Embed Size (px)
Citation preview

Composite low affinity interactions dictaterecognition of the cyclin-dependent kinase inhibitorSic1 by the SCFCdc4 ubiquitin ligaseXiaojing Tanga, Stephen Orlickya, Tanja Mittagb,1, Veronika Csizmokb, Tony Pawsona,c, Julie D. Forman-Kayb,d,2,Frank Sicheria,c,d,2, and Mike Tyersa,e,2
aCenter for Systems Biology, Samuel Lunenfeld Research Institute, Toronto, ON, Canada M5G 1X5; bProgram in Molecular Structure and Function, TheHospital for Sick Children, 555 University Avenue, Toronto, ON, Canada M5G 1X8; cDepartment of Molecular Genetics, University of Toronto, Toronto,ON, Canada M5S 1A8; dDepartment of Biochemistry, University of Toronto, Toronto, ON, Canada M5S 1A8; and eInstitute for Research in Immunologyand Cancer, Université de Montréal, Montréal, QC, Canada H3C 3J7
Edited by* Angelika Amon, Massachusetts Institute of Technology, Cambridge, MA, and approved November 21, 2011 (received for review October 7, 2011)
The ubiquitin ligase SCFCdc4 (Skp1/Cul1/F-box protein) recognizesits substrate, the cyclin-dependent kinase inhibitor Sic1, in a multi-site phosphorylation-dependent manner. Although short dipho-sphorylated peptides derived from Sic1 can bind to Cdc4 withhigh affinity, through systematic mutagenesis and quantitativebiophysical analysis we show that individually weak, dispersedSic1 phospho sites engage Cdc4 in a dynamic equilibrium. Theaffinities of individual phosphoepitopes serve to tune the overallphosphorylation site threshold needed for efficient recognition.Notably, phosphoepitope affinity for Cdc4 is dramatically wea-kened in the context of full-length Sic1, demonstrating the impor-tance of regional environment on binding interactions. Themultisite nature of the Sic1-Cdc4 interaction confers cooperativedependence on kinase activity for Sic1 recognition and ubiquitina-tion under equilibrium reaction conditions. Composite dynamicinteractions of low affinity sites may be a general mechanismto establish phosphorylation thresholds in biological responses.
Phosphorylation governs many of the protein interactions thatdictate cellular responses to growth factors, stress conditions,
and internal regulatory circuits. This control can be exertedthrough the generation of specific phosphoepitopes that arerecognized by cognate modular binding domains, the paradig-matic example of which is the recognition of pTyr epitopes by theSH2 domain (1). Multisite phosphorylation can serve to integratesignals and often confers nonlinear responses to signal inputs,referred to as cooperativity or ultrasensitivity (2, 3). The basis forultrasensitivity lies in the requirement for a threshold level ofphosphorylation, which, in the presence of a countervailing phos-phatase, confers a nonlinear dependence on kinase activity (3).Multisite phosphorylation thresholds can derive from enzymeactivation mechanisms (4), decoy phosphorylation sites (5), bulkelectrostatic effects (6, 7), and zero-order enzyme saturation (8).
Phosphorylation of the yeast cyclin-dependent kinase (CDK)inhibitor Sic1 in late G1 phase by G1 CDK (Cln-Cdc28) activitydrives its recognition by the yeast F-box protein Cdc4, whichserves to recruit Sic1 for ubiquitination by the core Skp1/Cul1/F-box protein (SCF) ubiquitin ligase machinery (9–12). Versionsof Sic1 that lack multiple CDK phosphorylation sites fail to bindCdc4, are stabilized in vivo, and cause cell cycle arrest (13). Sic1binds to the WD40 domain of Cdc4 only if a nominal set of sixof the nine CDK consensus sites are phosphorylated, termedCdc4 phosphodegron (CPDs) (14). This multisite threshold de-rives from hydrophobic and electrostatic conflicts of the naturalCPD sites with the Cdc4 binding pocket, which weakens the in-dividual site affinities (15). Seven of the nine CPD sites lie in theintrinsically disordered 90 residue N-terminal targeting region(NTR) of Sic1; at least six of the CPD sites in the NTR interactwith Cdc4 in rapid equilibrium (16). An ensemble model of theSic1-Cdc4 complex suggests that the NTR interconverts betweendramatically different conformers (17). This conformational
diversity may facilitate ubiquitination at multiple sites (18) and/or enable long-range electrostatic interactions with the positivelycharged binding pocket on Cdc4 (7).
The human Cdc4 ortholog, called Fbw7, targets phosphory-lated forms of cyclin E, Myc, Jun, Notch, SREBP, and other pro-teins (19). The phosphodegron described for Fbw7 is similar tothat of Cdc4, except that an additional phosphorylated residue ispreferred at the P þ 4 position with respect to the primary (P0)phosphorylation site (20). The structure of Fbw7 bound to adiphospho-CPD peptide from cyclin E revealed a secondary bind-ing surface that engages the P þ 4 phosphate and thereby confersan overall high affinity for isolated diphosphorylated peptides(21). As Cdc4 also favors a phosphorylated residue at the P þ 4position in isolated peptides, it has been suggested that Cdc4/Fbw7 substrates are recognized solely via diphospho-CPDs (21).However, it is not known if the P þ 4 interaction is required forSic1 elimination in vivo, nor how the P þ 4 requirement relatesto the apparent multisite site recognition threshold.
Here we investigate these fundamentally different recognitionmodels and show that the secondary phosphorylation sites in Sic1are dispensable for multisite recognition, that regional context inSic1 can dramatically reduce the interaction affinity of a shorttriphosphopeptide, and that Sic1 recognition and ubiquitinationexhibits cooperative dependence on targeting kinase activityunder equilibrium reaction conditions. The Sic1-Cdc4 multisiterecognition mechanism illustrates how a dynamic protein inter-action mode can engender complex system-level behavior.
