Embed Size (px)
Citation preview

Current Medicinal Chemistry, 2006, 13, 87-98 87
0929-8673/06 $50.00+.00 © 2006 Bentham Science Publishers Ltd.
Comparative Studies of the Antioxidant Effects of Cis- and Trans-Resveratrol
F. Orallo*
Departamento de Farmacología, Facultad de Farmacia, Universidad de Santiago de Compostela, Santiago deCompostela (La Coruña), Spain
Abstract: Resveratrol (3,4',5-trihydroxystilbene, RESV) is a natural phenolic compound that exists as cis andtrans isomers [c-RESV or (Z)-RESV and t-RESV or (E)-RESV, respectively]. t-RESV is a natural component ofVitis vinifera L. (Vitaceae), abundant in the skin of grapes (but not in the flesh) and in the leaf epidermis, andpresent in wines, especially red wines. In in vitro, ex vivo and in vivo experiments t-RESV exhibits a number ofbiological activities, including anti-inflammatory and anticarcinogenic properties. RESV also exists in winesas a cis isomer, which (unlike t-RESV) is not currently available commercially; as a result, little is knownabout this isomer’s pharmacological activity. In this review, I will focus on the few comparative studies of theantioxidant effects of the two RESV isomers in different experimental models.
Keywords: Resveratrol isomers, NAD(P)H oxidase, xanthine oxidase, inducible nitric oxide synthase, murine macrophages,antioxidant activity, free radicals, wine.
1. INTRODUCTION
Resveratrol [3,4',5-trihydroxystilbene, RESV, (Fig. 1)]is a natural phenolic compound that exists as cis and transisomers [the c-RESV or (Z)-RESV diasteromer, and t-RESVor (E)-RESV diasteromer, respectively], facilitated by thedouble bond in its chemical structure [1-3].
t-RESV is a natural component of Vitis vinifera L.(Vitaceae) that is abundant in the skin of grapes (but not inthe flesh) and in the leaf epidermis, and that is present inwines, especially red wines. t-RESV is not unique to Vitisbut is also present in at least 72 other plant species(distributed in 12 families and 31 genera, e.g. Veratrum,Arachis, Morus, Vaccinium and Trifolium), some of whichare components of the human diet, such as mulberries andpeanuts [1,4-6].
t-RESV was first identified in 1940 by Michio Takaoka[7] as a constituent of the roots of white hellebore [Veratrumgrandiflorum (Maxim. ex Baker) Loes. (Liliaceae)], and later(in 1963) found by Nonomura et al. [8] (together with its 3-O-β-D-glucoside β-glucoside, so-called trans-piceid or trans-polydatin) in the dried roots of Japanese knotweedPolygonum cuspidatum Sieb. et Zucc. (Polygonaceae), calledKo-jo-kon (or Itadori) in Japanese and Chinese folkmedicine, in which it is used for the treatment ofsuppurative dermatitis, gonorrhea favus, hyperlipemia,favus, athlete's foot (tinea pedis), arteriosclerosis, as well asin allergic and inflammatory diseases and other pathologies[see, e.g., refs. 1-4]. Later, in 1976, t-RESV was detected ingrapevines by Langcake and Pryce [9], who found that it issynthesized by leaf tissues in response to fungal infection(mainly by Botrytis cinerea) or exposure to ultraviolet light.
*Address correspondence to this author at the Departamento deFarmacología, Facultad de Farmacia, Universidad de Santiago deCompostela, Campus Universitario Sur, E-15782 Santiago de Compostela(La Coruña), Spain; Tel: +34-981-563100; Ext: 14895. Fax: +34-981-594595; E-mail: [email protected]
In 1992, Siemann and Creasy [10] reported the presenceof this polyphenol in wines (basically red wine). At thesame time, a number of large-scale epidemiological studieswere clearly suggesting that prolonged moderateconsumption of wine (especially red wine) by the SouthernFrench and other Mediterranean populations was associatedwith a very low incidence of cardiovascular diseases, notablycoronary heart disease, despite a high-fat diet, little exerciseand widespread smoking: this was the so-called “Frenchparadox” [see, e.g., ref. 11]. When it became known that theprotective effects of wine consumption were independent ofalcohol content, a number of studies were initiated with theaim of identifying the component(s) responsible.
Since then, the pharmacological activity of t-RESV hasbeen extensively investigated. In in vitro, ex vivo and in vivoexperiments t-RESV has shown a number of biologicalactivities including anti-inflammatory and anticarcinogenicproperties [for reviews, see, e.g., refs. 1,3,4,6,12-14].Several studies within the last few years have demonstratedthat t-RESV may protect against coronary heart disease as aresult of different effects, including significant antioxidantactivity, modulation of lipoprotein metabolism, andvasodilatory and platelet antiaggregatory properties [see,e.g., refs. 12,15-18].
