Embed Size (px)
Citation preview

1
Short title: Functional SNPs for leaf hair number 1
Corresponding author information: 2
Yuke He 3
Address: Fenglin Road 300, Shanghai 200032, China 4
E-mail: [email protected] 5
Title: Identification of functional single-nucleotide polymorphisms affecting leaf 6
hair number in Brassica rapa 7
All authors’ names and affiliations: 8
Wenting Zhang, Shirin Mirlohi, Xiaorong Li and Yuke He 9
National Key Laboratory of Plant Molecular Genetics, CAS Center for Excellence in 10
Molecular Plant Sciences, Shanghai Institute of Plant Physiology and Ecology, 11
Chinese Academy of Sciences, Fenglin Road 300, Shanghai 200032, China 12
13
Contribution of authors: 14
Y.H. conceived the project and research plan. W. Z. performed the experiments. W.Z. 15
and S.M. analyzed the data and wrote the article with contributions from all the 16
authors. X.L. co-supervised and complemented the writing. 17
18
One sentence summary: Functional SNPs for leaf hair number in Brassica rapa 19
were selected and non-functional SNPs excluded by intensive mutagenesis and 20
genetic transformation. 21
22
Funding information: 23
This work was supported by National Programs for Science and Technology Development 24
of China (Grant No. 2016YFD0101900) and Natural Science Foundation of China 25
(Grant No. 31571261) 26
27
Corresponding author email: [email protected] 28
29
Plant Physiology Preview. Published on April 25, 2018, as DOI:10.1104/pp.18.00025
Copyright 2018 by the American Society of Plant Biologists
www.plantphysiol.orgon August 26, 2018 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.

2
30
Identification of functional single-nucleotide polymorphisms 31
affecting leaf hair number in Brassica rapa 32
33
Wenting Zhang1,2,3, Shirin Mirlohi1,2,3, Xiaorong Li1,2,3and Yuke He1,3* 34
35 1National Laboratory of Plant Molecular Genetics, Institute of Plant Physiology and Ecology, 36
Chinese Academy of Sciences, 300 Fenglin Road, Shanghai 200032, China 37 2Graduate School of the Chinese Academy of Sciences, Shanghai, China 38
3CAS Center for Excellence in Molecular Plant Sciences, Institute of Plant Physiology and 39
Ecology, Chinese Academy of Sciences, Shanghai, China 40
41
*Corresponding author. E-mail: [email protected] 42
43
44
ABSTRACT 45
Leaf traits affect plant agronomic performance; for example, leaf hair number 46
provides a morphological indicator of drought and insect resistance. Brassica rapa 47
crops have diverse phenotypes and many B. rapa single-nucleotide polymorphisms 48
(SNPs) have been identified and used as molecular markers for plant breeding. 49
However, which SNPs are functional for leaf hair traits and therefore effective for 50
breeding purposes remains unknown. Here we identify a set of SNPs in the B. rapa 51
ssp. pekinenesis candidate gene BrpHAIRY LEAVES1 (BrpHL1) and a number of 52
SNPs of BrpHL1 in a natural population of 210 B. rapa accessions that have hairy, 53
margin-only hairy, and hairless leaves. BrpHL1 genes and their orthologs and 54
paralogs have many SNPs. By intensive mutagenesis and genetic transformation, we 55
selected the functional SNPs for leaf hairs by exclusion of non-functional SNPs and 56
the orthologous and paraologous genes. The residue Trp92
of BrpHL1a was essential 57
for direct interaction with GLABROUS3 (BrpGL3) and thus necessary for formation 58
of leaf hairs. The accessions with the functional SNP leading to substitution of the 59
Trp92
residue had hairless leaves. The orthologous BrcHL1b from B. rapa ssp. 60
chinensis regulates hair formation on leaf margins rather than leaf surfaces. The 61
selected SNP for the hairy phenotype could be adopted as a molecular marker for 62
www.plantphysiol.orgon August 26, 2018 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.

3
insect resistance in Brassica crops. Moreover, the procedures optimized here can be 63
used to explain the molecular mechanisms of natural variation and to facilitate 64
molecular breeding of many crops. 65
66
Key words: Brassica rapa; functional SNPs; leaf hairs; natural variation; SNPs; 67
molecular breeding by designing; trichome; 68
69
INTRODUCTION 70
Brassica is one of the most important branches of the Brassicaceae family, including 71
many varieties of common vegetable crops. By means of natural and artificial 72
selection through time, many crops in Brassica rapa have evolved that show obvious 73
differences in leaf traits such as heading Chinese cabbage (B. rapa ssp. pekinenesis), 74
non-heading Chinese cabbage (B. rapa ssp. chinensis), turnip (B. rapa ssp. rapifera), 75
and yellow sarson (B. rapa ssp. trilocularis). The Brassica database (BRAD) 76
website (http://brassicadb.org/brad/) has released the complete genome sequence of 77
several Brassica crops (Cheng et al., 2011). Traditionally, leaf shape, size and 78
curvature are the main traits in these crops that have been genetically selected for 79
improved yield and quality. Hence, studying the diversity of the leaf traits in 80
Brassica could provide valuable information to help understand leaf development 81
and leaf variation and how to genetically manipulate these vegetable crops in the 82
future. 83
A leaf hair (trichome) is an epidermal hair that serve as a physical barrier on 84
plant surfaces against biotic and abiotic stress, including insect herbivores, 85
pathogenic microorganisms, UV light, excessive transpiration, freezing, etc. (Harada 86
et al., 2010; Hegebarth et al., 2016; Nafisi et al., 2015; Van Cutsem et al., 2011). The 87
adaptive significance of leaf hairs for arid land plants has been documented 88
(Ehleringer and Mooney 1978). 89
Density and localization of leaf hairs vary with crops within B. rapa. In 90
Arabidopsis (Arabidopsis thaliana), leaf hairs usually exist throughout the whole 91
plant, except for the cotyledons and epicotyls. A range of mutants defining specific 92
www.plantphysiol.orgon August 26, 2018 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.

4
aspects of trichome development have been found in Arabidopsis. The genetic 93
analysis of these mutants has revealed a number of key genes controlling this 94
patterning process, and the mechanism by which trichome differentiation is triggered 95
in individual cells has been best studied in Arabidopsis (Galway et al., 1994; 96
Oppenheimer et al., 1991; Payne et al., 2000; Rerie et al., 1994; Wada et al., 1997). 97
As most biological traits are genetically complicated, mapping quantitative trait 98
loci (QTL) is a powerful means for estimating the framework of the genetic 99
architecture for a trait and potentially identifying the genes responsible for a specific 100
phenotype. Recombinant inbred lines (RILs) of two Arabidopsis ecotypes, hairy 101
Columbia and less hairy Landsberg erecta, have been constructed to identify QTL 102
contributing to trichome number, and a major locus named REDUCED TRICHOME 103
NUMBER (RTN) has been confirmed (Larkin et al., 1996). In another study, four 104
recombinant inbred mapping populations based on six Arabidopsis ecotypes have 105
been used to reveal QTL controlling trichome density, and nine QTL have been 106
identified as responsible for trichome initiation and development (Symonds et al., 107
2005). 108
Some studies report that the leaf hairs in Chinese cabbage are controlled by a 109
single dominant gene, whereas others have shown that leaf hairs are a quantitative 110
phenotype, controlled by several major QTL (Song et al., 1995; Zhang et al., 2009). 111
The mechanisms of trichome development in Brassica crops and in Arabidopsis 112
might be highly conserved (Alahakoon et al., 2016; Nayidu et al., 2014). A gene 113
(Bra009770) located on chromosome A06 in B. rapa is homologous with 114
TRANSPARENT TESTA GLABRA1 (TTG1) in Arabidopsis, and it controls trichome 115
formation and seed coat color (Zhang et al., 2009). Moreover, nucleotide 116
polymorphisms of four alleles in the GLABROUS1 (GL1) ortholog (BrGL1) are 117
associated with hairless leaves (Li et al., 2011). In addition, a 5 bp deletion in Brtri1 118
(BrGL1) is related to a glabrous phenotype (Ye et al., 2016). 119
The molecular understanding of the functional consequences of genetic 120
variation is critical for application of single nucleotide polymorphisms (SNPs) to 121
plant breeding. Progress towards this goal has been mostly successful when the 122
www.plantphysiol.orgon August 26, 2018 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.

5
genetic variation falls within a coding region. Unfortunately, most SNPs identified in 123
plants are located within large introns or are distal to coding regions. 124
SNPs have a wide distribution and can be found in any region of a gene, mRNA 125
or intergenic region. Although identification of SNPs is an important first step in 126
understanding the relationship between variation and phenotypes, a major challenge 127
in the post-GWAS era is to understand the functional significance of the identified 128
SNPs and to apply these SNPs to plant breeding. Usually, GWAS SNPs (SNPs at 129
QTL) in the coding sequences of the candidate genes are used for designing DNA 130
markers. In practice, many GWAS SNPs are not effective for selection of objective 131
traits and many breeders have experienced failures in the practice of molecular 132
marker-aided selection with GWAS SNPs. Among these SNPs, some are functional 133
for developmental events and traits because they affect the levels of gene expression 134
or translation, splicing, efficiency to enhance or inhibit mRNA stability and protein 135
function. Many polyploid plants have multiple gene copies and many SNPs. 136
Selection of the causal SNPs for certain traits largely depends on the exclusion of 137
non-functional SNPs. Identifying functional SNPs for objective traits from a large 138
number of SNPs presents a bottleneck in the process. Furthermore, successful 139
molecular breeding of crops relies largely on the accuracy of functional SNPs. 140
Recently, many SNPs have been identified in B. rapa (Kim et al., 2016; 141
Tanhuanpaa et al., 2016; Yu et al., 2016). However, which SNPs are responsible for 142
leaf hairs remain unknown. In this study, we took advantage of recent advances in 143
genome resequencing to perform QTL mapping using 150 RILs derived from the 144
cross between the hairy genotype Bre (B. rapa ssp. pekinensis) and the hairless 145
genotype Wut (B. rapa ssp. chinensis) of Chinese cabbage (Yu et al., 2012). We 146
identified functional SNPs from numerous SNPs in the candidate genes and their 147
duplicated copies, and optimized the procedures for selection of functional SNPs for 148
agronomic traits. The selected SNP for leaf hair is a molecular marker of insect 149
resistance in Brassica crops, and the optimized procedures can be used to explain the 150
molecular mechanism of natural variation and to manipulate molecular breeding of 151
many crops. 152
www.plantphysiol.orgon August 26, 2018 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.

