Embed Size (px)
Citation preview

General rights Copyright and moral rights for the publications made accessible in the public portal are retained by the authors and/or other copyright owners and it is a condition of accessing publications that users recognise and abide by the legal requirements associated with these rights.
• Users may download and print one copy of any publication from the public portal for the purpose of private study or research. • You may not further distribute the material or use it for any profit-making activity or commercial gain • You may freely distribute the URL identifying the publication in the public portal
If you believe that this document breaches copyright please contact us providing details, and we will remove access to the work immediately and investigate your claim.
Downloaded from orbit.dtu.dk on: Dec 20, 2017
Conversion of the biodiesel by-product glycerol by the non-conventional yeastPachysolen tannophilus
Liu, Xiaoying ; Jensen, Peter Ruhdal; Workman, Mhairi
Publication date:2012
Document VersionPublisher's PDF, also known as Version of record
Link back to DTU Orbit
Citation (APA):Liu, X., Jensen, P. R., & Workman, M. (2012). Conversion of the biodiesel by-product glycerol by the non-conventional yeast Pachysolen tannophilus. Kgs. Lyngby: Technical University of Denmark (DTU).

Conversion of the biodiesel by-product
glycerol by the non-conventional yeast
Pachysolen tannophilus
Ph.D. thesis
Xiaoying Liu
Supervisors:
Assoc. prof. Mhairi Workman
Prof. Peter Ruhdal Jensen
Center for Systems Microbiology
Department of Systems Biology
Technical University of Denmark
Lyngby, Denmark
2012, March

Konvertering af biodiesel affald glycerol
ved ikke-konventionelle gær Pachysolen
tannophilus
Ph.D. afhandling
Xiaoying Liu
vejledere:
Assoc. prof. Mhairi Workman
Prof. Peter Ruhdal Jensen
Center for Systems Microbiology
Institut for Systembiologi
Danmarks Tekniske Universitet
Lyngby, Danmark
2012, mart

I
Preface and Acknowledgements
This dissertation includes the results of my PhD study carried out at the Center for Systems
Microbiology (CSM), Department of Systems Biology in Technical University of Denmark
(DTU) during the period from March 2009 to March 2012. The PhD study was funded by the
European Community’s 7th Framework Research Programme under Grant Agreement
Number 213506 (Project GLYFINERY).
First of all, I would like to thank for my supervisor Peter Ruhdal Jensen who gave the great
opportunity to pursue my PhD study at DTU and guide me along the entire PhD project. I
would like to express my special gratitude to my supervisor Mhairi Workman. Whenever I
had problems with my project or in my daily life, she was always so kind to lead me to the
right direction and solve the problems. During my PhD study, her guidance, good advice,
support, encouragement and discussions regarding problems accompany me along with my
PhD. I also very appreciate that Mhairi helped me a lot with my publications and the PhD
thesis. In addition, thanks for the scientific discussions and help from Uffe Hasbro Mortensen
and Morten Kielland-Brandt at the Center for Microbial Biotechnology.
I would like to thank for my colleagues and friends who gave endless help during my study:
Søren Helmark, Christian Solem, Yuzhou Fan, Line Due Buron, Martin Engelhard Kornholt,
especially the technician Regina Åris Schürmann with assisting some of my lab work. Thank
you very much for the postdoc Arshnee Moodley in Copenhagen University for offering the
opportunity to run PFGE.
Finally, I would like to thank the support and comfort from my family and my boyfriend Tian
Ran Sun when I was struggling with my project and listen to my voice and give me so much
encouragement and confidence.
Without all those lovely and helpful people around, my PhD could not be completed!
Copenhagen, March 2012
Xiaoying Liu

II
Summary
The focus on developing new renewable energy in the transportation sector by the EU has
boosted the production of biodiesel from rapeseed and other vegetable oils in Europe. This
has led to an immense increase in the production of glycerol, which is an inevitable by-
product from the biodiesel production process. Since the volume of the glycerol by-product
has exceeded the current market need, biodiesel producers are looking for new methods for
sustainable glycerol management and improving the competitiveness of the biodiesel
industries. The EU Commission funded GLYFINERY project is one initiative targeted to
development of a novel technology based on biological conversion of the glycerol feedstocks
into known and new advanced liquid biofuels, bioenergy and valuable green chemicals in an
integrated biorefinery concept.
As part of the GLYFINERY project, the objective of this PhD project was to develop a
process for bioconversion of waste glycerol into biofuel ethanol, characterize and optimize
the process. The present thesis comprises of eight chapters. The project background, scope
and aims are introduced in Chapter 1. Besides, the related background knowledge for better
understanding the studies in the following chapters is also introduced in this chapter. Chapter
2-7 are comprised of the experimental results obtained during the whole PhD study.
The well characterized yeast Saccharomycees cerevisiae has been used for fermentation of
alcoholic beverages throughout thousands of years of human history, and is applied in many
areas of modern biotechnology. In this project the interest was in investigating non-
conventional yeasts which had the capability of conversion of glycerol primarily to liquid
biofuels. Chapter 2 i s about the initial results for screening of the potential candidates for
glycerol fermentation. Two candidates Pachia pastoris and Pachysolen tannophilus were
shown to be capable of producing ethanol with glycerol as the sole carbon source. After
growth comparison on glycerol and tests for extracellular metabolites in agitated flasks, P.
tannophilus was selected as the object of further studies for conversion of glycerol to ethanol.
In chapter 3, physiology studies in lab scale fermentation of the ethanol production process
with P. tannophilus were investigated on glycerol. The effect of aeration, pH and nitrogen
source was studied for improving the ethanol production and yield and designing a

III
competitive ethanol production process. The ethanol tolerance of P. tannophilus on glycerol
was studied for further characterizing the ethanol production process. A growth comparison
on crude glycerol and pure glycerol was performed to test if the impurities in the crude
glycerol inhibit the growth of P. tannophilus and affect product formation. Based on
optimized parameters, 28.1 g/L ethanol was produced by a staged batch process, which was
the maximum achieved so far for conversion of glycerol to ethanol by a microbial bioprocess.
The physiology study of ethanol tolerance of P. tannophilus showed that the ethanol
tolerance of this strain was relatively low. The low ethanol tolerance of P. tannophilus might
be the factor which inhibits further improvement of ethanol production process. Chapter 4
describes adaptive evolution studies performed to enhance the ethanol tolerance of P.
tannophilus on glycerol. The adapted strains isolated during the evolution process were
characterised according to the ethanol tolerance, growth rate on glycerol, ethanol production
and growth profile on glycerol.
For better understanding the genetic background, the genomic DNA of P. tannophilus
CBS4044 was isolated and sequenced. The draft genome sequencing results of P. tannophilus
are summarized in chapter 5. Raw data of short reads from genome sequencing results were
assembled together. The protein-coding genes were identified and the putative amino acid
sequences were analysed for the gene function annotation. Pulsed field gel electrophoresis
was performed to predict the chromosome numbers and approximate chromosome sizes in P.
tannophilus.
For the purpose of further improving the yields and production levels of ethanol produced, it
would be beneficial if P. tannophilus could be genetically engineered and the ethanol
synthesis pathway in P. tannophilus could be investigated. The whole-genome sequencing of
P. tannophilus also makes it possible to perform genetic engineering of this strain. Chapter 6
describes the attempts to set up the transformation system in P. tannophilus in order to know
more about the genetic background and further improve the ethanol production process. The
commonly applied methods using antibiotic resistance and auxotrophic markers URA3 were
used for transformation selection.
Since the genome of P. tannophilus CBS4044 was sequenced and the mechanism behind
glycerol metabolism is poorly understood in this strain. In chapter 7 focusses on studying the

IV
genes involved in glycerol metabolism in P. tannophilus, which were predicted by blasting
with the sequences of genes known to have these functions in S. cerevisiae. Quantitative real-
time PCR was performed to unveil the expression pattern of the genes during growth on
glycerol. The glycerol metabolism and pathways in P. tannophilus are discussed. The genes
involved in glycerol transport in P. tannophilus have been cloned and expressed in S.
cerevisae (CEN.PK 113-5D) strains to validate the function of the predicted glycerol
transporter genes.
Finally, the most relevant results from all the studies during the PhD are summarised and
future perspectives for continuing these studies are presented in Chapter 8.

V
Dansk Sammenfatning
Den øgede fokus på udvikling af nye vedvarende energikilder til transportsektoren fra
europæisk side har medført en øget produktion af biodiesel fra raps og andre vegetabilske
olier. Dette resulteret i en betydelig stigning i produktionen af glycerol, som er et uundgåeligt
biprodukt fra produktionen af biodiesel. Mængden af glycerol har efterhånden overskredet
det aktuelle behov på markedet, derfor er producenter af biodiesel på udkig efter nye metoder
til glycerol anvendelse og dermed forbedre konkurrenceevnen for biodiesel producenterne.
EU-kommissionen har finansieret GLYFINERY Projektet, et initiativ rettet mod udvikling af
nye teknologier baseret på biologisk omdannelse af glycerol til kendte og nye
biobrændstoffer, bioenergi og værdifulde grønne kemikalier i et integreret bioraffinaderi
koncept.
Som en del af GLYFINERY projektet, var formålet med dette ph.d.-projekt at udvikle en
proces til biologiskomdannelse af uraffineret glycerol til ethanol, samt karakterisere og
optimere processen. Denne afhandling består af otte kapitler; Kapitel 1 introducere projektets
baggrund, omfang og mål e, desuden er den tilhørende baggrundsviden for bedre at forstå de
undersøgelser i de følgende kapitler også indført i dette kapitel. Kapitel 2-7 er sammensat af
de eksperimentelle resultater opnået i løbet af hele ph.d.-studiet.
Den velkarakteriserede gær Saccharomyces cerevisiae er blevet brugt til fermentering af
alkoholholdige drikkevarer gennem tusinde år af menneskets historie, og anvendes inden for
mange områder af moderne bioteknologi. I dette projekt var interessen for at undersøge ikke-
konventionelle gær, som havde evnen til omdannelsen af glycerol primært til f lydende
biobrændstoffer. Kapitel 2 om handler de første resultater for screening af de potentielle
kandidater til glycerol gæring. To kandidater Pachia pastoris og Pachysolen tannophilus
viste sig at være i stand til at producere ethanol på baggrund af glycerol som eneste
kulstofkilde. Efter vækst sammenligning af glycerol og tests for eksterne metabolitter i
rystekolber blev P. tannophilus valgt som undersøgelsen genstand for yderligere
undersøgelse til omdannelse af glycerol til ethanol proces.
Kapitel 3 om handler fysiologiske undersøgelser af P. tannophilus vokset på glycerol i
laboratorieskala kultiveringer, med henblik på ethanol produktion. Virkningen af beluftning,

VI
pH og nitrogenkilde blev undersøgt for at forbedre produktionen, og udbyttet af ethanol samt
designe en konkurrencedygtig proces til ethanol produktion. P. tannophilus’ ethanol
tolerancemen den vokser på glycerol blev undersøgt for yderligere karakterisering af ethanol
produktionsprocessen. En vækst sammenligning af uraffineret glycerol og ren glycerol blev
udført for at teste om de urenheder i den uraffinerede glycerol har en inhiberende effekt på
væksten, og ethanol produktionen af P. tannophilus. Baseret på optimerede parametre blev
28.1 g/L ethanol fremstillet ved en trinvis batchproces, hvilket var den maksimale hidtidige
til omdannelse af glycerol til ethanol ved en mikrobiel bioprocess.
De fysiologiske undersøgelser af P. tannophilus’ ethanol toleranceviste at denne er relativt
lav, den lave ethanol tolerance P. tannophilus udviser, kan være den faktor der besværliggøre
en yderligere forbedring af ethanol produktionsprocessen. Kapitel 4 beskriver adaptive
evolution eksperimenter udført for at forbedre ethanol tolerancen hos P. tannophilus. De
tilpassede stammer der er blevet isoleret under eksperimentet karakteriseres ifølge ethanol
tolerance, væksthastighed på glycerol, ethanol og vækst profil glycerol.
For bedre forståelsen af den genetiske baggrund blev det genomiske DNA af P. tannophilus
CBS4044 isoleret og sekventeret. Udkastet genomsekvensering. Resultaterne af
sekvenseringen af P. tannophilus er sammenfattet i kapitel 5. Rådata med korte læsninger fra
genomsekvenseringen blev samlet sammen, de protein-kodende gener identificeret, og de
formodede aminosyresekvenser analyseret for genfunktion, og efterfølgende annoteret.
Pulsfeltgelelektroforese blev udført for at estimere kromosomtal og tilnærmelsesvise
kromosom størrelser i P. tannophilus.
Med henblik på yderligere at forbedre udbyttet og produktions niveauer af ethanol, vil det
være fordelagtigt, hvis P. tannophilus kan optimeres genetisk, og ethanol syntesevejen i P.
tannophilus undersøges. En komplet genom sekventering af P. tannophilus vil muliggøre
genetisk manipulation stammen. Kapitel 6 beskriver forsøg på at etablere et transformations
system til P. tannophilus for at bidrage yderligere til den tilgængelige viden om den genetiske
baggrund samt yderligere forbedre ethanol produktionen. De almindeligt anvendte metoder
såsom anvendelse af antibiotika resistens og den auxotrofe markør URA3 blev anvendt til
selektion for transformanter.

VII
Kapitel 7 f okuserer på at forstå og undersøge mekanismen bag glycerol metabolismen i
stammen, vedpå at studere de gener der er involveret i glycerol metabolisme i P. tannophilus.
Disse gener blev forudsagt ved blast mod sekvenser af gener, der vides at have disse
funktioner i S. cerevisiae. Kvantitativ real-time-PCR blev udført for at afsløre
ekspressionsmønsteret af generne under vækst på glycerol. Desuden er generne der
forventeligt er involveret i glycerol transport i P. tannophilus blevet klonet og udtrykt i S.
cerevisiae (CEN.PK 113-5D) for at validere deres funktion. Afslutningsvis diskuteres
glycerol metabolismen, og pathways i P. tannophilus.
I det afsluttende kapitel 8 a dresseres de relevante resultater fra hele ph.d. f orløbet, og
perspektiver for resultaterne, samt fremtidige studier diskuteres.

VIII
List of publications
Paper1
Liu, X., Jensen, P.R., Workman, M. 2012. Bioconversion of crude glycerol feedstocks into
ethanol by Pachysolen tannophilus. Bioresource Technology, 104(0), 579-586.
Paper2
Liu, X., Kaas, R.S, Jensen, P.R., Workman, M. 2012. Draft genome sequence of the yeast
Pachysolen tannophilus CBS 4044. Submitted to Eukaryotic Cell, April, 2012.
Paper3
Liu, X., Jensen, P.R., Workman, M. 2012. Expression and function studies of genes involved
in glycerol transport and metabolism in Pachysolen tannophilus. Submitted to Appl Environ
Microbiol, April, 2012.

IX
Table of Contents
Preface and Acknowledgements ................................................................................................ I
Summary ................................................................................................................................... II
Dansk Sammenfatning .............................................................................................................. V
List of publications ............................................................................................................... VIII
Chapter 1 Introduction ............................................................................................................. 1
1.1 Project overview and aim................................................................................................. 1
1.2 Biodiesel production and the glycerol glut ...................................................................... 3
Biodiesel production process .......................................................................................................... 3
The waste glycerol from biodiesel production ................................................................................ 5
Composition of biodiesel and crude glycerol.................................................................................. 6
1.3 Biotechnology .................................................................................................................. 8
Industrial biotechnology and the Microbial cell factory ................................................................. 8
Process optimization and Strain improvement ................................................................................ 9
1.4 Bioconversion of glycerol .............................................................................................. 10
Biological processes for conversion of glycerol by microorganisms ........................................... 10
Roles of glycerol in yeast metabolism .......................................................................................... 11
Glycerol metabolism ..................................................................................................................... 12
1.5 P. tannophilus as a non-conventional cell factory ......................................................... 16
Nature of P. tannophilus ............................................................................................................... 16
Substrate diversity of P. tannopphilus in biotechnology .............................................................. 17
P. tannophilus as an ethanol producer .......................................................................................... 18
References ............................................................................................................................ 19
Chapter 2 Initial screening results for ethanol producers on glycerol ..................................... 26
Chapter 3 Bioconversion of crude glycerol feedstocks into ethanol by Pachysolen
tannophilus ............................................................................................................................... 37

X
Chapter 4 Adaptive evolution studies for improving ethanol tolerance of Pachysolen
tannophilus CBS4044 ..............................................................
Chapter 5 Draft genome sequence of the yeast Pachysolen tannophilus CBS 4044 ............56...................................................................................................................................................
Chapter 6 Development of a transformation and selection system for Pachysolen tannophilus
.............................................................................................................................................. 60
Chapter 7 Functional studies of genes involved in transport and metabolism of glycerol in
Pachysolen tannophilus ........................................................................................................ 71
Chapter 8 Conclusions and future perspectives .....................................................................93

1
Chapter 1 Introduction
1.1 Project overview and aim
My PhD project forms part of the Europe Commission funded project GLYFINERY. The
background of the whole GLYFINERY project will be introduced in this chapter. The aim
and scope of my PhD project will be pointed out.
Among the different sources for the substitution of fossil fuels, bioethanol and biodiesel are
the most promising sources for biofuels. Biodiesel is a renewable fuel produced from
vegetable oils such as rape seed oil, sunflower seed oil, soybean oil and also used frying oils
or animal fats. Biodiesel has been demonstrated to have great environmental benefits
compared to fossil fuels. It can reduce CO2 emissions, particulate emissions and other
harmful emissions and decrease global warming impacts. In addition, biodiesel has a high
lubricity and fast biodegradability. It can be effectively used either by blending with fossil
diesel fuel or in a pure form as a transport fuel. In Europe, motor manufacturers have tested
biodiesel blended with diesel oil up to 5-10%, or 25-30% or 100% pure. But normally for use
at 100% pure biodiesel, minor modifications to seals and piping in existing engines are
required.
The European Union is the leading biodiesel production region worldwide, representing
about 55-60% of worldwide output and biodiesel accounted for over 75 % of the total
biofuels production in Europe in 2009 (Biofuels platform http://www.biofuels-
platform.ch/en/infos/eu-biodiesel.php). Due to increasing interest in biofuels, the European
biodiesel production has accelerated from approximately 500,000 tons in 1998 to 22 million
tonnes in July 2011 with 254 existing biodiesel facilities. Figure 1 p resents the biodiesel
production trends in the EU from 1998 to 2011 according to the European Biodiesel Board
(EBB) 2011 press release.

2
Figure 1. The biodiesel production trends in the EU from 1998 to 2011 (EBB press release).
Glycerol is the inevitable by-product in biodiesel production. It has seen a proportionally
dramatic increase in production with raising biodiesel production in recent years. In a given
biodiesel production process, approximately 10% of the reaction volume ends up as glycerol.
With the increased global biodiesel production, the problem with glycerol-waste will
continue to grow. The glycerol generated during biodiesel production contains impurities
such as salts, heavy metals, alcohols and water. Glycerol could be utilized in many industries
including cosmetic, soaps, pharmaceuticals, food, drinks and other uses. However, the crude
glycerol from biodiesel production must be purified for the traditional processing of glycerol,
which is not economically feasible for many biodiesel manufacturers. The market for
glycerol now has become saturated and it is considered as a waste by many biodiesel
producers. There is a lack of reliable methods for the efficient management of glycerol waste.
Bioconversion of glycerol into liquid biofuels, green chemicals and bioenergy on the basis of
fermentation processes might provide an efficient solution for sustainable management of
glycerol, which can improve the economics of biodiesel industries.
The GLYFINERY project aims at finding a solution for sustainable management of the
glycerol from both environmental and economic point of view. The target bioproducts
developed by the GLYFINERY concept are liquid biofuels, bioethanol and the long-chain
alcohol biobutanol, the green chemical (1,3-propanediol) and bioenergy in form of
biomethane. The concept of GLYFINERY is analogous to the concept of biorefinery, where
it integrates low cost biomass conversion processes and equipment to produce fuels, power,
heat, and value-added chemicals by virtue of microbes as the cell factory. As part of the

3
GLYFINERY project, the aim of my PhD project was developing a process for bioconversion
of waste glycerol into biofuel ethanol, characterization and optimization of the process.
1.2 Biodiesel production and the glycerol glut
Biodiesel production process
The simplified biodiesel production process is shown in the following flowchart (Figure 2).
The main reaction for converting oil to biodiesel is called transesterification. In the
transesterification process, the triglyceride oils contained in vegetable oils, animal fats, or
recycled greases react with an alcohol (typically methanol or ethanol) to form fatty acid alkyl
monoesters (biodiesel) and glycerol. The reaction requires heat and acid, alkali or enzymatic
catalysts.
Figure 2. An overview of a standard biodiesel production process. Biodiesel and glycerol are the two
main products produced in this process.
The basic biodiesel producing process is a ch emical transesterification reaction converting
triglycerides into fatty acid alkyl monoesters in the presence of a catalyst. An overview of the
stoichiometry of the transesterification reaction can be seen in Figure 3. Since the reaction is
a reversible reaction, the alcohol must be added in excess to drive the reaction towards the
right and ensure complete conversion.

4
Figure 3. Chemical process for methyl ester biodiesel production. The R groups represent fatty acid
radicals.
The most commonly used oils for biodiesel production are from rape seed, sunflower, palm,
canola, cotton seed and jatropha and soy bean. Other low cost feedstocks including waste
cooking oils or animal fats are also used for biodiesel production (Singh & Singh, 2010). The
feedstocks used for biodiesel production should fulfil two requirements: low production costs
and large production scale (Singh & Singh, 2010). In principle, any oleaginous material can
be used as a feedstock. The source of biodiesel usually depends on the crops amenable to the
regional climate. In Europe, rapeseed oil is the most common source for biodiesel while in
tropical countries palm oil is the most common source. In the United States, biodiesel is
mostly made from soybean oil or recycled cooking oils. Animal fats, other vegetable oils, and
other recycled oils can also be used to produce biodiesel, which depends on their costs and
availability. In reality biodiesel producers use a mixture of different vegetable oils. In the
future, blends of all kinds of fats and oils may be used to produce biodiesel.
Before the transesterification process some feedstocks must go through pretreatment, which
depends on the free fatty acid (FFA) concentration. Feedstocks having higher levels of FFA
and water might lead to problems with saponification during biodiesel production with an
alkali catalyst. If the feedstocks contain less than 4% free fatty acids, pretreatment is not
required, this is the case for vegetable oils and some food-grade animal fats. If the feedstocks
contain more than 4% free fatty acids, they must be pretreated in an acid esterification
process, such as when inedible animal fats and recycled greases are used. In this step, the
feedstock is reacted with an alcohol converting the free fatty acids into biodiesel in the
presence of a strong acid catalyst (sulfuric acid). The remaining triglycerides are converted to
biodiesel in the transesterification reaction.

5
Generally there is no di fference in the biodiesel yield with various types of alcohols. The
choice of alcohol to use is mainly an economic concern. Since methanol is often the cheapest
alcohol available and can be obtained in a very anhydrous formulation, it is often the alcohol
of choice. There are three major types of catalysts used for biodiesel production: acid
catalysts (e.g. HCl, H2SO4), alkali catalysts (e.g. NaOH, KOH) and enzymatic catalysts
(lipase). On an industrial scale only the acid and alkali catalysts are used. Alkali catalysts are
the predominant catalyst as they are cheap and give high yields and fast reaction times.
However, if the feedstock contains high levels of FFA’s and water, it might risk of
saponification during the reaction. Acid catalysts have higher tolerance for FFA’s and water
but require higher alcohol to oil ratios and have slower reaction times. Compared to chemical
catalysts, enzymatic catalysts have moderate reaction conditions and do not require the same
amount of excess alcohol ratios. The use of enzymatic catalysts also makes it easier for
downstream processing and product recovery. But, the cost of enzymatic catalysts is still very
high for application on a commercial scale. Alkali catalyzed transesterification with methanol
is the most prevalent process on an industrial scale.
The waste glycerol from biodiesel production
The main source of glycerol on t he market is associated with the increasing production of
biodiesel. In the biodiesel production, the two main by-products are glycerol and esters
(biodiesel). The glycerol phase and biodiesel phase can be separated by settling for several
hours or in some cases by centrifugation since the glycerol is much denser than the biodiesel.
Biodiesel is separated from the glycerol fraction after neutralization, washing and drying, and
then it is ready for use as a fuel. The crude glycerol stream leaving the separator typically is
about 50% glycerol or less and also contains unused catalyst, alcohol, soap, water and salts
(Singh & Singh, 2010). In some cases, the glycerol stream is treated by an inorganic acid
whereby soaps are split into salts. The alcohol in the glycerol phase could be removed by
vacuum flashing. The water will also be removed to form the concentrated glycerol
(approximately 80 - 88% purity) which can be sold to glycerol refiners. The concentrated
glycerol is normally regarded as crude glycerol, which comprises impurities like catalyst and
soaps and it was normally brown colour. Pure glycerol could be used in a variety of
applications in the food and pharmaceutical, cosmetic, textiles, paint, tobacco, waxes, pulp
and paper, leather industries (Wang et al., 2001). However, the purification of this crude
glycerol is not industrially feasible due to the high cost. Currently, only combustion is

6
considered as the utilized way of disposing crude glycerol. However, the salts present in the
crude glycerol create significant amounts of ash in the boiler and formation of acrolein by
thermal decomposition of glycerol is a concern. In view of the economic and environmental
concerns, therefore, utilization of crude glycerol in biorefineries can represent an alternative
and environmental friendly way to improve the economics of the biodiesel industry as well
as providing a means to handle the increasing increment of glycerol waste streams (Yazdani
& Gonzalez, 2007). Bioconversion of glycerol into liquid biofuels, green chemicals and
bioenergy on the basis of biological fermentation processes can provide an efficient solution
for sustainable management of glycerol (Lynd et al., 2005).
Composition of biodiesel and crude glycerol
The composition of biodiesel is closely related to the composition of the feedstocks used
since biodiesel production is a simple chemical transesterification reaction. Therefore, it
means that the biodiesel produced from animal fats which contain higher saturated fatty acid
or other saturated sources often have difficulty in achieving desired cold flow properties. This
problem can be partly remedied by the right choice of alcohol.
The composition of the crude glycerol depends on the parent feedstocks employed and the
biodiesel production process used. These characteristics together with the extent of post-
production purification of the crude glycerol, determines the final characteristics of the
commercially available glycerol feedstock. In general, the composition of the glycerol
depends on the process used and is varied from different biodiesel producers. Chemical and
physical properties of the crude glycerol from seven different vegetable oils: mustard,
rapeseed, canola, crambe, soybean, and waste cooking oils (WCO) were characterized by
Thompson and He (2006). In the final analysis there was very little variation with the
exception of WCO. The viscosity of the crude glycerol prior to any treatment ranged from
8.46 to 8.80 cs (centistokes) and 26.5 cs for the WCO. The carbon content averaged about
25% and the metals Ca, K, Mg, Na, P, and S were present in small quantities from 4 to 163
ppm with the exception of sodium, which averaged just over 1%. Protein levels ranged from
0.06% to 0.44%. Fat content ranged from 1% to 13% and carbohydrates ranged from 75% to
83%. Most crude glycerol feedstock falls within the range of 60-90% glycerol w/w with
varying amounts of the other components.

