Embed Size (px)
Citation preview

c o t 3 p o m HYDRIQUE DE 2 ESPECES D’ARJ3RES PIOHNIERS
< TRIWA HICKANTHA ET GOUPIA G M R 4 )
AU CIIUEG DE LA SAISON S m EN WANE FRANCAISE
D.Y. Alexandre
Orstom, Cayenne
Titre courant : Comportement hydrique - 1
I
l
I
...

,
Le p o t e n t i e l hydrique de base, le p o t e n t i e l hydrique minimum e t la
conductance stomatique maximale ont été mesurés sur deux arbres p ionniers
(Trema micrantha, Goupia glabra) au cours d e s deux pér iodes sèches de 1987,
à Cayenne, Guyane f ranca ise .
Trém:a es t localement spontané t and i s que les goupis proviennent de
sauvageons transplantés. Les p l a n t s des deux espèces ont environ 3 ans au
moment des mesures. a
Les résultats montrent des d i f f é rences de comportement importantes
entre les deux espèces. Tréma maintient un p o t e n t i e l hydrique e t une trans-
p i r a t ion é levés {mais perd une partie de sa sur face f o l i a i r e ) tandis que le
goupi accuse une f o r t e ba i s se de p o t e n t i e l e t d o i t réguler sa t ranspi ra -
t i o n .
Ces d i f f é rences sont en grande p a r t i e liées 8 la morphologie r a c i n a i r e
iles deux espèces.
Mots clés :
hydrique - Conductance stomatique.
Trema micrantha - Goupia glabra -Guyane f r ança i se - Poten t i e l

1, Y
tr'
Water characteristics of
m i c r a n t h ) in French Guiana.
two pimeer tree species (@ìa glatrra and T m
Predawn water po ten t i a l , mini" water p o t e n t i a l and m a x i m u m s tomatal
conductance were measured during t h e two d ry spells i n 1987 a t Cayenne fo r
tuo pioneer tree species : Trema micrantha 'and Goupia glahra.
Trema was spontaneous and Guupia grown from transplanted wide seed-
lings, Both were about 3 years old .
Results show marked di f fe rences between the two species. Trema main-
tains a high water po ten t i a l and t r ansp i r a t ion (but a decreased fol iar
srea) w h i l e Goupia endures a low water p o t e n t i a l and c loses its stomata.
The main reason f o r these d i f f e rences may be sought i n t h e root mor-
phologies.
.

La Guyane f rança ise appa r t i en t à l a zone climatique de la f o r ê t dense
humide pour ra i t se clef i n i r par l 'absence d e con t r a in t e s
hydriques majeures pour la végéta t ion . Ainsi , 3 Cayenne, la pluviométrie
moyenne annuelle e s t proche d e 3 O00 mm, s o i t deux f o i s supér ieure à la de-
mande Gvaporative. A p r i o r i , le f ac t eu r hydrique n ' e s t donc pas c e l u i q u i
cl6termine l e p l u s la vggétation, si ce n ' e s t par son excès.
subéquator ia le q u i
Globalement ce poin t d e vue est peu contes tab le : en revanche, la va-
r i a h i l i t 6 tant in t e r - que intra-annuel le de la p l u v i o s i t é (cf ALEXANDRE,
1938a), les f a c t e u r s s t a t i o n n e l s pédologiques ou biologiques peuvent confé-
rer une grande importance aux petites di f fgrences de comportement d e chaque
espèce. C'est s u r des plantes jeunes e t en milieu ouvert que la con t r a in t e
hydrique peut se révé ler f a c t e u r d e con t r a in t e e t d e d i f f é r e n c i a t i o n . Nous
en donnons i c i une i l l u s t r a t i o n .
Las mesures o n t . été condui tes à Cayenne, s u r un s i te en bordure d e la
rgserve f o r e s t i è r e du Hontabo, sur 2 espèces pionnières bien caractéris-
t iques : Trema micrankha e t Gbupia glabra ( le goupi). Ces deux arbres pré-
s en ten t 1 ' a v a n t k e , sur le plan de l 'expérimentation écophysiologique, de
posséder une a rch i t ec tu re ident ique (modèle d e Roux, HAUE & OLDEMAN, 1970)
e t des f e u i l l e s d e ta i l le e t d e d i spos i t i on semblables.
Les p l a n t s de Treina micrankha son t spontanés e t d a t e n t de f i n 1985,
&que à l aque l l e de jeunes plants d e Goupia g'lL&ra, prélevés en f o r ê t ,
p i s t e de Saint-Elie, ont été repiqués.
Des mesures complémentaires on t été e f f ec tuées s u r d ' au t r e s espèces
hé l ioph i l e s 6galement spontanées, p résentes s u r place (Cecropia palmata,
Dytiimopru7,a.x momtotoni, Solanum ,cririn,mense. . . >. Toutes ces mesures ont
apporté des r é s u l t a t s très proches d e ceux obtenus avec Trema micrankha e t
I
'i'

