Embed Size (px)
Citation preview

Eur. J. Biochem. 194, 9-18 (1990) 0 FEBS 1990
Cloning and nucleotide sequence of cDNA encoding human liver serine - pyruvate aminotransferase Kozo NISHIYAMA, Gabriel BERSTEIN, Toshiaki ODA and Arata ICHIYAMA
Department of Biochemistry, Hamamatsu University School of Medicine, Hamamatsu, Shizuoka, Japan
(Received April 2O/July 17, 1990) - EJB YO 0452
Cloned cDNAs for human liver serine - pyruvate aminotransferase (Ser - PyrAT) were obtained by screening of a human liver cDNA library with a fragment of cDNA for rat mitochondrial Ser-PyrAT as a probe. Two clones were isolated from 50000 transformants. Both clones contained approximately 1.5 kb cDNA inserts and were shown to almost completely overlap each other on restriction enzyme mapping and DNA sequencing. The nucleotide sequence of the mRNA coding for human liver Ser - PyrAT was determined from those of the cDNA clones. The mRNA comprises at least 1487 nucleotides, and encodes a polypeptide consisting of 392 amino acid residues with a molecular mass of 43 039 Da. The amino acid composition determined on acid hydrolysis of the purified enzyme showed good agreement with that deduced from the nucleotide sequence of the cDNA. In vitro translation of the mRNA derived from one of the isolated clones, pHsptl2, as well as that of mRNA extracted from human liver, yielded a product of 43 kDa which reacted with rabbit anti-(rat mitochondrial Ser - PyrAT) serum. Comparison of the deduced amino acid sequences of human Ser-PyrAT and the mature form of rat mitochondrial Ser - PyrAT revealed 79.3% identity. Although human Ser -PyrAT appears to be synthesized as the mature size, the 5’-noncoding region of human Ser-PyrAT mRNA contains a nucleotide sequence which would encode, if translated, an amino acid sequence similar to that of the N-terminal extension peptide of the precursor for rat mitochondrial Ser -PyrAT.
Serine -pyruvate aminotransferase (Ser - PyrAT) is an en- zyme whose organelle localization and hormone responsive- ness differ with the animal species. In human liver, this enzyme is entirely peroxisomal [l - 31, while in cat and dog liver, it is entirely mitochondrial [3]. In rat liver, there are two types of this enzyme in terms of organelle distribution and hormone responsiveness: one is a mitochondrial matrix enzyme which is markedly induced on the administration of glucagon [4, 51 or insulin [6], and the other a glucagon-insensitive peroxisomal enzyme [4, 5, 71.
It is well known that in eukaryotic cells each organelle participates in different cellular processes, and enzymes in- volved in these processes are synthesized being destined to be localized in a relevant subcellular compartment. Therefore, the localization of Ser - PyrAT in mitochondria and/or peroxisomes suggests that the enzyme in the different organelles participates in different metabolic processes.
Correspondence to K. Nishiyama, Department of Biochemistry, Hamamatsu University School of Medicine, 3600 Handa-cho, Hamamatsu, Shizuoka, Japan 431-31
Note. The novel amino acid sequence data published here has been deposited with the EMBL sequence data bank.
The novel nucleotide sequence data published here has been de- posited with the EMBL sequence data banks and is available under accession number X 56092.
Abbreviations. Ser - PyrAT, serine - pyruvate aminotransferase; pRspt, a plasmid containing the cDNA sequence for rat Ser- PyrAT; pHspt, a plasmid containing the cDNA sequence for human Ser- PyrAT; E64C, trans-epoxysuccinyl-~-leucylamido(3-methyl)butane.
Enzymes. Serine-pyruvate aminotransferase (EC 2.6.1.51); T7 RNA polymerase (EC 2.7.7.6).
With regard to species-specific subcellular distribution, some change(s) in the mechanism determining organelle local- ization may have occurred during the course of evolution according to metabolic needs. Studies from several laborato- ries have suggested a role for Ser - PyrAT in gluconeogenesis from serine [8 - lo]. On the other hand, this enzyme from all animal species so far examined utilizes glyoxylate as a good substrate [ l l - 161.
Ser - PyrAT from rodent liver has a broad substrate speci- ficity, catalyzing the transamination between various neutral L-a-amino acids and corresponding a-0x0 acids or glyoxylate, while the enzyme from human, cat and dog liver is relatively specific for alanine and serine, the corresponding a-0x0 acids, and glyoxylate [ l l , 13-16]. In fact, the substantial contri- bution of this enzyme to the metabolism of glyoxylate has recently been demonstrated [17, 181. Since glyoxylate is an immediate precursor of oxalate, an undesirable end product of metabolism [19], and a large portion of it is produced in peroxisomes via glycolate, the peroxisomal localization of Ser - PyrAT can be considered to be favorable for the elimin- ation of this precursor of oxalate.
Previously, we investigated the mechanism underlying the dual subcellular localization of rat Ser - PyrAT, and revealed that the mitochondrial and peroxisomal Ser - PyrATs are de- rived from respective mRNAs, which differ in the 5’-terminal sequence due to transcription from different initiation sites, about 70 nucleotides apart, in the first exon of the Ser -PyrAT gene [20]. The translation product of the longer mRNA is the precursor of mitochondrial Ser - PyrAT and contains an N- terminal extension peptide, which is essential for the specific translocation of the precursor into mitochondria. On the other

10
hand, most peroxisomal proteins are known to be synthesized as the mature size and then post-translationally transported into the organelle by means of a peroxisome-targeting sig- nal(s), probably located in the enzyme molecule [21, 221. The translation product encoded by the shorter mRNA for rat Ser - PyrAT had the same molecular mass as that of the ma- ture form of the enzyme determined on SDS/polyacrylamide gel electrophoresis. So far as rat Ser-PyrAT is concerned, it is thus indicated that the subcellular localization is directed by a mechanism which determines the transcription-initiation site. Therefore, the structures of Ser - PyrAT genes and the mechanism underlying the generation of Ser - PyrAT mRNAs in man (Ser-PyrAT is entirely peroxisomal), cat, dog and weasel (Ser - PyrAT is entirely mitochondrial) are of great interest in relation to the mechanism determining the subcellu- lar localization of this enzyme. In particular, in man, it was discovered that patients with primary hyperoxaluria type I are generally deficient in this enzyme [17, 181, and further, one type of this genetic disorder was recently shown to be caused, at least in part, by the abnormal localization of Ser-PyrAT in mitochondria [23]. We report in this paper the isolation of cDNA clones and the nucleotide sequence encompassing the whole coding region for human liver Ser - PyrAT.
MATERIALS AND METHODS Construction and screening of a human liver cDNA library
A piece of human liver was obtained at the time of surgical operation on a 62-year-old male who suffered from liver met- astasis of gall bladder carcinoma. RNA was extracted from macroscopically non-invasive parts of the sample by a modi- fied SDS/phenol method [24], followed by oligo(dT)-cellulose chromatography [25]. cDNA was synthesized from poly(A)- rich RNA using a cDNA synthesis kit, as described by the supplier (Pharmacia) [26, 271. Double-stranded cDNA was ligated to an EcoR1-digested, phosphatase-treated phagemid, pT7T318U (Pharmacia). Escherichia coli SCSl was trans- formed with the recombinant phagemid by the calcium chlor- ide procedure [28], followed by selection for ampicillin resist- ance. Since an Ah1 - AluI fragment (395 bp) of the cDNA for rat mitochondrial Ser - PyrAT (pRspt321) [29] was shown to hybridize with human Ser - PyrAT mRNA on preliminary RNA blot analysis, the human liver cDNA library (approxi- mately 50 000 transformants) was screened with the cDNA fragment of pRspt321 which had been 32P-labeled by the random-primer labeling method [30]. Prehybridization was carried out at 65 'C by incubating duplicate nitrocellulose filter lifts successively in 3 x NaCl/Cit (NaCl/Cit: 150 mM NaC1/15 mM sodium citrate) containing 0.1% SDS for 30 min, 10 x Denhardt's solution [31] containing 3 x NaCl/ Cit for 1 h, and 50 mM Tris/HCl (pH 7.8)/1 M NaCI/10 mM EDTAjlO x Denhardt's solution/O.l% SDS for 1 h. Hy- bridization was performed at 60°C for 14 h in 50 mM Tris/ HC1 (pH 7.8)/1 M NaCl/10 mM EDTA/10 x Denhardt's solution/O.l % SDS containing 100 pg/ml denatured salmon sperm DNA and the denatured cDNA probe (1 x lo9 cpm/ pg). The filters were washed three times, for 5 min each, with an excess of 2 x NaCl/Cit containing 0.1% SDS at room tem- perature, then twice, for 20 min each, with 0.1 x NaCl/Cit containing 0.1 YO SDS at 60 "C.