7
In the GLYFINERY project, three different batches of crude glycerol Batch 1 (B1), Batch 2
(B2), Batch 3 (B3) were received from the Slovakian biodiesel producer MEROCO A.S. The
details of the batches used for biodiesel production are listed below:
Figure 4. Samples of three different batches of crude glycerol B1, B2 and B3 from Meroco A.S.
Each batch was derived from different production runs. The crude glycerol feedstock
contains up to 7% ash, 0.5% methanol, 10% water and 82 ± 5% glycerol. A picture of the
three batches can be seen in Figure 4. Although the B1 and B3 are both based on 100% rape
seed oil, B1 seems much lighter, and thus possibly cleaner, compared to B3 and B2. This is a
indication that the composition of the crude glycerol varied even between different process of
the same biodiesel producer. The three batches were subjected to ion chromatography, gas
chromatography and high pressure liquid chromatography analytical procedures (HPLC). The
analysis with ion chromatography quantifies the amount of compound present in the crude
glycerol. The results can be seen in Table 1.
Table 1: Ion chromatography analysis results of the three batches of.crude glycerol: B1, B2 and B3.
It can be seen in Table 1 that chloride and citrate are present in fairly large amounts
compared to other components in all the three batches. From the analysis results of gas
chromatography, one peak is identified in sample B2 which was not present in the other
• B1: Based on 100% rape seed oil feedstock
• B2: Based on a mix of 90% rape seed oil
and 10% waste cooking oil
• B3: Based on 100% rape seed oil feedstock
B1 B2 B3

8
samples B1 and B3 (results not shown here). Further analysis would be needed to determine
the identity of this peak. Based on HPLC analysis, more peaks are evident in samples B2 and
B3 that are not present in B1 (figure not shown), which indicates that more complexity exists
in the samples B2 and B3 compared to B1. This corresponds to the visual inspection that the
first batch B1 looks much cleaner while the batches B2 and B3 appear more turbid (Figure 4).
More analysis is needed to determine the identity of the unknown compounds. There seems to
be a fair amount of variance within the batches of glycerol received from MEROCO A.S.
1.3 Biotechnology
Industrial biotechnology and the microbial cell factory
With the development of human civilization, the world in the future will be faced with an
energy crisis arising from rapid consumption of fossil fuel, global environmental pollution
and food crisis. Biotechnology could be one of the solutions to alleviate the problems for
sustainable development. Industrial biotechnology is often known as “white biotechnology”,
which is the application of microbial technology for processing of renewable resources to
produce biofuels, biomaterials and biochemical. Biotechnology could diminish our
dependency on fossil fuel, save energy in production processes and significantly reduce CO2
emissions (Sabra et al., 2010).
Long before the discovery of microorganisms, the yeast Saccharomyces cerevisiae had been
used for baking and fermentation of alcoholic beverages throughout thousands years of
human history. Different microorganisms were also exploited to produce cheeses, pickled
food, vinegar and soy sauce to meet human needs. Modern industrial biotechnology is
utilized to produce a bulk of fine chemicals, biofuels, biodegradable plastics, enzymes,
pharmaceuticals and food ingredients by microbial fermentation. The multi-step chemical
synthesis by the traditional chemistry industry could be replaced with a single step with
microbial as the cell factory. In recent years, several widely used microorganisms like the
filamentous fungi Aspergillus oryzae and Aspergillus niger, the bacterium E. coli and the
yeast S. cerevisiae have been used as cell factories for production of desired products.
With realizing the importance of bio-based production, the biorefinery concept analogous to
the petroleum refinery has emerged, where it integrates low cost biomass conversion
processes and equipment to produce fuels, power, heat, and value-added chemicals by virtue

9
of microbes as cell factories. The GLYFINERY project aims to achieve conversion of the low
cost glycerol feedstock into new bioproducts, which fits to the concept of biorefinery. In
addition, lignocellulosic biomass particularly agricultural and forestry residues is very
important low cost biomass used in the biorefinery. These are abundant, readily available and
renewable resources. To develop a robust and efficient production process in the biorefinery,
the characteristics concerned with the potential cell factory are:
• Growth rate on the low cost substrate;
• Tolerance towards elevated concentrations of substrate;
• Production of the desirable value-added products;
• Tolerance towards elevated concentrations of products;
• Substrate conversion efficiency;
• Products yields;
• Limited formation of by-products.
Process optimization and Strain improvement
The microbial fermentation platform is employed in biorefinery schemes to achieve the
conversion process. By controlling the process operating conditions and the medium
composition, it is possible to regulate the pattern of fermentation products in the fermentation
process. The bioreactor performance could be monitored under different conditions. In order
to optimize the production process, fermentation can be performed under different conditions
and at a variety of scales. In addition, different modes of fermentation such as batch, fed-
batch, continuous operation in chemostats could be operated for improving the production of
the compound of interest.
In order to further optimize the process and improve the strain performance for the desired
cellular properties, it is suitable to manipulate the microbial strains by genetic modification.
Traditional mutagenesis strategies by random mutagenesis and screening have been applied
for decades to increase product yield, production level, productivity, and elimination of the
side products. Mutagens such as chemical agents and short wavelength radiation (UV), X-
ray ionizing radiation have been extensively used. However, the screening and selection
process for isolating of stable mutants with desired characteristics demand in terms of time
and labor. Even though the mutants with desired properties may be obtained, the underlying

10
mechanisms may still be unknown, which is not beneficial for further research and
development.
The development of metabolic engineering has facilitated the expansion of industrial
biotechnology, which utilizes genetic tools to manipulate microbial metabolism to improve
the production of desired compounds. Metabolic engineering is through the modification of
biochemical reactions to improve the product formation and cellular properties by the use of
recombinant DNA technology. The applications of metabolic engineering greatly depend on
available molecular or functional knowledge. Compared to the traditional random
mutagenesis and screening, metabolic engineering is based on rational and directed genetic
modification, which improves the microbial production process in a more efficient way. In
addition, adaptive evolution, as a complementary strategy for strain development and process
optimization, simulates nature’s engineering principle where populations of cells adapt to
their environment over many generations by natural selection. During the evolutionary
process, the microbial cells undergo random mutation, recombination and continuous
evolution.
1.4 Bioconversion of glycerol
Biological processes for conversion of glycerol by microorganisms
A variety of (typically anaerobic) bacteria such as Klebsiella, Citrobacter, Enterobacter,
Clostridium, Lactobacillus, Bacillus, Propionibacterium, and Anaerobiospirillum have been
shown to be capable of fermenting glycerol. Glycerol can be fermented to 1,3-propanediol,
acetate, butyrate, acetone, butanol, ethanol, succinate and lactate by various species of
clostridia (Chatzifragkou et al., 2011; Dabrock et al., 1992). Anaerobic fermentative
production of 1,3-propanediol was considered as the most promising option for the biological
conversion of glycerol. It can be used for the synthesis of polyester with the necessary
properties for application in the textile industries. Pseudomonas aeruginosa was shown to
produce rhamnolipid which is a kind of biosurfactant (Silva et al., 2010). E. coli has been
genetically engineered for efficient conversion of crude glycerol into ethanol with the
coproducts hydorgen and formate (Shams Yazdani & Gonzalez, 2008). However, application
in industry of some of those organisms has been limited due to issues that include
pathogenicity, the need for strict anaerobic conditions and supplementation with rich

11
nutrients. The lack of genetic tools and physiological knowledge necessary for their
effective manipulation has also hampered progress.
A number of yeast strains also exhibit the ability to grow on g lycerol and produce value
added products. Glycerol can be assimilated into organic acids and single cell oil by
Yarrowia lipolytica (Makri et al., 2010; Papanikolaou et al., 2002). The basidiomycete yeast
Pseudozyma antarctica was reported to efficiently produce mannosylerythritol lipids as
glycolipid biosurfactants from glycerol (Morita et al., 2007). Glycerol has also been used as
the carbon source for producing biomass and recombinant proteins in Pichia pastoris (Celik
et al., 2008; Fieldhouse et al., 2009). Resting cells of Candida magnolia produce mannitol
from glycerol under aerobic conditions (Khan et al., 2009). Arabitol can be produced from
glycerol by Debaryomyces hansenii SBP-1 (Koganti et al., 2011). The industrial work horse
S. cerevisiae has been genetically engineered to produce and improve ethanol production
from glycerol (Yu et al., 2010). S. cerevisiae was also engineered to be as a platform for
production of fatty acid ethyl esters (FAEEs) from glycerol by esterifying exogenous fatty
acids with endogenously produced ethanol (Yu et al., 2012).
Roles of glycerol in yeast metabolism
Glycerol is involved in different metabolic pathways and plays an important role in yeast
metabolism. Glycerol can be utilized as the sole carbon and energy source for cell growth
under aerobic conditions and also can be formed as a product by many different types of
yeast. In S. cerevisiae, glycerol is formed with ethanol production as a by-product to maintain
the cytosolic redox balance and consume cellular NADH especially under anaerobic
conditions (Albertyn et al., 1994; Nevoigt & Stahl, 1997; van Dijken & Scheffers, 1986). In
addition to the function for maintaining the redox balance, the production of glycerol also has
the function of protecting against osmotic stress (Blomberg & Adler, 1989; Van Eck et al.,
1993). When cells are exposed to osmotic stress, the response appears to be the formation and
intracellular accumulation of osmoprotective solutes (osmolytes) such as glycerol, arabitol,
mannitol, and erythritol to survive in stressful environments (Kayingo et al., 2001; Van Eck
et al., 1993; Yancey et al., 1982). For example, Zygosaccharomyces rouxii releases glycerol
and also arabitol as the osmolytes upon h ypo-osmotic shock (Kayingo et al., 2001). Yeast
cells have developed adaptive mechanisms to control glycerol flux within limits suitable for
growth. These mechanisms include modulation of glycerol formation, degradation, and

12
increased retention by regulation glycerol transport under osmotic stress (Kayingo et al.,
2001). For instance, the production of glycerol is stimulated and accumulated intracellularly
under hyper-osmotic stress, and accumulated glycerol is secreted extracellularly when the
external osmolarity decreases (Hohmann, 2002). Rapid export of glycerol is an important
event in adaptation to low osmolarity.
In addition, glycerol is also involved in lipid metabolism in some yeasts (Meesters et al.,
1996; Nevoigt & Stahl, 1997) as well as in the regulation of inorganic phosphate recycling
(Alonso-Monge et al., 2003; Ansell et al., 1997). Moreover, glycerol appears to play a
protective role against heat shock (Siderius et al., 2000). It is crucial to understand the ways
glycerol crosses the cell membrane and how it is involved in different metabolic pathways
and cell responses.
Glycerol metabolism
The cell membrane is the first and also the last barrier for glycerol movement through the
cells. The existence of specific systems responsible for glycerol transport across the cell
membrane has been shown by several studies (Ferreira et al., 2005; Kayingo et al., 2009;
Sutherland et al., 1997; Tamás et al., 1999).
In E. coli, it w as investigated that glycerol was transported into the cells by faciliated
diffusion with the glycerol facilitator. It was thought that glycerol facilitator and glycerol
kinase are involved in the transporting of external glycerol into cellular metabolism (Voegele
et al., 1993). The glycerol facilitator GlpF is thought to function as a carrier to form a
selective pore in the cytoplasmic membrane, whereas the kinase traps the glycerol inside the
cell as glycerol-3-phosphate (Voegele et al., 1993). GlpF is also responsible transporting poly
alcohols and urea. The influx of glycerol through GlpF is 100- to 1000- fold greater than
expected for a transporter. Glycerol degradation may occur through oxidative and reductive
pathways in Klebsiella, Citrobacter, Clostridium and Enterobacter. In the oxidative pathways,
glycerol could be assimilated by a glycerol dehydrogenase and a dihydroxyacetone kinase to
dihydroxy-acetone phosphate (DHAP), which finally reaches the glycolytic pathway (Daniel
et al., 1995; Macis et al., 1998). In the reductive branch, glycerol is converted into 3-
hydroxypropionaldehyde (3-HPA) by coenzyme B12-dependent glycerol dehydratase and
related diol dehydratases (Talarico et al., 1990). The 3-HPA can be subsequently reduced to

13
1,3-propanediol (1,3-PDO) by a NADH-linked 1,3-propanediol dehydrogenase (Talarico et
al., 1990) or can be oxidized to 3-hydroxypropionic acid (3-HP) (Lonvaud-Funel, 2002).
For glycerol metabolism in yeast, S. cerevisiae was utilized as a model to investigate the
glycerol metabolism and regulation mechanisms (Brisson et al., 2001). It has been shown that
glycerol accumulation and export during osmoregulation in S. cerevisiae was controlled by a
MIP family membrane channel protein Fps1p (Tamás et al., 1999). The physiological role of
the facilitator Fps1p in S. cerevsiaie was described to be glycerol export rather than uptake
during hypo-osmotic shock. It appears that Fps1p channel closes during hyper-osmotic
conditions and opens during hypo-osmotic conditions thereby controlling the accumulation
and release of glycerol during osmoregulation. Similar transport systems and Fps1p
homologues have been found in many other yeasts (Kayingo et al., 2001; Neves et al., 2004).
Besides, Lages and Lucas (1997) discovered an active transport system with a high affinity
uptake for glycerol in S. cerevisiae. The active transport was described to be the proton
symport type and was shown to be under glucose repression and inactivation. The glycerol
proton symporter in S. cerevisiae is found to be encoded by STL1which is a member of the
sugar transporter family (Ferreira et al., 2005). It was demonstrated in S. cerevisae that
glycerol enters the cell by two different mechanisms: a low affinity transport system with
facilitated diffusion encoded by FPS1 and a high affinity proton symport system encoded by
STL1 (Figure 5). Active glycerol uptake systems have also been shown to be present in other
yeasts species like halotolerant yeasts Debaryomyces hansenii (Lucas et al., 1990), Pichia
sorbitophila (Lages & Lucas, 1995), Candida versatilis (Silva-Graça & Lucas, 2003) and in
Zygosaccharomyces rouxii (Zyl et al., 1990) and Candida albicans (Kayingo et al., 2009). In
these yeasts, glycerol was found to be actively transported along with protons or sodium ions.
For glycerol dissimilation it is known that two pathways exist in yeasts: the respiratory
pathway and the fermentative pathway. The first pathway is started by a phosphorylation of a
glycerol kinase and followed by a FAD-dependent glycerol 3-phosphate dehydrogenase on
the outer surface of the mitochondrial inner membrane (Klingenberg, 1970). The
fermentative pathway includes glycerol dehydrogenase and dihydroxyacetone kinase (May et
al., 1982). Dihydroxyacetone phosphate is formed as the glycolytic intermediate after the
two-branch pathways. In some yeast strains, all four enzymes in both pathways are present,
but only one pathway functions for glycerol dissimilation. In some strains, the enzyme
activity related with glycerol catabolism could be measured, but the strains are unable to

14
ferment glycerol or consume glycerol at very low rate. The wine yeast S. cerevisiae is able to
utilize glycerol as sole source of carbon. The catabolic pathway of glycerol involves a
glycerol kinase encoded by GUT1 (Pavlik et al., 1993) and a glycerol 3-phosphate
dehydrogenase encoded by GUT2 (Rønnow & Kielland-Brandt, 1993) (Figure 5). So far, it
has been shown that glycerol is degraded by the respiration pathway and the mutants lacking
one of the two genes are incapable of utilizing glycerol. The GUT1 and GUT2 genes were
repressed on glucose at the transcriptional level (Pavlik et al., 1993; Rønnow & Kielland-
Brandt, 1993). Although the fermentative pathway was also discovered to be present in S.
cerevisise, the function of them is unknown.
Glycerol H+ - Symporter STL1
FPS1 Glycerol faciliator
Glycerol Glycerol-3-phosphate
Dihydroxyacetone phosphate
GUT1 GUT2
GPP1/GPP2 GPD1/GPD2
Dihydroxyacetone Gcy1 Dak1/Dak2
Glycolysis
Ethanol Acetate
Lipid
H+ - Symporter STL1
FPS1 Glycerol faciliator
Figure 5. Glycerol Metabolism pathways in S.cerevisiae

15
Glycerol is synthesized with two steps by reduction of dihydroxyacetone phosphate to
glycerol 3-phosphate by a NAD dependent cytosolic G3P dehydrogenase, followed by
dephosphorylation by a specific phosphatase (Gancedo et al., 1968). In S. cerevisae, each
enzyme has two isoenzymes and encoded by GPD1/2 and GPP1/2 (Figure 5). The expression
of GPD1 at the mRNA level was induced under osmotic stress, however GPD2 gene was
constitutively expressed at a particularly low level (Ansell et al., 1997; Ohmiya et al., 1995).
The two genes GPP1 and GPP2 show 95% amino acid identity. Gpp2p was induced under
osmotic stress, while the concentration of Gpp1p in cells was not affected by changes of
external osmolarity (Norbeck et al., 1996). The synthesis of glycerol via glycerol-3-phosphate
is very common in yeasts since GPD1 or GPD2 genes have been cloned and sequenced from
many types of yeasts (Wang et al., 2001). Glycerol-3-phosphate and dihydroxyacetone
phosphate also function as the important metabolic intermediates for synthesis of other
substances besides glycerol. For instance, glycerol-3-phosphate and dihydroxyacetone
phosphate could be used as precursors for glycerolipids and phospholipids synthesis
(Athenstaedt et al., 1999; Daum et al., 1998; Racenis et al., 1992).

16
1.5 P. tannophilus as a non-conventional cell factory
Nature of P. tannophilus
P. tannophilus was selected as the host for ethanol production from glycerol after screening
of several yeast strains, discussed in detail in Chapter 2. H ere a general overview of this
species is summarized as basic knowledge for further study.
P. tannophilus was initially isolated from wood extracts used in leather tanning and described
by Boidin and Adzet in 1957 (Kurtzman, 1983). P. tannophilus was identified as a yeast and
the vegetative growth of P. tannophilus is mainly by budding and usually has one or two
buds as do many other types of yeast. Under certain conditions, it may form pseudohyphe.
However, P. tannophilus is one of the most unusual yeasts because of the unique way of asco
formation (Jeffries & Kurtzman, 1994). Asco formation starts when a vegetative cell
produces a stout tube which may be quite short or up to 60 µm in length and either straight or
curved (Figure 6a and 6b). The tip of the tube enlarges to form the ascus and the tube could
be regarded as an ascophore (Slininger et al., 1987). P. tannophilus is the only yeast known
so far to produce this unique structure. Four hemispheroidal ascospores are released from the
asci by the deliqueses of the ascus wall. Once this happens, the ascus has formed within a V-
shaped notch at the end of the ascophore. Ascophore walls normally are greatly thickened
and refractile, which make ascopores easily identified under microscope (Figure 6b). Asci
may be conjugated or unconjugated, and this is strain dependent to some extent. Cells
forming conjugated asci are haploid whereas unconjugated asci result from diploid cells. The
species of P. tannophilus is homothallic because single isolated ascospores produce
sporogenous colonies (Slininger et al., 1987).

17
Figure 6a. Life cycles of P. tannophilus cultures (Slininger et al., 1987); 6b. Asci with tubelike
ascophores under the microscope (Kurtzman, 1983).
Substrate diversity of P. tannopphilus in biotechnology
Plant biomass is considered as one of the most abundant renewable resources and refers
largely to lignocellulosic material as this makes up the majority of the cheap and abundant
nonfood materials available from plants. Plant biomass is seen as a promising source of
material for fuels and raw materials. Furthermore, glycerol is also regarded as a ch eap
substrate because of the increasing production of biodiesel as explained in section1 and 2 in
this chapter.
P. tannophilus was the first yeast shown to be capable of fermenting xylose sugars to ethanol
(Slininger et al., 1982) and the xylose utilisation pathway has been extensively studied in this
organism (Sathesh-Prabu & Murugesan, 2011; Slininger et al., 1987; Zhao et al., 2010). P.
tannophilus is one of the few yeasts that can ferment xylose to ethanol. Additionally, it was
reported that P. tannophilus also could ferment all the common sugars glucose, mannose and
galactose except L-arabinose occurring in the hemicelluloses hydrolysate mixture (Slininger
et al., 1987). Besides, it was reported that P. tannophilus could accumulate 4g/L ethanol on
glycerol under aerobic growth in a previous study (Maleszka et al., 1982). However, the
conditions for ethanol production were not precisely defined or controlled and the physiology
during growth on glycerol has not been extensively studied in this organism. P. tannophilus
has certain unique qualities that make it as the potential cell factory and worth of study as it
can naturally utilize xylose and glycerol, while the wild type of the brewing and baking yeast
a b

18
S. cerevisiae cannot utilize xylose (Olsson & Hahn-Hägerdal, 1993) and uses glycerol very
slowly (Ochoa-Estopier et al., 2011).
P. tannophilus as an ethanol producer
As P. tannophilus was the first yeast identified to have a significant capacity to convert
xylose to ethanol, most of previous studies of this species were about the ethanol production
process from xylose.
Ethanolic fermentation is traditionally regarded as occurring anaerobically like in S.
cerevisiae. It was noted that the cell growth of the xylose fermentation by P. tannophilus
depended on a erobic conditions (Slininger et al., 1987). A functioning mitochondrion is
essential to the growth of P. tannophilus. None of the xylose fermenting strains has been
shown capable of significant anaerobic growth on a ny substrate including glucose. By
addition of oxygen the fermentation rate of many yeasts could be enhanced and this
phenomenon is referred to as either a negative Pasteur effect or Custer’s effect (Slininger et
al., 1987). It was hypothesized that Custer’s effect might be due to a shortage of NAD
brought about by an imbalanced intracellular redox system. P. tannophilus displays a
Custer’s effect and an NAD shortage also diminished production of ethanol from xylose by P.
tannophilus (Slininger et al., 1987). Like in many yeasts and filamentous fungi, xylose is
converted to xylitol by a NADPH-linked reductase. By an NAD-linked reductase xylitol
dehydrogenase xylitol is further converted to xylulose in P. tannophilus. Xylulose is then
phosphorylated to xylulose-5-phosphated by xylulose kinase. It was shown that aeration was
necessary for successful induction of xylose reductase, xyilitol dehydrogenase and xylulose
kinase (Neirinck et al., 1982). It was demonstrated that with the decreasing of aeration rate
the ratio of NADH- to NADPH-linked xylose reductase activity increased (Verduyn et al.,
1985).
For the ethanol production process, the fermentation rate and the maximum ethanol
concentration accumulated ultimatedly depend on cellular resistance to ethanol toxicity. For
the xylose fermentation process by P. tannophilus, the specific ethanol production and xylose
consumption rates were affected when ethanol concentration was greater than 20 g/L . The
maximum growth rate of P. tannophilus on xylose without ethanol was 0.31 h-1. However, it
dropped to 0.08 w ith 34 g/L ethanol present in the medium. The maximum of ethanol

19
produced on xylose was 38 g/L at 25°C and P. tannophilus was able to produce > 50g/L
ethanol when glucose was the substrate.
Studies referring to the genetic engineering for further enhancing the ethanol production of
ethanol are few. Only one article has been published about transformation of the yeast P.
tannophilus by Wdelock and Thornton (1989). It was shown that lithium treated hexose
negative mutants of P. tannophilus have been successfully transformed with YRp7 plasmid.
However, the transformants lost the ability to grown on glucose after sub-culturing on non-
selective medium (Wedlock & Thornton, 1989). More information about the genetic
background and the effective genetic engineering tools of P. tannophilus are needed for
learning more about the strains and further improving the process. So far, a u seful
transformation system is still absent in P. tannophilus. Work in this project on developing a
transformation system in P. tannophilus is described in chapter 6.
References
A, L.-F. 2002. Lactic acid bacteria in winemaking: Influence on s ensorial and hygienic
quality. in: Progress in Industrial Microbiology, (Eds.) P. Singh Ved, D. Stapleton
Raymond, Vol. Volume 36, Elsevier, pp. 231-262.
Albertyn, J., Hohmann, S., Thevelein, J.M., Prior, B.A. 1994. G PD1, which encodes
glycerol-3-phosphate dehydrogenase, is essential for growth under osmotic stress in
Saccharomyces cerevisiae, and its expression is regulated by the high-osmolarity
glycerol response pathway. Molecular and Cellular Biology, 14(6), 4135-4144.
Alonso-Monge, R., Navarro-García, F., Román, E., Negredo, A.I., Eisman, B., Nombela, C.,
Pla, J. 2003. The Hog1 Mitogen-Activated Protein Kinase Is Essential in the
Oxidative Stress Response and Chlamydospore Formation in Candida albicans.
Eukaryotic Cell, 2(2), 351-361.
Ansell, R., Granath, K., Hohmann, S., Thevelein, J.M., Adler, L. 1997. The two isoenzymes
for yeast NAD+-dependent glycerol 3-phosphate dehydrogenase encoded by GPD1
and GPD2 have distinct roles in osmoadaptation and redox regulation. EMBO J,
16(9), 2179-2187.
Athenstaedt, K., Weys, S., Paltauf, F., Daum, G. 1999. Redundant Systems of Phosphatidic
Acid Biosynthesis via Acylation of Glycerol-3-Phosphate or Dihydroxyacetone

20
Phosphate in the Yeast Saccharomyces cerevisiae. Journal of Bacteriology, 181(5),
1458-1463.
Blomberg, A., Adler, L. 1989. R oles of glycerol and glycerol-3-phosphate dehydrogenase
(NAD+) in acquired osmotolerance of Saccharomyces cerevisiae. Journal of
Bacteriology, 171(2), 1087-1092.
Brisson, D., Vohl, M.-C., St-Pierre, J., Hudson, T.J., Gaudet, D. 2001. Glycerol: a neglected
variable in metabolic processes? BioEssays, 23(6), 534-542.
Celik, E., Ozbay, N., Oktar, N., Calık, P. 2008. Use of Biodiesel Byproduct Crude Glycerol
as the Carbon Source for Fermentation Processes by Recombinant Pichia pastoris.
Industrial & Engineering Chemistry Research, 47(9), 2985-2990.
Chatzifragkou, A., Papanikolaou, S., Dietz, D., Doulgeraki, A., Nychas, G.-J., Zeng, A.-P.
2011. Production of 1,3-propanediol by Clostridium butyricum growing on biodiesel-
derived crude glycerol through a non-sterilized fermentation process. Applied
Microbiology and Biotechnology, 91(1), 101-112.
Dabrock, B., Bahl, H., Gottschalk, G. 1992. P arameters Affecting Solvent Production by
Clostridium pasteurianum. Applied and Environmental Microbiology, 58(4), 1233-
1239.
Daniel, R., Stuertz, K., Gottschalk, G. 1995. Biochemical and molecular characterization of
the oxidative branch of glycerol utilization by Citrobacter freundii. Journal of
Bacteriology, 177(15), 4392-401.
Daum, G., Lees, N.D., Bard, M., Dickson, R. 1998. Biochemistry, cell biology and molecular
biology of lipids of Saccharomyces cerevisiae. Yeast, 14(16), 1471-1510.
Ferreira, C., van Voorst, F., Martins, A., Neves, L., Oliveira, R., Kielland-Brandt, M.C.,
Lucas, C., Brandt, A. 2005. A Member of the Sugar Transporter Family, Stl1p Is the
Glycerol/H+ Symporter in Saccharomyces cerevisiae. Molecular Biology of the Cell,
16(4), 2068-2076.
Fieldhouse, R., MacLennan, D.F., MacLennan, D.G., MacLennan, M.E. 2009. P ROCESS
FOR PREPARING NUTRITIONAL, THERAPEUTIC OR ORGANOLEPTIC
PRODUCTS FROM CRUDE GLYCEROL, Google Patents.
Gancedo, C., Gancedo, J.M., Sols, A. 1968. Glycerol Metabolism in Yeasts. European
Journal of Biochemistry, 5(2), 165-172.
Hohmann, S. 2002. Osmotic Stress Signaling and Osmoadaptation in Yeasts. Microbiology
and Molecular Biology Reviews, 66(2), 300-372.

21
Jeffries, T.W., Kurtzman, C.P. 1994. S train selection, taxonomy, and genetics of xylose-
fermenting yeasts. Enzyme and Microbial Technology, 16(11), 922-932.
Kayingo, G., Kilian, S., Prior, B. 2001. C onservation and release of osmolytes by yeasts
during hypo-osmotic stress. Archives of Microbiology, 177(1), 29-35.
Kayingo, G., Martins, A., Andrie, R., Neves, L., Lucas, C., Wong, B. 2009. A permease
encoded by STL1 is required for active glycerol uptake by Candida albicans.
Microbiology, 155(5), 1547-1557.
Khan, A., Bhide, A., Gadre, R. 2009. Mannitol production from glycerol by resting cells of
Candida magnoliae. Bioresource Technology, 100(20), 4911-4913.
Klingenberg, M. 1970. Localization of the Glycerol-Phosphate Dehydrogenase in the Outer
Phase of the Mitochondrial Inner Membrane. European Journal of Biochemistry,
13(2), 247-252.
Koganti, S., Kuo, T., Kurtzman, C., Smith, N., Ju, L.-K. 2011. Production of arabitol from
glycerol: strain screening and study of factors affecting production yield. Applied
Microbiology and Biotechnology, 90(1), 257-267.
Kurtzman, C. 1983. Biology and physiology of the Pentoses and Lignin, Vol. 27, Springer
Berlin / Heidelberg, pp. 73-83.
Lages, F., Lucas, C. 1995. Characterization of a glycerol/H+ symport in the halotolerant yeast
Pichia sorbitophila. Yeast, 11(2), 111-119.
Lages, F., Lucas, C. 1997. C ontribution to the physiological characterization of glycerol
active uptake in Saccharomyces cerevisiae. Biochimica et Biophysica Acta (BBA) -
Bioenergetics, 1322(1), 8-18.
Lonvaud-Funel, A., 2002. Lactic acid bacteria in winemaking: influence on s ensorial and
hygienic quality. In: Singh, V.P., Stapleton, R.D. (Eds.), Biotransformations:
bioremediation technology for health and environmental protection. Progress in
Industrial Microbiology, vol. 36. Elsevier Science, Amsterdam, pp. 231–262.
Lucas, C., Da Costa, M., Van Uden, N. 1990. Osmoregulatory active sodium-glycerol co-
transport in the halotolerant yeast Debaryomyces hansenii. Yeast, 6(3), 187-191.
Lynd, L., Wyman, C., Laser, M., Johnson, D., Landucci, R. 2005. Strategic biorefinery
analysis: Review of existing biorefinery examples. NREL Subcontract Report
(NREL/SR-510-34895).
Macis, L., Daniel, R., Gottschalk, G. 1998. Properties and sequence of the coenzyme B12-
dependent glycerol dehydratase of Clostridium pasteurianum. FEMS Microbiology
Letters, 164(1), 21-28.