il n’en sera p l u s f a i t & ta t i c i . Quelques mesures ont également été
conduites bmr Eperua grcmdiflora (ou Napa). Il s’agissait d e b r i n s i s s u s
d’un semis sur place en j anv ie r 1985. Cette p l an te est un grand a r b r e fo-
r e s t i e r dont l ‘écologie a f a i t l ’objet de travaux récents (FORGET, 1988).
La pet i te ta i l le des quelques b r i n s surv ivants d e cette espèce, i c i bien en
dehors de son h a b i t a t normal,. expl ique le pe t i t nombre de mesures effec-
t.i.ities, malgré l ’ i n t g r ë t qu’el les au ra i en t pu présenter .
11 f a u t noter que ¿;oupia g1abr.a est totalement absent d e l ’ î l e de
Cayenne e t d e la rgserve f o r e s t i e r e du Montabo en p a r t i c u l i e r , mais devien t
t r g s abondant dès sa pér iphér ie . Nous en avons a i n s i rencontré e t é t u d i é
d‘importantes populations dans l e perimètre O.N.F . d e Macouria ainsi que
Chemin Mogès à Stoupan, à une quinzaine d e kms d e Cayenne. Les caractsres
ecophysiologiques du goupi peuvent d i f f é r e r de façon sens ib l e se lon le si te
de croissance e t il importe donc d e soul igner que tous les paramètres rap-
por tés c i -dessous proviennent du s i te d e Cayenne.
. mesures e f fec tuées ont porté sur des paramètres très “c la s s iques” .
Ln situ, nous avons mesuré l e poten t ie1 .de base, l e po ten t i e l m i n i m u m e t la
conductance mqimale. Ces mesures ont été complétées au l abora to i r e par des
exp6riences conduisant à la courbe ”pression-volume”, au d e f i c i t s u b l é t a l
ou à La c iné t ique d e t r ansp i r a t ion de rameaux coupés.
Les
a) Hesures de potentiel
Le p o t e n t i e l hydrique a été mesuré à l ’ a ide d’une chambre à press ion
PMS. Quand les stomates son t fermés, la mesure ne pose pas de problèmes
( p o t e n t i e l d e Gase ou po ten t i e l minimum d e p l an te s s t r e s s é e s ) . Quand les
plantes t r ansp i r en t activement, le p o t e n t i e l s ‘ aba i s se très rapidement au
cours de la mesure ju squ’à la fermeture hydroactive des stomates. Afin
d‘obtenir malgré t o u t une est imat ion du p o t e n t i e l ìn vivo, on a e f f ec tué

i,
I
Les mesures d e façon très rapide e t chronométrée s u r d e s couples de ra-
neaux. En cor ré l an t la d i f fSrence de po ten t i e l observée avec l a d i f fé rence
de cilAr& de mesure, on o b t i e n t un f ac t eu r d e cor rec t ion g r o s s i e r q u i permet
a i n s i l ' e s t imat ion du p o t e n t i e l in vivo.
Sur goupi e t tréma, nous avons u t i l i s é pour les mesures des extrémités
CIF! rameaux comportant 3 ou 4 f e u i l l e s . Pour Eperua gmncl i f lora , on a
u t i l i s é la partie terminale d e la f e u i l l e avec 2 paires d e f o l i o l e s .
Les Inesures d e po ten t i e l de base s ' e f f e c t u e n t en f i n d e n u i t , avant l e
!?ver du ,jour. Leur i n t é r ê t a été souligné par AUSSENAC, 1985.
Les mesures d e p o t e n t i e l minimum ont été ef fec tuées ve r s 13 h, heure
loca le (= heure s o l a i r e ) .
b) Conductances storratiques naxinal e s .
Les mesures d e conductances stomatiques ont ét6 e f f ec tuées à l'aide du
pnrcrmètre à di f fus ion MK3 d e Delta-T Devices. Compte tenu de l 'humidi té
m b i a n t e , d!e mesure u t i l i s é e est celle q u i f a i t passer l 'humidi té
r e l a t i v e de l a chambre de O , 65 à O, 75.
l a gamme
Les mesures son t e f f ec tuées s u r la 4ème f e u i l l e en partant d e
l ' ex t r émi t é des rameaux. En e f f e t , en période de croissance, c'est souvent
la f e u i l l e d e ce rang q u i montre la conductance la p lus élevée e t la var ia -
b i l i t 6 la p lus f a i b l e ( f i g . 1). Sur chaque p l an te on mesure la conduct ivi té
de 4 f e u i l l e s s i t u é e s aux 4 p o i n t s cardinaux, à mi-hauteur du f eu i l l age .
Chmpte tenu d e l a v a r i a b i l i t é des mesures e t d e la préc is ion poss ib le ,
ún a mesuré la conduct ivi té s u r 4 individus pour l e goupi, 3 pour l e trema
e t 2 pour Eper~ia gmndi f lura . Tous les a rb res rencontrés ont des feuil les
a u s s i les mesures d e conductance ont -e l les été limitées à ' hjrpostomatiques,
la f ace abaxiale .
7

Sauf pour les valeurs très Blevées ou tres f a i b l e s de conductance oÙ
l e s problèmes techniques ne sont pas reso lus , la r ep roduc t ib i l i t é des me-
sures est tou t à f a i t s a t i s f a i s a n t e .
En saison sèche, chez un grand nombre d'espèces, l a conductance aug-
mente rapidement l e matin p u i s ba isse ensu i t e progressivement au cours de I
la. journke (non publ ié) . Afin d'est imer la conductance maximale, les me-
Sures an t donc 6t8 effectuées , dans la mesure du poss ib le , en m i l i e u de ma-
t.in6e.
c) Autres nesures
Les courbes pression-volume, les d é f i c i t s sublétaux e t les mesures d e
t ranspi ra t ion de rameaux coupés ont été obtenus avec une balance Mettler
glectronique sens ib le au 1 mg. Ce type de matériel permet un gain de temps
inestimable mais ne supporte pas les va r i a t ions rapides d'HR e t de tempéra-
ture d u t e r r a i n . Toutes les pesées ont donc dû être effectuées au la-
boratoire .
6 80°C. Pour le d é f i c i t sublétsrl (D.S.L.), les temps de ressa tura t ion adop-
i
L a sa tura t ion a été ef fec tuée Ei. l ' obscu r i t é e t 7°C e t le séchage $
t& après essais sont de 6h et 24h.
Le tréma et l e goupi ne posstkient pas de gomme ou de r é s ine suscep-
t i b l e s de boucher les vaisseaux e t de gêner la ressa tura t ion du matériel.
En revanche, chez l e Wapa, ce genre de problème peut éventuellement se pro-
dui re comme en témoignent quelques apparentes sursa tura t ions avec du maté-
r ie l b lessé . Une grande prudence reste de mise quant à l ' i n t e r p r é t a t i o n des
c h i f f r e s du Wapa.
La pluviométrie est mesurGe avec un pluviomètre standard relevé quo-
tidiennement a 18 h.
f