6
153
RESULTS 154
155
Genetic control of leaf hair number 156
The species B. rapa includes heading Chinese cabbage (B. rapa ssp. pekinensis), 157
non-heading Chinese cabbage (B. rapa ssp. chinensis), turnip (B. rapa ssp. rapa), 158
and yellow sarson (B. rapa ssp. trilocularis). Non-heading Chinese cabbage consists 159
of many crop types: baicai, caixin, caitai, purple caitai, taicai, wutacai. These crops 160
and crop types are characterized by their specialized product organs: curved leaves, 161
leafy heads, fleshy petioles, fleshy stems and fleshy roots. 162
To analyze the leaf variation of B. rapa, we collected 210 accessions of B. rapa. 163
The leaf hairs on young leaves of these accessions were observed under a 164
binocular stereo microscope. There were three types of leaves with regard to leaf 165
hairs (Table 1). Most of the accessions belonged to the "all hairy" leaves in which 166
the leaf hairs were visible on the surfaces and margins. Some accessions belonged to 167
the "margin-only" leaves in which the leaf hairs were visible only on the leaf 168
margins, and the rest of the accessions had hairless leaves. Among the 210 169
accessions observed, 99 showed the all-hairy phenotype while 16 displayed the 170
margin-only hairy phenotype (Table 1). Bre is a representative of the all-hairy 171
phenotype and Wut is a representative of the margin-only hairy phenotype: 172
numerous hairs were detected on the leaf surface and leaf margins of Bre whereas 173
only a few leaf hairs were seen on the leaf margins of Wut. (Fig. 1AB). Leaf hairs of 174
all-hairy and margin-only hairy phenotypes were not branched, in contrast with the 175
branched trichomes of Arabidopsis (Fig. 1CD). Most heading Chinese cabbage 176
accessions showed the all-hairy phenotype (Table 1, Fig. 1E, F, I, J), and most 177
non-heading Chinese cabbage accessions displayed a hairless or margin-only hairy 178
phenotype (Table 1, Fig. 1G, H, K, L). 179
180
Positional cloning of the HAIRY LEAVES1 (HL1a) loci 181
In the course of previous studies, the inbred lines of Bre and Wut were crossed to 182
www.plantphysiol.orgon August 26, 2018 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.

7
construct RIL populations for inheritance analysis and chromosomal mapping of the 183
related QTL. One major QTL, qTr, located on chromosome 6, was identified as a 184
locus for hairy leaves, and Bra025311 in the reference genome of Chiifu-401-42 was 185
selected as a candidate gene (Yu et al., 2013). Bra025311 belongs to the MYB gene 186
family (Supplementary Fig. 1) and is homologous to Arabidopsis GL1 with 59% 187
amino acid identity (Supplementary Fig. 2A). GL1 encodes an R2R3-MYB 188
transcription factor with a central function in the leaf hair patterning pathway, and is 189
involved in epidermal cell fate specification in leaves, promoting leaf hair formation 190
and endoreplication (Szymanski et al., 1998). 191
We named the candidate gene Bra025311 in Bre and Wut as BrpHL1a (B. rapa 192
ssp. pekinensis HL1a) and BrcHL1a (B. rapa ssp. chinensis HL1a), respectively. The 193
gene body (5' UTR, exons, introns and 3' UTR) and cDNAs of Bre BrpHL1a and 194
Wut BrcHL1a were cloned on the basis of genomic resequencing. Bre BrpHL1a 195
showed 56 SNPs in the gene body compared with that of Bra025311 of 196
Chiifu-401-42 (Supplementary Fig. 3). Fifty-five SNPs were identified in Wut 197
BrcHL1a (Supplementary Fig. 4). Compared to Chiifu-401-42 BrpHL1a, 11 SNPs 198
were detected in the exons of Bre BrpHL1a, causing 7 nonsynonymous substitutions; 199
and 13 SNPs were found in the Wut BrcHL1a gene, leading to 9 nonsynonymous 200
substitutions. Compared to Bre BrpHL1a, Wut BrcHL1a had 2 more SNPs in the 201
exons: 274T/C (274th dTMP of the coding sequence was changed to dCMP) and 202
403T/G (403rd T to dGMP) (Fig. 2A). SNPs 274T/C and 403T/G of Wut BrcHL1a 203
caused two nonsynonymous substitutions W92R and Y135D while 256C/T (256th 204
dCMP to dTMP) of Wut BrcHL1b leads to one nonsynonymous substitution Y86H. 205
B. rapa is a mesohexaploid and has more duplicated genes than Arabidopsis. To 206
check the other copies of the BrpHL1a gene, we searched for the genome sequences 207
of a collection of B. rapa accessions. BrpHL1b and BrcHL1b were detected on 208
chromosome 9 in Bre and Wut, respectively. The alignment shows that BrpHL1b and 209
BrcHL1b are homologous with GL1 of Arabidopsis (Supplementary Fig. 2B). Thus, 210
four members of the BrpHL1 gene family were related to the hairy phenotype of B. 211
www.plantphysiol.orgon August 26, 2018 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.

8
rapa. Our aim was to first discover which SNPs in the candidate BrpHL1a gene 212
were functional or causal for all hairy or hairless phenotypes. Then we aimed to 213
determine whether and how contributions of the other three members of the BrpHL1 214
gene family could be ruled out. 215
216
SNP analysis of BrpHL1a and BrpHL1b alleles in B. rapa 217
Compared to Chiifu-401-42 BrpHL1b, Bre BrpHL1b has an A/G SNP 85 bp 218
prior to the start codon, causing a shifted reading frame and prolongation of the first 219
exon. This gene also shows a 4.5 kb insertion in its second exon (Fig. 2B, 220
Supplementary Fig. 5A), which suggests that BrpHL1b was not functional in Bre. In 221
contrast, Wut BrcHL1b seemed to be functional as the A/G SNP and the large 222
insertion were not detected. 223
To find all the SNPs in B. rapa, we cloned and sequenced BrpHL1a and 224
BrpHL1b genes of 13 representative genotypes (Supplementary Table 1 and 2). A 225
subset of SNPs was detected. In total, 169 nucleotides (9%) of Chiifu-401-42 226
BrpHL1a and 27 nucleotides (2%) of Chiifu-401-42 BrpHL1b were substituted by 227
the other nucleotides of various BrpHL1a and BrpHL1b alleles, respectively. To 228
confirm the accuracy of SNPs, we cloned the full-length cDNA sequences of 229
BrpHL1a and BrpHL1b genes. Sequence analysis of these clones confirmed the 230
accuracy of the genomic sequences of BrpHL1a and BrpHL1b genes. 231
We then analyzed the association between SNPs of BrpHL1 genes and the hairy 232
phenotype. For BrpHL1a alleles, all the genotypes (Wut, Ripposinica and Qincai) 233
with SNP 274C showed the hairless phenotype (Supplementary Table 1), revealing a 234
association between 274C and the hairless phenotype. Among 10 genotypes with 235
274T, 7 showed all hairy phenotypes. Two of 7 genotypes with SNP 403G showed 236
the hairless phenotype. Four of 6 genotypes with 403T showed the all-hairy 237
phenotype. For BrpHL1b alleles, 2 of 3 genotypes with 255C/T showed the hairless 238
phenotype (Supplementary Table 2). Therefore, we were not certain that these SNPs 239
were associated with all-hairy phenotypes. 240
To clarify the relationship between SNPs and leaf hairs, we extended the SNP 241
www.plantphysiol.orgon August 26, 2018 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.

9
calling to a natural population of 210 B. rapa accessions. The re-sequencing of these 242
accessions generated two paired-end libraries with 150-bp reads (Supplementary 243
Table 3). According to the reference genome of B. rapa v1.5 (Ref), the sequencing 244
depth of the parental lines was more than 10-fold in each accession, and the mapped 245
depth was about 9-20. Each SNP supported by fewer than 4 reads was filtered out, 246
leading to 0.5~1.69 million high-quality SNPs (Supplementary Table 4). These SNPs 247
include many nucleotide substitutions, insertions and deletions. 248
Based on the genomic data in the BRAD, the SNPs were used to update the 249
genomic sequences of the BrpHL1a alleles (Supplementary Table 5) and separately 250
estimated by grouping the 210 accessions. Among 28 accessions with SNP 274C, 24 251
were hairless while 3 were margin-only hairy (Table 2; Supplementary Table 6), thus 252
showing the high association between SNP 274C and the hairless phenotype. Among 253
184 accessions with SNP 274T, 96 showed the all-hairy phenotype, revealing that 254
nearly half of the accessions with SNP 274T failed to show the all-hairy phenotype. 255
Surprisingly, the accessions with SNP 274T included a large proportion of hairless 256
accessions and a small proportion of margin-only hairy accessions. 257
258
Expression patterns of BrpHL1 genes 259
RT-qPCR and RT-PCR were used to examine the differences in expression of 260
BrpHL1a/BrcHL1a between Bre and Wut using the same pair of primers. The 261
expression level of BrpHL1a/BrcHL1a in Wut leaves was considerably higher than 262
that of Bre leaves (Fig. 3A). A similar result was obtained using RT-qPCR (Fig. 3B). 263
To investigate the expression patterns of BrpHL1a in the plants, we fused BrpHL1a 264
and BrcHL1a with the β-glucuronidase (GUS) gene. In the seedlings of the resultant 265
transgenic plants, the GUS signals of BrpHL1a::GUS and BrcHL1a::GUS were 266
visible in all organs, especially in cotyledons and rosette leaves (Fig. 3C). RT-PCR 267
showed that BrpHL1a and BrcHL1a genes were expressed in the leaf, stem, 268
cotyledon and root of Bre and Wut (Figure 3D). These results show that the temporal 269
and spatial expression patterns of two BrHL1a genes in Bre and Wut are similar. 270
271
www.plantphysiol.orgon August 26, 2018 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.

10
Functional analysis of the SNPs in BrpHL1a genes 272
To examine the GWAS SNPs of BrpHL1a alleles, we aligned gene bodies of 273
BrpHL1a with Wut BrcHL1a identified by GWAS. There were 44 GWAS SNPs in 274
BrpHL1a alleles (Supplementary Table 1), most of which were located in the introns. 275
We found that the two SNPs, 274T/C and/or 403T/G, cause non-synonymous 276
substitutions. To determine SNPs functional for leaf hairs, we mutagenized BrpHL1a 277
genomic DNA with 274C and/or 403G and BrcHL1a-g genomic DNA with 274T 278
and/or 403G and constructed a series of binary vectors of the mutated genes under 279
their control of the native promoters (Table 3). We then transferred them into the null 280
gl1 mutants of Arabidopsis that are deficient in trichome formation (Fig. 3E-F). 281
Firstly, the genomic BrpHL1a-g completely rescued the phenotype in terms of 282
trichome formation whereas the genomic BrcHL1a-g was unable to rescue the 283
phenotypes of the gl1 mutants (Table 3), revealing that Wut BrcHL1a-g is deficient 284
in formation of leaf hairs. Secondly, the C274T mutagenesis in BrcHL1a-g274C/T
285
plants completely rescued the gl1 phenotype, thus indicating that 274T/C is the 286
functional SNP for leaf hair. Thirdly, 403T/G mutagenesis in BrpHL1a-g403T/G
plants 287
also rescued the gl1 phenotype, showing that 403T/G is dispensable for leaf hair. 288
To exclude the possible effects of introns and the 3′-noncoding region on 289
function of BrcHL1a, we constructed cDNA sequences of BrpHL1a and BrcHL1a 290
under the control of their native promoters and the 3′-noncoding region and then 291
transferred them into the gl1 mutants. As expected, both BrpHL1a-c and BrcHL1a-c 292
were expressed equally at the transcriptional levels in all the transgenic lines (Fig. 293
3H). BrcHL1a-c did not rescue the phenotypes of the gl1 mutants, whereas 294
BrpHL1a-c completely rescued the phenotype in terms of trichome formation. 295
BrcHL1a-c274C/T
completely rescued the gl1 phenotype, while BrpHL1a-c274T/C
did 296
not rescue the gl1 phenotype. The phenotypic outcomes of BrpHL1a-c, BrcHL1a-c, 297
and BrcHL1a-c274C/T
in gl1 mutants were the same as the ones of BrpHL1a-g, 298
BrcHL1a-g, and BrcHL1a-g274C/T
, respectively (Fig. 3G). We conclude that the SNPs 299
in the intron and the 3′-noncoding regions of BrcHL1a are not the reason for the 300
alteration of leaf hairs. 301
www.plantphysiol.orgon August 26, 2018 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.