The resulting cDNA fragments were subcloned into the phagemid vector, pT7T318U. The resulting phagemids were amplified in E. coli NM522 and then recovered as single- stranded phage DNAs through the infection of helper phage M13K07 [32]. Sequence analysis was performed by the dideoxy-chain-termination method [33] using a Sequenase kit (United States Biochemical Corp.) or the Klenow fragment of E. coli DNA polymerase I.
RNA blot analysis
RNA blot analysis was performed according to Alwine et al. [34]. Poly(A)-rich RNA was prepared from human liver, rat liver and glucagon-treated rat liver after extraction by the SDS/phenol method [35], then denatured with 1 M glyoxal/ 50% dimethylsulfoxide. The denatured RNA (5 pg) was electrophoresed on a 1.2'?4 agarose gel, then transferred to a nylon membrane (Hybond N, Amersham). "P-labeled anti- sense transcript, synthesized using T7 RNA polymerase from the plasmid pAM19 containing the 177-bp Hinfl - Hinfl frag- ment of human Ser - PyrAT cDNA, was used as the hybrid- ization probe. Prehybridization and hybridization were performed at 55°C for 22 h in 5 x NaCl/P,/EDTA [NaCl/Pi/ EDTA: 0.18 M NaCl/10 mM sodium phosphate (pH 7.4)/ 1 mM EDTA] containing 5 x Denhardt's solution, 50% formamide and 100 pg/ml denatured salmon sperm DNA. The membrane was washed twice with 5 x NaCl/P,/EDTA, for 15 min each, at 55"C, then twice in 1 x NaCl/Pi/EDTA containing 0.1% SDS, for 10 min each, at 55"C, and finally twice in 0.1 x NaC1/Pi/EDTA containing 0.1 % SDS, for 15 min each, at room temperature.
In vitro translation
1 pg each of poly(A)-rich RNA obtained from human liver, control and glucagon-treated rat liver, and mRNA synthesized in vitro from an isolated clone (pHsptl2) using T7 RNA polymerase, were translated in a reticulocyte-lysate system in the presence of [35S]methionine as described pre- viously [36]. The translation products were immunoreacted with rabbit anti-(rat mitochondrial Ser - PyrAT) serum or rabbit non-immune serum then precipitated by adsorption to protein-A-bearing Staphylococcus aurrus cells or Staphylosorb (Sanraku Co., Japan). SDS/lOO/, polyacryl- amide gel electrophoresis of the precipitates and fluorography were carried out as previously described [36].
Enzyme purification and amino acid composition analysis
Male or female Wistar/ST rats of mass 150- 180 g were supplied by Japan SLC Inc. (Hamamatsu, Japan) and maintained on a standard laboratory chow until the exper- iments. Subcellular fractionation of rat liver into heavy and light mitochondrial fractions was carried out according to de Duve et al. [37], except that the homogenization medium contained 0.5 mM phenylmethylsulfonyl fluoride, 10 pg/ml of E64C [trans-epoxysuccinyl-~-leucylamido(3-methyl)butane, a thiol-protease inhibitor] and 0.1 YO ethanol. When peroxisomes were to be isolated, the homogenization was carried out in a loosely fitting Potter-Elvehjem homogenizer, with a single run upward against a teflon pestle rotating at 500 - 800 rpm. For isolation of the peroxisomal fraction, rats were maintained for 2-4 weeks on the laboratory chow
The isolated phagemid clones containing cDNA inserts containing 2% (by mass) di-(2-ethylhexyl)phthalate (a peroxisomal proliferator), and the light mitochondrial frac-
NucIeotide sequencing
were digested with appropriate pairs of restriction enzymes.

11
tion obtained was subjected to density gradient centrifugation (linear gradient between 36% and 57% sucrose, by mass) in a RPZ-35T zonal rotor (Hitachi, Japan) at 71 300 x g,, for 3 h at 4-12°C. The recoveries of catalase and glutamate dehydrogenase from the homogenate in the isolated peroxisomal fraction were 28.5% and 0.85%, respectively. Rat mitochondrial Ser - PyrAT was purified from the heavy mitochondrial fraction obtained from the livers of glucagon- treated rats, as described previously [5 , 291. Rat peroxisomal Ser - PyrAT and human Ser - PyrAT were purified from the peroxisomal fractions obtained from the livers of di-(2- ethylhexy1)phthalate-fed rats and the 70000 x g supernatant of the sonicated human liver homogenate, respectively, essen- tially by the same procedures [5, 291. The rat and human enzymes, thus purified, each gave a single band of protein on SDS/polyacrylamide gel electrophoresis performed according to Laemmli [38], with visualization by silver staining. The purity of the human Ser - PyrAT was also assessed by reverse- phase high-performance liquid chromatography, which was carried out on a pBondasphere 5 p c8 (3.9 mm x 150 mm) column, with linear gradient elution with between 20% and 60% (by vol.) acetonitrile in 0.01 M HC1.
The purified human enzyme was hydrolyzed in 6 M HCl at 110°C for 24 h. After evaporation, the hydrolysate was analyzed with a Hitachi 835 automatic amino acid analyzer.
Other methods RNA was determined spectrophotometrically with the as-
sumption that an absorbance reading of 1 .O at 260 nm corre- sponds to a concentration of40 pg/ml. Protein was determined by the method of Lowry et al. [39]. Ser - PyrAT was assayed as described previously [5].
Materials
E64C, a thiol-protease inhibitor, was a kind gift from Dr K. Hanada, Research Laboratories, Taisho Pharmaceutical Co., Ltd., Saitama, Japan. [u -~~PICTP (110 TBq/mmol) and [ u - ~ ~ P I ~ C T P (1 10 TBq/mmol) were obtained from the Hungarian Academy of Sciences (Hungary) ; the [ 14C]- methylated protein mixture (185 kBq/ml), ~-[~%]methionine (37 TBq/mmol), multiprime DNA-labeling system and plasmid pAM19 from Amersham (UK); the cDNA synthesis kit, oligo(dT)-cellulose (Type 7), phagemid vector pT7T31 8U, helper phage M13K07 and E. coli NM522 from Pharmacia (Sweden); the Sequenase kit from United States Biochemical Co. (USA), the silver stain kit from Bio-Rad Laboratories (USA); T7 RNA polymerase from New England Biolabs, Inc. (USA); E. coli SCSl from Stratagene (USA); Staphylosorb from Sanraku (Tokyo, Japan); EcoRI, PstI and other restric- tion enzymes from Toyobo Co., Ltd (Osaka, Japan); the PBondasphere 5 p c8 (3.9 mm x 150 mm) column from Waters (USA); the molecular mass protein markers from Sigma (USA) ; phenylmethylsulfonyl fluoride from Wako Pure Chemical Industries (Osaka, Japan) ; leupeptin and antipain from Peptide Institute, Inc. (Minoh-shi, Japan). Other re- agents were of analytical grade or the highest quality commer- cially available.
