Embed Size (px)
Citation preview

1
Crystal structure at 1.5 Å resolution of Pyrus pyrifolia pistil
ribonuclease responsible for gametophytic self-incompatibility*
Takanori Matsuura‡, Hiroaki Sakai‡, Masaki Unno‡, Koh Ida†, Mamoru Sato†,
Fumio Sakiyama¶, and Shigemi Norioka‡§
From the ‡Institute for Protein Research, Osaka University, Suita, Osaka 565-
0871, Japan, †Graduate School of Integrated Science, Yokohama City University,
Tsurumi-ku, Yokohama 230-0045, Japan, and ¶International Buddhist University,
Habikino, Osaka-538-8501, Japan
§Corresponding author:
Shigemi Norioka
Institute for Protein Research, Osaka University,
3-2 Yamadaoka, Suita, Osaka 565-0871, Japan
Tel. +81-6-6879-8618 Fax. +81-6-6879-8619
Email. [email protected]
Copyright 2001 by The American Society for Biochemistry and Molecular Biology, Inc.
JBC Papers in Press. Published on September 27, 2001 as Manuscript M107617200 by guest on M
arch 30, 2018http://w
ww
.jbc.org/D
ownloaded from

2
Running Title
Crystal Structure of Pyrus pyrifolia S3-RNase
by guest on March 30, 2018
http://ww
w.jbc.org/
Dow
nloaded from

3
SUMMARY
The crystal structure of the Pyrus pyrifolia pistil ribonuclease (S3-RNase)
responsible for gametophytic self-incompatibility was determined at 1.5 Å
resolution. It consists of eight helices and seven β-strands, and its folding
topology is typical of RNase T2 family enzymes. Based on a structural
comparison of S3-RNase with RNase Rh, a fungal RNase T2 family enzyme, the
active site residues of S3-RNase assigned were His33 and His88 as catalysts and
Glu84 and Lys87 as stabilizers of an intermediate in the transition state.
Moreover, amino acid residues that constitute substrate binding sites of the two
RNases could be superimposed geometrically. A hypervariable (HV) region
that has an S-allele-specific sequence comprises a long loop and short α-helix.
This region is far from the active site cleft, exposed on the molecule’s surface,
and positively charged. Four positively selected (PS) regions, in which the
number of nonsynonymous substitutions exceeds that of synonymous ones, are
located on either side of the active site cleft, and accessible to solvent. These
structural features suggest that the HV or PS regions may interact with a pollen
S-gene product(s) to recognize self and non-self pollen.
by guest on March 30, 2018
http://ww
w.jbc.org/
Dow
nloaded from

4
INTRODUCTION
Many flowering plants have a self-incompatibility system which recognizes
self or non-self between the pistil and pollen (tube) after pollination and
suppresses growth of the self-pollen tube to prevent self-fertilization (1,2).
Gametophytic self-incompatibility (GSI)1 is controlled genetically by a single
locus (S-locus) with multiple alleles (1,2). When a pollen grain lands on a
stigma of the pistil, a process which discriminates as to whether an S-allele of
the pollen matches one of the two S-alleles of the pistil takes place. The pollen
grain germinates on the stigma and grows into the style toward the embryo. If
its S-allele matches one of the two S-alleles of the pistil, pollen tube growth is
arrested in the style, and no fertilization takes place. In solanaceous,
scrophulariaceous, and rosaceous plants that have GSI, the pistil glycoproteins
that cosegregate with the S-alleles have been identified as ribonucleases of the
RNase T2 family (S-RNase) (3). McClure et al. reported that pollen rRNA is
degraded after self-pollination but not after cross-pollination and suggested that
GSI expression is mediated by degradation of the pollen rRNA of self-pollen
tubes by S-RNase, leading to depletion of protein biosynthesis and the eventual
arrest of tube growth (4). S-RNase has been confirmed necessary for GSI from
results of gain-of-function and loss-of-function experiments on transgenic plants
of solanaceous species (5,6). Transgenic experiments also have shown that
the RNase activity of S-RNase is necessary for GSI (7), that in petunia the
carbohydrate moiety is not responsible for GSI (8), and that mutant S-RNase
by guest on March 30, 2018
http://ww
w.jbc.org/
Dow
nloaded from

5
which has lost RNase activity acts as a dominant negative for GSI (9).
Based on these findings, two models have been proposed to explain S-
allele-specific inhibition of pollen tube growth; the receptor and inhibitor models
(10). In the receptor model, the pollen S-gene product(s) is the receptor that
incorporates the S-RNase that matches the pollen S-allele. In the inhibitor
model, S-RNase enters the pollen tube nonspecifically and is inhibited, except
for the enzyme that has the same pollen S-allele. Recent,
immunocytochemical research has shown that S-RNase enters and is
accumulated in the cytoplasm of all pollen tube haplotypes without S-allele
specificity, experimental support for the inhibitor model (11). The GSI
mechanism, however, is still not clear because no pollen S-gene product(s) that
interacts with S-RNase and causes self and non-self recognition between the
pollen and pistil has yet been found.
Amino acid sequence analyses of solanaceous S-RNases have identified two
hypervariable regions, HVa and HVb, with extremely high levels of divergence
between allelic sequences; candidates for sequences that would be recognized
as S-allele-specific by pollen (12). In fact, the S-allele specificity of the
Solanum chacoense S11-RNase was changed to the S13-allele by swapping the
HVa and HVb domains (13), and mutation in the HVa and HVb regions of S11-
RNase produced a new S-RNase with dual specificity, S11- and S13-alleles (14).
In contrast, only one hypervariable region, (HV)1, corresponding to the HVa of
the solanaceous S-RNase, has been detected in sequences of rosaceous S-
by guest on March 30, 2018
http://ww
w.jbc.org/
Dow
nloaded from

6
RNases (15,16), the HVb region apparently being deleted (16,17). This
structural feature suggests that the recognition mechanism of S-allele specificity
may differ for solanaceous and rosaceous S-RNases.
Knowledge of the three-dimensional structure is required for further
investigation of the recognition mechanism of rosaceous S-RNases at the
molecular level. We therefore made a three-dimensional structural analysis of
the S-RNases from Pyrus pyrifolia (Japanese pear), a member of the Rosaceae.
Seven S-RNases (S1 – S7-RNase) have been identified in and purified from the
styles of Pyrus pyrifolia (15,18). From these we chose S3-RNase for this X-ray
crystallographic analysis because it has only two N-glycosylation sites (19), and
its amino acid sequence is highly homologous to that of Pyrus pyrifolia S5-RNase
(95.5% identity) (15). Identification of the three-dimensional structures of the
S3- and S5-RNases will pave the way for a structural comparison that provides
high resolution information on molecular recognition between these S-RNases
and the pollen S-gene product(s). We report the crystal structure of Pyrus
pyrifolia S3-RNase at 1.5 Å resolution and discuss the recognition site(s) present
on S3-RNase and the structural basis of its enzymatic activity. This is the first
report on the three-dimensional structure of a rosaceous S-RNase.
by guest on March 30, 2018
http://ww
w.jbc.org/
Dow
nloaded from