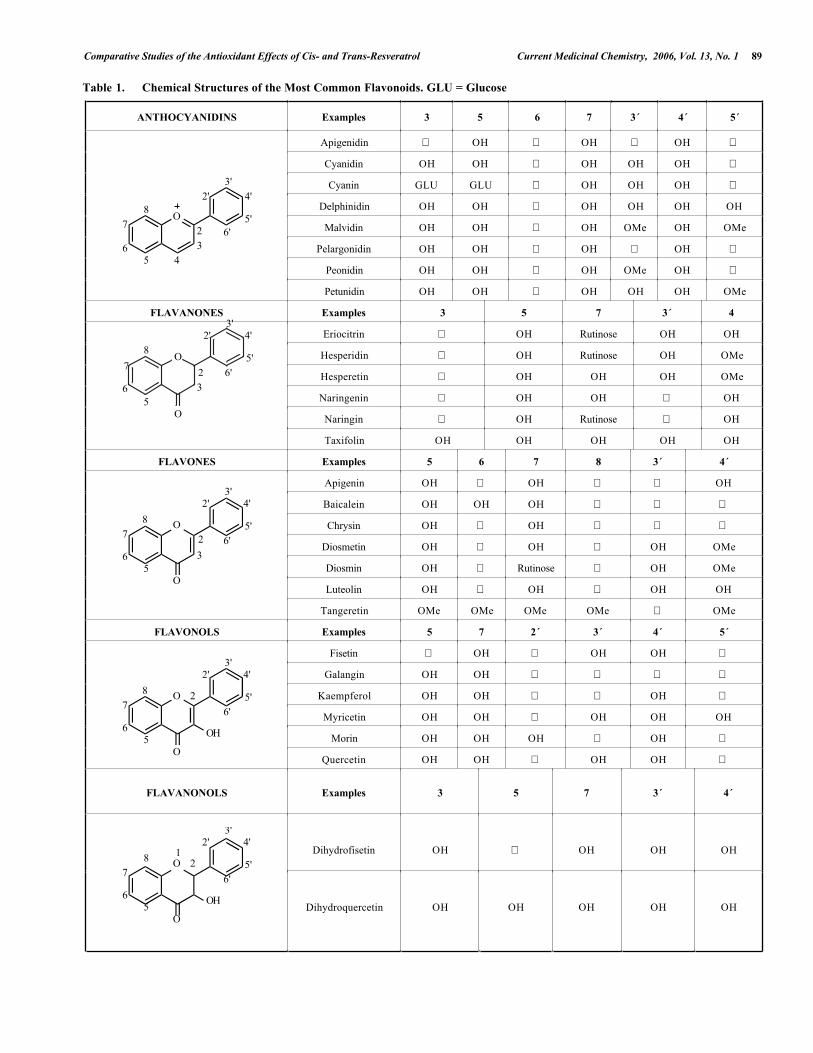
Besides t-RESV, other polyphenolic compounds (the so-called flavonoids) have been implicated in thecardioprotective effects of moderate wine consumption [19].Flavonoids are a large group of polyphenols present inplants, regularly consumed foods (e.g. vegetables and fruits),and beverages like tea and wine. These low molecular weightsubstances are phenylbenzo-γ-pyrones (phenyl-γ-chromones)with an assortment of structures based on a common three-ring nucleus. They are usually subdivided according to theirsubstituents into several subclasses includinganthocyanidins, flavanones, flavones, flavonols, flavanonols(or dihydroflavonols), chalcones, isoflavones and flavanols(also called catechins) (Table 1). The biological activities offlavonoids (including their antioxidant properties) have been
88 Current Medicinal Chemistry, 2006, Vol. 13, No. 1 F. Orallo
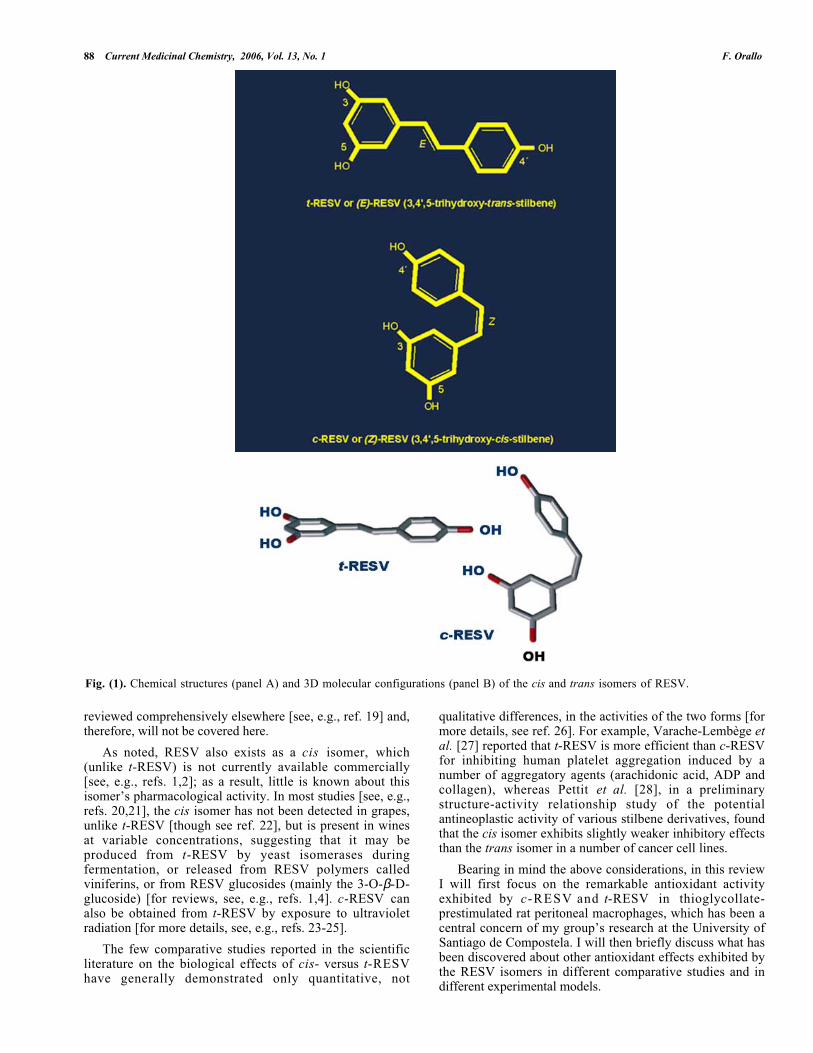
Fig. (1). Chemical structures (panel A) and 3D molecular configurations (panel B) of the cis and trans isomers of RESV.
reviewed comprehensively elsewhere [see, e.g., ref. 19] and,therefore, will not be covered here.
As noted, RESV also exists as a cis isomer, which(unlike t-RESV) is not currently available commercially[see, e.g., refs. 1,2]; as a result, little is known about thisisomer’s pharmacological activity. In most studies [see, e.g.,refs. 20,21], the cis isomer has not been detected in grapes,unlike t-RESV [though see ref. 22], but is present in winesat variable concentrations, suggesting that it may beproduced from t-RESV by yeast isomerases duringfermentation, or released from RESV polymers calledviniferins, or from RESV glucosides (mainly the 3-O-β-D-glucoside) [for reviews, see, e.g., refs. 1,4]. c-RESV canalso be obtained from t-RESV by exposure to ultravioletradiation [for more details, see, e.g., refs. 23-25].
The few comparative studies reported in the scientificliterature on the biological effects of cis- versus t-RESVhave generally demonstrated only quantitative, not
qualitative differences, in the activities of the two forms [formore details, see ref. 26]. For example, Varache-Lembège etal. [27] reported that t-RESV is more efficient than c-RESVfor inhibiting human platelet aggregation induced by anumber of aggregatory agents (arachidonic acid, ADP andcollagen), whereas Pettit et al. [28], in a preliminarystructure-activity relationship study of the potentialantineoplastic activity of various stilbene derivatives, foundthat the cis isomer exhibits slightly weaker inhibitory effectsthan the trans isomer in a number of cancer cell lines.
Bearing in mind the above considerations, in this reviewI will first focus on the remarkable antioxidant activityexhibited by c-RESV and t-RESV in thioglycollate-prestimulated rat peritoneal macrophages, which has been acentral concern of my group’s research at the University ofSantiago de Compostela. I will then briefly discuss what hasbeen discovered about other antioxidant effects exhibited bythe RESV isomers in different comparative studies and indifferent experimental models.
Comparative Studies of the Antioxidant Effects of Cis- and Trans-Resveratrol Current Medicinal Chemistry, 2006, Vol. 13, No. 1 89
Table 1. Chemical Structures of the Most Common Flavonoids. GLU = Glucose
ANTHOCYANIDINS Examples 3 5 6 7 3´ 4´ 5´
Apigenidin OH OH OH
Cyanidin OH OH OH OH OH
Cyanin GLU GLU OH OH OH
Delphinidin OH OH OH OH OH OH
Malvidin OH OH OH OMe OH OMe
Pelargonidin OH OH OH OH
O8
7
65 4
3
2'3'
4'
5'6'2
Peonidin OH OH OH OMe OH
Petunidin OH OH OH OH OH OMe
FLAVANONES Examples 3 5 7 3´ 4
Eriocitrin OH Rutinose OH OH
Hesperidin OH Rutinose OH OMe
Hesperetin OH OH OH OMe
Naringenin OH OH OH
O8
7
65
3
O
2
2'3'
4'
5'6'
Naringin OH Rutinose OH
Taxifolin OH OH OH OH OH
FLAVONES Examples 5 6 7 8 3´ 4´
Apigenin OH OH OH
Baicalein OH OH OH
Chrysin OH OH
Diosmetin OH OH OH OMe
Diosmin OH Rutinose OH OMe
O87
65
O
2'3'
4'
5'6'
3
2
Luteolin OH OH OH OH
Tangeretin OMe OMe OMe OMe OMe
FLAVONOLS Examples 5 7 2´ 3´ 4´ 5´
Fisetin OH OH OH
Galangin OH OH
Kaempferol OH OH OH
Myricetin OH OH OH OH OH
Morin OH OH OH OH
O87
65
O
2
2'3'
4'
5'6'
OH
Quercetin OH OH OH OH
FLAVANONOLS Examples 3 5 7 3´ 4´
Dihydrofisetin OH OH OH OHO8
7
65
O
2
2'3'
4'
5'6'
OH
1
Dihydroquercetin OH OH OH OH OH
90 Current Medicinal Chemistry, 2006, Vol. 13, No. 1 F. Orallo
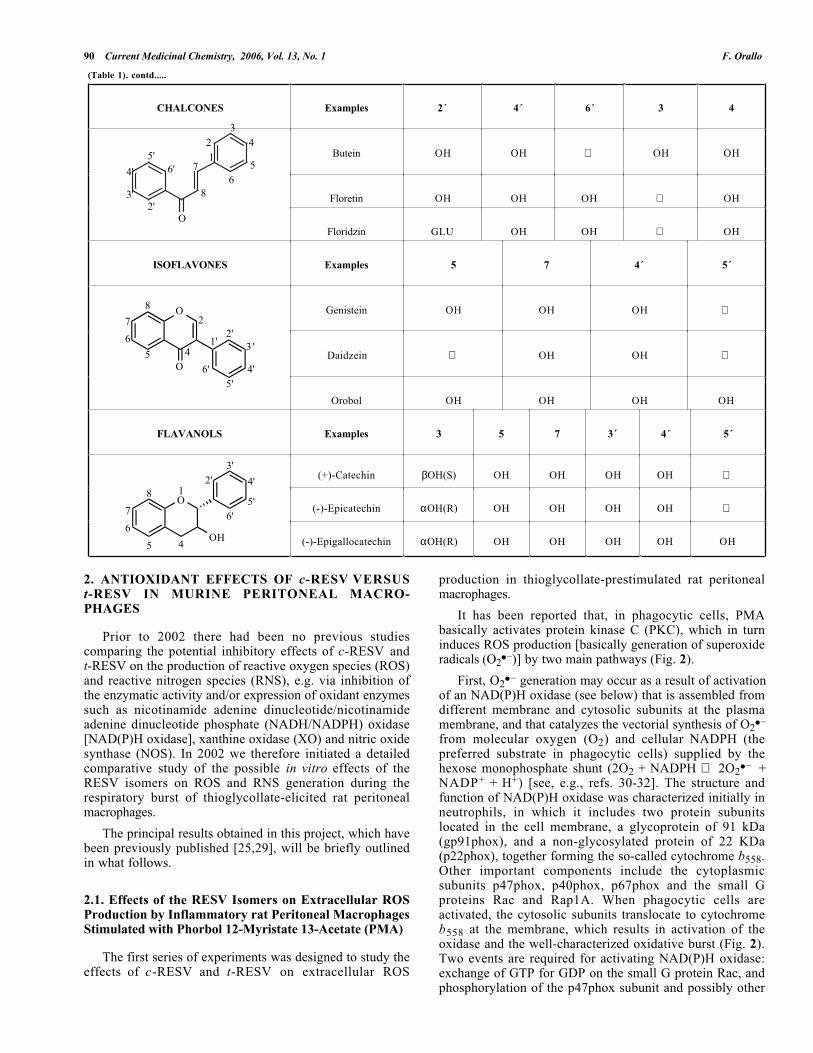
(Table 1). contd.....