ResultsAn Abrupt Phosphorylation-Dependent Transition in the Affinity ofSic1 for Cdc4. The nine weak CPD sequences in Sic1 all containmismatches to an optimal CPD motif (14, 15). Consistently, eachof the seven natural CPD phosphopeptides derived from theNTR bound to a monomeric Skp1-Cdc4 complex with a Ki valueof about 100 μM or greater (Fig. 1A). Pairwise analysis of diphos-phopeptides derived from the pS69/pS76/pS80 serine cluster
Author contributions: X.T., S.O., T.M., V.C., J.D.F.-K., F.S., and M.T. designed research; X.T.,S.O., T.M., and V.C. performed research; X.T., S.O., T.M., and V.C. contributed new reagents/analytic tools; X.T., S.O., T.M., V.C., J.D.F.-K., F.S., and M.T. analyzed data; and X.T., T.P.,J.D.F.-K., F.S., and M.T. wrote the paper.
The authors declare no conflict of interest.
*This Direct Submission article had a prearranged editor.
Data deposition: Crystallography, atomic coordinates, and structure factors have beendeposited in the Protein Data Bank, www.pdb.org (PDB ID code 3V7D).1Present address: Department of Structural Biology, St. Jude Children's Research Hospital,Memphis, TN 38105.
2To whom correspondence should be addressed. E-mail: [email protected], [email protected], or [email protected].
This article contains supporting information online at www.pnas.org/lookup/suppl/doi:10.1073/pnas.1116455109/-/DCSupplemental.
www.pnas.org/cgi/doi/10.1073/pnas.1116455109 PNAS ∣ February 28, 2012 ∣ vol. 109 ∣ no. 9 ∣ 3287–3292
BIOCH
EMISTR
Y
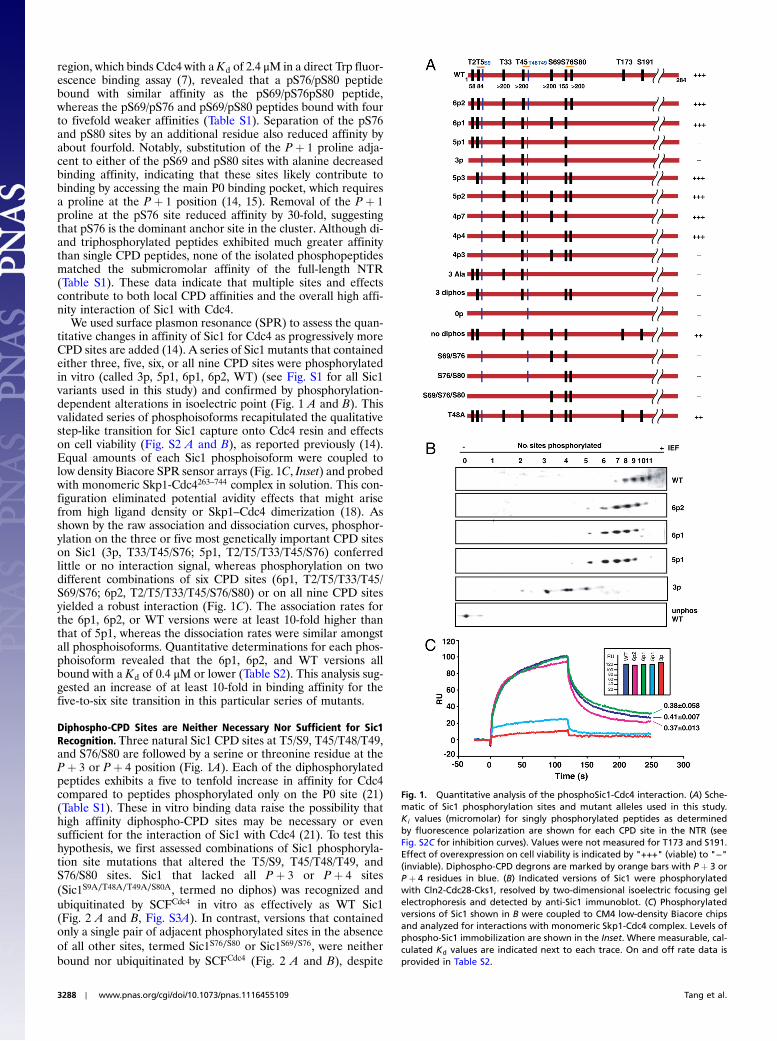
region, which binds Cdc4 with a Kd of 2.4 μM in a direct Trp fluor-escence binding assay (7), revealed that a pS76/pS80 peptidebound with similar affinity as the pS69/pS76pS80 peptide,whereas the pS69/pS76 and pS69/pS80 peptides bound with fourto fivefold weaker affinities (Table S1). Separation of the pS76and pS80 sites by an additional residue also reduced affinity byabout fourfold. Notably, substitution of the P þ 1 proline adja-cent to either of the pS69 and pS80 sites with alanine decreasedbinding affinity, indicating that these sites likely contribute tobinding by accessing the main P0 binding pocket, which requiresa proline at the P þ 1 position (14, 15). Removal of the P þ 1proline at the pS76 site reduced affinity by 30-fold, suggestingthat pS76 is the dominant anchor site in the cluster. Although di-and triphosphorylated peptides exhibited much greater affinitythan single CPD peptides, none of the isolated phosphopeptidesmatched the submicromolar affinity of the full-length NTR(Table S1). These data indicate that multiple sites and effectscontribute to both local CPD affinities and the overall high affi-nity interaction of Sic1 with Cdc4.
