upper cortical layers during development evolved
(Marin Padilla 2001; Hill and Walsh 2005; Kriegstein et al. 2006;
Martinez-Cerdeno et al. 2006). A larger cortex implies that
precursor cells undergo more rounds of division. In addition,
the extended period of neurogenesis adds the upper layer to
the cortex. Indeed, upper cortical layer neurons are highly
represented in the primate cerebral cortex, especially in
humans (reviewed in Hill and Walsh 2005). During develop-
ment and evolution, inhibition of Cux-2-mediated cell cycle
exit in the SVZ could be a target mechanism to expand the
number of upper layer neurons without affecting the number
of deep layer neurons. Thus, the functions of Cux-2 fulfill both
of the developmental and evolutionary necessities of expand-
ing the number of layers. In this regard, evolution might have
required Cux-2 functions in processes such as the addition of
upper layer neurons to the 3 layers of the reptilian cortex, the
later selective expansion of upper layers in higher mammals
(Reiner 1991; Marin Padilla 2001; Kriegstein et al. 2006), or the
focal expansion of the upper layers and the SVZ that has been
proposed to occur during the formation of the gyrencephalic
cortex (Kriegstein et al. 2006).
Supplementary Material
Supplementary figures 1--3 can be found at: http://www.cercor.
oxfordjournals.org/.
Funding
Ministerio de Educacion y Ciencia (SAF2005--0094); Mutua
Madrilena Automovilistica (2004); RYC-2003-006143 to M.N.;
JdC-05-162-74c to B.C.; the Ministerio de Educacion y Ciencia
(BES-2006-13901) to A.S.-S.; National Institute of Neurological
Disorders and Stroke (2RO1 NS032457) to C.A.W.; the Spanish
Ministry of Health and Consumer Affairs and the Pro-
CNIC Foundation to Centro Nacional de Investigaciones
Cardiovasculares.
Notes
We thank B. Alarcon, S. Bartlett, F. Guillemot, M. Mellado, J. M.
Rodrıguez-Frade, P. Bovolenta, M. Guzman, and H. M. van Santen for
critical reading of the manuscript and for their experimental advice.
We thank A. Nepveu (McGill University, Canada) for the anti-Cux-2
antibody, M. Sheng (Picower Institute, MIT, Cambridge, MA) for the
Tbr-1 antiserum, and A. J. van Wijnen (University of Massachusetts
Medical School, Worcester, MA) for the Cux-1 mutant mice. We are
grateful to S. Montalban, S. Gutierrez-Erlandsson, and D. Esteban for
technical assistance. During the generation of theCux-2loxP conditional
mice,M.N. was a Lefler fellow in the laboratory of C.A.W.We are indebted
to C.A.W. for allowing us to continue the project at the Centro Nacional
de Biotecnologıa (Madrid, Spain). C.A.W. is an Investigator at the
Howard Hughes Medical Institute. Conflict of Interest : None declared.
Address correspondence to Email: [email protected].
References
Aten JA, Stap J, Hoebe R, Bakker PJ. 1994. Application and detection of
IdUrd and CldUrd as two independent cell-cycle markers. Methods
Cell Biol. 41:317--326.
Blochlinger K, Bodmer R, Jan LY, Jan YN. 1990. Patterns of expression
of cut, a protein required for external sensory organ development
in wild-type and cut mutant Drosophila embryos. Genes Dev.
4:1322--1331.
Bodmer R, Barbel S, Sheperd S, Jack JW, Jan LY, Jan YN. 1987.
Transformation of sensory organs by mutations of the cut locus of D.
melanogaster. Cell. 51:293--307.
Bulfone A, Smiga SM, Shimamura K, Peterson A, Puelles L, Rubenstein JL.
1995. T-brain-1: a homolog of Brachyury whose expression defines
molecularly distinct domains within the cerebral cortex. Neuron.
15:63--78.
Calegari F, Haubensak W, Haffner C, Huttner WB. 2005. Selective
lengthening of the cell cycle in the neurogenic subpopulation of
neural progenitor cells during mouse brain development. J Neuro-
sci. 25:6533--6538.
Cappello S, Attardo A, Wu X, Iwasato T, Itohara S, Wilsch-Brauninger M,
Eilken HM, Rieger MA, Schroeder TT, Huttner WB, et al. 2006. The
Rho-GTPase cdc42 regulates neural progenitor fate at the apical
surface. Nat Neurosci. 9:1099--1107.