CHALCONES Examples 2´ 4´ 6´ 3 4
Butein OH OH OH OH
Floretin OH OH OH OH
5'
4'
3'2'
23
4
56
17
8
O
6'
Floridzin GLU OH OH OH
ISOFLAVONES Examples 5 7 4´ 5´
Genistein OH OH OH
Daidzein OH OH
O
O
1'2'
3 '
4'5'
6'
2
45
6
7
8
Orobol OH OH OH OH
FLAVANOLS Examples 3 5 7 3´ 4´ 5´
(+)-Catechin βOH(S) OH OH OH OH
(-)-Epicatechin αOH(R) OH OH OH OH O
OH
2'3'
4'
5'6'
1
45
6
7
8
(-)-Epigallocatechin αOH(R) OH OH OH OH OH
2. ANTIOXIDANT EFFECTS OF c-RESV VERSUSt-RESV IN MURINE PERITONEAL MACRO-PHAGES
Prior to 2002 there had been no previous studiescomparing the potential inhibitory effects of c-RESV andt-RESV on the production of reactive oxygen species (ROS)and reactive nitrogen species (RNS), e.g. via inhibition ofthe enzymatic activity and/or expression of oxidant enzymessuch as nicotinamide adenine dinucleotide/nicotinamideadenine dinucleotide phosphate (NADH/NADPH) oxidase[NAD(P)H oxidase], xanthine oxidase (XO) and nitric oxidesynthase (NOS). In 2002 we therefore initiated a detailedcomparative study of the possible in vitro effects of theRESV isomers on ROS and RNS generation during therespiratory burst of thioglycollate-elicited rat peritonealmacrophages.
The principal results obtained in this project, which havebeen previously published [25,29], will be briefly outlinedin what follows.
2.1. Effects of the RESV Isomers on Extracellular ROSProduction by Inflammatory rat Peritoneal MacrophagesStimulated with Phorbol 12-Myristate 13-Acetate (PMA)
The first series of experiments was designed to study theeffects of c-RESV and t-RESV on extracellular ROS
production in thioglycollate-prestimulated rat peritonealmacrophages.
It has been reported that, in phagocytic cells, PMAbasically activates protein kinase C (PKC), which in turninduces ROS production [basically generation of superoxideradicals (O2
● –)] by two main pathways (Fig. 2).
First, O2● – generation may occur as a result of activation
of an NAD(P)H oxidase (see below) that is assembled fromdifferent membrane and cytosolic subunits at the plasmamembrane, and that catalyzes the vectorial synthesis of O2
● –
from molecular oxygen (O2) and cellular NADPH (thepreferred substrate in phagocytic cells) supplied by thehexose monophosphate shunt (2O2 + NADPH ⇒ 2O2
● – +NADP+ + H+) [see, e.g., refs. 30-32]. The structure andfunction of NAD(P)H oxidase was characterized initially inneutrophils, in which it includes two protein subunitslocated in the cell membrane, a glycoprotein of 91 kDa(gp91phox), and a non-glycosylated protein of 22 KDa(p22phox), together forming the so-called cytochrome b558.Other important components include the cytoplasmicsubunits p47phox, p40phox, p67phox and the small Gproteins Rac and Rap1A. When phagocytic cells areactivated, the cytosolic subunits translocate to cytochromeb558 at the membrane, which results in activation of theoxidase and the well-characterized oxidative burst (Fig. 2).Two events are required for activating NAD(P)H oxidase:exchange of GTP for GDP on the small G protein Rac, andphosphorylation of the p47phox subunit and possibly other
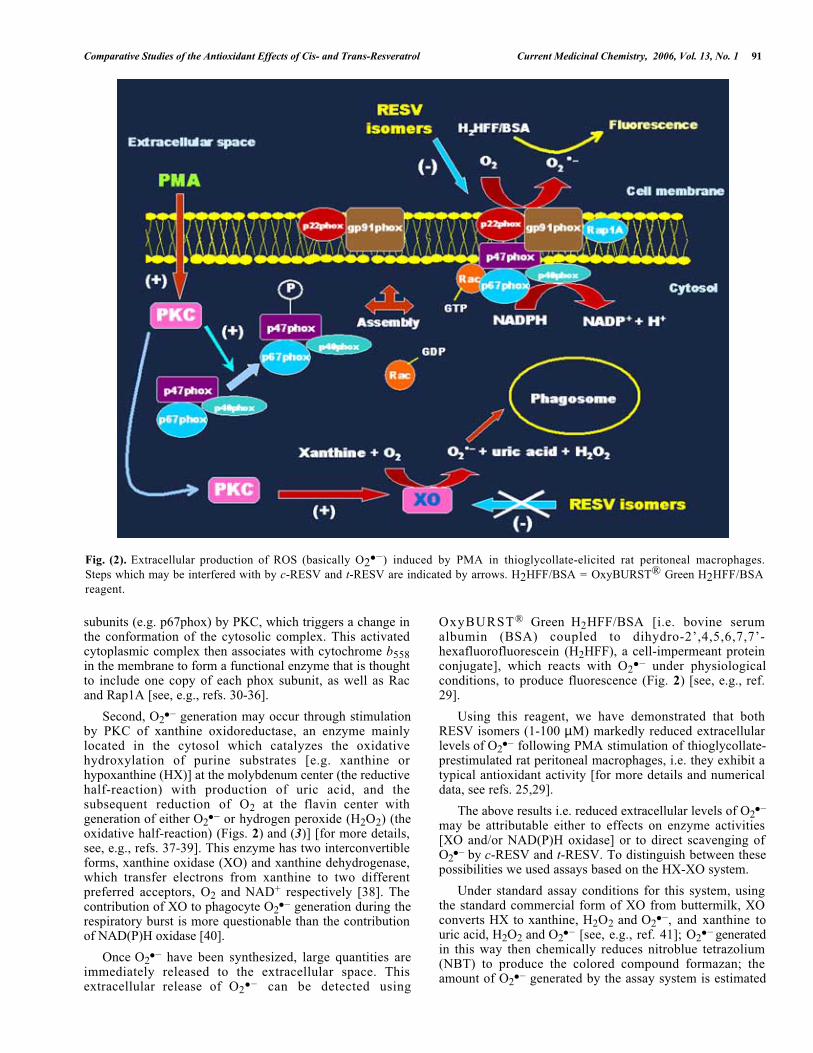
Comparative Studies of the Antioxidant Effects of Cis- and Trans-Resveratrol Current Medicinal Chemistry, 2006, Vol. 13, No. 1 91
Fig. (2). Extracellular production of ROS (basically O2● –) induced by PMA in thioglycollate-elicited rat peritoneal macrophages.Steps which may be interfered with by c-RESV and t-RESV are indicated by arrows. H2HFF/BSA = OxyBURST® Green H2HFF/BSAreagent.
subunits (e.g. p67phox) by PKC, which triggers a change inthe conformation of the cytosolic complex. This activatedcytoplasmic complex then associates with cytochrome b558in the membrane to form a functional enzyme that is thoughtto include one copy of each phox subunit, as well as Racand Rap1A [see, e.g., refs. 30-36].
Second, O2● – generation may occur through stimulation
by PKC of xanthine oxidoreductase, an enzyme mainlylocated in the cytosol which catalyzes the oxidativehydroxylation of purine substrates [e.g. xanthine orhypoxanthine (HX)] at the molybdenum center (the reductivehalf-reaction) with production of uric acid, and thesubsequent reduction of O2 at the flavin center withgeneration of either O2
● – or hydrogen peroxide (H2O2) (theoxidative half-reaction) (Figs. 2) and (3)] [for more details,see, e.g., refs. 37-39]. This enzyme has two interconvertibleforms, xanthine oxidase (XO) and xanthine dehydrogenase,which transfer electrons from xanthine to two differentpreferred acceptors, O2 and NAD+ respectively [38]. Thecontribution of XO to phagocyte O2
● – generation during therespiratory burst is more questionable than the contributionof NAD(P)H oxidase [40].
