Embed Size (px)
Citation preview

JOURNAL OF BACTERIOLOGY, Apr. 1978, p. 278-2860021-9193/78/0134-0278$02.00/0Copyright © 1978 American Society for Microbiology
Vol. 134, No. 1
Printed in U.S.A.
Conjugative Plasmids in Neisseria gonorrhoeaeTHOMAS E. SOX,`* WASSILA MOHAMMED,2 ELEANORE BLACKMAN,2 GOUR BISWAS,2 AND P.
FREDERICK SPARLING2
Departments ofMedicine and Bacteriology, School ofMedicine, University of North Carolina, Chapel Hill,North Carolina 27514
Received for publication 17 October 1977
A conjugation system initially discovered in fl-lactamase-producing gonococcimobilized small non-selftransmissible R plasmids encoding ,8-lactamase (penicil-linase) production into other gonococci, Neisseria, and Escherichia coli. Thisconjugation system was mediated by a separate selftransmissible plasmid of 23.9x 106 daltons, pFA2. Conjugative plasmids capable of mobilizing R plasmids werealso found in nearly 8% of the non-penicillinase-producing gonococci. These weresimilar to pFA2 in size, buoyant density, and restriction endonuclease digestpatterns but were less efficient than pFA2 in mobilization of the penicillinaseplasmid pFA3. The presence of conjugative plasmids in gonococci isolated beforethe appearance of penicillinase-producing strains indicates that a conjugationsystem for plasmid transfer predated the appearance of R plasmids in gonococci.
Recently, strains of Neisseria gonorrhoeaewith plasmid-mediated ,f-lactamase (penicillin-ase) production have been isolated in severalnations (6, 26). Although plasmid-mediated an-tibiotic resistance is common in many othergram-negative bacteria (3), antibiotic resistancein gonococci has been thought to be a result ofchromosomal loci (7). Chromosomal mutationsresult in only low-level resistance to penicillin(31). Acquisition of plasmid-mediated fi-lacta-mase production by a penicillin-sensitive strainof gonococcus, however, results in an approxi-mately 100- to 1,000-fold increase in resistanceto penicillin, depending on inoculum size (25).
,B-Lactamase-producing gonococcal clinicalisolates contain plasmids of about 4.7 or 3.4million daltons (Mdal) in addition to the 2.6-Mdal cryptic plasmids found in most gonococci(12, 28). The 4.7- and 3.4-Mdal plasmids areresponsible for ,B-lactamase production, as evi-denced by loss of these plasmids in sponta-neously cured (,8-lactamase-negative) deriva-tives, as well as by acquisition of these plasmidsin f-lactamase-positive transformants and trans-conjugants (12, 28). Transfer of the penicillinaseplasmids by conjugation to other gonococci,Neisseria flava, and Escherichia coli has beenreported (12, 15, 28). Since all gonococcal con-jugative donors of the f)-lactamase plasmids alsopossess plasmids of 24 Mdal, it has been postu-lated that conjugation might be mediated by the24-Mdal plasmids (12, 28).
In this communication, we show that the 24-Mdal plasmids are selftransmissible and are in-deed responsible for transfer of the R plasmids.
Similar conjugative plasmids were found in pen-icillin-sensitive strains isolated well before theappearance of penicillinase-producing gono-cocci.
MATERIALS AND METHODSStrains and materials. Table 1 presents a list of
relevant strains and their genotypes. Strain and plas-mid nomenclatures generally adhere to the proposalsof Novick et al. (23). Media and growth conditionswere as described previously (31). The presence of f-lactamase production was confirmed by a chromogeniccephalosporin indicator (Glaxo) (24) or by a phenolred-penicillin G indicator solution (30). Most antibiot-ics were obtained from sources previously described(31). Ampicillin was from Bristol Laboratories, andnalidixic acid was from Sterling-Winthrop.
