Embed Size (px)
Citation preview

Biocatalysis, 1987, Vol. 1, pp. 81-98 Photocopying permitted by license only
0 1987 hanvood academic publishers GmbH Printed in the United Kingdom
COMPUTER GRAPHICS AS A TOOL FOR THE PREDICTION OF THE STEREOSELECTIVITY
OF ENZYME CATALYZED REACTIONS.
OF SUBSTITUTED PROPANEDIOIC ACID DIESTERS.
a XHYMOTRY PSIN CATALYZED HYDROLYSIS
FREDRIK BJORKLING, TORBJORN NORIN* and PETER SZMULIK
Department of Organic Chemistry, Royal Institute of Technology, S-100 44 Stockholm, Sweden
JOHN BOUTELJE and KARL HULT*
Department of Biochemistry and Biotechnology, Royal Institute of Technology, S-100 44 Stockholm, Sweden
PER KRAULIS
Department of Molecular Biology, Box 590, Biomedical Center, S-751 24 Uppsala, Sweden
(Received 30 June 1986; in final form 30 September 1986)
Computer graphics can be used for the prediction of substrate specificity' and stereoselectivity in the a-chymotrypsin catalyzed hydrolysis of disubstituted propanedioic acid diesters. This enzyme catalyzed reaction is useful for the preparation of chiral starting materials, e.g. for the synthesis of optically pure cu-methylaminoacids.
KEY WORDS cu-Chymotrypsin, computer graphics, enantioselective hydrolysis, substituted malonate esters.
INTRODUCTION
Stereoselective hydrolysis of prochiral or racemic dicarboxylic acid diesters catalyzed by various hydrolytic enzymes has emerged recently as an important method for the preparation of optically active synthons (Bjorkling et al., 1985a, b, c; Whitesides and Wong, 1985 and references cited therein).
The family of serine proteases, of which a-chymotrypsin is a member, has been studied in detail by X-ray crystallography. Several structures of the native enzymes and the enzymes complexed with inhibitors have been determined at high resolution (Marquart et al., 1983; Walter and Bode, 1983; Tsukada and Blow, 1985). This forms a good basis for the modelling of different enzyme- substrate intermediates.
~~
* To whom correspondence should be addressed.
87
Bio
cata
l Bio
tran
sfor
mat
ion
Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
Hua
zhon
g U
nive
rsity
of
Scie
nce
& T
echn
olog
y on
11/
15/1
2Fo
r pe
rson
al u
se o
nly.
88 F. BJORKLING ET A L .
la
2a
3a
la
5a
60
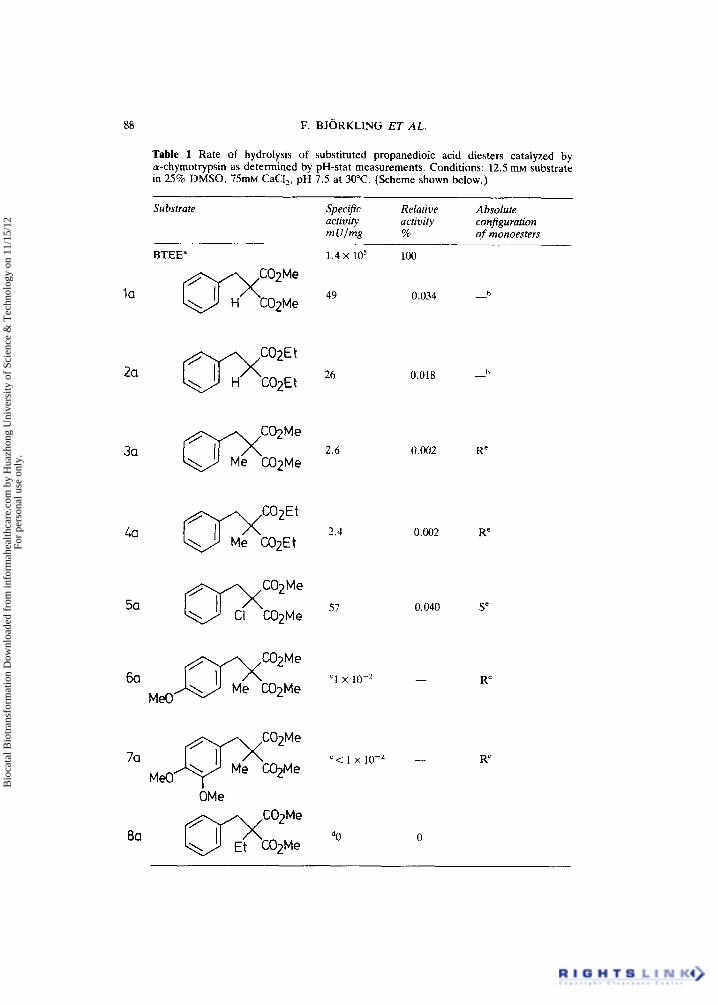
Table 1 Rate of hydrolysis of substituted propanedioic acid diesters catalyzed by tu-chymotrypsin as determined by pH-stat measurements. Conditions: 12.5 mM substrate in 25% DMSO, 75mM CaCI,, pH 7.5 at 30°C. (Scheme shown below.)