11
302
Effects of SNP 274T/C on direct interaction of BrcHL1a with BrpGL3 303
In Arabidopsis, a network of three classes of proteins consisting of TTG1 (a WD40 304
repeat protein), GL3 (a bHLH transcription factor) and GL1 (a MYB transcription 305
factor), activates trichome initiation and patterning (Zhang et al., 2003). GL3 306
functions together with GL1 and TTG1 to form a MYB-bHLH-WD40 (MBW) 307
activator complex. GL3 participates in the physical interactions with GL1, TTG1, 308
and itself, but GL1 and TTG1 do not interact. GL1 has the conserved 309
[DE]Lx2[RK]x3Lx6Lx3R amino acid signature in the R3 domain of R2R3 MYBs, 310
which is the structural basis for interaction between MYB and R/B-like bHLH 311
proteins (Zimmermann et al., 2004). We found that the W92R amino acid 312
substitution was within this conserved sequence. To further examine whether the 313
Trp92
mutation interferes with the interaction of BrcHL1a and GL3, we performed 314
pull-down assays. The result showed that the interaction of Arabidopsis 315
Maltose-binding protein (MBP)-AtGL3 with glutathione S-transferase 316
(GST)-BrpHL1a and GST-BrcHL1aR92W
(Fig. 4A) was strong, while that of 317
MBP-AtGL3 with GST-BrcHL1a and GST-BrpHL1aW92R
was weak. These results 318
suggest that Trp92
plays a critical role in direct interaction between BrpHL1a and 319
BrpGL3. 320
To further confirm the function of Trp92
, we performed a bimolecular 321
fluorescence complementation (BiFC) assay based on enhanced yellow fluorescent 322
protein (EYFP). The full-length coding sequences of AtGL3, BrpHL1a, BrcHL1a, 323
BrcHL1aR92W
and BrpHL1aW92R
were fused to the N- or C-terminal halves of EYFP. 324
Both types of fusion proteins were transiently introduced into mesophyll protoplasts 325
of Arabidopsis. The protein–protein interaction between the tester proteins resulted 326
in the proper folding of EYFP leading to its subsequent fluorescence in the 327
co-infiltrated protoplasts. The strong EYFP signals between BrpHL1a and AtGL3 328
and between BrcHL1aR92W
and AtGL3 were observed in the nucleus (Fig. 4B), 329
whereas much weaker BiFC signals were observed between BrpHL1aW92R
and 330
AtGL3 and between BrcHL1a and AtGL3. This result reveals that the interaction 331
www.plantphysiol.orgon August 26, 2018 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.

12
between BrcHL1a and AtGL3 was weak, thus confirming the critical role of Trp92
in 332
formation of leaf hairs. 333
334
Activation of BrpGL2 by BrpHL1a 335
In Arabidopsis, GL2 is required for normal trichome development. GL2 expression 336
is regulated by GL1 (Rerie et al., 1994). GL1 and GL3 bind directly to 337
the GL2 promoter (Wang and Chen, 2008). Dai et al., (2016) report that the 338
substitution of the 92nd
serine to phenylalanine (S92F) in the R3 domain of 339
Arabidopsis GL1 does not affect the interaction of GL1 and GL3 but affects the 340
binding of GL1 to the promoter of GL2. In Bre BrpHL1a, the 92nd
tryptophan 341
corresponds to the 91st tryptophan rather than the 92
nd serine in Arabidopsis GL1. 342
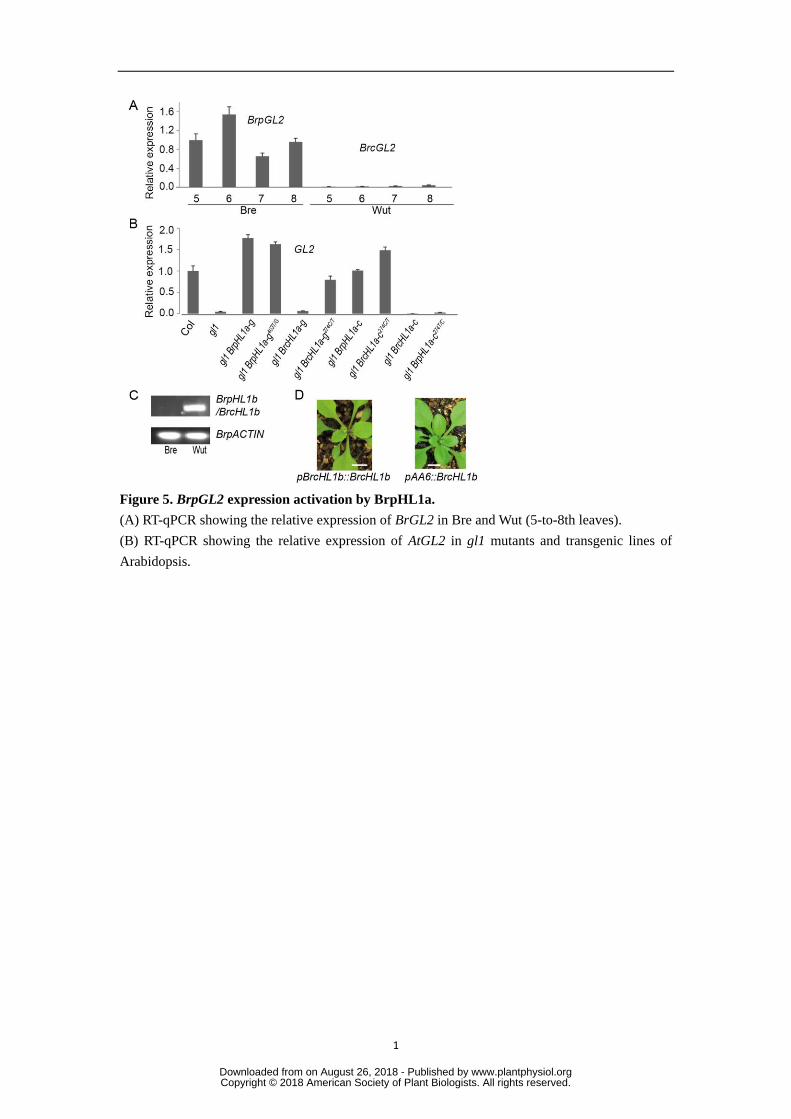
We supposed that the expression level of BrcGL2 in Wut would be reduced 343
compared with that of BrpGL2 in Bre if W92R in BrcHL1a was responsible for the 344
interaction between BrcHL1a and BrcGL3. To address this deduction, we performed 345
RT-qPCR using the same pair of primers whose sequences are conserved in Bre and 346
Wut. BrcGL2 expression was considerably lower than BrpGL2 expression in Bre 347
(Fig. 5A). 348
To confirm the role of the 92nd
tryptophan in the relevance of BrpHL1a to 349
BrpGL2, we analyzed the expression levels of GL2 in the Arabidopsis gl1 mutants 350
with exogenous BrpHL1a and BrcHL1a. GL2 expression was up-regulated in 351
pBrpHL1a::BrpHL1a plants, but not in pBrcHL1a::BrcHL1a plants (Fig. 5B), 352
indicating that BrcHL1a was not able to activate GL2. 353
354
Analysis of BrpHL1b gene functions 355
Considering that BrpHL1a functions in formation of leaf hairs in Bre while BrcHL1a 356
does not in Wut, we wondered whether and how BrpHL1b and BrcHL1b function in 357
formation of leaf hairs. So we analyzed the sequences of these two genes and found 358
that a 4.5 kb insertion was detected in the second exon of Bre BrpHL1b but not in 359
Wut BrcHL1b. RT-PCR result showed no expression of BrpHL1b in Bre (Fig. 5C), 360
meaning that BrpHL-2 is not functional. Also, BrpHL1a was the only functional 361
www.plantphysiol.orgon August 26, 2018 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.

13
gene of BrpHL1 genes in Bre. BrcHL1b of Wut did not contain any 362
insertions/deletions (InDels) that interrupt the protein sequence. If BrcHL1b of Wut 363
is functional for leaf hair, its hairless phenotype could be difficult to explain. To 364
verify the function of Wut BrcHL1b, we cloned the BrcHL1b gene body including 365
the promoter from Wut plants, and transferred pBrcHL1b::BrcHL1b (under the 366
control of the native promoter) and pAA6::BrcHL1b (under the control of 367
constitutive promoter AA6) constructs (Wang et al., 2014) into gl1 mutants. 368
Although no trichomes were observed on the leaf surface of pBrcHL1b::BrcHL1b 369
plants, a few trichomes were seen on the leaf margin. pAA6::BrcHL1b plants also 370
showed more trichomes on leaf margins than pBrcHL1b::BrcHL1b plants (Table 3; 371
Figure 5D). All together, these results indicated that BrcHL1b regulates hair 372
formation on leaf margins rather than the leaf surface. 373
374
DISCUSSION 375
376
Natural variation at the BrpHL1 locus is extensive 377
Genetic variation is brought about by mutation. Fundamentally, the numbers and 378
density of SNPs in a genome reflect the extent of natural variation in this species. In B. 379
rapa, our natural population of 210 accessions showed 0.5~1.69 million high-quality 380
SNPs compared with the reference genome of Chiifu-401. BrpHL1a on chromosome 381
6 of Bre shows 169 nucleotides that are substituted by its alleles of 13 representative 382
crop types, revealing that natural variation at BrpHL1 locus is extensive. However, 383
most of the SNPs are located in introns and may not be functional. The -85A/G 384
substitution of BrpHL1b on chromosome 9 in some accessions should change the start 385
codon at the 5' side and could thus affect the function of BrpHL1b. On the other hand, 386
BrpHL1b (the second copy of BrpHL1a) of Bre has a 4.5-kb insertion in the second 387
exon compared with BrcHL1b of Wut. This insertion causes a frame shift and 388
truncation of BrpHL1b in Bre. Although the SNP -85A/G and 4.5-kb insertion are not 389
GWAS SNPs, they substantially affect the functions of BrpHL1b in Bre. 390
www.plantphysiol.orgon August 26, 2018 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.