Once O2● – have been synthesized, large quantities are
immediately released to the extracellular space. Thisextracellular release of O2
● – can be detected using
OxyBURST® Green H2HFF/BSA [i.e. bovine serumalbumin (BSA) coupled to dihydro-2’,4,5,6,7,7’-hexafluorofluorescein (H2HFF), a cell-impermeant proteinconjugate], which reacts with O2
● – under physiologicalconditions, to produce fluorescence (Fig. 2) [see, e.g., ref.29].
Using this reagent, we have demonstrated that bothRESV isomers (1-100 µM) markedly reduced extracellularlevels of O2
● – following PMA stimulation of thioglycollate-prestimulated rat peritoneal macrophages, i.e. they exhibit atypical antioxidant activity [for more details and numericaldata, see refs. 25,29].
The above results i.e. reduced extracellular levels of O2● –
may be attributable either to effects on enzyme activities[XO and/or NAD(P)H oxidase] or to direct scavenging ofO2
● – by c-RESV and t-RESV. To distinguish between thesepossibilities we used assays based on the HX-XO system.
Under standard assay conditions for this system, usingthe standard commercial form of XO from buttermilk, XOconverts HX to xanthine, H2O2 and O2
● –, and xanthine touric acid, H2O2 and O2
● – [see, e.g., ref. 41]; O2● – generated
in this way then chemically reduces nitroblue tetrazolium(NBT) to produce the colored compound formazan; theamount of O2
● – generated by the assay system is estimated

92 Current Medicinal Chemistry, 2006, Vol. 13, No. 1 F. Orallo
Fig. (3). Summarized reaction steps in the HX/XO system. As noted in the text, our experiments indicate that neither c-RESV nort-RESV are scavengers of O2● – or inhibitors of commercial (buttermilk) XO.
on the basis of spectrophotometric determination offormazan at 560 nm (Fig. 3) [see, e.g., ref. 42]. In parallelassays, the production of uric acid by XO may be estimatedon the basis of spectrophotometric determination at 265 nm.When a test substance lowers the amount of O2
● – (i.e.decreases the rate of NBT reduction) and at the same timedoes not affect the formation of uric acid, it can beconsidered a selective scavenger of O2
● –; on the other hand,reduced levels of both O2
● – and uric acid indicate that thetest substance is inhibiting XO activity. In our experiments,and unlike the reference O2
● – scavenger superoxide dismutase(SOD, 0.1-10 U/ml), c-RESV and t-RESV (1-100 µM) didnot decrease the rate of NBT reduction. In addition, andunlike allopurinol (1-10 µM), a well-known referenceinhibitor of XO, neither c-RESV nor t-RESV (1-100 µM)had significant effects on uric acid production in this assaysystem [for more details and numerical data, see refs. 25,29].These results clearly indicate that c-RESV and t-RESV donot selectively scavenge O2
● – or directly inhibit commercialXO, suggesting that the reduction in extracellular O2
● – levelsinduced in inflammatory macrophages by the RESV isomersis due to inhibition of NAD(P) oxidase activity. Theseresults agree with those obtained by Hung et al. [43], whofound that low concentrations of t-RESV (10 µM) do notscavenge O2
● – generated by the HX/XO system. In contrast,our results do not agree with those of Zhou et al. [44], whofound that t-RESV inhibits the enzymatic activity of XOwith an IC50 of about 11 µM.
To confirm these findings, we performed experiments toassess the possible effects of the RESV isomers on XO andNAD(P)H oxidase activities (as measured bylucigenin-enhanced chemiluminescence) in extracts fromthioglycollate-prestimulated rat peritoneal macrophages. Inthese experiments, c-RESV and t-RESV (1-100 µM) [unlike
allopurinol (1-10 µM), a reference inhibitor of XO] had noeffects on O2
● – generation from xanthine (Fig. 2). These dataconfirm that c-RESV and t-RESV do not affect XO activity[in this case native XO, as opposed to commercial(buttermilk) XO]. However, both c-RESV and t-RESV (10-100 µM) [like diphenyleneiodonium (DPI, 10-100 µM), areference inhibitor of NAD(P)H oxidase] inhibited NAD(P)Hoxidase activity, i.e. the specific chemiluminescence signalemitted by the reaction between lucigenin and O2
● – generatedfrom NADPH (Fig. 2). These results strongly indicate that adecline in NAD(P)H oxidase activity may be responsible, atleast in part, for the observed inhibitory effects of the RESVisomers on extracellular ROS (basically O2
● –) production[for more details and numerical data, see ref. 25].
We have observed similar effects of c-RESV and t-RESVon NAD(P)H oxidase activity in rat aortic homogenates[45,46]. In addition, we have recently obtained similarresults using rat aortic myocytes and human umbilical-veinendothelial cells as sources of enzymes (unpublished data).These data agree with those obtained by Liu et al. [47], whofound that t-RESV inhibits mechanical-strain-inducedNAD(P)H oxidase activity in endothelial cells isolated fromhuman umbilical cords. However, our results disagree withthose obtained by Poolman et al. [48], who found thatt-RESV does not affect NAD(P)H oxidase activity in humanmonocytes (differentiated U937 cells).
2.2. Effects of c-RESV on Intracellular ROS Productionby Inflammatory Rat Macrophages Stimulated withKluyveromyces Lactis Cells
The second series of experiments was designed to studythe effects of the RESV isomers on intracellular ROSproduction in thioglycollate-prestimulated rat peritoneal
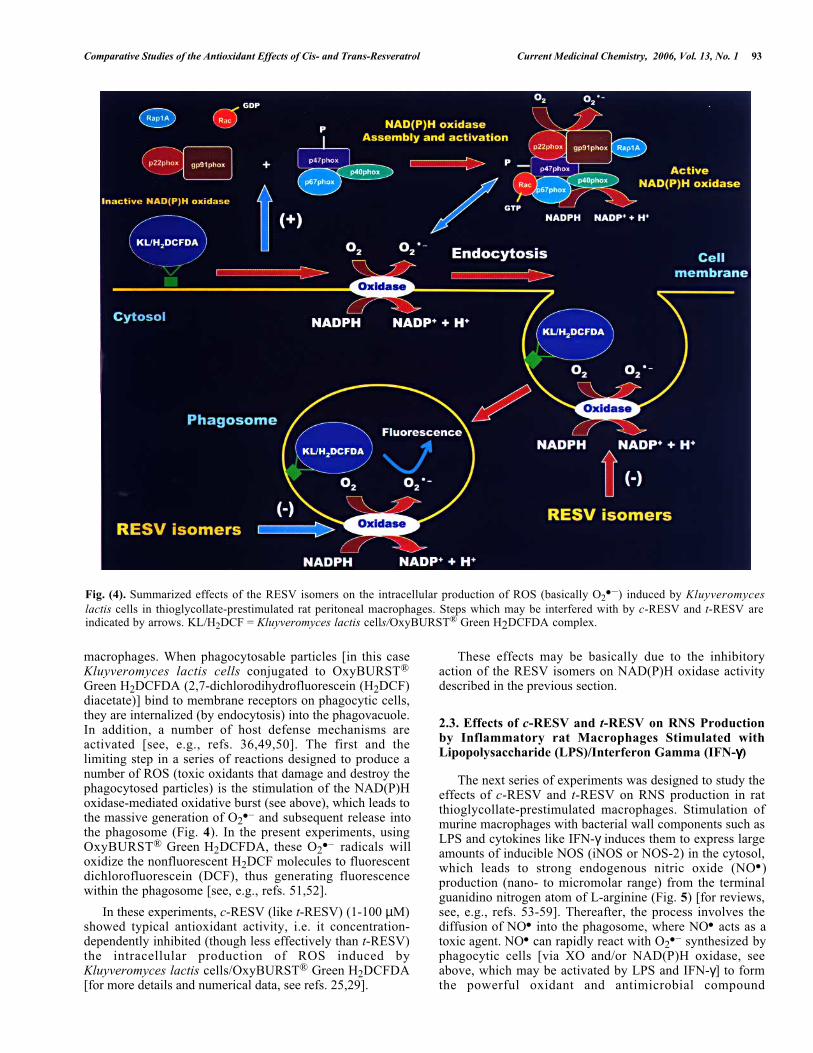
Comparative Studies of the Antioxidant Effects of Cis- and Trans-Resveratrol Current Medicinal Chemistry, 2006, Vol. 13, No. 1 93
Fig. (4). Summarized effects of the RESV isomers on the intracellular production of ROS (basically O2● –) induced by Kluyveromyces
lactis cells in thioglycollate-prestimulated rat peritoneal macrophages. Steps which may be interfered with by c-RESV and t-RESV areindicated by arrows. KL/H2DCF = Kluyveromyces lactis cells/OxyBURST® Green H2DCFDA complex.
macrophages. When phagocytosable particles [in this caseKluyveromyces lactis cells conjugated to OxyBURST®
Green H2DCFDA (2,7-dichlorodihydrofluorescein (H2DCF)diacetate)] bind to membrane receptors on phagocytic cells,they are internalized (by endocytosis) into the phagovacuole.In addition, a number of host defense mechanisms areactivated [see, e.g., refs. 36,49,50]. The first and thelimiting step in a series of reactions designed to produce anumber of ROS (toxic oxidants that damage and destroy thephagocytosed particles) is the stimulation of the NAD(P)Hoxidase-mediated oxidative burst (see above), which leads tothe massive generation of O2
● – and subsequent release intothe phagosome (Fig. 4). In the present experiments, usingOxyBURST® Green H2DCFDA, these O2
● – radicals willoxidize the nonfluorescent H2DCF molecules to fluorescentdichlorofluorescein (DCF), thus generating fluorescencewithin the phagosome [see, e.g., refs. 51,52].