Conjugation procedure. Conjugation was per-formed by the following two methods. (i) Donor andrecipient cells were cultivated together in GCB brothfor 2 to 16 h at 370C, washed in medium A of Davisand Mingioli (10), and spread on agar media containingappropriate antibiotics to select for transconjugants.(ii) Donor and recipient cells from plates inoculated16 to 20 h previously were suspended in GCB brothmedium to approximately 5 x 108 colony-forming unitsper ml. Portions (0.1 ml) of donor and recipient sus-pensions were placed on a sterile membrane filterresting on GCB agar (Difco Laboratories) that con-tained Kellogg supplements I and II. In some experi-ments, 0.05 ml of a solution containing 1 mg of pan-creatic deoxyribonuclease (Worthington BiochemicalsCorp.) per ml was added to the filter to inhibit trans-formation. Filters were incubated at 370C in the pres-ence of 3% C02 for 1 to 16 h, depending on theexperiment. After this incubation, filters were agitatedin 1.0 ml of medium A to suspend the cells. The cellswere pelleted by brief centrifugation and resuspended,
278
on May 12, 2021 by guest
http://jb.asm.org/
Dow
nloaded from

CONJUGATIVE PLASMIDS IN N. GONORRHOEAE
Strain and plasmidsaN. gonorrhoeaeFA192FA288(pFA2)(pFA3)FA293(pFA2)(pFA3-)FA305FA333(pFA3)FA453(pFA2)(pFA3)FA529(pFAl1)(pFA3)FA532(pFA12)(pFA3)FA539(pFA13)(pFA3)FA583FA589(pFA2)(pFA3)FA651(pFA11)FA652(pFA12)FA653(pFA13)
N. flavaFA530
E. coliC600.5
TABLE 1. Bacterial strains usedSource
Transformant of FA19 (31)C. Thornsberry (12)Spontaneous Pcf cure of FA288 (12)Construction from F62 (8, 12)Transformant of FA19 (12)Construction from FA305Construction from FA305Construction from FA305Construction from FA305Construction from F62Construction from F62Local isolate (1974)W. McCormackLocal isolate (1971)
Spontaneous mutant of ATCC 14221
C. A. Hutchison
Distinguishing characteristics
spc-3,B-Lactamase-producing (Pc'), conjugative donorLike FA288 but Pc"nal-) rif-1 str-7 fus-I met-I pro-IPcrLike FA305 but Pcr, conjugative donorLike FA305 but Pc', conjugative donorLike FA305 but Pcr, conjugative donorLike FA305 but Pcr, conjugative donornal-I arg-i met-I pro-irif-I str-7 spc-3 leu-i Pcr, conjugative donorConjugative donorConjugative donorConjugative donor
nal-2 rif-4
Res- Mod-
a All gonococcal strains in this table contained cryptic plasmids of approximately 2.6 Mdal. pFA2 and pFA3 had molecularweights of 23.9 x 10 and 4.7 x 106, respectively, as determined by electron microscopy. pFA11, pFA12, and pFA13 migrated on
agarose gels at the same rate as pFA2 and had similar EcoRI digest patterns. Their molecular weights were therefore estimatedto be 24 x 106. pFA2 is probably identical to the recently described pLE2450 (27); pFA3 is probably identical to pMR0360.
and portions were spread on selective media. RecipientNeisseria strains contained markers for nalidixic acid(Nal), fusidic acid (Fus), rifampin (Rif), or strepto-mycin (Str) resistance to allow selection against thedonor cells. Media for selection of penicillinase-pro-ducing (Pcr) gonococcal transconjugants contained 0.1to 0.5 ug of penicillin G per ml in addition to one or
more of the above antibiotics. E. coli recipients con-
tained one of the same chromosomal antibiotic resist-ance markers, and the MacConkey agar (Difco) usedfor selection of pCr E. coli transconjugants contained50 ug of ampicillin per ml in addition to antibiotics towhich the donor was sensitive. Background growth ofrecipient gonococci on original plates often necessi-tated replica plating for transconjugant colonies to beisolated. The filter conjugation method generallyyielded higher conjugation frequencies than did con-
jugation in broth culture. In the filter conjugationtechnique, the number of transconjugants obtainedvaried widely with the brand and lot of filters used.Gelman Metricel GA6 filters or Amicon Microporousfilters produced the highest conjugation frequencies.
Resistance mobilization test. Strains lacking re-sistance (R) plasmids were scored for the presence ofconjugative plasmids by a procedure similar to thatdescribed by Anderson (2). A strain lacking an Rplasmid served as the potential initial donor of theconjugative plasmid. FA333, a gonococcus that con-
tained a Pcr R plasmid but lacked a conjugative plas-mid, was used as an intermediate. An Nal' derivativeof E. coli C600.5 served as the final recipient. Transferof the R plasmid into E. coli could only occur if thetested strain introduced a conjugative plasmid intostrain FA333, thereby mobilizing its R plasmid fortransfer. A 0.1-ml amount of a suspension containingca. 5 x 10' colony-forming units of the strain beingtested for conjugative ability per ml was added to an
equal volume of strain FA333 (also ca. 5 x 101 colony-forming units per ml) on a membrane filter. After
several hours of incubation, 0.1 ml of a suspension ofNalr E. coli C600.5 was added, and the conjugationwas continued overnight. The cells were then resus-
pended and plated on media to select for pCr E. colitransconjugants. As a control, FA333 was usually in-cubated with Nalr E. coli C600.5 under the sameconditions.
Strain construction. A set of four homogenic de-rivatives of strain FA305 (Table 1) containing differentconjugative plasmids was constructed. FA288 was
conjugated directly with FA305 to yieldFA453(pFA2)(pFA3). The derivatives of strainsFA651, FA652, and FA653 were obtained by usingthese clinical isolates as conjugative donors andFA333(pFA3) as an intermediate in the resistancemobilization test, and scoring FA305 pcr transconju-gants for conjugative ability. This yieldedFA529(pFA11)(pFA3) from strain FA651 as the donor,FA532(pFA12)(pFA3) from strain FA652 as the donor,and FA539(pFA13)(pFA3) from strain FA653 as thedonor.