14
The SNPs of BrcHL1a and BrcHL1b in Wut may be related to the hairless 391
phenotype as Wut leaves are hairless. Among these SNPs, 403T/G is not a causal 392
element for leaf hair since T-to-C mutation in BrcHL1a is not able to rescue the gl1 393
mutant phenotype of Arabidopsis. Expression of BrcHL-2 under the control of its 394
native promoter causes marginal trichomes on the gl1 mutant, thus showing the 395
margin-specific expression of BrcHL-2. In this way, the contribution of the SNPs 396
to all hairy phenotypes is excluded except for SNP 274T/C in the BrcHL1a allele. 397
274T/C is a functional SNP for leaf hairs 398
In recent years, with the rapid development of next-generation sequencing 399
technologies and bioinformatics methods, crop breeding theory and technology has 400
undergone major changes. Numerous studies on genetic map construction and 401
marker-assisted selection have been carried out in Brassica crops. Genomic 402
resequencing is a method designed to sequence all regions of the genome aimed at 403
simplifying genome complexity. Marker-assisted selection is an effective 404
technology for obtaining large numbers of molecular markers and has been widely 405
used for high-throughput SNP discovery and for genotyping in different organisms 406
which are now widely used for large-scale high-throughput SNP genotyping, 407
particularly for de novo SNP discovery. In addition to the advantage of high density 408
and high throughput, our GWAS analysis of SNPs for leaf hairs in B. rapa was 409
effective. One major advantage of using the RIL populations is that researchers can 410
identify some QTL for the specific traits using low-covered resequencing. 411
Compared to GWAS in a natural population, GWAS in biparental cross populations 412
is more efficient and accurate (Yu et al., 2013). The QTL obtained in this way are 413
suitable for selection of candidate genes and GWAS SNPs relevant to leaf hairs, and 414
thereby establish the relationship between SNPs and leaf hairs in a natural 415
population. 416
The identification of causal SNPs is more difficult and should be combined with 417
the exclusion of other SNPs and the relevant alleles. The four members of the BrpHL1 418
gene family are relevant to leaf hairs. We resequenced 210 accessions of B. rapa and 419
www.plantphysiol.orgon August 26, 2018 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.

15
identified a SNP 274T/C in the candidate BrpHL1a gene. The function of the 420
candidate genes in two parents was identified by transgenic plants of suitable mutants. 421
Through point mutagenesis of SNPs in BrpHL1a genes and functional analysis of 422
these SNPs in the gl1 mutants of Arabidopsis, we excluded some SNPs in introns and 423
nonsynonymous SNPs in coding sequences. This selection criterion is more advanced 424
than that reported in many other previous studies. Nonetheless, a functional test in 425
Arabidopsis is not necessarily a proof of function in B. rapa. Therefore, we suggest 426
that the SNPs in the coding regions of B. rapa genes could be identified accurately by 427
gene transfer into Brassica crops. 428
The number of functional SNPs reported remains very limited, and some 429
functional SNPs should be embedded in the resequencing data. Thus, a great deal of 430
work is still needed to improve the SNP calling and QTL mapping accuracy by using 431
high-throughput sequencing technologies and making full use of the reference 432
Chinese cabbage genome. 433
434
Trp92
is essential for direct interaction with GL3 435
In Arabidopsis, a network of three classes of proteins consisting of TTG1 (a WD40 436
repeat protein), GL3 (a bHLH factor) and GL1 (a MYB transcription factor), 437
activates trichome initiation and patterning (Zhang et al., 2003). As positive 438
regulators, these three proteins form a MBW activator complex. GL3 participates in 439
physical interactions with GL1, TTG1, and itself, but GL1 and TTG1 do not interact 440
with each other. We also found the Trp92
in BrpHL1a is critical for interaction with 441
GL3. The interaction would be disrupted as soon as the critical amino acids in 442
BrpHL1a are mutated. In our BiFC experiments, the interactions between BrcHL1a 443
and GL3 and between BrpHL1aW92R
and GL3 were hardly detectable while 444
interactions between BrpHL1a and GL3 and between BrcHL1aR92W
and GL3 were 445
strong. In pull-down assays, the relative interaction strengths were similar. The hairy 446
phenotype of pBrpHL1a::BrpHL1a plants and hairless pBrcHL1a::BrcHL1a plants 447
in the gl1 background demonstrated that Trp92
of pBrpHL1a is essential for 448
formation of leaf hairs in B. rapa. Analyses of loss-of-function mutants reveal that 449
www.plantphysiol.orgon August 26, 2018 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.

16
single-repeat R3 MYB transcription factors act as negative regulators (Gan et al., 450
2011; Schellmann et al., 2002; Schnittger et al., 1999). In Arabidopsis, intron 1 and 451
the 3′-noncoding region of GL1 have been shown to be important for the expression 452
of GL1 (Larkin et al., 1993; Wang et al., 2004). Chromatin immune precipitation 453
results show that the single-repeat R3 MYB transcription factor TRICHOMELESS 1 454
(TCL1) can be recruited to the cis-acting regulatory elements of GL1, negatively 455
regulating trichome cell specification by directly suppressing the transcription of 456
GL1 (Wang et al., 2007). The importance of Trp92
for plant phenotype reveals that 457
intron 1 and the 3′-non-coding region of BrpHL1a are not as essential as Trp92
for 458
hair formation on the leaf surface. 459
The MYB (GL1)-bHLH(GL3/EGL3)-WDR(TTG1) proteins form a trimeric 460
MBW complex that activates the expression of the homo domain protein, GL2, 461
which, in turn, induces trichome formation (Rerie et al., 1994). Here, we noticed that 462
the expression of GL2 was reduced in Trp92
mutant plants, suggesting that blocking 463
the interaction between BrHL1a and GL3 affects hair formation on the leaf surface. 464
465
BrpHL1a regulates hair formation on both the leaf surface and leaf margin 466
while BrcHL1b functions only on the leaf margin 467
Usually, there are many SNPs in one gene, which makes it difficult for researchers to 468
select functional SNPs. It is very important to exclude the non-functional GWAS 469
SNPs, especially when duplicated genes are predicted to have biological functions. 470
Although the exclusion of non-functional SNPs is time-consuming and 471
labor-intensive, it is necessary for us to explain the complicated genetic process of 472
agronomic traits. Bre is a representative hairy crop type as the surface and margins of 473
leaves are hairy. By contrast, Wut is regarded as a representative hairless crop type as 474
hair is not seen on the surface of leaves and only a few hairs are detected on leaf 475
margins. The hairy phenotype in Bre corresponds to 274T and 403T in BrpHL1a and a 476
shift in the reading frame and a large insertion in the second exon in BrpHL1b. In 477
contrast, the hairless phenotype is concurrent with 274C and 403G in BrcHL1a and a 478
normal reading frame in BrpHL1b. By genetic transformation, we confirmed that 479
www.plantphysiol.orgon August 26, 2018 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.

17
BrpHL1a regulates hair formation on both the leaf surface and leaf margin while 480
BrpHL1b does not. The point mutation of BrpHL1a and BrcHL1a genes shows that 481
274T of BrpHL1a or Trp92
of BrpHL1a is essential for hair formation while 403T or 482
Tyr135
is dispensable. Comparison of intronic and non-intronic BrpHL1a transgenes 483
reveals that the first intron and 3’ UTR of BrpHL1a is not essential for its function, in 484
contrast with Arabidopsis GL1 whose first intron and 3’ UTR play roles in trichome 485
formation by interaction of GL1 with GL2 and GL3, respectively (Larkin et al., 1993; 486
Wang et al., 2004). Young developing gl3 leaves lack marginal trichomes, a 487
phenotype further enhanced in the tt8gl3 double mutant, indicating that both 488
TRANSPARENT TESTA8 (TT8) and GL3 are essential for trichome development on 489
leaf margins (Maes et al., 2008). 490
The function of BrpHL1b and BrcHL1b should be considered when that of 491
BrpHL1a and BrcHL1a is clarified. BrpHL1b in Bre is not functional due to the shift 492
in the reading frame and the large insertion in the coding region. However, BrcHL1b 493
in Wut is functional because its exogenous expression in gl1 mutant rescues the 494
trichome formation on leaf margins. Interestingly, hair formation on leaf margins in B. 495
rapa is not attributable to the promoter region of BrcHL1b as its native and 496
constitutive promoters generate leaf hairs on the same regions of leaf margins. 497
498
Functional SNPs are useful for molecular breeding by design 499
Traditional breeding is based on phenotype, and therefore depends primarily on 500
breeders’ experience. Since many traits of crops, such as disease resistance and yield, 501
cannot be observed easily, traditional breeding faces challenges and demands 502
high-throughput genotyping platforms. Molecular breeding by design is considered 503
the best option for breeders to improve their breeding efficiency. With the progress 504
in functional genomics research, increasing numbers of genes and QTL responsible 505
for agriculturally important traits have been identified, which provide valuable 506
genetic resources for molecular breeding. Resequencing and SNP genotyping are 507
two key strategies used in GWAS research and development of molecular markers to 508
target agronomic traits. To be suitable for molecular breeding by designing, we 509
www.plantphysiol.orgon August 26, 2018 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.

18
optimized the procedures for selection of functional SNPs for agronomic traits (Fig. 510
6). The segregation populations including RIL or doubled haploid lines are suitable 511
for QTL identification in which the genomic resequencing of different lines 512
generates the saturated SNPs. The SNPs located at the QTL are regarded as GWAS 513
SNPs because the candidate genes at the QTL locus are predicted according to 514
GWAS analysis. On the other hand, natural populations include major cultivars, 515
inbred lines and mutants, and, therefore, are very useful for variation analysis of 516
many agronomic traits. SNP genotyping on the basis of genomic resequencing 517
provides a strong tool for detection of SNPs in the large accession collections. 518
Through the comparison of GWAS SNPs from segregation and natural populations, 519
the candidates of functional SNPs are selected. They may be from exon-intron 520
junctures, DNA-RNA binding sites, protein-DNA binding sites, protein-protein 521
interaction domains, miRNAs and miRNA-target complementary sites. The 522
functional identification of the candidate SNPs is important but time-consuming. All 523
the binary vectors dedicated for genetic transformation should be designed to 524
exclude all of the non-functional SNPs and to select the functional SNPs. The null 525
mutants of the genes examined should be chosen for phenotypic rescue. The 526
molecular mechanism underlying the functional SNPs for agronomic traits could be 527
clarified. 528
In rice (Oryza sativa), a high-density SNP array with 51,478 markers has been 529
developed on the Illumina Infinium platform for use in functional genomics studies 530
and molecular breeding (Chen et al., 2014). However, many molecular makers 531
designed according to the GWAS SNPs are not effective in actual crop breeding, 532
largely due to non-functional SNPs. Among our accessions with SNP 274T/C, a 533
large proportion of accessions with 274T that are expected to have all hairy 534
phenotype show the hairless phenotype. One explanation is that some genes 535
downstream of BrpHL1 genes are mutated. Secondly, some cis- and trans-elements 536
exert effects on BrpHL1a. If 274T was used to design the molecular marker for leaf 537
hair, the high proportion of the false breeding materials would be selected. In 538
contrast, almost all accessions with 274C display the hairless phenotype. Therefore, 539
www.plantphysiol.orgon August 26, 2018 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.