7
EXPERIMENTAL PROCEDURES
Crystallization and Data collection— Pyrus pyrifolia S3-RNase was purified
and crystallized as described previously (20). The crystals belong to the P21
space group with unit cell dimensions of a=45.65, b=52.59, c=47.57 Å, and
β=106.45° (20). Diffraction data were collected with an image plate detector
Raxis IV (Rigaku) at the beam line of BL40B2 at SPring-8 to 1.5 Å resolution.
Image data were processed by the programs DENZO and SCALEPACK (21)
(Table I). Multiple isomorphous replacement was used to determine the crystal
structure of S3-RNase. Two ethylmercurithiosalicylic acid derivatives (Hg1,
Hg1’), a mercury (II) ammonium thiocyanate derivative (Hg2), and a lead (II)
acetate derivative (Pb) were prepared by soaking the crystals in heavy atom
reagents (Table I). All diffraction data on the derivative crystals were collected
at the Photon Factory with a CCD detector Quantum 4R (ADSC) at the beam
line of BL6A and processed by the MOSFLM (22) and SCALA (23) programs.
Crystal structure determination— Patterson maps were automatically
interpreted by the program SOLVE (24) to locate the heavy atom coordinates of
each derivative. By use of the solution coordinates, MIR phase calculation was
done at 2.5 Å resolution by the maximum-likelihood refinement program SHARP
(25) (Table I). The experimental MIR phases were improved by the DM (23)
and wARP (26) programs. First, the phases at 2.5 Å resolution gradually were
extended to 1.7 Å resolution by DM with solvent flattening and histogram
matching. The phases obtained were further refined and extended to 1.5 Å
by guest on March 30, 2018
http://ww
w.jbc.org/
Dow
nloaded from

8
resolution by weighted averaging of multiple-refined dummy atomic models
implemented in wARP. This technique for averaging the structure factors was
very effective for improving the phases and provided substantial improvement in
the resulting electron density map. The subsequent main chain trace was done
by wARP. Based on the automatically traced main chain coordinates, 200
amino acid residues were successfully built by program O (27). X-ray
crystallographic refinement was done by the CNS program (28). One cycle of
simulated annealing refinement followed by positional and temperature factor
refinements was repeated several times. Throughout the refinements bulk
solvent correction was applied, and the resolution range gradually was extended
from 2.3 to 1.5 Å. At each stage of crystallographic convergence, the model
was carefully checked and rebuilt into the simulated annealed omit map
calculated by CNS. Solvent water molecules were picked up from a difference
Fourier map by use of the automated scripts implemented in CNS. The lower
cut off level for picking up the solvent water molecules in the residual electron
densities was set at 3σ. High-resolution model coordinates of sugar chains
were obtained from the Uppsala web server, HIC-Up (29). The crystal structure
was refined to the respective R- and free R-factors of 17.2% and 20.2% in the
resolution range of 500-1.5 Å. The final model has 1643 protein atoms, 59
sugar atoms, and 266 water atoms. A Ramachandran plot from the program
PROCHECK (30) shows that 91.5% of the residues are in the most favorable
region, 8.5% in the additional allowed region, and none in the generally allowed
by guest on March 30, 2018
http://ww
w.jbc.org/
Dow
nloaded from

9
and disallowed regions. The coordinates have been deposited in the Protein
Data Bank with the accession code 1IQQ.
by guest on March 30, 2018
http://ww
w.jbc.org/
Dow
nloaded from

10
RESULTS
Overall Structure— The crystal structure of Pyrus pyrifolia S3-RNase was
determined by the multiple isomorphous replacement (MIR) and refined to an R-
factor of 17.2% at 1.5 Å resolution. Table I summarizes the data collected and
gives the final refinement statistics. The geometry of the current model is such
that the root mean square deviations (rmsds) from the ideal values are 0.022 Å
for bond length and 2.02° for bond angles.
A stereo view of the three-dimensional structure of S3-RNase is shown in Fig.
1. The molecule’s size is approximately 40 Å x 50 Å x 30 Å. This protein has
an α + β type structure consisting of eight helices (six α-helices [α1, α2, α4, and
α6-α8], two 310-helices [α3 and α5]), and seven β-strands (β1-β7) (Fig. 1a).
The folding topology of its main chain is very similar to the topologies of the
RNase T2 family enzymes: the RNase Rh from Rhizopus niveus (31,32), RNase
LE from cultured tomato cells (33), and RNase MC1 from bitter gourd seeds (34),
all of whose tertiary structures have been determined (Fig. 1b). Two secondary
structural element characteristics are present in S3-RNase; the length of the α2-
helix is shorter than the lengths of the other RNases, and a very short 310-helix
(α3), Arg74-Lys76, not seen in the RNase T2 family, is present (Fig. 2). The
electron density map for S3-RNase clearly shows four disulfide bridges: Cys15-
Cys22, Cys48-Cys91, Cys155-Cys193, and Cys170-Cys181. The topologies of these
bridges are the same as those of the Pyrus pyrifolia S4-RNase and Nicotiana
alata S2-RNase determined by mass spectrometry of fragmented peptides
by guest on March 30, 2018
http://ww
w.jbc.org/
Dow
nloaded from

11
(35,36).
Catalytic Site— The structure of S3-RNase was compared in detail with that
of RNase Rh to search for catalytic site residues, because many RNase Rh
studies on the structure-function relationship of this RNase T2 family enzyme
have been done by chemical modification (37-39), site-directed mutagenesis
(40-45), and X-ray crystallographic analysis (31,32). The main chain
frameworks, including the catalytic P1 site residues of RNase Rh (β2 and α4),
were well superimposable on those of S3-RNase (Fig. 1b), indicative that their P1
site topologies are similar. The P1 site is here defined according to the
nomenclature of the subsites of RNase A given by Richards and Wyckoff (46).
As shown in Fig. 3a, the His33 and His88 of S3-RNase were superimposable
geometrically on the general acid and base catalysts His(46) and His(109) of RNase
Rh (amino acids numbered in parentheses are those of RNase Rh) (32,38,41),
suggesting that His33 and His88 are the catalysts in S3-RNase. Glu84 and Lys87
are the respective counterparts of the Glu(105) and Lys(108) of RNase Rh which are
considered to stabilize a pentacovalent intermediate of substrate RNA in the
transition state (32,33,42-44). Glu84 and Lys87 therefore may have the same
respective roles as Glu(105) and Lys(108). Trp36 is the counterpart of the Trp(49) of
RNase Rh (Fig. 3a) which functions in the fixation of catalytically important
Glu(105) and His(109). Trp(49)Nε1 forms a hydrogen bond with the γ-carboxyl group
of Glu(105), and the indole ring of Trp(49) has a partial stacking interaction with the
imidazole ring of His(109) (32). In S3-RNase, a similar stacking interaction
by guest on March 30, 2018
http://ww
w.jbc.org/
Dow
nloaded from

