Embed Size (px)
Citation preview

JOURNAL OF BACTERIOLOGY, JUlY 1973, p. 98-106Copyright i 1973 American Society for Microbiology
Vol. 115, No. 1Printed in U.S.A.
Uptake and Utilization of Glutamic Acid byCryptococcus albidus
S. L. TANG AND D. H. HOWARD
Department of Microbiology and Immunology, School of Medicine, University of California, Los Angeles,Califomia 90024
Received for publication 6 February 1973
Cryptococcus albidus utilizes glutamate as a sole carbon source. The kineticsof uptake of this amino acid were studied. L-Glutamic acid was taken up by twosaturable systems: a high affinity system with a Michaelis constant (Kin) of 1.15x 10-' M and a Vmax of 0.049 ,umol per mg per h and a low affinity system with aKm of 2.5 x 10-3 M and a Vmax of 3.61 umol per mg per h. Both systems possessedcharacteristics of active transport which were dependent on temperature and pHand which required metabolic energy. Uptake was inhibited at 37 C but thetemperature-sensitive step was reversible. Chemical fractionation of cells with5% trichloroacetic acid showed that glutamic acid initially entered a soluble poolwhich decreased after 1 h as the amino acid was incorporated into the protein andnucleic acid fractions of the yeast. Some of the glutamate was completelyoxidized and could be recovered as '4CO2. Therefore, the amino acid wasalso used as an energy source.
Cryptococcus albidus is one of several sapro-phytic encapsulated yeasts belonging to thegenus Cryptococcus (22). This yeast utilizesglutamic acid as a sole source of carbon in asynthetic medium originally devised by Litt-man (21) to study amino acid assimilation by C.neoformans, a pathogenic member of the genus.Since amino acid uptake by encapsulatedyeasts has not previously been explored, physio-logical studies on uptake and utilization ofglutamic acid by C. albidus were done and arethe subject of this report which was presented,in part, at the 72nd Annual Meeting of theAmerican Society for Microbiology, 23-28 April1972, Philadelphia, Pa.
MATERIALS AND METHODSOrganism. The strain of C. albidus used in this
study (no. 378, stock culture collection of this depart-ment) was originally isolated from soil and wasidentified by standard procedures (22). Stock culturesof the yeast were maintained on slants of peptoneglucose medium (6) at room temperature for 3 daysand subsequently refrigerated. Subcultures weremade at monthly intervals.Growth and preparatDn of the fungal cells. C.
albidus was grown in Littman's capsular broth (21)incubated at room temperature on a rotary actionshaker (model V, New Brunswick Scientific Co., NewBrunswick, N.J.) for 24 h. Cells were harvested bycentrifugation at 2,600 x g for 15 min at 4 C, washedonce, and suspended in distilled water. A calibration
curve relating turbidity and dry weight was preparedaccording to standard methods (8, 15). Throughoutthese studies the cell suspension contained 0.4 mg(dry weight) of Littman thiamine-basal salts me-dium (thiamine-BSM) per ml (21).
Preparation of starved cells. Twenty-four-houryeast cell suspensions were harvested by centrifuga-tion, washed, suspended in thiamine-BSM, and in-cubated at room temperature on a rotary shaker for 12h. The starved cells were harvested, washed, andadjusted to contain 0.4 mg (dry weight) of thiamine-BSM per ml for the experiments.Measurement of L-glutaMic acid uptake and
incorporation. Uniformly labeled L-glutamic acid-14C with a specific activity of 260 mCi/mmol (Amer-sham/Searle Corp., Des Plaines, Ill.) was used. Theradioactive amino acid was diluted with unlabeledL-glutamic acid (Calbiochem, Los Angeles, Calif.) toachieve the desired molar concentrations of aminoacid. The uptake of amino acid was measured by theprocedure of Gupta and Pramer (15) with the follow-ing modifications. Equal volumes of 2 x 10-6 ML-glutamic acid- 4C and 0.4 mg of cell suspension perml were placed in separate 50-ml flasks in a waterbath shaker (New Brunswick, N. J.) at 23 C for 15min. After temperature equilibration, the glutamicacid solution was added to the cell suspension and theflask was shaken throughout the experiment. A 1-mlsample was withdrawn at determined intervals andfiltered through a 0.45-,um membrane filter and amicrofilter glass disk prefilter(AP 20 01500, MilliporeCorp., Bedford, Mass.). Five-tenths milliliter of eachfiltrate was then analyzed for radioactivity. Thedecrease in radioactivity of the filtrate at various time
38
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
b on
01
Dec
embe
r 20
21 b
y 11
8.15
8.18
8.12
2.
GLUTAMIC ACID UPTAKE BY C. ALBIDUS
intervals represented uptake of L-glutamic acid bycells (14, 15).The incorporation of 14C activity was measured by
the procedure of Gupta and Pramer (16). Briefly, 5-mlsamples of the reaction mixture described in thepreceding paragraph were withdrawn at determinedtime intervals and mixed with equal volumes of cold10% trichloroacetic acid for 30 min. The preparationwas then centrifuged at 2,600 x g for 10 min and thesupernatant fluid was tested for radioactivity. Theactivity of the cold trichloroacetic acid-soluble frac-tion was computed by subtracting the "IC activity offiltrate collected before the acid treatment from the"IC activity of the supematant fluid collected aftercold acid treatment. The pellet was extracted with 5ml of 5% trichloroacetic acid for 30 min at 90 C, andthe hot acid-soluble fraction was separated by centrif-ugation and analyzed for radioactivity (hot trichloroa-cetic acid-soluble fraction). The amount of hot tri-chloroacetic acid-insoluble material was calculatedby subtracting the "IC activity removed by cold andhot acid extraction from total "4C activity removedfrom the medium by the cells.Measurement of radioactivity. Five-tenths milli-
liter of each sample (filtrate or extract) was placed invials with 10 ml of scintillation fluid and counted in aliquid scintillation counter (Nuclear-Chicago, ModelMark I, Des Plaines, Ill.). The scintillation fluidcontained the following chemicals per liter of p-diox-ane: naphthalene, 70 g; 2, 5-diphenyloxazole, 7 g; and1, 4-bis-2-(5-phenyl oxazolyl)-benzene, 0.05 g. Thevials were counted for 10 min and the counts werecorrected for background.