19
274C is a functional SNP for designing the effective molecular markers to select the 540
hairless breeding materials. The application of the functional SNPs to designing 541
molecular markers will facilitate the selection of germplasms, parents and hybrids. 542
Further, the functional SNPs must be verified for their effectiveness in breeding of 543
new varieties with desirable traits. 544
545
MATERIALS AND METHODS 546
547
Plant materials 548
210 accessions of Brassica rapa were used in this study to survey leaf hairs. They 549
include many subspecies and varieties such as B. rapa subsp. chinensis, B. rapa var. 550
parachinensis, B. rapa var. purpuraria, B. rapa subsp. oleifera, B. rapa subsp. 551
narinosa, B. rapa var. perviridis, and B. rapa subsp. nipposinica. The seeds of these 552
crop types were sown in the field at the SIPPE Farm Station in Shanghai during 553
August 20-25 of 2008, 2009 and 2010. 554
Arabidopsis (Arabidopsis thaliana) gl1 (SALK_039478) mutants were kindly 555
provided by Prof. Xiaoya Chen (Wang et al., 2004). For phenotypic observation, 556
seeds were sown in pots with peat soil and incubated at 4°C in darkness for 3-4 days 557
and then moved to a growth chamber with 22°C temperature and 16/8 h of light/dark. 558
559
Phenotyping of leaf hairs 560
The third leaves at seedling stages were fixed and observed and the leaf hairs were 561
observed under an anatomical microscope, and the numbers of leaf hairs on surfaces 562
and margins of blades and petioles were observed. The mean value of the numbers of 563
leaf hairs per leaf in 10 plants was calculated. Plants with 1 or 2 hairs that were too 564
short to be recognized were regarded as hairless plants while plants with more than 2 565
hairs were considered hairy plants. 566
567
Sequencing data and alignment with reference genome 568
The DNA samples were sent to Novogene for sequencing by an Illumina 569
www.plantphysiol.orgon August 26, 2018 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.

20
hiseq-XTEN system, which produced the paired-end libraries with 2×150 bp read 570
length. All data were submitted to The Sequence Read Archive with BioProject ID 571
PRJNA421038. 572
After cutting adapters, the mean of the quality scores and the GC proportion of raw 573
reads were calculated. The first whole genome sequence of the Brassica A genome 574
species (B. rapa ssp. pekinensis vs Chiifu-401-42) was used as the reference 575
(http://brassicadb.org). The raw paired-end libraries of the 210 accessions were 576
aligned to the reference genome using SOAPalligner (SOAP2) software with the 577
parameter ‘‘-l 32 -s 40 -v 5 -m 10 -x 1000 -r 2’’, as well as bwa/samtools with the 578
default parameter. The effective depth of sequencing was calculated as follows: the 579
total length of clean reads minus that of the filter reads that could not match to the 580
reference genome, all divided by the length of the reference genome. 581
582
SNP calling and filtering 583
Based on the alignment file of SOAPalligner, the reads of genomic resequencing that 584
aligned with the 10 different chromosomes were separated into 10 files, and ordered 585
according to the physical location of the chromosome. SAMtools 586
(http://samtools.sourceforge.net) was used for SNP and InDel detection of each 587
chromosome using Bayesian theory. 588
The true SNPs were selected based on the following criteria: (1) no second 589
heterozygous base existed; (2) there was a quality score over 20; and (3) there were at 590
least five supported reads. The genes containing SNPs and short InDels were selected 591
by comparing the location of SNP and InDel with those of all Brassica gene models 592
v1.5 (http://blast.ncbi.nlm.nih.gov). SNPs were further determined as per whether 593
they were located in exon regions, and whether they caused synonymous/ 594
nonsynonymous mutation, premature termination, or abnormal termination. 595
596
Gene cloning and genetic transformation 597
The BrpHL1a promoter region (1824 bp upstream of the translation start site), 3’ 598
UTR region (1608 bp upstream of the translation termination site) and a full-length 599
www.plantphysiol.orgon August 26, 2018 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.

21
coding sequence (1596 bp) were amplified from Bre. Meanwhile, the BrcHL1a 600
promoter region (1886 bp upstream of the translation start site), 3’ UTR region 601
(1622 bp upstream of the translation termination site) and a full-length coding 602
sequence (1224 bp) were amplified from Wut. The promoter, 3’ UTR region, and a 603
full-length coding sequence were cloned into pCAMBIA1301 binary vectors to 604
obtain the pBrpHL1a::BrpHL1a and pBrcHL1a::BrcHL1a constructs, respectively. 605
To verify the function of mutation site, site-directed mutagenesis was performed. 606
The C274T nucleotide substitution of the BrcHL1a coding sequence resulted in the 607
mutated BrcHL1aR92W
while the T403G nucleotide substitution of the BrpHL1a 608
coding sequence gave rise to the mutated BrpHL1aY135D
. The primers used for 609
polymerase chain reactions (PCRs) are listed in Supplemental Table 7. 610
The Arabidopsis plants were transformed using the floral-dip method (Clough 611
and Bent, 1998). For selection of transgenic plants, the seeds were sterilized and 612
germinated on agar medium containing 50 mg/ml hygromycin. Seedlings conferring 613
resistance to the hygromycin were transplanted in a greenhouse and grown at 22°C 614
under an 8-h light regimen. 615
616
RNA analysis 617
For reverse transcription quantitative PCR (RT-qPCR), total RNA was extracted 618
using Trizol (Invitrogen) and treated with DNas I (TaKaRa), followed by a 619
phenol/chloroform extraction to remove contaminating DNA. Approximately 4 μg of 620
purified RNA was used for first-strand complementary DNA (cDNA) synthesis 621
using PrimeScript® Reverse Transcriptase (TaKaRa) with oligo (dT) primers. 622
RT-qPCR was performed using the specific primer pairs (Supplementary Table 7) in 623
the MyiQ2 Two-color Real-time PCR Detection System (Bio-Rad, Hercules, CA, 624
USA). The comparative threshold cycle (Ct) method was used to determine relative 625
transcript levels (MyiQ2 two-color real-time PCR detection system; Bio-Rad). 626
Expression was normalized relative to that of ACTIN. Two developing leaves in one 627
B. rapa seedling and ten shoots of Arabidopsis seedlings were harvested for RNA 628
sampling. Three biological replicates and three technical replicates were performed. 629
www.plantphysiol.orgon August 26, 2018 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.

22
Error bars indicate standard deviation. 630
631
GUS Staining 632
GUS staining was performed on 14-d-old plants. Seedlings of the transgenic plants 633
were placed in staining solution (50 mM Na3PO4, pH 7.0, 0.5 mM X-gluc 634
[5-bromo-4-chloro-3-indolyl glucuronide], and 20% [v/v] methanol), vacuum 635
infiltrated, and incubated at 37°C overnight. After staining, tissues were fixed in 636
alcohol for further analysis. 637
638
BiFC Assays 639
Paired cYFP-tagged and nYFP-tagged constructs were cotransformed into 640
Arabidopsis protoplasts. After incubation at 22°C in darkness for 12 h, GFP and 641
YFP fluorescence signals were excited with 488 or 514 nm argon laser lines, with an 642
emission band of 495–540 nm for GFP detection, 520–560 nm for YFP detection, 643
and 675–765 nm for chlorophyll autofluorescence by confocal microscopy. 644
645
In vitro pull-down assays 646
For MBP pull-down assays, MBP-tagged proteins were bound to amylose resin 647
(NEB) in binding buffer containing 25 mMTris, pH 7.4, 1 mM EDTA, 0.01% NP-40 648
and 2 M NaCl, and incubated with GST-tagged proteins overnight at 4°C. Then the 649
resin was washed 10 times in the binding buffer and eluted by boiling in sodium 650
dodecyl sulfate (SDS)-PAGE loading dye. Aliquots of eluents (20 μl) were resolved 651
on SDS-PAGE gels for immunoblotting with the GST antibody. 652
653
654
ACKNOWLEDGEMENT 655
The gl1 mutants were obtained from Dr. Wang (Wang et al., 2004). This work was 656
supported by National Programs for Science and Technology Development of China (Grant 657
No. 2016YFD0101900) and Natural Science Foundation of China (Grant No. 658
31571261) 659
www.plantphysiol.orgon August 26, 2018 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.

23
660
ACCESSION NUMBERS 661
Arabidopsis thaliana GL1:AT3G27920; GL2:AT1G79840 GL3:AT5G41315. 662
Brassica rapa BrpHL1a:Bra025311; BrpHL1b:Bra039065; BrpGL2:Bra003535. 663
664
665
SUPPLEMENTARY MATERIALS 666
Supplementary Fig. 1. Alignment of BrpHL1a with homologs in other plant 667
species. 668
Supplementary Fig. 2. Alignment of BrpHL1a and BrpHL1b amino acid 669
sequences with GL1. 670
Supplementary Fig. 3. Alignment of cloned BrpHL1a with a reference gene 671
sequence. 672
Supplementary Fig. 4. Alignment of cloned BrcHL1a with a reference gene 673
sequence. 674
Supplementary Fig. 5. Schematic diagram of BrpHL1b on chromosome 9 in Bre 675
and Wut. 676
677
Supplementary Table 1. Genomic sequences of the BrpHL1a alleles cloned from 678
13 Brassica rapa genotypes (inbred lines). 679
Supplementary Table 2. Genomic sequences of the BrpHL1b alleles cloned from 680
13 Brassica rapa genotypes (inbred lines). 681
Supplementary Table 3. Summary of genome resequencing and mapping data 682
in 210 Brassica rapa accessions. 683
Supplementary Table 4. Summary of SNP calling data from 210 Brassica rapa 684
accessions. 685
Supplementary Table 5. SNP genotyping of BrpHL1a alleles from genome 686
resequencing of 210 Brassica rapa accessions. 687
Supplementary Table 6. Association between SNPs 274T/C and 403T/G of 688
BrpHL1a with hairy phenotypes in 210 Brassica rapa accessions. 689
Supplementary Table 7. Primer sequences used in this study. 690
691
Figure legends 692
693
Figure 1. Leaf hairs of different crop types in Brassica rapa. 694
(A, B) Plants of Bre (A) and Wut (B) at the seedling stage. 695
(C, D) Scanning electron microscopy showing the leaf hairs of Bre (C; scale bar=500 µm) and 696
Wut (D; scale bar=200 µm). 697
(E, F) Hair distribution on adaxial surfaces of leaves in Da38 (E) and Da15 (F). 698
(G, H) Hair distribution on leaf margins in B26 (G) and W12 (H). 699
(I, J) Hair distribution on abaxial surfaces of leaves in Da102 (I) and Da203 (J). 700
www.plantphysiol.orgon August 26, 2018 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.