12
between Trp36 and His88 is possible because the distance between the indole
ring of Trp36 and the imidazole ring of His88 is about 3.50 Å, and both rings are
nearly parallel (Fig. 3a). Trp36Nε1 cannot, however, interact directly with Glu84Oε1
by hydrogen bonding because the distance between the two atoms is 4.59 Å.
Possibly, they may interact via a water molecule. Indeed, a water molecule that
can hydrogen bond to both Trp36Nε1 and Glu84Oε1 is present in the P1 pocket (Fig.
3a).
Substrate Binding Sites— Two pockets, which correspond to the base
binding B1 and B2 sites of RNase Rh (32,33), are present on either side of the P1
site in S3-RNase (Fig. 1c). These B1 and B2 sites also have the nomenclature
given by Richards and Wyckoff (46). The amino acid residues that constitute
these sites could be superimposed geometrically on S3-RNase and RNase Rh
(Figs. 3b and 3c).
As for the B1 site of S3-RNase, Trp36, Ser38, and Asp44 are the respective
counterparts of the Trp(49), Asp(51), and Tyr(57) of RNase Rh (Fig. 3b). The Trp(49)
of RNase Rh has two important functions; to fix the active side residues
(described above) and to recognize the base of the substrate RNA by an
aromatic π-π stacking interaction together with Tyr(57) (called double-sided
stacking) (32,33). Trp36 may bind to the base of the substrate in the same
manner as Trp(49), but the position corresponding to Tyr(57) is occupied by Asp44,
consequently double-sided stacking recognition is impossible in S3-RNase (Fig.
3b). The base therefore probably is recognized at the B1 site of S3-RNase by a
by guest on March 30, 2018
http://ww
w.jbc.org/
Dow
nloaded from

13
single-sided stacking interaction with Trp36, unlike RNase Rh and RNase LE
(32,33). X-ray crystallographic data on the RNase Rh / 2’-AMP complex shows
that Asp(51) contributes to the adenylic acid preference at the B1 site of RNase Rh
by hydrogen bonding to the adenine base of 2’-AMP (32,33). The position
corresponding to Asp(51), however, is occupied by Ser38 in S3-RNase (Fig. 3b).
Experiments on substrate specificity done with dinucleotide monophosphate
showed that the base specificity at the B1 site of S3-RNase is wider than that of
RNase Rh (data not shown). This extended base specificity may be due to
substitution of Asp(51) with Ser38, to single-sided stacking with Trp36, or both.
The B2 site of S3-RNase seems to be comprised of Gln9, Pro69, Asn70, Val71,
Phe72, and Phe80 (Fig. 3c), which geometrically correspond to the Gln(32), Pro(92),
Ser(93), Asn(94), Gln(95), and Phe(101) of RNase Rh that have been identified as
components of the B2 site based on X-ray crystallographic data for the complex
with d(ApC) (33). In RNase Rh, the base is thought to be recognized at the B2
site by an aromatic stacking interaction with Phe(101) and van der Waals contact
with the side-chains of Asn(94) and Gln(95) (33). Similar recognition should occur
at the B2 site of S3-RNase because Val71, Phe72, and Phe80, respectively, can be
superimposed geometrically on Asn(94), Gln(95), and Phe(101) (Fig. 3c).
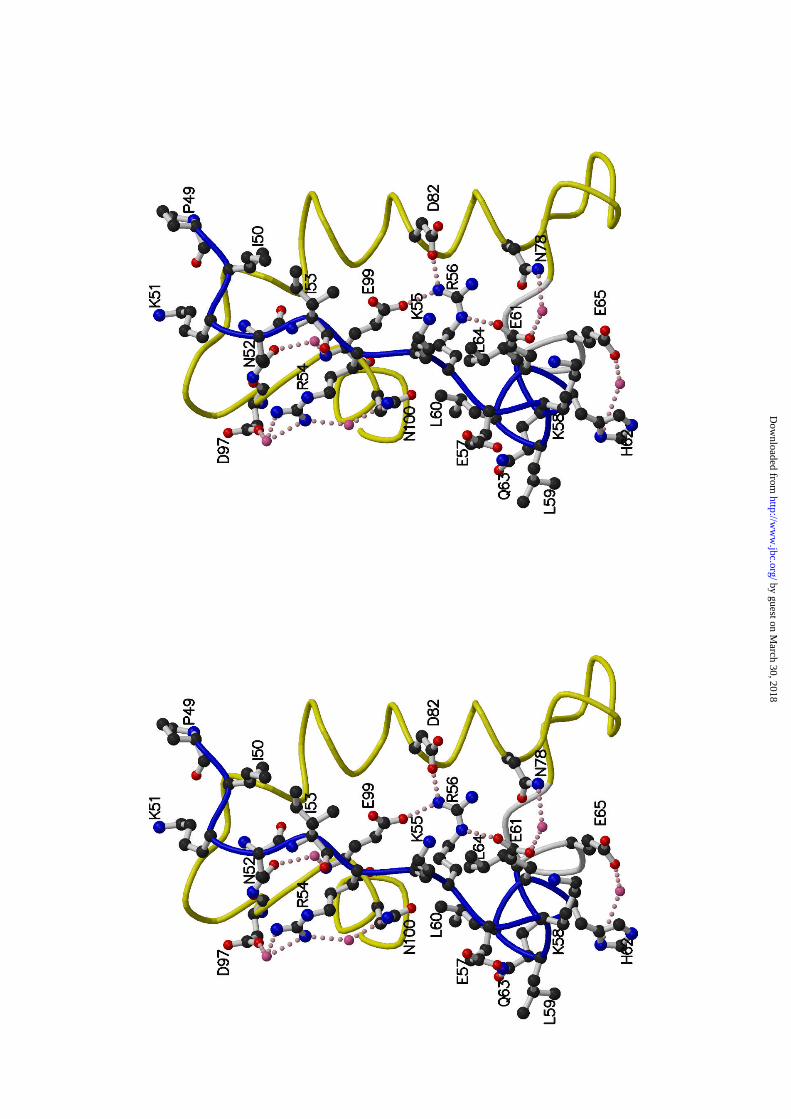
Hypervariable Region— One hypervariable (HV)1 region with an allele-
specific sequence is present in the rosaceous S-RNases (Fig. 4a) and is a
candidate for the recognition site of pollen S-gene product(s) (15). The HV
region of S3-RNase (Pro49-Gln63) is made up of a loop (Pro49-Glu57) and half of an
by guest on March 30, 2018
http://ww
w.jbc.org/
Dow
nloaded from

14
α2-helix (Lys58-Gln63) and is exposed on the molecule’s surface (Figs. 1a and 4b).
Although the exposed loop appears to be somewhat flexible, three of its nine
amino acid residues (Asn52, Arg54, and Arg56) interact with the amino acid
residues inside the molecule by hydrogen bonding (Fig. 5). Asn52Nδ2 is
hydrogen bonded to the carbonyl group of Asp97 directly, and Asn52Oδ1 to the
amide of Glu99 via a water molecule. The δ-guanidino group of Arg54 forms
water-mediated hydrogen bonds individually with Asn100Nδ2 and Asp97Oδ1. The
δ-guanidino group of Arg56 directly forms three hydrogen bonds with Asp82Oδ1,
Glu99Oε1, and Glu61Oε1 (Fig. 5).
In contrast, the half of the α2-helix (Lys58-Gln63), located within the HV region,
has six amino acid residues. Of these, the side-chains of Leu60 and Glu61
interact with other amino acids (Fig. 5). The side-chain of Leu60 is packed into a
hydrophobic space composed of the side-chains of Leu64, Asn100, Phe103, and
Ile107 and the main chain of Arg56-Glu57. The Oε1 and Oε2 of Glu61 respectively
are hydrogen bonded to Arg56Nε and Asn78Nδ2 (Fig. 5). These described
interactions may fix and stabilize the conformations of the loop and helix; i.e., the
HV region.
This HV region is comprised of 15 amino acid residues, ten of which, Pro49,
Ile50, Lys51, Ile53, Lys55, Glu57, Lys58, Leu59, His62, and Gln63 are widely exposed to
solvent. The side-chains of Ile53, Lys55, Glu57, Lys58, Leu59 and His62, in particular,
prominently extend into the solvent (Fig. 5). Given that the HV region is in
contact with the pollen factor(s), these residues may have importance in pollen
by guest on March 30, 2018
http://ww
w.jbc.org/
Dow
nloaded from