Since most gonococcal strains contain cryptic plas-mids of around 2.6 Mdal (7, 18, 33), purification ofconjugative plasmid DNA from the original hoststrains was difficult. This problem was circumventedby mating the homogenic derivatives of FA305(FA453, FA529, FA532) with N. flava (ATCC 14221),which contains no plasmids. pcr transconjugants wereselected and scored for their ability to mobilize pFA3to E. coli C600.5. Pc8 (pFA3-) derivatives of theseconjugative donors were then isolated and used toprepare conjugative plasmid DNA for restriction en-
zyme studies. We were unable to obtain N. flavatransconjugants when FA539(pFA13)(pFA3) was usedas a donor.
Multiply marked donor and recipient strains wereused in testing for conjugative transfer of chromo-somal markers. Strain F62, a proline auxotroph, was
mutagenized with ethyl methane sulfonate to yield a
VOL. 134, 1978 279
on May 12, 2021 by guest
http://jb.asm.org/
Dow
nloaded from

280 SOX ET AL.
methionine auxotroph (met-i) (8). The markers arg-1
and nal-l were added to this derivative by transfor-mation, yielding a strain (FA583) that was used as a
recipient. The donor strain FA589 was a leucine aux-
otroph of F62 obtained by ethyl methane sulfonatemutagenesis to which the markers rif-2, str-7, spc-3,and proline prototrophy were added by transformationand the plasmids pFA2 and pFA3 were introduced byconjugation.
Screening for plasmid content. A loopful of cells
(ca. 20 mg) scraped from an overnight plate was sus-
pended in 200 p1 of TES buffer [50 mM NaCl, 5 mMethylenediaminetetraacetate, 30 mM tris(hydroxy-methyl)aminomethane (pH 8.0)]. Ten microliters of a10-mg/ml lysozyme solution was added, and the sus-
pension was incubated at 37°C for 30 min. A 25-,l1amount of a 10% sodium dodecyl sulfate solution was
then added, and the mixture was shaken gently to lysethe cells. Five microliters of a 10-mg/ml ribonucleaseA (Sigma Chemical Co.) solution that had been heatedto 80°C for 20 min was added, and the mixture was
incubated at 37°C for 1 h. Ten microliters of a 1-mg/ml Pronase (Calbiochem) solution was thenadded, and the mixture was incubated at 37°C for an
additional 2 h. Seventy microliters of 5 M NaCl was
added, and the lysate was kept at 4°C overnight. Aftercentrifugation at 48,000 x g for 20 min, the superna-tant was decanted, and 5- to 25-,lI portions were sub-jected to electrophoresis on 0.8% agarose gels, using a
vertical slab gel apparatus (E-C Apparatus Corp.).Electrophoresis was performed by the procedure ofMeyers et al. (19). Gels were stained with a 1-,ug/mlethidium bromide solution and viewed with a long-wave UV transilluminator. Photographs were madethrough a Vivatar orange filter with Polaroid type 665film. The identity of plasmid bands was determinedby electron microscopy of DNA extracted from ethid-ium bromide-stained bands by the freeze-squeeze tech-nique (35).
Density gradient centrifugation. Cesium chlo-ride-ethidium bromide buoyant density gradients wereprepared as described previously (7). Neutral cesiumchloride gradients for buoyant density determinations(14) contained 14C-labeled plasmid DNA and 32P-la-beled E. coli chromosomal DNA. E. coli chromosomalDNA was assumed to have a specific gravity of 1.7035(11).
Electron microscopy. DNA was prepared for elec-tron microscopy by the Kleinschmidt technique (16)and viewed with an AE1 transmission electron micro-scope. Open circular simian virus 40 DNA was used as
an internal standard of 3.2 Mdal (36).Restriction endonuclease digestion. Restriction
endonucleases (EcoRI, BamHI, and HpaII) were ob-tained from Miles Laboratories, and digestions were
performed essentially by the supplier's instructions.
RESULTS
Identification of the 24-Mdal plasmid asa conjugative plasmid. Each of six tested clin-ical isolates functioning as conjugative donors ofthe ,B-lactamase plasmids contained plasmids ofabout 24 Mdal. This association suggested butdid not prove that these plasmids were conju-
gative. Proof was obtained in conjugation exper-
iments that used pcr strain FA288(pFA2) (pFA3)as a conjugal donor and FA305 (Pcs) as a recip-ient. Sixty-four pcr transconjugants were iso-lated and examined for their plasmid content byagarose gel electrophoresis and for their abilityto donate the pcr plasmid pFA3 to E. coli C600.5(Table 2). Six were found to have received pFA2from strain FA288; only these six clones were
able to mobilize pFA3 for transfer to E. coliC600.5 or to other gonococci. Thus, pFA2 was
responsible for conjugal transfer of pFA3 andcould mediate its own transfer between gono-cocci.
