Embed Size (px)
Citation preview

Deepika kritaniya
Ph.D scholar
Enrol.no-B1382/14
(College of biotechnology)
DUVASU

overview Introduction
Location of neural stem cell (NSC)
Development of NSC
Cell signalling pathway for NSC
Markesr’s of NSC
Factor affecting growth and multiplication of NSC
Isolation and culture of NSC
NSC for Therapeutic Use
Conclusion

Introduction

The term“neural stem cell” is used loosely to describe cells
that:
Can generate neural tissue or are derived from the nervous
system
Have some capacity for self-renewal, and
Can give rise to cells other than themselves through
asymmetric cell division

Neural stem cell
NSC
Glialprecursors
Sensory
neurons
Motor
neurons
Multipolar
neurons
Inter
neurons
Astroglia Oligodendroglia


The differentiation of the neuroepithelial stem cells intoneurons and glial then proceeds in a temporal specificmanner and in specific region of the developing neuraltube (McConnell,1995; Rao, 1999)
For decades, it was believed that most neurons in the adultcentral nervous system (CNS) were terminallydifferentiated and usually not replaced when they died
It has been established that active neurogenesis, a processof generating functionally integrated neurons fromundifferentiated, multipotent progenitor cells (Gage, 2000;Temple, 2001)

The concept of NSC plasticity and of their dependence onenvironmental cues is strengthened by transplantation andmanipulation studies in vivo (Gage et al., 1995)
This plasticity may perhaps outstretch the brainboundaries, so that NSCs (neuroectodermal in origin) cangive rise to cells that normally derive from germ layersother than the neuroectoderm (Jiang et al., 2002)
In addition, NSCs have been utilized as one of thepotential sources for the cell replacement therapy of CNSdisorders (Bjorklund et al., 2000; Falk, 2009)

Alternative sources of NSC progenitor cell for
cell therapy
Bone marrow
Blood cord
Hair follicles
Skin
Amniotic placental fluid
Human foetus

Location of (NSC)
NSC have been assumed to exist only in the embryonic
nervous system at the beginning of neural tube formation
NSC are present in the central nervous system (CNS), in the
peripheral nervous system (PNS) and in neural crest cell
populations
Germinal regions persist in the adult mammalian brain that is
capable of generating new neurons: the subventricular zone
(SVZ), and the subgranular zone (SGZ)

The spinal cord Progenitor cells in the mammalian fetal CNS
are considered to be concentrated in seven major areas:
Olfactory bulb
Ventricular zone (VZ)
Subventricular zone (SVZ) of the forebrain
Hippocampus
Cerebellum
Cerebral cortex
Spinal cord
Progenitor cells number and pattern of development vary in
different species (Gage, 2000)

Development of (NSCs)
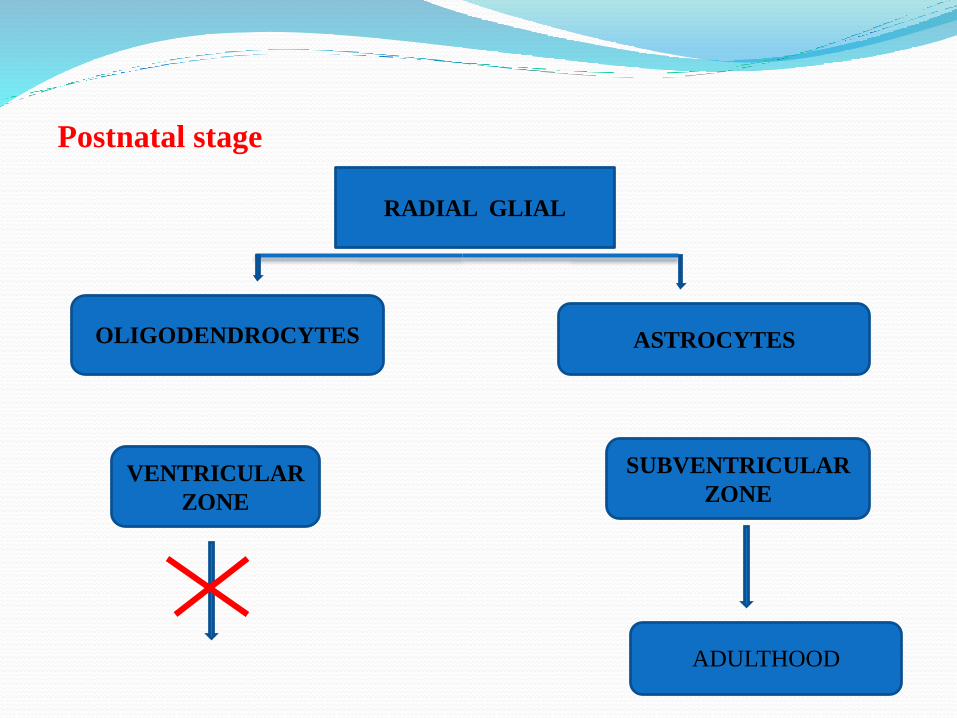
Postnatal stage
RADIAL GLIAL
ASTROCYTES
SUBVENTRICULAR
ZONEVENTRICULAR
ZONE
OLIGODENDROCYTES
ADULTHOOD