Substrate Specific Relative Absolute activity activity configuration m Ulme 70 of monoesters
BTEE"
CO2Me
7a MeO
OMe C02Me
8a m Q M e
1.4 x 105 100
49 0.034 - b
26
2.6
2.4
b 0.018 -
0.002 R'
0.002 R"
0.040 S" 57
c1 x 10-2 -
- '<1 x
*O 0
Re
R"
Bio
cata
l Bio
tran
sfor
mat
ion
Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
Hua
zhon
g U
nive
rsity
of
Scie
nce
& T
echn
olog
y on
11/
15/1
2Fo
r pe
rson
al u
se o
nly.

COMPUTER PREDICTION OF ENZYME REACTIONS 89
In order to investigate the use of interactive graphics molecular mechanics calculations to predict substrate specificity and enantioselectivity, we have correlated theoretical and experimental results for the a-chymotrypsin catalyzed hydrolysis of several disubstituted propanedioic acid diesters (see Table 1). The products of this hydrolytic reaction have been shown to be valuable intermediates in the synthesis -of optically pure (S)-a-methylaminoacids (Bjorkling et al., 1985a).
MATERIALS AND METHODS Synthesis of Substrates
Nuclear magnetic resonance (NMR) spectra were recorded on a Bruker 200 MHz instrument with CDC13 as solvent. Chemical shifts are reported relative to tetramethylsilane (TMS). Mass spectra were recorded on a VG 7070E instrument and high resolution mass spectra were recorded on a Kratos MS 50 instrument interfaced to a DS 55 data recording system. Gas chromatographic analyses were performed on a Pye Unicam 204 instrument equipped with a DB-WAX 30 N fused silica capillary column, obtained from J & W Scientific Inc. Determinations of enantiomeric excess e.e. = 100 I([R] - [ S ] ) / ( [ R ] + [S])l of the monoesters were made by NMR analyses of the diastereotopic protons of the corresponding salts with optically pure (+) and (-)-phenylethyl amine in CDC13 solution (Bjorkling et al., 1985b). Optical rotation measurements were made in CHC13 solution, and circular dicroism (CD) measurements in methanol solution. Benzylpropanedioic acid dimethyl ester (la), benzylpropanedioic acid diethyl ester (2a), benzylmethyl- propanedioic acid dimethyl ester (3a) , benzylmethylpropanedioic acid diethyl ester (4a), 4-methoxybenzylmethylpropanedioic acid dimethyl ester (6a) and 3,4-dimethoxybenzylmethylpropanedioic acid dimethyl ester (7a) were prepared as previously described (Bjorkling et al., 1985a,c).
Benzylchloropropanedioic acid dimethyl ester (5a) Benzyl bromide 30g was dissolved in dry tetrahydrofuran 100ml under argon atmosphere. To this solution was added sodium hydride 3.07g as a 80%
* N-benzoyl-L-thyrosine ethylester, a good wu-chymotrypsin substrate.
=The activity was too low to be measured by the pH-stat method. The activity was estimated from GC-analyses of long term incubations (18-56 days).
No hydrolysis product was isolated when using the normal incubation procedure even after 14 days of incubation. *The optical purity was larger than 98% e.e. according to NMR analysis.
The monoester racemices during acidic work-up.
Scheme for the hydrolysis of substituted propanedioic acid diesters
l a 2a 3a 4a 5a 69 7a 89
Bio
cata
l Bio
tran
sfor
mat
ion
Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
Hua
zhon
g U
nive
rsity
of
Scie
nce
& T
echn
olog
y on
11/
15/1
2Fo
r pe
rson
al u
se o
nly.

90 F. BJORKLING ET AL
dispersion in mineral oil and chloropropanedioic acid dimethyl ester 15 g in small portions under cooling. The mixture was stirred overnight, poured into cold water, extracted with diethyl ether (3 X 100 ml), washed successively with hydrochloric acid (2 N, 1 X 50 m1) followed by saturated sodium bicarbonate (1 X 50 ml), and dried with MgS04. The solvent was removed under reduced pressure, yielding an oil that crystallized upon treatment with a small portion of light petroleum (bp 40-60°C). Recrystallization from light petroleum gave benzylchloropropanedioic acid dimethyl ester (13.5 g, 74%), mp 50"C, 'H NMR 6: 3.57 (s,2H), 3.81 (s,6H), 7.2-7.3 (unres., 5H). "C NMR 6: 44.1, 54.5, 71.3, 128.1, 128.6, 130.8, 133.9, 167.0. Mass spectrum m/e (% relative intensity) M+ not detected, 221 (38.9) C12H1304, 189 (100) CllH9O3, 162 (6.91) C10H1002, 103 (14.4) C8H7, 91 (84.2) GH7, 77 (14.9) C6H5, 59 (9.73) C2H302; calculated for C12H1304 221.0813, measured 221.0810.