However, when pCr E. coli transconjugantsfrom a conjugation of C600.5 with FA288 were
examined for plasmid content, none of 24 was
found to contain pFA2. Also, resistance mobili-zation tests utilizing strain FA293 as a source ofpFA2, a pFA3-containing derivative of strainC600.5 as an R plasmid intermediate, and anStrr Nalr derivative of strain C600.5 as a finalrecipient failed to yield pcr transconjugants.Therefore, although pFA2 was able to mobilizepFA3 for transfer to C600.5, pFA2 itself waseither not transferred, could not replicate, or
was highly unstable in this strain of E. coli.R plasmid transfer was not due to stable
TABLE 2. Time dependence ofplasmid transfer
traqnsferyof pFA3 transcon- Frequency ofTime (h) tFA3n (x Jugants receiv- unselected re-
10-6) ing pFA2b ceipt of pFA2'
1 8.5 13/24 (54) 0/64 (<2)16 5.0 21/24 (88) 6/64 (9)d
a FA288(pFA2)(pFA3) (rift nal+) served as the pCrdonor, and FA305 (rif-1 nal-1) served as the Pc' recip-ient. Conjugation was performed on a membrane filter(type HA; Millipore Corp.) as described in the text;transfer frequencies in this experiment were unusuallylow due to the type of filter used. pcr transconjugantcolonies were selected on media containing 2.0,ug ofrifampin per ml, 2.0,ug of nalidixic acid per ml, and 0.2,ug of penicillin per ml. Frequencies are expressed perinput donor colony-forming units.
b Pcr transconjugants were scored for receipt ofpFA2 by agarose gel electrophoresis and by the abilityto mobilize pFA3 to suitable recipients. Numbers inparentheses are percentages.
'Recipient FA305 colonies were selected on mediacontaining 2.0 Mg of rifampin per ml and 2.0 ug ofnalidixic acid per ml. Receipt of pFA2 was determinedby the ability to function as a conjugative donor in aresistance mobilization test and by agarose gel electro-phoresis. Frequencies are expressed per recipient col-ony-forming unit. Numbers in parentheses are per-centages.
d None of the six clones receiving pFA2 also receivedpFA3.
J. BACTERIOL.
on May 12, 2021 by guest
http://jb.asm.org/
Dow
nloaded from

CONJUGATIVE PLASMIDS IN N. GONORRHOEAE
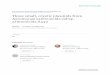
cointegrate formation. By analogy with thebehavior of plasmids from Proteus mirabilis(22) and other organisms (4), it was possible thatthe pcr plasmid (pFA3) formed a cointegratewith the conjugative plasmid (pFA2) in strainFA288. If this were the case, the conjugativeplasmid from the pcr strain would be larger, byabout 4.7 Mdal (the size of pFA3), than theconjugative plasmid from the Pcs derivativeFA293. Molecular weights of the conjugativeplasmids from strains FA288 and FA293 weredetermined from electron micrographs (Fig. 1).pFA2 molecules derived from FA288 were ap-proximately 23.9 Mdal (n, 19; standard devia-tion, 2.0); no measured molecule was as large asthe expected size of a cointegrate (28.6 Mdal).pFA2 molecules from FA293 had a similar size,23.8 Mdal (n, 13; standard deviation, 2.0). Thus,cointegrates of pFA2 and pFA3 do not appear tobe common in FA288. The possibility remainedthat a rare cointegrate was responsible for trans-
fer of the pFA3 genome during conjugation,although all transconjugants examined pos-sessed separate pFA2 and pFA3 plasmids. Thefact that many transconjugants received pFA3without receiving pFA2 (Table 2) confirmed thatpFA3 was transferred without stable linkage topFA2. The existence of transient (unstable) coin-tegrates cannot be excluded without further ex-periments.Conjugative plasmids in penicillin-sensi-
tive gonococci. The presence of a conjugativeplasmid in some ,f-lactamase-producing isolatesand previous reports of a 24.5-Mdal plasmid insome penicillin-sensitive strains of gonococci (18,33) indicated that conjugative plasmids mightbe present in non-,B-lactamase-producing gono-cocci. Therefore, a number of isolates from dif-ferent years and locales were examined for theirability to function as conjugative donors in theresistance mobilization test. Twelve of 156strains examined gave clearly positive results as
A-4~4FI _1 Plsmd oFA8. A pasmid ban from an ethdu broid ceiu chord gradient wasE J|8
preparedfo electron microscopy as described in the tt The largepld in te cnter is an on cr-
moleeop Oe cu pA is at t l
pFA3andpFAJ are also visible in the main figu BNLV6i*}.we;si- S
X Sss aB<k,> , ><,<s > t)sv*s*f.~~~~~~~~~5
iPt-; o ,' + tr.,>so,,%'/t.0 L., v
11!~~~~~~~ k.> *U;s < ;>i *; w;<$
Zla: 0§k''0'0' 'C''''s-" ' . ''' " .'>','~~~J
molecule MofPF2 Ope ciclap3 A isatl the lef sidle oftheistrliat thr ih sid of th iet
FIG.andPAlaremidsofvsilin88 thplasidbndfigre. Bar rpeenthidiu broidecsunclrd.raina
281VOL. 134, 1978
on May 12, 2021 by guest
http://jb.asm.org/
Dow
nloaded from