22
Makri, A., Fakas, S., Aggelis, G. 2010. M etabolic activities of biotechnological interest in
Yarrowia lipolytica grown on g lycerol in repeated batch cultures. Bioresource
Technology, 101(7), 2351-2358.
Maleszka, R., Wang, P., Schneider, H. 1982. E thanol production from D-galactose and
glycerol by Pachysolen tannophilus. Enzyme and Microbial Technology, 4(5), 349-
352.
May, J.W., Marshall, J.H., Sloan, J. 1982. G lycerol Utilization by Schizosaccharomyces
pombe: Phosphorylation of Dihydroxyacetone by a Specific Kinase as the Second
Step. Journal of General Microbiology, 128(8), 1763-1766.
Meesters, P.A.E.P., Huijberts, G.N.M., Eggink, G. 1996. High-cell-density cultivation of the
lipid accumulating yeast Cryptococcus curvatus using glycerol as a carbon source.
Applied Microbiology and Biotechnology, 45(5), 575-579.
Morita, T., Konishi, M., Fukuoka, T., Imura, T., Kitamoto, D. 2007. Microbial conversion of
glycerol into glycolipid biosurfactants, mannosylerythritol lipids, by a basidiomycete
yeast, Pseudozyma antarctica JCM 10317T. Journal of Bioscience and
Bioengineering, 104(1), 78-81.
Neirinck, L., Maleszka, R., Schneider, H. 1982. Alcohol production from sugar mixtures by
Pachysolen tannophilus.
Neves, L., Oliveira, R., Lucas, C. 2004. Yeast orthologues associated with glycerol transport
and metabolism. FEMS Yeast Research, 5(1), 51-62.
Nevoigt, E., Stahl, U. 1997. O smoregulation and glycerol metabolism in the yeast
Saccharomyces cerevisiae. FEMS Microbiology Reviews, 21(3), 231-241.
Norbeck, J., Påhlman, A.-K., Akhtar, N., Blomberg, A., Adler, L. 1996. Purification and
Characterization of Two Isoenzymes of DL-Glycerol-3-phosphatase from
Saccharomyces cerevisiae. Journal of Biological Chemistry, 271(23), 13875-13881.
Ochoa-Estopier, A., Lesage, J., Gorret, N., Guillouet, S.E. 2011. K inetic analysis of a
Saccharomyces cerevisiae strain adapted for improved growth on glycerol:
Implications for the development of yeast bioprocesses on g lycerol. Bioresource
Technology, 102(2), 1521-1527.
Ohmiya, R., Yamada, H., Nakashima, K., Aiba, H., Mizuno, T. 1995. Osmoregulation of
fission yeast: cloning of two distinct genes encoding glycerol-3-phosphate
dehydrogenase, one of which is responsible for osmotolerance for growth. Molecular
Microbiology, 18(5), 963-973.

23
Olsson, L., Hahn-Hägerdal, B. 1993. F ermentative performance of bacteria and yeasts in
lignocellulose hydrolysates. Process Biochemistry, 28(4), 249-257.
Papanikolaou, S., Muniglia, L., Chevalot, I., Aggelis, G., Marc, I. 2002. Yarrowia lipolytica
as a potential producer of citric acid from raw glycerol. J Appl Microbiol, 92(4), 737-
44.
Pavlik, P., Simon, M., Schuster, T., Ruis, H. 1993. T he glycerol kinase (<i>GUT1 )
gene of Saccharomyces cerevisiae : cloning and characterization. Current Genetics,
24(1), 21-25.
Racenis, P.V., Lai, J.L., Das, A.K., Mullick, P.C., Hajra, A.K., Greenberg, M.L. 1992. The
acyl dihydroxyacetone phosphate pathway enzymes for glycerolipid biosynthesis are
present in the yeast Saccharomyces cerevisiae. Journal of Bacteriology, 174(17),
5702-5710.
Rønnow, B., Kielland-Brandt, M.C. 1993. G UT2, a gene for mitochondrial glycerol 3-
phosphate dehydrogenase of Saccharomyces cerevisiae. Yeast, 9(10), 1121-1130.
Sabra, W., Dietz, D., Tjahjasari, D., Zeng, A.-P. 2010. Biosystems analysis and engineering
of microbial consortia for industrial biotechnology. Engineering in Life Sciences,
10(5), 407-421.
Sathesh-Prabu, C., Murugesan, A.G. 2011. P otential utilization of sorghum field waste for
fuel ethanol production employing Pachysolen tannophilus and Saccharomyces
cerevisiae. Bioresource Technology, 102(3), 2788-2792.
Shams Yazdani, S., Gonzalez, R. 2008. E ngineering Escherichia coli for the efficient
conversion of glycerol to ethanol and co-products. Metabolic Engineering, 10(6),
340-351.
Siderius, M., Van Wuytswinkel, O., Reijenga, K.A., Kelders, M., Mager, W.H. 2000. T he
control of intracellular glycerol in Saccharomyces cerevisiae influences osmotic stress
response and resistance to increased temperature. Molecular Microbiology, 36(6),
1381-1390.
Silva-Graça, M., Lucas, C. 2003. Physiological studies on long-term adaptation to salt stress
in the extremely halotolerant yeast Candida versatilis CBS 4019 (syn. C. halophila).
FEMS Yeast Research, 3(3), 247-260.
Silva, S.N.R.L., Farias, C.B.B., Rufino, R.D., Luna, J.M., Sarubbo, L.A. 2010. Glycerol as
substrate for the production of biosurfactant by Pseudomonas aeruginosa UCP0992.
Colloids and Surfaces B: Biointerfaces, 79(1), 174-183.

24
Singh, S.P., Singh, D. 2010. Biodiesel production through the use of different sources and
characterization of oils and their esters as the substitute of diesel: A review.
Renewable and Sustainable Energy Reviews, 14(1), 200-216.
Slininger, P.J., Bolen, P.L., Kurtzman, C.P. 1987. Pachysolen tannophilus: Properties and
process considerations for ethanol production from d-xylose. Enzyme and Microbial
Technology, 9(1), 5-15.
Slininger, P.J., Bothast, R.J., Van Cauwenberge, J.E., Kurtzman, C.P. 1982. Conversion of D-
xylose to ethanol by the yeast Pachysolen tannophilus. Biotechnology and
Bioengineering, 24(2), 371-384.
Sutherland, F.C., Lages, F., Lucas, C., Luyten, K., Albertyn, J., Hohmann, S., Prior, B.A.,
Kilian, S.G. 1997. C haracteristics of Fps1-dependent and -independent glycerol
transport in Saccharomyces cerevisiae. Journal of Bacteriology, 179(24), 7790-5.
Talarico, T.L., Axelsson, L.T., Novotny, J., Fiuzat, M., Dobrogosz, W.J. 1990. Utilization of
Glycerol as a H ydrogen Acceptor by Lactobacillus reuteri: Purification of 1,3-
Propanediol:NAD+ Oxidoreductase. Applied and Environmental Microbiology, 56(4),
943-948.
Tamás, M.J., Luyten, K., Sutherland, F.C.W., Hernandez, A., Albertyn, J., Valadi, H., Li, H.,
Prior, B.A., Kilian, S.G., Ramos, J., Gustafsson, L., Thevelein, J.M., Hohmann, S.
1999. Fps1p controls the accumulation and release of the compatible solute glycerol
in yeast osmoregulation. Molecular Microbiology, 31(4), 1087-1104.
van Dijken, J.P., Scheffers, W.A. 1986. R edox balances in the metabolism of sugars by
yeasts. FEMS Microbiology Letters, 32(3–4), 199-224.
Van Eck, J.H., Prior, B.A., Brandt, E.V. 1993. The water relations of growth and
polyhydroxy alcohol production by ascomycetous yeasts. Journal of General
Microbiology, 139(5), 1047-1054.
Verduyn C, Van Kleef R, Frank J, Schreuder H, Van Dijken JP, Scheffers WA. 1985.
Properties of the NAD(P)H-dependent xylose reductase from the xylose-fermenting
yeast Pichia stipitis. Biochem J 226:669–677.
Voegele, R.T., Sweet, G.D., Boos, W. 1993. Glycerol kinase of Escherichia coli is activated
by interaction with the glycerol facilitator. Journal of Bacteriology, 175(4), 1087-
1094.
Wang, Z., Zhuge, J., Fang, H., Prior, B.A. 2001. Glycerol production by microbial
fermentation: A review. Biotechnology Advances, 19(3), 201-223.

25
Wedlock, D.N., Thornton, R.J. 1989. T ransformation of a glucose negative mutant of
Pachysolen tannophilus with a plasmid carrying the cloned hexokinase PII gene from
Saccharomyces cerevisiae. Biotechnology Letters, 11(9), 601-604.
Yancey, P., Clark, M., Hand, S., Bowlus, R., Somero, G. 1982. Living with water stress:
evolution of osmolyte systems. Science, 217(4566), 1214-1222.
Yazdani, S.S., Gonzalez, R. 2007. Anaerobic fermentation of glycerol: a path to economic
viability for the biofuels industry. Current Opinion in Biotechnology, 18(3), 213-219.
Yu, K.O., Jung, J., Kim, S.W., Park, C.H., Han, S.O. 2012. S ynthesis of FAEEs from
glycerol in engineered Saccharomyces cerevisiae using endogenously produced
ethanol by heterologous expression of an unspecific bacterial acyltransferase.
Biotechnology and Bioengineering, 109(1), 110-115.
Yu, K.O., Kim, S.W., Han, S.O. 2010. Reduction of glycerol production to improve ethanol
yield in an engineered Saccharomyces cerevisiae using glycerol as a substrate.
Journal of Biotechnology, 150(2), 209-214.
Zhao, L., Yu, J., Zhang, X., Tan, T. 2010. The Ethanol Tolerance of Pachysolen tannophilus
in Fermentation on X ylose. Applied Biochemistry and Biotechnology, 160(2), 378-
385.
Zyl, P.J., Kilian, S.G., Prior, B.A. 1990. The role of an active transport mechanism in
glycerol accumulation during osmoregulation by Zygosaccharomyces rouxii. Applied
Microbiology and Biotechnology, 34(2), 231-235.

26
Chapter 2 Initial screening results for ethanol producers on
glycerol
Abstract
This chapter describes the initial results for screening of the potential candidates for glycerol
conversion to products of interest. Two candidates Pachia pastoris and Pachysolen
tannophilus were shown to be capable of producing ethanol with glycerol as the sole carbon
source. After growth comparison on glycerol and tests for ethanol production in shake flasks,
P. tannophilus was selected as the organism for further study for conversion of glycerol to
ethanol in the experimental work conducted in this PhD study.
Keywords: Screening; glycerol conversion; non-conventional yeasts.

27
1. Objective
In order to find new hosts capable of producing value-added products from glycerol, studies
focused on t he group of organisms known as non-conventional yeasts. In this project, the
interest was in investigating non-conventional yeasts which had the capability of converting
glycerol primarily to liquid biofuels. It is known from the literature that several non-
conventional yeasts can grow on glycerol and a number of these strains exhibit the ability to
produce value-added products, as was already introduced in Chapter 1. Screening for the
potential candidates for glycerol conversion was done by a thorough literature search, with a
shortlist of relevant organisms being subsequently tested for growth on agar plates with
glycerol as the sole carbon source. Then the potential candidates which showed growth on
glycerol plates were further tested in liquid medium. Potentially interesting strains were
assessed based on their profiles of secreted metabolites. Metabolites with primary interest
were alcohols.
2. Materials and Methods
2.1 Medium and strains
All the strains used in were obtained from CBS (Centraalbureau voor Schimmelcultures,
Utrecht, The Netherlands) named as CBS plus number or from the culture collection of the
Center for Microbial Biotechnology, Technical University of Denmark, named as IBT plus
number. The growth experiments were performed in defined medium, containing (per liter)
0.67g yeast nitrogen base w/o amino acids (Difco, USA) and glycerol at % (v/v) as stated.
Agar plates were prepared by adding 2% w/v agar. The strains were maintained in 15%
glycerol in frozen vials (-80°C).
2.2 Submerged cultivation
The strains were cultivated in 250ml shake flasks at 28°C, 150rpm with defined medium,
plus 2.5% (v/v) or 5% (v/v) glycerol as the sole carbon source.
2.3 Analytical methods
Growth was monitored by measuring OD450nm with a Shimadzu UV mini-1240
spectrophotometer (Shimadzu, Japan). Samples were taken periodically from the flasks and

28
filtered through a 0.22 µm syringe filter, and supernatants were preserved at -20°C for later
HPLC analysis. Concentrations of the substrate glycerol in supernatants were measured by
HPLC refractive index detector RID-10A using an Aminex87H column (Bio-Rad, USA).
Separations were performed at 60°C, flow rate of 0.6ml/min and 5mM H2SO4 as mobile
phase.
3. Results and Discussion
3.1 Initial screening results
For the purpose of selecting non-conventional yeast strains which could grow on glycerol and
produce biofuels, a thorough literature search was done in order to find the potential
candidates. The potential strains of interest were then obtained from different culture
collections and are listed in Table 1. The initial growth results on agar plates with different
concentrations of glycerol as the sole carbon source are also shown in Table 1. Results from
agar plates showed that all the yeast strains could grow with 1%, 2% or 5% glycerol as the
sole carbon source, except Zygosaccharomyces rouxii and Schizosacchromyces pombe. The
strains which showed growth on a gar plates were cultivated in liquid medium for further
physiological characterization and analysis of secreted metabolites.
From HPLC analysis, ethanol was only constantly produced from two strains Pichia pastoris
CBS704 and Pachysolen tannophils CBS4044. Candida tropicalis, Hansenula polymorpha,
Pichia stipitis, Debaromyces hansenii and Candida magnoliae exhibited growth on 2%
glycerol but without obvious products accumulated in defined medium except the biomass.
C. tropicalis and P. stipitis consumed only small amounts of glycerol, so the maximum
growth rate (µmax) on glycerol was not calculated with those two strains. P. pastoris and H.
polymorpha could produce relatively high levels of biomass compared to other screened
yeasts. The OD450nm of H. polymorpha reached 47 and P. pastoris reached 53 at 72 hours’
cultivation, while the OD450nm range of other yeasts was from 10 to 30 (Figure 1). Candida
utilis produced acetate on glycerol. Yarrowia lipolytica grew relatively fast compared to other
strains with µmax 0.33 h-1 and could generate mannitol, citric acid, erythritol in defined
medium, which indicated that Y. lipolytica has the potential to be developed for producing
biochemicals from glycerol. The products produced by the strains tested are listed in Table 1.

29
Figure 1. 250ml shake flask cultivations on 2.5% (v/v) glycerol as the sole carbon source with 100ml
medium showing OD450nm of the screened yeasts over 72 hours of cultivation. Results represent the
mean of at least duplicate experiments.
0
10
20
30
40
50
60
0 20 40 60 80Time (Hours)
OD
P. tannophilusP. pastoris
0
10
20
30
40
50
60
0 20 40 60 80Time (Hours)
OD Y. lipolyticaL. jadinii
0
10
20
30
40
50
60
0 20 40 60 80Time (Hours)
OD
C. tropicalisP.stipitis
0
10
20
30
40
50
60
0 20 40 60 80Time (Hours)
OD
H. polymorphaD. hanseniiC. magnolia

30
Table1. Results of initial screening of potential non-conventional yeast cell factories for a glycerol biorefinery (NC*: Not calculated).
Microorganisms Strain no. Growth 1/2/5%
glycerol plates
Products from glycerol (literature) Products in this study µmax
Candida magnoliae IBT235 Yes Mannitol (Khan et al., 2009)
Biomass 0.1
Candida tropicalis IBT249 Yes Ethanol produced from xylose/Glucose (Hahn-Hägerdal et al., 1985)
Biomass NC*
Candida utilis (Lindnera jadinii) (Pichia jadinii)
CBS 841 Yes Biomass (Fieldhouse et al, 2009)
Biomass, Acetate 0.2
Debaromyces hansenii IBT24 Yes Arabitol (Koganti et al., 2011)
Biomass 0.16
Hansenula polymorpha (Pichia angusta)
CBS 1976 Yes Biomass, phytase, alcohol oxidase (Eggeling & Sahm, 1980; Mayer et al., 1999)
Biomass 0.15
Pachysolen tannophilus CBS4044 Yes Ethanol (Maleszka et al, 1982b)
Ethanol, Acetate, 2,3-Butanediol, Biomass
0.33
Pichia pastoris CBS704 Yes Biomass, recombinant protein (Celik et al., 2008)
Ethanol, Acetate, Biomass 0.28
Yarrowia lipolytica IBT446 Yes Biomass, organic acids, lipids, α-amylase (Papanikolaou and Aggelis, 2002; Rywińska & Rymowicz, 2010)
Mannitol, Erythritol, Citric acid, Biomass
0.33
Pichia stipitis CBS5773 Yes Ethanol produced from xylose/Glucose (Silva et al., 2010)
Biomass NC*
Zygosaccharomyces rouxii
IBT565 No
Schizosacchromyces pombe
IBT8 No

31
The products produced by the organisms cultivated in previous studies, are dependent on the
mode of cultivation and the medium used. For instance, it was shown that mannitol can be
produced from glycerol in aqueous solution under aerobic condition without any other
nutrient by the resting cells of C. magnolia (Khan et al., 2009). However, another study
showed that C. magnoliae could only produce mannitol from fructose and sucrose but did not
produce mannitol from glycerol (Baek et al., 2003). The production of arabitol from glycerol
(Koganti et al., 2011) was not noticed in this study, probably because of differences in the
medium composition. S. pombe did not grow on the glycerol agar plate in this study, whereas
it has been shown that it can utilize glycerol in the presence of small amounts of ethanol and
1-propanol, which were recognized as inducers of glycerol metabolism. These results varied
between studies. In this study, a defined medium was used for all submerged cultivations, in
order to allow a relevant quantitative physiological comparison of results obtained under
different cultivation conditions.
3.2 Growth comparison of two ethanol producers on glycerol in shake flasks
For the purpose of selecting one strain which could be optimized for ethanol production from
glycerol, P. pastoris and P. tannophius were cultivated in defined medium on 5% (v/v)
glycerol as the sole carbon source in shake flasks.
P. pastoris was cultured with 50ml, 100ml, 150ml and 200ml of medium in 250 ml shake
flasks. This method allows a rough estimate of the influence of oxygen transfer in submerged
cultivation. It was shown (Figure 2) that the lower the volume of medium in the flasks, the
more biomass was accumulated and the faster glycerol was consumed. With regard to the
ethanol production level, the cells in 150ml medium produced the highest level of ethanol
(3.4 g/L and a yield of 0.13 g ethanol g-1 glycerol), which indicated that the ethanol
production level was likely to be related to oxygen availability.
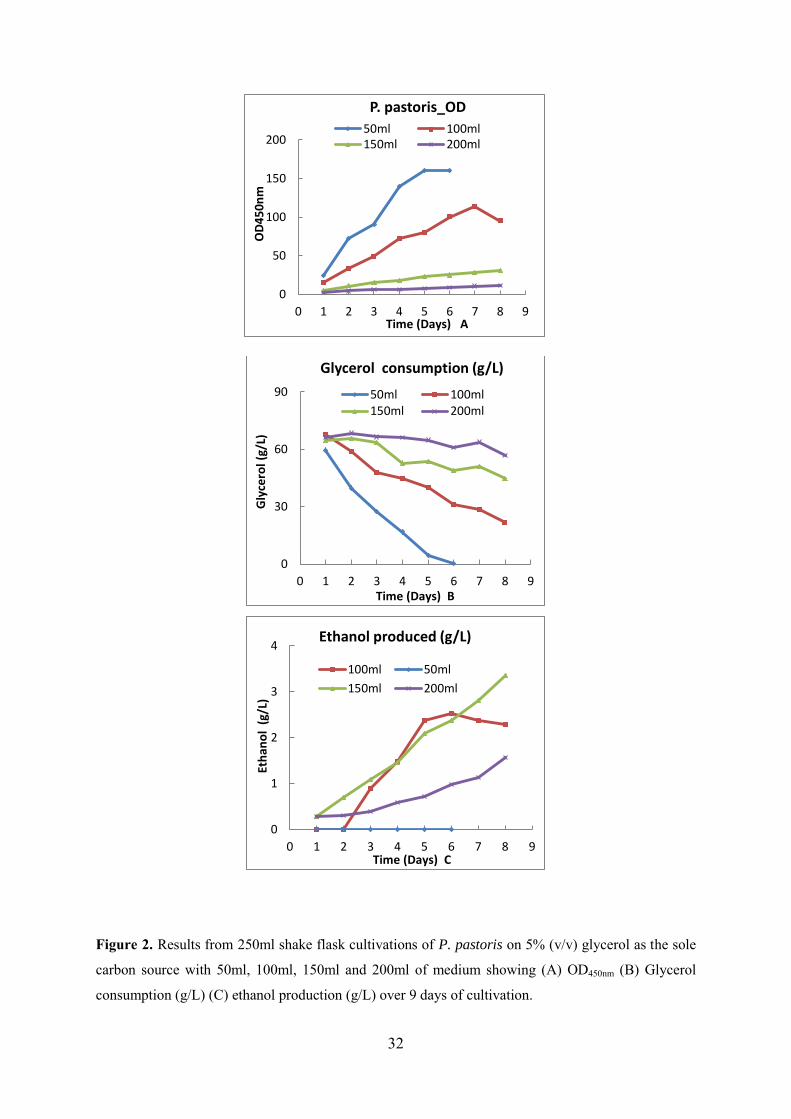
32
Figure 2. Results from 250ml shake flask cultivations of P. pastoris on 5% (v/v) glycerol as the sole
carbon source with 50ml, 100ml, 150ml and 200ml of medium showing (A) OD450nm (B) Glycerol
consumption (g/L) (C) ethanol production (g/L) over 9 days of cultivation.
0
50
100
150
200
0 1 2 3 4 5 6 7 8 9
OD4
50nm
Time (Days) A
P. pastoris_OD 50ml 100ml150ml 200ml
0
30
60
90
0 1 2 3 4 5 6 7 8 9
Gly
cero
l (g/
L)
Time (Days) B
Glycerol consumption (g/L)
50ml 100ml150ml 200ml
0
1
2
3
4
0 1 2 3 4 5 6 7 8 9
Etha
nol
(g/L
)
Time (Days) C
Ethanol produced (g/L)
100ml 50ml150ml 200ml

33
P. tannophilus was cultured with 100ml medium in 250ml shake flasks. It produced 22.7g/L
ethanol with a yield of 0.36 g ethanol g-1 glycerol after 6 days cultivation (Figure 3). Under
the same conditions, P. pastoris only accumulated 2.5 g /L ethanol with a yield of 0.1 g
ethanol g-1 glycerol over 8 days of cultivation. P. tannophilus grew on glycerol with µmax 0.33
h-1 while P. pastoris grew on glycerol with µmax 0.28 h-1 (Table 1). By comparing the ethanol
yield, production level and growth rate on glycerol, P. tannophilus showed more potential to
be selected as the ethanol production host on glycerol. The physiology studies and
characterization of the potential ethanol producer P. tannophilus was further investigated in
fermentor, where all the cultivation conditions, such as pH, dissolved oxygen concentration,
off-gas composition, temperature, stirrer speed and so on could be monitored and controlled
during the whole cultivation process. The results obtained from these studies are presented in
Chapter 3.
Figure 3. Results from 250ml shake flask cultivations of P. tannophilus and P. pastoris on 5% (v/v)
glycerol as the sole carbon source with 100ml medium showing OD450nm, glycerol consumption (g/L)
and ethanol production (g/L).
0
10
20
30
0
20
40
60
80
100
120
0 1 2 3 4 5 6 7 8 9
Gly
cero
l (g/
L) /
OD
Time (Days)
P. pastoris Glycerol OD Ethanol
P
0
10
20
30
0
20
40
60
80
100
120
0 1 2 3 4 5 6 7 8 9
Gly
cero
l (g/
L) /
OD
Time (Days)
P. tannophilus Glycerol OD Ethanol
P
Eth
anol
(g/L
) E
than
ol (g
/L)

34
In addition, prior to further investigation in the controlled fermentor, cultivation of P.
tannophilus on 2.5 % (v/v) glycerol were performed for confirmation of the previous shake
flask results. It was shown that P. tannophilus could produce 6.5 g/L ethanol with a yield of
0.282 g ethanol g-1 glycerol over 72 hours cultivation (Figure 4). The results from studies to
optimize this process are presented in Chapter 3.
Figure 4. Results from 250ml shake flask cultivations of P. tannophilus on 2.5% (v/v) glycerol as the
sole carbon source with 100ml medium showing OD450nm, glycerol consumption (g/L) and ethanol
production (g/L). Results represent the mean of triplicate experiments.
-2
0
2
4
6
8
-5
0
5
10
15
20
25
30
-2 8 18 28 38 48 58 68 78
Gly
cero
l (g/
L) /
OD
Time ( Hours)
P. tannophilus
OD450nm Glycerol (g/L) Ethanol (g/L)
Etha
nol (
g/L)

35
Reference
Baek, H., Song, K.-H., Park, S.-M., Kim, S.-Y., Hyun, H.-H. 2003. Role of glucose in the
bioconversion of fructose into mannitol by Candida magnoliae. Biotechnology
Letters, 25(10), 761-765.
Celik, E., Ozbay, N., Oktar, N., Calık, P. 2008. Use of Biodiesel Byproduct Crude Glycerol
as the Carbon Source for Fermentation Processes by Recombinant Pichia pastoris.
Industrial & Engineering Chemistry Research, 47(9), 2985-2990.
Eggeling, L., Sahm, H. 1980. R egulation of alcohol oxidase synthesis in Hansenula
polymorpha : Oversynthesis during growth on mixed substrates and induction by
methanol. Archives of Microbiology, 127(2), 119-124.
Fieldhouse, R., MacLennan, D.F., MacLennan, D.G., MacLennan, M.E. 2009. ROCESS FOR
PREPARING NUTRITIONAL, THERAPEUTIC OR ORGANOLEPTIC
PRODUCTS FROM CRUDE GLYCEROL, U.S. Patent 2011/0044972(Issued
Feb.24, 2011).
Hahn-Hägerdal, B., Jönsson, B., Lohmeier-Vogel, E. 1985. Shifting product formation from
xylitol to ethanol in pentose fermentations using Candida tropicalis by adding
polyethylene glycol (PEG). Applied Microbiology and Biotechnology, 21(3), 173-175.
Khan, A., Bhide, A., Gadre, R. 2009. Mannitol production from glycerol by resting cells of
Candida magnoliae. Bioresource Technology, 100(20), 4911-4913.
Koganti, S., Kuo, T., Kurtzman, C., Smith, N., Ju, L.-K. 2011. Production of arabitol from
glycerol: strain screening and study of factors affecting production yield. Applied
Microbiology and Biotechnology, 90(1), 257-267.
Maleszka, R., Wang, P., Schneider, H. 1982. E thanol production from D-galactose and
glycerol by Pachysolen tannophilus. Enzyme and Microbial Technology, 4(5), 349-
352.
Mayer, A.F., Hellmuth, K., Schlieker, H., Lopez-Ulibarri, R., Oertel, S., Dahlems, U.,
Strasser, A.W.M., van Loon, A.P.G.M. 1999. A n expression system matures: A
highly efficient and cost-effective process for phytase production by recombinant
strains of Hansenula polymorpha. Biotechnology and Bioengineering, 63(3), 373-381.
Papanikolaou, S., Muniglia, L., Chevalot, I., Aggelis, G., Marc, I. 2002. Yarrowia lipolytica
as a potential producer of citric acid from raw glycerol. J Appl Microbiol, 92(4), 737-
44.

36
Rywińska, A., Rymowicz, W. 2010. H igh-yield production of citric acid by Yarrowia
lipolytica on glycerol in repeated-batch bioreactors. Journal of Industrial
Microbiology & Biotechnology, 37(5), 431-435.
Silva, J., Mussatto, S., Roberto, I. 2010. The Influence of Initial Xylose Concentration,
Agitation, and Aeration on E thanol Production by Pichia stipitis from Rice Straw
Hemicellulosic Hydrolysate. Applied Biochemistry and Biotechnology, 162(5), 1306-
1315.