Benzylethylpropanedioic acid dimethyl ester (Sa) This compound was pre ared according to a previously described procedure (Bjorkling et al . , 1985~). H NMR 6: 0.89 (t, 3H), 1.81 (q,2H), 3.21 (s, 2H), 3.69 (s, 6H), 7.0-7.2 (unres., 5H) I3C NMR 6: 9.7, 25.9, 38.7, 52.9, 60.2, 127.2, 128.5, 130.1, 136.4, 171.7. Mass spectrum m/e (% relative intensity) M+ 250 (22.1)
(100.0) GH7, 77 (3.6) C6H5, 59 (5.3) GH30,; calculated for C14H1804 250.1205, measured 250.1196.
P
C14H1804, 221 (4.4) C13H1703, 190 (42.0) C12H1402, 189 (13.0) CllH903, 91
Hydrolysis of Disubstituted Propanedioic Acid Diesters
Enzymes a-Chymotrypsin (EC 3.4.21.1), 3 x crystallized from 4 x crystallized chymotryps- inogen, and pig liver esterase (EC 3.1.1.1) as a (NH4),S04 suspension, were obtained from Sigma.
Incubations : Typical Procedure Hydrolyses were carried out in 20 ml buffered batches containing dimethylsul- phoxide (DMSO; 25%), tris(hydroxymethy1)aminomethane (Tris-HC1) (0.375 M, pH 7.3, substrate (20 mM) and a-chymotrypsin (25-200 mg) or pig liver esterase (lmg). Reaction temperature was kept at 22-25°C. The reactions were moni- tored by GC analyses of diethyl ether extracts. After completion of the reaction (24-48 h with cr-chymotrypsin, and <1 h with pig liver esterase) the reaction mixture was acidified and the product was extracted with diethyl ether. The crude extract was extracted with saturated sodium bicarbonate (3 x 50 ml), washed with diethyl ether (1 x 50 ml), acidified with hydrochloric acid ( 2 ~ ) , extracted with diethyl ether (3 x 50ml) and dried with MgS04, and the solvent was removed under reduced pressure. Kinetics Reaction velocities were determined in a Radiometer pH-stat under N2 at 30°C; pH was maintained at 7.5 with lOmM NaOH. The reaction mixture contained substrate (0.5-12.5 mM), DMSO (25%), CaC1, (75 mM) and a-ch motrypsin in a total volume of 4 ml. The amount of enzyme added was 4 x 10- mg or 5 mg in 1 mM. HCl for N-benzoyl-L-thyrosine ethylester (BTEE) and the other substrates, respectively. One unit of enzyme activity was defined as the amount of enzyme
Y
Bio
cata
l Bio
tran
sfor
mat
ion
Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
Hua
zhon
g U
nive
rsity
of
Scie
nce
& T
echn
olog
y on
11/
15/1
2Fo
r pe
rson
al u
se o
nly.

COMPUTER PREDICTION OF ENZYME REACTIONS 91
catalyzing the hydrolysis of 1 pmol of substrate per minute under the conditions described. The apparent rate constant for the base catalysis was determined by measuring the rate of hydrolysis at pH 10 (pH 10.5 for BTEE) under the same conditions as given above but without enzyme.
Disubstituted propanedioic acid monoesters (3b and 5b) Benzylmethylpropanedioic acid monomethyl ester (3b) was obtained by standard incubation of the corresponding diester 3a with a-chymotrypsin according to Bjorkling et al. (1985~). CD: A,,, = 249 nm.
Benzylchloropropanedioic acid monomethyl ester (5b) was obtained by stand- ard incubation of the corresponding diester 5a with a-chymotrypsin. 'H NMR 6: 3.51 (s,2H), 3.75 (s,3H), 7.21 (unres., 5H). 13C NMR S: 43.94, 54.64, 71.94, 128.14, 128.64, 130.81, 133.62, 167.01, 169.93. CD: Ami, = 250 nm. Mass spectrum m/e (% relative intensity) 242 (1.2), 162 (55), 131 (79), 103 (59), 91 (loo), 77 (42). Due to the instability of this halfester no high resolution MS data was obtained.