282 SOX ET AL.
conjugative donors. These were analyzed forplasmid content and all were found to containplasmids of the same mobility on agarose gels asthat of pFA2. Seventeen clinical isolates thatfailed to act as conjugative donors were screenedfor plasmid content, and none was found tocontain a similar-sized plasmid. Three strainswith conjugative plasmids were selected for fur-ther study.Each of these conjugative plasmids, as well as
pFA2, was introduced into strain FA305 (Mate-rials and Methods), yielding a set of four homo-genic strains. Mobilization of pFA3 to othergonococci and E. coli C600.5 by this set wasexamined (Table 3). The plasmid derived fromthe naturally occurring pcr strain FA288, pFA2,consistently mobilized pFA3 to other gonococciand C600.5 at a higher frequency than did theother three plasmids investigated. Curiously,when the conjugative plasmids pFA12 andpFA13 were introduced into FA305, they wereunable to mobilize pFA3 to either E. coli or togonococcal recipients. The original clinical iso-lates containing pFA12 and pFA13 were reliable(but low-frequency) donors in the resistance mo-bilization test.The low frequencies of mobilization of the pcr
plasmid pFA3 by the other conjugative plasmidscould be due to inherently lower frequencies ofconjugation by these plasmids than by pFA2.Alternatively, all conjugative plasmids might beequally effective in establishing conjugation (asdetected by selftransfer), but mobilization ofpFA3 might occur more readily with pFA2 thanwith the other conjugative plasmids. To distin-guish between these possibilities, overnight con-jugations were performed by using the respec-tive FA305 derivative as the donor and FA192,an Spcr transformant of FA19, as the recipient.Receipt of pFA2 occurred at a significantlyhigher frequency (23%) than did receipt ofpFA13 (2%) (Table 3); pFA2 also mobilized
pFA3 much more readily than did pFA13. Re-ceipt of pFA11 by FA192 was as frequent asreceipt of pFA2, but pFAll mobilized pFA3 10-to 100-fold less frequently than did pFA2 (Table3). Thus, the differences in selftransfer did notfully account for the large differences in mobili-zation of pFA3; the conjugative plasmids appar-ently also differed in their ability to initiatetransfer of pFA3.Determination of buoyant density. The
buoyant density of pFA2 (Fig. 2) was 1.701g/cm3, corresponding to a guanine-plus-cytosinecontent of 48% (34). Chromosomal DNA of N.gonorrhoeae has been reported (11) to have thesame buoyant density as E. coli chromosomalDNA; this corresponds to a guanine-plus-cyto-sine content of 50.4%. The lower density ofpFA2compared with that of gonococcal chromosomalDNA was confirmed by the following experi-ment. Whole-cell DNA was prepared by themethod of Marmur (17) and centrifuged in neu-tral CsCl density gradients. One-drop fractionsfrom the region of pertinent density were col-lected, and portions of these fractions were sub-jected to electrophoresis on agarose gels to de-termine the location of chromosomal and plas-mid DNA in the gradient. The conjugative plas-mids pFA2, pFAll, pFA12, and pFA13 were allfound to band in similar regions that wereslightly less dense than the region of chromo-somal DNA.
Restriction endonuclease analysis. Thepurified conjugative plasmids pFA2, pFA1l, andpFA12 were digested separately with each of therestriction endonucleases EcoRI, BamHI, andHpaII, and the digestion products were sub-jected to electrophoresis on agarose gels. Allthree plasmids were cleaved by EcoRI into fourfragments, and the respective fragments fromeach plasmid had the same mobility (Fig. 3,tracks A to C). The BamHI digest of pFA12(track D in Fig. 3) differed from the BamHI
TABLE 3. Differences in function of conjugative plasmids in homogenic strains
Donor strain" Frequency of unselected trans- Frequency of mobilization of pFA3
fer of conjugative plasmid5 FA192 as recipiente C600.5 as recipient'
FA453(pFA2)(pFA3) 11/48 (23) 3.8 x 10-4 1.5 x 10-3FA529(pFA11)(pFA3) 5/16 (31) 3.7 x 10-5 1.3 x 10-5FA532(pFA12)(pFA3) 0/16 (<6) <1.0 x 10-7 <1.9 X 10-7FA539(pFA13)(pFA3) 1/48 (2) <3.2 x 10-7 <1.2 x 10-7
a These strains are homogenic derivatives of FA305, constructed as described in the text.b Overnight conjugations of homogenic Spc" FA305 derivatives as donors and Spcr FA192 as the recipient
were performed as described in the text. Recipient colonies were selected by plating dilutions of resuspendedcells on media containing 200,tg of spectinomycin per ml. The presence of a conjugative plasmid was determinedby agarose gel electrophoresis. Frequencies expressed per recipient colony-forming unit. Numbers in parenthesesare percentages.
c Mean of three experiments. Frequencies are expressed per input donor colony-forming unit.I Mean of two experiments.