Collection and measurement ofCO2 derived fromL-glutamic acid. The production by C. albidus 14CO2from uniformly labeled glutamic acid was measuredby the method of Gupta and Pramer (16). A total of80ml of cell suspension was placed in a 125-ml Erlen-meyer flask and treated with a final concentration of10-6 M L-glutamic acid. DL-glutamic acid-1-"4C (spact, 11.5 mCi/mmol), DL-glutamic acid-3,4-"4C (spact, 14.24 mCi/mmol), DL-glutamic acid-2-"4C (sp act,1.5 mCi/mmol), and DL-glutamic acid-5-"4C (sp act,15 mCi/mmol) were purchased from InternationalChemical and Nuclear Corp., Irvine, Calif.
RESULTSTime course of L-glutamic acid uptake. The
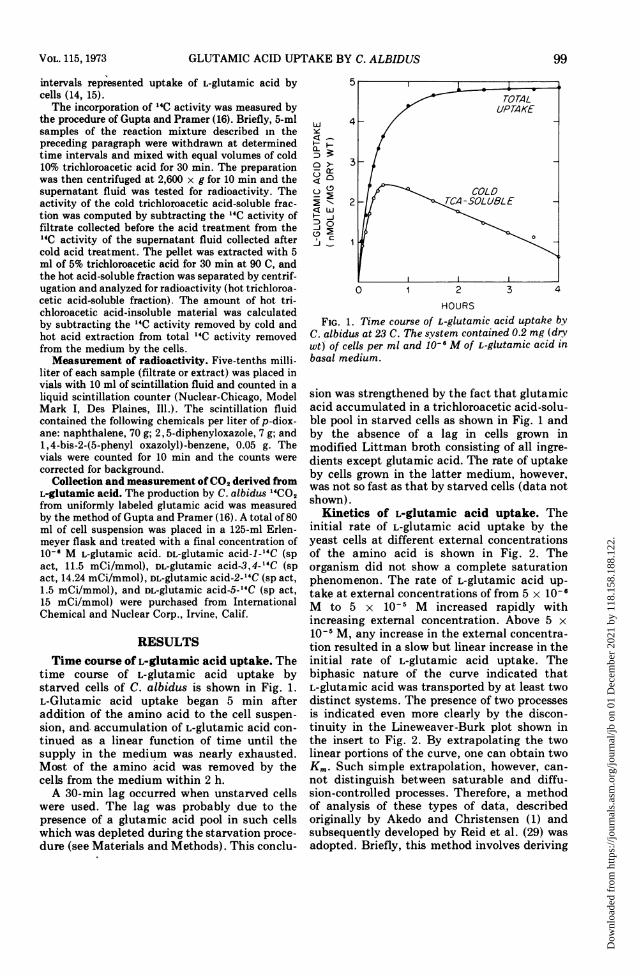
time course of L-glutamic acid uptake bystarved cells of C. albidus is shown in Fig. 1.L-Glutamic acid uptake began 5 min afteraddition of the amino acid to the cell suspen-sion, and. accumulation of L-glutamic acid con-tinued as a linear function of time until thesupply in the medium was nearly exhausted.Most of the amino acid was removed by thecells from the medium within 2 h.A 30-min lag occurred when unstarved cells
were used. The lag was probably due to thepresence of a glutamic acid pool in such cellswhich was depleted during the starvation proce-dure (see Materials and Methods). This conclu-
0- 3
'so
~-2 TCA -SQLULSE
<-LJIL? y' ? 0
0 1 2 3 4
HOURS
FIG. 1. Time course of L-glutamic acid uptake byC. albidus at 23 C. The system contained 0.2 mg (drywt) of cells per ml and 10-6 M of L-glutamic acid inbasal medium.
sion was strengthened by the fact that glutamicacid accumulated in a trichloroacetic acid-solu-ble pool in starved cells as shown in Fig. 1 andby the absence of a lag in cells grown inmodified Littman broth consisting of all ingre-dients except glutamic acid. The rate of uptakeby cells grown in the latter medium, however,was not so fast as that by starved cells (data notshown).