37
Chapter 3 Bioconversion of crude glycerol feedstocks into ethanol
by Pachysolen tannophilus
Xiaoying Liu, Peter Ruhdal Jensen, Mhairi Workman*
Published on Bioresource Technology

Bioconversion of crude glycerol feedstocks into ethanol by Pachysolen tannophilus
Xiaoying Liu, Peter Ruhdal Jensen, Mhairi Workman ⇑Center for Systems Microbiology, Department of Systems Biology, Building 301, Matematiktorvet, Technical University of Denmark, 2800 Kgs. Lyngby, Denmark
a r t i c l e i n f o
Article history:Received 17 August 2011Received in revised form 17 October 2011Accepted 18 October 2011Available online 28 October 2011
Keywords:Crude glycerolEthanolFermentationPachysolen tannophilus
a b s t r a c t
Glycerol, the by-product of biodiesel production, is considered as a waste by biodiesel producers. Thisstudy demonstrated the potential of utilising the glycerol surplus through conversion to ethanol bythe yeast Pachysolen tannophilus (CBS4044). This study demonstrates a robust bioprocess which wasnot sensitive to the batch variability in crude glycerol dependent on raw materials used for biodiesel pro-duction. The oxygen transfer rate (OTR) was a key factor for ethanol production, with lower OTR having apositive effect on ethanol production. The highest ethanol production was 17.5 g/L on 5% (v/v) crudeglycerol, corresponding to 56% of the theoretical yield. A staged batch process achieved 28.1 g/L ethanol,the maximum achieved so far for conversion of glycerol to ethanol in a microbial bioprocess. The fermen-tation physiology has been investigated as a means to designing a competitive bioethanol productionprocess, potentially improving economics and reducing waste from industrial biodiesel production.
� 2011 Elsevier Ltd. All rights reserved.
1. Introduction
In recent years, increasing focus on renewable and sustainableenergy has boosted the production of biodiesel from rapeseed oil,cooking oil and animal fats, especially in Europe. Glycerol, a by-product of biodiesel production, has seen a proportionally dramaticincrease in production. As approximately 10% of the reaction vol-ume in a given biodiesel production process ends up as glycerol(Johnson and Taconi, 2007), the market for glycerol has becomesaturated and it is now being considered as a waste by many bio-diesel producers. Bioconversion of glycerol into liquid biofuels,green chemicals and bioenergy on the basis of fermentation pro-cesses can provide an efficient solution for sustainable manage-ment of glycerol, which can improve the economics of biodieselindustries (Lynd et al., 2005). Among the different types of biofuelsfor replacement of fossil fuels, bioethanol is currently most used,and bioethanol is the only alcohol produced in large scale fermen-tation processes. Presently, ethanol is primarily produced fromsugarcane in Brazil, from corn starch in the USA and from sugarbeets in Europe (da Silva et al., 2009). Glycerol is considered asan inexpensive feedstock for biotechnology industries, and thusthe possibility for the production of sustainable biofuels from thissubstrate is attractive.
The composition of crude glycerol generated from biodieselprocesses varies dependent on the oil feedstock used and the cata-lysts employed in the transesterification process. The common
feedstocks are soybean (USA), rapeseed (Europe), sunflower, saf-flower, canola, and palm oils. In order to reduce the cost, wastecooking oil, animal fats, soapstocks and greases are also utilized(Canakci and Sanli, 2008). Oil from microalgae is also a competitivefeedstock for biodiesel production (Pokoo-Aikins et al., 2010).Increasing demand for sustainable biofuels such as biodiesel en-sures that glycerol will continue to be a potential substrate for fur-ther conversion into value-added products in the foreseeablefuture. This offers considerable scope for designing biorefineriesbased on glycerol and developing microbial cell factories for biolog-ical conversion of this substrate on large scale. The organisms em-ployed to convert the crude glycerol should be robust enough towithstand the variability in glycerol stock, thus ensuring the appli-cability of the conversion process to the widest range of biodieselproducers.
It has been shown that a number of (typically anaerobic) bacte-ria are capable of growing on glycerol as the sole carbon and energysource. Glycerol can be converted to a wide range of biochemicalsand biofuels such as ethanol, butanol, 1,3-propanediol, succinate,dihydroxyacetone, propionic acid and pigments (da Silva et al.,2009). The newly isolated bacterium, Kluyvera cryocrescens can pro-duce up to 27 g/L ethanol from crude glycerol under microaerobicbatch fermentation (Choi et al., 2011). Escherichia coli has beeninvestigated to be an ethanol production platform on glycerol, withup to 10 g/L achievable by engineered E. coli growing on 22 g/Lcrude glycerol and with hydrogen and formate as byproducts underanaerobic condition (Shams Yazdani and Gonzalez, 2008). An engi-neered Klebsiella pneumoniae strain has been shown to achieve25 g/L ethanol on crude glycerol (Oh et al., 2011). However, theseprocesses require a controlled anaerobic environment, maintainedthrough sparging with nitrogen.
0960-8524/$ - see front matter � 2011 Elsevier Ltd. All rights reserved.doi:10.1016/j.biortech.2011.10.065
⇑ Corresponding author. Present address: Center for Microbial Biotechnology,Department of Systems Biology, Building 223, Søltofts Plads, Technical University ofDenmark, 2800 Kgs. Lyngby, Denmark. Tel.: +45 4525 2700; fax: +45 4588 4148.
E-mail address: [email protected] (M. Workman).
Bioresource Technology 104 (2012) 579–586
Contents lists available at SciVerse ScienceDirect
Bioresource Technology
journal homepage: www.elsevier .com/locate /bior tech
38

In order to find new hosts capable of producing value-addedproducts from renewable substrates, it may be interesting to lookto non-conventional hosts for the production of biofuels and bio-chemicals. Here, a wider substrate range can be naturallyexploited, and more diverse product profiles are observed, withoutthe need for genetic engineering or expensive operating require-ments. It is known that several non-conventional yeasts can growon glycerol and a number of these strains exhibit the ability to pro-duce value added products. For example, glycerol can be assimi-lated into organic acids and single cell oil by Yarrowia lipolytica(Makri et al., 2010; Papanikolaou et al., 2002) and has beenexploited as carbon source for producing biomass and recombinantproteins in Pichia pastoris (Celik et al., 2008; Fieldhouse et al.,2009). For ethanol production from glycerol, only two geneticallyengineered yeasts have been reported which can convert glycerolinto ethanol. The industrial work horse Saccharomyces cerevisiaehas been genetically engineered to produce ethanol from glycerol,but only up to 4.4 g/L (Yu et al., 2010). The methylotrophic yeastHansenula polymorpha was engineered to improve ethanol produc-tion by expression of varied genes from bacteria, however afterseveral rounds of genetic engineering, the production levelachieved was only 3.1 g/L (Hong et al., 2010).
Pachysolen tannophilus was the first yeast shown to be capableof fermenting xylose sugars to ethanol (Kurtzman, 1983) and thexylose utilisation pathway has been extensively studied in thisorganism (Sathesh-Prabu and Murugesan, 2011; Slininger et al.,1987; Zhao et al., 2010). In a previous study, it was reported thatP. tannophilus could accumulate 4 g/L ethanol on glycerol underaerobic growth (Maleszka et al., 1982), however, the conditionsfor ethanol production were not precisely defined or controlledand the physiology during growth on glycerol has not been exten-sively studied in this organism. The possibility for studying thephysiology of glycerol conversion to ethanol in this organism pro-vides an interesting prospect for the future production of biofuels.In this study, therefore, the glycerol metabolism has been furtherinvestigated in this strain as a means of defining the parameterswhich affect ethanol production in controlled submerged cultiva-tions. The results may be used to design an optimised productionprotocol for ethanol production at lab scale, which could be imple-mented and tested at larger scale. In addition, we aim to providethe basis for an investigation of glycerol metabolism in P. tannophi-lus, with a view to highlighting targets for further improvements ofthis strain as a cell factory through genetic engineering.
2. Methods
2.1. Microorganism and culture medium
P. tannophilus CBS4044 (Centraalbureau voor Schimmelcultures,Utrecht, The Netherlands) was used throughout this study. Thegrowth experiments were performed in defined medium, contain-ing (per liter) 0.67 g yeast nitrogen base w/o amino acids (Difco,USA) and glycerol at % (v/v) as stated. The strain was maintainedin 15% glycerol in frozen vials (�80 �C).
2.2. Substrate source
Pure glycerol (Sigma–Aldrich, USA) or crude glycerol from bio-diesel production (MEROCO a.s., Leopoldov, Slovakia) was used asthe sole carbon source in media used in this study. Three differentbatches of crude glycerol were obtained from MEROCO, based ondifferent raw materials for biodiesel production. Batches 1 and 2were based on rapeseed oil (from production in December 2008and October 2009, respectively) and Batch 3 was based on 90% rape-seed oil and 10% cooking oil (from production in December 2009).
The crude glycerol feedstock contains up to 7% ash (includingformic acid, citric acid, acetic acid, nitrate and chloride), 0.5% meth-anol, 10% water and 82 ± 5% glycerol.
2.3. Submerged batch cultivations
Unless otherwise stated, experiments were carried out in fullyinstrumented and automatically controlled BIOSTAT� Q plus fer-mentors (Sartorius Stedim Biotech S.A., Germany). Temperature,stirrer speed, pH and dissolved oxygen tension (DOT) were con-stantly monitored and automatically controlled on-line. Carbondioxide and oxygen concentrations in the off-gas were monitoredby a Brüel and Kjær acoustic gas analyzer (Brüel and Kjær, Nærum,Denmark). The working volume was 600 mL.
All cultivations were performed at 28 �C. All batch processescontained 100 lL/L silicone oil to control the foaming during fer-mentation. Duplicates of all batches were performed; results shownare mean values (and standard error) for two identical processes.
To study the effect of aeration rate on growth and ethanol pro-duction, cultivations were performed on 2% (v/v) glycerol, at pH 5with a stirrer speed of 450 rpm with the following airflow (aera-tion) rates: 0.5 L/min (0.83 vvm), 0.2 L/min (0.33 vvm), 0.1 L/min(0.17 vvm), 0.05 L/min (0.083 vvm), and at 350 rpm with 0.05 L/min (0.083 vvm), and one process where the dissolved oxygen ten-sion (DOT) controlled at 20%. To convert from flow rate (L/min) toaeration rate (vvm) the following conversion factor was used: 1 L/min = 1.7 vvm (the working volume for the fermentator was 0.6 L).
The oxygen transfer rate (OTR) was assumed to be equal to theoxygen uptake rate (OUR) under dissolved oxygen limited condi-tion. From a mass balance, the OUR in batch fermentations couldbe calculated according to:
OUR ¼ F InO2 � FOutO2
V
where FIn and FOut are the molar flow rates based on the O2 concen-tration measured at the fermentor inlet and outlet, and V is workingvolume in the fermentor (Garcia-Ochoa and Gomez, 2009).
The effect of pH on ethanol production was investigated in sub-merged batch cultivations on 2% (v/v) glycerol. Cultivations wereperformed at pH 3, 4, 5, 6 and 7 with airflow (aeration) at 0.05 L/min (0.083 vvm) and 450 rpm stirring. In all cases, pH was auto-matically controlled by addition of 2 M HCl or 2 M NaOH.
To determine how ethanol production was affected by differentnitrogen sources, cultivations were performed in defined mediumwith (NH4)2SO4 or NaNO3 as the nitrogen source on 5% (v/v)glycerol in 1 L fermentors (Biostat, B. Braun Biotech International,Germany) at 150 rpm, 0.2 vvm, without pH control and 10 g succi-nic acid and 6 g NaOH (per liter) were supplemented as buffer.
The effect of three different batches of glycerol from biodieselproduction at Meroco was investigated in batch cultivations atpH 6, 450 rpm and 0.1 L/min (0.17 vvm) on 2% (v/v) crude glycerol.The effect of different concentrations of crude glycerol on cellgrowth and ethanol production was tested at 2% (v/v), 5% (v/v),and 10% (v/v) using Batch 3 crude glycerol based on 90% rapeseedoil and 10% cooking oil, pH 5, 450 rpm and 0.05 L/min (0.083 vvm).
A staged batch fermentation was performed on 5% (v/v) of Batch3 crude glycerol. At the end of batch phase where glycerol was ex-hausted, concentrated medium containing crude glycerol and YNBwere added the fermentor to a final concentration of 5% (v/v).
2.4. Ethanol tolerance studies
The cells were grown in defined medium on 2% (v/v) glycerol(Sigma–Aldrich, USA) in 250 mL Erlenmeyer flasks with 100 mLculture media with 0.9% (v/v), 1.7% (v/v), 2.6% (v/v), 3.4% (v/v),
580 X. Liu et al. / Bioresource Technology 104 (2012) 579–586
39
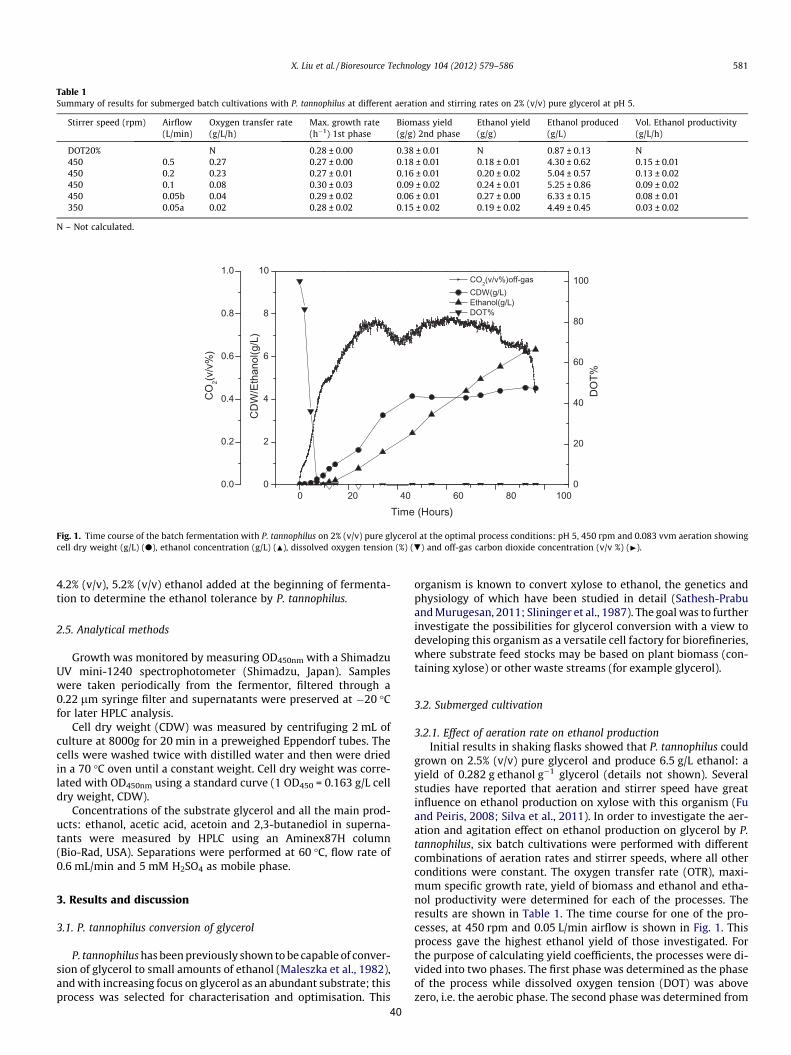
4.2% (v/v), 5.2% (v/v) ethanol added at the beginning of fermenta-tion to determine the ethanol tolerance by P. tannophilus.
2.5. Analytical methods
Growth was monitored by measuring OD450nm with a ShimadzuUV mini-1240 spectrophotometer (Shimadzu, Japan). Sampleswere taken periodically from the fermentor, filtered through a0.22 lm syringe filter and supernatants were preserved at �20 �Cfor later HPLC analysis.
Cell dry weight (CDW) was measured by centrifuging 2 mL ofculture at 8000g for 20 min in a preweighed Eppendorf tubes. Thecells were washed twice with distilled water and then were driedin a 70 �C oven until a constant weight. Cell dry weight was corre-lated with OD450nm using a standard curve (1 OD450 = 0.163 g/L celldry weight, CDW).
Concentrations of the substrate glycerol and all the main prod-ucts: ethanol, acetic acid, acetoin and 2,3-butanediol in superna-tants were measured by HPLC using an Aminex87H column(Bio-Rad, USA). Separations were performed at 60 �C, flow rate of0.6 mL/min and 5 mM H2SO4 as mobile phase.
3. Results and discussion
3.1. P. tannophilus conversion of glycerol
P. tannophilus has been previously shown to be capable of conver-sion of glycerol to small amounts of ethanol (Maleszka et al., 1982),and with increasing focus on glycerol as an abundant substrate; thisprocess was selected for characterisation and optimisation. This
organism is known to convert xylose to ethanol, the genetics andphysiology of which have been studied in detail (Sathesh-Prabuand Murugesan, 2011; Slininger et al., 1987). The goal was to furtherinvestigate the possibilities for glycerol conversion with a view todeveloping this organism as a versatile cell factory for biorefineries,where substrate feed stocks may be based on plant biomass (con-taining xylose) or other waste streams (for example glycerol).
3.2. Submerged cultivation
3.2.1. Effect of aeration rate on ethanol productionInitial results in shaking flasks showed that P. tannophilus could
grown on 2.5% (v/v) pure glycerol and produce 6.5 g/L ethanol: ayield of 0.282 g ethanol g�1 glycerol (details not shown). Severalstudies have reported that aeration and stirrer speed have greatinfluence on ethanol production on xylose with this organism (Fuand Peiris, 2008; Silva et al., 2011). In order to investigate the aer-ation and agitation effect on ethanol production on glycerol by P.tannophilus, six batch cultivations were performed with differentcombinations of aeration rates and stirrer speeds, where all otherconditions were constant. The oxygen transfer rate (OTR), maxi-mum specific growth rate, yield of biomass and ethanol and etha-nol productivity were determined for each of the processes. Theresults are shown in Table 1. The time course for one of the pro-cesses, at 450 rpm and 0.05 L/min airflow is shown in Fig. 1. Thisprocess gave the highest ethanol yield of those investigated. Forthe purpose of calculating yield coefficients, the processes were di-vided into two phases. The first phase was determined as the phaseof the process while dissolved oxygen tension (DOT) was abovezero, i.e. the aerobic phase. The second phase was determined from
Table 1Summary of results for submerged batch cultivations with P. tannophilus at different aeration and stirring rates on 2% (v/v) pure glycerol at pH 5.
Stirrer speed (rpm) Airflow(L/min)
Oxygen transfer rate(g/L/h)
Max. growth rate(h�1) 1st phase
Biomass yield(g/g) 2nd phase
Ethanol yield(g/g)
Ethanol produced(g/L)
Vol. Ethanol productivity(g/L/h)
DOT20% N 0.28 ± 0.00 0.38 ± 0.01 N 0.87 ± 0.13 N450 0.5 0.27 0.27 ± 0.00 0.18 ± 0.01 0.18 ± 0.01 4.30 ± 0.62 0.15 ± 0.01450 0.2 0.23 0.27 ± 0.01 0.16 ± 0.01 0.20 ± 0.02 5.04 ± 0.57 0.13 ± 0.02450 0.1 0.08 0.30 ± 0.03 0.09 ± 0.02 0.24 ± 0.01 5.25 ± 0.86 0.09 ± 0.02450 0.05b 0.04 0.29 ± 0.02 0.06 ± 0.01 0.27 ± 0.00 6.33 ± 0.15 0.08 ± 0.01350 0.05a 0.02 0.28 ± 0.02 0.15 ± 0.02 0.19 ± 0.02 4.49 ± 0.45 0.03 ± 0.02
N – Not calculated.
0 20 40 60 80 1000
2
4
6
8
10
CD
W/E
than
ol(g
/L)
DO
T%
CO
2(v/v
%)
Time (Hours)
CO2(v/v%)off-gas CDW(g/L) Ethanol(g/L) DOT%
0
20
40
60
80
100
0.0
0.2
0.4
0.6
0.8
1.0
Fig. 1. Time course of the batch fermentation with P. tannophilus on 2% (v/v) pure glycerol at the optimal process conditions: pH 5, 450 rpm and 0.083 vvm aeration showingcell dry weight (g/L) (d), ethanol concentration (g/L) (N), dissolved oxygen tension (%) (.) and off-gas carbon dioxide concentration (v/v %) (I).
X. Liu et al. / Bioresource Technology 104 (2012) 579–586 581
40

the point where DOT decreased to zero until all glycerol was con-sumed. This phase was considered to be microaerobic phase. Sim-ilar trends were observed for all processes, with the aerobic phasecontinuing until approximately 10 h after inoculation, typicallywithout ethanol production in this phase. The off-gas CO2 tracein the microaerobic phase clearly demonstrates oxygen limitationin the culture, and was accompanied by a lower growth rate andethanol accumulation.
The results showed that the maximum specific growth rate wasnot greatly influenced by the change in OTR in the range studied.For the set of processes operated at 450 rpm, the biomass yieldon the second phase on glycerol decreased as OTR decreased, witha maximum of 0.18 g cells g�1 glycerol at 0.5 L/min airflow, whileethanol yield increased to a maximum of 0.27 g ethanol g�1 glyc-erol at 450 rpm and 0.05 L/min airflow.
One additional process was performed where the DOT was con-trolled at stable value of 20% throughout the cultivation; the cellsgrew at maximum growth rate of 0.28 h�1 with a biomass yield of0.51 g cells g�1 glycerol. Only 0.87 ± 0.13 g/L of ethanol was pro-duced at the last 8 h fermentation.
The tendency for ethanol production from glycerol was thatwith increasing OTR the ethanol yield declines (Fig. 2A). It has beenreported that the aeration rate is one of the most important param-eters concerned with alcoholic fermentation in yeasts (Jeffries,1983b; Visser et al., 1990). In the ethanol production phase in thisstudy, the DOT was around 0% for all processes. Some studies haveshown that the optimal DOT values for ethanol production undermicroaerobic condition are below 1% of saturation (Agbogbo andCoward-Kelly, 2008). The cells begin to accumulate ethanol undermicroaerobic conditions when their growth is oxygen limited.Otherwise the cells will grow at maximum growth rate. HighOTR has negative effect on ethanol yield and production anddetermines the carbon flow distribution between cell growth andethanol production.
From HPLC analysis results, at higher airflow rates (0.5 L/min,0.2 L/min) the main products on glycerol were ethanol, acetic acid,and small amounts of acetoin and 2,3-butanediol. At loweraeration rates (0.1 L/min, 0.05 L/min), the products were mainlyethanol and acetic acid.
0.05aL/min0.05bL/min 0.1L/min 0.2L/min 0.5L/min DOT20%0.00
0.05
0.10
0.15
0.20
0.25
0.30(A)
Etha
nol y
ield
(g/g
)
Ethanol yield (g/g)
pH3 pH4 pH5 pH6 pH6 Batch1 Batch2 Batch30.00
0.05
0.10
0.15
0.20
0.25
0.30(B)
Etha
nol y
ield
(g/g
)
Ethanol yield (g/g)
Fig. 2. (A) Ethanol yield from batch cultivations with P. tannophilus at differentaeration and stirring rates on 2% (v/v) pure glycerol at pH 5. (B) Ethanol yields frombatch cultivations with P. tannophilus at different pH on 2% (v/v) pure glycerol andat pH 6 on different batches of crude glycerol from biodiesel production. Note: 0.05a– 0.05 L/min, 350 rpm; 0.05b – 0.05 L/min, 450 rpm.
0
5
10
15
20
25
30
0 10 20 30 40 50 60 70 80 90 1000
5
10
15
20
25
30
(A)
CD
W/E
than
ol(g
/L)
Gly
cero
l(g/L
)
01234567
(B)
Gly
cero
l(g/L
)
Time (Hours)
Glycerol
0
1
2
3
4
5
6
7
CDW Ethanol
CD
W/E
than
ol (g
/L)
Fig. 3. Time course for submerged batch cultivations of P. tannophilus on (A) 2% (v/v) pure glycerol and (B) 2% (v/v) Batch 3 crude glycerol at the optimal process conditions:pH 5, 450 rpm, and 0.083 vvm aeration, showing glycerol concentration (g/L) (j), ethanol concentration (g/L) (.), and cell dry weight (g/L) (d).
582 X. Liu et al. / Bioresource Technology 104 (2012) 579–586
41