In these experiments, c-RESV (like t-RESV) (1-100 µM)showed typical antioxidant activity, i.e. it concentration-dependently inhibited (though less effectively than t-RESV)the intracellular production of ROS induced byKluyveromyces lactis cells/OxyBURST® Green H2DCFDA[for more details and numerical data, see refs. 25,29].
These effects may be basically due to the inhibitoryaction of the RESV isomers on NAD(P)H oxidase activitydescribed in the previous section.
2.3. Effects of c-RESV and t-RESV on RNS Productionby Inflammatory rat Macrophages Stimulated withLipopolysaccharide (LPS)/Interferon Gamma (IFN-γγγγ)
The next series of experiments was designed to study theeffects of c-RESV and t-RESV on RNS production in ratthioglycollate-prestimulated macrophages. Stimulation ofmurine macrophages with bacterial wall components such asLPS and cytokines like IFN-γ induces them to express largeamounts of inducible NOS (iNOS or NOS-2) in the cytosol,which leads to strong endogenous nitric oxide (NO● )production (nano- to micromolar range) from the terminalguanidino nitrogen atom of L-arginine (Fig. 5) [for reviews,see, e.g., refs. 53-59]. Thereafter, the process involves thediffusion of NO● into the phagosome, where NO● acts as atoxic agent. NO● can rapidly react with O2
● – synthesized byphagocytic cells [via XO and/or NAD(P)H oxidase, seeabove, which may be activated by LPS and IFN-γ] to formthe powerful oxidant and antimicrobial compound
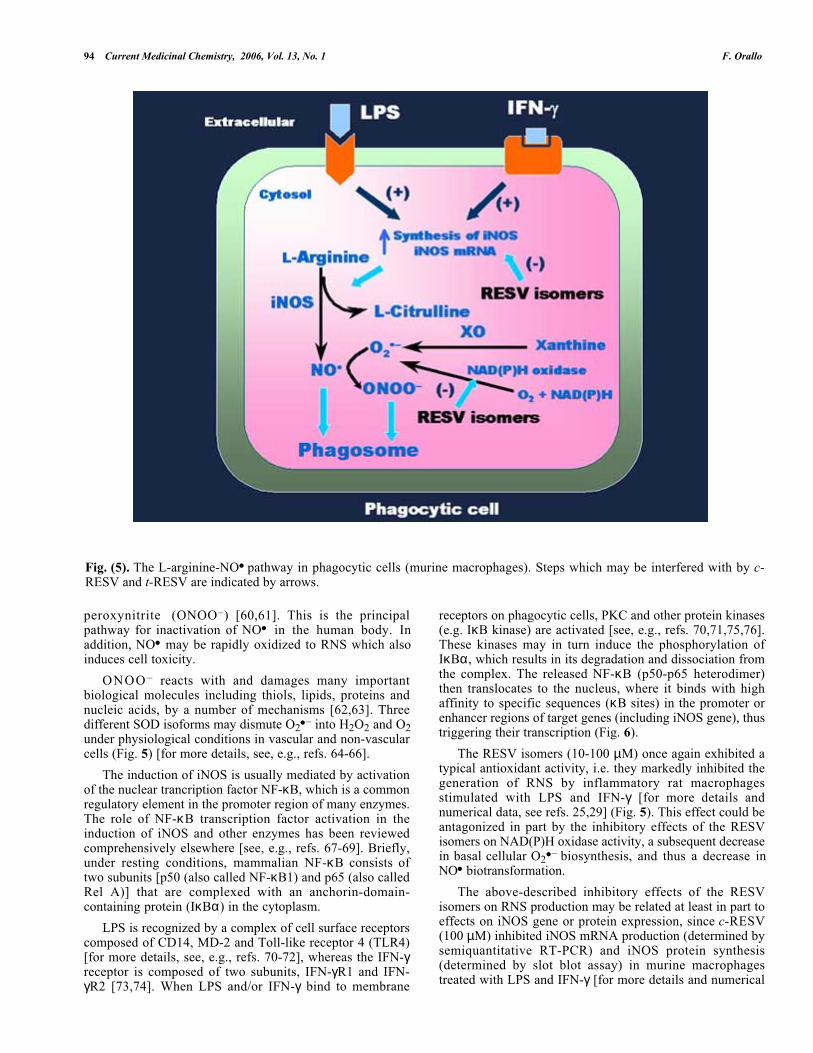
94 Current Medicinal Chemistry, 2006, Vol. 13, No. 1 F. Orallo
Fig. (5). The L-arginine-NO● pathway in phagocytic cells (murine macrophages). Steps which may be interfered with by c-RESV and t-RESV are indicated by arrows.
peroxynitrite (ONOO–) [60,61]. This is the principalpathway for inactivation of NO● in the human body. Inaddition, NO● may be rapidly oxidized to RNS which alsoinduces cell toxicity.
ONOO – reacts with and damages many importantbiological molecules including thiols, lipids, proteins andnucleic acids, by a number of mechanisms [62,63]. Threedifferent SOD isoforms may dismute O2
● – into H2O2 and O2under physiological conditions in vascular and non-vascularcells (Fig. 5) [for more details, see, e.g., refs. 64-66].
The induction of iNOS is usually mediated by activationof the nuclear trancription factor NF-κB, which is a commonregulatory element in the promoter region of many enzymes.The role of NF-κB transcription factor activation in theinduction of iNOS and other enzymes has been reviewedcomprehensively elsewhere [see, e.g., refs. 67-69]. Briefly,under resting conditions, mammalian NF-κB consists oftwo subunits [p50 (also called NF-κB1) and p65 (also calledRel A)] that are complexed with an anchorin-domain-containing protein (IκBα) in the cytoplasm.
LPS is recognized by a complex of cell surface receptorscomposed of CD14, MD-2 and Toll-like receptor 4 (TLR4)[for more details, see, e.g., refs. 70-72], whereas the IFN-γreceptor is composed of two subunits, IFN-γR1 and IFN-γR2 [73,74]. When LPS and/or IFN-γ bind to membrane
receptors on phagocytic cells, PKC and other protein kinases(e.g. IκB kinase) are activated [see, e.g., refs. 70,71,75,76].These kinases may in turn induce the phosphorylation ofIκBα, which results in its degradation and dissociation fromthe complex. The released NF-κB (p50-p65 heterodimer)then translocates to the nucleus, where it binds with highaffinity to specific sequences (κB sites) in the promoter orenhancer regions of target genes (including iNOS gene), thustriggering their transcription (Fig. 6).
The RESV isomers (10-100 µM) once again exhibited atypical antioxidant activity, i.e. they markedly inhibited thegeneration of RNS by inflammatory rat macrophagesstimulated with LPS and IFN-γ [for more details andnumerical data, see refs. 25,29] (Fig. 5). This effect could beantagonized in part by the inhibitory effects of the RESVisomers on NAD(P)H oxidase activity, a subsequent decreasein basal cellular O2
● – biosynthesis, and thus a decrease inNO● biotransformation.
The above-described inhibitory effects of the RESVisomers on RNS production may be related at least in part toeffects on iNOS gene or protein expression, since c-RESV(100 µM) inhibited iNOS mRNA production (determined bysemiquantitative RT-PCR) and iNOS protein synthesis(determined by slot blot assay) in murine macrophagestreated with LPS and IFN-γ [for more details and numerical

Comparative Studies of the Antioxidant Effects of Cis- and Trans-Resveratrol Current Medicinal Chemistry, 2006, Vol. 13, No. 1 95
Fig. (6). Hypothesized interference of c-RESV and t-RESV with the LPS and IFN-γ/NF-κB/NO● signal transduction pathway in murinemacrophages (for more details see text).
data, see ref. 25]. Similar results have been obtained for t-RESV by other authors [see, e.g., refs. 77-78] and by us(unpublished results).