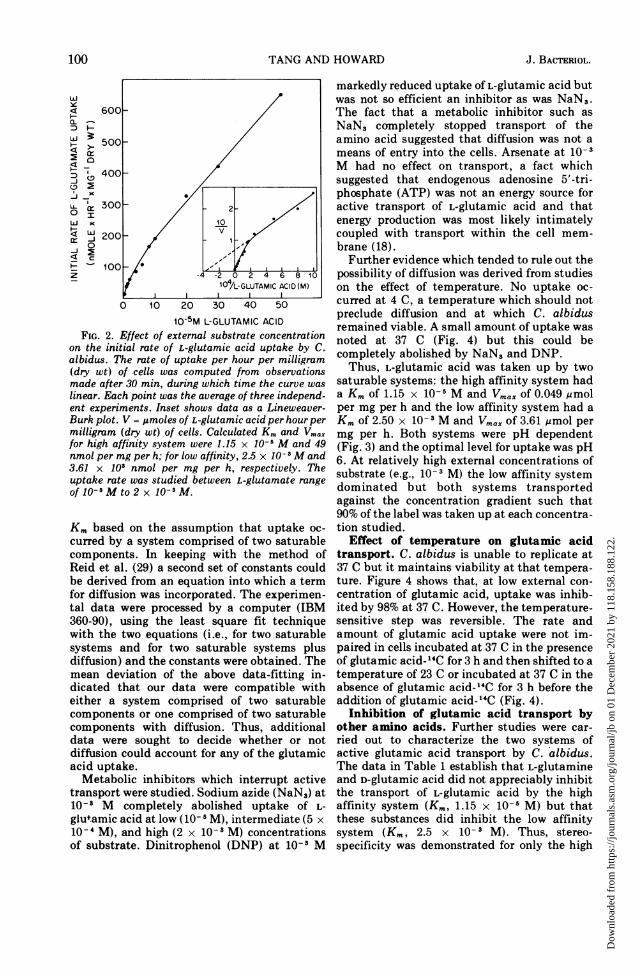
Kinetics of L-glutamic acid uptake. Theinitial rate of L-glutamic acid uptake by theyeast cells at different external concentrationsof the amino acid is shown in Fig. 2. Theorganism did not show a complete saturationphenomenon. The rate of L-glutamic acid up-take at external concentrations of from 5 x 10-6M to 5 x 10- 5 M increased rapidly withincreasing external concentration. Above 5 x10-5 M, any increase in the external concentra-tion resulted in a slow but linear increase in theinitial rate of L-glutamic acid uptake. Thebiphasic nature of the curve indicated thatL-glutamic acid was transported by at least twodistinct systems. The presence of two processesis indicated even more clearly by the discon-tinuity in the Lineweaver-Burk plot shown inthe insert to Fig. 2. By extrapolating the twolinear portions of the curve, one can obtain twoKm. Such simple extrapolation, however, can-not distinguish between saturable and diffu-sion-controlled processes. Therefore, a methodof analysis of these types of data, describedoriginally by Akedo and Christensen (1) andsubsequently developed by Reid et al. (29) wasadopted. Briefly, this method involves deriving
VOL. 115, 1973 99
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
b on
01
Dec
embe
r 20
21 b
y 11
8.15
8.18
8.12
2.
TANG AND HOWARD
Li
< 600/
LU~500-
400-
u- 300 2-
( 104/-uA l CD(
0
0 10 20 30 40 50
10-5M L-GLUTAMIC ACID
FIG. 2. Effect of external substrate concentrationon the initial rate of L-glutamic acid uptake by C.albidus. The rate of uptake per hour per milligram(dry wt) of cells was computed from observationsmade after 30 min, during which time the curve was
linear. Each point was the average of three independ-ent experiments. Inset shows data as a Lineweaver-Burk plot. V = ,umoles of L-glutamic acid per hour permilligram (dry wt),of cells. Calculated Km and Vmaxfor high affinity system were 1.15 x 10-5 M and 49nmol per mg per h; for low affinity, 2.5 x 10-3 M and3.61 x 103 nmol per mg per h, respectively. Theuptake rate was studied between L-glutamate rangeof 10- M to 2 x 10-3M.
Km based on the assumption that uptake oc-curred by a system comprised of two saturablecomponents. In keeping with the method ofReid et al. (29) a second set of constants couldbe derived from an equation into which a termfor diffusion was incorporated. The experimen-tal data were processed by a computer (IBM360-90), using the least square fit techniquewith the two equations (i.e., for two saturablesystems and for two saturable systems plusdiffusion) and the constants were obtained. Themean deviation of the above data-fitting in-dicated that our data were compatible witheither a system comprised of two saturablecomponents or one comprised of two saturablecomponents with diffusion. Thus, additionaldata were sought to decide whether or notdiffusion could account for any of the glutamicacid uptake.Metabolic inhibitors which interrupt active
transport were studied. Sodium azide (NaN3) at10-3 M completely abolished uptake of L-glutamic acid at low (10-5 M), intermediate (5 x
10-' M), and high (2 x 10-' M) concentrationsof substrate. Dinitrophenol (DNP) at 10-' M
markedly reduced uptake of L-glutamic acid butwas not so efficient an inhibitor as was NaN,.The fact that a metabolic inhibitor such asNaN3 completely stopped transport of theamino acid suggested that diffusion was not ameans of entry into the cells. Arsenate at 10-3M had no effect on transport, a fact whichsuggested that endogenous adenosine 5'-tri-phosphate (ATP) was not an energy source foractive transport of L-glutamic acid and thatenergy production was most likely intimatelycoupled with transport within the cell mem-brane (18).Further evidence which tended to rule out the
possibility of diffusion was derived from studieson the effect of temperature. No uptake oc-curred at 4 C, a temperature which should notpreclude diffusion and at which C. albidusremained viable. A small amount of uptake wasnoted at 37 C (Fig. 4) but this could becompletely abolished by NaN3 and DNP.Thus, L-glutamic acid was taken up by two
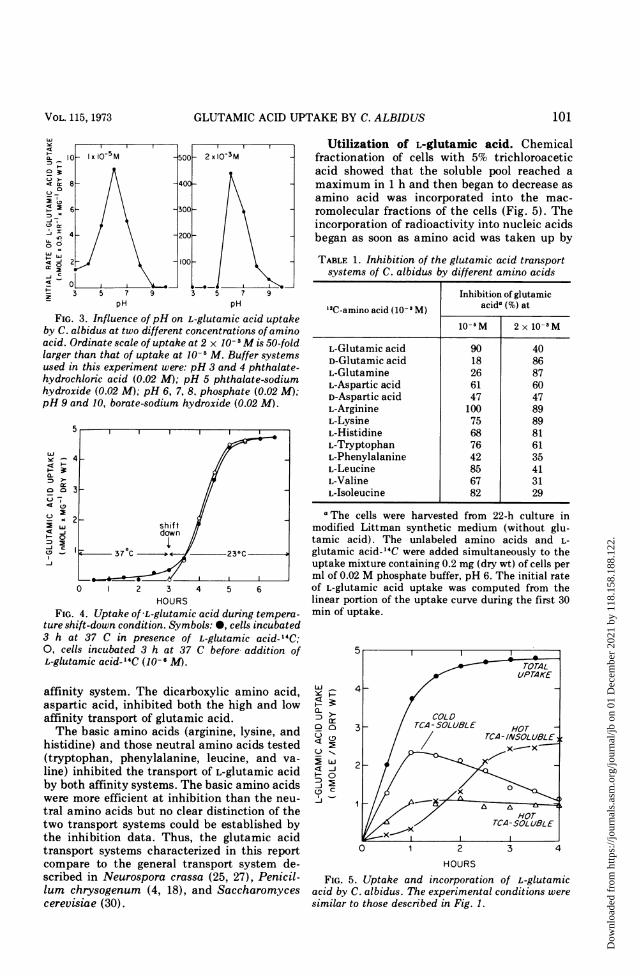
saturable systems: the high affinity system hada Km of 1.15 x 10-s M and Vmax of 0.049 ,molper mg per h and the low affinity system had aKm of 2.50 x 10-i M and Vmax of 3.61 mmol permg per h. Both systems were pH dependent(Fig. 3) and the optimal level for uptake was pH6. At relatively high external concentrations ofsubstrate (e.g., 10-3 M) the low affinity systemdominated but both systems transportedagainst the concentration gradient such that90% of the label was taken up at each concentra-tion studied.