We used surface plasmon resonance (SPR) to assess the quan-titative changes in affinity of Sic1 for Cdc4 as progressively moreCPD sites are added (14). A series of Sic1 mutants that containedeither three, five, six, or all nine CPD sites were phosphorylatedin vitro (called 3p, 5p1, 6p1, 6p2, WT) (see Fig. S1 for all Sic1variants used in this study) and confirmed by phosphorylation-dependent alterations in isoelectric point (Fig. 1 A and B). Thisvalidated series of phosphoisoforms recapitulated the qualitativestep-like transition for Sic1 capture onto Cdc4 resin and effectson cell viability (Fig. S2 A and B), as reported previously (14).Equal amounts of each Sic1 phosphoisoform were coupled tolow density Biacore SPR sensor arrays (Fig. 1C, Inset) and probedwith monomeric Skp1-Cdc4263–744 complex in solution. This con-figuration eliminated potential avidity effects that might arisefrom high ligand density or Skp1–Cdc4 dimerization (18). Asshown by the raw association and dissociation curves, phosphor-ylation on the three or five most genetically important CPD siteson Sic1 (3p, T33/T45/S76; 5p1, T2/T5/T33/T45/S76) conferredlittle or no interaction signal, whereas phosphorylation on twodifferent combinations of six CPD sites (6p1, T2/T5/T33/T45/S69/S76; 6p2, T2/T5/T33/T45/S76/S80) or on all nine CPD sitesyielded a robust interaction (Fig. 1C). The association rates forthe 6p1, 6p2, or WT versions were at least 10-fold higher thanthat of 5p1, whereas the dissociation rates were similar amongstall phosphoisoforms. Quantitative determinations for each phos-phoisoform revealed that the 6p1, 6p2, and WT versions allbound with a Kd of 0.4 μM or lower (Table S2). This analysis sug-gested an increase of at least 10-fold in binding affinity for thefive-to-six site transition in this particular series of mutants.
Diphospho-CPD Sites are Neither Necessary Nor Sufficient for Sic1Recognition. Three natural Sic1 CPD sites at T5/S9, T45/T48/T49,and S76/S80 are followed by a serine or threonine residue at theP þ 3 or P þ 4 position (Fig. 1A). Each of the diphosphorylatedpeptides exhibits a five to tenfold increase in affinity for Cdc4compared to peptides phosphorylated only on the P0 site (21)(Table S1). These in vitro binding data raise the possibility thathigh affinity diphospho-CPD sites may be necessary or evensufficient for the interaction of Sic1 with Cdc4 (21). To test thishypothesis, we first assessed combinations of Sic1 phosphoryla-tion site mutations that altered the T5/S9, T45/T48/T49, andS76/S80 sites. Sic1 that lacked all P þ 3 or P þ 4 sites(Sic1S9A∕T48A∕T49A∕S80A, termed no diphos) was recognized andubiquitinated by SCFCdc4 in vitro as effectively as WT Sic1(Fig. 2 A and B, Fig. S3A). In contrast, versions that containedonly a single pair of adjacent phosphorylated sites in the absenceof all other sites, termed Sic1S76∕S80 or Sic1S69∕S76, were neitherbound nor ubiquitinated by SCFCdc4 (Fig. 2 A and B), despite
Fig. 1. Quantitative analysis of the phosphoSic1-Cdc4 interaction. (A) Sche-matic of Sic1 phosphorylation sites and mutant alleles used in this study.Ki values (micromolar) for singly phosphorylated peptides as determinedby fluorescence polarization are shown for each CPD site in the NTR (seeFig. S2C for inhibition curves). Values were not measured for T173 and S191.Effect of overexpression on cell viability is indicated by "+++" (viable) to "−"(inviable). Diphospho-CPD degrons are marked by orange bars with P þ 3 orP þ 4 residues in blue. (B) Indicated versions of Sic1 were phosphorylatedwith Cln2-Cdc28-Cks1, resolved by two-dimensional isoelectric focusing gelelectrophoresis and detected by anti-Sic1 immunoblot. (C) Phosphorylatedversions of Sic1 shown in B were coupled to CM4 low-density Biacore chipsand analyzed for interactions with monomeric Skp1-Cdc4 complex. Levels ofphospho-Sic1 immobilization are shown in the Inset. Where measurable, cal-culated Kd values are indicated next to each trace. On and off rate data isprovided in Table S2.
3288 ∣ www.pnas.org/cgi/doi/10.1073/pnas.1116455109 Tang et al.

the moderately high affinity of the isolated diphosphorylatedpeptides for Cdc4 (21) (Table S1).
We then determined if the diphospho-CPD sites influencedelimination of Sic1 in vivo. Overexpression of partially stabilizedforms of Sic1 causes a lethal G1 phase arrest, which is a sensitivereadout for Sic1 elimination in vivo (14). Strains that over-expressed the SIC1S9A, SIC1T49A, or SIC1S9A/S80A alleles were in-distinguishable from WT (Fig. S3B), whereas overexpression ofeither SIC1T48A or SIC1S9A/T48A/S80A (no diphos) caused a mildgrowth defect (Fig. 2C, Fig. S3B). Conversely, SIC1 alleles thatencoded only the three diphospho-CPD sites, the high affinityserine cluster (S69/S76/S80) alone, or that lacked only the serinecluster (3 Ala) caused inviability when overexpressed, similar to acontrol that lacked all single CPD sites (Fig. 2 C and D). Theseeffects on viability were paralleled by effects on Sic1 stability invivo (Fig. S3C). Addition of the T45 site to the serine cluster(4p3) did not rescue viability, whereas further addition of thelow affinity T33 site (5p2) restored growth (Fig. 2D). Surprisingly,removal of the T2/T5 diphospho-CPD from the 6p1 and 6p2alleles (T33/T45/S69/S76 and T33/T45/S76/S80, termed 4p7 and4p4, respectively) actually improved viability, whereas the mini-mal T33/T45/S76 allele (3p) caused lethality (Fig. 2D, Fig. S2A).These data demonstrate that the originally inferred six site re-quirement for Sic1 recognition is not absolute, but depends onthe particular combination of four or more sites. Although thepT2/pT5 diphospho-CPD site binds to Cdc4 with moderateaffinity in vitro (Table S1), these sites may antagonize Sic1 degra-dation in vivo. Overall, we conclude that diphospho-CPD motifsare not required for the recognition and degradation of Sic1 invivo, despite the moderately high affinity interactions betweendiphospho-CPD peptides and Cdc4 in vitro.