,J. BACTERIOL.
on May 12, 2021 by guest
http://jb.asm.org/
Dow
nloaded from

CONJUGATIVE PLASMIDS IN N. GONORRHOEAE
1.75
a-1.70
a 1650
ILcia-C,)
2500
2000 f
(n1500
z0
1000 9z"I0z
500 8u
60 70 80 90 100 110 120 130 14085.2 89.3
FRACTION NUMBERFIG. 2. Neutral cesium chloride density gradient of ["'Cladenine-labeled pFA2 DNA and "P-labeled E.
coli C600.5 DNA. Solid line, "'C counts; broken line, 32p counts. One-drop fractions were collected from the topof the gradient. Points at 85.2 and 89.3 mark the calculated means of the 'IC and 32ppeaks, respectively.
digests of the other two plasmids (tracks E andF). The pFA12 digest lacked the fifth and sev-enth bands from the top found in the other twodigests; instead there was a band migrating be-tween the fourth and fifth bands of the othertwo BamHI digests. In partial BamHI digests ofpFA2, a band was seen at a similar position; thisdisappeared upon further digestion (data notshown). Addition of excess amounts of enzymedid not result in the disappearance of this bandin the pFA12 digest. Apparently this fragmentlacked a BamHI cleavage site present in pFA2and pFA11. The HpaII digests of pFA2, pFA11,and pFA12 were identical for all of 17 resolvablebands (Fig. 4).We were unable to obtain purified pFA13, the
conjugative plasmid of strain FA652 (Materialsand Methods). However, bands similar to thosein tracks A to C of Fig. 3 were present in anEcoRI digest of the plasmids of strain FA539,which contained pFA13 obtained from strainFA652. These bands were not present in a digestof plasmids from the homogenic strain FA305,which lacked pFA13 (data not shown).Failure to detect chromosomal transfer.
The mobilization of the non-selftransmissibleplasmid pFA3 by pFA2 suggested that pFA2might also mobilize the chromosome. Experi-ments to detect such mobilization were per-formed using multiply marked conjugative do-nor (FA589) and recipient (FA583) strains thatwere not competent for genetic transformation(Materials and Methods). There was no differ-ence in the number of apparent Arg+, Met', Strr,or Spcr recombinants when the recipient wasmated overnight on membrane filters with donorcells compared with overnight incubation of the
recipient with DNA isolated from the donor(data not shown). In both cases, recombinantsfor individual donor markers occurred at a fre-quency of less than lo-. Recombinants fromboth parts of this experiment were examined forthe acquisition of linked markers. Both groupsshowed similar low linkage frequencies (data notshown). Other experiments, not presented here,showed that low-frequency transformation oc-curred during prolonged matings despite initialaddition of deoxyribonuclease; only by addingdeoxyribonuclease hourly could transformationbe prevented.
DISCUSSIONAbout 8% of the non-f3-lactamase-producing
clinical isolates ofN. gonorrhoeae that we testedcontained conjugative plasmids. The actual oc-currence of such plasmids may be greater than8%, since the resistance mobilization test wouldnot detect conjugative plasmids unable to mo-bilize the R plasmid used (pFA3). Conjugativeplasmids are known to be quite common in othergram-negative bacteria; Falkow reported that asmany as 40% of E. coli isolates possessed con-jugative ability (13). Previous attempts to dem-onstrate conjugation in gonococci (32) were un-successful partly because of the lack of an Rplasmid whose transfer could be readily de-tected.Our failure to detect mobilization of chromo-
somal markers is not surprising. Transfer ofchromosomal markers by most conjugative plas-mids is apparently quite rare (13). Also, thepaucity of genetic markers presently known ingonococci allowed us to search for transfer ofonly a few chromosomal regions. The isolation
283VOL. 134, 1978
on May 12, 2021 by guest
http://jb.asm.org/
Dow
nloaded from

A B C D E F G pFA2 and pFA13 in overnight matings. Thus,plasmids that appear to be very similar by re-striction endonuclease digest patterns can bequite different functionally. A number of expla-nations could account for these differences. Theconjugative plasmids may differ in the amountof repression exerted on sex pilus formation orthey may vary in the frequency with whichplasmid DNA is activated for transfer. Thesedifferences could also be due to differences inthe copy numbers of these plasmids; however,we did not notice any marked difference in theintensity of these plasmid bands on agarose gels.
FIG. 3. Agarose gel (0.8%) of EcoRI and BamHIdigests of conjugal plasmids. Tracks A to C, EcoRIdigests of pFAII, pFA12, and pFA2, respectively;tracks D to F, BamHI digests ofpFAll, pFA12, andpFA2, respectively; track G, EcoRI digest ofphagelambda DNA. Arrow on left indicates position of afaint band present in tracks A, B, and C; arrow onright indicates position of a faint band in E and F.