Chenn A, Walsh CA. 2002. Regulation of cerebral cortical size by control
of cell cycle exit in neural precursors. Science. 297:365--369.
Cubelos B, Gimenez C, Zafra F. 2005. Localization of the GLYT1 glycine
transporter at glutamatergic synapses in the rat brain. Cereb Cortex.
15:448--459.
Englund C, Fink A, Lau C, Pham D, Daza RA, Bulfone A, Kowalczyk T,
Hevner RF. 2005. Pax6, Tbr2, and Tbr1 are expressed sequentially
by radial glia, intermediate progenitor cells, and postmitotic
neurons in developing neocortex. J Neurosci. 25:247--251.
Ferland RJ, Cherry TJ, Preware PO, Morrisey EE, Walsh CA. 2003.
Characterization of Foxp2 and Foxp1 mRNA and protein in the
developing and mature brain. J Comp Neurol. 460:266--279.
Gingras H, Cases O, Krasilnikova M, Berube G, Nepveu A. 2005.
Biochemical characterization of the mammalian Cux2 protein.
Gene. 344:273--285.
Glickstein SB, Alexander S, Ross ME. 2007. Differences in cyclin D2 and
D1 protein expression distinguish forebrain progenitor subsets.
Cereb Cortex. 17:632--642.
Gotz M, Huttner WB. 2005. The cell biology of neurogenesis. Nat Rev
Mol Cell Biol. 6:777--788.
Grueber WB, Jan LY, Jan YN. 2003. Different levels of the homeodomain
protein cut regulate distinct dendrite branching patterns of
Drosophila multidendritic neurons. Cell. 112:805--818.
Guillemot F, Molnar Z, Tarabykin V, Stoykova A. 2006. Molecular
mechanisms of cortical differentiation. Eur J Neurosci. 23:857--
868.
Haubensak W, Attardo A, Denk W, Huttner WB. 2004. Neurons arise in
the basal neuroepithelium of the early mammalian telencephalon:
a major site of neurogenesis. Proc Natl Acad Sci USA. 101:
3196--3201.
Cerebral Cortex August 2008, V 18 N 8 1769

Hevner RF, Shi L, Justice N, Hsueh Y, Sheng M, Smiga S, Bulfone A,
Goffinet AM, Campagnoni AT, Rubenstein JL. 2001. Tbr1 regulates
differentiation of the preplate and layer 6. Neuron. 29:353--366.
Hill RS, Walsh CA. 2005. Molecular insights into human brain evolution.
Nature. 437:64--67.
Iulianella A, Vanden Heuvel G, Trainor P. 2003. Dynamic expression
of murine Cux2 in craniofacial, limb, urogenital and neuronal
primordia. Gene Expr Patterns. 3:571--577.
Kriegstein A, Noctor S, Martinez-Cerdeno V. 2006. Patterns of neural
stem and progenitor cell division may underlie evolutionary cortical
expansion. Nat Rev Neurosci. 7:883--890.
Luong MX, van der Meijden CM, Xing D, Hesselton R, Monuki ES,
Jones SN, Lian JB, Stein JL, Stein GS, Neufeld EJ, et al. 2002.
Genetic ablation of the CDP/Cux protein C terminus results in hair
cycle defects and reduced male fertility. Mol Cell Biol. 22:
1424--1437.
Malatesta P, Hartfuss E, Gotz M. 2000. Isolation of radial glial cells by
fluorescent-activated cell sorting reveals a neuronal lineage. De-
velopment. 127:5253--5263.
Marin Padilla M. 2001. [The evolution of the structure of the neocortex
in mammals: a new theory of cytoarchitecture]. Rev Neurol.
33:843--853.
Martinez-Cerdeno V, Noctor SC, Kriegstein AR. 2006. The role of
intermediate progenitor cells in the evolutionary expansion of the
cerebral cortex. Cereb Cortex. 16(Suppl 1):i152--i161.
McEvilly RJ, de Diaz MO, Schonemann MD, Hooshmand F,
Rosenfeld MG. 2002. Transcriptional regulation of cortical neuron
migration by POU domain factors. Science. 295:1528--1532.
Miyata T, Kawaguchi A, Saito K, Kawano M, Muto T, Ogawa M. 2004.