L’année 1987 a S t é par t icul i6rement “sèche“, avec s u r t o u t un “ p e t i t
de comportement e n t r e l e goupi e t le tréma se re t rouve l o r s des deux épi-
sodes secs d e l‘année 1987. Cependant des d i f f é rences appara issent quant à
l a reprise d ’ a c t i v i t k normale au re tour des p lu i e s . Lors du p e t i t été d e ,
eté de mars“ très accusé. La f a i b l e s s e de l a p luv ios i t é a des répercussions
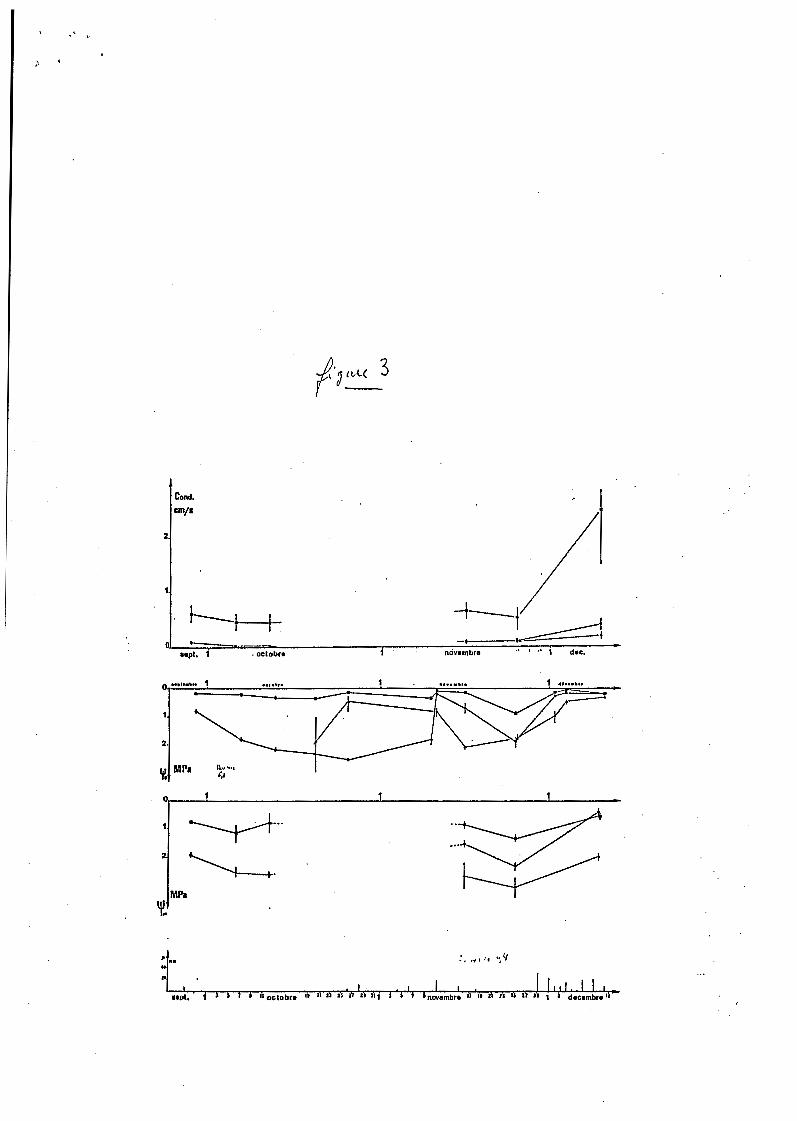
manifestes s u r la végétat ion comme on l e v o i t s u r les f i g u r e s 2 e t 3.
Le comportement du tréma e t c e l u i du goupi sont bien d i f f 6 r e n t s . Tréma
(::onserve un p o t e n t i e l de base proche d e zéro e t une t r ansp i r a t ion élevée
contrairement au goupi q u i v o i t son p o t e n t i e l diminuer fortement et qu i
f i n i t par fermer compl6tement ses stomates. L e comportement du wapa semble
i n t e r r i d i a i r e e n t r e c e l u i du trema e t du goupi, ce q u i peut paraître
paradoxal si l’on considère q u ‘ i l y a un gradien t a l l a n t ’ d e trema, espèce
tres hél iophi le , au wapa s c i a p h i l e en passant par l e goupi. La d i f fe rence
mars où la sécheresse du mil ieu n ’ e s t que relative, la reprise de transpi-
r a t ion ou la remontée des potentiels sont très rapides , par t icul ièrement
dans le cas du tréma. Au c o n t r a i r e , au caurs d e la sa ison sèche d’août à
dGcembre, l e d é f i c i t hydrique est considérable e t les a r b r e s ne reprennent
que lentement l eu r p l e ine activité t r a n s p i r a t o i r e . En f i n d e saison sèche,
[:in cons ta te que le p o t e n t i e l du wapa descend au-dessous d e c e l u i du goupi
e t que sa reprise est par t icul ièrement l en te . La plante semble touchée dans
ses mécanismes in te rnes a l o r s que lo r s de l‘épiscde précédent, e l l e avait
pu en p le ine période “sèche“, émettre des . f e u i l l e s neuves t raduisant ainsi
un bon 6 t a t de s a n t é , Les tests d e labora to i re vont confirmer cette d i f f é -
rence de s e n s i b i l i t é à la sécheresse.
Considerons tou t d’abord les teneurs en matière sèche des f e u i l l e s
les ca lcu le r à p a r t i r des nombreux échan t i l l ons prélevés te l les qu’on peut
pour les d ive r ses expériences. C e paramètre vaut en moyenne 31,O % chez
Y

