Embed Size (px)
Citation preview

Virus Research 118 (2006) 16–22
Completion of the genome sequence of Lettuce necrotic yellows virus,type species of the genus Cytorhabdovirus�
Ralf G. Dietzgen a,b,∗, Ben Callaghan a,b, Thierry Wetzel c,d, James L. Dale c
a Department of Primary Industries and Fisheries, Queensland Agricultural Biotechnology Centre, Queensland Bioscience Precinct,The University of Queensland, 306 Carmody Rd, St. Lucia, Qld. 4072, Australia
b School of Molecular and Microbial Sciences, The University of Queensland, St. Lucia, Qld. 4072, Australiac Science Research Centre, Queensland University of Technology, Faculty of Science, Brisbane, Qld 4001, Australia
d RLP Agroscience, AlPlanta-Institute for Plant Research, Breitenweg 71, 67435 Neustadt a.d.W., Germany
Received 7 September 2005; received in revised form 31 October 2005; accepted 31 October 2005Available online 28 November 2005
Abstract
We completed the genome sequence of Lettuce necrotic yellows virus (LNYV) by determining the nucleotide sequences of the 4a (putativepsarmcLsm©
K
1
aaRasjtgm
u
0d
hosphoprotein), 4b, M (matrix protein), G (glycoprotein) and L (polymerase) genes. The genome consists of 12,807 nucleotides and encodesix genes in the order 3′ leader-N-4a(P)-4b-M-G-L-5′ trailer. Sequences were derived from clones of a cDNA library from LNYV genomic RNAnd from fragments amplified using reverse transcription-polymerase chain reaction. The 4a protein has a low isoelectric point characteristic forhabdovirus phosphoproteins. The 4b protein has significant sequence similarities with the movement proteins of capillo- and trichoviruses anday be involved in cell-to-cell movement. The putative G protein sequence contains a predicted 25 amino acids signal peptide and endopeptidase
leavage site, three predicted glycosylation sites and a putative transmembrane domain. The deduced L protein sequence shows similarities with theproteins of other plant rhabdoviruses and contains polymerase module motifs characteristic for RNA-dependent RNA polymerases of negative-
trand RNA viruses. Phylogenetic analysis of this motif among rhabdoviruses placed LNYV in a group with other sequenced cytorhabdoviruses,ost closely related to Strawberry crinkle virus.2005 Elsevier B.V. All rights reserved.
eywords: Plant rhabdovirus; LNYV; Phylogeny
. Introduction
Rhabdoviruses can infect vertebrates, invertebrates and plantsnd represent a large virus family of importance to agriculturend human health. They have a single-stranded, negative-senseNA genome of ca. 12–15 kb, which encodes five function-lly conserved proteins. Axillary proteins are encoded by somepecies. Conserved intergenic regions are located at the geneunctions and partially complementary untranslated regions,ermed 3′ leader and 5′ trailer are located at either end of theenome. In rhabdovirus-infected cells, the viral RNA poly-erase facilitates the transcription of distinct mRNA species
� The sequence data have been deposited in the GenBank/EMBL databanksnder accession number no. AJ867584.∗ Corresponding author. Tel.: +61 7 3346 2703; fax: +61 7 3346 2727.
E-mail address: [email protected] (R.G. Dietzgen).
from each gene and genome replication (Tordo et al., 2005;Jackson et al., 2005).
Lettuce necrotic yellows virus (LNYV) is the type speciesof the genus Cytorhabdovirus (Tordo et al., 2005), membersof which are characterised by accumulation of enveloped viri-ons in the cytoplasm of infected cells (Dietzgen, 1995; Jacksonet al., 2005). LNYV causes a serious disease of lettuce inAustralia and is transmitted in a persistent, propagative man-ner by the aphid Hyperomyzus lactucae (Francki et al., 1989).The LNYV genome consists of a monopartite, negative-sense,single-stranded RNA of about 13,000 nucleotides (Wetzel et al.,1994a). The physical map of the LNYV genome is 3′ leader-N-P-4b-M-G-L-5′ trailer, where N is the nucleocapsid gene, Pis the putative phosphoprotein gene, 4b encodes a protein ofunknown function, M is the matrix protein gene, G is the gly-coprotein gene and L is the polymerase gene (Wetzel et al.,1994a). The sequences of the 3′ leader, N gene, 5′ trailer and allintergenic sequences have been reported previously (Wetzel et
168-1702/$ – see front matter © 2005 Elsevier B.V. All rights reserved.oi:10.1016/j.virusres.2005.10.024

R.G. Dietzgen et al. / Virus Research 118 (2006) 16–22 17
al., 1994a,b). In this paper, we present the completed nucleotidesequence of LNYV and compare it to the genomes of other plantrhabdoviruses.