15
factor binding.
Positively Selected Regions— Recognition sites in some proteins (e.g., the
major histocompatibility complex (MHC) [47-49], antigenic surface proteins of
parasites and viruses (49), and acrosomal proteins of the abalone [50,51]) are
reported to be regions in which the number of nonsynonymous nucleotide
substitutions (dN)1 exceeds that of synonymous substitutions (dS)1, and positive
selection probably takes place in these regions. Window analysis of the dS and
dN in rosaceous S-RNases detected four regions with an excess of dN over dS,
in which positive selection may operate (PS1-PS4) (Fig.4) (52). PS1 nearly
duplicates the HV region. It is interesting that the four positively selected (PS)1
regions were detected by window analysis of the rosaceous S-RNase genes but
that there is only a single region with an allele-specific amino acid sequence.
PS1 (HV) and PS2, as well as PS3 and PS4 respectively are close in the three-
dimensional structure of S3-RNase, on either side of the active site cleft
(including the P1, B1, and B2 sites) and accessible to solvent (Figs. 1c and 4b).
Sugar Chains— Pyrus pyrifolia S3-RNase has two potential N-glycosylation
sites, Asn18 and Asn116 (Fig. 1a). The Asn18 site is specific to the S3- and S5-
RNases, whereas the Asn116 site is conserved among all the resaceous S-
RNases (Fig. 4a). A two-dimensional sugar map and mass spectrometry of
fragmented peptides showed that both sites have heterogeneous N-glycans,
Asn18 mainly glycosylated by a chitobiose (GlcNAcβ1-4GlcNAc) and Asn116
primarily occupied by a xylomannose type sugar chain (19). The core structure,
by guest on March 30, 2018
http://ww
w.jbc.org/
Dow
nloaded from

16
Manβ1-4GlcNAcβ1-4GlcNAc, of the xylomannose type sugar chain at Asn116,
clearly visible in the difference Fourier map, was modeled to fit the densities
(data not shown). No clear electron densities corresponding to the chitobiose
at Asn18 and the mannose and xylose moieties of the sugar chain at Asn116 were
observed, evidence that their conformations are highly disordered. As Asn18
and Asn116 respectively are located at the end of and on the opposite side of the
active site cleft (Figs.1a and 1c), the S3-RNase sugar chains probably are not
involved in its enzymatic properties and the recognition of self and non-self.
by guest on March 30, 2018
http://ww
w.jbc.org/
Dow
nloaded from

17
DISCUSSION
The structure of the active site (the catalytic P1 site and substrate binding B1
and B2 sites) of S3-RNase is typical of the structures of the RNase T2 family
enzymes. Probably, His33 and His88 function as general acid and base catalysts,
and Glu84 and Lys87 stabilize the pentacovalent intermediate in the transition
state. One marked difference between the P1 sites of S3-RNase and RNase Rh
is that the imidazole ring of His33 is rotated about 90° from that of His(46) (Fig. 3a),
even though the distance between His33Nε2 and His88Nε2 (7.15 Å) is nearly equal
to that between His(46)Nε2 and His(109)Nε2 (6.71 Å) (32). As S3-RNase has
ribonuclease activity (20), rotation of the ring of His33 is not crucial for its catalytic
activity. Another difference is that the His(104) of RNase Rh, which is expected to
bind to the phosphate group of the substrate RNA (32,41), is replaced by Lys83 in
S3-RNase (Fig. 3a). As the ε-amino group of Lys83 extends outside the P1 site
and is about 8 Å from the imidazole ring of His(104), it is unlikely that it interacts
with the phosphate group of the substrate unless a large induced fit occurs on
the side-chain of Lys83 during catalysis. A more detailed X-ray crystallographic
study of the S3-RNase complex with nucleotides is required to clarify how the
rotation of the imidazole ring of His33 and substitution of His(104) for Lys83 affect the
ribonuclease activity of S3-RNase.
Although it has not been shown experimentally what the in vivo substrate of
S-RNase during the GSI reaction is, it must be pollen (tube) rRNA according to
the receptor and inhibitor models (10). As the overall shape of the active site of
by guest on March 30, 2018
http://ww
w.jbc.org/
Dow
nloaded from

18
S3-RNase is very similar to the active sites of the other RNase T2 family enzymes
(except for a few amino acid substitutions), S3-RNase is not likely to have a strict
substrate specificity corresponding to the S-alleles. Actually, S3-RNase can
hydrolyze dinucleotide monophosphate nonspecifically (data not shown),
therefore it is reasonable that S-allele specificity in the inhibition of pollen tube
growth is not expressed due to the enzyme’s restricted substrate specificity but
to its interaction with the pollen S-gene product(s); consistent with either the
receptor or inhibitor model.
Domain swapping experiments proved that in the solanaceous S-RNases,
two hypervariable regions, HVa and HVb, essentially are responsible for S-
allele-specific pollen recognition (13,14). Only one hypervariable (HV) region
appears to be present in the rosaceous S-RNases, and it most likely is a
recognition site for the pollen factor(s) (Fig.4a). As compared with the
sequences of the solanaceous S-RNases (16,17), the HVa and HVb regions
correspond respectively to the HV region and vicinity of the α3-helix (Arg74-Lys76),
tentatively called the HVb’, of S3-RNase. As HVb’ is much shorter than HVb, it
has not been identified as an allele-specific sequence (Fig.4a). The three-
dimensional structure around the α3-helix therefore also is assumed to differ in
the solanaceous and rosaceous S-RNases.
The HV region of S3-RNase is located on the molecule’s surface, easily
accessible to external molecules such as the pollen factor(s). As shown in Fig.
1b, the main chain frameworks of RNase T2 family enzymes, including S3-RNase,
by guest on March 30, 2018
http://ww
w.jbc.org/
Dow
nloaded from