TANNER, 1985 ; RYCHNOVSKA, 1963). C'est su r tou t un ind ice relativement fa-
c i l e à obteni r avec des moyens de l abora to i r e élémentaires, a tou t d é t e m i -
n m t i c i . L ' i n t e rp ré t a t ion des va leurs obtenues reste assez d é l i c a t e et
trema, 33,l % chez le goupi e t 41,O chez le wapa. Ces d i f fkrences e n t r e
c h i f f r e s moyens sont s ta t is t iquement hautement s i ,gn i f ica t ives .
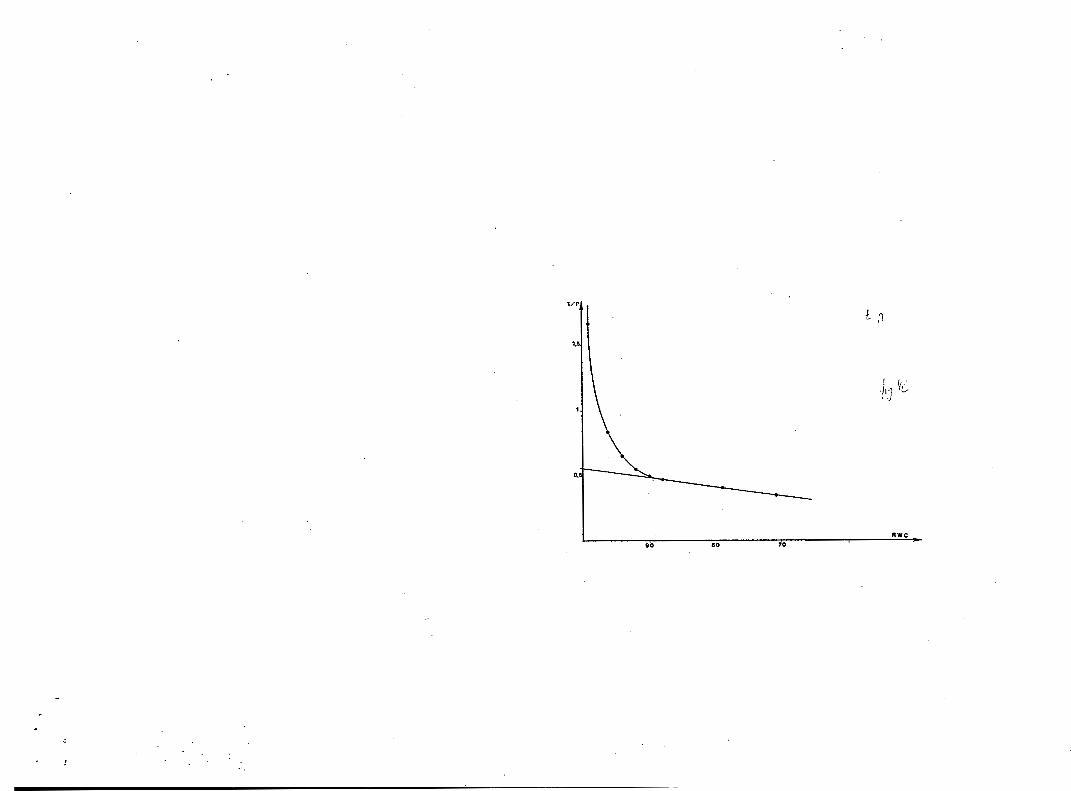
Les courbes pression-volume ( f i g . 4) sont su r tou t u t i l e s pour dé te r -
miner l a pression osmotique à sa tu ra t ion (TYREE & HAMMEL, 1972). Elles mon-
t r e n t que le Wapa a un p o t e n t i e l osmotique d i f f é r e n t de c e l u i du Tréma ou
du @". A partir cle l 'ensemble des courbes, nous avons en e f f e t :
Goupi : TP moyen = 13,O k U,6 bars
Trima : = 13,8 k 0,6 bar s
Napa : = 16,O k 1 , 7 bars.
La d i f f é rence n ' e s t cependant pas s u f f i s a n t e pour être s i g n i f i c a t i v e . Le d é f i c i t s u b l e t a l e s t un assez bon indice d e la résistance cyto-
à l 'abaissement de la teneur en eau (OPPENHEIMER, 1963 in KAPOS &
I t
plasmique
par fo is subjec t ive . On ob t i en t p a r f o i s , au bout de.temps de t r ansp i r a t ion
très longs , une ressa tura t ion , vo i re une sursa tura t ion , rapide des f e u i l l e s
q u i n ' a p l u s ; r i e n d e physiologique. I1 importe donc de v é r i f i e r
l'homogénéité d e l ' évolu t ion des va leurs en fonction du temps de dessèche-
ment.
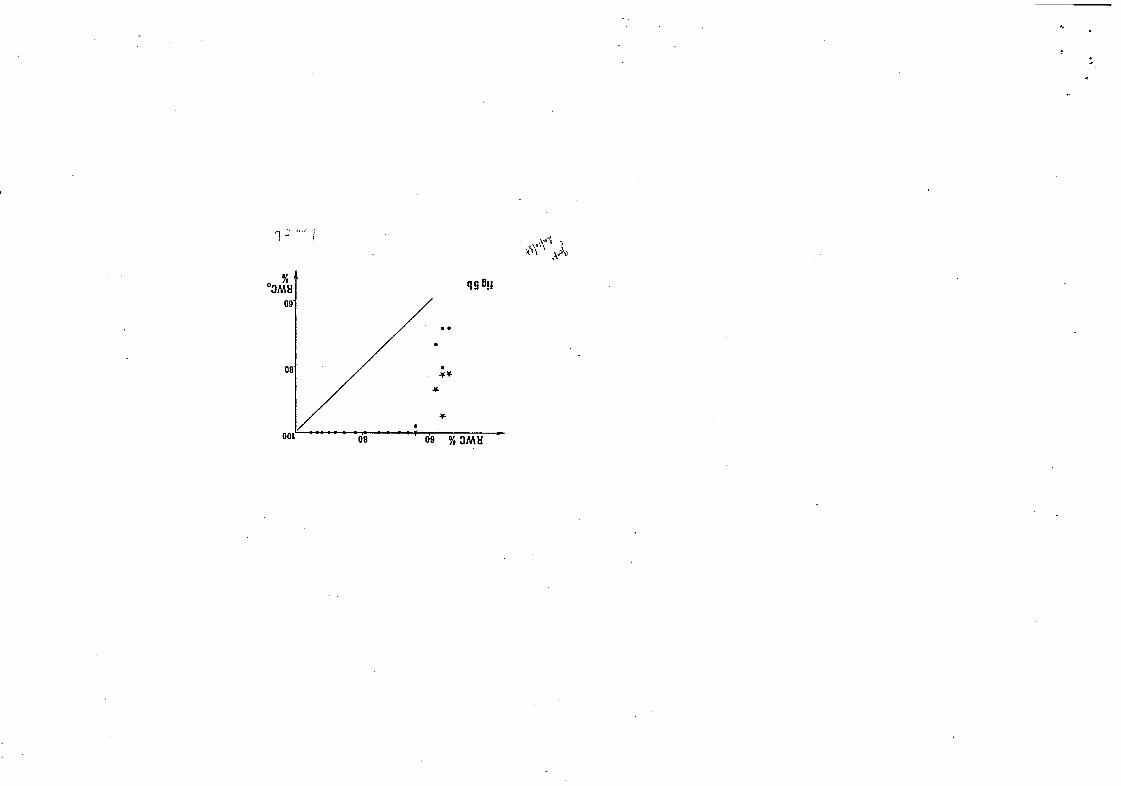
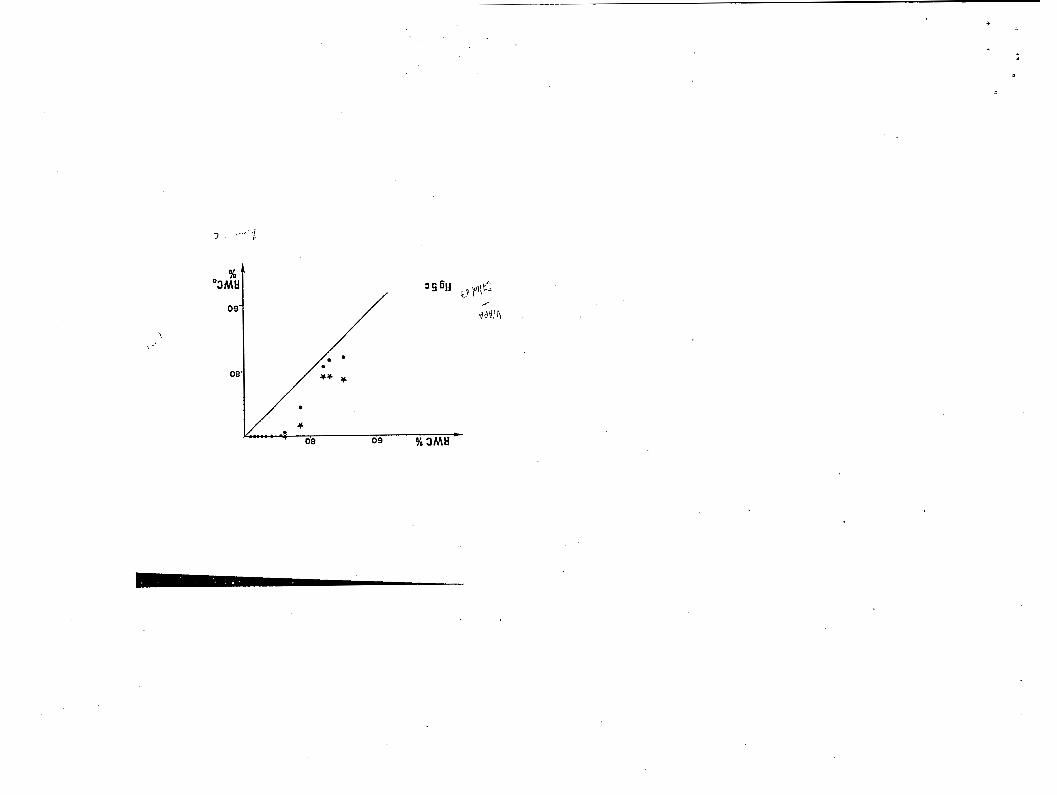
Compte tenu de cette remarque, les d é f i c i t s sublétaux observés au
cours des séries de mesures les p lus r égu l i è re s ( f i g . 5), chez nos 3
arbres , son t de :
Trema guineensis : 48 X
Goiipia glabra : 66 %
Eperua gr,wdlflora : 88 x .
i
i