RESULTS Isolation and characterization of human Ser - PyrAT cDNA
Two clones were isolated from the human liver cDNA library using the 395-bp AluI - AluI fragment (positions 185 -
Base Pairs
1 500 1000 1500
pHsptl2
- pHsptl6
Fig. 1. Restriction sites of and sequencing strategy for the human Ser - PyrAT cDNA clones, pHsptl2 and pHsptl6. The restriction enzyme map only shows the relevant restriction endonuclease sites used for sequencing by the method of Sanger [33]: Hf, HinfI; Hp, HpaII; P, PstI; R, RsaI; D, DdeI. The open box corresponds to the coding region, and the thin lines represent the 5’- and 3‘-noncoding regions. The directions and extents of the sequences determined are shown by horizontal arrows under each clone
579; cf. [29]) of rat mitochondrial Ser-PyrAT cDNA as a probe. Both clones contained approximately 1.5-kb cDNA inserts and were shown to almost completely overlap each other on restriction enzyme mapping and DNA sequencing (Fig. 1). The insert of one clone, designated as pHsptl2, was 36 bp longer at the 5’ end and that of the other clone, desig- nated as pHsptl6, was 6 bp longer at the 3‘ end. The nucleotide sequence of human Ser -PyrAT cDNA was obtained by over- lapping the sequences of the two isolated clones, and is shown together with the deduced amino acid sequence in Fig. 2 . DNA sequencing revealed an open reading frame of 1200 nucleotides (positions 1 - 1200). The first ATG codon was found at position 22, the sequence around the initial ATG codon (positions 19 - 25) being in agreement with the consen- sus sequence (A/G N N A T G G) for the eukaryotic initiation site reported by Kozak [40]. The open reading frame termin- ates with a TGA stop codon (positions 1198 - 1200), followed by 287 bp of the 3’-untranslated end, which terminates at the EcoRI adaptor sequence. The resulting reading frame encodes a translation product of 392 amino acids with a calculated molecular mass of 43039 Da. A 3’-poly(A) tail was not ob- served, but there was a sequence corresponding to a canonical polyadenylation signal at a site 272 bp beyond the stop codon.
mRNA and in-vitro-translation product for human liver Ser - PyrAT
On RNA blot analysis of human liver poly(A)-rich RNA, a single band of 1.7-kb mRNA, hybridizing to an antisense RNA probe prepared from human Ser-PyrAT cDNA, was observed (Fig. 3, lanes 1 and 1’). The mobility of the human Ser - PyrAT mRNA was almost the same as that of the 1.7-kb mRNA from normal rat liver, which was electrophoresed on the same gel as a size marker (Fig. 3, lane 3). With poly(A)- rich RNA from the livers of glucagon-treated rats, a longer mRNA of approximately 1.9 kb was observed, as previously reported [6, 411 (Fig. 3, lanes 2 and 2’).
In vitro translation was carried out with a rabbit re- ticulocyte-lysate system and the products were analyzed by SDS/polyacrylamide gel electrophoresis after immuno-

12 1 2 1 * GCGGCAGGTTGGGTGCGGACC
9 3* ATG.GCC.TCT.CAC.AAG.CTG.CTG.GTG.ACC.CCC.CCC.AAG.GCC.CTG.CTC.AAG.CCC.CTC.TCC.ATC.CCC.AAC.CAG.CTC. Met-Ala-Ser-His-Lys-Leu-Leu-Val-Thr-Pro-Pro-Lys-Ala-Leu-Le~-Lys-Pro-Le~-Ser-Ile-Pro-Asn-Gln-Leu- 1
165* CTG.CTG.GGG.CCT.GGT.CCT.TCC.AAC.CTG.CCT.CCT.CGC.ATC.ATG.GCA.GCC.GGG.GGG.CTG.CAG.ATG.ATC.GGG.TCC. Leu-Leu-Gly-Pro-Gly-Pro-Ser-Asn-Leu-Pro-Pro-Arg-Ile-Met-Ala-Ala-Gly-Gly-Leu-Gln-l~et-Ile-Gly-Ser-
30 2 3 7 *
ATG.AGC.AAG.GAT.ATG.TAC.CAG.ATC.ATG.GAC.GAG.ATC.AAG.GAA.GGC.ATC.CAG.TAC.GTG.TTC.CAG.ACC.AGG.AAC. Met-Ser-Lys-Asp-Met-Tyr-Gln-Ile-Met-Asp-Glu-Ile-Lys-Glu-Gly-Ile-Gln-Tyr-Val-Phe-Gln-Thr-Arg-Asn-
60 3 0 9 *
CCA.CTC.ACA.CTG.GTC.ATC.TCT.GGC.TCG.GGA.CAC.TGT.GCC.CTG.GAG.GCC.GCC.CTG.GTC.AAT.GTG.CTG.GAG.CCT, Pro-Leu-Thr-Leu-Val-Ile-Ser-Gly-Ser-Gly-His-Cys-Ala-Leu-Glu-Ala-Ala-Leu-Val-Asn-Val-Leu-Glu-Pro-
90 3 8 1 *
GGG.GAC.TCC.TTC.CTG.GTT.GGG.PYCC.AAT.GGC.ATT.TGG.GGG.CAG.CGA.GCC.GTG.GAC.ATC.GGG.GAG.CGC.ATA.GGA. Gly-Asp-Ser-Phe-Leu-Val-Gly-Ala-Asn-Gly-Ile-Trp-Gly-Gln-Arg-Ala-Val-Asp-Ile-Gly-Gl~-Arg-Ile-Gly-
1 2 0 4 5 3 *
GCC.CGA.GTG.CAC.CCG.ATG.ACC.AAG.GAC.CCT.GGA.GGC.CAC.TAC.ACA.CTG.CAG.GAG.GTG.GAG.GAG.GGC.CTG.GCC. Ala-Arg-Val-His-Pro-Met-Thr-Lys-Asp-Pro-Gly-Gly-His-Tyr-Thr-Leu-Gln-Glu-Val-Glu-Glu-Gly-Leu-Ala-
5 2 5 * CAG.CAC.AAG.CCA.GTG.CTG.CTG.TTC.TTA.ACC.CAC.GGG.GAG.TCG.TCC.ACC.GGC.GTG.CTG.CAG.CCC.CTT.GAT.GGC. Gln-His-Lys-Pro-Val-Leu-Leu-Phe-Leu-Thr-His-Gly-Glu-Ser-Ser-Thr-Gly-Val-Leu-Gln-Pro-Leu-Asp-Gly-
150 5 9 7 *
TTC.GGG.GAA.CTC.TGC.CAC.AGG.TAC.AAG.TGC.CTG.CTC.CTG.GTG.GAT.TCG.GTG.GCA.TCC.CTG.GGC.GGG.ACC.CCC. Phe-Gly-Glu-Leu-Cys-His-Arg-Tyr-Lys-Cys-Leu-Leu-Leu-Val-Asp-Ser-Val-Ala-Ser-Leu-Gly-Gly-Thr-Pro-
180 6 6 9 *
CTT.TAC.ATG.GAC.CGG.CAA.GGC.ATC.GAC.ATC.CTG.TAC.TCG.GGC.TCC.CAG.AAG.GCC.CTG.AAC.GCC.CCT.CCA.GGG. Leu-Tyr-Met-Asp-Arg-Gln-Gly-Ile-Asp-Ile-Leu-Tyr-Ser-Gly-Ser-Gln-Lys-Ala-Leu-Asn-Ala-Pro-Pro-Gly-
2 1 0 7 4 1 *
ACC.TCG.CTC.ATC.TCC.TTC.AGT.GAC.AAG.GCC.AAA.AAG.AAG.ATG.TAC.TCC.CGC.AAG.ACG.AAG.CC~.TTC.TCC.TTC. Thr-Ser-Leu-Ile-Ser-Phe-Ser-Asp-Lys-Ala-Lys-Lys-Lys-Met-Tyr-Ser-Arg-Lys-Thr-Lys-Pro-Phe-Ser-Phe-
2 4 0 8 1 3 *
TAC.CTG.GAC.ATC.AAG.TGG.CTG.GCC.AAC.TTC.TGG.GGC.TGT.GAC.GAC.CAG.CCC.AGG.ATG.TAC.CAT.CAC.ACA.ATC. Tyr-Leu-Asp-Ile-Lys-Trp-Leu-Ala-Asn-Phe-Trp-Gly-Cys-Asp-Asp-Gln-Pro-Arg-Met-Tyr-Hls-His-Thr-Ile-
8 8 5 * CCC.GTC.ATC.AGC.CTG.TAC.AGC.CTG.AGA.GAG.AGC.CTG.GCC.CTC.ATT.GCG.GAA.CAG.GGC.CTG.GAG.AAC.AGC.TGG. Pro-Val-Ile-Ser-Leu-Tyr-Ser-Leu-Arg-Glu-Ser-Leu-Ala-Leu-Ile-Ala-Glu-Gln-Gly-Leu-Glu-Asn-Ser-Trp-
270 9 5 7 *
CGC.CAG.CAC.CGC.GAG.GCC.GCG.GCG.TAT.CTG.CAT.GGG.CGC.CTG.CAG.GCA.CTG.GGG.CTG.CAG.CTC.TTC.GTG.AAG. Arg-Gln-His-Arg-Glu-Ala-Ala-Ala-Tyr-Leu-His-Gly-Arg-Leu-Gln-Ala-Leu-Gly-Leu-Gln-Leu-Phe-Va~-Lys-
300 1 0 2 9 *