19
superimpose well on one another; in particular their core structures composed of
three α-helices (α4, α6 and α7) and four β-strands (β1, β2, β4 and β5). The
backbone structures, however, are notably very variable in what corresponds to
the HV region (Fig. 1b and 4b) even though it is composed of the same
secondary elements, a loop and an α-helix, as in all the RNases (Fig. 2). This
suggests that the HV region of S3-RNase has an allele-specific conformation.
Five basic amino acids, Lys51, Arg54, Lys55, Arg56, and Lys58 and two acidic
amino acids, Glu57 and Glu61, are present in the HV region of S3-RNase (Fig. 4a).
Although they seem to form a positively or negatively charged cluster on the
molecule’s surface, no such cluster was clearly detected because the side
chains of the basic or acidic amino acids are oriented in different directions (Figs.
5 and 6). As the δ-guanidino groups of Arg54 and Arg56 are neutralized by the
formation of hydrogen bonds, the surface of HV carries a weak rather than
strong positive charge (Figs. 5 and 6). If the other six Pyrus pyrifplia S-RNases
(S1, S2, and S4~S7) have the same main chain framework as S3-RNase, their HV
regions also would similarly have a weak positive charge, which might be
important for binding to the pollen factor(s) (Figs. 4 and 5).
PS regions with an excess of dN over dS, like the HV region a candidate for
the binding site to the pollen factor(s), are located on either side of the active site
cleft (Fig. 4b). Although there is no marked charge cluster in any of these
regions, PS1 (HV), PS2, and PS3 are hydrophilic and weakly basic (Fig. 6). In
contrast, PS4 is neutral and hydrophobic in spite of its exposure to solvent.
by guest on March 30, 2018
http://ww
w.jbc.org/
Dow
nloaded from

20
Why the rosaceous S-RNase has been positively selected on such a wide area
of its molecular surface is not clear, but location topology of the PS regions
suggests that S3-RNase simultaneously interacts with multiple pollen factors and
the substrate RNA.
The amino acid sequence identity between the Pyrus pyrifolia S3- and S5-
RNases is 95.5%, and there are only nine substitutions in their 200 amino acid
residues, all concentrated on the N-terminal half of S3-RNase (Fig. 4a). Two of
the nine substitutions, Lys(Arg)51 and His(Pro)62, are the most likely to contribute
to recognition between the two alleles because they are located on the
molecular surface of the HV region (amino acids in parentheses are those of S5-
RNase) (Fig. 4b). The side-chain of Lys(Arg)51 is fully exposed to solvent and
therefore has no interaction with the other residues (Fig. 5). In contrast,
His(Pro)62 are close to Glu(Ala)65 in the three-dimensional structure although at
separate locations in the primary structure. His62Nε2 forms a water-mediated
hydrogen bond with Glu65Oε1 and both are appreciably exposed to solvent (Fig.
5). Two possibilities are suggested as to how the pollen factor(s) discriminates
S3-RNase from S5-RNase. The pollen factor(s) must come in contact with Lys51,
the site consisting of His62 and Glu65, or both in order to recognize the difference
between the S3- and S5-RNases. Or, substitution of His62 for Pro produces a
large conformational change in the α2-helix and the pollen factor(s) recognize
such a conformational change rather than the amino acid substitutions. In
general, substitution for proline tends to bend the helix structure (53). An X-ray
by guest on March 30, 2018
http://ww
w.jbc.org/
Dow
nloaded from

21
crystallographic study of Pyrus pyrifolia S5-RNase is in progress in our laboratory.
If the three-dimensional structure of S5-RNase can be determined, a detailed
structural comparison can be made of the two S-Rnases which should shed light
on the recognition mechanism that operates between S3- and S5-alleles.
by guest on March 30, 2018
http://ww
w.jbc.org/
Dow
nloaded from

22
Acknowledgments
We thank Prof. T. Tsukihara of the Institute for Protein Research, Osaka
University for his helpful advice on the crystallization and the X-ray
crystallographic analysis. We also thank K. Miura of SPring-8, Hyogo and N.
Igarashi of the Photon Factory, Ibaragi, for their useful advice on the data
collection.
by guest on March 30, 2018
http://ww
w.jbc.org/
Dow
nloaded from

23
References
1. Newbigin E., Anderson, M.A., and Clarke, A.E. (1993) Plant Cell 5, 1315-
1324
2. de Nettancourt, D. (2001) Incompatibility and Incongruity in Wild and
Cultivated Plants, 2nd Ed., Springer, Heidelberg, Germany
3. Kawata, Y., Sakiyama, F., Hayashi, F., and Kyogoku, Y. (1990) Eur. J.
Biochem. 187, 255-262
4. McClure, B.A., Gray, J.E., Anderson, M.A., and Clarke, A.E. (1990) Nature
347, 757-760
5. Lee, H.S., Huang, S., and Kao T.-H. (1994) Nature 367, 560-563
6. Murfett, J., Atherton, T.L., Beiquan, M., Gasser, C.S., and McClure, B.A.
(1994) Nature 367, 563-566
7. Huang, S., Lee. H.S., Karunanaadaa, B., and Kao, T.-H. (1994) Plant Cell 6,
1021-1028
8. Karunanandaa, B., Huang, S., and Kao, T.-H. (1994) Plant Cell 6, 1933-
1940
9. McCubbin, A.G., Chung, Y.-Y., and Kao, T.-H. (1997) Plant Cell 9, 85-95
10. Dodds, P.N., Clarke, A.E., and Newbigin, E. (1996) Cell 85, 141-144
11. Luu, D.-T., Qin, X., Morse, D., and Cappadocia, M. (2000) Nature 407, 649-
651
12. Kao, T.H. and McCubbin, A.G. (1996) Proc. Natl. Acad. Sci. USA 93, 12059-
12065
by guest on March 30, 2018
http://ww
w.jbc.org/
Dow
nloaded from

24
13. Matton, D.P., Maes, O., Laublin, G., Xike, Q., Bertrand, C., Morse,
D., and Cappadocia, M. (1997). Plant Cell 9, 1757-1766
14. Matton, D.P., Luu, D.T., Xike, Q., Laublin, G., O’Brien, M., Maes, O.,
Morse, D., and Cappadocia, M. (1999). Plant Cell 11, 2087-2097
15. Ishimizu, T., Shinkawa, T., Sakiyama, F., and Norioka, S. (1998) Plant Mol.
Biol. 37, 931-941
16. Norioka, N., Norioka, S., Ohnishi, T., Ishimizu, T., Oneyama, C., Nakanishi,
T., and Sakiyama, F. (1996) J. Biochem. 120, 335-345
17. Richman, A.D., Broothaerts, W., and Kohn, J.R. (1997) American Journal of
Botany 87, 912-917
18. Ishimizu, T., Sato, Y., Saito, T., Yoshimura, Y., Norioka, S., Nakanshi, T., and
Sakiyama, F. (1996) J. Biochem. 120, 326-334
19. Ishimizu, T., Mitsukami, Y., Miyagi, M., Shinkawa, T., Natsuka, S., Hase, S.,
Sakiyama, F., and Norioka S. (1999) Eur. J. Biochem. 263, 624-634
20. Matsuura, T., Unno, M., Sakai, H., Tsukihara, T., and Norioka S. (2001) Acta
Crystallogr. Sect. D 57, 172-173
21. Otwinowski, Z. and Minor, W. (1997) Methods Enzymol. 276, 307-326
22. Leslie, A. G. W. (1992) in Joint CCP4 and ESF_EAMBC Newsletter on
Protein Crystallography, 26
23. Collaborative Computational Project 4. (1994) Acta Crystallogr. D50, 760-
763
24. Terwilliger, T. C. and Berendzen, J. (1999) Acta Crystallogr. D55, 849-861
by guest on March 30, 2018
http://ww
w.jbc.org/
Dow
nloaded from