I1 y a u r a i t donc un Btalement des valeurs avec une cor ré la t ion en t re
hSLiophilie e t rés is tance physiologique à l 'abaissement de l a teneur en
eau.
Remarquons que l e d é f i c i t sub lé t a l de tréma, bien que p l u s bas, e s t
a t t e i n t beaucoup p l u s v i t e par c e t t e plante (1 h environ dans l e s condi-
ticsns irle l 'expérience, c 'es t -à-dire au s o l e i l ) que par les 2 au t r e s espèces
i mviron 3 h ) .
incidemment il convient de signaler les abservations sur le f l é t r i s -
sement i fanaison) faites au cours des expériences de d é f i c i t sub lé t a l . I1
apparaî t une d i f fé rence marquée en t r e les 3 a rb res svr le plan de
l a fanaison, différence en rapport avec la r i g i d i t é des f e u i l l e s :
en e f f e t
- Tréma fane bien avant d ' a t t e ind re le d é f i c i t s u b l é t a l .
- Goupi fane (faiblement) quand il a t t e i n t l e d é f i c i t sub lé t a l .
- Wapa n e fane jamais ( f e u i l l e s adul tes ) .
Bien @.videment, le d é f i c i t sub lé t a l n ' a de valeur que r e l a t i v e : il 1
p l u s ou moins facilement a t t e i n t par la p lan te selon son apt i tude s o i t es t
à p u i s e r l 'eau (fonction essentiellement de son enracinement e t de son po-
t e n t i e l ) , s o i t $ l imi te r son dépar t , ce qu i e s t fonction de l a surface fo-
l i a i r e et, ~ u r t o u t , du controle stomatique e t cu t i cu la i r e .
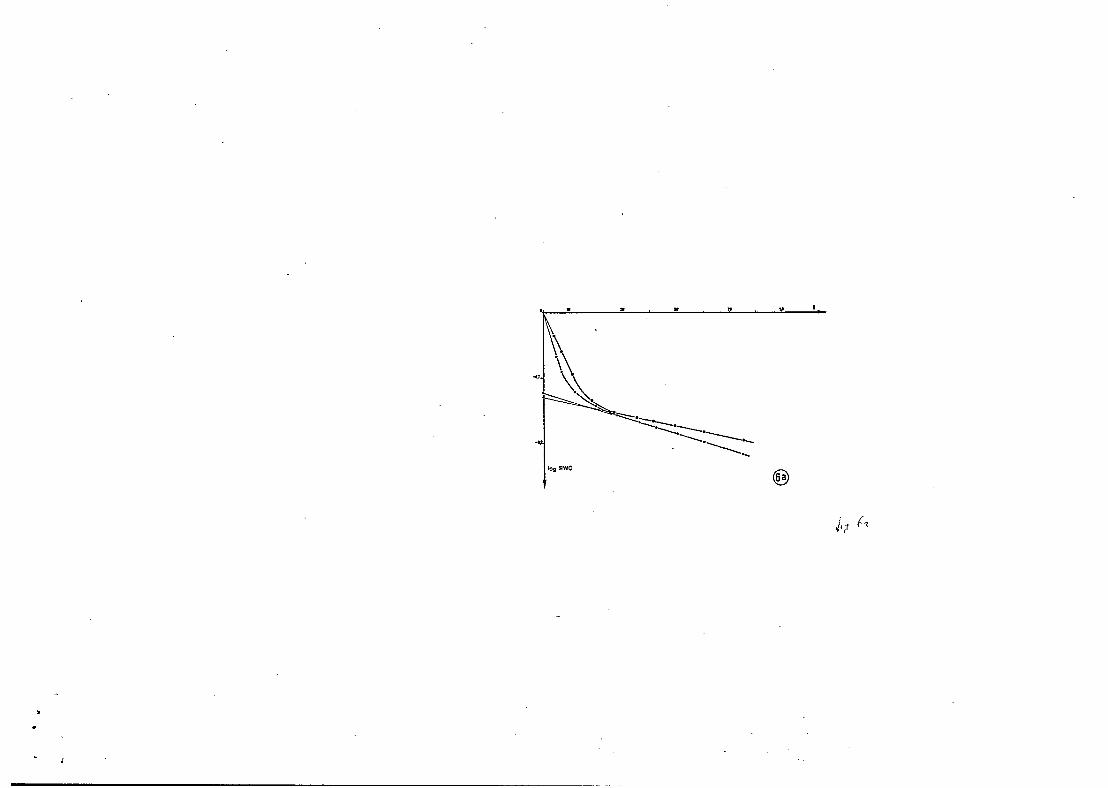
I c i la pesée des rameaux coupés est r iche d' informations ( f i g . 6 ) .
Nous avons regroupé sur le tableau 1 les pr inc ipa les caractéristiques
d a u i t e s des courbes : le point de début e t d e f i n de fermeture stomatique
exprimé en pourcentage de contenu en eau, le temps pour a t t e ind re la p le ine I
fermeture, l a pente de la première e t de l a deuxième partie d r o i t e de l a
isclurbe, anfin le temps pour a t t e i n d r e la d é f i c i t sub lé t a l .
~
On constate que l e goupi a très exactement les mêmes carac té r i s t iques
que l e wapa sauf pour la t ranspi ra t ion cu t i cu la i r e , p lus f a i b l e chez l e
wapa.