GAC.CCG.GCG.CTC.CGG.CTT.CCC.ACA.GTC.ACC.ACT.GTG.GCT.GTA.CCC.GCT.GGC.TAT.GAC.TGG.AGA.GAC.ATC.GTC. Asp-Pro-Ala-Leu-Arg-Leu-Pro-Thr-Val-Thr-Thr-Val-Ala-Val-Pro-Ala-Gly-Tyr-Asp-Trp-Arg-Asp-I~e-Va~-
330 1101*
AGC.TAC.GTC.ATA.GAC.CAC.TTC.GAC.ATT.GAG.ATC.ATG.GGT.GGC.CTT.GGG.CCC.TCC.ACG.GGG.AAG.GTG.CTG.CGG. Ser-Tyr-Val-Ile-Asp-His-Phe-Asp-Ile-Glu-Ile-Met-Gly-Gly-Leu-Gly-Pro-Ser-Thr-Gly-Lys-Val-Leu-Arg-
3 6 0 1 1 7 3 *
ATC.GGC.CTG.CTG.GGC.TGC.AAT.GCC.ACC.CGC.GAG.AAT.GTG.GAC.CGC.GTG.ACG.GAG.GCC.CTG.AGG.GCG.GCC.CTG. Ile-Gly-Leu-Leu-Gly-C~s-Asn-Ala-Thr-Arg-Glu-Asn-Val-Asp-Arg-Val-Thr-Glu-Ala-Leu-Arg-Ala-Ala-Leu-
1200* 1 2 5 9 * CAG.CAC.TGC.CCC.AAG.AAG.AAG.CTG.TGA CCTGCCCACTGGCACACAGCTGGCACTGGCACACACCTGTCCCATGCCCACCCTGAGGG Gln-His-Cys-Pro-Lys-Lys-Lys-Leu
ATCAGGAGCAAACAGACCCTGCAAGGTCCTCCAGGCCTGGGGACAGGAAAGCCACTGACCCAGCCCGGGAGGCAGAACCAGGCAGCCTCCCTGGC
CCCAGGCAGCCCTTTTCCCTCCAGTGGCACCTCCTGGAAACAGTCCACTTGGGCGCAAARCCCAGTGCCTTCCAAATGAGCTGCAGTCCCCAGGC
1 3 5 4 *
1 4 4 9 *
1487* CATGAGCCTCCCGGGAATGTTTAATAAAAGGGCCTGGCC
Fig. 2. Nucleotide .~equence of’thc. cDNAfor human Ser - PyrAT and the deduced amino acid sequence. The nucleotide sequence comprises those of the two human Ser-PyrAT cDNA clones, pHsptl2 and pHsptl6. Nucleotides are numbered in the 5’-3’ direction, beginning with the first nucleotide after the EcoRI adaptor. The deduced amino acid residues are numbered from the methionine corresponding to the first appearing ATG codon. The putative polyadenylation signal in the 3’-noncoding region is underlined
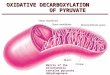
precipitation with anti-(rat mitochondrial Ser -PyrAT) serum or non-immune serum. As shown in Fig. 4A, an immuno- precipitated band of approximately 43 kDa was observed as the translation product of human liver poly(A)-rich RNA (Fig. 4A, lane 2). When the translation products were incu- bated with non-immune serum, this band was not detectable
(Fig. 4A, lane 1). The immunological cross reactivity of the translation product for human Ser-PyrAT with anti-(rat mitochondrial Ser-PyrAT) serum was also supported by the fact that an excess of purified rat mitochondrial Ser - PyrAT competed efficiently with the in-vim-translation product with regard to interaction with the antibody (Fig. 4A, lanes 3 and

13
Origin- 1 2 3 1’2’
28S-
18S-
/1.9 kb
1.7 kb 1
Fig. 3. RNA blot analysis using an antisense RNA probe prepared from human Ser - PyrAT cDNA. Poly(A)-rich RNA was prepared from livers of man (lane I), glucagon-treated rats (glucagon was injected intraperitoneally at the dose of 300 pg/lOO g body mass 3.5 h before sacrifice; lane 2) or untreated rats (lane 3). 5 pg of each poly(A)-rich RNA was loaded on a 1.2% agarose gel. Hybridization was carried out using a 32P-labeled antisense transcript derived from the 177-bp HinfI - Hinff fragment o f human Ser - PyrAT cDNA as a probe. The details of the procedures are given under Materials and Methods. Ribosomal RNAs were used as size markers. Lanes 1’ and 2‘, shorter exposure (approximately 25%) than that for lanes 1 and 2, respectively
4). In the case of translation from rat liver poly(A)-rich RNA, on the other hand, two immunoprecipitable products of differ- ent sizes (45 kDa and 43 kDa) were formed, in accord with the dual distribution of Ser-PyrAT in mitochondria and peroxisomes. The larger product (45 kDa) was predominant in the case of poly(A)-rich RNA from the livers of glucagon- treated rats (Fig. 4A, lane 5) , while the majority of the product migrated as a 43-kDa band when the translation was carried out with RNA from the livers of untreated rats (Fig. 4A, lane 6). Our previous data demonstrated that the larger product in glucagon-treated rats was the precursor for rat mitochondrial Ser - PyrAT [42]. In a separate experiment, cell-free trans- lation was carried out with mRNA synthesized in vitro from the T7 RNA polymerase promoter in an isolated clone, pHsptl2. The translation product of the synthesized mRNA reacted with anti-(rat mitochondrial Ser - PyrAT) serum (Fig. 4B, lane 5 ) and had the same molecular mass as the in- vitro-translation product of human liver poly(A)-rich RNA (Fig. 4B, lane 3). These findings indicate that the isolated clone, pHsptl2, has a cDNA insert which encompasses the whole coding region of human Ser - PyrAT mRNA. In lanes 3 and 5 in Fig. 4B, a minor band with a smaller molecular mass (39 kDa) was observed, in addition to the major band. The minor band was also seen when the in vitro translation was carried out in the presence of several protease inhibitors (phenylmethylsulfonyl fluoride, leupeptin, antipain and E64C; cf. Fig. 5, lane 4). Thus the smaller band may have been derived through translation from a downstream AUG codon (second appearing methionine) rather than degradation of the synthesized protein.
Molecular size, amino acid composition and deduced amino acid sequence of human liver Ser - PyrAT
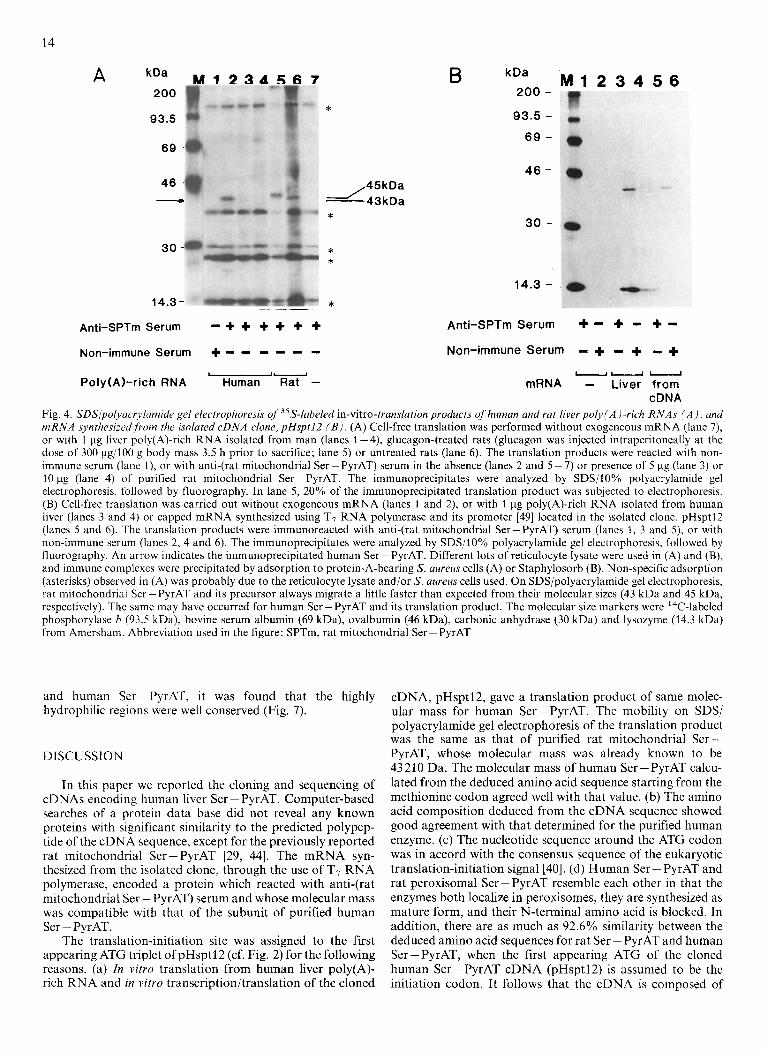
The purified human Ser - PyrAT and rat Ser - PyrAT were electrophoresed on an SDS/lO% polyacrylamide gel then vis- ualized by silver staining. Rat mitochondrial (Fig. 5, lane l), rat peroxisomal (Fig. 5, lane 2) and human Ser-PyrAT (Fig. 5, lane 3) each gave a single band at the same position, whose molecular mass corresponded to 43 kDa. In this exper- iment, the 35S-labeled in-vitro-transcription/translation prod- uct derived from pHsptl2 was electrophoresed in adjacent lanes and found to migrate to the same position as the purified enzymes (Fig. 5, lanes 4 and 5) , indicating that human Ser- PyrAT is synthesized as the mature size, as in the cases of rat peroxisomal Ser - PyrAT [20] and many other peroxisomal proteins [21, 221.