of derepressed transfer mutants of the conjuga-tive plasmid (20) or the use of the technique ofintegrative suppression (21) might result in adetectable mobilization of chromosomalmarkers, which would greatly facilitate futuregenetic studies of gonococci.The four conjugative plasmids that we exam-
ined were similar in size, buoyant density, andrestriction endonuclease digest patterns, indicat-ing a strong degree of nucleotide homology. Wefound similar-sized conjugative plasmids in localstrains isolated as long ago as 1971; also, theseplasmids were found in a group of strains col-lected in Boston, Mass., in 1975. Whatever thesource of the genetic material of the fl-lactamaseplasmids, a conjugative system capable of pro-moting the transfer of these R plasmids wasalready present in the gonococcus.The data of Table 3 show that the conjugative
plasmids differed widely in their ability to mo-bilize the Pcr R plasmid pFA3 from a homogenicdonor. Also, there was a significant difference inthe selftransfer of the conjugative plasmids
FIG. 4. Agarose gel (2%) of HpaII digests of con-jugal plasmids. First track, pFA1i1 second track,pFA12 third track, pFA2.
.i
284 SOX ET AL. J. BACTERIOL.
on May 12, 2021 by guest
http://jb.asm.org/
Dow
nloaded from

CONJUGATIVE PLASMIDS IN N. GONORRHOEAE
Although the mobilization of the Pcr plasmidpFA3 was often accompanied by transfer of theconjugative plasmid, we could find no evidenceof stable cointegrate formation. Rather, the re-sults suggested that these plasmids were trans-ferred separately, as in the class 2 transfer sys-tem described by Anderson and Natkin (5).The 24-Mdal pasnids described in this paper
are among the smallest conjugative plasmidsthat have been reported (29). However, a con-jugative plasmid of 17.3 Mdal has been described(9), and the portion of the E. coli F plasmidencoding transfer functions may occupy onlyabout 15 Mdal (1). Thus, these gonococcal plas-mids may be large enough to encode geneticinformation unrelated to transfer functions. Wefound that these plasmids did not confer entryexclusion (37) upon the host cells, since recipientstrains differing only in the presence or absenceof the conjugative plasmid served equally wellas recipients ofthe pcr plasmid (data not shown).The natural selection that maintained theseplasmids in gonococcal strains before the ap-pearance of R plasmids remains obscure. How-ever, these conjugative plasmids now provide a
means oftransfer of antibiotic resistance in vitro,.and this transfer may occur in vivo as well.
ACKNOWLEDGMENTS
We thank W. M. McCormack, Marie Roberts, C. Thoms-berry, and E. Stolz for donating strains.
This work was supported by Public Health Service grantAI10646 from the National Institute of Allergy and InfectiousDiseases and by a grant from The John A. Hartford Founda-tion, Inc.
LITERATURE CITED
1. Achtman, M., and R. Helmuth. 1975. The F factorcaries an operon of more than 15 x 10i daltons codingfor deoxyribonucleic acid transfer and surface exclusion,p. 95-103. In D. Schlessinger (ed.), Microbiology-1974.American Society for Microbiology, Washington, D.C.
2. Anderson, E. S. 1965. A rapid screening test for transferfactors in drug-sensitive Enterobacteriaceae. Nature(London) 208:1016-1017.
3. Anderson, E. S. 1968. The ecology of transferable drugresistance in the enterobacteria. Annu. Rev. Microbiol.22:132-180.
4. Anderson, E. S., and M. J. Lewis. 1965. Characteriza-tion of a transfer factor asociated with drug resistancein Salmonella typhimurium. Nature (London)208:843-849.
5. Anderson, E. S., and E. Natkin. 1972. Transduction ofresistance determinants and R factors of the A transfersystems by phage Plkc. Mol. Gen. Genet. 114:261-265.
6. Ashford, W. A., Ri G. Golash, and V. G. Hemming.1976. Penicillinase-producing Neisseria gonorrhoeae.Lancet ii:657-658.
7. Biewas, G., S. Comer, and P. F. Sparling. 1976. Chro-mosomal location of antibiotic resistance genes in Neis-seria gonorrhoeae. J. Bacteriol. 125:1207-1210.
8. Biswas, G. D., T. Sox, E. Blackman, and P. F. Spar-ling. 1977. Factors affecting genetic transformation ofNeisseria gonorrhoeae. J. Bacteriol. 129:983-992.
9. Crosa, J. H., L K. Luttropp, F. Heffron, and S. Fal-kow. 1975. Two replication initiation sites on R-plasmid
DNA. Mol. Gen. Genet. 140:39-50.10. Davis, B. D., and E. S. Mingioli. 1950. Mutants of
Escherichia coli requiring methionine or vitamin B12. J.Bacteriol. 60:17-28.
11. DeLey, J. 1970. Reexamination ofthe association betweenmelting point, buoyant density, and chemical base com-position of deoxyribonucleic acid. J. Bacteriol.101:738-754.
12. Eisenstein, B. I., T. Sox, G. Biewas, E. Blackman, andP. F. Sparling. 1977. Conjugal transfer of the gonococ-cal penicillinase plasmid. Science 195:998-1000.