3.2.2. Effect of pH on ethanol productionPrevious studies investigating the effect of pH on xylitol produc-
tion in P. tannophilus have shown that the optimum to be betweenpH 6 and pH 7.5 (Converti et al., 1999). With the purpose of optimi-zation ethanol production on glycerol, the effect of pH on ethanolproduction was investigated. Results are shown in Fig. 2B. pH 5was found to be the optimal pH for ethanol yield and productionon glycerol by P. tannophilus. The deviation from optimal pH af-fected the ethanol yield to varying degrees. Only very slow growthof the culture was observed at pH 7 on glycerol. These results weredifferent from the value (pH 6) reported for the optimal growth andethanol production on sorghum stove for this yeast (Sathesh-Prabuand Murugesan, 2011), however, the optimal value is likely tochange with varied substrates, products, medium and strains uti-lized for different processes. The pH effects probably resulted fromthe different enzyme activity in the metabolic pathways for the spe-cific process (Sathesh-Prabu and Murugesan, 2011).
3.3. Conversion of crude glycerol
3.3.1. Growth comparison on crude glycerol and pure glycerolDue to a wide variety of feedstocks being utilized in biodiesel
production, the composition of crude glycerol can vary due to res-idues and impurities being carried over to the by-product. Chemi-cals used in the transesterification process can also be present inthe glycerol. These residues can inhibit the growth of micro-organ-isms and thus affect product formation (GLYFINERY project report,
results not shown). The process for converting glycerol should berobust to the impurities and variability of crude glycerol offeringa cost-effective solution for converting this waste. In order todetermine the sensitivity of the strain P. tannophilus and the etha-nol production process to impurities and variations in crude glyc-erol supply, three different batches of glycerol from a biodieselproducer were tested. The growth and ethanol yield varied onlyslightly compared to the values on pure glycerol at pH 6 and nogreat difference was observed on the three different crude glycerolbatches (Fig. 2B).
Previous results (Table 1) showed that the optimal conditions forbatch ethanol production on pure glycerol were pH 5, 0.05 L/min air-flow (0.083 vvm), and 28 �C. Ethanol production using these condi-tions and the third batch of crude glycerol from rapeseed oil andcooking oil, which is likely to contain more impurities and residues,was almost the same as on the pure glycerol (Fig. 3A and B). The eth-anol yield on crude glycerol was 0.26 g ethanol g�1 glycerol and onpure glycerol was 0.27 g ethanol g�1 glycerol. The ethanol produc-tion was around 6.3 g/L for both processes. Therefore, from the re-sults we can conclude that the impurities (ash, methanol, salts,etc.) and variability of crude glycerol have no negative effect onthe growth and ethanol production of P. tannophilus. The P. tannophi-lus process is thus robust and can be adapted for ethanol productionfrom different sources of crude glycerol feedstocks from biodieselproduction. This means that for large scale production of ethanol,it would not be necessary to purify the crude glycerol which willgreatly reduce the production cost.
Table 2Summary of results for submerged batch cultivations with P. tannophilus grown on 2% (v/v) Batch 1, Batch 2, Batch 3 crude glycerol from biodiesel production with different oilfeedstocks and on 2% (v/v), 5% (v/v), 10% (v/v) Batch 3 crude glycerol. Results are compared to the standard batch process on 2% (v/v) pure glycerol.
Glycerol source pH Airflow (L/min) Glycerol conc. (v/v%) Glycerol utilized (g/L) Ethanol produced (g/L) Ethanol yield (g/g) Dry cell weight (g/L)
Pure 5 0.05 2 24.2 ± 1.41 6.33 ± 0.15 0.27 ± 0.00 4.67 ± 0.26Batch 3 5 0.05 2 26.1 ± 0.71 6.28 ± 0.15 0.26 ± 0.02 4.40 ± 0.02Batch 3 5 0.05 5 62.9 ± 0.95 17.5 ± 0.88 0.28 ± 0.03 5.90 ± 0.11Batch 3 5 0.05 10 113.8 ± 1.02 17.5 ± 1.15 0.16 ± 0.04 5.40 ± 0.03
Note: Glycerol conc. 1 (v/v%) = 1.261 g/cm3 = 12.61 g/L.
0
10
20
30
40
50
60
0 25 50 75 100 125 150 175 2000
10
20
30
40
50
60
(A)
Gly
cero
l(g/L
)
(B)
Glycerol
0
4
8
12
16
CDW Ethanol
Gly
cero
l(g/L
)
Time (Hours)
0
4
8
12
16
CD
W/E
than
ol (g
/L)
CD
W/E
than
ol (g
/L)
Fig. 4. Time course of the batch fermentation with P. tannophilus on 5% (v/v) pure glycerol with different nitrogen sources (A) NO�3 (B) NHþ4 , showing glycerol concentration(g/L) (j), ethanol concentration (g/L) (.), and cell dry weight (g/L) (d).
X. Liu et al. / Bioresource Technology 104 (2012) 579–586 583
42
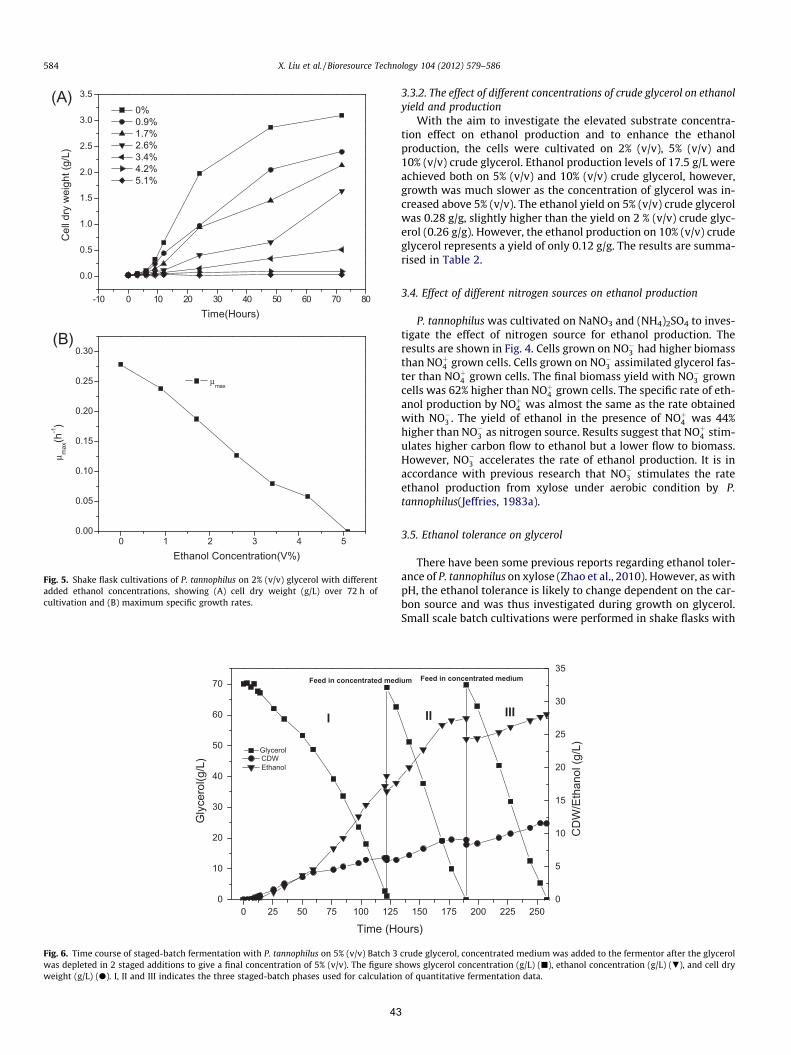
3.3.2. The effect of different concentrations of crude glycerol on ethanolyield and production
With the aim to investigate the elevated substrate concentra-tion effect on ethanol production and to enhance the ethanolproduction, the cells were cultivated on 2% (v/v), 5% (v/v) and10% (v/v) crude glycerol. Ethanol production levels of 17.5 g/L wereachieved both on 5% (v/v) and 10% (v/v) crude glycerol, however,growth was much slower as the concentration of glycerol was in-creased above 5% (v/v). The ethanol yield on 5% (v/v) crude glycerolwas 0.28 g/g, slightly higher than the yield on 2 % (v/v) crude glyc-erol (0.26 g/g). However, the ethanol production on 10% (v/v) crudeglycerol represents a yield of only 0.12 g/g. The results are summa-rised in Table 2.
3.4. Effect of different nitrogen sources on ethanol production
P. tannophilus was cultivated on NaNO3 and (NH4)2SO4 to inves-tigate the effect of nitrogen source for ethanol production. Theresults are shown in Fig. 4. Cells grown on NO�3 had higher biomassthan NOþ4 grown cells. Cells grown on NO�3 assimilated glycerol fas-ter than NOþ4 grown cells. The final biomass yield with NO�3 growncells was 62% higher than NOþ4 grown cells. The specific rate of eth-anol production by NOþ4 was almost the same as the rate obtainedwith NO�3 . The yield of ethanol in the presence of NOþ4 was 44%higher than NO�3 as nitrogen source. Results suggest that NOþ4 stim-ulates higher carbon flow to ethanol but a lower flow to biomass.However, NO�3 accelerates the rate of ethanol production. It is inaccordance with previous research that NO�3 stimulates the rateethanol production from xylose under aerobic condition by P.tannophilus(Jeffries, 1983a).
3.5. Ethanol tolerance on glycerol
There have been some previous reports regarding ethanol toler-ance of P. tannophilus on xylose (Zhao et al., 2010). However, as withpH, the ethanol tolerance is likely to change dependent on the car-bon source and was thus investigated during growth on glycerol.Small scale batch cultivations were performed in shake flasks with
-10 0 10 20 30 40 50 60 70 80
0.0
0.5
1.0
1.5
2.0
2.5
3.0
3.5(A)
Cel
l dry
wei
ght (
g/L)
Time(Hours)
0% 0.9% 1.7% 2.6% 3.4% 4.2% 5.1%
0 1 2 3 4 50.00
0.05
0.10
0.15
0.20
0.25
0.30(B)
µ max
(h-1)
Ethanol Concentration(V%)
µmax
Fig. 5. Shake flask cultivations of P. tannophilus on 2% (v/v) glycerol with differentadded ethanol concentrations, showing (A) cell dry weight (g/L) over 72 h ofcultivation and (B) maximum specific growth rates.
0 25 50 75 100 125 150 175 200 225 2500
10
20
30
40
50
60
70
Gly
cero
l(g/L
)
Time (Hours)
Glycerol
0
5
10
15
20
25
30
35
I IIIII
Feed in concentrated medium
CDW Ethanol
CD
W/E
than
ol (g
/L)
Feed in concentrated medium
Fig. 6. Time course of staged-batch fermentation with P. tannophilus on 5% (v/v) Batch 3 crude glycerol, concentrated medium was added to the fermentor after the glycerolwas depleted in 2 staged additions to give a final concentration of 5% (v/v). The figure shows glycerol concentration (g/L) (j), ethanol concentration (g/L) (.), and cell dryweight (g/L) (d). I, II and III indicates the three staged-batch phases used for calculation of quantitative fermentation data.
584 X. Liu et al. / Bioresource Technology 104 (2012) 579–586
43

varying amounts (0–5.1%) of ethanol being added to the cultures atthe point of inoculation. As is shown in Fig. 5A, biomass increasedmore slowly as the ethanol concentration increased. When theorganism was cultivated without ethanol addition, 3.11 g/Lbiomass was achieved after 72 h cultivation, where only 0.097 g/Lbiomass was obtained at 4.2% (v/v) ethanol. The maximum specificgrowth rate decreased with an increasing ethanol concentration(Fig. 5B). The maximum specific growth rate at 4.2% (v/v) ethanolwas 0.058 h�1 compared to 0.279 h�1 without ethanol added. Re-sults show no growth occurred on glycerol with 5.2 V% (41 g L�1)ethanol in the medium, which is consistent with the study wheregrowth of P. tannophilus ceased when 40 g L�1 ethanol was includedin xylose medium (Zhao et al., 2010). The ethanol tolerance for P.tannophilus could be improved in further studies for examplethrough UV mutagenesis and selection for more ethanol tolerantstrains (Watanabe et al., 2011).
3.6. Staged-batch fermentation on crude glycerol
In order to further improve glycerol conversion to ethanol andincrease yields in the fermentation processes, a staged-batch fer-mentation was performed (Fig. 6). In this process, medium wasadded to the fermenter after the glycerol was depleted in the initialbatch phase, and a second batch addition was made again onglycerol depletion. After 2 stages of batch addition of medium, anethanol production of 28.1 g/L was achieved on crude glycerol(Fig. 6, Table 3). This value is the highest reported so far in the lit-erature for microbial conversion of glycerol to ethanol. However, asTable 3 shows the productivity was still very low compared to bac-terial processes for ethanol production from glycerol. In order tofurther optimise conditions with a view to industrial scale produc-tion, further improvements in ethanol production could beachieved by designing a fed-batch process where feeding followsgrowth rate and substrate utilisation. Improving ethanol tolerancein the strain (e.g. by adaptive evolution) would be a furtherrequirement prior to larger scale development of this process, inorder to achieve the higher ethanol concentrations required forthe currently used recovery methods. However, this study repre-sents considerable steps towards developing a process for conver-sion of crude glycerol to ethanol, which could improve theeconomic and environmental burden for future biofuel production.
4. Conclusions
P. tannophilus is known for its ability to convert xylose, a com-ponent of plant biomass, to ethanol. This investigation shows thatcrude glycerol, the biodiesel by-product, can also be utilized as apotential low cost substrate by P. tannophilus. The organism wasnot sensitive to variations in crude glycerol batches from biodieselproduction based on different oil feedstocks. An ethanol produc-tion of 28.1 g/L was achieved in a staged-batch process, the highest
value for glycerol conversion to ethanol reported to date. P.tannophilus can, therefore, be considered as a potential versatilehost for biorefineries based on a variety of feedstocks.
Acknowledgements
This work was funded by the European Community’s 7th Frame-work Research Programme under Grant Agreement Number 213506(Project GLYFINERY) providing financial support to X.L. and M.W.The authors acknowledge the Technical University of Denmark Fer-mentation Platform at the Department of Systems Biology for pro-viding the fermentation equipment utilised in this study.
References
Agbogbo, F., Coward-Kelly, G., 2008. Cellulosic ethanol production using thenaturally occurring xylose-fermenting yeast, Pichia stipitis. Biotechnol. Lett.30, 1515–1524.
Canakci, M., Sanli, H., 2008. Biodiesel production from various feedstocks and theireffects on the fuel properties. J. Ind. Microbiol. Biotechnol. 35, 431–441.
Celik, E., Ozbay, N., Oktar, N., Calık, P., 2008. Use of biodiesel byproduct crudeglycerol as the carbon source for fermentation processes by recombinant Pichiapastoris. Ind. Eng. Chem. Res. 47, 2985–2990.
Choi, W., Hartono, M., Chan, W., Yeo, S., 2011. Ethanol production from biodiesel-derived crude glycerol by newly isolated Kluyvera cryocrescens. Appl. Microbiol.Biotechnol. 89, 1255–1264.
Converti, A., Perego, P., Domínguez, J.M., 1999. Microaerophilic metabolism ofPachysolen tannophilus at different pH values. Biotechnol. Lett. 21, 719–723.
da Silva, G.P., Mack, M., Contiero, J., 2009. Glycerol: a promising and abundantcarbon source for industrial microbiology. Biotechnol. Adv. 27, 30–39.
Durnin, G., Clomburg, J., Yeates, Z., Alvarez, P.J.J., Zygourakis, K., Campbell, P.,Gonzalez, R., 2009. Understanding and harnessing the microaerobic metabolismof glycerol in Escherichia coli. Biotechnol. Bioeng. 103, 148–161.
Fieldhouse, R., MacLennan, D.F., MacLennan, D.G., MacLennan, M.E., 2009. Processfor preparing nutritional, therapeutic or organoleptic products from crudeglycerol. Google Patents.
Fu, N., Peiris, P., 2008. Co-fermentation of a mixture of glucose and xylose to ethanolby Zymomonas mobilis and Pachysolen tannophilus. World J. Microbiol.Biotechnol. 24, 1091–1097.
Garcia-Ochoa, F., Gomez, E., 2009. Bioreactor scale-up and oxygen transfer rate inmicrobial processes: an overview. Biotechnol. Adv. 27, 153–176.
Hong, W.-K., Kim, C.-H., Heo, S.-Y., Luo, L., Oh, B.-R., Seo, J.-W., 2010. Enhancedproduction of ethanol from glycerol by engineered Hansenula polymorphaexpressing pyruvate decarboxylase and aldehyde dehydrogenase genes fromZymomonas mobilis. Biotechnol. Lett. 32, 1077–1082.
Jeffries, T.W., 1983a. Effects of nitrate on fermentation of xylose and glucose byPachysolen tannophilus. Bio-Technology 1, 503–506.
Jeffries, T.W., 1983b. Utilization of xylose by bacteria, yeasts, and fungi. Adv.Biochem. Eng. Biotechnol. 27, 1–32.
Johnson, D.T., Taconi, K.A., 2007. The glycerin glut: options for the value-addedconversion of crude glycerol resulting from biodiesel production. Environ. Prog.26, 338–348.
Kurtzman, C., 1983. Biology and physiology of the D-xylose fermenting yeastPachysolen tannophilus. Pentoses and Lignin, vol. 27. Springer, Berlin/Heidelberg,pp. 73–83.
Lynd, L., Wyman, C., Laser, M., Johnson, D., Landucci, R., 2005. Strategic biorefineryanalysis: review of existing biorefinery examples. NREL Subcontract Report(NREL/SR-510-34895).
Makri, A., Fakas, S., Aggelis, G., 2010. Metabolic activities of biotechnologicalinterest in Yarrowia lipolytica grown on glycerol in repeated batch cultures.Bioresour. Technol. 101, 2351–2358.
Table 3Comparison of quantitative data from ethanol production processes on glycerol.
Organism Fermentation method Total ethanol production(g/L)
Vol. ethanol productivity(g/L/h)
Escherichia coli EH05 (Durnin et al., 2009) Batch 20.7 0.22Klebsiella pneumoniae GEM167/pBR-pdc-adh (Oh et al., 2011) Fed-batch 25.0 0.78Kluyvera cryocrescens S26 (Choi et al., 2011) Batch 27.0 0.61Hansenula polymorpha HpDL1-L/pYH-pdc-adhB-dhaDKLM (Hong et al., 2010) Batch 3.1 0.02Saccharomyces cerevisiae YPH499fps1Dgpd2 (Yu et al., 2010) Batch 4.4 0.04
Pachysolen tannophilus CBS4044 (this study) Staged-batchPhase I 18.7 0.16Phase II 27.5 0.18Phase III 28.1 0.06
X. Liu et al. / Bioresource Technology 104 (2012) 579–586 585
44

Maleszka, R., Wang, P., Schneider, H., 1982. Ethanol production from D-galactoseand glycerol by Pachysolen tannophilus. Enzyme Microb. Technol. 4, 349–352.
Oh, B.-R., Seo, J.-W., Heo, S.-Y., Hong, W.-K., Luo, L.H., Joe, M.-h., Park, D.-H., Kim,C.H., 2011. Efficient production of ethanol from crude glycerol by a Klebsiellapneumoniae mutant strain. Bioresour. Technol. 102, 3918–3922.
Papanikolaou, S., Muniglia, L., Chevalot, I., Aggelis, G., Marc, I., 2002. Yarrowialipolytica as a potential producer of citric acid from raw glycerol. J. Appl.Microbiol. 92, 737–744.
Pokoo-Aikins, G., Nadim, A., El-Halwagi, M., Mahalec, V., 2010. Design and analysisof biodiesel production from algae grown through carbon sequestration. CleanTechnol. Environ. Policy 12, 239–254.
Sathesh-Prabu, C., Murugesan, A.G., 2011. Potential utilization of sorghum fieldwaste for fuel ethanol production employing Pachysolen tannophilus andSaccharomyces cerevisiae. Bioresour. Technol. 102, 2788–2792.
Shams Yazdani, S., Gonzalez, R., 2008. Engineering Escherichia coli for theefficient conversion of glycerol to ethanol and co-products. Metab. Eng. 10,340–351.
Silva, J.P.A., Mussatto, S.I., Roberto, I.C., Teixeira, J.A., 2011. Ethanol production fromxylose by Pichia stipitis NRRL Y-7124 in a stirred tank bioreactor. Braz. J. Chem.Eng. 28, 151–156.
Slininger, P.J., Bolen, P.L., Kurtzman, C.P., 1987. Pachysolen tannophilus: propertiesand process considerations for ethanol production from D-xylose. EnzymeMicrob. Technol. 9, 5–15.
Visser, W., Scheffers, W.A., Batenburg-van der Vegte, W.H., van Dijken, J.P., 1990.Oxygen requirements of yeasts. Appl. Environ. Microbiol. 56, 3785–3792.
Watanabe, T., Watanabe, I., Yamamoto, M., Ando, A., Nakamura, T., 2011. A UV-induced mutant of Pichia stipitis with increased ethanol production from xyloseand selection of a spontaneous mutant with increased ethanol tolerance.Bioresour. Technol. 102, 1844–1848.
Yu, K.O., Kim, S.W., Han, S.O., 2010. Reduction of glycerol production to improveethanol yield in an engineered Saccharomyces cerevisiae using glycerol as asubstrate. J. Biotechnol. 150, 209–214.
Zhao, L., Yu, J., Zhang, X., Tan, T., 2010. The ethanol tolerance of Pachysolentannophilus in fermentation on xylose. Appl. Biochem. Biotechnol. 160, 378–385.
586 X. Liu et al. / Bioresource Technology 104 (2012) 579–586
45

46
Chapter 4 Adaptive evolution studies for improving ethanol
tolerance of Pachysolen tannophilus CBS4044
Abstract
P. tannophilus CBS4044 can be considered as a potentially versatile host for biorefineries as
it can utilize both glycerol and pentoses (mainly xylose) in bioconversions. The studies in this
thesis have focused on the production of ethanol. Ethanol toxicity has been found to be a
limiting factor in ethanol production and it is therefore hypothesized that fermentation
performance could be improved in more ethanol tolerant strains. In the present study,
ethanol-tolerant mutants were obtained by adaptive evolution by increasing concentrations of
ethanol in the growth medium with glycerol as the sole carbon source. Two adapted cultures,
Evo4 and Evo2 of P. tannophilus CBS4044 were obtained by sequentially transferring 20 and
43 times respectively. Adapted mutant E243 was isolated by streaking out Evo2 culture on
YPD plates, and selecting a single colony. The ethanol tolerance and maximum specific
growth rate (µmax) of Evo4, Evo2 and E243, were improved at higher concentrations of
ethanol (> 4 % v/v) compared to the wild type. However, the µmax and the initial glycerol
consumption rate of both adapted cultures and mutant when ethanol was not present in the
medium were decreased compared to the wild type strain. Additionally, the ethanol
production from glycerol when ethanol was absent was decreased with the mutant and
adapted cultures. E243 only produced 4.5 g/L ethanol while the wild type strain could
produce 5.8 g/L ethanol.
Keywords: Pachysolen tannonphilus; ethanol tolerance; adaptive evolution; mutant.

47
1. Introduction
The yeast Pachysolen tannophilus has been investigated for ethanol production in previous
studies due to its ability to convert xylose and glycerol into ethanol (Liu et al., 2012;
Slininger et al., 1987). Any potential cell factory to be employed in a biorefinery should be
robust and tolerant to high concentrations of substrate and product. P. tannophilus was shown
not to be sensitive to variations in the crude glycerol batches supplied from one biodiesel
producer and can grow on relatively high concentrations of glycerol (up to 20% v/v).
However, the strain was sensitive to higher ethanol concentrations with the maximum
specific growth rate dropping to 0.058 h-1 when 4.2% v/v ethanol was present in the medium,
compared to 0.28 h-1 without ethanol present. The accumulation of ethanol during
cultivations causes stress to yeast cells, leading to a decrease in cell growth and reduced the
production of desired products (Dinh et al., 2008). The low ethanol tolerance of P.
tannophilus might be the factor which has inhibited further improvement in ethanol
production in the work presented in this thesis. It is believed that improving ethanol tolerance
will lead to improved ethanol productivities and yields (van Voorst et al., 2006).
Using genetic engineering to improve the ethanol tolerance of yeast is limited by the lack of
knowledge and the complexity of the ethanol stress-related mechanisms that inhibit cell
performance (Hu et al., 2007). The approaches for generating ethanol-tolerant yeast mutants
could be mutagenic or non-mutagenic adaptive evolution with ethanol stress as a selection
pressure (Stanley et al., 2010). Isolation of stable mutants with desired characteristics from
random mutagenesis could be very difficult, since the isolation procedure is time consuming
and mutants always contain undesired characteristics in addition to the desired traits. An
effective means of obtaining ethanol-tolerant yeast strains is by exposing yeast cells to a
stepwise increase in the level of ethanol stress (Dinh et al., 2008). Adaptive evolution works
on the principle that populations of cells adapt to their environment over time by natural
selection. Previous investigations about adaptive evolution for improved ethanol tolerance in
yeast (Dinh et al., 2008) showed that the ethanol-adapted cultures of S. cerevisiae had
slightly higher specific growth rates at higher concentrations of ethanol. By observing the
morphology and analyzing the fatty acid compositions of the cell membranes it was revealed
that the cells from the ethanol adapted cultures were larger and had a lower proportion of
C16:0 fatty acids in their cytoplasmic membrane (Dinh et al., 2008). Stanley et al. (2010)

48
generated ethanol tolerant mutants by chemical mutagenesis and adaptive evolution with
ethanol stress as a selective pressure. It was concluded that the mutants from both approaches
had increased growth rates when the cells were cultivated in sub-lethal ethanol
concentrations. It was suggested that the increased ethanol tolerance of the mutants might be
due to their elevated glycerol production rates (Stanley et al., 2010).
In this study, adaptive evolution with ethanol stress as the selection pressure was used as a
strategy to obtain the ethanol tolerant-mutants of P. tannophilus. The cells were cultivated in
medium without ethanol and then transferred to fresh medium containing a low ethanol
concentration followed by repetitive cultivation. After the cells adapted to the low ethanol
concentration, they were inoculated into medium containing a higher ethanol concentration
followed by repetitive cultivation. Adaptive evolution was used as an approach for increasing
ethanol tolerance of P. tannophilus CBS4044, potentially leading to improvement of
fermentation performance, high ethanol yield and productivity.
2. Materials and Methods
2.1 Microorganism and culture medium
Pachysolen tannophilus CBS4044 was used in this study. Both the adapted evolution and
growth experiments with the mutants were performed in defined medium, containing (per
liter) 0.67g yeast nitrogen base w/o amino acids (Difco, USA), 10g succinic acid and 6g
NaOH as a buffer system, and glycerol at 2% (v/v) as the sole carbon source.
2.2 Adaptive evolution under ethanol stress
All the cultivations were performed in test tubes with a working volume of 8ml defined
medium at 28oC under aerobic conditions. Adaptive evolution of the yeast was performed by
sequentially transferring and growing cells in defined media containing incrementally
increased concentrations of ethanol step by step (0%, 2%, 4%, 4.5%, 5%, 5.5%, 6% v/v). The
wild type strain of P. tannophilus was inoculated without ethanol in the medium and
cultivated overnight. Then the cells were transferred into a new medium with 2 % (v/v)
ethanol present with 2.5 % (v/v) inoculating rate. When the cells grew up after three days
(OD450nm around 10), the cells were transferred into medium with 4 % (v/v) ethanol present
with 2.5 % (v/v) inoculating rate. To obtain a strain with improved ethanol tolerance on

49
glycerol, the strain was repeatedly transferred into fresh minimal medium containing the
same ethanol concentration. After that, the cells were transferred to medium containing a
higher ethanol concentration followed by repetitive cultivations. As a back-up cells continued
to be cultivated at the lower ethanol concentration, while sequential transfer to higher ethanol
concentrations was performed. As a standard, achieving an OD450nm of 10 was required after
3 days of growth at the higher ethanol concentration. In some cases, growth took longer. If
the cells were not grown after one week in the medium with the higher concentration of
ethanol, further transfers were performed at the lower concentration before trying again to
increase the ethanol concentration. Four independent test tube parallels were performed for
adaptive evolution from 0% to 6% v/v of ethanol concentration, the respective culture series
obtained were named as Evo1, Evo2, Evo3, and Evo4.
2.3 Ethanol tolerance studies
The cells were grown at 28oC in defined medium on 2% (v/v) glycerol in 250ml Erlenmeyer
flasks with 100ml culture media with 0% (v/v), 2% (v/v), 4% (v/v), 5% (v/v), 6% (v/v)
ethanol added at the beginning of cultivation to characterize the growth profile and determine
the ethanol tolerance of adaptive cultures and mutant. All the growth experiments last for 72
hours with 100ml medium in the flasks.
2.4 Analytical methods
Growth was monitored by measuring OD450nm with a Shimadzu UV mini-1240
spectrophotometer (Shimadzu, Japan). Samples were taken periodically from the fermentor,
filtered through a 0.22 µm syringe filter and supernatants were preserved at -20°C for later
HPLC analysis. Concentrations of the substrate glycerol and all the main products: ethanol,
acetic acid, acetoin and 2, 3-butanediol in supernatants were measured by HPLC using an
Aminex87H column (Bio-Rad, USA). Separations were performed at 60°C, flow rate of
0.6ml/min and 5mM H2SO4 as mobile phase.
3. Results and discussion
3.1. Adaptive evolution to generate ethanol-tolerant mutants
In order to enhance the ethanol tolerance of P. tannophilus CBS 4044, an adaptive evolution
approach was used to generate ethanol-tolerant yeast mutants. The strain was repeatedly

50
transferred (up to 43 times) into fresh minimal medium with incremental increases in ethanol
concentration as shown in Figure 1. Variants with an advantageous phenotype are likely to
emerge when a microorganism is growing under a strong pressure of selection (for example
high temperature, uncommon substrate) (Dykhuizen, 1990). Along the adaptive evolution, the
cultures obtained after 20 transfers (Evo4) and 43 transfers (Evo2) were analyzed with
respect to their for ethanol tolerance, maximum growth rate, glycerol consumption and
ethanol production in submerged batch cultivation in shake flasks. The “adapted strain” used
for the ethanol tolerance analysis was isolated from the Evo2 culture by single colony
selection from colonies grown on YPD agar plate inoculated from the last cultivation (43
transfer no.). The strain was named as E243.
Figure 1. Adapted evolution was performed for enhancing ethanol tolerance with Pachysolen
tannophilus CBS4044. 4 independent lineages of P. tannophilus were transferred in defined medium
with increasing concentration of ethanol for 43 transfers. No ethanol was added to the defined
medium in the first transfer.
3.2. Growth characterization of adapted cultures Evo4, Evo2 and mutant E243
Figure 2 shows the biomass concentration after 72 hours cultivation of the 4 strains in the
presence of ethanol up to 6% (v/v). Higher biomass concentrations were obtained for the
adapted cultures and mutant in the presence of higher concentration of ethanol. For example,

51
at 5.2 % (v/v) ethanol the wild type strain showed almost no growth, while Evo4 produced
0.17g/L biomass at 5 % (v/v) ethanol and Evo2 produced 0.95g/L biomass at 4.9 % (v/v)
ethanol present. The mutant strain E243 produced 0.82 g/L biomass at 5.2 % (v/v) ethanol.
Figure 3 shows the maximum specific growth rates for each of the strains at each of the
ethanol concentrations added to the growth medium. At low ethanol concentrations the
adapted cultures and mutant performed generally poorer than the wild type strain. At higher
concentrations, however, the adapted cultures and mutant strain performed better compared
to the wild type.
0 1 2 3 4 5 6 7
0
1
2
3
4
Biom
ass
(g/L
)
Ethanol concentration (v/v %)
Evo4 Wide type Evo2 E243
Figure 2. Comparison of biomass concentrations obtained in shake flask cultivations with
P.tannophilus CBS4044 wild type strain, adapted cultures Evo4, Evo2 and mutant E243 after 72 hours
cultivation in defined medium containing 2% (v/v) glycerol and a range of ethanol concentrations at
28°C. Results represent the means of duplicate experiments.