Determination of absolute configurations of 3b and 5b The absolute configuration of benzylmethylpropanedioic acid monomethyl ester (3b) was assigned to be (R) by comparison of the optical rotation ( [ @ I D = 4.8"(c = 1.86, CHC13)) with an authentic sample of (R)-(+)-benzylmethyl- propanedioic acid monomethyl ester of identical [a], (Bjorkling et al., 1985~).
In order to obtain a reference sample of nearly racemic benzylchloropropane- dioic acid monomethyl ester (5b), a pig liver esterase hydrolysis (see Incubations above) of the corresponding dimethyl ester 5a was performed, thus yielding the (S)-monoester in 9% e.e. as determined by the NMR method. The corresponding a-chymotrypsin catalyzed hydrolysis yielded a product of >98% e.e.
The benzylchloropropanedioic acid monomethyl ester (5b) obtained by (Y-
chymotrypsin hydrolysis exhibits a negative Cotton effect in its CD-spectrum, similar to that of (R)-benzylmethylpropanedioic acid monomethyl ester (3b). This indicates that the chloro derivative is configurationally related to (R)-( +)- benzylmethylpropanedioic acid monomethyl ester (Nilsson, 1985). This assign- ment is supported by the similar NMR shifts of the diastereotopic protons of the (+) and (-)-phenylethylamine salts of 3b and 5b. Attempts were made to transform 5b into a compound of known absolute configuration. So far, these attempts have been unsuccessful due to the instability of 5b.
Computer Graphics The interactive molecular graphics computer program used to model the different states of the reaction was TOM (Cambillau, Horjales and Jones, 1984) interfaced to FRODO (Jones, 1978; 1982; Kollman, 1985) on an Evans & Sutherland PS300 display, running under VAX/VMS on a MicroVAX 11. The program works in dihedral conformational space only, and uses the energy potentials for charge- charge, torsion and van der Waals interaction from the molecular dynamics program package GROMOS (Hermans et al., 1984). The energy of the system is minimized using the conjugate gradient method.
The models of the substrates were built using standard bond lengths and bond angles. The energy minimization was performed by allowing the dihedral angles
Bio
cata
l Bio
tran
sfor
mat
ion
Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
Hua
zhon
g U
nive
rsity
of
Scie
nce
& T
echn
olog
y on
11/
15/1
2Fo
r pe
rson
al u
se o
nly.

92 F. BJORKLING ET AL.
at non-carboxylic single bonds in the substrate and at side-chain bonds of the enzyme to be Aexible. In the model of the non-covalent complex (the Michaelis complex) the substrate was allowed to move relative to the enzyme. The covalent complex (the tetrahedral intermediate) was modelled by introducing a covalent bond between the y-oxygen of Ser-195 and the carboxylic carbon of the substrate while changing to tetrahedral bond angles. Hydrogen bonds are implicitly included in the calculation by introduction of all hydrogens bonded to polar atoms (nitrogen and oxygen) in the coordinate set.
The geometries of the different states were obtained by energy minimization from reasonable starting positions as guided by the crystal structures of inhibitor complexes of the closely related enzyme trypsin (Marquart et al., 1983; Walter and Bode, 1983; Tsukada and Blow, 1985). The phenyl ring of the substrate was placed in the hydrophobic pocket of the enzyme formed mainly by residues 190-195 and 213-217. The methyl-ester group to be hydrolyzed by the enzyme was positioned for nucleophilic attack by y-oxygen of Ser-195 with the carboxylic oxygen oriented towards the potentially hydrogen bonding peptide amide groups of Gly-193 and Ser-195. At different stages of the minimization, the models were adjusted manually to check against trapping in local energy minima.
The coordinates for a-chymotrypsin were those from the determination at 1.68 A resolution and refined to a crystallographic R-factor of 0.234 (Tsukada and Blow, 1985). There are two independently determined molecules in this structure determination. There is no significant difference between the structures of the active sites in the two molecules. The first molecule in the set (designated A ) was used. During energy minimization, only the side-chains of the residues within 3.5 A from any atom of the substrate were allowed to move. In no case except for His-57 and Ser-195 was the movement of the side-chains significant.
RESULTS AND DISCUSSION
Since the structure of a-chymotrypsin is well determined, it is possible to use computer graphics for predicting the stereochemical outcome of the a- chymotrypsin catalyzed hydrolysis of prochiral esters. The accuracy of the active site-substrate interaction energy values, as obtained by the present method, allow only qualitative conclusions of the relative stability of the enzyme-substrate complexes. This is mainly because of the exclusion of water in the calculations, and also due to the approximations made when bond angles and bond lengths are constrained.