The amino acid composition deduced from the cDNA sequence showed good agreement with that determined on acid hydrolysis of the purified protein, when the first appear- ing ATG codon (nucleotide positions 22 - 24) of the isolated cDNA was assumed to be the initiation methionine codon (Table 1). On amino acid sequencing from the N-terminus of the purified human liver Ser - PyrAT by means of automated Edman degradation, no amino acid was liberated, suggesting that the N-terminus of the enzyme was blocked, as in the case of rat peroxisomal Ser - PyrAT [20].
Comparison of the deduced amino acid sequences for human Ser - PyrAT and rat Ser - PyrAT
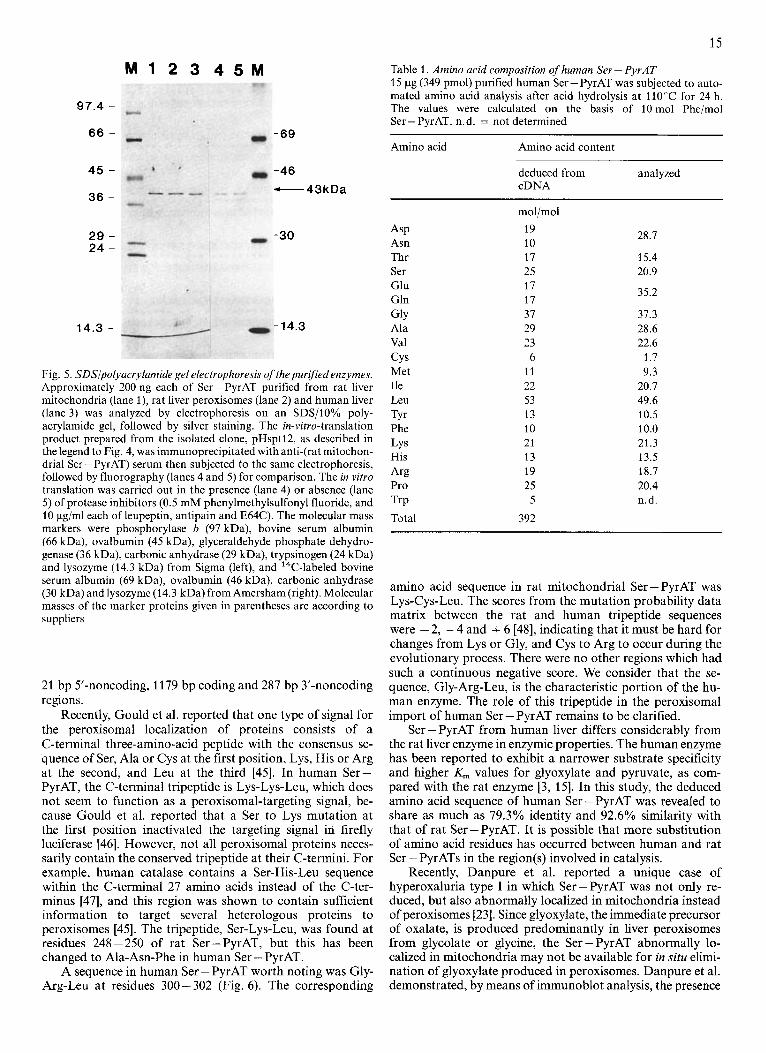
The amino acid sequences deduced from cDNAs for hu- man Ser-PyrAT (pHsptl2) and the precursor of rat mito- chondrial Ser - PyrAT (pRspt321) were compared (Fig. 6). In rat liver, in which Ser-PyrAT is localized in both mitochon- dria and peroxisomes [4, 5, 71, mitochondrial Ser-PyrAT is synthesized from a longer mRNA (1.9 kb mRNA) as a precur- sor which contains an N-terminal extension peptide. The mito- chondrial-targeting peptide is cleaved off when the precursor is transported into mitochondria, resulting in the formation of the mature form of mitochondrial Ser - PyrAT [42], whose N-terminal amino acid, methionine, is numbered 1 in Fig. 6. On the other hand, peroxisomal Ser - PyrAT is believed to be translated from a shorter mRNA (1.7 kb mRNA), the first appearing AUG codon being that corresponding to the N-terminal methionine of the mature mitochondrial Ser - PyrAT, yielding a product whose primary structure is the same as that of the precursor for mitochondrial Ser - PyrAT, except that the N-terminal extension peptide is lacking. This trans- lation product has been assumed to be transported into peroxisomes without being processed. Thus the structures of rat mitochondrial and peroxisomal Ser - PyrATs have been believed to be the same, except that the N-terminal amino acid of the peroxisomal Ser - PyrAT is blocked.
In Fig. 6, the first 22 amino acid residues underlined of the precursor of rat mitochondrial Ser -PyrAT represent the N-terminal extension peptide. The predicted amino acid sequence of human Ser - PyrAT shares 79.3% identity with that determined for the mature form of rat mitochondrial or peroxisomal Ser - PyrAT. When conservative amino acid substitutions are included, the similarity is as much as
Hopp and Woods [43] reported that the region of highest local average hydrophilicity in proteins is located in, or immediately adjacent to, an antigenic determinant. On hydrophilicity analysis of the rat mitochondrial Ser - PyrAT
92.6%.
14
M 1 2 3 4 5 6 7 A kDa
200 -
93.5 -
69 -
4 6 - - 30 -
*
* *
M 1 2 3 4 5 6 kDa 200 -
B
93.5 -
6 9 -
46 -
30 -
14.3 - *
~ -_ 14.3 -
Anti-SPTm Serum - + + + + + + Anti-SPTm Serum + - + - + - Non-immune Serum + - - - - - - Non-immune Serum - + - + I + -- Poly(A)-rich RNA Human Rat - ---
mRNA - Liver from cDNA
Fig. 4. SDSlpolyacrylarnide gel electrophoresis of 35S-labeled in-vitro-translation products of human and rat liver poly(A) -rich RNAs ( A ) , und rn R N A synthesized,frorn the isoluted cDNA clone, pHsptl2 ( B ) . (A) Cell-free translation was performed without exogeneous mRNA (lane 7), or with 1 pg liver poly(A)-rich RNA isolated from man (lanes 1 -4), glucagon-treated rats (glucagon was injected intraperitoneally at the dose of 300 pg/IOO g body mass 3.5 h prior to sacrifice; lane 5 ) or untreated rats (lane 6) . The translation products were reacted with non- immune serum (lane I), or with anti-(rat mitochondrial Ser-PyrAT) serum in the absence (lanes 2 and 5-7) or presence of 5 pg (lane 3) or 10 pg (lane 4) of purified rat mitochondrial Ser-PyrAT. The immunoprecipitates were analyzed by SDS/lO% polyacrylamide gel electrophoresis, followed by fluorography. In lane 5 , 20% of the immunoprecipitated translation product was subjected to electrophoresis. (B) Cell-free translation was carried out without exogeneous mRNA (lanes 1 and 2), or with 1 pg poly(A)-rich RNA isolated from human liver (lanes 3 and 4) or capped mRNA synthesized using T7 RNA polymerase and its promoter [49] located in the isolated clone, pHsptl2 (lanes 5 and 6). Thc translation products were immunoreacted with anti-(rat mitochondrial Ser-PyrAT) serum (lanes 1, 3 and 5 ) , or with non-immune serum (lanes 2, 4 and 6). The immunoprecipitates were analyzed by SDS/10% polyacrylamide gel electrophoresis, followed by fluorography. An arrow indicates the immunoprecipitated human Ser - PyrAT. Different lots of reticulocyte lysate were used in (A) and (B), and immune complexes were precipitated by adsorption to protein-A-bearing S. aureus cells (A) or Staphylosorb (B). Non-specific adsorption (asterisks) observed in (A) was probably due to the reticulocyte lysate and/or S. aweus cells used. On SDS/polyacrylamide gel electrophoresis, rat mitochondrial Ser - PyrAT and its precursor always migrate a little faster than expected from their molecular sizes (43 kDa and 45 kDa, respectively). The same may have occurred for human Ser-PyrAT and its translation product. The molecular size markers were 14C-labeled phosphorylase h (93.5 kDa), bovine serum albumin (69 kDa), ovalbumin (46 kDa), carbonic anhydrase (30 kDa) and lysozyme (14.3 kDa) from Amersham. Abbreviation used in the figure: SPTm, rat mitochondrial Ser-PyrAT
and human Ser-PyrAT, it was found that the highly hydrophilic regions were well conserved (Fig. 7).