The P þ 4 Binding Surface is Dispensable but Adjusts the Phosphoryla-tion Threshold. To ascertain that the diphospho-CPD bindingmode of yeast Cdc4 is analogous to that of human Fbw7 (21),
we determined the structure of a pS69/pS76/pS80 20-mer CPDtriphosphopeptide bound to Skp1-Cdc4263–744 at 2.3-Å resolution(Table S3). The pS76 and pS80 sites and adjacent residues werevisible in the structure (Fig. S4A). The core CPD sequence G73-L-T-pS-P77 engaged the primary P0 pocket in a similar manner asin the Cdc4-CPDcyclinE structure (15), but with weaker interac-tions of the nonoptimal T75 residue in the important hydropho-bic P-1 pocket. Notably, pS80 engaged the positively chargedsurface adjacent to the primary P0 binding pocket, with hydrogenbond interactions between the phosphate and residues R443,S464, T465, and R485 (Fig. S4 A and B). The Cdc4/Fbw7 residuesthat form the P þ 4 engagement surface are highly conserved,although the fission yeast homologue Pop2 contains isoleucineinstead of threonine at position 465, and may exhibit less favor-able interactions with the P þ 4 phosphate. The Sic1 diphospho-CPD interaction with Cdc4 thus closely resembles the humancyclin E diphospho-CPD interaction with Fbw7.
To evaluate the role of the P þ 4 site in Sic1 recognition, wemutated the three nonessential residues in Cdc4 that engage theP þ 4 phosphate (R443A, S464A, T465I), either alone or in com-bination. Each Cdc4 mutant bound to a singly phosphorylatedCPD peptide as well as WT Cdc4 but had reduced affinity for adiphospho-CPD peptide (Fig. 3A), as shown previously for simi-lar mutations in Fbw7 (21). Despite this defect in CPD recogni-tion in vitro, each CDC4 allele, including the CDC4AAI triplemutant, complemented CDC4 function in vivo (Fig. 3B). TheCDC4AAI strain had normal cell morphology, but exhibited a slightaccumulation of cells with 1N DNA content (Fig. 3C). These re-sults demonstrate that the P þ 4 binding surface is not essentialfor Cdc4 activity in vivo.
We then assessed the performance of the CDC4AAI allele in theface of excess SIC1 dosage in vivo. To titrate CDC4 capacity, aseries of SIC1 phosphorylation site mutants expressed from theGAL1 promoter were evaluated for effects on viability. WhereasWT CDC4 allowed cells to cope with overexpression of versionsof SIC1 that contained six or more phosphorylation sites, theCDC4AAI strain was unable to cope even with the loss of the weakCPD sites at T173 and S191 (Fig. 3D). As a control, a version ofSIC1 that was targeted solely by a high affinity singly phosphory-lated CPD derived from cyclin E, termed SIC1CPD (14), did notcompromise growth of the CDC4AAI strain. From these genetictitration experiments, we conclude that the P þ 4 pocket servesto tune the number of CPD sites needed for efficient Sic1 elim-ination, and that the weak CPD sites at T173 and S191 contributeto Sic1 recognition in vivo.
Effect of Regional Sequence Context on CPD Interaction Affinities.Weused NMR to examine interactions of the isolated WD40 domainof Cdc4 (Cdc4WD40) with each CPD site in N15-labeled phos-phorylated full-length Sic1. Resonances of each phosphorylatedresidue have a pronounced downfield chemical shift due to theelectronegative influence of the phosphate (Fig. 4A). The titra-tion of Cdc4WD40 against full-length Sic1 caused a similar degreeof resonance broadening for each CPD site in the NTR (Fig. 4B,Fig. S5), as previously observed (16). Additional resonances cor-responding to T173 and S191, which lie outside of the NTR andimmediately proximal to the CDK inhibitory domain of Sic1,were also broadened by Cdc4WD40 (Fig. 4B), consistent with thegenetic effects of T173 and S191 (Fig. 3D). This NMR analysisdemonstrates that the dynamic equilibrium engagement of multi-ple CPD sites occurs in the context of full-length Sic1.
Despite the relatively high affinity interaction of the isolatedpS69/pS76/pS80 triphosphopeptide with Cdc4 (7), this extendeddegron was insufficient for Sic1 elimination in vivo (Fig. 2C).We speculated that the inherent affinity of the serine cluster de-gron might be reduced in its natural regional context. We used anintrinsic Trp fluorescence binding assay to directly compare theaffinity of the pS69/pS76/pS80 triphosphopeptide to that of
A
DC
B
bound
input
WT
S76/S
80
Sic1-
no dip
hos
S69/S
76
- (Ub)n
2 3 4 5 6 7 8 9 10
S76/S80
S69/S76
ladder
WT
ladder
0 1 2 3 4 5 6
unphosS76/S80
no diphos
No. sites phosphorylated + IEF
2 3 4 5
0 1 2 3 4 5 7
- +
diphos
WT
0p0p 3 diphos
WT
T48A
no diphos
3 diphos
4p3
5p2
WT
S69/S76/S80
3 Ala
WT
5p3
4p7
6p2
4p4
Sic1
Fig. 2. Interaction of Sic1 with SCFCdc4 does not depend on P þ 4 phosphor-ylation sites. (A) The indicated Sic1 proteins were phosphorylated and eithercaptured onto Cdc4FLAG resin or incubated with complete SCFCdc4 reactionmixture and detected by anti-Sic1 immunoblot. (B) Input Sic1 phosphoiso-forms were detected by two-dimensional isoelectric focusing gel electrophor-esis followed by Coomassie blue stain. Partial Sic1 phosphorylation reactionswere used to generate a marker ladder. (C, D) Strains bearing the indicatedGAL1-HASIC1 (C) or GAL1-SIC1 (D) alleles were grown for 2 d on 2%galactose medium at 30 °C.