When the neural plate first emerges, does it consist solely of
stem cells ?
Analysis of adherent clone production suggests stem cells are
prevalent at early stages
In spinal neural tube from embryonic day 8 (E8) rat, over 50%
of the viable cells at 24 hours are stem cells (Kalyani et al.,
1997, 1998; Qian et al., 2000)
In telencephalon of E10 mouse, estimates of stem cells range
from 5 to 20% (Kilpatrick and Bartlett, 1993)

But the frequency of stem cells declines rapidly (Kalyani et al.,
1997, 1998)
Stem cells seem to be much rarer when neurosphere production
is used as the assay: only 0.3% of E8.5 mouse anterior neural
plate cells make neurospheres
Perhaps neurosphere-generating cells are a subpopulation of
early stem cells

If stem cells are, or rapidly become, a subset of early neural
progenitor cells in vivo, how are they distributed ?
Clonal studies suggest that most glial, both astrocytes and
oligodendrocytes, originate from stem cells (Rao, 1999)
Neurospheres generated from different CNS regions express
region- specific markers
Regulatory sequences control region-specific expression of the
transcription factor Sox2, so that expression is seen in
telencephalic but not spinal cord stem cells (Zappone, 2000)

Neural stem cells cultured from early to mid-gestation give rise
to more neurons than those cultured at later periods (Qian et al.,
2000)
Nestin is an intermediate filament expressed by the
neuroepithelial cells of the neural tube (Lendahl et al., 1990)
Nestin-positive cells make contact with the ventricular surface
and have radially oriented processes

Developmental changes in stem cells are accomplished by
changes in their growth factor
Thus signaling molecules, such as FGF, bone
morphogenetic proteins and Noggin can influence neural
stem cells from neural induction
Growth factor concentrations vary during development of
NSC (Wolpert, 1994)


Cell signalling pathway for neural stem cells
Notch signaling pathway
It has been found that Notch signaling plays a significant rolein neurogenesis in both embryonic and adult brains (Hitoshi etal., 2002; Yoon et al., 2008)
Upon activation of Notch by its ligands, the Notch intracellulardomain (NICD) is released from the membrane andtranslocates to the nucleus
It was reported that subsets of GFAP+ cells function as neuralstem cells in the adult SVZ (Imura et al., 2003)

Transcriptional genes
NICD
Hes1
Hes3
Hes5

WNT Signaling Pathway
The Wnt family of signaling proteins participates in multiple
developmental events during embryogenesis
Mitogenic stimulation
Prevent differentiation
Cell fate determination
Removal of Wnt1 results in severe defects of the midbrain,
cerebellum and spinal cord

Marker’s of Neural Stem Cells
Cell specific markers are a valuable tool in tracing neural
stem cells during development
A reliable marker should identify NSC not only in the
embryonic brain but also in the adult brain
Markers have been described which are either cell surface
proteins such as CD133, Nestin, an intermediate filament
molecule, or Musashi, an RNA binding protein

Nestin
Highly accepted marker for NSC (Frederikson et al., 1988;Naresh k. et al., 2012 )
Nestin expression, neither restricted to the embryonic CNS, northe progenitor cells of neurons, but can be found in the PNS
Nestin seems to play a role in the structural organisation ofcells where it probably participates in remodelling processesand isolated from human fetal striatum and from rat brain(Michalczyk and Ziman, 2005,Li et al. 2005, Zhanget al. 2006)