Effect of temperature on glutamic acidtransport. C. albidus is unable to replicate at37 C but it maintains viability at that tempera-ture. Figure 4 shows that, at low external con-centration of glutamic acid, uptake was inhib-ited by 98% at 37 C. However, the temperature-sensitive step was reversible. The rate andamount of glutamic acid uptake were not im-paired in cells incubated at 37 C in the presenceof glutamic acid- 14C for 3 h and then shifted to atemperature of 23 C or incubated at 37 C in theabsence of glutamic acid-14C for 3 h before theaddition of glutamic acid-14C (Fig. 4).
Inhibition of glutamic acid transport byother amino acids. Further studies were car-ried out to characterize the two systems ofactive glutamic acid transport by C. albidus.The data in Table 1 establish that L-glutamineand D-glutamic acid did not appreciably inhibitthe transport of L-glutamic acid by the highaffinity system (K, 1.15 x 10-5 M) but thatthese substances did inhibit the low affinitysystem (Ki,, 2.5 x 10- 3 M). Thus, stereo-specificity was demonstrated for only the high
100 J. BACTERIOL.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
b on
01
Dec
embe
r 20
21 b
y 11
8.15
8.18
8.12
2.
GLUTAMIC ACID UPTAKE BY C. ALBIDUS
10 IXI0-5M 500- 2x10-3M
6 /300_
4- -200-
2 100
3 5 7 9 3 5 7 9pH pH
FIG. 3. Influence of pH on L-glutamic acid uptakeby C. albidus at two different concentrations of aminoacid. Ordinate scale of uptake at 2 x 10-3M is 50-foldlarger than that of uptake at 10-i M. Buffer systemsused in this experiment were: pH 3 and 4 phthalate-hydrochloric acid (0.02 M); pH 5 phthalate-sodiumhydroxide (0.02 M); pH 6, 7, 8, phosphate (0.02 M);pH 9 and 10, borate-sodium hydroxide (0.02 M).
4
0 2 3 4 5 6HOURS
FIG. 4. Uptake of L-glutamic acid during tempera-ture shift-down condition. Symbols: 0, cells incubated3 h at 37 C in presence of L-glutamic acid- 14C;0, cells incubated 3 h at 37 C before addition ofL-glutamic acid-'4C (10-6 M).
affinity system. The dicarboxylic amino acid,aspartic acid, inhibited both the high and lowaffinity transport of glutamic acid.The basic amino acids (arginine, lysine, and
histidine) and those neutral amino acids tested(tryptophan, phenylalanine, leucine, and va-line) inhibited the transport of L-glutamic acidby both affinity systems. The basic amino acidswere more efficient at inhibition than the neu-tral amino acids but no clear distinction of thetwo transport systems could be established bythe inhibition data. Thus, the glutamic acidtransport systems characterized in this reportcompare to the general transport system de-scribed in Neurospora crassa (25, 27), Penicil-lum chrysogenum (4, 18), and Saccharomycescerevisiae (30).