2. Materials and methods
2.1. Virus propagation, purification and isolation of viralRNAs
The previously described garlic isolate (Sward, 1990) wasused throughout. LNYV was propagated in Nicotiana gluti-nosa and leaves were collected 10–12 days post inoculation(dpi). Virus was purified and the genomic RNA extractedas described previously (Francki et al., 1989; Dietzgen etal., 1989). Total RNA from LNYV-infected N. glutinosa wasextracted as described by Rezaian et al. (1983) or Higginsand Dietzgen (2000), or using RNeasy® Plant Mini kits (Qia-gen, Hilden, Germany) following the manufacturer’s instruc-tions. Poly(A)+ RNA was fractionated on oligo(dT)-celluloseusing a mRNA purification kit (Pharmacia Biotech, Freiburg,Germany).
2.2. Oligonucleotide primers
Oligonucleotide primers for determination of the P, 4b andMssPbT
2.3. PCR amplification and RACE
Poly(A)+ RNA preparations from LNYV-infected N. gluti-nosa, and purified LNYV genomic RNA were used forcDNA synthesis and PCR amplification as described previously(Wetzel et al., 1994a). For the P, 4b and M genes, the reversetranscription mixture was denatured for 5 min at 94 ◦C prior toPCR amplification for 40 cycles of 94 ◦C for 20 s, 42 ◦C for 20 sand 72 ◦C for 30 s.
For amplification of the G and L genes, primers G1 and G3,respectively were heat-denatured prior to cDNA synthesis andPfu Turbo DNA polymerase (Stratagene, La Jolla, CA) was usedfor PCR. Primers G1 and G2 were used to amplify the completeG gene between the G-M and the G-L intergenic regions (PCR-1; Fig. 1) using 30 cycles of 94 ◦C for 1 min, 50 ◦C for 1 min,and 72 ◦C for 2 min. Primers L11 and L12 were used in PCRfor 30 cycles of 94 ◦C for 30 s, 50 ◦C for 30 s and 72 ◦C for6 min to amplify the majority of the L gene (PCR-3; Fig. 1). Lgene cDNA was also used to amplify additional DNA fragments,which overlapped each of the two internal EcoRI sites of thePCR-3 DNA fragment (L4/L5 and L6/L7) or linked PCR-3 tothe G gene (G3/L3), yielding clones PCR-4, PCR-5 and PCR-2,respectively (Fig. 1).
For 3′ RACE (rapid amplification of cDNA ends), anoligo(dT)12–18 primer was used for cDNA synthesis. The subse-quent PCR amplification was done with oligo(dT) and theLftwst
TL
C ide se
CTATTTGATTCAATGTGAAAC
CCGTGTTG
P AGAGTTCT
P GCGAGA
P TTCTTTCT
P ACTTCATC
P TGCCCGTG
“ a”, “4a
gene sequences were synthesized using a PCR-MATE DNAynthesizer (Applied Biosystems). Primers for amplification andequencing of the G and L genes were synthesized by Bresatecty Ltd. (Adelaide, South Australia) and Sigma-Genosys (Mel-ourne, Australia). Primer names and sequences are listed inable 1.
able 1NYV-specific oligonucleotide primers used in this study
lone Primer name Nucleot
g4a TAGAGm4a GATTGg4b CGCTCm4b1 GAGGGm4b2 AGTGCgM ACGGAmM TGTTTgG ACCTT
CR-1 G1 GGTCTG2 CGGAA
CR-2 G3 GTCAAL3 TATCAT
CR-3 L11 GCGAAL12 GCGAA
CR-4 L4 CTTTAL5 CTCTT
CR-5 L6 CGATAL7 GCTTC
g” corresponds to the genomic sense, “m” to the messenger RNA sense and “4re underlined.
12–18NYV-specific primers m4a, m4b1 or m4b2, and mM (Table 1)
or 3′ RACE of the LNYV 4a, 4b, and M mRNAs, respec-ively. For 5′ RACE, cDNA was synthesized, purified, tailedith dCTP or dGTP and PCR-amplified using the 5′ RACE
ystem (Life Technologies), according to protocols provided byhe supplier. For dG-tailing, an oligo-dC15 primer was used in
quence 5′ → 3′ Remark
CATCAGAAGTTTTAGG 5′ RACECTTCCTTGTCC 3′ RACETAGCATCCAGATCCTC 5′ RACE
ATTGCGTGATCACGTTG 3′ RACECATTGATGAGTTAGGGG 3′ RACEAAGAACTCACATCG 5′ RACECTGAAGTGTTC 3′ RACECATCTAGTCAC 5′ RACE
ATTCACATATAGACGAGTTATATCCG XbaITTTCTTAAATCACACATGCCACTTGG EcoRI
ACATGGATCTGAGAGATTGAAGG
TGGAGGTTGTTGAGATTTGAC EcoRITGGTTCTAATTGCATACACC EcoRI
GTATGGACTCACTCATCAACC
AAAGGAGCCCGTTGGTGTC
b”, “M”, “G” and “L” correspond to the respective genes; non-viral sequences

18 R.G. Dietzgen et al. / Virus Research 118 (2006) 16–22
Fig. 1. Organisation of the LNYV genome and location of clones that were used for sequence analysis. The location and relative size of the LNYV genes are shown.Clone � and the multigenic clones A, B, C, D and X were identified previously (Dietzgen et al., 1989; Wetzel et al., 1994a). PCR clones 1–5 covering the G and Lgenes were generated using the primer pairs listed in Table 1.
place of the AAP primer. The LNYV-specific primers g4a, g4b,gM and gG (Table 1) were used for 5′ RACE of LNYV 4a,4b, M and G mRNAs, respectively. At least two clones fromindependent PCR reactions were sequenced for each 5′ RACEexperiment.