Tang et al. PNAS ∣ February 28, 2012 ∣ vol. 109 ∣ no. 9 ∣ 3289
BIOCH
EMISTR
Y

full-length Sic1 that was phosphorylated only on the same threeresidues (Sic1S69∕S76∕S80). This comparison revealed that althoughthe isolated peptide had a Kd of 2.4 μM for Cdc4, the affinity ofthe serine cluster degron was reduced by at least 25-fold, to anestimated Kd > 50 μM, when placed in the context of full-lengthSic1 (Fig. 4C). The constraint on local degron affinity imposed bythe NTR reveals the important role of regional context in disor-dered protein interactions.
Ultrasensitive Dependence of Sic1 Recognition on CDK Activity. Themultisite phosphorylation threshold should render Sic1 recogni-tion and ubiquitination ultrasensitive with respect to CDK kinaseactivity, provided that phosphorylation is distributive and that acompeting phosphatase back-reaction establishes an equilibriumdistribution of phosphorylation states (3). We developed an invitro recombinant system for equilibrium phosphorylation of Sic1by titrating input Cln2-Cdc28-Cks1 against a fixed level of Cdc14,which is a physiological phosphatase for Sic1 (22). Phosphoryla-tion of full-length untagged Sic1 by Cln2-Cdc28-Cks1 in thesereactions was distributive (Fig. S6A) and equilibrium levels ofphosphorylation were obtained over prolonged incubation peri-ods in the presence of Cdc14 (Fig. S6 B and C), provided a con-taminating insect phosphatase was inactivated with okadaic acid(Fig. S6D). The dependence of Sic1 phosphoisoform distribu-tions on input kinase concentration was confirmed by isoelectricfocusing, although the distribution balance varied in differentkinase preparations (compare Fig. 5A to Fig. S6C). Sic1 fromeach phosphorylation reaction was captured on monomericSCF-Cdc4Flag resin and quantitated by immunoblot. This analysisrevealed a step transition in the Sic1-Cdc4 interaction from neg-ligible to near maximal levels over a mere twofold increase inCDK concentration, with a calculated Hill coefficient (nH) of 4.8(Fig. 5C, Table S4). As a control, we carried out an analogous
experiment with Sic1CPD, which requires only a single phosphor-ylation event for efficient capture by Cdc4 (14). Despite somenonspecific binding in the absence of phosphorylation in theseexperiments, the Sic1CPD-Cdc4 interaction exhibited a moregraded dependence on CDK activity with an estimated nH of 1.8(Fig. 5 B and C). We then subjected a series of Sic1 phosphor-ylation reactions to a simultaneous SCFCdc4 ubiquitination reac-tion over a prolonged incubation period. Conversion of Sic1 tohighly ubiquitinated species also exhibited a strongly cooperativedependence on input CDK concentration (Fig. 5D). This steptransition in ubiquitination was manifest despite the opportu-nity for transiently phosphorylated forms of Sic1 to become ubi-quitinated by SCFCdc4 and trapped as ubiquitinated species atlower kinase concentrations. These data demonstrate that Sic1recognition and ubiquitination by SCFCdc4 exhibits ultrasensitivedependence on the targeting kinase under the equilibrium reac-tion conditions expected to exist in vivo.
DiscussionOur biochemical, genetic and structural data suggest that themultiple weak CPD sites in Sic1 engage Cdc4 in rapid dynamicequilibrium. This binding mode depends on manifold variablesincluding the number, spacing, and individual affinities of differ-ent CPD sites, decoy sites, and regional sequence context effects.These variables allow the recognition threshold to be tuned in asubstrate-specific fashion, such that interactions can be dictatedprimarily by high affinity diphospho-CPD sites (19–21, 23), or bya combination of individually weak CPD sites, such as in Sic1 andother proteins (24, 25). This type of dynamic interaction contrastsstarkly with static engagement models that are typically invokedto explain regulated protein interactions (26). In this dynamicview, the X-ray structure of Cdc4 in complex with the pS76 andpS80 sites represents only a snapshot of one of many possible
Fig. 3. The P þ 4 binding site on Cdc4 is dis-pensable in vivo but helps establish the pre-cise phosphorylation threshold for Sic1recognition. (A) Binding affinities of singlyand doubly phosphorylated cyclin E-derivedpeptides with wild type and P þ 4 bindingsurface mutants of Cdc4. Values are themean of three independent experiments�SEM. (B) The indicated P þ 4 binding surfacealleles on a hCDC4 TRP1 CENi plasmid wereintroduced into a cdc4ΔHIS3 hCDC4 URA3CENi strain, streaked on either 5-fluorooro-tic acid medium to select against the WT al-lele or, as a control, on -Trp or -Ura mediumand grown at 30 °C for 3 d. (C) WT andCDC4S464AT465IR443A strains were assessed forDNA content by FACS analysis (Upper) andmicroscopy (Lower) at early log phase in richglucose medium. Approximately 1 and 4%of the cell population had an elongated-bud phenotype in WT and CDC4S464AT465IR443A
strains, respectively. (D) Serial dilutions ofWT or Cdc4S464AT465IR443A strains bearing the in-dicated GAL1-SIC1 alleles were spotted onglucose or 0.5% galactose medium and incu-bated at 30 °C for 3 d. Schematic showsphosphorylation sites in each allele; Insetshows sequence surrounding the high affi-nity CPD site.