25
25. de La Fortelle, E. and Bricogne, G. (1997) Methods Enzymol. 276, 472-493
26. Perrakis, A., Sixma, T.K., Wilson, K.S., and Lamzin, V.S. (1997) Acta
Crystallogr. D53, 448-455
27. Jones, T.A., Zou, J.Y., Cowan, S.W., and Kjeldgaard, M. (1991) Acta
Crystallogr. D47, 110-119
28. Brunger, A.T., Adams, P.D., Clore, G.M., DeLano, W.L., Gros, P., Grosse-
Kunstleve, R.W., Jiang, J.S., Kuszewski, J., Nilges, M., Pannu, N.S., Read,
R.J., Rice, L.M., Simonson, T., and Warren, G.L. (1998) Acta Crystallogr.
Sect. D 54, 905-921
29. Kleywegt, G.J. and Jones, T.A. (1998) Acta Crystallogr. Sect. D 54, 1119-
1131
30. Laskowski, R. A., MacArthur, M. W., Moss, D. S., and Thornton, J. M. (1993)
J. Appl. Cryst. 26, 283-291
31. Kurihara, H., Mitsui, Y., Ohgi, K., Irie, M., Mizuno, H., and Nakamura, K.T.
(1992) FEBS Letters 306, 189-192
32. Kurihara, H., Nonaka, T., Mitsui, Y., Ohgi, K., Irie, M, and Nakamura, K.T.
(1996) J. Mol. Biol. 255, 310-320
33. Tanaka, N., Arai, J., Inokuchi, N., Koyama, T., Ohgi, K., Irie, M, and
Nakamura, K.T. (2000) J. Mol. Biol. 298, 859-873
34. Nakagawa, A., Tanaka, I., Sakai, R., Nakashima, T., Funatsu, G., and
Kimura, M. (1999) Biochim. Biophys. Acta 1433, 253-260
35. Ishimizu, T., Norioka, S., Kanai, M., Clarke, A.E., and Sakiyama, F. (1996)
by guest on March 30, 2018
http://ww
w.jbc.org/
Dow
nloaded from

26
Eur. J. Biochem. 242, 627-635
36. Oxley, D. and Bacic, A. (1996) Eur. J. Biochem. 242, 75-80
37. Sanda, A. and Irie, M. (1980) J. Biochem. 87, 1079-1087
38. Sanda, A., Takizawa, Y., and Irie, M. (1985) Chem. Pharm. Bull. 33, 4515-
4521
39. Sanda, A., Takizawa, Y., Iwama, M., and Irie, M. (1985) J. Biochem. 98,
125-132
40. Ohgi, K., Horiuchi, H., Watanabe, H., Takagi, M., Yano, K., and Irie, M.
(1991) J. Biochem. 109, 776-785
41. Ohgi, K., Horiuchi, H., Watanabe, H., Iwama, M., Takagi, M., and Irie, M.
(1992) J. Biochem. 112, 132-138
42. Ohgi, K., Horiuchi, H., Watanabe, H., Iwama, M., Takagi, M., and Irie, M.
(1993) J. Biochem. 113, 219-224
43. Ohgi, K., Iwama, M., Tada, K., Takizawa, R., and Irie, M. (1995) J. Biochem.
117, 27-33
44. Ohgi, K., Iwama, M., Ogawa, Y., Hagiwara, C., Ono, E., Kawaguchi, R.,
Kanazawa, C., and Irie, M. (1996) Biol. Pharm. Bull. 19, 1080-1082
45. Ohgi, K., Shiratori, Y., Nakajima, A., Iwama, M., Kobayashi, H., Inokuchi, N.,
Koyama, T., Kock, M., Loffler, A., Glund, K., and Irie, M. (1997) Biosci.
Biotech. Biochem. 61, 432-438
46. Richards, F.M. and Wyckoff, H.W. (1971) in The Enzymes (Boyer, P.D., ed.),
vol.4, pp. 647-806, Academic Press, New York, USA
by guest on March 30, 2018
http://ww
w.jbc.org/
Dow
nloaded from

27
47. Hughes, A.L. and Nei, M. (1988) Nature 335, 167-170
48. Hughes, A.L. and Nei, M. (1989) Proc. Natl. Acad. Sci. USA 86, 958-962
49. Endo, T., Ikeo, K., and Gojobori, T. (1996) Mol. Biol. Evol. 13, 685-690
50. Swanson, W.J. and Vacquier, V.D. (1995) Proc. Natl. Acad. Sci. USA 92,
4957-4961
51. Lee, Y.-H., Ota, T., and Vacquier, V.D. (1995) Mol. Biol. Evol. 12, 231-238
52. Ishimizu, T., Endo, T., Yamaguchi-Kabata, Y., Nakamura, K.T., Sakiyama, F.,
and Norioka, S. (1998) FEBS letters 440, 337-342
53. Branden, C. and Tooze, J. (1999) Introduction to Protein Structure, 2nd Ed.,
Garland Publishing, New York, USA
54. Kraulis, P.J. (1991) J. Appl. Crystallogr 24, 946-950
55. Merrit, E.A. and Murphy, M.E.P. (1994) Acta Crystallogr. Sect. D 50, 869-873
56. Nicholls, A., Sharp, K. A., and Honig, B. (1991) Protein: Struct. Funct. Genet.
1, 281-296
57. Kabsch, W. and Sander, C. (1983) Biopolymers 22, 2577-2637
by guest on March 30, 2018
http://ww
w.jbc.org/
Dow
nloaded from

28
Footnotes
*This work was supported in part by a Grant-in-Aid for Special Research on
Priority Areas (B) (11238204) from the Ministry of Education, Culture, Sports,
Science and Technology, Japan.
The atomic coordinates and structure factors (code 1IQQ) have been deposited
in the Protein Data Bank, Research Collaboratory for Structural Bioinformatics,
Rutgers University, New Brunswick, NJ (http://www.rcsb.org/pdb).
1The abbreviations used are GSI, gametophytic self-incompatibility; HV,
hypervariable; PS, positively selected; dN, the number of nonsynonymous
nucleotide substitutions; dS, the number of synonymous nucleotide
substitutions.
by guest on March 30, 2018
http://ww
w.jbc.org/
Dow
nloaded from

29
Figure Legends
Figure 1. Stereo diagram showing the overall structure of Pyrus pyrifolia S3-
RNase. (a) Stereo ribbon diagram of S3-RNsae. N-Glycosylated asparagines,
N18 and N116, are shown as ball-and-stick models. (b) Stereo diagram
showing the superposition of RNase T2 family enzymes. S3-RNase is in red,
RNase Rh in cyan (32), RNase LE in yellow (33), and RNase MC1 in green (34).
(c) Stereo drawing of the space filling model of S3-RNase. Amino acid residues
that constitute the P1, B1, and B2 sites (46) respectively are colored red, blue,
and green. Orientations of the S3-RNase molecules in figures (a), (b) and (c)
are the same. Figures (a) and (b) were prepared by the programs MOLSCRIPT
(54) and RASTER3D (55), figure (c) by the program GRASP (56).
Figure 2. Sequence alignment of RNase T2 family enzymes and their
secondary structural elements. Amino acid residue numbering is based on that
for Pyrus pyrifolia S3-RNase. Amino acid residues that form secondary
structures are the α-helix (red), 310-helix (red, slanted), and β-strand (blue).
The secondary S3-RNase structures were defined by the program DSSP (57).
Figure 3. Superposition of the active sites of Pyrus pyrifolia S3-RNase and
RNase Rh. (a), Catalytic P1 site; (b), base-binding B1 site; (c), base-binding B2
site. Names of the P1, B1, and B2 sites are based on the nomenclature of the
by guest on March 30, 2018
http://ww
w.jbc.org/
Dow
nloaded from