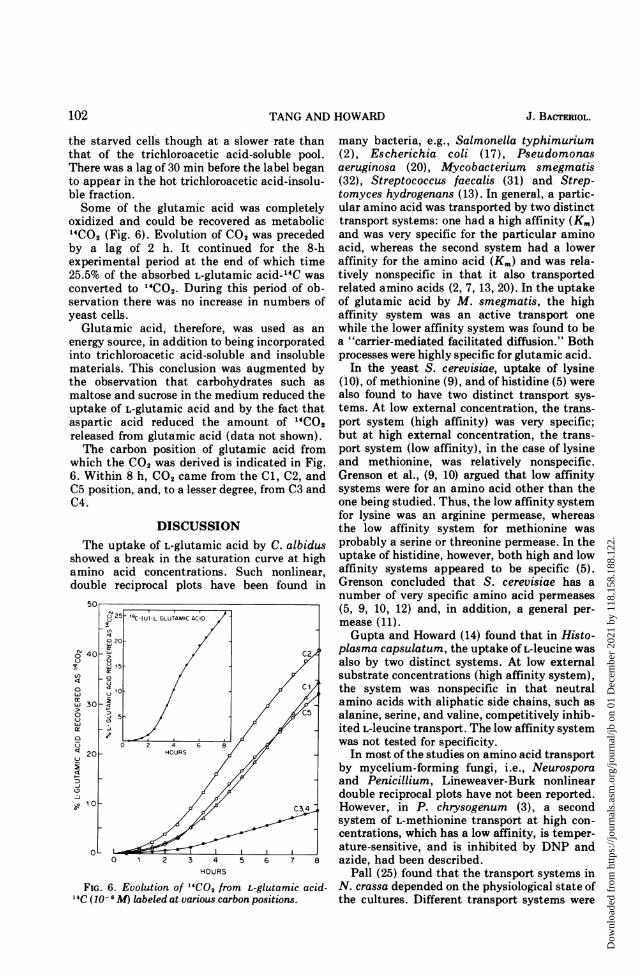
Utilization of L-glutamic acid. Chemicalfractionation of cells with 5% trichloroaceticacid showed that the soluble pool reached amaximum in 1 h and then began to decrease asamino acid was incorporated into the mac-romolecular fractions of the cells (Fig. 5). Theincorporation of radioactivity into nucleic acidsbegan as soon as amino acid was taken up by
TABLE 1. Inhibition of the glutamic acid transportsystems of C. albidus by different amino acids
Inhibition of glutamic'2C-amino acid (10-3 M) acida (%) at
10-'M 2 x 10-3M
L-Glutamic acid 90 40D-Glutamic acid 18 86L-Glutamine 26 87L-Aspartic acid 61 60D-Aspartic acid 47 47L-Arginine 100 89L-Lysine 75 89L-Histidine 68 81L-Tryptophan 76 61L-Phenylalanine 42 35L-Leucine 85 41L-Valine 67 31L-Isoleucine 82 29
aThe cells were harvested from 22-h culture inmodified Littman synthetic medium (without glu-tamic acid). The unlabeled amino acids and L-glutamic acid-14C were added simultaneously to theuptake mixture containing 0.2 mg (dry wt) of cells perml of 0.02 M phosphate buffer, pH 6. The initial rateof L-glutamic acid uptake was computed from thelinear portion of the uptake curve during the first 30min of uptake.
5 § ;
Lii~ ~4JTA
a-F 4 _/4_
C] 3 TC-SOLUBLE HOT
TCA-INSOLaUL BE
ui 2
oc
HOURS
FIG. 5. Uptake and incorporation of L-glutamicacid by C. albidus. The experimental conditions were
similar to those described in Fig. 1.
a.
n a:
<c_^a:
u I
O °
-
Z
I-a
3.I0
2 c
<_jJ
shif tdown
101VOL. 115,1973
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
b on
01
Dec
embe
r 20
21 b
y 11
8.15
8.18
8.12
2.
TANG AND HOWARD
the starved cells though at a slower rate thanthat of the trichloroacetic acid-soluble pool.There was a lag of 30 min before the label beganto appear in the hot trichloroacetic acid-insolu-ble fraction.Some of the glutamic acid was completely
oxidized and could be recovered as metabolic14CO2 (Fig. 6). Evolution of CO2 was precededby a lag of 2 h. It continued for the 8-hexperimental period at the end of which time25.5% of the absorbed L-glutamic acid-14C wasconverted to 14CO2. During this period of ob-servation there was no increase in numbers ofyeast cells.
Glutamic acid, therefore, was used as anenergy source, in addition to being incorporatedinto trichloroacetic acid-soluble and insolublematerials. This conclusion was augmented bythe observation that carbohydrates such asmaltose and sucrose in the medium reduced theuptake of L-glutamic acid and by the fact thataspartic acid reduced the amount of 14CO2released from glutamic acid (data not shown).The carbon position of glutamic acid from
which the CO2 was derived is indicated in Fig.6. Within 8 h, CO2 came from the Cl, C2, andC5 position, and, to a lesser degree, from C3 andC4.
DISCUSSIONThe uptake of L-glutamic acid by C. albidus
showed a break in the saturation curve at highamino acid concentrations. Such nonlinear,double reciprocal plots have been found in
0 25 4C-(U)-L GLUTAMIC ACID
o20-c 40 -ui C2
o SDC>Li
U
o _j5 C5
0 2 4 6 820 -HOURS
0010 C3.4
0 1 2 3 4 5 6 7 8HOURS
FIG. 6. Evolution of 14CO, from L-glutamic acid-4C (10-6 AlM labeled at various carbon positions.
many bacteria, e.g., Salmonella typhimurium(2), Escherichia coli (17), Pseudomonasaeruginosa (20), Mycobacterium smegmatis(32), Streptococcus faecalis (31) and Strep-tomyces hydrogenans (13). In general, a partic-ular amino acid was transported by two distincttransport systems: one had a high affinity (Ki)and was very specific for the particular aminoacid, whereas the second system had a loweraffinity for the amino acid (Ki) and was rela-tively nonspecific in that it also transportedrelated amino acids (2, 7, 13, 20). In the uptakeof glutamic acid by M. smegmatis, the highaffinity system was an active transport onewhile the lower affinity system was found to bea "carrier-mediated facilitated diffusion." Bothprocesses were highly specific for glutamic acid.
In the yeast S. cerevisiae, uptake of lysine(10), of methionine (9), and of histidine (5) werealso found to have two distinct transport sys-tems. At low external concentration, the trans-port system (high affinity) was very specific;but at high external concentration, the trans-port system (low affinity), in the case of lysineand methionine, was relatively nonspecific.Grenson et al., (9, 10) argued that low affinitysystems were for an amino acid other than theone being studied. Thus, the low affinity systemfor lysine was an arginine permease, whereasthe low affinity system for methionine wasprobably a serine or threonine permease. In theuptake of histidine, however, both high and lowaffinity systems appeared to be specific (5).Grenson concluded that S. cerevisiae has anumber of very specific amino acid permeases(5, 9, 10, 12) and, in addition, a general per-mease (11).Gupta and Howard (14) found that in Histo-
plasma capsulatum, the uptake of L-leucine wasalso by two distinct systems. At low externalsubstrate concentrations (high affinity system),the system was nonspecific in that neutralamino acids with aliphatic side chains, such asalanine, serine, and valine, competitively inhib-ited L-leucine transport. The low affinity systemwas not tested for specificity.