The reason for the enantioselectivity in the hydrolysis of prochiral disubstituted propanedioic acid diesters by a-chymotrypsin is shown to be the different steric interactions experienced by the non-hydrolyzed ester group of the substrate in the two possible tetrahedral intermediates. This is evident from the large difference in stability between the tetrahedral intermediates of ( R ) - and (S)-configurations as calculated by the molecular mechanics program TOM. In the Michaelis (non- covalent) complex, the difference in stability between the two possible (pro-R and pro-S) enzyme-substrate complexes is, however, insignificant.
Figure l a shows the tetrahedral intermediate of (S)-configuration with substrate 3a. Because of nomenclature reasons, the intermediate with (S)-configuration
Bio
cata
l Bio
tran
sfor
mat
ion
Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
Hua
zhon
g U
nive
rsity
of
Scie
nce
& T
echn
olog
y on
11/
15/1
2Fo
r pe
rson
al u
se o
nly.

COMPUTER PREDICTION OF ENZYME REACTIONS 93
Figure l a Stereoscopic view of the tetrahedral intermediate of (S)-configuration of substrate 3a with a-chymotrypsin (see color plate I).
Figure l b Stereoscopic view of the two tetrahedral intermediates of substrate 3a with a- chymotrypsin. The @)-intermediate is drawn in green and the @)-intermediate in red (see color plate 11).
Bio
cata
l Bio
tran
sfor
mat
ion
Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
Hua
zhon
g U
nive
rsity
of
Scie
nce
& T
echn
olog
y on
11/
15/1
2Fo
r pe
rson
al u
se o
nly.
94 F. BJORKLING E T A L .
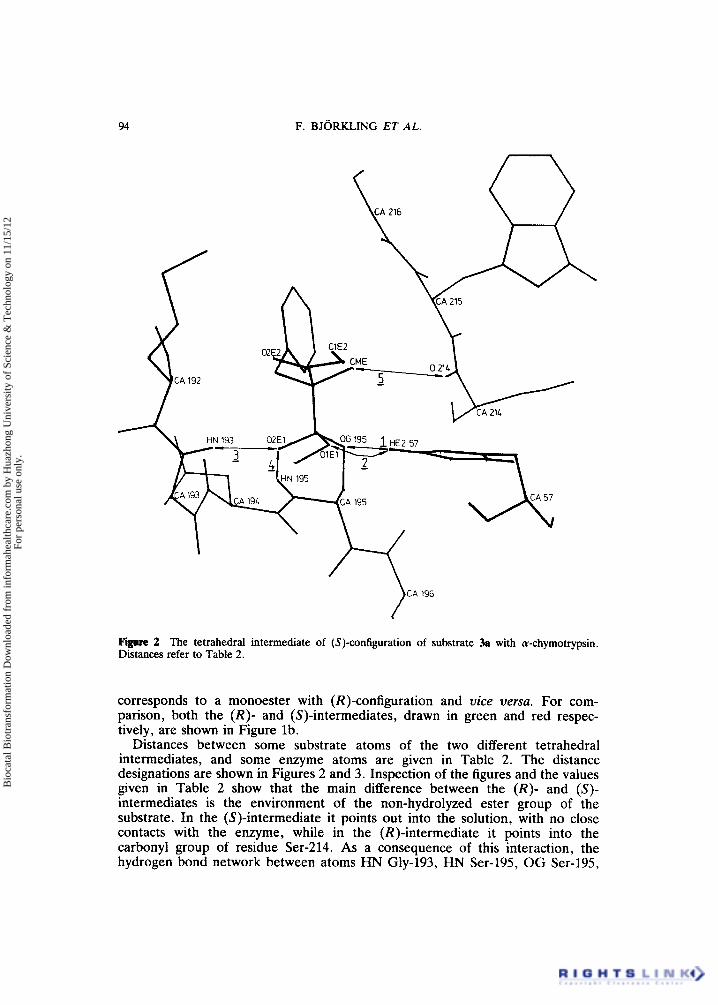
IGgure 2 The tetrahedral intermediate of (S)-configuration of substrate 3a with a-chymotrypsin. Distances refer to Table 2.
corresponds to a monoester with (R)-configuration and vice versa. For com- parison, both the ( R ) - and (S)-intermediates, drawn in green and red respec- tively, are shown in Figure lb.
Distances between some substrate atoms of the two different tetrahedral intermediates, and some enzyme atoms are given in Table 2. The distance designations are shown in Figures 2 and 3. Inspection of the figures and the values given in Table 2 show that the main difference between the ( R ) - and (S)- intermediates is the environment of the non-hydrolyzed ester group of the substrate. In the (S)-intermediate it points out into the solution, with no close contacts with the enzyme, while in the (R)-intermediate it points into the carbonyl group of residue Ser-214. As a consequence of this interaction, the hydrogen bond network between atoms HN Gly-193, HN Ser-195, OG Ser-195,
Bio
cata
l Bio
tran
sfor
mat
ion
Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
Hua
zhon
g U
nive
rsity
of
Scie
nce
& T
echn
olog
y on
11/
15/1
2Fo
r pe
rson
al u
se o
nly.