52
0 1 2 3 4 5 6 7
0.00
0.05
0.10
0.15
0.20
0.25
0.30
µ max
(h-1)
Ethanol concentration (v/v%)
Evo4 Wide type Evo2 E243
Figure 3. µmax comparison of P.tannophilus CBS4044 wild type strain, adapted cultures Evo4 Evo2
and mutant E243 in defined medium containing 2% (v/v) glycerol and a range of ethanol
concentrations at 28°C in agitated flasks. Results represent the means of duplicate experiments.
With the continuation of adaptive evolution, the ethanol tolerance of P.tannophilus was
indeed improved when higher concentrations of ethanol were present in the medium.
However, the ethanol yield on glycerol and ethanol production levels were decreased (Figure
4B). When the organism was cultivated without ethanol addition, the yield of ethanol from
glycerol was around 0.25 g/g, while the yield for Evo4 was 0.23 g/g, Evo2 was 0.19 g/g and
E243 was also 0.19 g/g. The ethanol production level for wild type strains was 5.8 g/L, while
for Evo4 it was 5.5 g/L, Evo2 was 4.3 g/L and E243 was 4.5 g/L. It was noticed that for the
wild type strain and Evo4 at 24 hours after inoculation, there was some amount of ethanol
produced in the medium. However, Evo2 culture and E243 mutant started to produce ethanol
after 24 hours. For glycerol consumption (Figure 4A), it seems that the mutants consume
glycerol at lower rate at the initial 24 hours after inoculation compared to the wild type strain.
Evo2, E243 and wild type strain consumed all the glycerol at 72h cultivations. For the
biomass, the adaptive cultures Evo2, Evo4 and mutant E243 accumulated biomass at slower
rate compared to the wild type strain at the beginning of fermentation, however at 72 hours
cultivation they will end up with almost the same level of biomass (Figure 4C). The mutant
E243 accumulates a slightly higher biomass compared to wild type and adaptive cultures
Evo4 and Evo2.
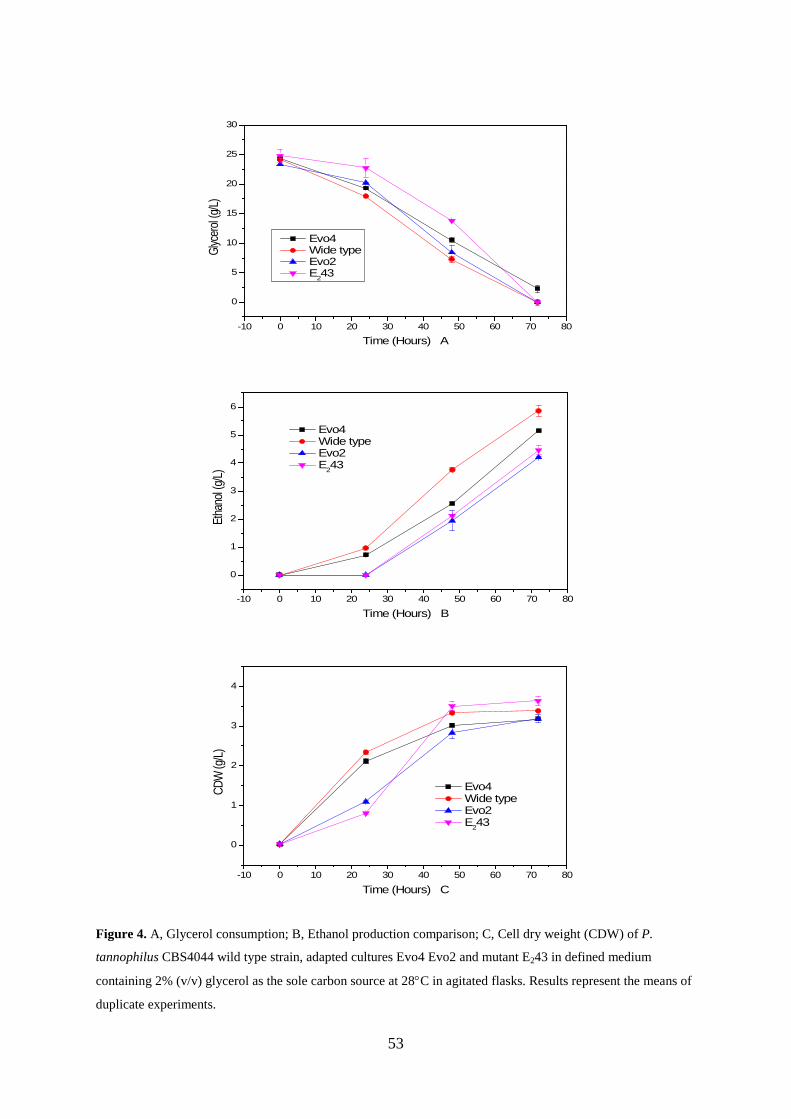
53
-10 0 10 20 30 40 50 60 70 80
0
5
10
15
20
25
30
Glyc
erol (g
/L)
Time (Hours) A
Evo4 Wide type Evo2 E243
-10 0 10 20 30 40 50 60 70 80
0
1
2
3
4
5
6
Etha
nol (g
/L)
Time (Hours) B
Evo4 Wide type Evo2 E243
-10 0 10 20 30 40 50 60 70 80
0
1
2
3
4
CDW
(g/L)
Time (Hours) C
Evo4 Wide type Evo2 E243
Figure 4. A, Glycerol consumption; B, Ethanol production comparison; C, Cell dry weight (CDW) of P.
tannophilus CBS4044 wild type strain, adapted cultures Evo4 Evo2 and mutant E243 in defined medium
containing 2% (v/v) glycerol as the sole carbon source at 28°C in agitated flasks. Results represent the means of
duplicate experiments.

54
4. Conclusion
In this study, the adaptive evolution of P. tannophilus towards improved ethanol tolerance
was investigated by exposing the cells to a stepwise increase of ethanol concentration in the
medium with repetitive cultivations. Using this adaptive evolution strategy two adapted
cultures (Evo4 and Evo2) and one mutant (E243) were investigated. Increases in biomass
concentration after 72 hours of growth were seen when the strains were cultivated in shake
flasks in the presence of increased concentrations of ethanol. The lethal ethanol concentration
for the wild type strain was 5.2% (v/v), however, the mutant and adapted cultures were able
to grow at this concentration.
The present study mainly focuses on characterization of the adapted cultures and mutant of P.
tannophilus. It has been shown in other studies that the lipids in the cell membrane and
ergosterol, which is one of the components of cell surface, are responsible for ethanol stress
tolerance in S. cerevisiae and other yeasts (Aguilera et al., 2006; Alexandre et al., 1994;
Thomas et al., 1978). Some studies also showed that the addition of ethanol causes a cell-
cycle delay and results in an increase in cell size (Dinh et al., 2008; Kubota et al., 2004). Due
to the time limitation of the PhD study period, the mechanism of ethanol tolerance has not
been investigated further in with P. tannophilus. It would be interesting to look into the lipid
composition, cell size and perform genome and transcriptional analysis in order to unveil the
mechanism behind the performance of ethanol-tolerant mutants of P. tannophilus.
References
Aguilera, F., Peinado, R.A., Millán, C., Ortega, J.M., Mauricio, J.C. 2006. Relationship
between ethanol tolerance, H+-ATPase activity and the lipid composition of the
plasma membrane in different wine yeast strains. International Journal of Food
Microbiology, 110(1), 34-42.
Alexandre, H., Rousseaux, I., Charpentier, C. 1994. Relationship between ethanol tolerance,
lipid composition and plasma membrane fluidity in Saccharomyces cerevisiae and
Kloeckera apiculata. FEMS Microbiology Letters, 124(1), 17-22.
Dinh, T.N., Nagahisa, K., Hirasawa, T., Furusawa, C., Shimizu, H. 2008. Adaptation of
Saccharomyces cerevisiae Cells to High Ethanol Concentration and Changes in Fatty
Acid Composition of Membrane and Cell Size. PLoS ONE, 3(7), e2623.

55
Dykhuizen, D.E. 1990. Experimental Studies of Natural Selection in Bacteria. Annual Review
of Ecology and Systematics, 21(ArticleType: research-article / Full publication date:
1990 / Copyright © 1990 Annual Reviews), 373-398.
Hu, X.H., Wang, M.H., Tan, T., Li, J.R., Yang, H., Leach, L., Zhang, R.M., Luo, Z.W. 2007.
Genetic Dissection of Ethanol Tolerance in the Budding Yeast Saccharomyces
cerevisiae. Genetics, 175(3), 1479-1487.
Kubota, S., Takeo, I., Kume, K., Kanai, M., Shitamukai, A., Mizunuma, M., Miyakawa, T.,
Shimoi, H., Iefuji, H., Hirata, D. 2004. Effect of Ethanol on Cell Growth of Budding
Yeast: Genes That Are Important for Cell Growth in the Presence of Ethanol.
Bioscience, Biotechnology, and Biochemistry, 68(4), 968-972.
Liu, X., Jensen, P.R., Workman, M. 2012. Bioconversion of crude glycerol feedstocks into
ethanol by Pachysolen tannophilus. Bioresource Technology, 104(0), 579-586.
Slininger, P.J., Bolen, P.L., Kurtzman, C.P. 1987. Pachysolen tannophilus: Properties and
process considerations for ethanol production from d-xylose. Enzyme and Microbial
Technology, 9(1), 5-15.
Stanley, D., Fraser, S., Chambers, P., Rogers, P., Stanley, G. 2010. Generation and
characterisation of stable ethanol-tolerant mutants of Saccharomyces cerevisiae.
Journal of Industrial Microbiology & Biotechnology, 37(2), 139-149.
Thomas, D.S., Hossack, J.A., Rose, A.H. 1978. Plasma-Membrane lipid composition and
ethanol tolerance in Saccharomyces cerevisiae. Archives of Microbiology, 117(3),
239-245.
van Voorst, F., Houghton-Larsen, J., Jønson, L., Kielland-Brandt, M.C., Brandt, A. 2006.
Genome-wide identification of genes required for growth of Saccharomyces
cerevisiae under ethanol stress. Yeast, 23(5), 351-359.

56
Chapter 5 Draft genome sequence of the yeast Pachysolen
tannophilus CBS 4044
Xiaoying Liu, Rolf Sommer Kaas, Peter Ruhdal Jensen, Mhairi Workman*
Submitted to Eukaryotic Cell
Abstract
A draft genome sequence of the non-conventional yeast P. tannophilus CBS 4044 is
presented. The organism has potential to be developed as a cell factory for biorefineries, due
to its ability to utilize agricultural and industrial wastes. The sequenced genome size was
12,238,196 bp, consisting of 34 scaffolds. 4,463 genes were annotated with function from
5,346 predicted open reading frames. The genome sequence data provides a better
understanding of the genetic background of this strain and will facilitate improved industrial
applications of P. tannophilus.
Keywords: Pachysolen tannophilus; genome; annotation; chromosome.

57
The yeast, Pachysolen tannophilus was first isolated from wood extracts used in leather
tanning by Boidin and Adzet in 1957 (Kurtzman, 1983). This yeast has gained considerable
attention due to its ability to ferment D-xylose, the main component in hemicellulose plant
residues (Fu & Peiris, 2008; Schneider et al., 1981; Slininger et al., 1987). A number of
studies have investigated fuel ethanol production from D-xylose by P.tannophilus (Sathesh-
Prabu & Murugesan, 2011; Slininger et al., 1987; Zhao et al., 2010). Additionally, it has been
reported that P. tannophilus could ferment the common sugars (glucose, mannose and
galactose) occurring in hemicellulose hydrolysate mixtures, with the exception of L-
arabinose (Slininger et al., 1987). It has been shown recently that P. tannophilus could
produce ethanol from glycerol, indicating the potential application of this organism in
converting other waste streams (Liu et al., 2012). The whole-genome sequencing of P.
tannophilus was performed in this study to provide genetic information on this yeast. This is
a necessary step towards genetic engineering of the strain and its potential further
development as a cell factory for industrial applications.
The genome of Pachysolen tannohpilus CBS4044 was sequenced by a whole-genome
shotgun sequencing strategy with an Illumina Genome Analyser at Beijing Genomics
Institute (BGI, Shenzhen, China). Raw data of short reads was assembled into 279 contigs
and the contigs were ordered into 34 scaffolds (>2kb) with N50 size of 1.1Mb by using the
SOAPdenovo package (Li et al., 2010) which is self-developed by BGI. Augustus v2.5
software (Stanke et al., 2004), which was trained for predicting genes in Debaryomyces
hansenii, was utilized to identify protein-coding genes in the genome and the putative amino
acid sequences were used for subsequent gene functional annotation analysis. The functional
annotation was accomplished by BlastP analysis (E-value < 1x10-5) of protein sequences in
the databases (COG, KEGG) and the best hit was selected. Pulsed field gel electrophoresis
(PFGE) (Maringele & Lydall, 2006) was performed to predict the chromosome numbers and
approximate sizes in P. tannophilus. The program for PFGE was 48h at 3V/cm with a 500s
switch time at an included angle of 106o, 0.5xTBE on 0.75% LMP (Low melting point)
Agarose and at 14°C.
The total length of the sequenced genome was 12,238,196 bp (without N) with a GC content
of 29.82%. Total 1970.8 Mb raw data was sequenced representing around 145 fold coverage
of the P.tannophilus genome. 5,346 protein-coding genes (CDSs) set were predicted, with
4,463 (83.5%) genes annotated with function.

58
Four coding sequences of P. tannphilus were retrieved from Genbank to compare with the
annotated CDS sequences in the sequenced genome. There were 2 bp differences among
4803bp length. Furthermore, 13 full-length coding sequences were PCR amplified and
resequenced to estimate the accuracy of the genome. The sequencing results showed that the
genome sequencing results were very accurate. The total resequenced lengths were 21,111bp.
Based on PFGE results, six chromosomal bands were separated with two of the bands
probably migrating as doublets. The sizes of chromosomal bands were estimated to be
around 2.9±0.05Mb, 2.1±0.04Mb, 1.9±0.05Mb, 1.6±0.08Mb (doublet), 1.3±0.07Mb
(doublet), 0.98±0.02Mb based on comparison with the yeast marker Hansenula wingei. The
estimated genome size of P. tannophilus (CBS4044) was approximately 13.6±0.4Mb with an
estimated 8 chromosomes in P. tannophilus.
P. tannophilus was characterized during ethanol production from the uncommon carbon
sources xylose and glycerol. It produced ethanol, acetic acid, acetoin and 2, 3-butanediol on
glycerol as the sole carbon source (Liu et al., 2012). The genome analysis revealed that all the
genes coding for enzymes involved in glycolysis, xylose degradation, glycerol assimilation
and formation, products synthesis were present in the genome sequences.
Nucleotide sequence accession number. The draft genome sequences of P. tannophilus
CBS 4044 were deposited in GenBank.
This work was funded by the European Community’s 7th Framework Research Programme
under Grant Agreement Number 213506 (Project GLYFINERY) providing financial support
to XL and MW.
References
Fu, N., Peiris, P. 2008. Co-fermentation of a mixture of glucose and xylose to ethanol by
Zymomonas mobilis and Pachysolen tannophilus. World Journal of Microbiology and
Biotechnology, 24(7), 1091-1097.
Kurtzman, C. 1983. Biology and physiology of the Pentoses and Lignin, Vol. 27, Springer
Berlin / Heidelberg, pp. 73-83.

59
Li, R., Zhu, H., Ruan, J., Qian, W., Fang, X., Shi, Z., Li, Y., Li, S., Shan, G., Kristiansen, K.,
Li, S., Yang, H., Wang, J., Wang, J. 2010. De novo assembly of human genomes with
massively parallel short read sequencing. Genome Research, 20(2), 265-272.
Liu, X., Jensen, P.R., Workman, M. 2012. Bioconversion of crude glycerol feedstocks into
ethanol by Pachysolen tannophilus. Bioresource Technology, 104, 579-586.
Maringele, L., Lydall, D. 2006. Pulsed-Field Gel Electrophoresis of Budding Yeast
Chromosomes Yeast Protocol, (Ed.) W. Xiao, Vol. 313, Humana Press, pp. 65-73.
Sathesh-Prabu, C., Murugesan, A.G. 2011. Potential utilization of sorghum field waste for
fuel ethanol production employing Pachysolen tannophilus and Saccharomyces
cerevisiae. Bioresource Technology, 102(3), 2788-2792.
Schneider, H., Wang, P.Y., Chan, Y.K., Maleszka, R. 1981. Conversion of D-xylose into
ethanol by the yeast Pachysolen tannophilus. Biotechnology Letters, 3(2), 89-92.
Slininger, P.J., Bolen, P.L., Kurtzman, C.P. 1987. Pachysolen tannophilus: Properties and
process considerations for ethanol production from d-xylose. Enzyme and Microbial
Technology, 9(1), 5-15.
Stanke, M., Steinkamp, R., Waack, S., Morgenstern, B. 2004. AUGUSTUS: a web server for
gene finding in eukaryotes. Nucleic Acids Research, 32(suppl 2), W309-W312.
Zhao, L., Yu, J., Zhang, X., Tan, T. 2010. The Ethanol Tolerance of Pachysolen tannophilus
in Fermentation on Xylose. Applied Biochemistry and Biotechnology, 160(2), 378-
385.

60
Chapter 6 Development of a transformation and selection system
for Pachysolen tannophilus
Abstract
Pachysolen tannophilus has been studied in this PhD project as a host for production of
ethanol from glycerol. Much time has been devoted to performing submerged cultivations
with this strain at a variety of scales and under different cultivation conditions with a view to
optimizing the ethanol production process. Certain progress has been made by this process
optimization strategy. However, in order to further improve the organism as a cell factory,
genetic engineering could be a useful tool. Specifically, for the purposes of this study, it
would be advantageous to investigate the glycerol consumption and ethanol synthesis
pathways. A first step for genetic engineering is the development of a suitable transformation
and selection system in P. tannophilus, as such genetic tools do not exist for this strain at
present. In this chapter, the creation and selection of antibiotic (Geneticin) resistance mutants
and uracil auxotrophic mutants is described. Unfortunately, the strategies and methods
employed here did not yield successful results. Much more effort and knowledge about the
genetic background of P. tannophilus is needed in order to develop a successful
transformation procedure.
Keywords: Pachysolen tannophilus; G418; URA3; transformation.

61
1. Introduction
P. tannophilus was considered in this study as a potential versatile host for biorefineries
based on its ability to convert the carbon sources xylose and glycerol to ethanol. While a lot
of research has been performed on physiological characterization of the strain, very little is
known about the genetic background of P. tannophilus. With the aim of further improving
ethanol production, it is necessary to better understand the ethanol synthesis pathway in P.
tannophilus and to design tools for subsequent genetic engineering of this yeast species. The
whole-genome sequencing of P. tannophilus aided a better understanding of the genetic
background, which opens the possibility of genetically engineering the strain.
In order to work towards metabolic engineering of the yeast strain, a first step is development
of an efficient transformation and selection system in P. tannophilus. Such genetic tools do
not exist for this strain at present. However, it was reported once that that the lithium treated
hexose negative mutants of the yeast P. tannophilus survived in the presence of D-glucose or
D-fructose by the introduction of a S. cerevisiae HXK2 gene (Wedlock & Thornton, 1989). It
indicates the possibility of transformation of this strain.
With regard to development of a transformation selection system, antibiotic resistance is a
simple and powerful tool. The only requirement is that the wide type cells should be killed or
prevented from growing by the antibiotic. The fact that the transformed cells harbouring the
antibiotic resistance gene grow in the presence of antibiotic could be utilized as the selection
principle. It has been shown that the Kanr gene from the bacterial transposable element Tn903
confers resistance to the antibiotic geneticin (G418) in yeast (Jimenez & Davies, 1980).
Many prokaryotes and eukaryotes including plants and yeasts are sensitive to G418. Jimenez
and Davies (1980) demonstrated that Sacchromyces cerevisiae expressing the Kanr gene of
Tn903 (Tn601) become resistant to G418. The aminoglycoside phosphotransferase encoded
by Kanr inactivates the antibiotic G418 and renders transformed cells resistant to the G418
(Sreekrishna et al., 1984). The similar antibiotic resistance selection has been successfully
employed in many different types of yeasts and fungi (Sreekrishna et al., 1984; Suárez &
Eslava, 1988). In developing the G418 selection procedure, we have constructed the
recombinant shuttle vectors containing the KanMX module, which are hybrids of the coding
sequence of the kanr gene of transposon Tn903 and transcriptional and translational control
sequences from the TEF gene of the filamentous fungus Ashbya gossypii (Steiner &

62
Philippsen, 1994). The KanMX module with long flanking sequences homologous to P.
tannophilus GUT1 was also constructed in order to obtain G418-resistant P. tannophilus.
In addition to the antibiotic selection system, most transformation systems are based on
auxotrophic markers. The most commonly used method is based on the orotidine-5-phosphate
decarboxylase (URA3) gene which is widely used in yeast and fungi (Boeke et al., 1984;
François et al., 2004; Goosen et al., 1987). URA3 is a gene that encodes orotidine 5-
phosphate decarboxylase (ODCase), which is an enzyme that catalyzes the last step in the
synthesis of pyrimidine ribonucleotides in yeast RNA (Rose et al., 1984). The URA3 gene
works as a counter selectable marker and allows positive and negative selections. The yeast
cells which cannot grow lack oODCase activity (ura3) unless uracil or uridine is added into
the medium. After the URA3 gene was reintroduced into cells, the auxotrophic cells can start
to grow (positive selection). In contrast, if 5-Fluoroorotic acid (5-FOA) is added into the
media, the cells could convert 5-FOA into the toxic compound 5-fluorouracil leading to death
(negative selection). Since URA3 allows two different selection systems, it is one of the most
important genetic markers in yeast genetic modification. Here the methods and results for
making URA3 selection marker in P. tannophilus are also described.
2. Materials and Methods
2.1 Strains and plasmids
Pachysolen tannophilus CBS4044 was used throughout this study. S. cerivisae CEN.PK 113-
5D (MATa MAL2-8c SUC2 ura3-52) was used for the control experiments of the
transformation procedure. Plasmid PUG6 (Güldener et al., 1996) harbouring the KanMX
module (1473bp) was used as a template for amplification of the G418 resistance cassette. A
centromeric plasmid pCM188 (Garí et al., 1997) was used for constructing the G418
resistance vector used for transformation.
2.2 Medium and culture conditions
P. tannophilus was cultivated on yeast peptone dextrose (YPD) medium (1% yeast extract,
2% peptone, and 2% glucose) and synthetic dextrose medium (SD) (0.67% yeast nitrogen
base without amino acid and 2% glucose) media supplemented with 0.03% Uracil and 0.15%
5-FOA were used for selection of mutants. Different concentrations of G418 were

63
supplemented in YPD agar medium for selection of G418 resistance transformants. For the
preparation of solid plates, 2% agar was added.
2.3 Sensitivity of P. tanniphilus to G418 and 5-FOA
The sensitivity of P. tannophilus with antibiotic G418 was determined by plating cell
suspensions (approximately 1x 106 cells/plate, 100ul OD450=1) on YPD containing various
concentrations of G418 (0, 25, 50, 75, 100 µg/ml) and plates were incubated at 30°C for 2-3
days. The sensitivity of P. tannophilus with 5-FOA was performed by plating cell
suspensions (approximately 1x 107 cells/plate, l ml OD450=1) on SD media containing various
concentrations of 5-FOA (0, 0.5, 1, 1.5, 2 mg/ml) and plates were incubated at 30°C for 5-10
days.
2.4 Construction of vectors for selection of G418 resistance transformants
In this study, two vectors carrying the KanMX cassette were constructed. The first vector was
constructed by combination of the pCM188 plasmid and KanMX cassette. The pCM188
plasmid was digested with BamHI/SalI enzymes (Fermentas). The Phusion® High-Fidelity
DNA Polymerase (Fermentas) was used for amplification of the KanMX cassette (1473bp)
with the PUG6 plasmid as the template. The amplified KanMX cassette was then digested
with BamHI/SalI enzymes for cloning into the digested pCM188 plasmid. The digested
KanMX and pCM188 plasmid were ligated and the reaction mix was used directly to
transform chemically competent E. coli DH5α cells. The construct was named as pCM188-
KanMX and was verified by sequencing. Lithium chloride (Gietz et al., 1995) treated cells
were transformed with the pCM188-KanMX vector. After transformation, the cells were
cultivated in YPD liquid medium without G418 for 4 hours and then plated on YPD + G418
agar plates for transformants with G418 resistance selection. Meanwhile, S. cerevisiae
CEN.PK 113-5D was used for control experiments.
The second vector was constructed as an integrative vector with the KanMX cassette flanking
with PtGUT1 partial sequences. Gene targeting substrates were constructed based on PCR
methods using the cloning-free PCR-based allele replacement strategy (Erdeniz et al., 1997).
In total, 4 individual pieces of DNA need to be fused to create the substrates as is shown in
Figure 1.

64
Figure 1. Schematic representation of the homologous recombination in yeast cells during the gene
targeting experiment. The gene targeting substrates should be integrated into the predestined locus of
P. tannophilus by homologous recombination.
The primers Pt_GUT1_Up_F and D_Pt_GUT1_Up_R, C_Pt_GUT1_Dw_F and
Pt_GUT1_Dw_R were used to amplify the PtGUT1 upstream sequences and downstream
sequences. The primers dKanMX_5" and KanMX_3"_int, KanMX_5"_int and cKanMX_3"
were utilized to amplify the KanMX N-term and KanMX C-term sequences. To construct the
first bipartite substrate, the fragment upstream of PtGUT1 was fused to the KanMX N-term
fragment using primers Pt-GUT1-Up_F and KanMX_3"_int. The second bipartite substrate
was obtained by fusion of the KanMX C-term fragment with the downstream PtGUT1
fragment using primers Pt-GUT1-Dw_F and cKanMX_3". The transformation was
performed by the lithium acetate method (Gietz et al., 1995) for chromosomally homologous
integration. After transformation, the cells were cultivated in YPD liquid medium without
G418 for 4 hours and then plated on YPD + G418 agar plates for transformants with G418
resistance selection.