2.4. Cloning of PCR products
PCR products of the P, 4b and M mRNA sequences were lig-ated into pCR II (TA cloning kit, Invitrogen), or into dTTP-tailedpBluescript (Stratagene) and used to transform INVaF’ (Invit-rogen) or DH5aF’ E. coli cells. The G gene PCR product wasdigested with EcoRI and XbaI, gel-purified and ligated into sim-ilarly digested pGEM-3Z (Promega, Madison, WI). The PCR-3product of the L gene was digested with EcoRI, the 3 fragmentswere isolated using QIAEX II gel extraction kit (Qiagen), lig-ated into EcoRI-digested, shrimp alkaline phosphatase-treatedpBluescript SK, and transformed into E. coli (JM109 strain)(Promega, Madison, WI). Plasmid miniprep extractions weredone by alkaline lysis (Sambrook and Russell, 2001) or using aQIAprep® Spin Miniprep Kit (Qiagen). The order and orienta-tion of the L gene EcoRI fragments was determined by restrictionenzyme analysis. Additional overlapping PCR products were A-tailed with Taq DNA polymerase, ligated into pCR2.1-TOPOand transformed into TOP10 cells (Invitrogen).
2
Spatld(b(Ro3e(A
2.6. Sequence analysis and phylogeny
The putative G and L gene mRNA sequences were identifiedbased on the published sequence data from multigenic clones(Wetzel et al., 1994a). Open reading frame (ORF) analysis andhydropathy plots were carried out using MacVector. Pair-wise(“GAP”) and multiple sequence (“CLUSTALW”) alignmentsand phylogenetic analysis utilizing the “Phylip” and GCG soft-ware packages were facilitated through the Australian NationalGenome Information Service (Sydney, Australia). ORF sizeswere calculated excluding the stop codon. ORFs were trans-lated into amino acid sequences using “TRANSLATE” andqueried against the Swiss-Prot database using “BLAST P”. Pre-dicted protein molecular weight, isoelectric point and chargewere determined using “PEPSTATS”. Some putative proteinmotifs and secondary structure elements were identified usingprograms on the Predict Protein website (http://www.embl-heidelberg.de/predictprotein/predictprotein.html).
3. Results
3.1. The complete nucleotide sequence of LNYV
We have completed the genome sequence of LNYV fromoverlapping clones derived from a cDNA library from LNYVvf(g1natod
3
psdGs
.5. Sequencing of cDNA clones and PCR products
Some plasmid DNA was sequenced manually using theequenase kit (USB) and thio-[a-35S]dATP (Dupont). For thereviously obtained LNYV cDNA clones (Dietzgen et al., 1989),ddition of 0.5 mg of T4 gene 32 protein (Pharmacia) was some-imes required during the labelling reaction to read throughong G- or C-tails. Gel-purified PCR products were sequencedirectly following the protocol described by Casanova et al.1990), except that 10% DMSO was added to the annealinguffer. Automatic sequencing of plasmid DNA or PCR productsat least 3 clones on both strands) was done using the PrismTM
eady Reaction DyeDeoxy Termination Cycle sequencing kitr Big Dye Terminator Ready Reaction Mix versions 2.0 or.1 (PE Applied Biosystems, Foster City, CA). Sequences weredited and aligned using the IBI Sequence Analysis, MacVectorOxford Molecular Ltd.) or Sequencher 3.0 (Gene Codes Corp.,nn Arbor, MI) software packages.
irion RNA generated previously (Wetzel et al., 1994a,b) androm fragments of the G and L genes amplified by RT-PCRFig. 1). Recently PCR-amplified sequences of the N and Penes differed from the previously determined sequences by
(Callaghan and Dietzgen, 2005) and 7 nucleotides (dataot shown), respectively. Clones of the P, 4b and M mRNAsnd 5′ and 3′ RACE confirmed the transcribed sequences ofhese genes. The complete LNYV genome sequence comprisedf 12,807 nucleotides has been deposited in GenBank/EMBLatabases with the accession number AJ867584.