These effects of the RESV isomers on iNOS gene andprotein expression may be mediated by inhibition of NF-κBactivation, mainly as a result of inhibition of the degradationand phosphorylation of IκBα , as previously reported for t-RESV by Murakami et al. [79] (Fig. 6). The precise effectsof c-RESV and t-RESV on the NF-κB signaling pathwayare currently being evaluated in our laboratory. Thepreliminary results obtained (basically using a DNAhybridization array containing 96 NF-κB-related genes)indicate that c-RESV provokes an increase in the IκBα geneexpression and a down-regulation of two genes of the NF-κB family, NF-κB1 (p50) and NF-κB2 (p52) [for moredetails, see ref. 80]. Similar results have been obtained witht-RESV (unpublished results).
3. DPPH-SCAVENGING ACTIVITY OF c-RESV ANDt-RESV
The potential capacity of c-RESV and t-RESV toscavenge free radicals other than O2
● – has been studied usingthe stable free radical 1,1-diphenyl-2-picryl-hydrazyl(DPPH). In these assays, the free-radical-scavenging activityof an antioxidant is evaluated by measurement of thedecrease in absorbance at 515 nm. Because of its oddelectron, the DPPH radical shows a strong absorbance bandat this visible-light wavelength (a deep purple color). When
this electron is paired off (by acceptance of an electron orhydrogen radical from an antioxidant compound), a stableDPPH-H molecule is generated, leading to a reduction inabsorbance (color change from purple to yellow) proportionalto the number of electrons taken up [see, e.g., refs. 81,82].
Several authors have studied the free-radical-scavengingproperties of t-RESV and c-RESV using DPPH assay. Baslyet al. [83] found that c-RESV is more efficient than t-RESVas a scavenger of DPPH. However, this finding contradictsthe results of Fauconneau et al. [84] and Waffo-Teguo et al.[85], who found that c-RESV is less efficient than t-RESVfor scavenging DPPH, as well as the results of Stivala et al.[24], who found that the two isomers scavenge DPPH withsimilar efficiency.
4. EFFECTS OF c-RESV AND t-RESV ON LIPIDPEROXIDATION
Lipid peroxidation is initiated by the attack on a fattyacid or fatty acyl side chain by any chemical species (usuallya free radical) that has sufficient reactivity to abstract ahydrogen atom from a methylene carbon in the side chain.
Most biological studies of lipid peroxidation involvetransition metal ions, added to the reaction mixtures. Whenferrous ions, cuprous ions, or certain chelates of these ions(e.g. Fe2+-ADP) are added to liposomes, lipoproteins,cultured cells or isolated biological membranes (such asmicrosomes, mitochondrias, plasma-membrane fractions,

96 Current Medicinal Chemistry, 2006, Vol. 13, No. 1 F. Orallo
etc.), peroxidation occurs [see, e.g., refs. 86,87]. Theoxidized forms of these transition metal ions (e.g. Fe3+,Cu2+) can also accelerate peroxidation if a reducing agent(e.g. ascorbate) is present. Free radical generators such as2,2'-azobis(2-amidinopropane) dihydrochloride (AAPH) [88]and tert-butylhydroperoxide (TBHP) [89] are also widelyused in assays of this type.
A number of free radicals (mainly peroxyl radicals whichthen initiate peroxidation) can usually be detected in thesevarious reaction mixtures [for more details, see, e.g., ref.87].
Several authors have studied the effects of the isomers ofRESV on lipid peroxidation under different experimentalconditions. Fauconneau et al. [84] found that t-RESV ismore effective than c-RESV for reducing Fe2+-induced lipidperoxidation in rat liver microsomes and Cu2+-induced lipidperoxidation in human low-density lipoproteins (LDL).Similar results were obtained by Waffo-Teguo et al. [85] inhuman LDL, by Belguendouz et al. [23] in porcine LDL,and by Stivala et al. [24] in rat liver microsomes (usingFe2+-ascorbate in place of Fe2+ alone) and in culturednormal human fibroblasts (using TBHP). By contrast,Belguendouz et al. [23] found that c-RESV and t-RESVinhibit AAPH-mediated porcine LDL oxidation with similarpotency. These antioxidant effects of the RESV isomers maybe due to one or more of the following mechanisms:
1. Direct quelation by RESV of metal ions (e.g. Cu2+)[see refs. 1,23];
2. Inhibition by RESV of free radical generation bymetal ions (e.g. Fe2+) or other generators (e.g.TBHP);
3. Scavenging of the free radicals by RESV.
Further studies are required to clarify the precisemechanisms by which the RESV isomers inhibit lipidperoxidation.
5. OTHER ANTIOXIDANT EFFECTS OF c-RESVVERSUS t-RESV
Finally, Stivala et al. [24] have studied the possibleantioxidant effects of the RESV isomers using citronealthermo-oxidation. In this assay, the aldehyde (-)-citronellalis used as the oxidation substrate: it is subjected to heatingand intensive oxygenation in chlorobenzene (which acts asan oxidant agent), and its disappearance with the consequentformation of degradation products is monitored by gaschromatography. The antioxidant efficacy of the RESVisomers was measured by determining the efficient quantity(EQ), i.e. the concentration required for each isomer todouble the half-life of citronellal with respect to the controlreaction (citronellal without antioxidant). The resultsindicated that t-RESV inhibited citroneal thermo-oxidationmore effectively than c-RESV (EQ = 135 ± 8.81 and 241 ±38.0 µM, respectively).
6. CONCLUDING REMARKS
In summary, the studies described in this review indicatethat c-RESV and t-RESV at micromolar concentrations
significantly interfere with macrophage function.Specifically, both c-RESV and t-RESV exhibit typicalantioxidant activity, i.e. they block extra- and intracellularproduction of ROS (basically O2
● –) by inflammatory ratperitoneal macrophages, through inhibition of NAD(P)Hoxidase activity, and inhibit the production of NO● (at leastin part by inhibition of iNOS gene and protein expression).In adition, in in vitro studies, c-RESV and t-R E S Vscavenge the stable free radical DPPH and inhibit both lipidperoxidation (measured under different experimentalconditions) and citroneal thermo-oxidation.
These inhibitory effects of c-RESV and t-RESV arequalitatively similar. Therefore, the different spatialconformation of c-RESV [versus that of the trans isomer,(see Fig. 1 )] does not seem to markedly modify itsinteraction with the potential cellular targets.
c-RESV and t-RESV seem to have different hepaticmetabolisms (specifically, regio- and stereoselectiveglucuronidation, catalyzed by different UDP-glucuronosyltransferase isoforms) [90]; but notwithstandingthis, c-RESV (like t-RESV) has been reported to beeffectively absorbed after oral administration in rats, and toaccumulate in rat tissues such as the heart, liver and kidney[91,92].
In addition to free RESV isomers (c-RESV and t-RESV)present at variable concentrations in red wines, a number ofRESV derivatives (mainly the 3-O-β-D-glucosides) are alsopresent [see, e.g., refs. 1,4]. These may be absorbed directly,as reported for the rat small intestine [93,94], and/orhydrolysed before absorption by glucosidases present in thehuman intestinal tract, with subsequent release of free RESV[95,96; for reviews, see, e.g., refs. 4,97]. These RESVderivatives may contribute to the biologically availableRESV dose.
For these reasons, Bertelli et al. [98] have concluded thatan average drinker of wine can absorb a sufficient amount ofRESV, at least in the long term, to explain the beneficialeffects of red wine on health.
In addition, it is interesting to note that increasedmacrophage activation [e.g. as a result of overproduction ofROS and RNS, or abnormally high NAD(P)H oxidase oriNOS activity] and abnormally high oxidative stress (e.g.LDL oxidation) have been observed in different stages ofcoronary heart disease, and in a number of inflammatoryprocesses (including atherosclerosis) [see, e.g., ref. 99].Bearing in mind the above considerations and assuming thatthe RESV isomers show similar behaviour in humans and invivo, it can be concluded that:
1. The apparent beneficial (cardioprotective) effects ofmoderate red wine consumption may be due to thecombined effects of t-RESV and c-RESV;
2. The RESV isomers may be of value as structuraltemplates for the design and development of newdrugs useful for reducing pathogenesis ofcardiovascular diseases and other oxidative-stress-related pathologies.