Sox2
Sox2 is a “founder member” of the Sox gene family
Sox2 can also re-establish pluripotency in terminallydifferentiated cells reprogramming them to induced pluripotentstem cells (iPS) (Silva J et al.,2009)
Sox2 express in the developing central nervous system (CNS)(Ellis et al., 2004, Collignon etal., 1996)
Regulate the Notch pathway which is responsible formaintenance of neural stem cells (Bani-Yaghoub et al., 2006)

RNA-binding protein Musashi-1
Musashi is an evolutionarily conserved family of RNA-binding
proteins that is expressed in the nervous system (Okano et al.,
2002)
Level of expression is selectively higher in NSCs than in neural
precursor cells (Kaneko et al., 2000)
Musashi-1 protein has been found to function in cooperation
with Musashi-2 protein to activate Notch signaling

Pax 6
Members of Pax family proteins are HD (homeodomain)containing transcription factors
Transcription factor Pax6 plays an important role in fatedetermination of neural progenitor cells in animal models(Gehring and Ikeo, 1999)
Pax6 is expressed in ventral region (Spinal Cord) andplays crucial roles in generation of ventral neurons(Kuldeep k. et al., 2014)

CD 133
Widely used as a marker for identification and isolation ofneural precursor cells from normal brain or tumor tissue (Sunet al.,2009)
CD133 five membrane domain glycoprotein and is expressedon immature hematopoietic and progenitor cells (Uchida et al.,2000)
When these CD133+cells were isolated, they were able to formClonal neurospheres and produce new tumours after serialtransplantation (Yuan et al., 2004; Li et al., 2005)

Genes Primers Product
No Temp(°C) (bp)
Nestin F: 5’-AACGCTGAGTCATTGAGAAC-3’
R: 5'-CACTGCCTCCTGGTCTTC-3’
276bp
Sox-2 F: 5'-CGAGTCAAGCGGCCCATGAAC-3‘
R: 5'-TGGCAGCCATCTTGCGTAGG-3'
187bp
Pax6 F:5’AACAGAGTTCTTCGCAACCTGGCTA
G-3‘
R: 5'-TGGCAGCCATCTTGCG TAGG-3'
164bp
Mushashi F: 5-GGTGAAGGAGTGTCTGGTGATGC-3R: 5-TCGAGTCACCATCTTAGGCTGTGC-3
187

Factor Affecting Growth And Multiplication Of
Neural Stem Cells
The most commonly used methods for the isolation and cultureof stem cells use serum-free culture medium supplementedwith various hormones and nutrients and mitogenic growthfactors EGF or FGF-2 (Bottenstein and Sato, 1979, Naresh k.et al., 2012 )
EGF has been used to culture forebrain stem cells asneurospheres from embryonic and adult mouse (Reynolds etal., 1992)
A combination of EGF and FGF-2 is needed to cultureembryonic and adult mouse spinal cord, striatum andsubventricular zone (SVZ) derived progenitor cells asneurospheres (Gritti et al.,1999; Kuldeep k. et al., 2014)

Neurospheres, are kept proliferating by adding growth
factors (EGF), (bFGF) and (LIF) (Palmer et al., 2001)
It also suggest signaling of FGF together with Wnt
signaling regulates late features of the dorsal
telencephalon (Gunhaga et al., 2003)