Tréma t r ansp i r e p l u s que l e goupi ou le wapa et malgré une régulation
s tomatique précoce dans l a temps, il a t t e i n t rapidement des valeurs très
basses de teneur en eau. Sa v é r i t a b l e régulat ion t r ansp i r a to i r e se s i t u e au
niveau d e la surface f o l i a i r e t o t a l e q u i , grâce à un renouvellement très
rapide d e s f e u i l l e s , s ’a jus te aux p o s s i b i l i t é s t r ansp i r a to i r e s ; nous avons
ainsi estimé l ’ i n d i c e f o l i a i r e 9 2 en saison humide e t moins de 1 en saison
sèche. L e goupi e t le wapa jeunes ne présentent pas de réduction sens ib le
\:!e l a sur face f o l i a i r e . La fanaison des f e u i l l e s de tréma fonctionne comme
une r4gulation supplémentaire en réduisant l ’ in te rcept ion de la lumière (cf
CHIARIELM & al . ,1987).
III - DlSCUSSLW
Le f a i t premier q u i r e s so r t de cette série de mesures, c‘est la grande
d i f fgrence de comportement e n t r e espèces. Ces d i f fé rences sont accentuées
par l a secheresse, ce qu i revient à d i r e que celle-ci est bien réelle.
Cependant, les espèces spontanées, avec l’exemple de tréma, sou f f r en t
I t
bien moins que c e l l e s qu i sont i n t rodu i t e s , i c i goupi e t wapa. Ceci dé-
montre l ’ex is tence d’une nécessaire succession végétale au sens de
CLEMENTS. I1 y a f a c i l i t a t i o n (au sens de CONNELL & SLATYER) dans la mesure
où la végétation pionnière contribue à créer une “ambiance f o r e s t i g r e ” p l u s
humide que le macroclimat.
Le deuxième f a i t es t peut-être méthodologique. Les indices c lass iques
de 1’6cophysiologie sont u t i l i s a b l e s en mi l ieu f o r e s t i e r &quatorial, mais
ces indices n’ont pas de valeur isolément. L e s t a t u t hydrique es t la résul-
t.ante d-i-in grand nombre de composantes e t c‘est bien évidemment le cont i -
nuum sol-plante-atmosphère q u ’ i l f a u t considérer : il f a u t éviter de couper
la f e u i l l e de ses racines . Précisément, l es t r o i s p lan tes i c i considérées
d i f f è r e n t nettement quant à leur morphologie racinaire (fig. 7) .