.2. The phosphoprotein gene
The 4a gene (AF209035), thought to represent the phos-hoprotein (P) gene, based on its location in the genome, wasequenced from both genomic RNA and mRNA. 5′ RACE andG-tailing of the cDNA identified the start of the mRNA as 5′-GAAACC-3′, indicating the presence of an additional guano-
ine (G cap). The P gene was 1082 nucleotides in length and
R.G. Dietzgen et al. / Virus Research 118 (2006) 16–22 19
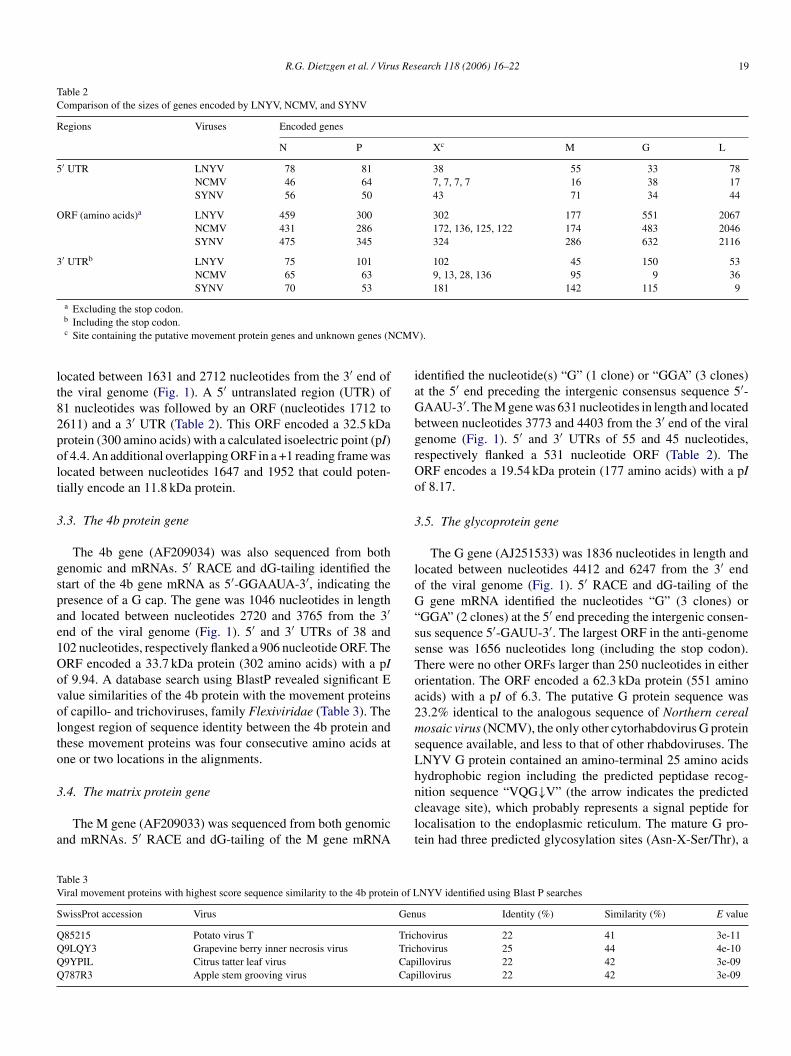
Table 2Comparison of the sizes of genes encoded by LNYV, NCMV, and SYNV
Regions Viruses Encoded genes
N P Xc M G L
5′ UTR LNYV 78 81 38 55 33 78NCMV 46 64 7, 7, 7, 7 16 38 17SYNV 56 50 43 71 34 44
ORF (amino acids)a LNYV 459 300 302 177 551 2067NCMV 431 286 172, 136, 125, 122 174 483 2046SYNV 475 345 324 286 632 2116
3′ UTRb LNYV 75 101 102 45 150 53NCMV 65 63 9, 13, 28, 136 95 9 36SYNV 70 53 181 142 115 9
a Excluding the stop codon.b Including the stop codon.c Site containing the putative movement protein genes and unknown genes (NCMV).
located between 1631 and 2712 nucleotides from the 3′ end ofthe viral genome (Fig. 1). A 5′ untranslated region (UTR) of81 nucleotides was followed by an ORF (nucleotides 1712 to2611) and a 3′ UTR (Table 2). This ORF encoded a 32.5 kDaprotein (300 amino acids) with a calculated isoelectric point (pI)of 4.4. An additional overlapping ORF in a +1 reading frame waslocated between nucleotides 1647 and 1952 that could poten-tially encode an 11.8 kDa protein.
3.3. The 4b protein gene
The 4b gene (AF209034) was also sequenced from bothgenomic and mRNAs. 5′ RACE and dG-tailing identified thestart of the 4b gene mRNA as 5′-GGAAUA-3′, indicating thepresence of a G cap. The gene was 1046 nucleotides in lengthand located between nucleotides 2720 and 3765 from the 3′end of the viral genome (Fig. 1). 5′ and 3′ UTRs of 38 and102 nucleotides, respectively flanked a 906 nucleotide ORF. TheORF encoded a 33.7 kDa protein (302 amino acids) with a pIof 9.94. A database search using BlastP revealed significant Evalue similarities of the 4b protein with the movement proteinsof capillo- and trichoviruses, family Flexiviridae (Table 3). Thelongest region of sequence identity between the 4b protein andthese movement proteins was four consecutive amino acids atone or two locations in the alignments.