However, further studies are required to confirm that theantioxidant effects of the RESV isomers observed in vitrolikewise occur in vivo (after long-term treatment), with the

Comparative Studies of the Antioxidant Effects of Cis- and Trans-Resveratrol Current Medicinal Chemistry, 2006, Vol. 13, No. 1 97
aim of confirming the possible therapeutic applicationsmentioned above. Until conclusive long-term clinical,epidemiological and toxicological studies with RESV havebeen yet carried out in healthy and unhealthy humanvolunteers, the RESV isomers should be considered asfashionable dietary components with interesting antioxidantand other biological properties (e.g. antineoplastic activity)which may be therapeutically beneficial.
7. ACKNOWLEDGMENTS
I apologize for failing to cite many relevant primarypapers because of space constraints. The work in mylaboratory was supported in part by grants from the SpanishMinisterio de Ciencia y Tecnología (SAF2002-0245),Almirall-Prodesfarma Laboratories (Pharmacology Award2003) and the Xunta de Galicia (PGIDIT02BTF20301PRand PGIDIT05BTF20302PR), Spain. I am especiallygrateful to Almirall-Prodesfarma Laboratories and theSpanish Pharmacological Society for granting me the 2003Pharmacology Award.
NON-STANDARD ABBREVIATIONS
AAPH = 2,2'-azobis(2-amidinopropane) dihy-drochloride
BSA = Bovine serum albumin
c-RESV = Resveratrol, cis isomer
DCF = Dichlorofluorescein
DPI = Diphenyleneiodonium chloride
DPPH = 1,1-diphenyl-2-picryl-hydrazyl
EQ = Efficient quantity
H2DCFDA = 2,7-dichlorodihydrofluorescein (H2DCF)diacetate
H2HFF = Dihydro-2’,4,5,6,7,7’-hexafluorofluor-escein
HX = Hypoxanthine
IFN-γ = Interferon gamma
LDL = Low-density lipoproteins
LPS = Lipopolysaccharide
NADH/NADPH = Nicotinamide adenine dinucleotide/nicotinamide adenine dinucleotide phosphate oxidase
NBT = Nitroblue tetrazolium
NF-κB = Nuclear factor-κB
NO• = Nitric oxide
iNOS = Inducible nitric oxide synthase
O2● – = Superoxide radical
ONOO– = Peroxynitrite
PKC = Protein kinase C
PMA = Phorbol 12-myristate 13-acetate
RESV = Resveratrol
RNS = Reactive nitrogen species
ROS = Reactive oxygen species
RT-PCR = Reverse transcription-polymerase chain reaction
SOD = Superoxide dismutase
TBHP = Tert-butylhydroperoxide
t-RESV = Resveratrol, trans isomer
XO = Xanthine oxidase
REFERENCES
[1] Frémont, L. Life. Sci., 2000, 66, 663.[2] Pervaiz, S. FASEB J., 2003, 17, 1975.[3] Aggarwal, B.B.; Bhardwaj, A.; Aggarwal, R.S.; Seeram, N.P.;
Shishodia, S.; Takada, Y. Anticancer Res., 2004, 24, 2783.[4] Soleas, G.J.; Diamandis, E.P.; Goldberg, D.M. Clin. Biochem.,
1997, 30, 91.[5] Soleas, G.J.; Diamandis, E.P.; Goldberg, D.M. Adv. Exp. Med.
Biol., 2001, 492, 159.[6] Alarcón de la Lastra, C.; Villegas, I. Mol. Nutr. Food Res., 2005,
49, 405.[7] Takaoka, M.J. J. Fac. Sci. Hokkaido Imp. Univ., 1940, 3, 1.[8] Nonomura, S.; Kanagawa, H.; Makimoto, A. Yakugaku Zasshi.,
1963, 83, 988.[9] Langcake, P.; Pryce, R. J. Physiol. Plant Pathol., 1976, 9, 77.[10] Siemann, E.H.; Creasy, L.L. Am. J. Enol. Vitic., 1992, 43, 49.[11] Renaud, S.; De Lorgeril, M. Lancet, 1992, 339, 1523.[12] Wu, J.M.; Wang, Z.R.; Hsieh, T.C.; Bruder, J.L.; Zou, J.G.;
Huang, Y.Z. Int. J. Mol. Med., 2001, 8, 3.[13] Granados-Soto, V. Drug News Perspect., 2003, 16, 299.[14] Ulrich, S.; Wolter, F.; Stein, J.M. Mol. Nutr. Food Res., 2005, 49,
452.[15] Hao, H.D.; He, L.R. J. Med. Food, 2004, 7, 290.[16] Bradamante, S.; Barenghi, L.; Villa, A. Cardiovasc. Drug. Rev.,
2004, 22, 169.[17] Delmas, D.; Jannin, B.; Latruffe, N. Mol. Nutr. Food Res., 2005,
49, 377.[18] Olas, B.; Wachowicz, B. Platelets, 2005, 16, 251.[19] Middleton, E.Jr.; Kandaswami, C.; Theoharides, T.C. Pharmacol.
Rev., 2000, 52, 673.[20] Palomino, O.; Gómez-Serranillos, M.P.; Slowing, K.; Carretero,
E.; Villar, A. J. Chromatogr. A, 2000, 870, 449.[21] Burns, J.; Yokota, T.; Ashihara, H.; Lean, M.E.; Crozier, A. J.
Agric. Food Chem., 2002, 50, 3337.[22] Wang, Y.; Catana, F.; Yang, Y.; Roderick, R.; van Breemen, R.B.
J. Agric. Food Chem., 2002, 50, 431.[23] Belguendouz, L.; Frémont, L.; Linard, A. Biochem. Pharmacol.,
1997, 53, 1347.[24] Stivala, L.A.; Savio, M.; Carafoli, F.; Perucca, P.; Bianchi, L.;
Maga, G.; Forti, L.; Pagnoni, U.M.; Albini, A.; Prosperi, E.;Vannini, V. J. Biol. Chem., 2001, 276, 22586.
[25] Leiro, J.; Álvarez, E.; Arranz, J.A.; Laguna, R.; Uriarte, E.;Orallo, F. J. Leukoc. Biol., 2004, 75, 1156.
[26] Orallo, F. In Resveratrol in Health and Disease, Aggarwal, B.B.;Shishodia, S., Eds.; CRC Press: Boca Raton, USA, 2005, pp. 577-600.
[27] Varache-Lembège, M.; Waffo-Teguo, P.; Richard, T.; Monti, J.P.;Deffieux, G.; Vercauteren, J.; Mérillon, J.M.; Nuhrich, A. Med.Chem. Res., 2000, 10, 253.
[28] Pettit, G.R.; Grealish, M.P.; Jung, M. K.; Hamel, E.; Pettit, R.K.;Chapuis, J.C.; Schmidt, J.M. J. Med. Chem., 2002, 45, 2534.
[29] Leiro, J.; Álvarez, E.; García, D.; Orallo, F. I n t .Immunopharmacol., 2002, 2, 767.
[30] Cai, H.; Griendling, K.K.; Harrison, D.G. Trends Pharmacol. Sci.,2003, 24, 471.
[31] Griendling, K.K. Heart, 2004, 90, 491.[32] El-Benna, J.; Dang, P.M.; Gougerot-Pocidalo, M.A.; Elbim, C.
Arch. Immunol. Ther. Exp. (Warsz), 2005, 53, 199.

98 Current Medicinal Chemistry, 2006, Vol. 13, No. 1 F. Orallo
[33] Lassègue, B.; Clempus, R.E. Am. J. Physiol. Regul. Integr. Comp.Physiol., 2003, 285, R277.
[34] Geiszt, M.; Leto, T.L. J. Biol. Chem., 2004, 279, 51715.[35] Brandes, R.P.; Kreuzer, J. Cardiovasc. Res., 2005, 65, 16.[36] Segal, A.W. Annu. Rev. Immunol., 2005, 23, 197.[37] Harrison, R. Free Radic. Biol. Med., 2002, 33, 774.[38] Berry, C.E.; Hare, J.M. J. Physiol., 2004, 555, 589.[39] Hille, R. Arch. Biochem. Biophys., 2005, 433, 107.[40] Martin, H.M.; Hancock, J.T.; Salisbury, V.; Harrison, R. Infect.
Immun., 2004, 72, 4933.[41] Cos, P.; Ying, L.; Calomme, M.; Hu, J.P.; Cimanga, K.; Van Poel,
B.; Pieters, L.; Vlietinck, A.J.; Berghe, D.V. J. Nat. Prod., 1998,61, 71.