65
Table 1. Primers used for constructing vectors
Primer Name Sequence 5' to 3'
KanMX_F_BamHI CGCGGATCCTAGGTCTAGAGATCTGTTTAGC
KanMX_R_SalI TGCGGTCGACATTAAGGGTTCTCG AGAGC
Pt_GUT1_Up_F CAATCCAACTTTCAACCCTCAA
D_Pt_GUT1_Up_
R
GCAGGGATGCGGCCGCTGACTCACAACTAAAGGTTCAAACGAC
C_Pt_GUT1_Dw_
CCGCTGCTAGGCGCGCCGTGTGACTTCTAAACCTCAATTTGC
Pt_GUT1_Dw_R TGAGCGACCTGAGAAAAATAC
dKanMX_5" GTCAGCGGCCGCATCCCTGCTACGCTGCAGGTCGACAA
KanMX_3"_int GACTCGTCCAACATCAATACA
KanMX_5"_int AAAAGACTCACGTTTCGAG
cKanMX_3" CACGGCGCGCCTAGCAGCGGGAGTCAGTGAGCGAGGAA
2.5 Ura- Mutants selection
P. tannophilus was grown overnight in YPD liquid medium, centrifuged, washed twice with
sterile water, and resuspended in 1 ml of sterile water. Aliquots (200µl) of cells were then
spread on 5-FOA plates and exposed to 0, 30J and 40J UV (around 20 – 40% killing
efficiency). Colonies were picked after 5 days at 28°C and tested for a uracil requirement by
assessing growth on minimal medium with and without uracil.
3. Results
3.1 G418 antibiotics as the transformation marker
P. tannophilus showed no growth when the concentration of antibiotic G418 in medium was
greater than 50µg/ml. Therefore, G418 was chosen for being used as a transformation
marker. It has been shown that the ARS (autonomously replication sequences) of S. cerevisae
in YRp7 plasmid functioned in the lithium treated hexose negative mutants of the yeast P.
tannophilus (Wedlock & Thornton, 1989). The ARS of the pCM188 plasmid is also from
S.cerevisiae, which indicates that the ARS should also work in P.tannophilus. Lithium
chloride treated cells were transformed with the pCM188-KanMX vector. Meanwhile, S.
cerevisiae was used for control experiments. KanMX gave resistance to G418 if this cassette

66
could be successfully expressed. It was shown from the results that S. cerevisiae with the
KanMX grew on the YPD+G418 (100 µg/ml) plates. However, no transformants of P.
tannophilus were obtained on YPD+G418 (50 µg/ml) selection plates. Lithium chloride
treated cells were transformed with KanMX flanking PtGUT1 upstream and downstream
fragments of P.tannophilus for homologous recombination. However, no transformants were
obtained on YPD+G418 (50 µg/ml) selection plates.
3.2 URA3 as the transformation marker
The possibility of making a uracil auxotrophic marker was also investigated. The first step in
the development of the transformation system was the isolation of a stable ura3 auxotrophic
mutant. The mutants could be selected on media containing 5-FOA. The growth of wide-type
P. tannophilus was inhibited when 1.5 mg/ml 5-FOA was supplemented in the medium. The
cells of P. tannophilus were UV mutated and then selected onto SD supplemented with uracil
and 5-FOA plates for 5 days cultivation (Figure 2). The mutants from the plates were re-
streaked onto the same plates for isolating single colonies and confirming the mutants’
resistance to 5-FOA. Then the single colonies were streaked onto SD plates to check if they
were ura3 auxotrophic mutants. The ura3 auxotrophic mutants are expected to have no
growth on SD plates and grow only on the SD + Uracil plates. However, from the results
obtained here, several mutants exhibited growth when the cells were streaked out on SD
plates. Several mutants which lacked of growth on SD plates also exhibited no growth when
the cells were streaked out onto SD + Uracil plates.
Figure 2. P. tannophilus cells were mutated with different doses of UV radiation on SD + Uracil +
5FOA plates.
4. Discussion
oJ 40J 30J

67
The aim of this study was the development of a transformation and selection system for the
ethanol producing yeast P. tannophilus based on the G418 antibiotic resistance marker and
uracil auxotrophic marker. However, no right mutants were obtained during the time frame of
the work.
Many factors might influence the transformation procedure including the vector composition,
replicons, genetic markers, transformation methods and efficiency, vector stability and copy
numbers, the promoters and the host strain (Wang et al., 2001). The success of a
transformation procedure depends on all the factors working properly. It has been shown that
the ARS from S. cerevisae functioned in the lithium treated hexose negative mutants of the
yeast P. tannophilus (Wedlock & Thornton, 1989). Since the ARS in the pCM188 plasmid is
also from S. cerevisiae, the vector of pCM188-KanMX should replicate in the cells of P.
tannophilus once the vector gets inside of the cells. For the chromosomally homologous
integration, it also can eliminate the ARS factor. However, the vector stability, TEF promoter
activity, transformation methods and efficiency and the incubation time for expression G418
resistance might still influence the transformation results. In order to successfully transform
P. tannophilus, the strategy employed could be further improved. This could involve
constructing a new plasmid with the original automatic replication sites, and original
promoter from P. tannophilus and trying to transform P. tannophilus with this plasmid. This
may eliminate the uncertain factors influencing the transformation procedure. In addition,
other transformation methods such as electroporation of P. tannophilus could also be tested.
For obtaining the ura3 auxotrophic mutants, selection of more 5-FOA mutants might be
necessary for obtaining mutants with right phenotype. The difficulty of obtaining
auxotrophic mutants might be related to the complex life cycle of P. tannophilus. For
instance, low concentrations of carbon source in the medium leads to vegetative cells
transition to the mating process (Bolotnikova et al., 2005). An excess of microelements,
vitamins, potassium and phosphorous activates the sexual stage of the yeast (Bolotnikova et
al., 2005). In comparison, yeasts of the genera Saccharomyces and Schizosaccharomyces
which have a simple life cycle are subject to genetic and biotechnological studies.
Since the approaches of trying to set up a transformation and selection system did not yield
successful results, an alternative strategy could be transferring the glycerol assimilation
pathways in P. tannophilus into the well characterized yeast S. cerevisise, where the genetic

68
background are well understood and the genetic engineering tools are available. Heterologous
expression of the enzyme or the metabolic pathways involved in glycerol metabolism in S.
cerevisiae was investigated in the following chapter.
References
Boeke, J.D., Croute, F., Fink, G.R. 1984. A positive selection for mutants lacking orotidine-
5′-phosphate decarboxylase activity in yeast: 5-fluoro-orotic acid resistance.
Molecular and General Genetics MGG, 197(2), 345-346.
Bolotnikova, O.I., Mikhailova, N.P., Shabalina, M.V., Bodunova, E.N., Ginak, A.I. 2005.
Conditions Favoring Differentiation and Stabilization of the Life Cycle of the Yeast
Pachysolen tannophilus. Microbiology, 74(4), 415-419.
François, F., Chapeland-Leclerc, F., Villard, J., Noël, T. 2004. Development of an integrative
transformation system for the opportunistic pathogenic yeast Candida lusitaniae using
URA3 as a selection marker. Yeast, 21(2), 95-106.
Garí, E., Piedrafita, L., Aldea, M., Herrero, E. 1997. A Set of Vectors with a Tetracycline-
Regulatable Promoter System for Modulated Gene Expression in Saccharomyces
cerevisiae. Yeast, 13(9), 837-848.
Gietz, R.D., Schiestl, R.H., Willems, A.R., Woods, R.A. 1995. Studies on the transformation
of intact yeast cells by the LiAc/SS-DNA/PEG procedure. Yeast, 11(4), 355-360.
Goosen, T., Bloemheuvel, G., Gysler, C., Bie, D.A., Broek, H.W.J., Swart, K. 1987.
Transformation of Aspergillus niger using the homologous orotidine-5′-phosphate-
decarboxylase gene. Current Genetics, 11(6), 499-503.
Güldener, U., Heck, S., Fiedler, T., Beinhauer, J., Hegemann, J.H. 1996. A New Efficient
Gene Disruption Cassette for Repeated Use in Budding Yeast. Nucleic Acids
Research, 24(13), 2519-2524.
Jimenez, A., Davies, J. 1980. Expression of a transposable antibiotic resistance element in
Saccharomyces cerevisiae. Nature, 287(5785), 869-871.
Rose, M., Grisafi, P., Botstein, D. 1984. Structure and function of the yeast URA3 gene:
expression in Escherichia coli. Gene, 29(1–2), 113-124.
Sreekrishna, K., Webster, T.D., Dickson, R.C. 1984. Transformation of Kluyveromyces lactis
with the kanamycin (G418) resistance gene of Tn905. Gene, 28(1), 73-81.

69
Steiner, S., Philippsen, P. 1994. Sequence and promoter analysis of the highly expressed TEF
gene of the filamentous fungus Ashbya gossypii. Molecular and General Genetics
MGG, 242(3), 263-271.
Suárez, T., Eslava, A.P. 1988. Transformation of Phycomyces with a bacterial gene for
kanamycin resistance. Molecular and General Genetics MGG, 212(1), 120-123.
Wang, T.-T., Choi, Y.-J., Lee, B.H. 2001. Transformation Systems of non-Saccharomyces
Yeasts. Critical Reviews in Biotechnology, 21(3), 177-218.
Wedlock, D.N., Thornton, R.J. 1989. Transformation of a glucose negative mutant of
Pachysolen tannophilus with a plasmid carrying the cloned hexokinase PII gene from
Saccharomyces cerevisiae. Biotechnology Letters, 11(9), 601-604.

70
Chapter 7 Functional studies of genes involved in transport and
metabolism of glycerol in Pachysolen tannophilus
Xiaoying Liu, Peter Ruhdal Jensen, Mhairi Workman*
Submitted to Applied and Environmental Microbiology
Abstract
Pachysolen tannophilus is considered as a potential microbial cell factory since it can
metabolize a variety of the carbon sources found in low cost feedstocks including glycerol
and xylose. The xylose utilisation pathways have been extensively studied in this organism.
However, the mechanism behind glycerol metabolism is poorly understood. The genome
sequence of P. tannophilus CBS4044 has become available recently. The genes related to
glycerol transport and metabolism in P. tannophilus have been identified here by blasting the
genome sequence with the sequences of genes known to have these functions in S. cerevisiae.
Quantitative real-time PCR was performed to unveil the expression pattern of the genes
during growth on glycerol and glucose as sole carbon source. The genes involved in glycerol
transport in P.tannophilus have been cloned and successfully expressed in S. cerevisiae
(CEN-PK 1135D) strains. The transformed S. cerevisiae strains with heterologous genes
showed improved growth and glycerol consumption rates with glycerol as the sole carbon
source.
Key works: P. tannophilus; genes involved in glycerol metabolism; glycerol transporter;
FPS1; STL1.

71
1. Introduction Pachysolen tannophilus is known for its ability to ferment D-xylose, one of the major
components in hemicellulose plant residues, to ethanol (Kurtzman, 1983). In recent years,
due to the dramatic increase in production of biodiesel, glycerol (a by-product of biodiesel
production) has also been considered as a potential alternative carbon source for industrial
bioprocesses. P. tannophilus has been shown to be capable of converting this crude glycerol
to ethanol under microaerobic conditions (Liu et al.). Further optimization of the previously
described processes would be beneficial and could be achieved through genetic modification
of the organism directed towards improved glycerol uptake and processing.
The whole genome of P. tannophilus CBS4044 has been sequenced (Chapter 5), and with
this, the possibility for genetic comparison of P. tannophilus with other better characterized
yeasts is possible. A great number of studies have investigated glycerol transport,
consumption and production by many different types of yeast, but the glycerol transport and
metabolic pathways in P. tannophilus have not been studied so far. Since the genome
sequence is now known, it is possible to look at the mechanism behind glycerol metabolism
in P. tannophilus.
Glycerol transport is the first barrier for glycerol utilization in a microbial cell. In
Saccharomyces cerevisiae, glycerol enters the cell by two different mechanisms: a low
affinity transport system and a high affinity proton symport system (Lages and Lucas, 1997).
When glucose is present, glycerol diffuses into the cells through a glycerol channel by
facilitated transport, a process encoded by the FPS1 gene (Sutherland et al., 1997; Tamás et
al., 1999). However, during growth on non-fermentable carbon sources (glycerol, acetate,
ethanol) an active uptake system driven by a proton motive force ensures the uptake of
glycerol (Holst et al., 2000; Lages & Lucas, 1997). Two multi-membrane-spanning proteins
encoded by GUP1 and GUP2 (Glycerol Uptake Protein), were first identified as being
involved in active glycerol uptake in S. cerevisiae (Holst et al., 2000). However, in later
studies Gup1/2p were proposed to have different roles than glycerol transport (Neves et al.,
2004). A gene involved in active glycerol uptake in S. cerevisiae was identified by screening
for genes encoding membrane proteins involved in glycerol assimilation. It was concluded
that the glycerol proton symporter in S. cerevisiae is encoded by STL1. The protein which is
localized in the plasma membrane, is a member of the sugar transporter family and is glucose

72
repressed (Ferreira et al., 2005). Stl1p is present in the plasma membrane of S. cerevisiae
when the glycerol symport is functional. Stl1p has also been shown to have a function in
glycerol uptake in several other yeasts. It has been shown that in Candida albicans glycerol
was actively transported into the cells by a proton symporter encoded by the C. albicans
STL1 (Kayingo et al., 2009). It was also reported that S. cerevisiae strains harboring the STL1
gene from D. hansenii improved their growth and doubling times slightly (González-
Hernández, 2010).
After glycerol is transported into the cells, two different routes have been identified in yeasts
for further assimilation: phosphorylation route and oxidation route. Within the first pathway,
glycerol is dissimilated by glycerol kinase encoded by GUT1 and then by mitochondrion-
located glycerol-3-phosphate dehydrogenase encoded by GUT2 (Nevoigt & Stahl, 1997).
Another pathway is catalyzed via glycerol dehydrogenase encoded by GCY1 and followed by
dihydroxyacetone kinase encoded by DAK1 and DAK2 to dihydroxyacetone phosphate and
then enter the glycolysis. The pathways involved in glycerol utilization differ in different
yeasts. For glycerol formation, glycerol is commonly produced in the cytosol of yeasts from
the glycolytic intermediate dihydroxyacetone phosphate in two steps that are catalyzed by
glycerol-3- phosphate dehydrogenase (Gpd) and glycerol-3-phosphatase (Gpp), respectively.
Each enzyme activity has two isoenzymes, Gpd1p and Gpd2p, Gpp1p and Gpp2p (Wang et
al., 2001). The production of glycerol has two functions in S. cerevisiae: redox balance and
protection against osmotic stress as a compatible solute (osmolyte).
Although there have been several studies on glycerol metabolism in S. cerevisiae, many
industrially used baker’s yeast strains grow poorly on glycerol. For example, the specific
growth rate of S. cerevisiae CBS 8066 was found to be very low at 0.010 ± 0.002 h-1 during
shake flask cultivations in mineral medium containing glycerol as sole carbon source under
aerobic conditions (Ochoa-Estopier et al., 2011). It was reported that the specific growth rate
of S. cerevisiae BY 4741 was 0.05 h-1 grown on glycerol in a minimal medium (Lu et al.,
2005). Adapted evolution, overexpression or knock out of the genes involved in glycerol
metabolism have been performed to increase the host strain’s growth on glycerol and
production of different value-added products (Ochoa-Estopier et al., 2011; Yu et al., 2010).
However, one possible rate-limiting step during glycerol metabolism, glycerol transport, was
not addressed in those approaches (Burd et al., 2009).

73
In this study, a comparison of the genome sequences of S. cerevisiae and P. tannophilus has
been performed to compare the genes involved in glycerol transport and glycerol metabolism
in these two organisms. Quantitative real-time PCR was performed to compare transcript
levels during growth on glycerol. The genes involved in glycerol transport in P. tannophilus
have been heterologously expressed in a S. cerevisiae (CEN.PK 113-5D) to assess their
function and also the possible contribution of transporter genes to the growth of S. cerevisiae
on glycerol.
2. Materials and methods
2.1 Strains and plasmids
The Pachysolen tannophilus strain used in this study was CBS4044. S. cerevisaie CEN.PK
113-5D was used as a host for heterologous expression of glycerol transporters from P.
tannophilus. The integrative USER vector pXI-5 (Mikkelsen et al., 2012) was used in this
study for constructing the expression vectors in S. cerevisiae. Plasmid pSP-G1 (Partow et al.,
2010) was used as a template for amplifying the bidirectional promoter TEF1/PGK1. All
plasmids were propagated in Escherichia coli strain DH5α. S. cerevisiae strains and plasmids
used in this study were listed in Table1.
Table 1. Strains and plasmids used in this study.
Plasmid or strain Relevant characteristics Source or reference
Plasmids
pSP-G1 2µ-based URA3 plasmid with TEF1/PGK1 promoter Partow et al., 2010
pXI-5 Integrative USER vector, with reusable URA3 Marker Mikkelsen et al., 2012
pXI-5-PtFPS1 pXI-5 with TEF1/PGK1 promoter and PtFPS1 gene This study
pXI-5-PtFPS2 pXI-5 with TEF1/PGK1 promoter and PtFPS2 gene This study
pXI-5-PtSTL1 pXI-5 with TEF1/PGK1 promoter and PtSTL1 gene This study
pXI-5-PtSTL2 pXI-5 with TEF1/PGK1 promoter and PtSTL2 gene This study
S.cerevisiae Strains
CEN.PK 113-5D MATa MAL2-8c SUC2 ura3-52 Peter Kötter
XI-5_XYC1 CEN.PK 113-5D with empty plasmid pXI-5 This study
PtFPS1_XY1 CEN.PK 113-5D with plasmid pXI-5-PtFPS1 This study
PtFPS2_XY2 CEN.PK 113-5D with plasmid pXI-5-PtFPS2 This study
PtSTL1_XY3 CEN.PK 113-5D with plasmid pXI-5-PtSTL1 This study
PtSTL2_XY4 CEN.PK 113-5D with plasmid pXI-5-PtSTL2 This study

74
2.2 Medium and culture conditions
For quantitative real-time PCR (qPCR) experiments, P. tannophilus was grown at 30°C in
defined medium, containing (per liter) 0.67g yeast nitrogen base w/o amino acids, 10g
succinic acid and 6g NaOH as buffer system, 2% (v/v) glycerol or 2% (w/v) glucose as the
carbon source. Cells were harvested at mid-log exponential growth phase at the same optical
density by centrifugation at 5000g, 4°C for 5mins, resuspended in 1ml ice cold ddH2O, and
the cell pellet was stored at -80°C. S. cerevisiae CEN.PK 113-5D transformants harbouring
glycerol transporter genes from P. tannophilus were cultivated at 30°C for 96 hours in
agitated flasks with defined medium, containing (per liter) 7.25g synthetic complete (SC)
powder, 10g succinic acid and 6g NaOH as buffer system, 2% (v/v) glycerol as the sole
carbon source.
2.3 Sequence retrieval and analysis
The genome sequence from P. tannophilus CBS4044 was obtained from previous work and is
available in the GenBank database. The sequences of genes involved in glycerol transport and
metabolism in S.cervisiae FPS1, STL1, GUT1, GUT2, GCY1, Dak1/2, GPD1/2, GPP1/2 were
blasted against the genome sequence of P. tannophilus. The genes with high similarities and
high identities were listed as potential orthologous genes. Gene sequences were registered in
GeneBank at NCBI. The multiple sequence alignments with the amino acid sequences of
FPS1 and STL1 transporters from different yeast strains and E. coli were performed by using
ClustalW2 free program at PDBe. Phylogenetic analyses were performed by using PAUP*
4.0b10 (Swofford, 2003). Unweighted parsimony analysis was performed. Trees were
inferred using the heuristic search option with tree bisection-reconnection branch swapping
and 1000 random sequence additions. Maxtrees were unlimited, branches of zero length were
collapsed and all multiple parsimonious trees were saved. Clade stability was assessed in a
bootstrap analysis with 1000 replicates. Trees were visualized in Treeview (Page, 1996).
2.4 Quantitative real-time PCR
Total RNA isolation and cDNA synthesis
Total RNA was isolated from frozen cells using an RNeasy Mini Kit (QIAGEN, USA)
according to the manufacturer’s protocol. The quantity and quality of the isolated RNA were
measured by NanoDrop ND-1000. The total RNA was treated with DNaseI (Fermentas) prior

75
to cDNA synthesis. Five µg total RNA were used to synthesize cDNA employing the
RevertAidTM First Strand cDNA Synthesis kit (Fermentas) following the manufacturer’s
recommendations using oligo(dT)18 primer. The cDNA was used as the template for
quantitative real-time PCR for determining the transcript levels of the target genes under
different growth conditions.
qPCR was performed on a Stratagene Mx3005P using the SYBR Green technology. The
qPCR reaction mixture was prepared with 5µl of 5 times dilution of cDNA as template, 10µl
SYBR Green master mix, 2µl of 1µM forward primer and 2µl of 1µM reverse primer and
ddH2O to 20µl. The PCR program for qPCR was as follows: 10 min of incubation at 95°C,
followed by 40 cycles of 95°C for 30s, 58 °C for 30 s and 72 °C for 30s, and finally the
temperature was increased from 55°C to 94°C to check for unspecific products. The number
of fluorescence threshold cycles (Ct) was calculated with the set threshold value by using
Mx3005P software. Results presented are mean values of three independent experiments.
Suitable primer pairs for all the genes investigated were designed using the Primer Express
software v3.0 (Applied Biosystems) software with the following parameters: product size 140
- 180bp and melting temperature (Tm) 57-59°C. The primers used in this work are listed in
Table 2. In addition, control reactions which include all components for qPCR except for the
reverse transcriptase were performed to detect the genomic DNA contaminating. The absence
of products under these conditions meant the absence of enough genomic DNA for successful
amplification.
The relative expression levels (RE) were calculated approximately based on 2-ΔΔCT, where
ΔΔCT = (CT gene of interest - CT internal control) sample A - (CT gene of interest - CT internal
control) sample B. CT represents the cycle number at which a sample reaches a predetermined
threshold signal value for the specific target gene. Relative expression data were normalized
to the relative expression value of the housekeeping gene TAF12 in each sample, thus giving
normalized relative expression for a target gene.

76
Table 2. Primers used for Semi-quantitative RT-PCR and USER Cloning.
Primer Name Sequence 5' to 3'
PtFPS1_qF CTGCCGTTTACAGAGATTTT
PtFPS1_qR AATATCTGCACCACCTTCGTA
PtFPS2_qF GGTTTTGCTGTTATGATGGGTT
PtFPS2_qR CAACATAAGCACCAATAAGACC
PtSTL1_qF AATTAGGTTGTATGACTGGTGCATTT
PtSTL1_qR TAGCAACTGCCAATTGTGCAA
PtSTL2_qF TGGTTATGCAACTTCTGTTTGGA
PtSTL2_qR AACCATCTTGGAGTTTCAACAATG
PtGcy1_qF AACTGCCTTACAAAATGGGTACAG
PtGcy1_qR TGCTTTAGCAACATCGTCTTGA
PtGcy2_qF TGGTAACAAGATTCCGGCATT
PtGcy2_qR ATTGCTTTGCCGACATCTTCT
PtDak_qF GTAGTACTTTTGCTTCTCCTTCCACTAA
PtDak_qR AACCTTCTGCTTTTGCTCTTTCA
PtGPD_qF AGGTGTTTCAAAAATGAGTACGGTAT
PtGPD_qR CCGGACCCTATCACTGCAA
PtGPP_qF CAGTGGAAAGAGCTTGGAGTGT
PtGPP_qR ACTCATCGGAAGCAGCATTG
PtGUT1_qF CCAATTTGTGGTTGTTTAGGTG
PtGUT1_qR CACCATGACGAGAGATCAAT
PtGUT2_qF TCAAGAATGAATGTCGCGTT
PtGUT2_qR CATCAGCACCAACCAAATATTC
PtTAF12_qF CAAACCTTCATTACCACTGCC
PtTAF12_qR GTTGCCATCTCCTTCATCG
TEF1_F_U CGTGCGAUTTGTAATTAAAACTTAGATTAGATTGC
PGK1_R_U AGCGTTAGUTTGTTTTATATTTGTTGTAAAAAGTAGA
PtFPS1_F_U ACTAACGCUATGTCAAATTCATCCGGGAAC
PtFPS1_R_U CACGCGAUTTAAGTAGAGTCAACTTCTTTTTCAAG
PtFPS2_F_U ACTAACGCUATGCAAATAGAGAATGTTCAAGGA

77
PtFPS2_R_U CACGCGAUTTAGTATTGATTTTCTACGTCTTTTGTT
PtSTL1_F_U ACTAACGCUATGTTCAAAAAAATCGATAAAATTG
PtSTL1_R_U CACGCGAUTCACTCTTTCTTTTCGGGTT
PtSTL2_F_U ACTAACGCUATGGATTCGAATATAGATGATACAG
PtSTL2_R_U CACGCGAUCTAATGACGACCGCTATTG
2.5 Constructs harboring glycerol transporters from P. tannophilus
In this study, four expression plasmids carrying the glycerol transporter genes from P.
tannophilus were constructed. The USER vector pXI-5 was digested with AsiSI and then the
nicking endonuclease Nb.BsmI for making the AsiSI/Nb.BsmI USER cassettes (Hansen et
al., 2011). The proofreading PfuX7 (Norholm, 2010) was used for amplification of
fragments PtFPS1, PtFPS2, PtSTL1, PtSTL2 and bidirectional promoter TEF1/PGK1 with
appropriate USER tails for insertion into the designated USER cassette AsiSI/Nb.BsmI. PCR
was performed with PfuX7 DNA polymerase according to manufacturer’s instructions. DNA
mixtures were prepared from purified digested vector, glycerol transporter gene, and
bidirectional promoter TEF1/PGK1, 5× Phusion HF buffer (Fermentas), and 1 U of USER
enzyme (New England BioLabs), adjusted to 10µl by adding ddH2O. The mixture was
incubated at 37°C for 20 min, followed by 25°C for 20 min. The 10µl reaction mix was used
directly to transform chemically competent E. coli DH5α cells. All the glycerol transporter
genes were cloned under control of the PGK1 promoter. Constructs were named as: pXI-5-
PtFPS1, pXI-5-PtFPS2, pXI-5-PtSTL1, pXI-5-PtSTL2 for plasmids (Table 1). The plasmids
were verified by sequencing and digested with NotI (Fermentas) for 1h at 37°C and then
subjected to gel purification. Each linearized construct was homologously integrated into the
S. cerevisiae CEN.PK 113-5D genome by transformation (Gietz et al., 1995); five yeast strain
lines were constructed as summarized in Table 1. Transformants were plated onto SC-Ura for
selection. Sequences and descriptions of primers are presented in Table 2.
2.6 Analysis of S. cerevisiae transformants by colony PCR and RT-PCR
The S. cerevisiae transformants were restreaked on SC-Ura plates for single colonies, and the
desired homologous integration was checked by colony PCR. Cells of confirmed integrants
grown in SC minimal medium at 30°C for 96 hours were harvested and used for total RNA
extraction and cDNA synthesis. RT-PCR was used to analyze expression of the transporter

78
genes from P. tannophilus. The program for RT-PCR: 94°C for 2min, followed by 34 cycles:
50°C for 30s, 72°C for 2min, finally 72°C for 5min.
2.7 Analytical methods
Growth was monitored by measuring OD450nm with a Shimadzu UV mini-1240
spectrophotometer (Shimadzu, Japan). Samples were taken periodically from the flasks and
filtered through a 0.22 µm syringe filter, and supernatants were preserved at -20°C for later
HPLC analysis. Concentrations of the substrate glycerol in supernatants were measured by
HPLC refractive index detector RID-10A using an Aminex87H column (Bio-Rad, USA).
Separations were performed at 60°C, flow rate of 0.6ml/min and 5mM H2SO4 as mobile
phase.
3. Results
3.1 Identification of genes in glycerol metabolism and analysis of glycerol transporters
in P. tannophilus
In order to find genes related to glycerol transport and metabolism in P. tannophilus, the
genome sequence was blasted with the sequences of genes known to have these functions in S.
cerevisiae. In P. tannophilus, two glycerol facilitator and two glycerol symporter genes were
found related with glycerol transport and named respectively PtFPS1, PtFPS2, PtSTL1 and
PtSTL2. In the glycerol consumption pathways, the genes with high similarity to S. cerevisae
were found and named PtGUT1, PtGUT2, PtGCY1, PtGCY2, PtDAK (Table 3). In the
glycerol production pathways, the genes found were named PtGPD and PtGPP. It was
noticed that the genes PtFPS2 and PtGUT1 were located close to each other in the P.
tannophilus genome. With the aim of predicting the functions of the putative genes,
bioinformatics tools Blastx and Blastp were used, and functional domain predictions were
applied. The results are presented in Table 3.

79
Table 3. Discovery of genes potentially involved in glycerol metabolism in P. tannophilus by a
BLAST search with known S. cerevisiae genes.
Annotated function Gene Name
S.cerevisiae
Gene Name
P.tannophilus
Size
(bp)
Identity
/Similarity
Accession
no.
Transport
Glycerol Faciliator ScFPS1 PtFPS1 1170 35/49 JQ481631
PtFSP2 972 32/51 JQ481632
Glycerol Symporter ScSTL1 PtSTL1 1728 34/51 JQ481633
PtSTL2 1905 31/50 JQ481634
Consumption
Glycerol kinase ScGUT1 PtGUT1 1848 53/67 JQ481635
G3P dehydrogenase ScGUT2 PtGUT2 1998 52/66 JQ481636
Glycerol dehydrogenase ScGCY1 PtGCY1 936 54/73 JQ481637
PtGCY2 933 51/70 JQ481638
Dihydroxyacetone kinase ScDAK1
PtDAK 1767 42/58 JQ481639
ScDAK2
Production
G3P dehydrogenase ScGPD1 PtGPD 1314 67/80 JQ481640
ScGPD2
G3-phosphatase ScGPP1 PtGPP 705 33/53 JQ481641
ScGPP2
In order to investigate the phylogenetic relationship among putative glycerol transporters in P.
tannophilus and their homologues from other yeast strains, an alignment was performed with
Ptfps1p, Ptfps2p, Ptstl1p ,Ptstl2p as well as other predicted or published transporter proteins
available from GenBank, Génolevures, SGD (Saccharomyces Genome Database), CGD
(Candida Genome Database) databases of other yeasts. Unrooted phylogenetic trees are
presented in Figure 1.
For FPS1, the lengths of PtFps1p, PtFps2p, PpFps1p, YlFps1p and GlpF were relatively short
compared to other Fps1p proteins. PtFps1p was shown to be 54% identical to PpFps1p, 45%

80
to YlFps1p, 35% to ScFps1p and only 31% to GlpF. PtFps2p was shown to be 60% identical
to PpFps1p, 49% to YlFps1p, 34% to GlpF and only 32% to ScFps1p. PtFps1p and PtFps2p
were grouped together with YlFps1p and PpFps1p, while they were in separate branch with
ScFps1p. For STL1, the identity between PtStl1p and ScPtStl1p was only 34%. However, the
identity between PtStl1p and YlPtStl1p was 56%.
Fps1
Stl1

81
Figure 1. Unrooted phylogenetic tree based on alignment of predicted protein sequences. The tree
was bootstrapped 1000 times. Accession numbers are presented after protein names. Ca, Candida
albicans; Dh, Debaryomyces hansenii; Kl, Kluyveromyces lactis; Km, Kluyveromyces marxianus; Pp,
Pichia pastoris; Pt, Pachysolen tannophilus; Sc, Sacchromyces cerevisiae; Yl, Yarrowia lipolytica;
Zr, Zygosaccharomyces rouxii.
3.2 Quantitative real time RT-PCR
qPCR was used to compare the levels of transcripts during growth on glycerol to that on
glucose as the sole carbon source. It was found that the genes involved in glycerol transport
and assimilation PtFPS1, PtFPS2, PtGUT1, PtGUT2, PtGCY1, PtDAK were up-regulated on
glycerol to different degrees compared to on glucose, while the genes involved with glycerol
production PtGPD and PtGPP were down-regulated on glycerol (Figure 2). Among all the
genes PtFPS2 and PtGUT1 were most highly over-expressed and were up-regulated by a
factor of 19.6 ± 2.3 and 17.6 ± 1.3, respectively, on glycerol compared to glucose. The
transcript levels of PtFPS1, another facilitator, was up-regulated by a factor of 5.4 ± 0.9 on
glycerol compared to glucose, while the glycerol symporter PtSTL1, PtSTL2 genes were
expressed almost at the same level on glycerol and glucose. PtGUT2, which is a putative
mitochondrial glycerol phosphate ubiquinone oxidoreductase, was also up-regulated at 4.2 ±
0.04 fold on glycerol.