Asymmetric production of surface-dividing and non-surface-dividing
cortical progenitor cells. Development. 131:3133--3145.
Nieto M, Monuki ES, Tang H, Imitola J, Haubst N, Khoury SJ,
Cunningham J, Gotz M, Walsh CA. 2004. Expression of Cux-1 and
Cux-2 in the subventricular zone and upper layers II-IV of the
cerebral cortex. J Comp Neurol. 479:168--180.
Noctor SC, Flint AC, Weissman TA, Dammerman RS, Kriegstein AR.
2001. Neurons derived from radial glial cells establish radial units in
neocortex. Nature. 409:714--720.
Noctor SC, Martinez-Cerdeno V, Ivic L, Kriegstein AR. 2004. Cortical
neurons arise in symmetric and asymmetric division zones and
migrate through specific phases. Nat Neurosci. 7:136--144.
Nowakowski RS, Caviness VS, Jr., Takahashi T, Hayes NL. 2002. Pop-
ulation dynamics during cell proliferation and neuronogenesis in
the developing murine neocortex. Results Probl Cell Differ. 39:1--25.
Paxinos G, Watson C. 1982. The rat brain in stereotaxis coordinates.
New York: Academic Press.
Quaggin SE, Heuvel GB, Golden K, Bodmer R, Igarashi P. 1996. Primary
structure, neural-specific expression, and chromosomal localization
of Cux-2, a second murine homeobox gene related to Drosophila
cut. J Biol Chem. 271:22624--22634.
Reiner A. 1991. A comparison of neurotransmitter-specific and
neuropeptide-specific neuronal cell types present in the dorsal cor-
tex in turtles with those present in the isocortex in mammals: im-
plications for the evolution of isocortex. Brain Behav Evol. 38:53--91.
Rubenstein JL, Anderson S, Shi L, Miyashita-Lin E, Bulfone A, Hevner R.
1999. Genetic control of cortical regionalization and connectivity.
Cereb Cortex. 9:524--532.
Sterio DC. 1984. The unbiased estimation of number and sizes of
arbitrary particles using the disector. J Microsc. 134:127--136.
Sugitani Y, Nakai S, Minowa O, Nishi M, Jishage K, Kawano H, Mori K,
Ogawa M, Noda T. 2002. Brn-1 and Brn-2 share crucial roles in the
production and positioning of mouse neocortical neurons. Genes
Dev. 16:1760--1765.
Takahashi T, Nowakowski RS, Caviness VS, Jr. 1995. The cell cycle of
the pseudostratified ventricular epithelium of the embryonic
murine cerebral wall. J Neurosci. 15:6046--6057.
Tarabykin V, Stoykova A, Usman N, Gruss P. 2001. Cortical upper layer
neurons derive from the subventricular zone as indicated by Svet1
gene expression. Development. 128:1983--1993.
Wu SX, Goebbels S, Nakamura K, Kometani K, Minato N, Kaneko T,
Nave KA, Tamamaki N. 2005. Pyramidal neurons of upper cortical
layers generated by NEX-positive progenitor cells in the subven-
tricular zone. Proc Natl Acad Sci USA. 102:17172--17177.
Zimmer C, Tiveron MC, Bodmer R, Cremer H. 2004. Dynamics of Cux2
expression suggests that an early pool of SVZ precursors is fated to
become upper cortical layer neurons. Cereb Cortex. 14:1408--1420.
1770 Cux-2 Controls Upper Layer Precursor Proliferation d Cubelos et al.






![Grignard and Organozinc Reagents - Sigma-Aldrich · 2021. 2. 23. · R' R R' X Br R Br Ar R O X X R X R Ar X Ar R CuX CuX CuX [Pd] CuX Ar Cl If you are unable to find a reagent for](https://img.pdfslide.us/doc/110x75/6148d92a2918e2056c22f4b2/grignard-and-organozinc-reagents-sigma-aldrich-2021-2-23-r-r-r-x-br-r-br.jpg)




















![Networks of Neuronal Genes Affected by Common and Rare ... · development, cellular proliferation, neuronal migration and projection [15,16]. Another way to identify the connection](https://img.pdfslide.us/doc/110x75/60012353304f7f1b1a79c1fb/networks-of-neuronal-genes-affected-by-common-and-rare-development-cellular.jpg)