In most of the studies on amino acid transportby mycelium-forming fungi, i.e., Neurosporaand Penicillium, Lineweaver-Burk nonlineardouble reciprocal plots have not been reported.However, in P. chrysogenum (3), a secondsystem of L-methionine transport at high con-centrations, which has a low affinity, is temper-ature-sensitive, and is inhibited by DNP andazide, had been described.
Pall (25) found that the transport systems inN. crassa depended on the physiological state ofthe cultures. Different transport systems were
o L
102 J. BACTERIOL.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
b on
01
Dec
embe
r 20
21 b
y 11
8.15
8.18
8.12
2.
GLUTAMIC ACID UPTAKE BY C. ALBIDUS
r: DC14 c1
0~~~~~~~~~~~144'0 .0
o ~ ~ ~ ~ ~ C)C) C.
>~~~~~a).-~ ~ ~ ~ ~ ~~Lrj~~~~~~~~~~~~~.
00
16.,-4 Q U).- a Q m = 0 .<
C: <: ao 00o 00 00 00_- _4 _-
~~~~~~~~~~0~~ ~ ~ ~ ~ ~ 6a) a
,>a," 0 ) a
-6j. ", . QbD 5
(L)~~~~~~~~~~~~~~~~-w rA
boo
CZ)
-u ,
aL) 0 0 1*4+.) 0.w o
a):3
>u ) 0 "0 co
yy4) u ~~~~~~~.0
.4 ,3 0-:8)
-40 a)JLDt +IfA
a)->
0 c E-"5
.04i 6;
a0 0
-4<.- -4- Q
: e.
bD
¢)
103
o6.0 0
VOL. 115, 1973
%
U)
."
0
L.
C.)
0
0
a)
CC
00.
Cs
ca0
-0
.R
.a
a)
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
b on
01
Dec
embe
r 20
21 b
y 11
8.15
8.18
8.12
2.
TANG AND HOWARD
.= O
'20.-0.0A
lo'
-o ;.0 4)0 t0 &
.r. .
E j.,!C
4. V.0 o 5K = 4)
r. an0, .6-3,p v 9.-oi
Cr ci
4)
0U
*_ Co _:z > e. >0-
.0, _.A4 &- 0 CO)
4)
Q Z Q) c)
-U)a 4
X~~~~~ 4)
,eAx,: -o@
4) CZ
n .404~~~~~~~~4
_4 m- c9
a EE E E
_ to 0 0.2, CL) za C) C)
UZ;4)
ci t0&
'
0 S.4) 4
0. >
4 '. mEv EA
co
0 00
a
0 0.
0
J-
0 -
40
bb
~,
0 0
+4> 1 16
~00
0
CD0
0 p. 0
044) 0 l
- j
C) XbD
O-
4) 0
0
0U.Ooe; _ b
~ca ~HCI
104 J. BACTERIOL.
0
z~
0'I
.4c4
I)
V3
*0
4)4
0C')
cII0AH4
I
I
-
-
'r'I
F
00 oo
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
b on
01
Dec
embe
r 20
21 b
y 11
8.15
8.18
8.12
2.

GLUTAMIC ACID UPTAKE BY C. ALBIDUS
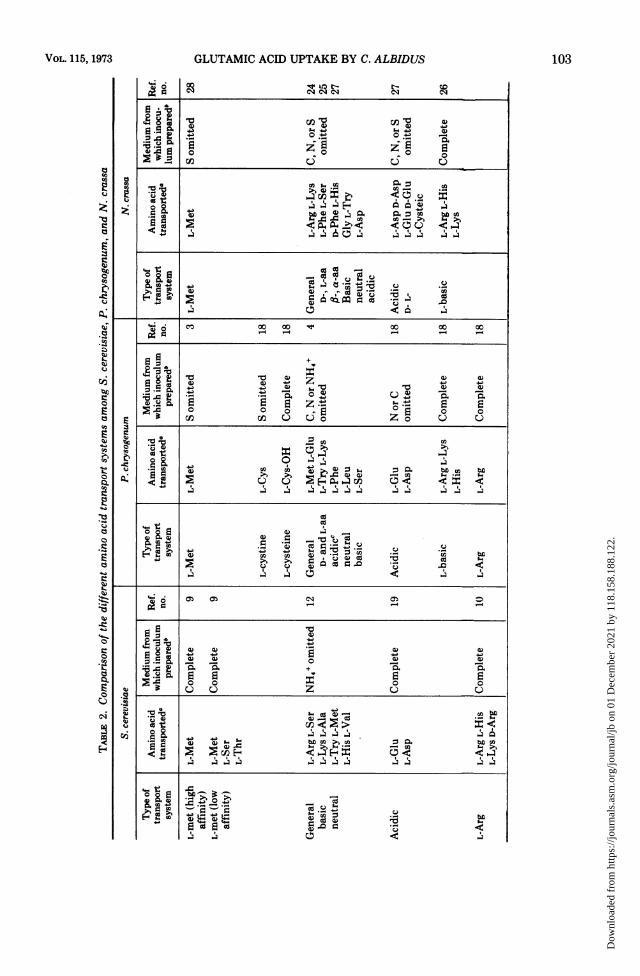
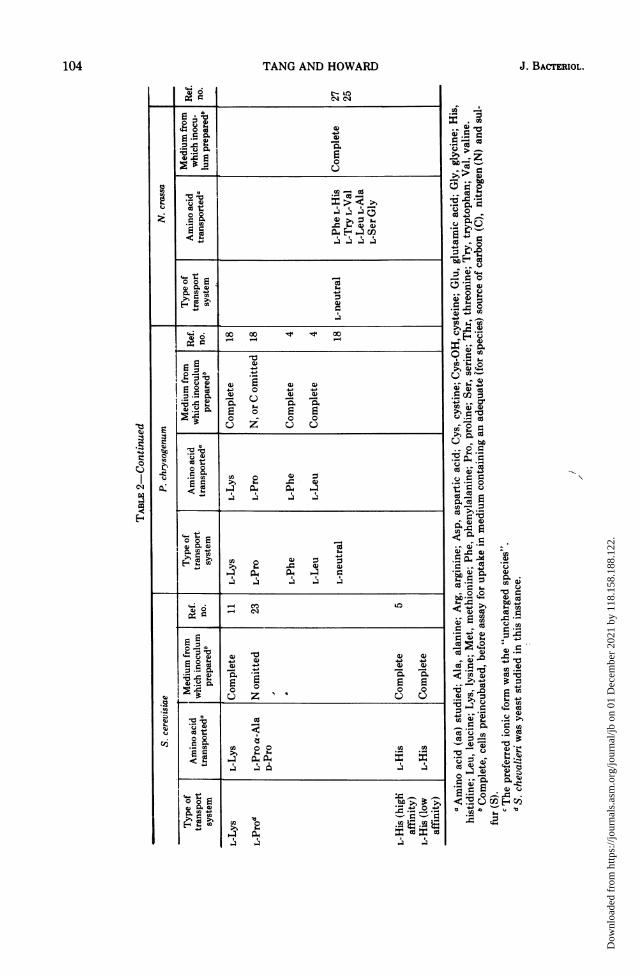
manifested, depending on the age of the inocu-lum and on the media employed. He showedthat most of the amino acids were transportedby four systems: (i) a neutral amino acidtransport system (which he designated as sys-tem I); (ii) a general nonspecific system (whichhe designated as system II); (iii) a basic aminoacid transport system (which he designated assystem III); and (iv) an acidic amino acidtransport system (which he designated as sys-tem IV). System I was highly active in young,rapidly growing cultures, whereas system II washighly active in ungerminated conidia and oldmycelia. The low Km values of system II made itquite efficient in removing traces of amino acidsfrom the external media. Pall suggested that itmay serve as a scavenger taking up any remain-ing exogenous amino acid (25).Table 2 shows a comparison of amino acid