3290 ∣ www.pnas.org/cgi/doi/10.1073/pnas.1116455109 Tang et al.

binding modes of pSic1. We speculate that the rapidly fluctuatingstructure of Sic1 weakens the interactions of individual sites, suchas the triphosphorylated serine cluster, in part through net repul-sive electrostatic interactions (7). Two additional points areworthy of emphasis. First, although Sic1 is a highly disorderedprotein, it nevertheless exhibits a surprisingly compact structure(17), which may contribute to its dynamic interactions with Cdc4through electrostatic averaging effects (7). Second, although theSCFCdc4 holocomplex is a dimer in solution, the affinity of Cdc4for Sic1 does not depend on dimerization, which precludes amodel based on simultaneous engagement of two spatially sepa-rated binding pockets (18).
Multisite Phosphorylation and Switch-Like Biological Responses. En-zyme activation or protein interactions that depend on phosphor-
ylation of more than one site generate a cooperative responsecurve (3). Multisite phosphorylation can also lead to ultrasen-sitive responses through preferred decoy sites, which serve toincrease the concentration of kinase needed to phosphorylatecritical effector sites of lower affinity (5). In a variation on thisphenomenon, the T2/T5 sites in Sic1 may act as decoys that con-tribute to binding but adversely affect a downstream step in theelimination of Sic1. Multiple phosphorylation events can alsoengender switch-like effects by causing transitions in electrostaticpotential. Charge neutralization of the highly basic NTR by theaddition of multiple phosphates has been posited to help estab-lish the threshold for Sic1 recognition (7). Similarly, multisitephosphorylation of the Ste5 scaffold protein cooperatively dis-rupts its membrane localization through electrostatic repulsionbetween the multiple phosphates and the negatively chargedphospholipid membrane surface (6). Finally, the biochemicalphenomenon of zero-order ultrasensitivity, first described for me-tabolic enzymes (2), can also operate in signaling complexeswhere the local concentration of phosphorylation sites exceedsthe catalytic capacity of the kinase (8). These different typesof multisite dependencies all serve to enable cellular decisionmaking processes. Although models based on entropic, electro-static, and/or kinetic considerations have been proposed for theSic1-Cdc4 interaction (7, 27–29), the physicochemical basis forthe precise multisite threshold remains to be determined.
Fig. 4. Dynamics of full-length Sic1-Cdc4WD40 interaction and regional con-text effects on CPD interaction affinity. (A) NMR 1HN-15N correlation spectraof phosphorylated residues of full-length Sic1 in the absence (black) and pre-sence (red) of Cdc4WD40. Residues with multiple resonances from cis- andtrans-prolyl isomers are denoted with a prime. (B) Intensity ratios of amideresonances of full-length Sic1 upon titration with unlabeled Cdc4WD40 atpSic1∶Cdc4WD40 molar ratios of 1∶0, 1∶0.1, 1∶0.25, 1∶0.5, 1∶0.75, and1∶1.2, represented by lines of increasing darkness. Red stars indicate phos-phorylation sites; red bar indicates serine cluster degron. Intensity ratios ofamide resonances of residues T173 and S191 are reported separately. (C) Ex-perimental binding curves for an isolated pS69/pS76/pS80 triphosphopeptide(blue) and phosphorylated full-length Sic1S69∕S76∕S80 (red) compared to theo-retical fluorescence binding curves at 500 nM protein concentration for theindicated affinity constants. As the titration of Skp1-Cdc4263–744 with phos-pho-Sic1S69∕S76∕S80 did not reach saturation, a numerical Kd could not be cal-culated. Data represent two independent experiments.
Sic1
Sic1-(Ub)n
Cdc14Flag
DCkinase units
Cdc4Flag
(SCF)0 2 4 6 8 10 12 14 16 18
5 CPD, nH = 1.8
WT, nH = 4.8
35
30
25
20
15
10
Kinase units
Sig
nal
inte
nsi
ty
Sic1 WT
unbound
1 2 4 8 16 1 2 4
bound
0 8 16
kinase units
Sic1CPD
B
0 0.25 1 2 4 8 16
A No. sites phosphorylated +- IEF
4
0
1
kin
ase
un
its
0 1 2 3 4 5 6 7 8 9
Fig. 5. Dependence of Sic1 recruitment and ubiquitination by SCFCdc4 onCDK activity under equilibrium reaction conditions. (A) Sic1 or Sic1CPD wereincubated with the indicated arbitrary units of Cln2-Cdc28-Cks1 kinase in thepresence of fixed Cdc14 phosphatase and SCFCdc4 resin at 30 °C for 60 min.Phospho-Sic1 species for the indicated reactions were resolved by two-dimensional isoelectric focusing gel electrophoresis and Coomassie bluestain. Minor Sic1 species detected between phosphoisoforms are due to par-tial carbamoylation during sample preparation. (B) Capture of Sic1 or Sic1CPD
from reactions in A onto Cdc4 resin and detection by anti-Sic1 immunoblot(Upper) or Coomassie blue stain (Lower). (C) Quantitation of Sic1 or Sic1CPD
captured onto Cdc4 and estimation of Hill coefficients (nH). Plots are averageof duplicate experiments. (D) Ubiquitination of Sic1 under equilibrium phos-phorylation conditions. A similar but independent experiment as in A wasperformed except that components for SCFCdc4-dependent ubiquitinationwere included in the reaction and incubated at 30 °C for 60 min.