COMPUTER PREDICTION OF ENZYME REACTIONS 95
Figure 3 The tetrahedral intermediate of @)-configuration of substrate 3a with a-chymotrypsin. Distances refer to Table 2.
O l E l substrate and HE2 His-57 is substantially altered, since the tetrahedral carbon of the substrate has been forced from His-57 towards Gly-193. The proton HE2 His-57 should transfer to OlEl to facilitate the leaving of a methanol molecule from the tetrahedral intermediate. This is made more difficult in the @)-intermediate by the increased distance between those atoms.
In the (R)-intermediate, there is also a close steric intrasubstrate contact between the atoms 02E2 and CPH2. This decreases further the stability of the (R)-intermediate. The ethyl group of substrate 8a comes in contact with the CA Trp-215 atom and thereby decreases the stability of the (S)-intermediate of this substrate, so that in this case neither intermediate is capable of forming.
These results are in good agreement with those of Wipff et al. (1983), who have investigated the a-chymotrypsin catalyzed hydrolysis of L- and D-N-
Bio
cata
l Bio
tran
sfor
mat
ion
Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
Hua
zhon
g U
nive
rsity
of
Scie
nce
& T
echn
olog
y on
11/
15/1
2Fo
r pe
rson
al u
se o
nly.
96 F. BJORKLING ET AL.
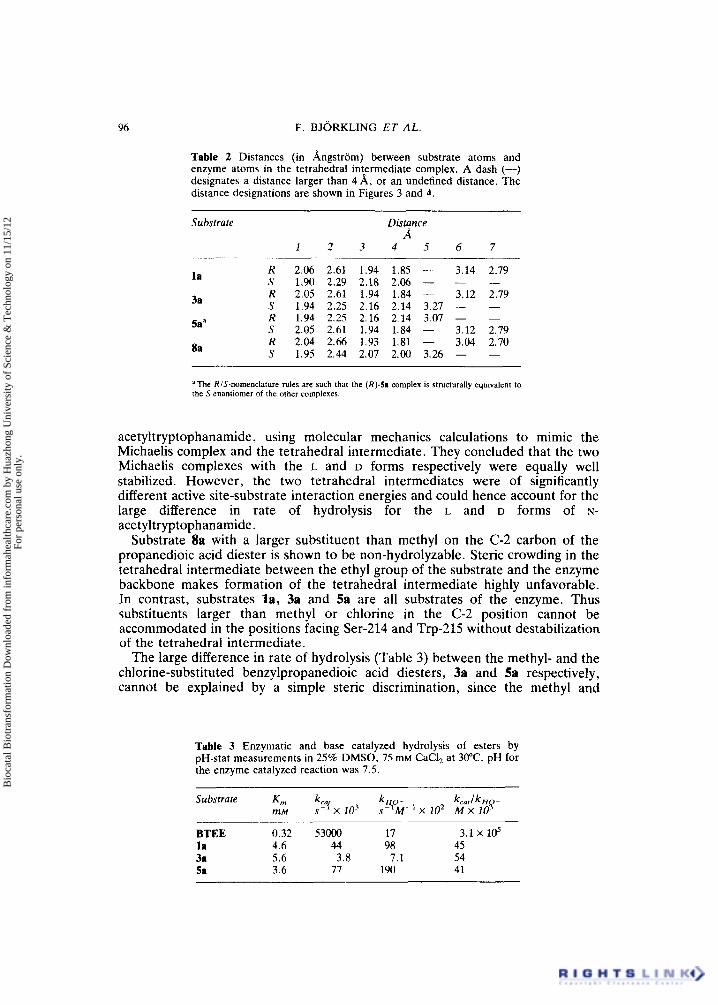
Table 2 Distances (in Angstrom) between substrate atoms and enzyme atoms in the tetrahedral intermediate complex. A dash (-) designates a distance larger than 4 A, or an undefined distance. The distance designations are shown in Figures 3 and 4.
~
Substrate Distance A
1 2 3 4 5 6 7
R 2.06 2.61 1.94 1.85 - 3.14 2.79 S 1.90 2.29 2.18 2.06 - - - R 2.05 2.61 1.94 1.84 - 3.12 2.79 S 1.94 2.25 2.16 2.14 3.27 - - R 1.94 2.25 2.16 2.14 3.07 - - S 2.05 2.61 1.94 1.84 - 3.12 2.79 R 2.04 2.66 1.93 1.81 - 3.04 2.70 S 1.95 2.44 2.07 2.00 3.26 - -
la
3a
5a"
8a
aThe RIS-nomenclature rules are such that the (R)-5 i complex is structurally equivalent to the S enantiomer of the other complexes.
acetyltryptophanamide , using molecular mechanics calculations to mimic the Michaelis complex and the tetrahedral intermediate. They concluded that the two Michaelis complexes with the L and D forms respectively were equally well stabilized. However, the two tetrahedral intermediates were of significantly different active site-substrate interaction energies and could hence account for the large difference in rate of hydrolysis for the L and D forms of N- acetyltryptophanamide.