24
(K, L) The leaf surface without hairs in B55 (K) and B61 (L). 701
702
Figure 2. cDNA and amino acid sequences of BrpHL1a and BrpHL1b. 703
(A) SNPs of BrpHL1a and nonsynonymous substitutions of BrpHL1a between Bre, Wut and 704
Chiifu-401-42. 705
(B) SNPs of BrpHL1b and nonsynonymous substitutions of BrpHL1b between Bre, Wut and 706
Chiifu-401-42. 707
SNPs are shown on a white background. Nonsynonymous substitutions are boxed. Black lines 708
indicate 5’UTR, exon or 3’UTR; Red arrows show R2 and R3 domains of MYB transcription 709
factors. 710
711
Figure 3. Temporal and spatial expression of BrpHL1a and BrpHL1b and the phenotypic 712
rescue of Arabidopsis gl1 by BrpHL1a and BrpHL1b and their mutated versions. 713
(A, B) RT-PCR (A) and RT-qPCR (B) showing the expression of BrpHL1a/BrcHL1a in Bre and 714
Wut. 715
(C, D) GUS fusion signals in Arabidopsis (C) and RT-PCR (D) in B. rapa showing the 716
expression patterns of BrpHL1a and BrcHL1a in 20-day-old seedlings. Bars=5 mm. 717
(E) Seedling phenotypes of the wild-type (Col) and gl1 mutants of Arabidopsis. Bars=10 mm. 718
(F, G) Seedling phenotypes of gl1 mutants transgenic for genomic BrpHL1a-g and BrcHL1a-g (F) 719
and BrpHL1a-c and BrcHL1a-c cDNAs (G) under the control of the native promoters. Bars=10 720
mm. 721
(H) RT-qPCR showing the expression level of BrpHL1a/BrcHL1a in gl1 mutants and all 722
transgenic Arabidopsis lines. Three biological replicates were used. Error bars indicate standard 723
deviation. 724
725
Figure 4. Physical interaction between BrpHL1a and GL3 proteins. 726
(A) Pull-down assay showing protein–protein interaction between BrpHL1a versions and GL3 727
tagged with GST and MBP respectively. 728
(B) BiFC analysis showing protein–protein interaction between BrpHL1a versions and GL3 in 729
protoplasts. Number of cells with GFP is shown in the table beneath. 730
731
Figure 5. BrpGL2 expression activation by BrpHL1a. 732
(A) RT-qPCR showing the relative expression of BrGL2 in Bre and Wut (5-to-8th leaves). 733
(B) RT-qPCR showing the relative expression of AtGL2 in gl1 mutants and transgenic lines of 734
Arabidopsis. 735
(C) RT-PCR showing expression of BrcHL1b/BrcHL1b in Bre and Wut. 736
(D) Seedling phenotypes of gl1 mutants transgenic for genomic BrcHL1b under the control of 737
the native promoter and PAA6 promoter. Bars=10 mm. 738
Three biological replicates were used for PCR. Error bars indicate standard deviation. 739
740
Figure 6. Procedures optimized for selection of functional SNPs. (1) Building of 741
segregation populations and natural populations. (2) Phenotyping, DNA 742
resequencing, SNP calling with segregation populations and variation analysis of 743
agronomic traits with natural populations. (3) QTLs identification with segregation 744
www.plantphysiol.orgon August 26, 2018 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.

25
populations and DNA resequencing with natural populations. (4) Selection of 745
GWAS SNPs with natural populations and SNP genotyping with natural 746
populations.(5) Functional identification of the candidate genes and SNPs. (6) 747
Prediction of functional SNPs on the basis of SNP mutagenesis and phenotype 748
rescue. (7) Exclusion of non-functional SNPs. (8) Selection of functional SNPs. (9) 749
Designing of effective markers. (10) Molecular breeding by design. DH, Doubled 750
haploid lines; GWAS, genome-wide association study; QTL, Quantitative trait locus; 751
SNP, Single nucleotide polymorphism; RIL, Recombinant inbred lines. 752
753
754
www.plantphysiol.orgon August 26, 2018 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.

26
REFERENCES 755
756
Alahakoon, U.I., Taheri, A., Nayidu, N.K., Epp, D., Yu, M., Parkin, I., Hegedus, 757
D., Bonham-Smith, P., Gruber, M.Y., 2016. Hairy Canola (Brasssica napus) 758
re-visited: Down-regulating TTG1 in an AtGL3-enhanced hairy leaf background 759
improves growth, leaf trichome coverage, and metabolite gene expression 760
diversity. Bmc Plant Biol 16, 12. 761
Chen, H., Xie, W., He, H., Yu, H., Chen, W., Li, J., Yu, R., Yao, Y., Zhang, W., 762
He, Y., Tang, X., Zhou, F., Deng, X.W., Zhang, Q., 2014. A high-density SNP 763
genotyping array for rice biology and molecular breeding. Molecular Plant 7, 764
541-553. 765
Cheng, F., Liu, S., Wu, J., Fang, L., Sun, S., Liu, B., Li, P., Hua, W., Wang, X., 766
2011. BRAD, the genetics and genomics database for Brassica plants. Bmc Plant 767
Biol 11, 136. 768
Clough, S.J., and Bent, A.F. 1998. Floral dip: a simplified method for 769
Agrobacterium-mediated transformation of Arabidopsis thaliana. Plant J. 16, 770
735–743. 771
Dai, X.M., Zhou, L.M., Zhang, W., Cai, L., Guo, H.Y., Tian, H.N., Schiefelbein, 772
J., Wang, S.C., 2016. A single amino acid substitution in the R3 domain of 773
GLABRA1 leads to inhibition of trichome formation in Arabidopsis without 774
affecting its interaction with GLABRA3. Plant Cell Environ 39, 897-907. 775
Ehleringer J. R., Mooney H. A. 1978. Leaf hairs: Effects on physiological activity 776
and adaptive value to a desert shrub. Oecologia 37, 183-200. 777
Galway, M.E., Masucci, J.D., Lloyd, A.M., Walbot, V., Davis, R.W., 778
Schiefelbein, J.W., 1994. The Ttg Gene Is Required to Specify Epidermal-Cell 779
Fate and Cell Patterning in the Arabidopsis Root. Developmental biology 166, 780
740-754. 781
Gan, L.J., Xia, K., Chen, J.G., Wang, S.C., 2011. Functional characterization of 782
TRICHOMELESS2, a new single-repeat R3 MYB transcription factor in the 783
regulation of trichome patterning in Arabidopsis. Bmc Plant Biol 11. 784
Harada, E., Kim, J.A., Meyer, A.J., Hell, R., Clemens, S., Choi, Y.E., 2010. 785
Expression profiling of tobacco leaf trichomes identifies genes for biotic and 786
abiotic stresses. Plant Cell Physiol 51, 1627-1637. 787
Hegebarth, D., Buschhaus, C., Wu, M., Bird, D., Jetter, R., 2016. The 788
composition of surface wax on trichomes of Arabidopsis thaliana differs from wax 789
on other epidermal cells. The Plant journal : for cell and molecular biology 88, 790
762-774. 791
Kim, J., Kim, D.S., Park, S., Lee, H.E., Ahn, Y.K., Kim, J.H., Yang, H.B., Kang, 792
B.C., 2016. Development of a high-throughput SNP marker set by transcriptome 793
sequencing to accelerate genetic background selection in Brassica rapa. Hortic 794
Environ Biote 57, 280-290. 795
Larkin, J.C., Oppenheimer, D.G., Pollock, S., Marks, M.D., 1993. Arabidopsis 796
Glabrous1 Gene Requires Downstream Sequences for Function. The Plant cell 5, 797
1739-1748. 798
www.plantphysiol.orgon August 26, 2018 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.

27
Larkin, J.C., Young, N., Prigge, M., Marks, M.D., 1996. The control of trichome 799
spacing and number in Arabidopsis. Development 122, 997-1005. 800
Li, F., Kitashiba, H., Nishio, T., 2011. Association of sequence variation in 801
Brassica GLABRA1 orthologs with leaf hairiness. Mol Breeding 28, 577-584. 802
Maes, L., Inze, D., Goossens, A., 2008. Functional specialization of the 803
TRANSPARENT TESTA GLABRA1 network allows differential hormonal 804
control of laminal and marginal trichome initiation in Arabidopsis rosette leaves. 805
Plant physiology 148, 1453-1464. 806
Nafisi, M., Stranne, M., Fimognari, L., Atwell, S., Martens, H.J., Pedas, P.R., 807
Hansen, S.F., Nawrath, C., Scheller, H.V., Kliebenstein, D.J., Sakuragi, Y., 808
2015. Acetylation of cell wall is required for structural integrity of the leaf surface 809
and exerts a global impact on plant stress responses. Frontiers in plant science 6. 810
Nayidu, N.K., Kagale, S., Taheri, A., Withana-Gamage, T.S., Parkin, I.A.P., 811
Sharpe, A.G., Gruber, M.Y., 2014. Comparison of Five Major Trichome 812
Regulatory Genes in Brassica villosa with Orthologues within the Brassicaceae. 813
Plos One 9. 814
Oppenheimer, D.G., Herman, P.L., Sivakumaran, S., Esch, J., Marks, M.D., 815
1991. A Myb Gene Required for Leaf Trichome Differentiation in Arabidopsis Is 816
Expressed in Stipules. Cell 67, 483-493. 817
Payne, C.T., Zhang, F., Lloyd, A.M., 2000. GL3 encodes a bHLH protein that 818
regulates trichome development in arabidopsis through interaction with GL1 and 819
TTG1. Genetics 156, 1349-1362. 820
Rerie, W.G., Feldmann, K.A., Marks, M.D., 1994. The Glabra2 Gene Encodes a 821
Homeo Domain Protein Required for Normal Trichome Development in 822
Arabidopsis. Gene Dev 8, 1388-1399. 823
Schellmann, S., Schnittger, A., Kirik, V., Wada, T., Okada, K., Beermann, A., 824
Thumfahrt, J., Jurgens, G., Hulskamp, M., 2002. TRIPTYCHON and 825
CAPRICE mediate lateral inhibition during trichome and root hair patterning in 826
Arabidopsis. The EMBO journal 21, 5036-5046. 827
Schnittger, A., Folkers, U., Schwab, B., Jurgens, G., Hulskamp, M., 1999. 828
Generation of a spacing pattern: the role of triptychon in trichome patterning in 829
Arabidopsis. The Plant cell 11, 1105-1116. 830
Song, K., Slocum, M.K., Osborn, T.C., 1995. Molecular marker analysis of genes 831
controlling morphological variation in Brassica rapa (syn. campestris). TAG. 832
Theoretical and applied genetics. Theoretische und angewandte Genetik 90, 1-10. 833
Symonds, V.V., Godoy, A.V., Alconada, T., Botto, J.F., Juenger, T.E., Casal, 834
J.J., Lloyd, A.M., 2005. Mapping quantitative trait loci in multiple populations of 835
Arabidopsis thaliana identifies natural allelic variation for trichome density. 836
Genetics 169, 1649-1658. 837
Szymanski, D.B., Jilk, R.A., Pollock, S.M., Marks, M.D., 1998. Control of GL2 838
expression in Arabidopsis leaves and trichomes. Development 125, 1161-1171. 839
Tanhuanpaa, P., Erkkila, M., Tenhola-Roininen, T., Tanskanen, J., Manninen, 840
O., 2016. SNP diversity within and among Brassica rapa accessions reveals no 841
geographic differentiation. Genome 59, 11-21. 842
www.plantphysiol.orgon August 26, 2018 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.