30
subsites of RNase A (46). The amino acid residues of S3-RNase and RNase
Rh respectively are shown as ball-and-stick and wireframe constructions. The
figures were prepared by the program MOLSCRIPT (54).
Figure 4. Location of the HV and PS regions on the primary and tertiary
structures of Pyrus pyrifolia S3-RNase. (a) Sequence alignment of rosaceae S-
RNases; P. pyrifolia, Japanese pear; M. domestica, apple. HV and PS regions
are represented aligned as colored bars: HV (blue), HVb’ (black), PS1 (red), PS2
(yellow), PS3 (green), PS4 (purple). Amino acid residues that are the same as
those of S3-RNase are shaded, and positions of the amino acid substitutions
between S3- and S5-RNases are indicated by the symbol # over the alignment.
Numbering is the same that for S3-RNase. Potential N-glycosylation sites are in
red with white letters. (b) Location of HV and PS regions in the tertiary structure
of S3-RNase. These regions are in the same colors as in Figure 4(a); HV (blue),
PS1 (red), PS2 (yellow), PS3 (green), PS4 (purple). The substituted amino
acids in the S3- and S5-RNases are labeled and shown as ball-and-stick
constructions. The S3-RNase molecule orientation is the same as in Fig. 1.
This figure was prepared by the program GRASP (56).
Figure 5. Stereo diagram of the HV and PS2 regions of Pyrus pyrifolia S3-
RNase. The main chains of the HV and PS2 regions are shown as thick wires,
respectively in blue and yellow. A water molecule is indicated by a pink ball and
by guest on March 30, 2018
http://ww
w.jbc.org/
Dow
nloaded from

31
a hydrogen bond by a dotted pink line. This figure was prepared by the
program MOLSCRIPT (54).
Figure 6. Electrostatic surface potential of the HV and PS regions of Pyrus
pyrifolia S3-RNase. (a) Surfaces of the HV (PS1) and PS2 regions; (b) surfaces
of the PS3 and PS4 regions. The potential displayed represents a range of -10
to +10 kBT, red being negative and blue positive. This figure was prepared by
the program GRASP (56).
by guest on March 30, 2018
http://ww
w.jbc.org/
Dow
nloaded from

32
Table I
Data collection and refinement statistics for Pyrus pyrifolia S3-RNase
Rmerge = ∑|Iobs-<I>|/∑Iobs
Riso = ∑||Fdeliv|-|Fnative||/∑|Fnative|, Diffraction data were processed by DENZO &
SCALEPACK (20) (native) or MOSFLM (21) & SCALA (22) (derivative).
Rcullis = ∑||Fhobs|-|Fhcalc||/∑|Fhobs| for centric reflections, where Fhobs and Fhcalc are the
observed and calculated heavy atom structure factor amplitudes.
Phasing power = rms(|Fh|/E), where.Fh is the heavy atom structure factor amplitude,
and E the residual lack of closure error.
Figure of merit = |F(hkl)best|/|F(hkl) |
R-factor = ∑||Fobs|-|Fcalc||/∑|Fobs|
Diffraction data statisticsData set Native Hg1 Hg1’ Hg2 PbResolution (Å) 1.5 2.5 2.5 2.5 2.5Completeness (%) 96.3 99.7 100.0 98.2 99.7Unique reflection 33441 7483 7536 7450 7667Rmerge 5.7 8.5 10.1 5.2 11.5Riso 0.125 0.131 0.320 0.181Phasing statisticsRcullis 0.88 0.92 0.85 0.97Phasing power(Centrics/Acentrics)
0.97/0.96 1.18/1.45 1.64/1.68 1.04/0.9
Figure of merit(Centrics/Acentrics)
0.57/0.66
Refinement statisticsResolution range (Å) 500-1.5R-factor (%) 17.2Free- R-factor (%) 20.2Rms deviation from ideality Bond distance (Å) 0.022 Bond angles (º) 2.02
by guest on March 30, 2018
http://ww
w.jbc.org/
Dow
nloaded from

(b)
(c)
12 5 4
7
1
3
2
6
4
5
6
8
3
7
N116
N18
12 5 4
7
1
3
2
6
4
5
6
8
3
7
N116
N18
(a)
by guest on March 30, 2018
http://ww
w.jbc.org/
Dow
nloaded from

1 10 20 30 40
* * * * *
11
2S3 YDYFQFTQQYQLAVCNSNRT---LCKDPPDKLFTVHGLWPSNM
LE AKDFDFFYFVQQWPGSYCDTKQSCCYPTTGKPAADFGIHGLWPNNN
MC1 FDSFWFVQQWPPAVCSFQKS--GSCPGSGLRTFTIHGLWPQQ-
Rh SSCSSTALSCSNSANSDTCCSPEYGLVVLNMQWAPGYG-------------PDNAFTLHGLWPDKC
50 60 70 80 90
* * * * *
23
45
S3 VGPD-PSK-C-P-IKNI--RKREKL----LEHQLEIIWPNVFD-RTKNNLFWDKEWMKHGSCGYPT
LE DGTY-PSN-CDPNSPYD--QSQISD----LISSMQQNWPTLACPSGSGSTFWSHEWEKHGTCAESV
MC1 SGTSLT-N-C-PGSPFD--ITKISH----LQSQLNTLWPNVLR--ANNQQFWSHEWTKHGTCSEST
Rh SGAYAPSGGCDSNRASSSIASVIKSKDSSLYNSMLTYWPSNQG---NNNVFWSHEWSKHGTCVSTY
100 110 120 130 140 150
* * * * * *
67
38
S3 ID----------NENHYFETVIKMYISKKQNVSRILSKAKIEPDGKKRALLDIENAIRNGADNKKP
LE LT----------NQHAYFKKALDLKNQID--LLSILQGADIHPDGESYDLVNIRNAIKSAIG-YTP
MC1 F-----------NQAAYFKLAVDMRNNYD--IIGALRPHAAGPNGRTKSRQAIKGFLKAKFG-KFP
Rh DPDCYDNYEEGEDIVDYFQKAMDLRSQYN---VYKAFSSNGITPGGTYTATEMQSAIESYFG-AKA
160 170 180 190 200
* * * * *
45
67
S3
KLKCQKK--GTTTELVEITLCSDKSG---EHFIDCPHPFEPISPHYCPTNNIKY
LE
WIQCNVD-QSGNSQLYQVYICVDGSG---SSLIECPIF----PGGKCGTSIEFPTF
MC1
GLRCRTDPQTKVSYLVQVVACFAQDG---STLIDCTRD-------TCGANFIF
Rh
KIDCSS------GTLSDVALYFYVRGRDTYVITDALST------GSCSGDVEYPTK
by guest on March 30, 2018
http://ww
w.jbc.org/
Dow
nloaded from