Tang et al. PNAS ∣ February 28, 2012 ∣ vol. 109 ∣ no. 9 ∣ 3291
BIOCH
EMISTR
Y

Intrinsically Disordered Regions and Protein Recognition. Proteinswith disordered regions often lie at the nexus of cellular regula-tory networks (26, 30). Disordered regions can interact viacoupled folding and binding, via simple modular recognition ofshort linear sequence motifs or, as shown here, via low affinitylinear motifs that interact in rapid equilibrium. The spectrumof potential binding modes afforded by disordered regions is par-ticularly suited to control by phosphorylation, which has a strongpropensity to occur in such regions (31). The rapid equilibriummode for multisite engagement is readily tuned to different bio-logical thresholds and also inherently able to integrate differentsignals through phosphorylation of different sites. Many disor-dered regions harbor multiple short linear motifs that mediatemoderate to low affinity interactions (30, 32, 33). The elucidationof the biophysical mechanisms that underlie the combinatorialweak interactions of linear motifs in disordered proteins withtheir receptors will be critical for understanding how the cell con-structs precise thresholds in various biological responses (3, 26).
Materials and MethodsYeast Culture. Strains and plasmids used in this study are listed in Tables S5 andS6. Methods for cell culture, genetic manipulation, protein extraction, immu-noblot, and immunoprecipitation were carried out as described (15, 18).
Binding and Ubiquitination Assays. Recombinant proteins were produced,phosphorylated, and dephosphorylated as described (15, 18, 22). Sic1 wasphosphorylated by Cln2-Cdc28-Cks1 for 1 h at 30 °C in kinase buffer [50 mMHepes (pH 7.2), 55 mM NaCl, 1 mM ATP, 10 mM MgCl2, 1 mM DTT]. For equi-librium binding, Sic1, FLAGCdc4, and Cdc14 were mixed before addition ofkinase. For ubiquitination, reactions were initiated by adding Cln2-Cdc28-Cks1 to premixed Sic1, Cdc14, human E1, Cdc34, SCFCdc4, and ubiquitin inkinase buffer. For SPR assays, purified Sic1 isoforms bearing a C-terminalcysteine were phosphorylated then diluted approximately 10-fold with im-mobilization buffer (10 mM NaOAc, pH 5.0) to 25 μg∕mL and immobilizedto a CM4 sensor chip (GE Healthcare) at 5 μL∕min using thiol coupling toa level of approximately 100 RU. Analyte Skp1-Cdc4263–744 monomeric com-
plex was equilibrated in running buffer [10mMHepes (pH 7.3), 150 mMNaCl,3 mM EDTA, 0.005% Surfactant P2] and injected at between 29 and 1,860 nMat 100 μL∕min for 2 min. Following 4 min of dissociation, two 1 min pulses of2 M MgCl2 were applied to regenerate the surface. Sensorgrams were re-corded on a Biacore 3000 and kinetic parameters extracted using BIAevalua-tion Software v3.1. Binding curves were fit with a Langmuir 1∶1 interactionmodel with Chi2 ¼ 2 or better. For fluorescence polarization, unlabeled pep-tides were competed against an N-terminal fluorescein-labeled cyclin Epeptide (GLLpTPPQSG) in 50 mM Hepes (pH 7.5), 50 mM NaCl, 1 mM DTT,and 0.1 mg∕mL BSA (14). Intrinsic tryptophan fluorescence changes uponpeptide addition to 50–500 nM Skp1-Cdc4263–744 were measured as described(7). Immunoblot signal intensity was quantitated on a Fluor-S MultiImagerand curve fitting performed with GraphPad Prism software.
X-ray Structure and NMR Analysis. Skp1-Cdc4263–744 was purified and crystal-lized as previously reported (15). Crystals of the apo form of Skp1-Cdc4263–744
were grown and transferred to a solution containing 100 mM Tris (pH 8.5),1.5 M ammonium sulfate, 15% glycerol, and 1 mM of Ac-GMTpSPFNGLT-pSPQRpSPFPKS-NH2 peptide for 10min prior to freezing. Data were collectedat BioCars 14-MBC at 0.9 Å and the structure solved by molecular replace-ment with 1NEX. For NMR experiments, the WD40 domain of Cdc4 wasexpressed from pET-Sumo (Invitrogen), isolated by Ni-nitrilotriacetate (NTA)affinity chromatography, digested with Ulp protease, and further purified byNi-NTA and Superdex 200 chromatography. NMR titrations of the phosphory-lated NTR and full-length Sic1 with Cdc4WD40 were performed on a VarianInova 500 MHz spectrometer at 5 °C. Assignments for NTR resonances weretransferred to full-length Sic1. T173 and S191 were assigned by mutagenesis,and peak intensities in the heteronuclear single quantum correlation spectrain the presence and absence of Cdc4WD40 were analyzed by FuDA (16).
ACKNOWLEDGMENTS. We thank Lewis Kay, Mikael Borg, and Hue-Sun Chanfor discussions. Supported by grants from the Canadian Cancer SocietyResearch Institute (J.D.F.-K.) and the Canadian Institutes of Health Research(CIHR) (T.P., J.D.F.-K., F.S., and M.T.). Support was provided by the Terry FoxFoundation (T.M.), the CIHR Strategic Training Program in Protein Foldingand Interaction Dynamics (V.C.), and by Canada Research Chairs in StructuralBiology (F.S.) and in Systems and Synthetic Biology (M.T.).
1. Pawson T (1995) Protein modules and signaling networks. Nature 373:573–580.2. Goldbeter A, Koshland DE, Jr (1984) Ultrasensitivity in biochemical systems controlled
by covalent modification. Interplay between zero-order and multistep effects. J BiolChem 259:14441–14447.