\
Tréma possède un systgme de racines extr6mement simple : 1 disque ou
plateau de rac ines & la sur face du 3.01 e t un cour t pivot q u i se prolonge,
vraisemblablement t rès profondément, par des rac ines f i n e s .
Chez le goupi ( c f ALEXANDRE, 1988b), les rac ines hor izonta les sont
P.gaiement les p lus développées mais elles ne forment pas un disque comme
(:hez tréma e t e l les sont moins s u p e r f i c i e l l e s . I1 n ’ y a pas de p ivot pro-
prement d i t mais un grand nombre d’axes rayonnants. La profondeur a t t e i n t e
par ces axes obliques e s t f a i b l e chez la jeune plante. Le goupi présente
souvent, e t c’est le cas i c i , d e s f e u i l l e s petites, minces e t claires, par-
f o i s rougeâtres. L’hypothèse d’une nématose a pu être Bcartée après tes t
avec un nématicide e t observation d i r e c t e de Scot to la Massese (com.
pers . ) . Un apport de f e r t i l i s a n t s du commerce n’a pas eu d ’ e f f e t s . Nous
pensons que la p l an te peut présenter une d i f f i c u l t é de mycorrhization q u i
a c c r o î t les r isques de d é f i c i t hydrique.
Les quelques observations e f fec tuées sur , l e wapa montrent un système
rac ina i r e composé d‘.assez grosses racines peu nombreuses mais profondes. La
profondeur d’enracinement es t directement en r e l a t ion avec la p o s s i b i l i t é
d’a l imenta t ion e n eau en période aÙ il ne p l eu t pas.
Un peut parler de r é s i s t ance à la sécheresse dans les mêmes termes que
ceux u t i l i s é s pour la r é s i s t ance aux maladies. I l existe a l o r s t r o i s types
de r é s i s t ance au sens large : l’immunité où la plante ignore le manque
d‘eau en ayant une absorption s u f f i s a n t e e t éventuellement d e s réserves , la
rés i s tance proprement d i t e QÙ la plante limite ses pertes, enf in la
tolérance où la plante fonctionne encore malgré un b i l an hydrique dé f i -
c i taire.
S i l‘on peut parler d e résistance a la sécheresse chez nos t r o i s
a rb res q u i bien sûr n ’en ont qu‘une bien modeste, chacun d’eux i l l u s t r e une
de ces tendances. Tréma est immun grâce su r tou t à ses rac ines profondes

mais également gràce à la forte teneur en eau de ses tissus. En conservant
iine. transpiration élevée, trema peut continuer à croître malgré la séche-
resse relative e t peut ainsi conserver dans les recrûs sa posi t ion domi-
n a n t e , ce q u i es t nécessaire à sa survie car c ' es t une espèce strictement
héliophile. Cette stratégie écophysiologique l e met en revanche à l a merci
d'une skheresse trop accentuée. La stratégie de tréma semble caractéris-
tique de très nombreuses espèces pionnières.
Le ,soupi, l u i , est p l u t ô t tolérant ; en effet il continue à transpirer
inaLgr6 un b i l a n déf ic i ta i re e t le wapa e s t p lu t e t résistant avec son '
@paisse cuticule qui se retrouve chez la grande majorité des espèces l e s
p l u s "forestières". Ce caractère bien connu (cf RICHARDS, 1952) ne. semble
@&re avoir d ' u t i l i t é , ii moins de considérer que son rôle e s t la conserva-
t i o n du CO2 e t non de H z O (cf RAPOS & TANNER, loc. c i t . ) .
. .