Isolation and culture of fetal
brain stem cells
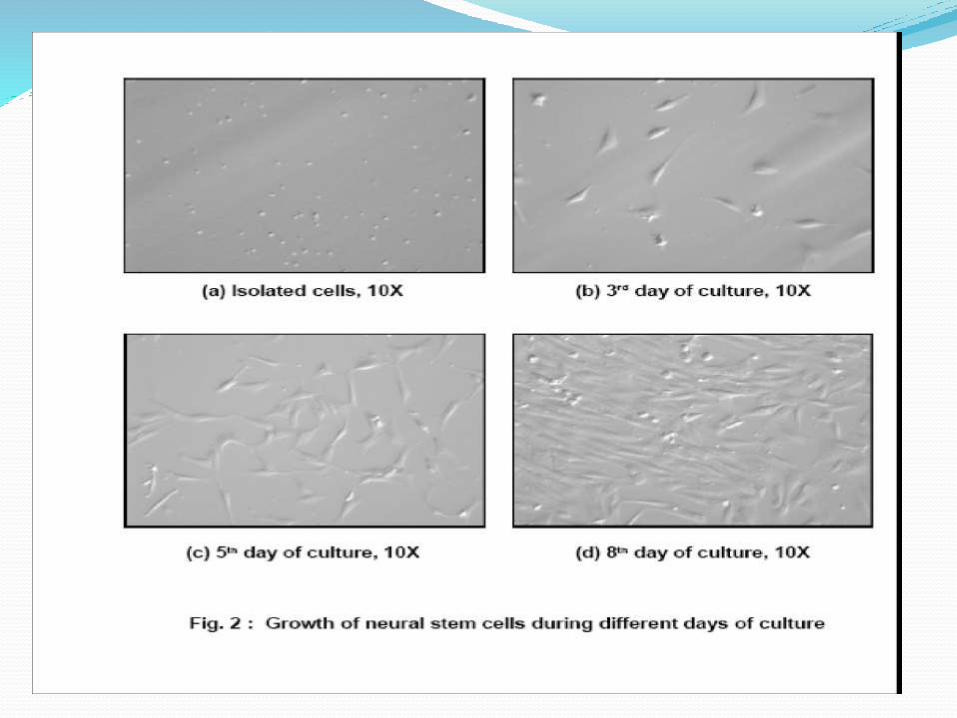

Structure of Neurospheres Karyotype of NSC CELL (P6)

Neural Stem Cells for Therapeutic Use
In the nervous system, replacement of neurons is often
considered to be the main goal of cell therapy
But cells, including stem cells, are already being used as gene
delivery tools and for rescuing neurons rather than replacing
them
NSC can be genetically transduced currently, by the most
efficient and popular way of introducing genes into NSC is by
means of mutagenesis

Clinical trials of neural stem cells
Stem cell
source
Disease Delivery Year References Current
state of the
art
Fetal neural
stem cells
Batten disease,
or neuronal
ceroid
lipofuscinosis
Brain
neurosurgery
2006 Taupin
P.(2006)
phaseI
ongoing
Embryonic
stem cell-
oligodendroc
yte
progenitors
Spinal cord
injury
Spinal cord
injection
2009 Alper
J.(2009)
Withdrawn
because of
risk of
economic
failure
Fetal neural
stem cells
(8-week-old
fetus)
Amyotrophic
lateral
sclerosis
Multisite
injection into
the
spinal cord
2010 Raore
B.(2011)
ongoing (14
patients
transplanted)

Stem cell source Disease Delivery Year References Current state of
the art
Human
embryonic
stem
cells
Stroke Brain
neurosurgery2010 Stroemer
P.(2009)
Under way
Genetically
modified
human
neural stem
cells (Seung
U. Kim,
University
of British
Columbia)
Glioma Intravenous
delivery
2010 Aboody
KS.(2008)
Recruitment
of
patients
Neural fetal
stem cells
(Stem
Cell Factory,
Hospital S.
Maria,
Terni, Italy)
Amyotrophic
lateral
sclerosis
Multisite
injection
into the
spinal cord
2012 Vescovi
AL.(1999)
Recruitment
of patients

Cell Therapy Treatments in Development
Amyotrophic lateral sclerosis (ALS)
Spinal Cord Injury
Ischemic Stroke
Multiple Sclerosis
Alzheimer’s Disease
Traumatic Brain Injury
Peripheral Nerve Injury
Parkinson’s Disease

Drug Treatments in Development
Major Depressive Disorder
Alzheimer’s Disease
Stroke
Traumatic Brain Injury
Post-Traumatic Stress Disorder
Neurodegeneration

CONCLUSION The development of methods to establish NSC lines in vitro has
been one of the main goals of researchers since the discovery ofactive neurogenesis in the adult mammalian CNS
Current preclinical studies strongly suggest that the therapeuticefficacy of stem cell transplantation
For clinical application, it is important that these protective strategiesare proven safe and effective in humans
Several clinical trials using human embryonic stem-derived NSCs orfetal NSCs are currently under way

Our greatest limitation in treating many neurodegenerative disorders is
the lack of understanding of what causes the onset or drives the
progression of sporadic and idiopathic pathologies
They show tropism towards brain pathology, which appears to be
mediated at least in part by chemokines
We will be benefit from repetive and unconvential concept and
unexpected result that will lead us to future discoveries that we cannot
imagine today