[42] Robak, J.; Gryglewski, R.J. Biochem. Pharmacol., 1988, 37, 837.[43] Hung, L.M.; Su, M.J.; Chu, W.K.; Chiao, C.W.; Chan, W.F.; Chen,
J.K. Br. J. Pharmacol., 2002, 135, 1627.[44] Zhou, C.X.; Kong, L.D.; Ye, W.C.; Cheng, C.H.; Tan, R.X. Planta
Med., 2001, 67, 158.[45] Orallo, F.; Álvarez, E.; Camiña, M.; Leiro, J.M.; Gómez, E.;
Fernández, P. Mol. Pharmacol., 2002, 61, 294.[46] Rivadulla, E.; Álvarez, E.; Leiro, J.M.; Quezada, E.; Uriarte, E.;
Orallo, F. The Pharmacologist, 2002, 44, suppl. 2, A144, 92.5(abstract).
[47] Liu, J.C.; Chen, J.J.; Chan, P.; Cheng, C.F.; Cheng, T.H.Hypertension, 2003, 42, 1198.
[48] Poolman, T.M.; Ng, L.L.; Farmer, P.B.; Manson, M.M. FreeRadic. Biol. Med., 2005, 39, 118.
[49] Allen, L.A. Microbes Infect., 2003, 5, 1329.[50] Voyich, J.M.; Musser, J.M.; DeLeo, F.R. Microbes Infect., 2004,
6, 1117.[51] Ischiropoulos, H.; Gow, A.; Thom, S.R.; Kooy, N.W.; Royall, J.A.;
Crow, J.P. In Nitric Oxide. Part C. Biological and Antioxidantactivities. Methods in Enzymology, Packer, L., Ed.; AcademicPress: New York, 1999; Vol. 301, pp. 367-373.
[52] O'Malley, Y.Q.; Reszka, K.J.; Britigan, B.E. Free Radic. Biol.Med., 2004, 36, 90.
[53] Förstermann, U.; Gath, I.; Schwarz, P.; Closs, E.I.; Kleinert, H.Biochem. Pharmacol., 1995, 50, 1321.
[54] Marín, J.; Rodríguez-Martínez, M.A. Pharmacol. Ther., 1997, 75,111.
[55] Hobbs, A.J.; Higgs, A.; Moncada, S. Annu. Rev. Pharmacol.Toxicol., 1999, 39, 191.
[56] Domenico, R. Curr. Pharm. Des., 2004, 10, 1667.[57] Mariotto, S.; Menegazzi, M.; Suzuki, H. Curr. Pharm. Des., 2004,
10, 1627.[58] Ghafourifar, P.; Cadenas, E. Trends Pharmacol. Sci., 2005, 26,
190.[59] Korhonen, R.; Lahti, A.; Kankaanranta, H.; Moilanen, E. Curr.
Drug Targets Inflamm. Allergy, 2005, 4, 471.[60] Tiefenbacher, C.P.; Kreuzer, J. Curr. Vasc. Pharmacol., 2003, 1,
1231.[61] Lindgren, H.; Stenman, L.; Tarnvik, A.; Sjöstedt, A. Microbes
Infect., 2005, 7, 467.[62] Murphy, M.P.; Packer, M.A.; Scarlett, J.L.; Martin, S.W. Gen.
Pharmacol., 1998, 31, 179.[63] Dröge, W. Physiol. Rev., 2002, 82, 47.[64] Muscoli, C.; Cuzzocrea, S.; Riley, D.P.; Zweier, J.L.;
Thiemermann, C.; Wang, Z.Q.; Salvemini, D. Br. J. Pharmacol.,2003, 140, 445.
[65] Faraci, F.M.; Didion, S.P. Arterioscler. Thromb. Vasc. Biol., 2004,24, 1367.
[66] Petersen, S.V.; Enghild, J.J. Biomed. Pharmacother., 2005, 59,175.
[67] Hayden, M.S.; Ghosh, S. Genes Dev., 2004, 18, 2195.[68] Shishodia, S.; Aggarwal, B.B. Biochem. Pharmacol., 2004, 68,
1071.[69] Xiao, C.; Ghosh, S. Adv. Exp. Med. Biol., 2005, 560, 41.[70] Beutler, B. Nature, 2004, 430, 257.[71] Takeda, K.; Akira, S. Int. Immunol., 2005, 17, 1.[72] Theofilopoulos, A.N.; Baccala, R.; Beutler, B.; Kono, D.H. Annu.
Rev. Immunol., 2005, 23, 307.[73] Schroder, K.; Hertzog, P.J.; Ravasi, T.; Hume, D.A. J. Leukoc.
Biol., 2004, 75, 163.[74] Rosenzweig, S.D.; Holland, S.M. Immunol. Rev., 2005, 203, 38.[75] Blanchette, J.; Jaramillo, M.; Olivier, M. Immunology, 2003, 108,
513.[76] Viatour, P.; Merville, M.P.; Bours, V.; Chariot, A. Trends
Biochem. Sci., 2005, 30, 43.[77] Tsai, S.H.; Lin-Shiau, S.Y.; Lin, J.K. Br. J. Pharmacol., 1999, 126,
673.[78] Chan, M.M.; Mattiacci, J.A.; Hwang, H.S.; Shah, A.; Fong, D.
Biochem. Pharmacol., 2000, 60, 1539.[79] Murakami, A.; Matsumoto, K.; Koshimizu, K; Ohigashi, H.
Cancer Lett., 2003, 195, 17.[80] Leiro, J.; Arranz, J.A.; Fraiz, N.; Sanmartin, M.L.; Quezada, E.;
Orallo, F. Int. Immunopharmacol., 2005, 5, 393.[81] Matthäus, B. J. Agric. Food Chem., 2002, 50, 3444.[82] Aruoma, O.I. Mutat. Res., 2003, 523-524, 9.[83] Basly, J.P.; Marre-Fournier, F.; Le Bail J.C.; Habrioux, G.; Chulia,
A.J. Life Sci., 2000, 66, 769.[84] Fauconneau, B.; Waffo-Teguo, P.; Huguet, F.; Barrier, L.;
Decendit, A.; Mérillon, J.M. Life Sci., 1997, 61, 2103.[85] Waffo-Teguo, P.; Fauconneau, B.; Deffieux, G.; Huguet, F.;
Vercauteren, J.; Mérillon, J.M. J. Nat. Prod., 1998, 61, 655.[86] Halliwell, B.; Chirico, S. Am. J. Clin. Nutr., 1993, 57, 715S.[87] Rice-Evans, C.; Leake, D.; Bruckdorfer, K.R.; Diplock, A.T. Free
Radic. Res., 1996, 25, 285.[88] Yoshida, Y.; Itoh, N.; Saito, Y.; Hayakawa, M.; Niki, E. Free
Radic. Res., 2004, 38, 375.[89] Azorín, I.; Bella, M.C.; Iborra, F.J.; Fornas, E.; Renau-Piqueras, J.
Free Radic. Biol. Med., 1995, 19, 795.[90] Aumont, V.; Krisa, S.; Battaglia, E.; Netter, P.; Richard, T.;
Mérillon, J.M.; Magdalou, J.; Sabolovic, N. Arch. Biochem.Biophys., 2001, 393, 281.
[91] Bertelli, A.A.; Giovannini, L.; Stradi, R.; Bertelli, A.; Tillement,J.P. Int. J. Tissue React., 1996, 18, 67.
[92] Bertelli, A.A.; Giovannini, L.; Stradi, R.; Urien, S.; Tillement, J.P.;Bertelli, A. Int. J. Clin. Pharmacol. Res., 1996, 16, 77.
[93] Andlauer, W.; Kolb, J.; Siebert, K.; Furst, P. Drugs Exp. Clin. Res.,2000, 26, 47.
[94] Kuhnle, G.; Spencer, J.P.; Chowrimootoo, G.; Schroeter, H.;Debnam, E.S.; Srai, S.K.; Rice-Evans, C.; Hahn, U. Biochem.Biophys. Res. Commun., 2000, 272, 212.
[95] Goldberg, D.M.; Ng, E.; Karumanchiri, A.; Diamandis, E.P.;Soleas, G.J. Am. J. Enol. Vitic., 1996, 47, 415.
[96] Soleas, G.J.; Angelini, M.; Grass, L.; Diamandis, E.P.; Goldberg,D.M. Methods Enzymol., 2001, 335, 145.
[97] Wenzel, E.; Somoza, V. Mol. Nutr. Food Res., 2005, 49, 472.[98] Bertelli, A.; Bertelli, A.A.E.; Gozzini, A.; Giovannini, L. Drugs
Exp. Clin. Res., 1998, 24, 133.[99] Stocker, R.; Keaney, J.F.Jr. Physiol. Rev., 2004, 84, 1381.
Received: May 17, 2005 Revised: September 5, 2005 Accepted: September 10, 2005

Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.