Substrate 8a with a larger substituent than methyl on the C-2 carbon of the propanedioic acid diester is shown to be non-hydrolyzable. Steric crowding in the tetrahedral intermediate between the ethyl group of the substrate and the enzyme backbone makes formation of the tetrahedral intermediate highly unfavorable. In contrast, substrates la , 3a and 5a are all substrates of the enzyme. Thus substituents larger than methyl or chlorine in the C-2 position cannot be accommodated in the positions facing Ser-214 and Trp-215 without destabilization of the tetrahedral intermediate.
The large difference in rate of hydrolysis (Table 3) between the methyl- and the chlorine-substituted benzylpropanedioic acid diesters, 3a and 5a respectively, cannot be explained by a simple steric discrimination, since the methyl and
Table 3 Enzymatic and base catalyzed hydrolysis of esters by pH-stat measurements in 25% DMSO, 75 mM CaCI, at 30°C. pH for the enzyme catalyzed reaction was 7.5.
BTEE 0.32 53000 17 3.1 x 105 la 4.6 44 98 45 3a 5.6 3.8 7.1 54 Sa 3.6 77 190 41
Bio
cata
l Bio
tran
sfor
mat
ion
Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
Hua
zhon
g U
nive
rsity
of
Scie
nce
& T
echn
olog
y on
11/
15/1
2Fo
r pe
rson
al u
se o
nly.

COMPUTER PREDICTION OF ENZYME REACTIONS 97
chlorine groups are of similar size. To determine? whether this is due to differences in the interaction with the enzyme or due to the inherent properties of the substrates, we determined the rate constant kHO- for the base catalyzed hydrolysis of substrates la, 3a and 5a in a homogeneous aqueous medium. As can be seen from Table 3, the ratios kcar/kHo- are very similar for the various substrates. This means that the enzyme is equally efficient in catalysing the hydrolysis of these substrates and that the relative rates of hydrolysis are determined by the inherent properties of the substrates. Neither are there any large differences in binding constants for the benzylpropanedioic acid diesters as is obvious from the K , values in Table 3. It can also be seen (Table 3) that the rate enhancement for the good substrate BTEE catalyzed by the enzyme, compared to the base catalyzed hydrolysis, is much larger than for the benzylpropanedioic acid diesters.
In contrast to previous observations on pig liver esterase catalyzed hydrolysis of propanedioic acid diesters (Bjorkling et al., 1985c), the ester function of the substrate in a-chymotrypsin catalyzed hydrolysis exhibits no influence on the stereochemical outcome of the reaction and only limited influence on the reaction rate. As evident from computer graphics, the alcohol part of the non-hydrolysed ester function experiences no serious steric interaction with the enzyme, which explains the results obtained for methyl and ethyl diesters.
Substituents on the phenyl ring also effect the stability of the enzyme-substrate intermediates. The 3,4-dimethoxy derivative 7a fits precisely in the hydrophobic pocket, and there is no space for larger substituents in the 3- and 4-positions, nor for substituents in the 2- and 6-positions of the phenyl ring. As evidenced by X-ray crystallography (Tsukada and Blow, 1985), in the bottom of the hydropho- bic binding site there is a tightly bonded water molecule (designated H605 in the coordinate set, hydrogen-bonded to 0 Val-227 and OH Tyr-228) situated in a surrounding important for the interaction of the para-methoxy group of the substrate with the enzyme. The hydrolysis of 6a and 7a were highly stereoselec- tive but the reaction rates were very low (Table 1); this may be due to slow release of water from the hydrophobic pocket. Since water was excluded from the molecular mechanics calculations, no such effect could be studied by this approach. Unfortunately, there are today no established methods for calculating the influence of solvents on the enzyme-substrate interaction (Chothia, 1974). However, there are attempts at empirical descriptions aiming at rough estimates of the entropic effect of solvent accessibility. This may yield useful complemen- tary information to the quantitative results of sophisticated molecular mechanics calculations (Eisenberg and McLachlan, 1986).
For the comparison of the specific activities, N-benzoyl-L-tyrosine eth lester (BTEE) is included in Table 3. This substrate exhibits activity up to 10 times higher than that of the propanedioic acid diesters. It is thus clear that modifications proposed from computer graphics studies of either the enzyme or the substrate could lead to large improvements in catalytic rate.