3
a
identified the nucleotide(s) “G” (1 clone) or “GGA” (3 clones)at the 5′ end preceding the intergenic consensus sequence 5′-GAAU-3′. The M gene was 631 nucleotides in length and locatedbetween nucleotides 3773 and 4403 from the 3′ end of the viralgenome (Fig. 1). 5′ and 3′ UTRs of 55 and 45 nucleotides,respectively flanked a 531 nucleotide ORF (Table 2). TheORF encodes a 19.54 kDa protein (177 amino acids) with a pIof 8.17.
3.5. The glycoprotein gene
The G gene (AJ251533) was 1836 nucleotides in length andlocated between nucleotides 4412 and 6247 from the 3′ endof the viral genome (Fig. 1). 5′ RACE and dG-tailing of theG gene mRNA identified the nucleotides “G” (3 clones) or“GGA” (2 clones) at the 5′ end preceding the intergenic consen-sus sequence 5′-GAUU-3′. The largest ORF in the anti-genomesense was 1656 nucleotides long (including the stop codon).There were no other ORFs larger than 250 nucleotides in eitherorientation. The ORF encoded a 62.3 kDa protein (551 aminoacids) with a pI of 6.3. The putative G protein sequence was23.2% identical to the analogous sequence of Northern cerealmosaic virus (NCMV), the only other cytorhabdovirus G proteinsequence available, and less to that of other rhabdoviruses. TheLNYV G protein contained an amino-terminal 25 amino acidshnclt
TV n of L
S Gen
Q TricQ TricQ CapQ Cap
.4. The matrix protein gene
The M gene (AF209033) was sequenced from both genomicnd mRNAs. 5′ RACE and dG-tailing of the M gene mRNA
able 3iral movement proteins with highest score sequence similarity to the 4b protei
wissProt accession Virus
85215 Potato virus T9LQY3 Grapevine berry inner necrosis virus9YPIL Citrus tatter leaf virus787R3 Apple stem grooving virus
ydrophobic region including the predicted peptidase recog-ition sequence “VQG↓V” (the arrow indicates the predictedleavage site), which probably represents a signal peptide forocalisation to the endoplasmic reticulum. The mature G pro-ein had three predicted glycosylation sites (Asn-X-Ser/Thr), a
NYV identified using Blast P searches
us Identity (%) Similarity (%) E value
hovirus 22 41 3e-11hovirus 25 44 4e-10illovirus 22 42 3e-09illovirus 22 42 3e-09

20 R.G. Dietzgen et al. / Virus Research 118 (2006) 16–22
putative carboxy-terminal transmembrane domain and 18 cys-teine residues, which could potentially be involved in the for-mation of disulfide bridges (data not shown).
3.6. The polymerase (L) gene
The L gene (AJ746199) was 6335 nucleotides in length andlocated between nucleotides 6278 and 12,613 from the 3′ endof the viral genome (Fig. 1). There was a single large ORF of6204 nucleotides (including the stop codon) commencing fromthe first AUG. No other ORFs larger than 250 nucleotides wereidentified. The ORF encoded a protein of 236.4 kDa (2067 aminoacids), which had a sequence similarity at the nucleotide level of44.5% with NCMV and of 42.3% with either Sonchus yellow netvirus (SYNV) or Rice yellow stunt virus (RYSV). The L proteinsequence contained polymerase module motifs characteristicfor RNA-dependent RNA polymerases of negative-strand RNAviruses, including the conserved “GDN” motif, which is thoughtto represent the catalytic centre (data not shown).
3.7. Phylogeny of plant rhabdoviruses based on the L gene
Phylogenetic analysis of the conserved polymerase module(block III) of the L gene comprising motifs pre-A to E (Poch
FsEmranS(vyn(J
et al., 1989; Muller et al., 1994; Callaghan, 2005) placed theplant rhabdoviruses into two distinct clades, which correspondto the cyto- and nucleorhabdovirus genera, respectively (Fig. 2).According to this analysis, the plant rhabdoviruses appear tobe more closely related to the novirhabdoviruses than to theother animal rhabdoviruses. LNYV clustered with the other twosequenced cytorhabdoviruses, but appears to be evolutionarilymore closely related to Strawberry crinkle virus (SCV) than toNCMV. The five nucleorhabdoviruses fell into two distinct sisterclades with Maize mosaic virus (MMV) and taro vein chloro-sis virus (TaVCV) forming one sub-clade and SYNV and maizefine streak virus (MFSV) another. The association of RYSV witheither sister clade varied depending on whether maximum par-simony or neighbour-joining methods were used.