DISCUSSION
In this paper we reported the cloning and sequencing of cDNAs encoding human liver Ser - PyrAT. Computer-based searches of a protein data base did not reveal any known proteins with significant similarity to the predicted polypep- tide of the cDNA sequence, except for the previously reported rat mitochondrial Ser-PyrAT [29, 441. The mRNA syn- thesized from the isolated clone, through the use of T7 RNA polymerase, encoded a protein which reacted with anti-(rat mitochondria1 Ser - PyrAT) serum and whose molecular mass was compatible with that of the subunit of purified human Ser - PyrAT.
The translation-initiation site was assigned to the first appearing ATG triplet of pHsptl2 (cf. Fig. 2) for the following reasons. (a) In v i m translation from human liver poly(A)- rich RNA and in virro transcription/translation of the cloned
cDNA, pHsptl2, gave a translation product of same molec- ular mass for human Ser-PyrAT. The mobility on SDS/ polyacrylamide gel electrophoresis of the translation product was the same as that of purified rat mitochondrial Ser - PyrAT, whose molecular mass was already known to be 43210 Da. The molecular mass of human Ser-PyrAT calcu- lated from the deduced amino acid sequence starting from the methionine codon agreed well with that value. (b) The amino acid composition deduced from the cDNA sequence showed good agreement with that determined for the purified human enzyme. (c) The nucleotide sequence around the ATG codon was in accord with the consensus sequence of the eukaryotic translation-initiation signal [40]. (d) Human Ser - PyrAT and rat peroxisomal Ser - PyrAT resemble each other in that the enzymes both localize in peroxisomes, they are synthesized as mature form, and their N-terminal amino acid is blocked. In addition, there are as much as 92.6% similarity between the deduced amino acid sequences for rat Ser - PyrAT and human Ser-PyrAT, when the first appearing ATG of the cloned human Ser-PyrAT cDNA (pHspt12) is assumed to be the initiation codon. It follows that the cDNA is composed of
15
M 1 2 3 4 5 M
97.4 -
66 - - 69
45 -
36 -
29 - 24 -
14.3 -
-46 -43kDa
-30
- 14.3
Fig. 5. SDSlpolyacrylamide gel electrophoresis of the purified enzymes. Approximately 200 ng each of Ser-PyrAT purified from rat liver mitochondria (lane 1), rat liver peroxisomes (lane 2) and human liver (lane 3) was analyzed by electrophoresis on an SDS/10% poly- acrylamide gel, followed by silver staining. The in-vitro-translation product prepared from the isolated clone, pHsptl2, as described in the legend to Fig. 4, was immunoprecipitated with anti-(rat mitochon- drial Ser - PyrAT) serum then subjected to the same electrophoresis, followed by fluorography (lanes 4 and 5) for comparison. The in vitro translation was carried out in the presence (lane 4) or absence (lane 5) of protease inhibitors (0.5 mM phenylmethylsulfonyl fluoride, and 10 pg/ml each of leupeptin, antipain and E64C). The molecular mass markers were phosphorylase b (97 kDa), bovine serum albumin (66 kDa), ovalbumin (45 kDa), glyceraldehyde phosphate dehydro- genase (36 kDa), carbonic anhydrase (29 kDa), trypsinogen (24 kDa) and lysozyme (14.3 kDa) from Sigma (left), and I4C-labeled bovine serum albumin (69 kDa), ovalbumin (46 kDa), carbonic anhydrase (30 kDa) and lysozyme (14.3 kDa) from Amersham (right). Molecular masses of the marker proteins given in parentheses are according to suppliers
21 bp 5’-noncoding, 1179 bp coding and 287 bp 3’-noncoding regions.
Recently, Gould et al. reported that one type of signal for the peroxisomal localization of proteins consists of a C-terminal three-amino-acid peptide with the consensus se- quence of Ser, Ala or Cys at the first position, Lys, His or Arg at the second, and Leu at the third [45]. In human Ser- PyrAT, the C-terminal tripeptide is Lys-Lys-Leu, which does not seem to function as a peroxisomal-targeting signal, be- cause Gould et al. reported that a Ser to Lys mutation at the first position inactivated the targeting signal in firefly luciferase [46]. However, not all peroxisomal proteins neces- sarily contain the conserved tripeptide at their C-termini. For example, human catalase contains a Ser-His-Leu sequence within the C-terminal 27 amino acids instead of the C-ter- minus [47], and this region was shown to contain sufficient information to target several heterologous proteins to peroxisomes [45]. The tripeptide, Ser-Lys-Leu, was found at residues 248-250 of rat Ser-PyrAT, but this has been changed to Ala-Asn-Phe in human Ser - PyrAT.
A sequence in human Ser - PyrAT worth noting was Gly- Arg-Leu at residues 300- 302 (Fig. 6). The corresponding
Table 1. Amino acid composition of human Ser - PyrAT 15 pg (349 pmol) purified human Ser - PyrAT was subjected to auto- mated amino acid analysis after acid hydrolysis at 110°C for 24 h. The values were calculated on the basis of 10 mol Phelmol Ser - PyrAT. n. d. = not determined
Amino acid Amino acid content
deduced from analyzed cDNA
ASP Asn Thr Ser Glu Gln GlY Ala Val CYS Met Ile Leu TYr Phe LYS His Arg Pro TrP Total
mol/mol 19 10 17 25 17 17 37 29 23 6
11 22 53 13 10 21 13 19 25
5
392
28.7
15.4 20.9
35.2
37.3 28.6 22.6
1.7 9.3
20.7 49.6 10.5 10.0 21.3 13.5 18.7 20.4 n.d.
amino acid sequence in rat mitochondria1 Ser-PyrAT was Lys-Cys-Leu. The scores from the mutation probability data matrix between the rat and human tripeptide sequences were - 2, - 4 and + 6 [48], indicating that it must be hard for changes from Lys or Gly, and Cys to Arg to occur during the evolutionary process. There were no other regions which had such a continuous negative score. We consider that the se- quence, Gly-Arg-Leu, is the characteristic portion of the hu- man enzyme. The role of this tripeptide in the peroxisomal import of human Ser - PyrAT remains to be clarified.
Ser - PyrAT from human liver differs considerably from the rat liver enzyme in enzymic properties. The human enzyme has been reported to exhibit a narrower substrate specificity and higher K, values for glyoxylate and pyruvate, as com- pared with the rat enzyme [3, 151. In this study, the deduced amino acid sequence of human Ser-PyrAT was revealed to share as much as 79.3% identity and 92.6% similarity with that of rat Ser-PyrAT. It is possible that more substitution of amino acid residues has occurred between human and rat Ser - PyrATs in the region(s) involved in catalysis.