A ckno wledgernents Financial support from the Swedish Natural Science Research Council and the Swedish Board for Technical Development is gratefully acknowledged. We thank
Y
t We thank the referee for a valuable comment on the different rates of hydrolysis of substrates 3a and 5a.
Bio
cata
l Bio
tran
sfor
mat
ion
Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
Hua
zhon
g U
nive
rsity
of
Scie
nce
& T
echn
olog
y on
11/
15/1
2Fo
r pe
rson
al u
se o
nly.

98 F. BJORKLING E T A L .
Dr Eduardo Horjales and Dr Christian Cambillau for their help in the use of the program TOM. Professor Jan Sandstrom and Dr Ingemar Nilsson are acknow- ledged for the CD-measurements. We thank Dr Jaceck Bielawski at the Swedish Tobacco Co. and Dr Borje Egestad at the Karolinska Institute for running high resolution MS and MS, respectively.
References Bjorkling, F., Boutelje, J . , Gatenbeck, S. , Hult, K. and Norin, T. (1985a) Enzyme catalyzed
hydrolysis of dialkylated propanedioic acid diesters, synthesis of optically pure (S)-a- methylphen ylalanin, (S)-a-methylthyrosine and (S)- a-methyl-3,4-dimethoxyphenylalanin. Terruhed- ron Len., 26, 4957-4958
Bjorkling, F. , Boutelje, J. , Gatenbeck. S. , Hult. K. and Norin, T. (1985b) Enzyme catalyzed hydrolysis of the esters of cis- and rrum- cyclohexanedicarboxylic acids. Appl. Microbiol. Biofechnol., 21, 16- 19
Bjorkling, F., Boutelje, J., Gatenbeck, S . , Hult, K., Norin, T. and Szmulik, P. (198%) Enzyme catalyzed hydrolysis of dialkylated propanedioic acid diesters, chain length dependent reversal of enantioselectivity. Tetrahedron, 41, 1347- 1352
Cambillau, C., Horjales, E . and Jones, T. A. (1984) TOM, a display program for fitting ligands into protein receptors and performing interactive energy minimization. J . Mol. Graph., 2, 53-54
Chothia, C. (1974) Hydrophobic bonding and accessible surface area in proteins. Nature, 248, 338-339
Eisenberg, D. and McLachlan, A. D. (1986) Solvation energy in protein folding and binding. Nature, 319, 199-203
Hermans, J., Berendsen, H. J . C.. van Gunsteren, W. F. and Postma, J . P. M. (1984) A consistent empirical potential for water-protein interactions. Biopolymers, 23, 1513-1518
Jones, T. A . (1978) A graphics model building and refinement system for macromolecules. J. Appl. Crystallogr., 11, 268-272
Jones, T. A. (1982) FRODO: A graphics fitting program for macromolecules. In Computational Crystallography, edited by D. Sayre, pp. 303-317. Oxford: Oxford University Press.
Kollman, P. (1985) Theory of complex molecular interactions: computer graphics, distance geometry, molecular mechanics, and quantum mechanics. Acc. Chem. Res., 18, 105-111
Marquart, M., Walter, J . , Deisenhofer, J . , Bode, W. and Huber, R. (1983) The geometry of the reactive site and of the peptide groups in trypsin. trypsinogen and its complexes with inhibitors. Acra Crystallogr. Secf. B , B39, 480-490
Nilsson, I. (1985) The conformational dependence of the circular dichroism spectra of chiral indoles. PhD Thesis, University of Lund, Sweden
Tsukada, H. and Blow, D. M. (1985) Structure of alpha-chymotrypsin refined at 1.68 8, resolution. J. Mol. Biol., 184, 703-711
Walter, J. and Bode, W. (1983) The X-ray crystal structure analysis of the refined complex formed by trypsin and p-amidinophenylpyruvate at 1.4 Angstroms resolution. Hoppe-Seyler’s Z. Physiol., 364, 949-959
Whitesides, G. M. and Wong, C. (1985) Enzymes as catalysts in organic chemistry. Angew. Chem. Int. Ed. Engl., 24, 617-638
Wipff, G., Dearing, A., Weiner, P. K.. Blaney, J. M. and Kollman, P. A. (1983) Molecular mechanics studies of enzyme-substrate interactions: the interaction of L- and D-N- acetyltryptophanamide with a-chymotrypsin. J. Am. Chem. SOC. , 105,997- 1005
Bio
cata
l Bio
tran
sfor
mat
ion
Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
Hua
zhon
g U
nive
rsity
of
Scie
nce
& T
echn
olog
y on
11/
15/1
2Fo
r pe
rson
al u
se o
nly.





