4. Discussion
LNYV isolates can be divided into two subgroups based onN gene sequence variability—the isolate we sequenced belongsto subgroup I (Callaghan and Dietzgen, 2005). The order ofthe genes on the LNYV genome 3′-N-P-4b-M-G-L-5′ and thenucleotide sequences of all intergenic regions, nucleocapsid pro-tein gene, 3′ leader and 5′ trailer have been reported previously(Wetzel et al., 1994a,b). The N gene sequence did not showgenetic drift from the previously reported sequence since itonly differed by a single nucleotide (Callaghan and Dietzgen,2egtsr12tagdotnhN“abTi
ig. 2. Phylogenetic tree constructed from alignment of the deduced amino acidequences of the conserved rhabdovirus L protein polymerase module (pre-A to
domains of box III). The tree was generated using the maximum parsimonyethod. Bootstrap values of 100 tree replicas are shown at each branch node. The
habdoviruses and GenBank sequence accession numbers used in this analysis
re bovine ephemeral fever virus (BEFV; AF234533), infectious hemopoieticecrosis virus (IHNV; X89213), Hirame rhabdovirus (HIRRV; AF104985),nakehead rhabdovirus (SHRV; AF147498), viral hemorrhagic septicemia virusVHSV; Y18203), maize mosaic virus (MMV; AY618418), taro vein chlorosisirus (TaVCV; AY674964), rice yellow stunt virus (RYSV; AB011257), sonchusellow net virus (SYNV; L32603), maize fine streak virus (MFSV; AY618417),orthern cereal mosaic virus (NCMV; AB030277), strawberry crinkle virusSCV; AY005146), rabies virus (RABV; M31046), and vesicular stomatitis Newersey virus (VSNJV; M29788).tRmcS5“p
005). All six LNYV transcripts have been detected by north-rn blot analysis (Wetzel et al., 1994a). We have completed theenome sequence by determining the nucleotide sequences ofhe remaining genes, namely P, 4b, M, G and L. The genomeize of 12,807 nucleotides is within the range of 12 to 14.5 kbeported for other sequenced plant rhabdoviruses (Choi et al.,994; Tanno et al., 2000; Huang et al., 2003; Revill et al.,005; Tsai et al., 2005; C.D. Schoen, personal communica-ion), but there is considerable variability in the number of ORFsmong and between cyto- and nucleorhabdoviruses. The LNYVenome organisation is most similar to that of the nucleorhab-oviruses SYNV and TaVCV, but differs considerably from thatf the other cytorhabdoviruses in that LNYV has six ORFs inhe antigenome sense, while SCV has seven and NCMV hasine ORFs (Tanno et al., 2000). The LNYV and SCV genomesave one ORF located between the P and M genes, whereasCMV carries four ORFs in this position which we termed
X” (Table 2). Among the sequenced cytorhabdoviruses, SCVnd NCMV, appear to have an additional small putative ORFetween the G and L genes (C.D. Schoen, pers. communication;anno et al., 2000), while no ORF was evident in this location
n the LNYV genome.The start sites for the transcription of all LNYV genes, except
he L gene mRNA have been experimentally determined by 5′ACE. The 5′ terminus of each LNYV mRNA sequence com-enced with a guanine residue, which probably represents a 5′
ap structure (Wetzel et al., 1994b), similar to that suggested forYNV mRNAs (Scholthof et al., 1994). Some of the dG-tailed′ RACE clones of the M and G mRNAs had the 5′ sequenceGGA” instead of “G”. The additional nucleotides “GA” hadreviously also been identified in one of the dG-tailed clones of

R.G. Dietzgen et al. / Virus Research 118 (2006) 16–22 21
the N mRNA (Wetzel et al., 1994a). They may be due to poly-merase slippage resulting in transcription of the two nucleotidesimmediately prior to the predicted mRNA start at the 5′ end(viral sense) of the conserved intergenic region. Polymerase slip-page at Vesicular stomatitis virus (VSV) gene junctions has beenreported (Barr and Wertz, 2001). It is possible, but seems lesslikely that the additional nucleotides are of non-viral origin ashas been suggested for the 5′ termini of the RYSV M, G and Lgenes (Luo and Fang, 1998).
The deduced amino acid sequences of the P, 4b, M, G andL proteins shared a number of features with analogous rhab-dovirus genes. The putative P gene, previously referred to as“4a”, contained a second ORF which could potentially encode asmall highly basic protein in an overlapping +1 reading frame,similar to the overlapping P gene ORF conserved among thevesiculoviruses (Spiropoulou and Nichol, 1993; Kretzschmaret al., 1996). This additional protein is expressed during VSVinfection in tissue culture, but it appears to be dispensable forvirus replication and a role in viral pathogenesis or vector trans-mission has been proposed (Kretzschmar et al., 1996). It isunknown if this internal P gene ORF is expressed in plant orinsect hosts during LNYV infection. Overlapping +1 ORFs havealso been identified in NCMV, MFSV and MMV, but appear tobe absent from the P genes of SYNV and RYSV (Callaghan,2005).
Genes located between the P and M genes are found inaila(d(tmmpstHomeeIc
rhahwaFrit
reducing and non-reducing condition (Dietzgen and Francki,1988) and a detailed examination of the amino acid sequence(Callaghan, 2005).