transport in N. crassa, P. chrysogenum, and S.cerevisiae. It is obvious that, under the usualculture conditions, fungi may display a numberof transport systems with narrow specificity.Under conditions of starvation, they have anactive transport system with very broad speci-ficity. Some deviations do occur, e.g., in young,actively growing mycelia or freshly germinatedNeurospora conidia, an additional relativelynonspecific system is present which has little orno affinity for basic amino acid (25). No distinctneutral amino acid system was detected in P.chrysogenum (18). The basic amino acid trans-port systems of Penicillium and Neurospora(26) have a very low affinity for L-histidine. S.cerevisiae, however, possesses two distincttransport systems for histidine (5). Also, in S.cerevisiae, a specific methionine transport sys-tem has been characterized by Gits and Gren-son (9) which is similar to those of Neurosporaand Penicillium in substrate specificity but isactive in cells grown in sulfur-rich medium.
In the uptake of glutamic acid by C. albidus,kinetic studies suggest that the amino acid mayenter the yeast cells by two saturable systems:one of high and one of low affinity. Both systemssatisfied the following criteria for active trans-port: (i) each system can be described by theMichaelis-Menten equation; (ii) each requiredmetabolic energy, as indicated by more than50% inhibition of uptake by 2,4-dinitrophenoland almost 100% by sodium azide; (iii) eachaccumulated against a concentration gradient,and (iv) each was dependent on temperature.Uptake was severely inhibited at 37 C. Glu-tamic acid transport systems in C. albiduscorrespond to the general nonspecific transportsystem like that described in N. crassa (25, 27),P. chrysogenum (4, 18), and S. cerevisiae (30).
Once it was inside the cells, glutamic acidwas incorporated into the protein and nucleicacid fractions. It was also used as an energysource. The metabolism of glutamic acid didnot appear to be affected at 37 C (unpublisheddata) although its uptake was severely inhibitedat that temperature.
ACKNOWLEDGMENTSThis investigation was supported by a Public Health
Service training grant 2 T01-AI-00249, and in part by a PublicHealth Service Grant AI-10478-01 from the National Instituteof Allergy and Infectious Disease.We thank Rishab K. Gupta, Department of Surgery,
School of Medicine, UCLA, for his constant helpful advicethroughout the course of the work.
LITERATURE CITED
1. Akedo, H., and H. N. Christensen. 1962. Nature of insulinaction on amino acid uptake by the isolated dia-phragm. J. Biol. Chem. 237:118-122.
2. Ames, G. F. 1964. Uptake of amino acids by Salmonellatyphimurium. Arch. Biochem. Biophys. 104:1-18.
3. Benko, P. V., T. C. Wood, and I. H. Segel. 1967.Specificity and regulation of methionine transport infilamentous fungi. Arch. Biochem. Biophys.122:783-804.
4. Benko, P. V., T. C. Wood, and I. H. Segel. 1969.Multiplicity and regulation of amino acid transport inPenicillium chrysogenum. Arch. Biochem. Biophys.129:498-508.