Reference
Alvarez-Buylla, A. and Lim, D.A. (2004). For the long run:
Maintaining germinal niches in the adult brain. Neuron, 41: 683-686.
Bani-Yaghoub, M., Tremblay, R.G., Lei, J.X., Zhang, D., Zurakowski, B.
and Sandhu, J.K. (2006). Role of SOX2 in the development of the mouse
neocortex. Dev. Biol., 295: 52-66.
Bjorklund, A. and Lindvall, O. (2000). Cell replacement
therapies for central nervous system disorders. Nat Neurosci., 3: 537-
44.
Bottenstein, J.E. and Sato, G. (1979). Growth of rat neurobalstoma cell line
in serum-free supplemented medium. Proc. Natl. Acad. Sci.,76: 514–517.

Collignon, J., Sockanathan, S., Hacker, A., Cohen-Tannoudji, M., Norris,D.and Rastan, S. (1996). A comparison of the properties of Sox-3 with Sry andtwo related genes Sox-1 and Sox-2. Development,122: 509-520.
Doetsch. F., (2003). The glial identity of neural stem cells., Nat.Neurosci.,6: 1127–1134.
Ellis, P., Fagan, B.M., Magness, S.T., Hutton, S., Taranova, O. and Hayashi,S. (2004). SOX2, a persistent marker for multipotential neural stem cellsderived from embryonic stem cells, the embryo or the adult. Dev. Neurosci.,26: 148-65.
Falk, S. and Sommer, L. (2009). Stage- and area-specific control of stemcells in the developing nervous system. Curr. Opini. Genet. Dev., 19: 454-60.
Frederikson, K., Jat, P.S.,Valtz, N., Levy, D. and McKay, R. (1988).Immortalization of precursor cells from the mammalian CNS. Neuron, 6:439-448.
Gage, F.H. (1995). Survival and differentiation of adult neuronal progenitorcells transplanted to the adult brain. Proc. Natl. Acad. Sci. USA, 92: 11879-11883.

Gage, F.H. (2000). Mammalian neural stem cells. Science, 287: 1433-
1438.
Gehring W.J. and Ikeo, K. (1999). Pax6: mastering eye morphogenesis and
eye evolution. Trends. Genet., 15: 371-377.
Gritti, A., Frolichsthal-Schoeller, P., Galli, R., Parati, E., Cova, L., Pagano,
S., Bjornson, C. and Vescovi, A. (1999). Epidermal and fibroblast growth
factors behave as mitogenic regulators for a single multipotent stem cell-like
population from the subventricular region of the adult mouse forebrain. J.
Neurosci., 19: 3287–3297.
Gunhaga, L., Marklund, M., Sjodal, M., Hsieh, J. C., Jessell, T. M. and
Edlund, T. (2003). Specification of dorsal telencephalic character by
sequential Wnt and FGF signaling. Nat. Neurosci., 6: 701-707.
Hitoshi, S., Alexson, T., Tropepe, V., Donoviel, D., Elia, A.J., Nye, J.S.,
Conlon, R.A., Mak T.W., Bernstein, A. and Van Der Kooy, D. (2002). Notch
pathway molecules are essential for the maintenance, but not the generation,
of mammalian neural stem cells. Genes Dev., 16: 846- 858.

Imura, T., Kornblum, H.I. and Sofroniew, M.V. (2003). Thepredominantneural stem cell isolated from postnatal and adultforebrain but not early embryonic forebrain expresses GFAP., J.Neurosci., 23:2824 –2832.
Jiang, X., Iseki, S., Maxson, R.E., Sucov, H.M. and Morriss-Kay, G.M.(2002). Tissue origins and interactions in the mammalian skull vault. Dev.Biol., 241: 106-116.
Kalyani, A., Hobson, K., and Rao, M. S. (1997). Neuroepithelial stem cellsfrom the embryonic spinal cord: Isolation, characterization and clonalanalysis. Dev. Biol., 186: 202–223.
Kalyani, A.J., Piper, D., Mujtaba, T., Lucero, M.T. and Rao, M.S. (1998).Spinal cord neuronal precursors generate multiple neuronal phenotypes inculture. J. Neurosci., 18: 7856-7868.
Kaneko, Y., Sakakibara, S., Imai, T., Suzuki, A., Nakamura, Y., Sawamoto,K., Ogawa Y., Toyama, Y., Miyata, T. and Okano, H. (2000). Musashi1: anevolutionally conserved marker for CNS progenitor cells including neuralstem cells, Dev. Neurosci., 22139- 22153.