The phylogeny of rhabdovirus L proteins suggests a mono-phyletic origin (Hogenhout et al., 2003). Based on phylogeneticanalysis of the conserved L gene polymerase module, plant rhab-doviruses appear to be evolutionarily closely related, but with aclear differentiation into two sister clades. This grouping con-firms the current two genera of plant rhabdoviruses, which isbased on the site of virus replication and maturation. Futureavailability of additional plant rhabdovirus sequences as well asknowledge of sequence variability between isolates (Callaghanand Dietzgen, 2005; Klerks et al., 2004; Revill et al., 2005) canbe expected to lead to a better understanding of the evolution ofthis important group of viruses.
Acknowledgements
We thank Margaret Bernard for technical assistance and PeterRevill for provision of TaVCV sequence data prior to publi-cation. This work was supported by the Australian ResearchCouncil, Horticulture Australia Limited and the Department ofPrimary Industries and Fisheries, Queensland.
References
B
C
C
C
C
D
D
D
F
H
H
H
ll plant rhabdoviruses analysed to date, but not in vertebrate-nfecting rhabdoviruses. In the LNYV genome the 4b gene wasocated in this position, as were the genes that encode the sc4nd P3 proteins in the SYNV and RYSV genomes, respectivelyScholthof et al., 1994; Huang et al., 2003). Evidence from ouratabase searches and refined secondary structure predictionshttp://opbs.okstate.edu/Virevol/web/Rhabdo.html) suggest thathe 4b protein belongs to the 30 kDa superfamily of plant viral
ovement proteins (Melcher, 2000), most similar to the move-ent proteins of capillo- and trichoviruses. Secondary structure
redictions for the SYNV sc4 protein and RYSV P3 also showome similarities to the 30 kDa superfamily, but distinct fromhat of the LNYV 4b protein (Melcher, personal communication;uang et al., 2005). The membrane and cell wall associationsf SYNV sc4 protein also suggest a role in cell-to-cell move-ent (Scholthof et al., 1994; Goodin et al., 2002) and recent
xperimental evidence suggests that RYSV P3 has some prop-rties typical for viral movement proteins (Huang et al., 2005).t therefore appears likely that the 4b protein has a role in LNYVell-to-cell movement.
The LNYV G gene encoded a protein with features typical ofhabdoviral glycoproteins, including three glycosylation sites, aydrophobic, N-terminal signal peptide for membrane targetingnd putative C-terminal transmembrane domain. The G proteinas previously been shown to be glycosylated in N. glutinosaith a complex network of oligosaccharides containing �-N-
cetylchitobiose N-linked to asparagine residues (Dietzgen andrancki, 1988). The assumption that some of the 18 cysteineesidues in the G protein sequence may form disulphide bondss supported by previously observed differences in the migra-ion rate of the G protein from purified LNYV particles under
arr, J.N., Wertz, G.W., 2001. Polymerase slippage at vesicular stomatitisvirus gene junctions to generate poly (A) is regulated by the upstream 3′-AUAC05′ tetranucleotide: implications for the mechanism of transcriptiontermination. J. Virol. 75, 6901–6913.
allaghan, B., 2005. Sequence analysis and variability study of Lettucenecrotic yellows virus. Ph.D thesis. The University of Queensland, Bris-bane, Australia.
allaghan, B., Dietzgen, R.G., 2005. Nucleocapsid gene variability revealstwo subgroups of Lettuce necrotic yellows virus. Arch. Virol. 150,1661–1667.
asanova, J.L., Pannetier, C., Jaulin, C., Kourilsky, P., 1990. Optimal condi-tions for directly sequencing double-stranded PCR products with Seque-nase. Nucleic Acids Res. 18, 4028.
hoi, T.J., Wagner, J.D., Jackson, A.O., 1994. Sequence analysis of the trailerregion of sonchus yellow net virus genomic RNA. Virology 185, 32–38.
ietzgen, R.G., 1995. Rhabdoviridae. In: Kohmoto, K., Singh, RP., Singh,US., Zeigler, R. (Eds.), Pathogenesis and Host–Parasite Specificity inPlant Diseases: Histopathological, Biochemical, Genetic and MolecularBasis, vol. 3. Pergamon Press, Oxford, pp. 177–197.
ietzgen, R.G., Francki, R.I.B., 1988. Analysis of lettuce necrotic yellowsvirus structural proteins with monoclonal antibodies and concanavalin A.Virology 166, 486–494.
ietzgen, R.G., Hunter, B.G., Francki, R.I.B., Jackson, A.O., 1989. Cloningof lettuce necrotic yellows virus RNA and identification of virus-specificpolyadenylated RNAs in infected Nicotiana glutinosa leaves. J. Gen.Virol. 70, 2299–2307.
rancki, R.I.B., Randles, J.W., Dietzgen, R.G., 1989. Lettuce necrotic yellowsvirus. In “AAB Descriptions of Plant Viruses” No. 343.
iggins, C.M., Dietzgen, R.G., 2000. Genetic transformation, regenerationand analysis of transgenic peanut. ACIAR Technical Report no. 48, 86pp.
ogenhout, S.A., Redinbaugh, M.G., Ammar, E.D., 2003. Plant and animalrhabdovirus host range: a bug’s view. Trends Microbiol. 11, 264–271.
uang, Y., Zhao, H., Luo, Z., Chen, X., Fang, R.-X., 2003. Novel structureof the genome of Rice yellow stunt virus: identification of the gene 6-encoded virion protein. J. Gen. Virol. 84, 2259–2264.