5. Crabeel, M., and M. Grenson. 1970. Regulation of histi-dine uptake by specific feedback inhibition of twohistidine permeases in Saccharomyces cerevisiae. Eur.J. Biochem. 14:197-204.
6. Emmons, C. W., C. H. Binford, and J. P. Utz. 1970.Medical mycology, 2nd Edition, Lea & Febiger, Phila-delphia.
7. Friede, J. D., D. P. Gilboe, K. C. Triebwasser, and L. M.Henderson. 1972. Transport of lysine and hydroxyly-sine in Streptococcus faecalis. J. Bacteriol.109:179-185.
8. Gilbert, B. E., and D. H. Howard. 1970. Uptake of cystineby the yeast phase of Histoplasma capsulatum. Infect.Immunity 2:139-144.
9. Gits, J. J., and M. Grenson. 1967. Multiplicity of theamino acid permeases in Saccharomyces cerevisiae.Biochim. Biophys. Acta 135:507-516.
10. Grenson, M. 1966. Multiplicity of the amino acid per-meases in Saccharomyces cerevisiae. IR. Evidence for aspecific lysine-transporting system. Biochim. Biophys.Acta 127:339-346.
11. Grenson, M., C. Hou, and M. Crabeel. 1970. Multiplicityof the kmino acid permeases in Saccharomyces cerevi-siae. IV. Evidence for a general amino acid permease.J. Bacteriol. 103:770-777.
12. Grenson, M., M. Mousset, J. M. Wiame, and J. Bechet.1966. Multiplicity of the amino acid permeases inSaccharomyces cerevisiae. I. Evidence for a specificarginine-transporting system. Biochim. Biophys.Acta 127:325-338.
13. Gross, W., and K. Ring. 1971. Active transport ofglutamate in Streptomyces hydrogenans. I. Studies onuptake and pool size, and their interrelationship.Biochim. Biophys. Acta 233:652-665.
14. Gupta, R. K., and D. H. Howard. 1971. Comparativephysiological studies of the yeast and mycelial forms ofHistoplasma capsulatum: Uptake and incorporation ofL-leucine. J. Bacteriol. 105:690-700.
105VOL. 115, 1973
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
b on
01
Dec
embe
r 20
21 b
y 11
8.15
8.18
8.12
2.

TANG AND HOWARD
15. Gupta R. K., and D. Pramer. 1970. Amino acid transportby the filamentous fungus Arthrobotrys conoides. J.Bacteriol. 103:120-130.
16. Gupta, R. K., and D. Pramer. 1970. Metabolism of valineby the filamentous fungus Arthrobotrys conoides. J.Bacteriol. 103:131-139.
17. Halpern, Y. S., and A. Even-Shoshan. 1967. Properties ofthe glutamate transport system in Escherichia coli. J.Bacteriol. 93:1009-1016.
18. Hunter, D. R., and I. H. Segel. 1971. Acidic and basicamino acid transport systems of Penicillium chrysoge-num. Arch. Biochem. Biophys. 144:168-183.
19. Joiris, C. R., and M. Grenson. 1969. Specificite etregulation d'une permease des acides amines dicarbox-yliques chez Saccharomyces cerevisiae. Arch. Int. Phy-siol. Biochim. 77:154-156.
20. Kay, W. W., and A. F. Gronlund. 1969. Proline transportby Pseudomonas aeruginosa. Biochim. Biophys. Acta193:444-455.
21. Littman, M. L. 1958. Capsule synthesis by Cryptococcusneoformans. Transactions, N. Y. Acad. Sci., Series II20:623-648.
22. Lodder, J. (ed.). 1970. The yeasts. Second Edition.North-Holland Publishing Co., Amsterdam, London.
23. Magania-Schwencke, N., and J. Schwencke. 1969. Aproline transport system in Saccharomyces chevalieri.Biochim. Biophys. Acta 173:313-323.
24. Magill, C. W., H. Sweeney, and V. W. Woodward. 1972.Histidine uptake in strains of Neurospora crassa with
normal and mutant transport systems. J. Bacteriol.110:313-320.
25. Pall, M. L. 1969. Amino acid transport in Neurosporacrassa. I. Properties of two amino acid transportsystems. Biochim. Biophys. Acta 173:113-127.
26. Pall, M. L. 1970. Amino acid transport in Neurosporacrassa. II. Properties of a basic amino acid transportsystem. Biochim. Biophys. Acta 203:139-149.
27. Pall, M. L. 1970. Amino acid transport in Neurosporacrassa. III. Acidic amino acid transport. Biochim.Biophys. Acta 211:513-520.
28. Pall, M. L. 1971. Amino acid transport in Neurosporacrassa. IV. Properties and regulation of a methioninetransport system. Biochim. Biophys. Acta 233:201-214.
29. Reid, K. G., N. M. Utech, and J. T. Holden. 1970.Multiple transport components for dicarboxylic aminoacids in Streptococcus faecalis. J. Biol. Chem.245:5261-5272.
30. Surdin, Y., W. Sly, J. Sire, A. M. Bordes, and H. deRobichon-Szulmajster. 1965. Proprietes et contr6le ge-netique du systeme d'accumulation des acides amineschez Saccharomyces cerevisiae. Biochim. Biophys.Acta 107:546-566.
31. Utech, N. M., K. G. Reid, and J. T. Holden. 1970.Properties of a dicarboxylic amino acid transport-defi-cient mutant of Streptococcus faecalis. J. Biol. Chem.245:5273-5280.
32. Yabu, K. 1970. Amino acid transport in Mycobacteriumsmegmatis. J. Bacteriol. 102:6-13.
106 J. BACTERIOL.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
b on
01
Dec
embe
r 20
21 b
y 11
8.15
8.18
8.12
2.





