Kilpatrick, T.J. and Bartlett, P.F. (1993). Cloning and growth of
multipotential neural precursors: requirements for proliferation and
differentiation. Neuron, 10: 255-265.
Kilpatrick, T.J., Richards, L.J. and Bartlett, P.F. (1995). The regulation of
neural precursor cells within the mammalian brain. Mol. Cell.Neurosci.,
6(1): 2-15.
Kriegstein, A. and Alvarez-Buylla, A. (2009). The glial nature of embryonic
and adult neural stem cells. Annu. Rev. Neurosci., 32:149 –184.
Kuldeep. K,Renu Singh,Manish .K, P S Mahapatra,Ajay Kumar., Dhruba
Malakar, and Sadhan Bag . International Journal of Neuroscience, 2014;
124(6): 450–456.
Lendahl, U. and McKay, R.D.G. (1990). The use of cell lines in
neurobiology. Trends Neurosci., 13: 132-137.

Li, X. J., Du, Z.W., Zarnowska, E. D., Pankratz, M., Hansen, L. O., Pearce,
R.A. and Zhang, S.C. (2005). Specification of motoneurons from human
embryonic stem cells. Nature Biotechnol., 23: 215–221.
Mujtaba, T., Mayer-Proschel, M.and Rao, M.S. (1998). A common neural
progenitor for the CNS and PNS. Dev Biol., 200:1–15.
McConnell, S.K. (1995). Constructing the cerebral cortex:
neurogenesis and fate determination. Neuron, 15: 761-768.
Naresh k., Nilesh Bari., Kuldeep k.,B C Das., Sadhan Bag.
(2013). Isolation and characterization of neural stem cells from
caprine, Indian Journal of Animal Sciences 83 (2): 146–149,

Okano, H., Imai, T., Okabe, M. (2002). Musashi: Atranslational regulator of cell fates. J. Cell Sci., 115: 1355–1359.
Palmer, T.D., Schwartz, P.H., Taupin, P., B. Kaspar, Stein, S.Aand Gage, F.H. (2001). Progenitor cells from human brain afterdeath. Nature, 411: 42–43.
Pinson, K.,I., Brennan, J., Monkley, S., Avery, B.J., andSkarnes, W.C. (2000). An LDL-receptor-related proteinmediates Wnt signalling in mice. Nature, 407: 535–538.
Qian, X., Shen, Q., Goderie, S.K., He, W., Capela, A., Davis,A.A. and Temple, S. (2000). Timing of CNS cell generation: aprogrammed sequence of neuron and glial cell production fromisolated murine cortical stem cells. Neuron, 28: 69-80.

Rao, M.S. (1999). Multipotent and restricted precursors in the central nervoussystem. Anat. Rec., 257: 137-148.
Reynolds, B.A. and Weiss, S. (1992). Generation of neurons and astrocytesfrom isolated cells of the adult mammalian nervous system. Science, 255: 1707-1710.
Stemple, D.L. and Anderson, D.J. (1992). Isolation of a stem cell for neuronsand glia from the mammalian neural crest. Cell, 71: 973-985.
Sun, Y, Kong. W, Falk. A, Hu. J, Zhou L, Pollard. S, and Smith A. (2009).CD133 (Prominin) negative human neural stem cells are clonogenic andtripotent. Epub. 4(5):5498.
Temple, S. (2001). The development of neural stem cells. Nature, 414: 112-
117. Uchida, N., Buck, D.W., He, D., Reitsma, M.J., Masek, M., Phan,
T.V.,Tsukamoto, A.S., Gage, F.H. and Weissman, I.L. (2000). Directisolation of human central nervous system cells. Proc. Natl. Acad. Sci.USA., 97(26): 14720-14725.





![Creating permissive microenvironments for stem cell ... · neuronal differentiation from NSPCs [33]. Elastic modulus Cell differentiation can be influenced, in part, by the mechanical](https://img.pdfslide.us/doc/110x75/5fcf1181a9c1051993304a8a/creating-permissive-microenvironments-for-stem-cell-neuronal-differentiation.jpg)