22 R.G. Dietzgen et al. / Virus Research 118 (2006) 16–22
Huang, Y.-W., Geng, Y.-F., Ying, X.-B., Chen, X.-Y., Fang, R.-X., 2005.Identification of a movement protein of rice yellow stunt rhabdovirus. J.Virol. 79, 2108–2114.
Goodin, M.M., Dietzgen, R.G., Schichnes, D., Ruzin, S., Jackson, A.O.,2002. pGD vectors: versatile tools for the expression of green and redfluorescent protein fusions in agroinfiltrated plant leaves. Plant J. 31,375–383.
Jackson, A.O., Dietzgen, R.G., Goodin, M.M., Bragg, J.N., Deng, M., 2005.Biology of plant rhabdoviruses. Annu. Rev. Phytopathol. 43, 623–660.
Klerks, M.M., Lindner, J.L., Vaskova, D., Spak, J., Thompson, J.R., Jelkmann,W., Schoen, C.D., 2004. Detection and tentative grouping of Strawberrycrinkle virus isolates. Eur. J. Plant Pathol. 110, 45–52.
Kretzschmar, E., Peluso, R., Schnell, M.J., Whitt, M.A., Rose, J.K., 1996.Normal replication of vesicular stomatitis virus without C proteins. Virol-ogy 216, 309–316.
Luo, Z.L., Fang, R.-X., 1998. Structure analysis of the rice yellow stunt rhab-dovirus glycoprotein gene and its mRNA. Arch. Virol. 143, 2453–2459.
Melcher, U., 2000. The ‘30K’ superfamily of viral movement proteins. J.Gen. Virol. 81, 257–266.
Muller, R., Poch, O., Delarue, M., Bishop, D.H., Bouloy, M., 1994. RiftValley fever virus L segment: correction of the sequence and possiblefunctional role of newly identified regions conserved in RNA-dependentpolymerases. J. Gen. Virol. 75, 1345–1352.
Poch, O., Sauvaget, I., Delarue, M., Tordo, N., 1989. Identification of fourconserved motifs among the RNA-dependent polymerase encoding ele-ments. EMBO J. 8, 3867–3874.
Revill, P., Trinh, X., Dale, J., Harding, R., 2005. Taro vein chlorosis virus:characterization and variability of a new nucleorhabdovirus. J. Gen. Virol.86, 491–499.
Rezaian, M.A., Heaton, L.A., Pederson, K., Milner, J.J., Jackson, A.O., 1983.Size and complexity of polyadenylated RNAs induced in tobacco infectedwith sonchus yellow net virus. Virology 131, 221–229.
Sambrook, J., Russell, D.W., 2001. Molecular cloning: a laboratory manual,third ed. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY.
Scholthof, K.-B.G., Hillman, B.I., Modrell, B., Heaton, L.A., Jackson, A.O.,1994. Characterization and detection of sc4: a sixth gene encoded bysonchus yellow net virus. Virology 204, 279–288.
Spiropoulou, C.F., Nichol, S.T., 1993. A small highly basic protein is encodedin overlapping frame within the P gene of vesicular stomatitis virus. J.Virol. 67, 3103–3110.
Sward, R.J., 1990. Lettuce necrotic yellows rhabdovirus and other virusesinfecting garlic. Australas. Plant Path. 19, 46–51.
Tanno, F., Nakatsu, A., Toriyama, S., Kojima, M., 2000. Complete nucleotidesequence of northern cereal mosaic virus and its genome organization.Arch. Virol. 145, 1373–1384.
Tordo, N., Benmansour, A., Calisher, C., Dietzgen, R.G., Fang, R.-X., Jack-son, A.O., Kurath, G., Nadin-Davies, S., Tesh, R.B., Walker, P.J., 2005.Family Rhabdoviridae Virus Taxonomy, Classification and Nomenclatureof Viruses. Academic Press, San Diego.
Tsai, C.-W., Redinbaugh, M.G., Willie, K.J., Reed, S., Goodin, M., Hogen-hout, S.A., 2005. Complete genome sequence and in planta subcellularlocalization of maize fine streak virus proteins. J. Virol. 79, 5304–5314.
Wetzel, T., Dietzgen, R.G., Dale, J.L., 1994a. Genomic organization of lettucenecrotic yellows rhabdovirus. Virology 200, 401–412.
Wetzel, T., Dietzgen, R.G., Geering, A.D.W., Dale, J.L., 1994b. Analysis ofthe nucleocapsid gene of lettuce necrotic yellows rhabdovirus. Virology202, 1054–1057.





