Recently, Danpure et al. reported a unique case of hyperoxaluria type I in which Ser-PyrAT was not only re- duced, but also abnormally localized in mitochondria instead of peroxisomes [23]. Since glyoxylate, the immediate precursor of oxalate, is produced predominantly in liver peroxisomes from glycolate or glycine, the Ser - PyrAT abnormally lo- calized in mitochondria may not be available for in situ elimi- nation of glyoxylate produced in peroxisomes. Danpure et al. demonstrated, by means of immunoblot analysis, the presence
16
Human
Rat
Human
Rat
Human
Rat
Human
Rat
Human
Rat
Human
Rat
Human
Rat
1 27 MASHKLLVTPPKALLKPLSIPNQLLLG *+**+*** ** ** ******++****
MFRNIAKASVTLGSRAASWVRNMGSHQLLVPPPEALSKPLSIPKRLLLG
98 PGPSNLPPRIMAAGGLQMIGSMSKDMYQIMDEIKEGIQWFQTRNPLTLVISGSGHCALEAALVNVLEPGD ******+**++***+*+*** * *+*+*******+***************+*******+*+** *+***** PGPSNLAPRVLAAGSLRMIGHMQKEMFQIMDEIKQGIQWFQTRNPLTLWSGSGHCAMETALFNLLEPGD
169 SFLVGANGIWGQRAVDIGERIGARVHPMTKDPGGHYTLQEVEEGLAQHKPVLLFLTHGESSTGVLQPLDGF *****+***** ** +*+******** * * ** . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . SFLVGTNGIWGIRAAEIAERIGARVHQMIKKPGEHYTLQEVEEGLAQHKPVLLFLTHGESSTGVLQPLDGF
240 GELCHRYKCLLLVDSVASLGGTPLYMDRQGIDILYSGSQ~N~PGTSLISFSDKAKKKMYSRKTKPFSF . . . . . . . . . . . . . . . . . . . . . *+***+************ ****** *****+**** *+******* ** GELCHRYQCLLLVDSVASLGGVPIYMDQQGIDILYSGSQKVLNAPPGISLISFNDKAXSKVYSRKTKPVSF
.em 311 YLDIKWLANFWGCDDQPRMYHHTIPVISLYSLRESLALIAEQGLENSWRQHREAAAYLHGRLQALGLQL~ * ** *+++***+++ *+ ***+****** ********+*********+****+* ** *+ ***++** YTDITYLSKLWGCEGKTRVIHHTLPVISLYCLRESLALISEQGLENSWRRHREATAHLHKCLRELGLKFFV
382 K D P A L R L P ~ T ~ A V P A G Y D ~ D I V S ~ I D H F D I E I M ~ L G P S T G K V L R I G L L G C N A T R E ~ D R V T E ~ ~ * * * +****+***+*****+********+***+*** ****** +********* *** ** ***+**** KDPEIRLPTITTVTVPAGYNWRDIVSYVLDHFNIEISGGLGPSEDKVLRIGLLGYNATTENADRVAEALRE
... 392
ALQHCPKKKL *******+** ALQHCPKNKL
Fig. 6 . Comparison ofthe amino acid sequences of human Ser - PyrATand the rat mitochondrial Ser - PyrATprecursor. Human and rat represent the sequences deduced from the cDNAs for human Ser - PyrAT (pHsptl2, Fig. 2) and the precursor of rat mitochondrial Ser - PyrAT (pRspt321) [29], respectively. Amino acid residues are aligned using the one-letter amino acid notation, and numbered from the expected initiation methioninc in the case of the human enzyme, and from the N-terminal methionine residue of the mature form in the case of the rat mitochondrial enzyme. The N-terminal amino acid of human Ser -PyrAT was blocked, as described in the text, and has not yet been identified. The N-terminal extension peptide of the precursor for rat mitochondrial Ser - PyrAT is underlined. Dotted arc internal tripeptides, Ser-Lys- Leu in rat Ser - PyrAT and Gly-Arg-Leu in human Ser - PyrAT, discussed in the text in relation to peroxisomal targeting. Asterisks indicate identical amino acid residues in the human Ser- PyrAT and rat mitochondrial Ser- PyrAT. Crosses indicate conservative amino acid changes. judged on the basis of a score of greater than zero for the mutation data matrix [48]
in the cytoplasmic fraction of an immunoreactive protein with a molecular mass 2-3 kDa larger than that of the normal human liver Ser - PyrAT [23]. This larger protein may corre- spond to the precursor of rat mitochondrial Ser -PyrAT judg- ing from the molecular mass. In this respect, it is especially noteworthy that the 5'-noncoding region of human Ser- PyrAT cDNA contains a nucleotide sequence corresponding to the C-terminal seven amino acids of the mitochondrial- targeting N-terminal extension peptide of the precursor of rat mitochondrial Ser - PyrAT. Among the seven amino acids tentatively deduced from the cDNA sequence, five were ident- ical with those in the rat mitochondria]-targeting signal and one was conservative (Fig. 8). One possibility is that the human Ser-PyrAT gene also contains, upstream of the coding region, a nucleotide sequence for targeting to mito- chondria. If the nucleotide sequence of the 5'-flanking or 5'- noncoding region of the human Ser - PyrAT gene shares high similarity with that of the entire N-terminal extension peptide of rat mitochondrial Ser-PyrAT, it may well be that the human Ser - PyrAT gene is normally transcribed from an initiation site corresponding to that for the shorter rat Ser - PyrAT mRNA. It follows that, in man, transcription from the initiation site corresponding to that for the longer rat mRNA (mRNA for the precursor of mitochondrial Ser - PyrAT) does not occur due to some mechanism. A disorder of the mechan- ism for the determination of the transcription-initiation site
3.0] Human h
a, 0.0 3 (0 > -
200 300 3921 c >, 1 100 '6 -3.0 .- - a 3.01 Rat
2 1 0.0 -0 >,
1 100 200 300 392 -3.0
Amino acid number
Fig. 7. Hydrophilicity profiles of human Ser- PyrAT and the mature form of rat mitochondrial Ser-PyrAT. The amino acid sequences of human Ser - PyrAT and the mature form of rat mitochondrial Ser - PyrAT were deduced from cDNAs, pHsptl2 and pRspt321, respec- tively. The average hydrophilicity values for hexapeptides are plotted versus the positions of the amino acid residues. Each point corre- sponds to the center of the respective hexapeptide. The hydrophilicity values for individual amino acids were taken from [43]. Amino acid residues are numbered as described in the legend to Fig. 6

17
Rat (pRSpt321)
50 60 I I I 7 0 80 ATG.TTC.CGG.ATG.TTG.GCC.AAG.GCC.AGT.GTG.ACG.CTG.GGC.TCC. Met-Phe-Arg-Met-Leu-Ala-Lys-Ala-Ser-Val-Thr-Leu-Gly-Ser- -22 ( + ) ( + )
90 100 110 y 120 130 AGG.GCA.GCA.AGT.TGG.GTA.CGG.AAC. ATG.GGC.TCG.CAC.CAG.CTG. Arg-Ala-Ala-Ser-Trp-Val-Arg-Asn- Met-Gly-Ser-His-Gln-Leu-
( + ) -’ L R a t mitochondrial SPT (Rat peroxisomal SPT)
Human (pHsptl2)
1 10 20 30 GCG.GCA.GGT.TGG.GTG.CGG.ACC. ATG.GCC.TCT.CAC.AAG.CTG. (Ala-Ala-Gly-Trp-Val-Arg-Thr) Met-Ala-Ser-His-Lys-Leu-
* * + * * *
L H u m a n SPT
Fig. 8. Comparison of the 5’-terminal regions of the cDNAs for rat Ser - PyrAT and human Ser - PyrAT. The nucleotide sequence of the cDNA for the precursor of rat mitochondrial Ser - PyrAT (pRspt321) was taken from [29], and that for human Ser -PyrAT (pHsptl2) from Fig. 2. The 5’-terminal nucleotide of each cDNA is numbered 1. The 5’-terminal portion of the coding region of pRspt321 and the 5’- terminal portion of pHsptl2, including the noncoding region, are shown together with the amino acid sequences deduced from the nucleotide sequences. Amino acids tentatively deduced from the 5’- noncoding region of the human cDNA are given in parentheses. The amino acid residues are numbered from the N-terminal amino acid of the mature Ser-PyrAT. The N-terminal extension peptide of rat mitochondrial Ser - PyrAT is underlined, the positively charged amino acid residues in the N-terminal extension peptide being indi- cated (+). The downward-pointing arrows and the arrowhead indicate the transcription-initiation site for the shorter mRNA (1.7 kb mRNA) for rat Ser - PyrAT [20] and the site of processing during the translo- cation of rat mitochondrial Ser -PyrAT into mitochondria [50], re- spectively. * and (+) indicate identical and conserved amino acids, respectively. SPT, Ser - PyrAT
may lead to the formation of a translation product which contains the mitochondrial-targeting N-terminal extension peptide. At present the sequence of the human Ser-PyrAT gene preceding the initiator codon is not known, and therefore the above discussion on transcription of human Ser - PyrAT gene is still a matter of conjecture.