82
PtFPS1 PtFPS2 PtSTL1 PtSTL2 PtGUT1 PtGUT2 PtGcy1 PtGcy2 PtDak PtGPP PtGPD PtTAF12
0
5
10
15
20
Rela
tive
expr
essio
n le
vels
Figure 2. Relative expression levels (RE) of glycerol metabolism related genes in P. tannophilus on
glycerol compared to that on glucose. RE estimation was based on 2-ΔΔCT, where ΔΔCT= (CT gene of
interest - CT internal control) sample A - (CT gene of interest - CT internal control) sample B. CT
represents the cycle number at which a sample reaches a predetermined threshold signal value for the
specific target gene. All the experiments were performed in triplicate.
3.3 Performance of S. cerevisiae harboring glycerol transporter genes from P.
tannophilus on glycerol
All the predicted transporter genes from P. tannophilus were heterologously expressed under
a strong and constitutive promoter PGK1 in S. cerevisiae. To analyze the performance of S.
cerevisiae recombinant strains, growth in defined medium with 2% (v/v) glycerol as the sole
carbon source of CEN.PK 113-5D strain harboring plasmids pXI-5PtFPS1, pXI-5PtFPS2,
pXI-5PtSTL1 and pXI-5PtSTL2 were tested under aerobic conditions in shake flasks.
CEN.PK 113-5D containing the empty plasmid pXI-5 was grown aerobically in the same
medium as a control.
The recombinant strains expressing the transporter genes PtFPS1, PtFPS2 and PtSTL1 from
P. tannophilus showed improved growth on glycerol to different degrees compared to the
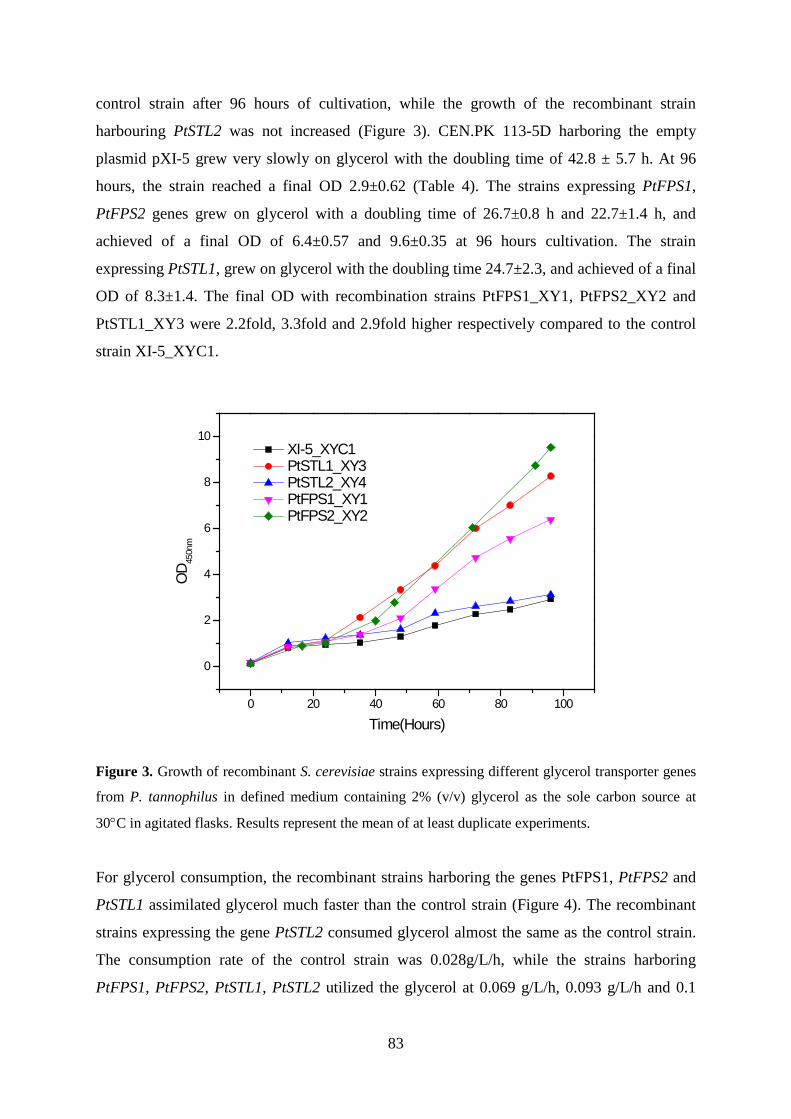
83
control strain after 96 hours of cultivation, while the growth of the recombinant strain
harbouring PtSTL2 was not increased (Figure 3). CEN.PK 113-5D harboring the empty
plasmid pXI-5 grew very slowly on glycerol with the doubling time of 42.8 ± 5.7 h. At 96
hours, the strain reached a final OD 2.9±0.62 (Table 4). The strains expressing PtFPS1,
PtFPS2 genes grew on glycerol with a doubling time of 26.7±0.8 h and 22.7±1.4 h, and
achieved of a final OD of 6.4±0.57 and 9.6±0.35 at 96 hours cultivation. The strain
expressing PtSTL1, grew on glycerol with the doubling time 24.7±2.3, and achieved of a final
OD of 8.3±1.4. The final OD with recombination strains PtFPS1_XY1, PtFPS2_XY2 and
PtSTL1_XY3 were 2.2fold, 3.3fold and 2.9fold higher respectively compared to the control
strain XI-5_XYC1.
0 20 40 60 80 100
0
2
4
6
8
10
OD 45
0nm
Time(Hours)
XI-5_XYC1 PtSTL1_XY3 PtSTL2_XY4 PtFPS1_XY1 PtFPS2_XY2
Figure 3. Growth of recombinant S. cerevisiae strains expressing different glycerol transporter genes
from P. tannophilus in defined medium containing 2% (v/v) glycerol as the sole carbon source at
30°C in agitated flasks. Results represent the mean of at least duplicate experiments.
For glycerol consumption, the recombinant strains harboring the genes PtFPS1, PtFPS2 and
PtSTL1 assimilated glycerol much faster than the control strain (Figure 4). The recombinant
strains expressing the gene PtSTL2 consumed glycerol almost the same as the control strain.
The consumption rate of the control strain was 0.028g/L/h, while the strains harboring
PtFPS1, PtFPS2, PtSTL1, PtSTL2 utilized the glycerol at 0.069 g/L/h, 0.093 g/L/h and 0.1
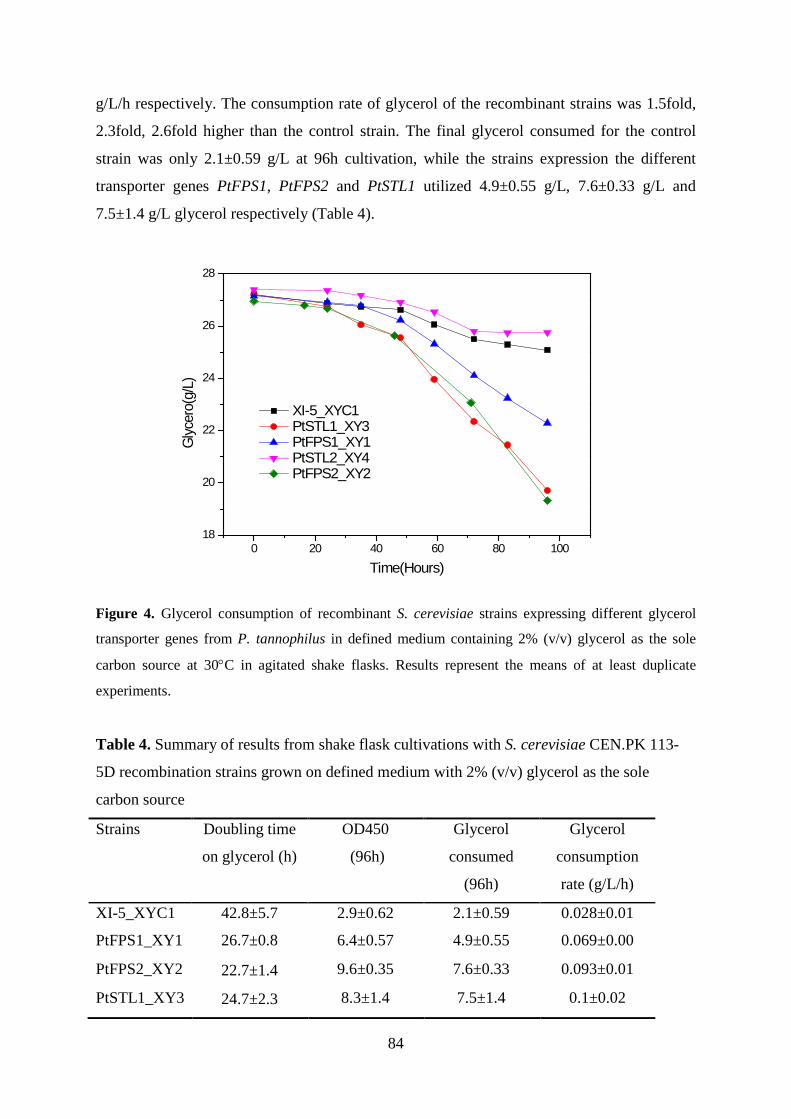
84
g/L/h respectively. The consumption rate of glycerol of the recombinant strains was 1.5fold,
2.3fold, 2.6fold higher than the control strain. The final glycerol consumed for the control
strain was only 2.1±0.59 g/L at 96h cultivation, while the strains expression the different
transporter genes PtFPS1, PtFPS2 and PtSTL1 utilized 4.9±0.55 g/L, 7.6±0.33 g/L and
7.5±1.4 g/L glycerol respectively (Table 4).
0 20 40 60 80 10018
20
22
24
26
28
Glyc
ero(
g/L)
Time(Hours)
XI-5_XYC1 PtSTL1_XY3 PtFPS1_XY1 PtSTL2_XY4 PtFPS2_XY2
Figure 4. Glycerol consumption of recombinant S. cerevisiae strains expressing different glycerol
transporter genes from P. tannophilus in defined medium containing 2% (v/v) glycerol as the sole
carbon source at 30°C in agitated shake flasks. Results represent the means of at least duplicate
experiments.
Table 4. Summary of results from shake flask cultivations with S. cerevisiae CEN.PK 113-
5D recombination strains grown on defined medium with 2% (v/v) glycerol as the sole
carbon source
Strains Doubling time
on glycerol (h)
OD450
(96h)
Glycerol
consumed
(96h)
Glycerol
consumption
rate (g/L/h)
XI-5_XYC1 42.8±5.7 2.9±0.62 2.1±0.59 0.028±0.01
PtFPS1_XY1 26.7±0.8 6.4±0.57 4.9±0.55 0.069±0.00
PtFPS2_XY2 22.7±1.4 9.6±0.35 7.6±0.33 0.093±0.01
PtSTL1_XY3 24.7±2.3 8.3±1.4 7.5±1.4 0.1±0.02

85
3.4 Analysis of S. cerevisiae transformants by PCR and RT-PCR
To validate the function of glycerol transporters from P. tannophilus, four expression vectors
harboring genes PtFPS1, PtFPS2, PtSTL1 and PtSTL2 under control of the PGK1 promoter
were constructed and integrated into S. cerevisiae CEN.PK 113-5D. The engineered strains
carrying the plasmid pXI-5-PtFPS1, pXI-5-PtFPS2, pXI-5-PtSTL1 and pXI-5-PtSTL2 were
confirmed by colony PCR to have undergone correct insertion of the target genes. Expression
of the target genes in the recombinant strains was confirmed by RT-PCR. The colony PCR
and RT-PCR showed that all the transporter genes from P. tannophilus were successfully
inserted and transcribed in the recombination strains of S. cerevisiae CEN.PK 113-5D (Figure
5 a and b).
Figure 5 a. Results for colony PCR performed on the S.cerevisiae recombinant strains. The expected
PCR products are 1170bp for PtFPS1 (lane1), 972bp for PtFPS2 (lane2/3) and 1728bp for PtSTL1
(lane4/5), 1905bp for PtSTL2 (lane6/7). M represents marker.
b. Results for RT-PCR performed on the control and recombinant strains. mRNA was extracted from
the cells grown in defined medium at 30°C for 96 h. PtFPS1(lane3/4); PtFPS2(lane5/6);
PtSTL1(lane1/2); PtSTL2(lane7/8). Lane2/4/6/8 was the results from the control strain XI-5_XYC1.
4. Discussion
With regard to glycerol transport, two glycerol facilitator homologues PtFPS1, PtFPS2 and
two glycerol symporter homologues PtSTL1, PtSTL2 were found in the genome of P.
tannophilus. Based on qPCR results, all the genes involved in glycerol transport and
assimilation were induced on glycerol compared to glucose except the predicted glycerol
symporters PtSTL1 and PtSTL2 which were constitutively expressed on both glycerol and
glucose as carbon source. It was found that the two most strongly up-regulated genes PtFPS2
and PtGUT1 (19.6 fold and 17.6 fold) were located nearby each other in the scaffold of

86
genome sequences, which could fit with the possibility that those two genes might be
functionally related and thus possibly most relevant for glycerol assimilation in P.
tannophilus. Since attempts to investigate the function of those genes by knock-out in P.
tannophilus have failed so far (data not shown), the pathway actually responsible for glycerol
assimilation in P. tannophilus remains uncertain. Therefore, further focus has been placed on
glycerol transport. In this paper the function of the two types of glycerol transporters from P.
tannophilus was addressed by transferring them individually to the well characterized yeast S.
cerevisae.
The low affinity transporter Fps1p is a glycerol facilitator protein and belongs to the major
intrinsic protein (MIP) family of channel proteins with six putative transmembrane domains
(TMDs). Fps1p is responsible for transporting water, small molecules like glycerol, urea,
NH3, CO2 or ions without consuming energy. The physiological role of the facilitator Fps1p
in S. cerevsiaie was described to be glycerol export rather than uptake during hypo-osmotic
shock and the Fps1p channel closed and retained the glycerol inside the cells in response to
hyperosmostic shock (Tamás et al., 1999). An N-terminal domain 225LYQNPQTPTVLP236
and a C-terminal domain 535HESPVNWSLPVY546 were found to have an important role in
controlling Fps1p function. The N-terminus was required for closing of the channels and
restricted transport through Fps1p. It was found that the rate of glycerol efflux was higher
than that for uptake (Tamás et al., 1999). In the present study, the similarity of PtFps1p and
PtFps2p to ScFps1p were 35% and 32% respectively, but the homology was only restricted to
the core of the protein with the six putative TMDs. ScFps1p (669 amino acid residues) was
much longer than PtFps1p (389 residues) and PtFps2p (323 residues), the size of which was
more similar to the E. coli GlpF (281 amino acid). The size difference was mainly due to long
N- and C-terminal domains of ScFps1p. By searching the protein sequences and also the
alignment results, both the N- and the C-terminal hydrophilic extensions in ScFps1p were
missing in the sequences of PtFps1p, PtFps2p and GlpF (Figure 6). By expressing PtFPS2
and PtFPS1 in S. cerevisiae the facilitators from P. tannophilus increased the glycerol influx
and glycerol consumption, presumably due to improved glycerol transport ability. Previously
it was also reported that the glycerol transport was approximately 2.5-fold increased in S.
cerevisiae by introduction of the bacterial gene GlpF (Tamás et al., 1999).

87
Figure 6. Multiple Sequence Alignments with Fps1p from different yeast strains and E.coli by
CLUSTALW2, showing the N-terminal and C-terminal extension domain.
The high-affinity transporter Stl1p was identified as the glycerol proton symporter in S.
cerevisiae (Ferreira et al., 2005), which is a member of the sugar permease family of the
major facilitator superfamily (MFS) (Zhao et al., 1994). It was demonstrated that the
transcription of STL1 gene was significantly induced with glycerol as the sole carbon source,
and the STL1 gene was subject to glucose repression based on microarray-based
transcriptome analysis (Roberts and Hudson, 2006; Roberts and Hudson, 2009). However,
the glycerol uptake by STL1 from C. albicans was not affected by the carbon source and salt
stress (Kayingo et al., 2009). In D. hansenii, it was shown that the active glycerol transport
system was constitutively expressed and not subject to glucose repression (Lucas et al., 1990).
In our study, the glycerol symporters PtSTL1 and PtSTL2 in P. tannophilus also showed
constitutive expression on glycerol and glucose based on qPCR expression analysis. However,
the presence of the PtSTL2 gene had no obvious effect on the physiology of S. cerevisae,
while the glycerol consumption and growth with PtSTL1 were improved. The symporters
PtStl1p showed a low degree of sequence identities to ScStl1p with 34%. However, the
functional PtStl1p from P. tannophilus exhibited 52% identity to the Stl1p from D. hansenii
(DEHA2A12364g) and 56% to the Stl1p from Yarrowia lipolytica (YALI0C16522g).
It has been reported that Y. lipolytica can grow on glycerol with µmax around 0.3h-1
(Papanikolaou & Aggelis, 2002; Papanikolaou et al., 2002). P. tannophilus can grow on
glycerol with µmax around 0.29h-1 (Liu et al.), while S. cervisiae grows relatively slowly on
glycerol with µmax of 0.017h-1 (CEN.PK 113-5D). Both the facilitator and symporter
similarity between P. tannophilus and Y. lipolytica were higher than that compared to S.
cerevisiae. The protein sequence similarity to some degree was relevant to the biological

88
functions. The facilitator and symporter from P. tannophilus might have higher affinity and
efficiency for transporting glycerol through cell membrane into the cells. However, more
protein sequence data with verified functions are needed to provide a definitive conclusion.
Heterologous expression of the glycerol transporters from yeasts growing well on glycerol
was used as the approach for efficient glycerol assimilation in S. cerevisiae in this paper.
Besides, many consider that the glycerol transport might be the rate-limiting step for glycerol
utilization. Improved glycerol transport ability was demonstrated here to increase the glycerol
consumption rate and growth rate under aerobic conditions with S. cerevisae. PtFps2 and
PtStl1 had similar effects on growth and glycerol consumption in the recombinant strains. It
is known that the secondary active transport (glycerol active symporter) consumes more
energy than facilitation. Although, with the exception of PtSTL2, all the putative transporters
from P. tannophilus improved the growth and glycerol consumption to different degrees, the
quantitative contribution of each transporter gene to the growth of P. tannophilus on glycerol
has not yet been ascertained and it may be relevant to investigate this under different growth
conditions and metabolic regulation of the cells. The current study demonstrates the influence
of the glycerol transporter from P. tannophilus on growth and glycerol utilization with S.
cerevisiae. It would be of interest to further study whether there is a synergy of different
transporters. Furthermore, all the experiments were performed under aerobic conditions, and
it could be relevant to investigate the physiological performance under anaerobic conditions.
Our studies open new possibilities for further improving glycerol fermentation in industrial
yeast strains. Heterologous expression of glycerol transporters from the glycerol utilising P.
tannophilus could aid development of glycerol-based bioprocesses in S. cerevisae.
Acknowledgement
This work was funded by the European Community’s 7th Framework Research Programme
under Grant Agreement Number 213506 (Project GLYFINERY) providing financial support
to XL and MW.
References
Burd,G., Bhattacharyya, A. 2009. COMPOSITIONS AND METHODS FOR ENHANCING
GLYCEROL UTILIZATION, U.S. Patent 2009/0176285 A1(Issued Jul.9, 2009).

89
Ferreira, C., van Voorst, F., Martins, A., Neves, L., Oliveira, R., Kielland-Brandt, M.C.,
Lucas, C., Brandt, A. 2005. A Member of the Sugar Transporter Family, Stl1p Is the
Glycerol/H+ Symporter in Saccharomyces cerevisiae. Molecular Biology of the Cell,
16(4), 2068-2076.
Gietz, R.D., Schiestl, R.H., Willems, A.R., Woods, R.A. 1995. Studies on the transformation
of intact yeast cells by the LiAc/SS-DNA/PEG procedure. Yeast, 11(4), 355-360.
Hansen, B.G., Salomonsen, B., Nielsen, M.T., Nielsen, J.B., Hansen, N.B., Nielsen, K.F.,
Regueira, T.B., Nielsen, J., Patil, K.R., Mortensen, U.H. 2011. Versatile Enzyme
Expression and Characterization System for Aspergillus nidulans, with the
Penicillium brevicompactum Polyketide Synthase Gene from the Mycophenolic Acid
Gene Cluster as a Test Case. Applied and Environmental Microbiology, 77(9), 3044-
3051.
Kayingo, G., Martins, A., Andrie, R., Neves, L., Lucas, C., Wong, B. 2009. A permease
encoded by STL1 is required for active glycerol uptake by Candida albicans.
Microbiology, 155(5), 1547-1557.
Kurtzman, C. 1983. Biology and physiology of the. in: Pentoses and Lignin, Vol. 27,
Springer Berlin / Heidelberg, pp. 73-83.
Liu, X., Jensen, P.R., Workman, M. Bioconversion of crude glycerol feedstocks into ethanol
by Pachysolen tannophilus. Bioresource Technology(0).
Lu, L., Roberts, G., Oszust, C., Hudson, A. 2005. The YJR127C gene product is involved in
glycerol-based respiratory growth of the yeast Saccharomyces cerevisiae. Current
Genetics, 48(4), 235-246.
Mikkelsen, M.D., Buron, L.D., Salomonsen, B., Olsen, C.E., Hansen, B.G., Mortensen, U.H.,
Halkier, B.A. 2012. Microbial production of indolylglucosinolate through engineering
of a multi-gene pathway in a versatile yeast expression platform. Metabolic
Engineering, 14(2), 104-111.
Nevoigt, E., Stahl, U. 1997. Osmoregulation and glycerol metabolism in the yeast
Saccharomyces cerevisiae. FEMS Microbiology Reviews, 21(3), 231-241.
Norholm, M. 2010. A mutant Pfu DNA polymerase designed for advanced uracil-excision
DNA engineering. BMC Biotechnology, 10(1), 21.
Ochoa-Estopier, A., Lesage, J., Gorret, N., Guillouet, S.E. 2011. Kinetic analysis of a
Saccharomyces cerevisiae strain adapted for improved growth on glycerol:
Implications for the development of yeast bioprocesses on glycerol. Bioresource
Technology, 102(2), 1521-1527.

90
Partow, S., Siewers, V., Bjørn, S., Nielsen, J., Maury, J. 2010. Characterization of different
promoters for designing a new expression vector in Saccharomyces cerevisiae. Yeast,
27(11), 955-964.
Tamás, M.J., Luyten, K., Sutherland, F.C.W., Hernandez, A., Albertyn, J., Valadi, H., Li, H.,
Prior, B.A., Kilian, S.G., Ramos, J., Gustafsson, L., Thevelein, J.M., Hohmann, S.
1999. Fps1p controls the accumulation and release of the compatible solute glycerol
in yeast osmoregulation. Molecular Microbiology, 31(4), 1087-1104.
Wang, Z., Zhuge, J., Fang, H., Prior, B.A. 2001. Glycerol production by microbial
fermentation: A review. Biotechnology Advances, 19(3), 201-223.
Yu, K.O., Kim, S.W., Han, S.O. 2010. Reduction of glycerol production to improve ethanol
yield in an engineered Saccharomyces cerevisiae using glycerol as a substrate.
Journal of Biotechnology, 150(2), 209-214.
Zhao, S., Douglas, N.W., Heine, M.J.S., Williams, G.M., Winther-Larsen, H.C., Meaden,
P.G. 1994. The STL1 gene of Saccharomyces cerevisiae is predicted to encode a
sugar transporter-like protein. Gene, 146(2), 215-219.

91
Chapter 8 Conclusions and future perspectives
In this PhD project, possible hosts for conversion of the biodiesel by-product glycerol were
selected through screening of the literature and were studied with regard to growth and
external metabolite production on glycerol. Pachysolen tannophilus was selected as the
investigation subject for further study of ethanol production on glycerol.
In chapter 3, the fermentation physiology was investigated as a means to designing a
competitive bioethanol production process by P. tannophilus. It was shown that the oxygen
transfer rate (OTR) was a key factor for ethanol production, with lower OTR having a
positive effect on ethanol production. The effect of pH on ethanol production showed that pH
5 was the optimal pH for ethanol yield and production on glycerol by P. tannophilus. The
deviation from optimal pH affected the ethanol yield to varying degrees. Compared to NO3-
as the nitrogen source, NH4+ stimulated higher carbon flow to ethanol but a lower flow to
biomass. However, NO3- accelerated the rate of ethanol production. The highest ethanol
production with batch fermentation was 17.5 g/L on 5% (v/v) crude glycerol, corresponding
to 56% of the theoretical yield. This study demonstrated that the ethanol production process
from crude glycerol was a robust bioprocess and was not sensitive to the batch variability in
crude glycerol dependent on raw materials used for biodiesel production. A staged batch
process achieved 28.1 g/L ethanol, the maximum achieved so far for conversion of glycerol
to ethanol in a microbial bioprocess.
In chapter 4, adaptive evolution was performed to enhance the ethanol tolerance of P.
tannophilus on glycerol by exposing the cells to a stepwise increase of ethanol concentration
in the medium with repetitive cultivations. The growth performance and the ethanol tolerance
of the mutants were investigated. The ethanol tolerance and maximum specific growth rate
(µmax) of isolated mutant E243, were improved at higher concentrations of ethanol (> 4 % v/v)
compared to the wild type. However, the µmax and the initial glycerol consumption rate of the
mutant when ethanol was not present in the medium were decreased compared to the wild
type strain. The ethanol yield on glycerol and ethanol production levels of E243were also
decreased.

92
A draft genome sequence of the non-conventional yeast P. tannophilus CBS 4044 was
presented in Chapter 5. The total length of the sequenced genome was 12,238,196 bp
(without N) with a GC content of 29.82%. Total 1970.8 Mb raw data was sequenced
representing around 145 fold coverage of the P. tannophilus genome. 5,346 protein-coding
genes (CDSs) set were predicted, with 4,463 (83.5%) genes annotated with function. Based
on PFGE results, the estimated genome size of P. tannophilus (CBS4044) was approximately
13.6 ± 0.4Mb with an estimated of 8 chromosomes.
Chapter 6 was mainly about the attempts to set up the transformation and selection system in
P. tannophilus in order to know more about the genetic background and further improve the
ethanol production process. Antibiotic (Geneticin) resistance marker and uracil auxotrophic
marker were tested for creating the transformation and selection system. Unfortunately, the
strategies and methods employed did not yield successful results.
In chapter 7, based on genome sequencing results the genome of P. tannophilus has been
blasted to find genes similar to FPS1, STL1, GUT1, GUT2, GPD1/2, GPP1/2, GCY1,
DAK1/2 in S. cerevisiae. Eleven genes were found with the predicted function in glycerol
metabolism of P. tannophilus. Quantitative real-time PCR showed that the genes PtFPS1,
PtFPS2, PtGUT1, PtGUT2, PtGCY1, PtDAK involved in glycerol transport and assimilation
were up-regulated on glycerol to different degrees compared to on glucose, while the genes
involved with glycerol production PtGPD and PtGPP were down regulated on glycerol. It
was shown that the two most strongly up-regulated genes PtFPS2 and PtGUT1 (19.6 fold and
17.6 fold) were located close each other in the P. tannophilus genome and might be
functionally related and possibly most relevant for glycerol assimilation in P. tannophilus. A
phylogenetic analysis with the transporter genes revealed that the similarity of PtFps1p and
PtFps2p to ScFps1p were 35% and 32% respectively. The symporters PtStl1p also showed a
low degree of sequence identities to Scstl1p with 34%. The genes involved in glycerol
transport in P. tannophilus have been cloned and successfully expressed in S. cerevisiae
(CEN-PK 1135D) strains. The transformed S. cerevisiae strains with heterologous genes
showed improved growth and glycerol consumption rates with glycerol as the sole carbon
source.
Based on the results of my PhD study, the following suggestions could be recommended for
further developing the glycerol utilization platform and ethanol production process:

93
1. Rational fed-batch fermentation should be designed as growth and ethanol production
is correlated to oxygen transfer rate. This could further increase ethanol yields and
production level.
2. More effort should be focused on mapping the 34 scaffolds from genome sequencing
results into the chromosomes of P. tannophilus.
3. More experiments such as testing different transformation methods would be needed
for setting up a suitable transformation and selection system in P. tannophilus.
4. The physiological performance of the S. cerevisiae transformants under anaerobic
conditions should be investigated.
5. The S. cerevisiae recombination transformant harboring the combination transporters
of PtFPS1 and PtSTL1 from P. tannophilus should be constructed to test if there is
any synergistic effect for growth and glycerol assimilation with S. cerevisiae.
Hopefully this thesis will help clarify the glycerol metabolism in yeast and provide readers
with more understanding in the area of conversion of glycerol by different microorganisms
and particularly P. tannophilus.





