3. Ferrell JE, Jr (1996) Tripping the switch fantastic: How a protein kinase cascade canconvert graded inputs into switch-like outputs. Trends Biochem Sci 21:460–466.
4. Ferrell JE, Jr, Machleder EM (1998) The biochemical basis of an all-or-none cell fateswitch in Xenopus oocytes. Science 280:895–898.
5. Kim SY, Ferrell JE, Jr (2007) Substrate competition as a source of ultrasensitivity in theinactivation of Wee1. Cell 128:1133–1145.
6. Strickfaden SC, et al. (2007) A mechanism for cell-cycle regulation of MAP kinasesignaling in a yeast differentiation pathway. Cell 128:519–531.
7. Borg M, et al. (2007) Polyelectrostatic interactions of disordered ligands suggest aphysical basis for ultrasensitivity. Proc Natl Acad Sci USA 104:9650–9655.
8. MalleshaiahMK, Shahrezaei V, Swain PS, Michnick SW (2010) The scaffold protein Ste5directly controls a switch-like mating decision in yeast. Nature 465:101–105.
9. Bai C, et al. (1996) SKP1 connects cell cycle regulators to the ubiquitin proteolysismachinery through a novel motif, the F-box. Cell 86:263–274.
10. Feldman RM, Correll CC, Kaplan KB, Deshaies RJ (1997) A complex of Cdc4p, Skp1p,and Cdc53p/cullin catalyzes ubiquitination of the phosphorylated CDK inhibitor Sic1p.Cell 91:221–230.
11. Skowyra D, et al. (1997) F-box proteins are receptors that recruit phosphorylatedsubstrates to the SCF ubiquitin-ligase complex. Cell 91:209–219.
12. Petroski MD, Deshaies RJ (2005) Function and regulation of cullin-RING ubiquitinligases. Nat Rev Mol Cell Biol 6:9–20.
13. Verma R, et al. (1997) Phosphorylation of Sic1p by G1 Cdk required for its degradationand entry into S phase. Science 278:455–460.
14. Nash P, et al. (2001) Multisite phosphorylation of a CDK inhibitor sets a threshold forthe onset of DNA replication. Nature 414:514–521.
15. Orlicky S, et al. (2003) Structural basis for phosphodependent substrate selection andorientation by the SCFCdc4 ubiquitin ligase. Cell 112:243–256.
16. Mittag T, et al. (2008) Dynamic equilibrium engagement of a polyvalent ligand with asingle-site receptor. Proc Natl Acad Sci USA 105:17772–17777.
17. Mittag T, et al. (2010) Structure/function implications in a dynamic complex of theintrinsically disordered Sic1 with the Cdc4 subunit of an SCF ubiquitin ligase. Structure18:494–506.
18. Tang X, et al. (2007) Suprafacial orientation of the SCFCdc4 dimer accommodatesmultiple geometries for substrate ubiquitination. Cell 129:1165–1176.
19. Welcker M, Clurman BE (2008) FBW7 ubiquitin ligase: A tumor suppressor at thecrossroads of cell division, growth and differentiation. Nat Rev Cancer 8:83–93.
20. Welcker M, et al. (2003) Multisite phosphorylation by Cdk2 and GSK3 controls cyclin Edegradation. Mol Cell 12:381–392.
21. Hao B, et al. (2007) Structure of a Fbw7-Skp1-cyclin E complex: Multisite-phosphory-lated substrate recognition by SCF ubiquitin ligases. Mol Cell 26:131–143.
22. Visintin R, et al. (1998) The phosphatase Cdc14 triggers mitotic exit by reversal ofCdk-dependent phosphorylation. Mol Cell 2:709–718.
23. Bao MZ, et al. (2010) Multisite phosphorylation of the Saccharomyces cerevisiae fila-mentous growth regulator Tec1 is required for its recognition by the E3 ubiquitin li-gase adaptor Cdc4 and its subsequent destruction in vivo. Eukaryotic Cell 9:31–36.
24. Moberg KH, et al. (2004) The Drosophila F box protein archipelago regulates dMycprotein levels in vivo. Curr Biol 14:965–974.
25. Liu Q, et al. (2011) SCFCdc4 enables mating type switching in yeast by cyclin-dependentkinase-mediated elimination of the Ash1 transcriptional repressor. Mol Cell Biol31:584–598.
26. Boehr DD, Nussinov R, Wright PE (2009) The role of dynamic conformational ensem-bles in biomolecular recognition. Nat Chem Biol 5:789–796.
27. Varedi KS, Ventura AC, Merajver SD, Lin XN (2010) Multisite phosphorylation providesan effective and flexible mechanism for switch-like protein degradation. PLoS One 5:e14029.
28. Lenz P, Swain PS (2006) An entropic mechanism to generate highly cooperative andspecific binding from protein phosphorylations. Curr Biol 16:2150–2155.
29. Klein P, Pawson T, Tyers M (2003) Mathematical modeling suggests cooperativeinteractions between a disordered polyvalent ligand and a single receptor site. CurrBiol 13:1669–1678.
30. Gibson TJ (2009) Cell regulation: Determined to signal discrete cooperation. TrendsBiochem Sci 34:471–482.
31. Iakoucheva LM, et al. (2004) The importance of intrinsic disorder for protein phosphor-ylation. Nucleic Acids Res 32:1037–1049.
32. Mittag T, Kay LE, Forman-Kay JD (2010) Protein dynamics and conformational disorderin molecular recognition. J Mol Recognit 23:105–116.
33. Hazy E, Tompa P (2009) Limitations of induced folding in molecular recognition byintrinsically disordered proteins. ChemPhysChem 10:1415–1419.
3292 ∣ www.pnas.org/cgi/doi/10.1073/pnas.1116455109 Tang et al.





