28
Van Cutsem, E., Simonart, G., Degand, H., Faber, A.M., Morsomme, P., 843
Boutry, M., 2011. Gel-based and gel-free proteomic analysis of Nicotiana 844
tabacum trichomes identifies proteins involved in secondary metabolism and in 845
the (a)biotic stress response. Proteomics 11, 440-454. 846
Wada, T., Tachibana, T., Shimura, Y., Okada, K., 1997. Epidermal cell 847
differentiation in Arabidopsis determined by a Myb homolog, CPC. Science 277, 848
1113-1116. 849
Wang, S., Kwak, S.H., Zeng, Q., Ellis, B.E., Chen, X.Y., Schiefelbein, J., Chen, 850
J.G., 2007. TRICHOMELESS1 regulates trichome patterning by suppressing 851
GLABRA1 in Arabidopsis. Development 134, 3873-3882. 852
Wang, S., Wang, J.W., Yu, N., Li, C.H., Luo, B., Gou, J.Y., Wang, L.J., Chen, 853
X.Y., 2004. Control of plant trichome development by a cotton fiber MYB gene. 854
The Plant cell 16, 2323-2334. 855
Wang, S.C., Chen, J.G., 2008. Arabidopsis Transient Expression Analysis Reveals 856
that Activation of GLABRA2 May Require Concurrent Binding of GLABRA1 857
and GLABRA3 to the Promoter of GLABRA2. Plant Cell Physiol 49, 1792-1804. 858
Ye, X.L., Hu, F.Y., Ren, J., Huang, S.N., Liu, W.J., Feng, H., Liu, Z.Y., 2016. 859
Fine mapping and candidate gene analysis of Brtri1, a gene controlling trichome 860
development in Chinese cabbage (Brassica rapa L. ssp pekinensis). Genetics and 861
molecular research : GMR 15. 862
Yu, F.Q., Zhang, X.G., Huang, Z., Chu, M.G., Song, T., Falk, K.C., Deora, A., 863
Chen, Q.L., Zhang, Y., McGregor, L., Gossen, B.D., McDonald, M.R., Peng, 864
G., 2016. Identification of Genome-Wide Variants and Discovery of Variants 865
Associated with Brassica rapa Clubroot Resistance Gene Rcr1 through Bulked 866
Segregant RNA Sequencing. Plos One 11. 867
Yu, X., Wang, H., Lu, Y.Z., de Ruiter, M., Cariaso, M., Prins, M., van Tunen, 868
A., He, Y.K., 2012. Identification of conserved and novel microRNAs that are 869
responsive to heat stress in Brassica rapa. J Exp Bot 63, 1025-1038. 870
Yu, X., Wang, H., Zhong, W., Bai, J., Liu, P., He, Y., 2013. QTL mapping of 871
leafy heads by genome resequencing in the RIL population of Brassica rapa. Plos 872
One 8, e76059. 873
Zhang, F., Gonzalez, A., Zhao, M.Z., Payne, C.T., Lloyd, A., 2003. A network of 874
redundant bHLH proteins functions in all TTG1-dependent pathways of 875
Arabidopsis. Development 130, 4859-4869. 876
Zhang, J., Lu, Y., Yuan, Y., Zhang, X., Geng, J., Chen, Y., Cloutier, S., 877
McVetty, P.B., Li, G., 2009. Map-based cloning and characterization of a gene 878
controlling hairiness and seed coat color traits in Brassica rapa. Plant Mol Biol 69, 879
553-563. 880
Zimmermann, I.M., Heim, M.A., Weisshaar, B., Uhrig, J.F., 2004. 881
Comprehensive identification of Arabidopsis thaliana MYB transcription factors 882
interacting with R/B-like BHLH proteins. Plant Journal 40, 22-34. 883
884
www.plantphysiol.orgon August 26, 2018 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.

1
Table 1. Number of accessions with and without leaf hairs in a collection of 210
Brassica rapa accessions.
Crop types
Number of accessions
Total All
hairy
Margin-only
hairy Hairless
Heading Chinese cabbage 116 85 12 19
Non-heading Chinese
cabbage
Baicai 70 10 2 58
Caitai 5 1 0 4
Caixin 8 0 0 8
Taicai 2 0 0 2
Tacai 4 0 1 3
Turnip 3 1 1 1
Yellow sarson 2 2 0 0
Total 210 99 16 95
www.plantphysiol.orgon August 26, 2018 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.

1
Table 2. Correlation between the accessions with the SNPs 274T/C and 403T/G of
BrpHL1a and hairy phenotypes.
Genotype Number of accessions
274 site 403 site Total All hairy Margin-only
hairy
Hairless
T T 111 63 7 41
T G 67 31 5 31
C T 0 0 0 0
C G 28 1 3 24
T X 4 2 1 1
Total 210 97 16 97
Note: “X”indicates the unknown nucleotide.
www.plantphysiol.orgon August 26, 2018 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.
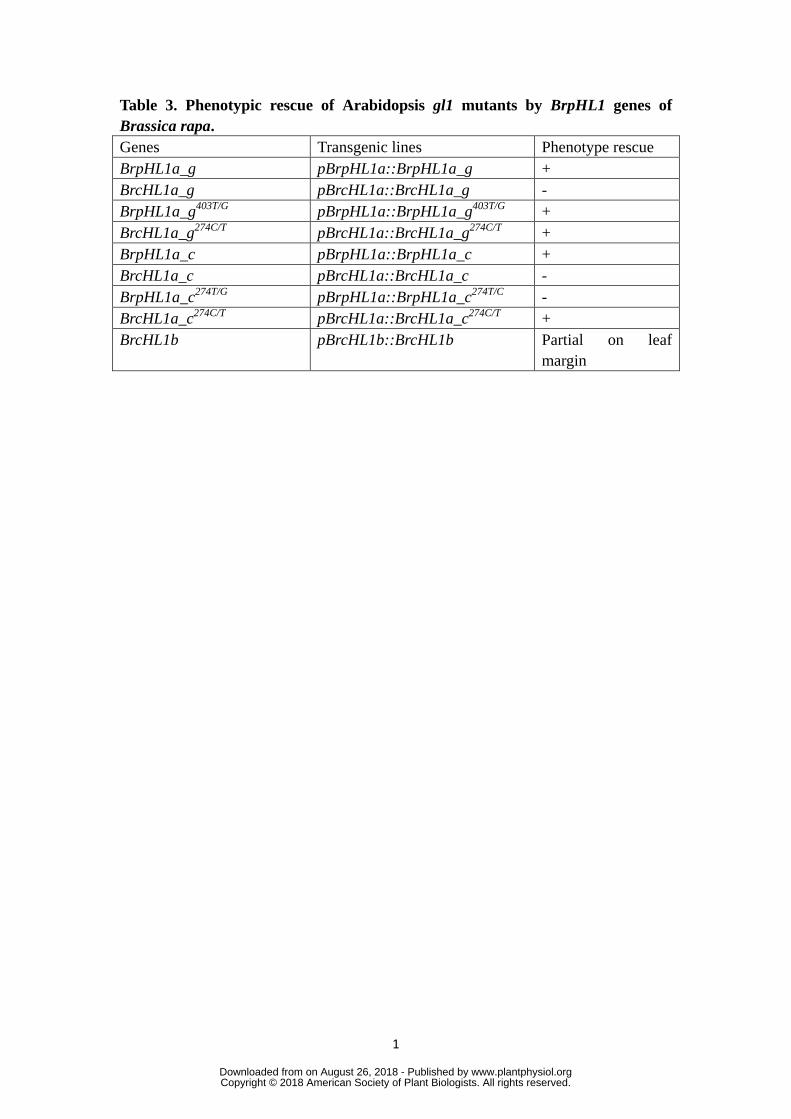
1
Table 3. Phenotypic rescue of Arabidopsis gl1 mutants by BrpHL1 genes of
Brassica rapa.
Genes Transgenic lines Phenotype rescue
BrpHL1a_g pBrpHL1a::BrpHL1a_g +
BrcHL1a_g pBrcHL1a::BrcHL1a_g -
BrpHL1a_g403T/G
pBrpHL1a::BrpHL1a_g403T/G
+
BrcHL1a_g274C/T
pBrcHL1a::BrcHL1a_g274C/T
+
BrpHL1a_c pBrpHL1a::BrpHL1a_c +
BrcHL1a_c pBrcHL1a::BrcHL1a_c -
BrpHL1a_c274T/G
pBrpHL1a::BrpHL1a_c274T/C
-
BrcHL1a_c274C/T
pBrcHL1a::BrcHL1a_c274C/T
+
BrcHL1b pBrcHL1b::BrcHL1b Partial on leaf
margin
www.plantphysiol.orgon August 26, 2018 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.

1
1 Figure 1. Leaf hairs of different crop types in Brassica rapa. 2
(A, B) Plants of Bre (A) and Wut (B) at the seedling stage. 3
(C, D) Scanning electron microscopy showing the leaf hairs of Bre (C; scale bar=500 µm) and 4
Wut (D; scale bar=200 µm). 5
(E, F) Hair distribution on adaxial surfaces of leaves in Da38 (E) and Da15 (F). 6
(G, H) Hair distribution on leaf margins in B26 (G) and W12 (H). 7
(I, J) Hair distribution on abaxial surfaces of leaves in Da102 (I) and Da203 (J). 8
(K, L) The leaf surface without hairs in B55 (K) and B61 (L). 9
10
www.plantphysiol.orgon August 26, 2018 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.

1
Figure 2. cDNA and amino acid sequences of BrpHL1a and BrpHL1b.
(A) SNPs of BrpHL1a and nonsynonymous substitutions of BrpHL1a between Bre, Wut and
Chiifu-401-42.
(B) SNPs of BrpHL1b and nonsynonymous substitutions of BrpHL1b between Bre, Wut and
Chiifu-401-42.
SNPs are shown on a white background. Nonsynonymous substitutions are boxed. Black lines
indicate 5’UTR, exon or 3’UTR; Red arrows show R2 and R3 domains of MYB transcription
factors.
www.plantphysiol.orgon August 26, 2018 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.

2
www.plantphysiol.orgon August 26, 2018 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.

1
Figure 3. Temporal and spatial expression of BrpHL1a and BrpHL1b and the phenotypic
rescue of Arabidopsis gl1 by BrpHL1a and BrpHL1b and their mutated versions.
(A, B) RT-PCR (A) and RT-qPCR (B) showing the expression of BrpHL1a/BrcHL1a in Bre and
Wut.
(C, D) GUS fusion signals in Arabidopsis (C) and RT-PCR (D) in B. rapa showing the expression
patterns of BrpHL1a and BrcHL1a in 20-day-old seedlings. Bars=5 mm.
(E) Seedling phenotypes of the wild-type (Col) and gl1 mutants of Arabidopsis. Bars=10 mm.
(F, G) Seedling phenotypes of gl1 mutants transgenic for genomic BrpHL1a-g and BrcHL1a-g (F)
and BrpHL1a-c and BrcHL1a-c cDNAs (G) under the control of the native promoters. Bars=10
mm.
(H) RT-qPCR showing the expression level of BrpHL1a/BrcHL1a in gl1 mutants and all
transgenic Arabidopsis lines. Three biological replicates were used. Error bars indicate standard
deviation.
www.plantphysiol.orgon August 26, 2018 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.

1
Figure 4. Physical interaction between BrpHL1a and GL3 proteins.
(A) Pull-down assay showing protein–protein interaction between BrpHL1a versions and GL3
tagged with GST and MBP respectively.
(B) BiFC analysis showing protein–protein interaction between BrpHL1a versions and GL3 in
protoplasts. Number of cells with GFP is shown in the table beneath.
www.plantphysiol.orgon August 26, 2018 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.
1
Figure 5. BrpGL2 expression activation by BrpHL1a.
(A) RT-qPCR showing the relative expression of BrGL2 in Bre and Wut (5-to-8th leaves).
(B) RT-qPCR showing the relative expression of AtGL2 in gl1 mutants and transgenic lines of
Arabidopsis.
www.plantphysiol.orgon August 26, 2018 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.

1
Figure 6. Procedures optimized for selection of functional SNPs. (1) Building of
segregation populations and natural populations. (2) Phenotyping, DNA resequencing,
SNP calling with segregation populations and variation analysis of agronomic traits
with natural populations. (3) QTLs identification with segregation populations and
DNA resequencing with natural populations. (4) Selection of GWAS SNPs with
natural populations and SNP genotyping with natural populations. (5) Functional
identification of the candidate genes and SNPs. (6) Prediction of functional 745 SNPs
on the basis of SNP mutagenesis and phenotype rescue. (7) Exclusion of
non-functional SNPs. (8) Selection of functional SNPs. (9) Designing of effective
markers. (10) Molecular breeding by design. DH, Doubled haploid lines; GWAS,
genome-wide association study; QTL, Quantitative trait locus; SNP, Single nucleotide
polymorphism; RIL, Recombinant inbred lines.
www.plantphysiol.orgon August 26, 2018 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.