This work was supported in part by a Grant-in-Aid (01480153) for Scientific Research and a Grant-in-Aid (01 7701 54) for the Encour- agement of Young Scientists from the Ministry of Education, Science and Culture of Japan. The authors wish to thank Mr Michio Fujie, of the Equipment Center of their medical school, for performing the amino acid analysis, and Dr Kaneo Ohori, of the Department of Public Health, for his help in the computer searches. The authors are also grateful to Dr Tsuneyoshi Funai, of their department, for the valuable discussions and help.
REFERENCES
7600. I . Noguchi, T. & Takada, Y. (1978) J . Biol. Chem. 253, 7598-
2. Yokota, S., Oda, T. & Ichiyama, A. (1987) Histochemistry 87,
3. Noguchi, T. (1987) in Peroxisomes in biology and medicine (Fahimi, H. D. & Sies, H., eds) pp. 234-243, Springer-Verlag, Berlin, Heidelberg.
4. Noguchi, T., Minatogdwa, Y., Takada, Y., Okuno, E. & Kido, R. (1978) Biochem. J. 170, 173-175.
601 - 606.
5. Oda, T., Yanagisawa, M. & Ichiyama, A. (1982) J. Biochem.
6. Miyajima, H., Oda, T. & Ichiyama, A. (1 989) J . Biochem. (Tokyo)
7. Yokota, S. & Oda, T. (1984) Histochemistry 80, 591 -595. 8. Rowsell, E. V., Snell, K., Carnie, J. A. & Al-Tai, A. H. (1969)
9. Snell, K. (1974) Biochem. J . 142, 433-436.
(Tokyo) 91, 219-232.
105, 500 - 504.
Biochem. J . 115, 1071-1073.
10. Sallach, H. J., Sanborn, T. A. &Bruin, W. J. (1972) Endocrinology
11. Thompson, J. S. & Richardson, K . E. (1967) J . Bid. Chem. 242,
12. Feld, R. D. & Sallach, H. J. (1973) Arch. Biochem. Biophys. 159,
13. Noguchi, T., Takada, Y. & Kido, R. (1977) Biochem. J . 161,
14. Yanagisawa, M., Higashi, S., Oda, T. & Ichiyama, A. (1983) in Biochemistry of metabolic processes (Lennon, D. L. F., Stratman, F. W. & Zahlten, R. N., eds) pp. 41 3 -426, Elsevier Biomedical, New York, Amsterdam, Oxford.
91, 1054-2063.
3614-3619.
757 - 766.
609-614.
15. Noguchi, T. & Takada, Y. (1978) Biochem. J . 175,765-768. 16. Okuno, E., Minatogawa, Y., Nakamura, M., Kamoda, N.,
Nakanishi, J., Makino, M. & Kido, R. (1980) Biochem. J. 189,
17. Danpure, C. J. & Jennings, P. R. (1986) FEBS Lett. 201, 20-
18. Wise, P. J., Danpure, C. J. & Jennings, P. R. (1987) FEBS Lett.
19. Richardson, K. E. & Tolbert, N. E. (1961) J. Biol. Chem. 236,
20. Oda, T., Funai, T. & Ichiyama, A. (1990) J . Bid. Chem. 265,
21. Lazarow, P. B. & Fujiki, Y. (1985) Annu. Rev. Cell Biol. 1, 489-
22. Borst, P. (1986) Biochim. Biophys. Acta 866, 179-203. 23. Danpure, C. J., Cooper, P. J., Wise, P. J. & Jennings, P. R. (1989)
24. Chomczynski, P. & Sacchi, N. (1987) Anal. Biochem. 162, 156-
25. Aviv, H. & Leder, P. (1972) Proc. Natl Acad. Sci. USA 69,1408 -
26. Okayama, H. & Berg, P. (1982) Mol. Cell. Biol. 2, 161 - 170. 27. Gubler, U. & Hoffman, B. J. (1983) Gene 25,263-269. 28. Mandel, M. & Higa, A. (1970) J . Mol. Biol. 53, 159- 162. 29. Oda, T., Miyajima, H., Suzuki, Y. & Ichiyama, A. (1987) Eur. J .
Biochem. 168, 537- 542. 30. Feinberg, A. P. & Vogelstein, B. (1983) Anal. Biochem. 132, 6-
13. 31. Maniatis, T., Fritsch, E. F. & Sambrook, J. (1982) Molecular
cloning, a laboratory manual, p. 448, Cold Spring Harbor Lab- oratory, New York.
581 -590.
24.
222, 17 - 20.
1280 - 1284.
7513-7519.
530.
J . Cell Biol. 108, 1345 - 1352.
159.
141 2.
32. Vieira, J. & Messing, J. (1987) Methods Enzymol. 153, 3 - 11. 33. Sanger, F., Nicklen, S. & Coulson, A. R. (1977) Proc. Natl Acad.
34. Alwine, J. C., Kemp, D. J. & Stark, G. R. (1977) Proc. Nut1 Acad.
35. Mori, M., Miura, S., Tatibana, M. & Cohen, P. P. (1979) Proc.
36. Oda, T., Ichiyama, A,, Miura, S., Mori, M. & Tatibana, M. (1981)
37. de Duve, C., Pressman, B. C., Gianetto, R., Wattiaux, R. &
38. Laemmli, U. K. (1970) Nature 227,680-685. 39. Lowry, 0. H., Rosebrough, N. J., Farr, A. L. & Randall, R. J.
(1951) J . Biol. Chem. 193, 265-275. 40. Kozak, M. (1981) Nucleic Acids Res. 9, 5233-5252. 41. Oda, T., Kitamura, N., Nakanishi, S. & Ichiyama, A. (1985) Eur.
42. Oda, T., Ichiyama, A,, Miura, S. & Mori, M. (1984) J . Biochem.
43. Hopp, T. P. & Woods, K. R. (1981) Proc. Nut1 Acad. Sci. USA
Sci. USA 74, 5463 - 5467.
Sci. USA 74, 5350- 5354.
Natl Acad. Sci. USA 76, 5071 - 5075.
Biochem. Biophys. Res. Commun. 102, 568 - 573.
Appelmans, F. (1955) Biochem. J . 60, 604-617.
J . Biochem. 150,415 -421.
(Tokyo) 95, 81 5 - 824.
78, 3824-3828.

18
44. Pearson, W. R. & Lipman, D. J. (1988) Proc. Nut1 Acad. Sci.
45. Gould, S. J., Keller, G.-A. & Subramani, S . (1988) J. Cell Biol.
46. Gould, S. J., Keller, G.-A., Hosken, N., Wilkinson, J. & Sub-
47. Quan, F., Korneluk, R. G., Tropak, M. B. & Gravel, R. A. (1986)
USA 85, 2444-2448.
107, 897 - 905.
ramani, s. (1989) J . Cell Biol. 108, 1657- 1664.
Nucleic Acid7 Res. 14. 5321 - 5335.
48. Dayhoff, M. 0. (1978) Atlas of protein sequence and structure, vol. 5 , suppl. 3, National Biochemical Research Foundation, Washington.
49. Melton, D. A., Krieg, P. A., Rebagliati, M. R., Maniatis, T., Zinn, K. & Green, M. R. (1984) Nucleic Acids Res. 12, 7035- 7056.
50. Oda, T., Miyajima, H., Suzuki, Y., Ito, T., Yokota, S., Hoshino, M. & Ichiyama, A. (1989) J . Biochem. (Tokyo) 106, 460- 467.


![Tyrosine Aminotransferase Contributes to ... · Tyrosine Aminotransferase Contributes to Benzylisoquinoline Alkaloid Biosynthesis in Opium Poppy1[W] Eun-Jeong Lee and Peter J. Facchini*](https://img.pdfslide.us/doc/110x75/5b85532a7f8b9a4a488e07e1/tyrosine-aminotransferase-contributes-to-tyrosine-aminotransferase-contributes.jpg)





![Research Paper The Prognostic Value of aspartate aminotransferase … · 2019-05-22 · Aspartate aminotransferase (AST) to lymphocyte ratio (ALRI) [17], systemic immune-inflammation](https://img.pdfslide.us/doc/110x75/5f0222077e708231d402bbfe/research-paper-the-prognostic-value-of-aspartate-aminotransferase-2019-05-22-aspartate.jpg)