LEGENDE DES F ICjUEES
Figure 1. Distr ibut ion de la résistance stomatique en fonct ion du rang de la f e u i l l e à partir de l‘extrémité du rameau. Exemple obtenu à p a r t i r de Goupia glabra, mai 1985.
Figure 2. Pluviométrie, po ten t i e l de base ( B I , po ten t i e l minimum ( m) e t conductance maximale de Trema micrantha (carrés), Goiipia glabra (ronds) e t Eperua grandiflom ( 6 t o i l e s ) au cours du “ p e t i t été de mars“ en 1977, à Cayenne.
Figure 3. Pluviométrie, po ten t i e l de base ( B ) , po ten t i e l m i n i m u m ( m) e t conductance maximale de Trema micmntha ( c a r r é s ) , Gaipia g h b r a (ronds) et E p e r m grandiflora (triangles) au cours de la saison sèche de 1977, à Cayenne.
Figure 4. Exemples de “courbes pression-volume” obtenues à partir de Trema micrantlia ( f i g . 4a) , Goupia &labra ( f i g . 4b) e t Eperua grand i f lo ra ( f i g . 4C).
Figure 5. Déf i c i t s u b l é t a l : réhydratation au bout de 6 heures (ronds) e t 24 heures ( é t o i l e s ) en fonction du contenu en eau i n i t i a l : 5a, Trema micrantha ; Sb, ~ ‘ ; o ~ p i a glabra ; 5c, Eperua grandi f lora .
“ I Figure 6. Courbe de t r ansp i r a t ion de rimeaux coupés en condi t ions contrô- l5es = logarithme du contenu relatif en eau, en fonction du temps. F i g . 6a : Trema micrantha sous éclairement de 350 ( é t o i l e s ) e t 5 W/m2 ( ronds ) , température 30°C, HR 70-75 %. F i g . 6b : t ranspi ra t ion de Goupia g1,abra (ronds) e t Trema micrantha (triangles), éclairement 250 W/m2, température 28”C, HR 60-70 %. Absence de contr8le stomatique fréquemment observée chez Trema e t de nombreuses a u t r e s hé l iophi les (Cecropia, Inga, Terminalia., . . .. ) . F i g . 6c : Eperua grandi f lora , éclairement 350 W/mz, température 30”C, HR 70 %.
Figure 7. Dessin schématique de l’appareil rac ina i re , à t r o i s ans, de Trema micrantha (Tm), Goipia glLabra (Gg) e t Eperua grandi f lora (Eg) .

i
L , 7
Debut Fin Temps al a2 Temps ferraeture ferreture fermeture D,S,L.
x x minutes heures
Trema 7’1 t 2 b7 t 4 28 f 6 b l i 5 10 f 5 2
û o u p i 87 i 2 77 f a JO f 10 45 f 10 10 f 3 b
Mapa 83 i 3 75 ,, 5 40 f 10 j 45 i 15 5 f 3 6
Tableau I : Carac tér i s t iques d e la t r ansp i r a t ion par la technique de pesée des rameaux coupés : - Turgescence r e l a t i v e en début e t f i n de fermeture stomatique. - Conduct.ances stomatiques (al) e t c u t i c u l a i r e s (U) (pentes d e s parties droi tes des courbes de t ranspi ra t ion , u n i t é s a r b i t r a i r e s ) . - Temps pour a t t e i n d r e la teneur en eau correspondant au d é f i c i t s u b l é t a l . Les rmeaux coupés sont placés à l’ombre dans des condi t ions d’évaporation modérée.

BIBLIOGRAPHIE
ALFXANljRE D.Y., 1988a.- Pluies e t alimentation en eau des plantes dans la région de Sinnamary (Guyane française) . - Múltigr. ORSMM Cayenne, à para î t r e in Nature guyanaise, 10 p.
ALEXANDRE D.Y., 1988b.- Note : aperçu sur la morphologie rad icu la i re de deux espèces pionnières de Guyane française : Gbupia glabra e t Vismia guimensis. - Multigr . ORSMM Cayenne, 6 p.
FORGET P.M., 1988.- Dissémination e t régénération na tu re l l e de h u i t espèces d'arbres en f o r ê t guyanaise.- Th. Paris 6, 232 p .
AUSSENAC C., 1985.- Le po ten t ie l hydrique de l ' a r b r e : une donnée essen- t i e l l e pour la compr6hension de l 'écophysiologie des essences fores- t i è r e s . - Science du Sol 4 : 217-226.
CHIARIELM N . R . et a l . , 1987.- Midday wil t ing in a t rop ica l pioneer tree.-
HALLE F. & OLDEMAN R.A.A., 1970.- Essai sur l ' a r ch i t ec tu re e t l a dynamique
Functional Ecology 1 : 3-11.
de croissance des arbres tropicaux.- Par is , Masson, 178 p.
V. & TANNER E.V.J., 1985.- Water r e l a t ions of Jamaican upper montane rain fo re s t trees.- Ecology 66 (1) : 241-250.
OPPENHEIMER H . R . , 1963.- Zur Kenntnis k r i t i s che r Wasser sat t igungsdefizi te i n Blattern und ih re r Bestimmung.- Planta 60 : 51-69.
RICHARDS P.W., 1952.- The t ropica l r a in fo re s t . - Cambridge University Press, 450 p.
RYCHNOVSKá-SOUDKOVá M., 1963.- Study of the r e v e r s i b i l i t y of the water sa- turat ion d e f i c i t as one of the methods of causal photogeography.- Biol. Plant.;5 : 175-180.
TYREE M.T. & HAMMEL H.T. , 1972.- The measurement of the turgor pressure and the water r e l a t ions of p lan ts by the pressure bomb technique.- J . Ekp. Bot. 23 : 267-282.

YO
110.
IQ.
.!i
-3
.1
'
n 10 1 2 3 4 1 i 7 8 9 1 0 ~ 1 1 2
-
f ig 1

I S II c
I , I I I l I I - I I... 1,111 il
cond.
cm/s I: ' I
I .
1 I
HPt. 1 . oclobrm i ' ' 1 dmc. .. I .. novmmbre o *---
MPa I
.. .

I RWC so sb 70 10

I RWC o 8'0 70 Il0
. .

1 %
O3Mtl o 9'
O 8
. . O r .$ . r *
.* "
001, 08 o9 0 2 %lMU /
i
o91
- 1

c Y

k
i






