1 10 20 30 40 50 60 70 80 90 * * * * * * * * * * # # # # # # # # #P. pyrifolia S3 YDYFQFTQQYQLAVCNSNRTLCKDPPDKLFTVHGLWPSNMVGPDPSKCPIKNIRK--REKLLEHQLEIIWPNVFDRTKNNLFWDKEWMKHGSCG-YPTIDNE S5 YDYFQFTQQYQLAVCNSNRTPCKDPPDKLFTVHGLWPSSMAGPDPSNCPIRNIRK--REKLLEPQLAIIWPNVFDRTKNKLFWDKEWMKHGTCG-YPTIDNE S1 YDYFQFTQQYQPAVCNSNPTPCNDPTDKLFTVHGLWPSNRNGPDPEKCKTTALNSQK-IGNMTAQLEIIWPNVLNRSDHVGFWEKEWIKHGTCG-YPTIKDD S2 ARYDYFQFTQQYQQAFCNSNPTPCKDPPDKLFTVHGLWPSTKVGRDPEYCKTKRYRK---IQRLEPQLEIIWPNVSDRKANRGFWRKQWYKHGSCA-SPALPNQ S4 FDYFQFTQQYQPAVCNSNPTPCNDPTDKLFTVHGLWPSNRNGPDPEKCKTTTMNSQK-IGNMTAQLEIIWPNVLNRSDHVGFWEREWLKHGTCG-YPTIKDD S6 YNYFQFTQQYQPAVCNSNPTPCKDPPDKLFTVHGLWPSNDVGDDPIYCKNKTIKSQQ-IGNLTAQLIIIWPNVLDRTDHVGFWNRQWNKHGSCGKAPTIKDE S7 YDYFQFTQQYQPAVCNSKPTPCKDPPDKLFTVHGLWPSNLNGPHPENCTNATVNPHR-IKNIQAQLKIIWPNVLDRTNHVGFWNKQWIKHGSCG-YPAIMNDM. domestica S2 YDYFQFTQQYQPAACNSNPTPCKDPPDKLFTVHGLWPSNMNRSELFNCSSSNVTYAK-IQNIRTQLEMIWPNVFNRKNHLGFWNREWNKHGACG-YPTIRND S3 VKFDYFQFTQQYQPAVCSSNPTPCKDPPDKLFTVHGLWPSNVNGSDPKKCKTTILNPQT-ITNLTAQLEIIWPNVLNRKAHARFWRKQWRKHGTCG-YPTIADD S4 FDYFQFTQQYQPAVCNFvPTPCKDPTDKLFTVHGLWPSNNVGGDPESCKIRNHRT--RAKALEPQLEIIWPNVLDRTNHTAFWRRQWIKHGTCG-YPTIQNE S7 YDYFQFTQQYQLAACNSKPIPCKDPPDKLFTVHGLWPSDSNGHDPVNCSKSTVDAQK-LGNLTTQLEIIWPNVYNRTDHISFWDKQWNKHGTCG-HPTIMND S9 YDYFQFTQQYQPAVCHFNPTPCRDPPDKLFTVHGLWPSNSSGNDPIYCKNTTMNSTK-IANLTARLEIIWPNVLDRTDHITFWNKQWNKHGSCG-HPAIQND
HV HVb' PS1 PS2
100 110 120 130 140 150 160 170 180 190 200 * * * * * * * * * * *
P. pyrifolia S3 NHYFETVIKMYISKKQNVSRILSKAKIEPDGKKRALLDIENAIRNGADNKKPKLKCQKK-GTTTELVEITLCSDKSGEHFIDCPHPFEPISPHYCPTNNIKY S5 NHYFETVIKMYISKKQNVSRILSKAKIEPDGKKRALLDIENAIRNGADNKKPKLKCQKK-GTTTELVEITLCSDKSGEHFIDCPHPFEPISPHYCPTNNIKY S1 MHYLQTVIRMYITQKQNVSAILSKAAIQPNGTNRPLVDIENAIRRGTNNTKPKFKCQKNTRTTTELVEVTLCSDRDLKKFINCPHGPPQGSRFSCPSS-VQY S2 KHYFETVIRMFLAEKQNVSRILSMATIEPEGKNRTLLEIQNAIRAGTTNMIPKLKCQKV-NGMTELVEVTLCHDSNLTQFINCPRPLPQASPYFCPIDDIQY S4 MHYLKTVIKMYITQKQNVSAILSKATIQPNGNNRSLVDIENAIRSGNNNTKPKFKCQKNTRTTTELVEVTLCSNRDLTKFINCPHGPPKGSRYFCPAN-VKY S6 MHYFKTVIKMYITQKQNVSEILSRAKIEPEGKIRRRDDIINAIRLGTKDKKPKLKCQKN-NQTTELVEITICSDRNLTQFIDCPRSSFKGSPFHCPTNHILY S7 THYFQTVINMYITQKQNVSEILSKAKIEPLGIQRPLVHIENAIRNSTNNKKPKFKCQKN-SGVTELVEVGLCSDGSLTQFRNCPHPPP-GSPYLCPAD-VKYM. domestica S2 LHYFQTVIKMYITQKQNVSDILSKAKIEPDGNIRTQKEIVDAIRKGIHGKEPNLKCQKN-TQMTELVEVTLCSDGNLKQFIDCPHHFPNGSRHNCPTNHILY S3 MHYFSTVIEMYITKKQNVSEILSKAKIKPEKKFRTRDDIVNAISQSIDYKKPKLKCKNN-NQITELVEVGLCSDNNLTQFIDCPRPFPQGSPFFCPTNNIQY S4 NDYFETVVKMYITEKQNVSKILSNAKIEPDGIKRTLADLEIAIRSGTDNKKPKFKCQKK-RRVTELVEVTLCSDKNRAHFIDCPNPFQPGSPYLCPTNNILY S7 IHYFQTVIKMYITQKQNVSKILSRAKIEPEGKPRKQVDIVNAIRKGTNDKEPKLKCQKN-NQVTELVEVTLCSNRNLTGFINCPRHIPNGSRYSCPTKNILY S9 MHYLQTVIKMYITQKQNVSEILSKAKIEPVGKFRTQKEIEKAIRKGTNNKEPKLKCQKN-SQRTELVEVTLCSDRNLNQFIDCPRPILNGSRYYCPTNNILY
PS3 PS4
(a)
(b)
by guest on March 30, 2018
http://ww
w.jbc.org/
Dow
nloaded from

HV
(P
S1)
PS
2
PS
4
PS
3
(a)
(b)
by guest on March 30, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Sakiyama and Shigemi NoriokaTakanori Matsuura, Hiroaki Sakai, Masaki Unno, Koh Ida, Mamoru Sato, Fumio
responsible for gametophytic self-incompatibilityCrystal structure at 1.5 A resolution of Pyrus pyrifolia pistil ribonuclease
published online September 27, 2001J. Biol. Chem.
10.1074/jbc.M107617200Access the most updated version of this article at doi:
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
by guest on March 30, 2018
http://ww
w.jbc.org/
Dow
nloaded from