13. Falkow, S. 1975. Infectious multiple drug resistance. PionLimited, London.
14. Freifelder, D. 1971. Isolation of extrachromosomal DNAfrom bacteria. Methods Enzymol. 21D:153-163.
15. Kirvin, L A., and C. Thorneberry. 1977. Transfer ofbeta-lactamase genes of Neisseria gonorrhoeae by con-jugation. Antimicrob. Agents Chemother.11:1004-1006.
16. Kleinschmidt, A. K. 1968. Monolayer techniques in elec-tron microscopy of nucleic acid molecules. MethodsEnzymol. 12B:361-379.
17. Marmur, J. 1961. A procedure for the isolation of deoxy-ribonucleic acid from micro-organisms. J. Mol. Biol.3:208-218.
18. Mayer, L. W., K. K. Holmes, and S. Falkow. 1974.Characterization of plasmid deoxyribonucleic acid fromNeisseria gonorrhoeae. Infect. Immun. 10:712-717.
19. Meyers, J. A., D. Sanchez, L P. Elwell, and S. Fal-kow. 1976. Simple agarose gel electrophoretic methodfor the identification and characterization of plasmiddeoxyribonucleic acid. J. Bacteriol. 127:1529-1537.
20. Meyneli, E., and N. Datta. 1967. Mutant drug resistancefactors of high transmissibility. Nature (London)214:885-887.
21. Nishimura, A., Y. Nishimura, and L. Caro. 1973. Iso-lation of Hfr strains from RI and ColV2+ strains ofEscherichia coli and derivation of an R' lac factor bytransduction. J. Bacteriol. 116:1107-1112.
22. Nisioka, T., L. Michiko, and R. Clowes. 1969. Compos-ite circular forms of R factor deoxyribonucleic acidmolecules. J. Bacteriol. 97:376-385.
23. Novick, Ri P., R. C. Clowes, S. N. Cohen, R. Curtissm, N. Datta, and S. Falkow. 1976. Unifonn nomen-clature for bacterial plasmids: a proposal. Bacteriol.Rev. 40:168-189.
24. O'Callaghan, C. H., A. Morris, S. M. Kirby, and A. H.Shindler. 1972. Novel method for detection off,-lac-tamase by using a chromogenic cephalosporin substrate.Antimicrob. Agents Chemother. 1:283-288.
25. Percival, A., J. Rowlands, J. E. Corkill, C. D. Aler-gant, 0. P. Arya, E. Rees, and E. aL Annels. 1976.Penicillinase-producing gonococci in Liverpool. Lancetli:1379-1382.
26. Phillips, L. 1976. Beta-lactamase-producing, penicillin-re-sistant gonococcus. Lancet ii:656-657.
27. Roberts, M., L. P. Elwell, and S. Falkow. 1977. Molec-ular characterization of two beta-lactamase-specifyingplasmids isolated from Neisseria gonorrhoeae. J. Bac-teriol. 131:557-563.
28. Roberts, M., and S. Falkow. 1977. Conjugal transfer ofR plasmids in Neisseria gonorrhoeae. Nature (London)266:630-631.
29. Rowbury, R. J. 1977. Bacterial plasmids with particularreference to their replication and transfer properties.Prog. Biophys. Mol. Biol. 31:271-318.
30. Rubin, F. A., and D. H. Smith. 1973. Characterizationof R factor ,B-lactamases by the acidimetric method.Antimicrob. Agents Chemother. 3:68-73.
31. Sarubbi, F. A., Jr., E. Blackman, and P. F. Sparling.1974. Genetic mapping of linked antibiotic resistanceloci in Neisseria gonorrhoeae. J. Bacteriol. 120:1284-1292.
VOL. 134, 1978 285
on May 12, 2021 by guest
http://jb.asm.org/
Dow
nloaded from

286 SOX ET AL.
32. Sarubbi, F. A., Jr., and P. F. Sparling. 1974. Transferof antibiotic resistance in mixed cultures of Neisseriagonorrhoeae. J. Infect. Dis. 130:660-663.
33. Stiffler, P. W., S. A. Lerner, M. Bohnhoff, and J. A.Moreilo. 1975. Plasmid deoxyribonucleic acid in clinicalisolates of Neisseria gonorrhoeae. J. Bacteriol.122:1293-1300.
34. Szybalski, W. 1968. Use of cesium sulfate for equilibriumdensity gradient centrifugation. Methods Enzymol.12B:330-360.
35. Thuring, R. W. J., J. M. Sanders, and P. Borst. 1975.A freeze-squeeze method for recovering long DNA fromagarose gels. Anal. Biochem. 66:213-220.
36. Weliauer, P. K., R. H. Reeder, D. Carroll, D. D.Brown, A. Deuter, T. Higashima-kagawa, and 1.B. Dawid. 1974. Amplified ribosomal DNA from Xen-opus laevis has heterogenous spacer lengths. Proc. Natl.Acad. Sci. U.S.A. 71:2823-2827.
37. Willetts, N. 1972. The genetics of transmissable plasmids.Annu. Rev. Genet. 6:257-268.
J. BACTERIOL.
on May 12, 2021 by guest
http://jb.asm.org/
Dow
nloaded from