Parsed CitationsAlahakoon, U.I., Taheri, A., Nayidu, N.K., Epp, D., Yu, M., Parkin, I., Hegedus, D., Bonham-Smith, P., Gruber, M.Y., 2016. Hairy Canola(Brasssica napus) re-visited: Down-regulating TTG1 in an AtGL3-enhanced hairy leaf background improves growth, leaf trichomecoverage, and metabolite gene expression diversity. Bmc Plant Biol 16, 12.
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Chen, H., Xie, W., He, H., Yu, H., Chen, W., Li, J., Yu, R., Yao, Y., Zhang, W., He, Y., Tang, X., Zhou, F., Deng, X.W., Zhang, Q., 2014. Ahigh-density SNP genotyping array for rice biology and molecular breeding. Molecular Plant 7, 541-553.
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Cheng, F., Liu, S., Wu, J., Fang, L., Sun, S., Liu, B., Li, P., Hua, W., Wang, X., 2011. BRAD, the genetics and genomics database forBrassica plants. Bmc Plant Biol 11, 136.
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Clough, S.J., and Bent, A.F. 1998. Floral dip: a simplified method for Agrobacterium-mediated transformation of Arabidopsis thaliana.Plant J. 16, 735–743.
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Dai, X.M., Zhou, L.M., Zhang, W., Cai, L., Guo, H.Y., Tian, H.N., Schiefelbein, J., Wang, S.C., 2016. A single amino acid substitution in theR3 domain of GLABRA1 leads to inhibition of trichome formation in Arabidopsis without affecting its interaction with GLABRA3. PlantCell Environ 39, 897-907.
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Ehleringer J. R., Mooney H. A. 1978. Leaf hairs: Effects on physiological activity and adaptive value to a desert shrub. Oecologia 37,183-200.
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Galway, M.E., Masucci, J.D., Lloyd, A.M., Walbot, V., Davis, R.W., Schiefelbein, J.W., 1994. The Ttg Gene Is Required to SpecifyEpidermal-Cell Fate and Cell Patterning in the Arabidopsis Root. Developmental biology 166, 740-754.
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Gan, L.J., Xia, K., Chen, J.G., Wang, S.C., 2011. Functional characterization of TRICHOMELESS2, a new single-repeat R3 MYBtranscription factor in the regulation of trichome patterning in Arabidopsis. Bmc Plant Biol 11.
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Harada, E., Kim, J.A., Meyer, A.J., Hell, R., Clemens, S., Choi, Y.E., 2010. Expression profiling of tobacco leaf trichomes identifies genesfor biotic and abiotic stresses. Plant Cell Physiol 51, 1627-1637.
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Hegebarth, D., Buschhaus, C., Wu, M., Bird, D., Jetter, R., 2016. The composition of surface wax on trichomes of Arabidopsis thalianadiffers from wax on other epidermal cells. The Plant journal : for cell and molecular biology 88, 762-774.
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Kim, J., Kim, D.S., Park, S., Lee, H.E., Ahn, Y.K., Kim, J.H., Yang, H.B., Kang, B.C., 2016. Development of a high-throughput SNP markerset by transcriptome sequencing to accelerate genetic background selection in Brassica rapa. Hortic Environ Biote 57, 280-290.
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Larkin, J.C., Oppenheimer, D.G., Pollock, S., Marks, M.D., 1993. Arabidopsis Glabrous1 Gene Requires Downstream Sequences forFunction. The Plant cell 5, 1739-1748.
Pubmed: Author and TitleCrossRef: Author and Title www.plantphysiol.orgon August 26, 2018 - Published by Downloaded from
Copyright © 2018 American Society of Plant Biologists. All rights reserved.

Google Scholar: Author Only Title Only Author and Title
Larkin, J.C., Young, N., Prigge, M., Marks, M.D., 1996. The control of trichome spacing and number in Arabidopsis. Development 122,997-1005.
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Li, F., Kitashiba, H., Nishio, T., 2011. Association of sequence variation in Brassica GLABRA1 orthologs with leaf hairiness. MolBreeding 28, 577-584.
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Maes, L., Inze, D., Goossens, A., 2008. Functional specialization of the TRANSPARENT TESTA GLABRA1 network allows differentialhormonal control of laminal and marginal trichome initiation in Arabidopsis rosette leaves. Plant physiology 148, 1453-1464.
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Nafisi, M., Stranne, M., Fimognari, L., Atwell, S., Martens, H.J., Pedas, P.R., Hansen, S.F., Nawrath, C., Scheller, H.V., Kliebenstein, D.J.,Sakuragi, Y., 2015. Acetylation of cell wall is required for structural integrity of the leaf surface and exerts a global impact on plantstress responses. Frontiers in plant science 6.
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Nayidu, N.K., Kagale, S., Taheri, A., Withana-Gamage, T.S., Parkin, I.A.P., Sharpe, A.G., Gruber, M.Y., 2014. Comparison of Five MajorTrichome Regulatory Genes in Brassica villosa with Orthologues within the Brassicaceae. Plos One 9.
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Oppenheimer, D.G., Herman, P.L., Sivakumaran, S., Esch, J., Marks, M.D., 1991. A Myb Gene Required for Leaf Trichome Differentiationin Arabidopsis Is Expressed in Stipules. Cell 67, 483-493.
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Payne, C.T., Zhang, F., Lloyd, A.M., 2000. GL3 encodes a bHLH protein that regulates trichome development in arabidopsis throughinteraction with GL1 and TTG1. Genetics 156, 1349-1362.
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Rerie, W.G., Feldmann, K.A., Marks, M.D., 1994. The Glabra2 Gene Encodes a Homeo Domain Protein Required for Normal TrichomeDevelopment in Arabidopsis. Gene Dev 8, 1388-1399.
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Schellmann, S., Schnittger, A., Kirik, V., Wada, T., Okada, K., Beermann, A., Thumfahrt, J., Jurgens, G., Hulskamp, M., 2002.TRIPTYCHON and CAPRICE mediate lateral inhibition during trichome and root hair patterning in Arabidopsis. The EMBO journal 21,5036-5046.
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Schnittger, A., Folkers, U., Schwab, B., Jurgens, G., Hulskamp, M., 1999. Generation of a spacing pattern: the role of triptychon intrichome patterning in Arabidopsis. The Plant cell 11, 1105-1116.
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Song, K., Slocum, M.K., Osborn, T.C., 1995. Molecular marker analysis of genes controlling morphological variation in Brassica rapa(syn. campestris). TAG. Theoretical and applied genetics. Theoretische und angewandte Genetik 90, 1-10.
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Symonds, V.V., Godoy, A.V., Alconada, T., Botto, J.F., Juenger, T.E., Casal, J.J., Lloyd, A.M., 2005. Mapping quantitative trait loci inmultiple populations of Arabidopsis thaliana identifies natural allelic variation for trichome density. Genetics 169, 1649-1658.
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
www.plantphysiol.orgon August 26, 2018 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.

Szymanski, D.B., Jilk, R.A., Pollock, S.M., Marks, M.D., 1998. Control of GL2 expression in Arabidopsis leaves and trichomes.Development 125, 1161-1171.
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Tanhuanpaa, P., Erkkila, M., Tenhola-Roininen, T., Tanskanen, J., Manninen, O., 2016. SNP diversity within and among Brassica rapaaccessions reveals no geographic differentiation. Genome 59, 11-21.
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Van Cutsem, E., Simonart, G., Degand, H., Faber, A.M., Morsomme, P., Boutry, M., 2011. Gel-based and gel-free proteomic analysis ofNicotiana tabacum trichomes identifies proteins involved in secondary metabolism and in the (a)biotic stress response. Proteomics 11,440-454.
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Wada, T., Tachibana, T., Shimura, Y., Okada, K., 1997. Epidermal cell differentiation in Arabidopsis determined by a Myb homolog, CPC.Science 277, 1113-1116.
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Wang, S., Kwak, S.H., Zeng, Q., Ellis, B.E., Chen, X.Y., Schiefelbein, J., Chen, J.G., 2007. TRICHOMELESS1 regulates trichomepatterning by suppressing GLABRA1 in Arabidopsis. Development 134, 3873-3882.
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Wang, S., Wang, J.W., Yu, N., Li, C.H., Luo, B., Gou, J.Y., Wang, L.J., Chen, X.Y., 2004. Control of plant trichome development by a cottonfiber MYB gene. The Plant cell 16, 2323-2334.
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Wang, S.C., Chen, J.G., 2008. Arabidopsis Transient Expression Analysis Reveals that Activation of GLABRA2 May Require ConcurrentBinding of GLABRA1 and GLABRA3 to the Promoter of GLABRA2. Plant Cell Physiol 49, 1792-1804.
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Ye, X.L., Hu, F.Y., Ren, J., Huang, S.N., Liu, W.J., Feng, H., Liu, Z.Y., 2016. Fine mapping and candidate gene analysis of Brtri1, a genecontrolling trichome development in Chinese cabbage (Brassica rapa L. ssp pekinensis). Genetics and molecular research : GMR 15.
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Yu, F.Q., Zhang, X.G., Huang, Z., Chu, M.G., Song, T., Falk, K.C., Deora, A., Chen, Q.L., Zhang, Y., McGregor, L., Gossen, B.D.,McDonald, M.R., Peng, G., 2016. Identification of Genome-Wide Variants and Discovery of Variants Associated with Brassica rapaClubroot Resistance Gene Rcr1 through Bulked Segregant RNA Sequencing. Plos One 11.
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Yu, X., Wang, H., Lu, Y.Z., de Ruiter, M., Cariaso, M., Prins, M., van Tunen, A., He, Y.K., 2012. Identification of conserved and novelmicroRNAs that are responsive to heat stress in Brassica rapa. J Exp Bot 63, 1025-1038.
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Yu, X., Wang, H., Zhong, W., Bai, J., Liu, P., He, Y., 2013. QTL mapping of leafy heads by genome resequencing in the RIL population ofBrassica rapa. Plos One 8, e76059.
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Zhang, F., Gonzalez, A., Zhao, M.Z., Payne, C.T., Lloyd, A., 2003. A network of redundant bHLH proteins functions in all TTG1-dependentpathways of Arabidopsis. Development 130, 4859-4869.
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Zhang, J., Lu, Y., Yuan, Y., Zhang, X., Geng, J., Chen, Y., Cloutier, S., McVetty, P.B., Li, G., 2009. Map-based cloning and characterization www.plantphysiol.orgon August 26, 2018 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.

of a gene controlling hairiness and seed coat color traits in Brassica rapa. Plant Mol Biol 69, 553-563.Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Zimmermann, I.M., Heim, M.A., Weisshaar, B., Uhrig, J.F., 2004. Comprehensive identification of Arabidopsis thaliana MYB transcriptionfactors interacting with R/B-like BHLH proteins. Plant Journal 40, 22-34.
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
www.plantphysiol.orgon August 26, 2018 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.





























