Embed Size (px)
Citation preview

Cloning and molecular characterization of the citrate utilizationcitMCDEFGRP cluster of Leuconostoc paramesenteroides
Mauricio Mart|èn a;b, Maria Angeles Corrales a, Diego de Mendoza b,Paloma Loèpez a;*, Christian Magni b
a Centro de Investigaciones Bioloègicas (CSIC), Velaèzquez 144, 28006 Madrid, Spainb Programa Multidisciplinario de Biolog|èa Experimental (PROMUBIE, CONICET) and Departamento de Microbiolog|èa, Facultad de Ciencias
Bioqu|èmicas y Farmaceuèticas, Universidad Nacional de Rosario, Suipacha 531, 2000 Rosario, Argentina
Received 11 February 1999; received in revised form 17 March 1999; accepted 17 March 1999
Abstract
The citMCDEFGRP cluster from Leuconostoc paramesenteroides involved in citrate utilization was cloned and its nucleotidesequence determined. Homology of the inferred gene products with characterized enzymes reveals that citP encodes the citratepermease P, citC the citrate ligase and citDEF the subunits of the citrate lyase of Leuconostoc. Moreover, it suggests that citMencodes a Leuconostoc malic enzyme. Analysis of citrate consumption by and citrate lyase activity of Lc. paramesenteroidesJ1[pCITJ1] showed that its citrate permease and its citrate lyase are induced by the presence of citrate in the growth medium.Southern blot analysis demonstrated that the citMCDEFGRP cluster is located in a plasmid. z 1999 Federation of EuropeanMicrobiological Societies. Published by Elsevier Science B.V. All rights reserved.
Keywords: Lactic acid bacterium; Leuconostoc ; Citrate permease; Citrate lyase; Citrate utilization
1. Introduction
Metabolism of citrate by a limited number of lac-tic acid bacteria (LAB) is relevant to the food indus-try, since the production of CO2 and diacetyl fromcitrate is essential for the development of the aromaand £avor of dairy products [1]. The citrate is trans-ported inside the cell by a permease and is convertedto pyruvate by the concerted action of citrate lyaseand oxaloacetate decarboxylase [1]. The citrate per-mease P (CitP) encoded by the citP gene is respon-sible for citrate uptake in Lactococcus lactis biovar
diacetylactis (L. diacetylactis) [2] and Leuconostoc(lactis and mesenteroides) [3]. The citP genes ofboth genera are almost identical [4]. The regulationof expression of citP has been extensively studied inLactococci (reviewed in [5]). The lactococcal citP isharbored by the 8-kb pCIT264 plasmid [2] and in-cluded in the citQRP operon [6], which encodes thecitrate transport system. Transcription of this operonis mainly driven from the pH-regulated P1 promoter[6], and it is induced by acidi¢cation of the growthmedium [7]. The P1 promoter is located 1 kb up-stream of the citQ gene, and this 1-kb DNA regionincludes the insertion sequence (IS) element IS982.The introduction of this IS has provided the citQRPoperon with a new promoter (P2), which increases its
0378-1097 / 99 / $20.00 ß 1999 Federation of European Microbiological Societies. Published by Elsevier Science B.V. All rights reserved.PII: S 0 3 7 8 - 1 0 9 7 ( 9 9 ) 0 0 1 4 5 - 7
FEMSLE 8740 29-4-99
* Corresponding author. Tel. : +34 (1) 5611 800, ext. 4203;Fax: +34 (1) 562 7518; E-mail: [email protected]
FEMS Microbiology Letters 174 (1999) 231^238
expression in Lactococci [8]. It has been establishedthat the citP from Leuconostoc is localized in a 22-kbuncharacterized plasmid [9]. The genes encodingCitP [4] and the citrate lyase [10] have been inde-pendently cloned from genomic DNA of two di¡er-ent Lc. mesenteroides strains. Moreover, it has been
suggested that in this bacteria citrate utilization isinduced by the presence of this compound in thegrowth medium [11]. However, the genetic organiza-tion and the molecular regulation of the citratetransport system from Leuconostoc as well as its con-nection with the citrate lyase modulation are un-
FEMSLE 8740 29-4-99
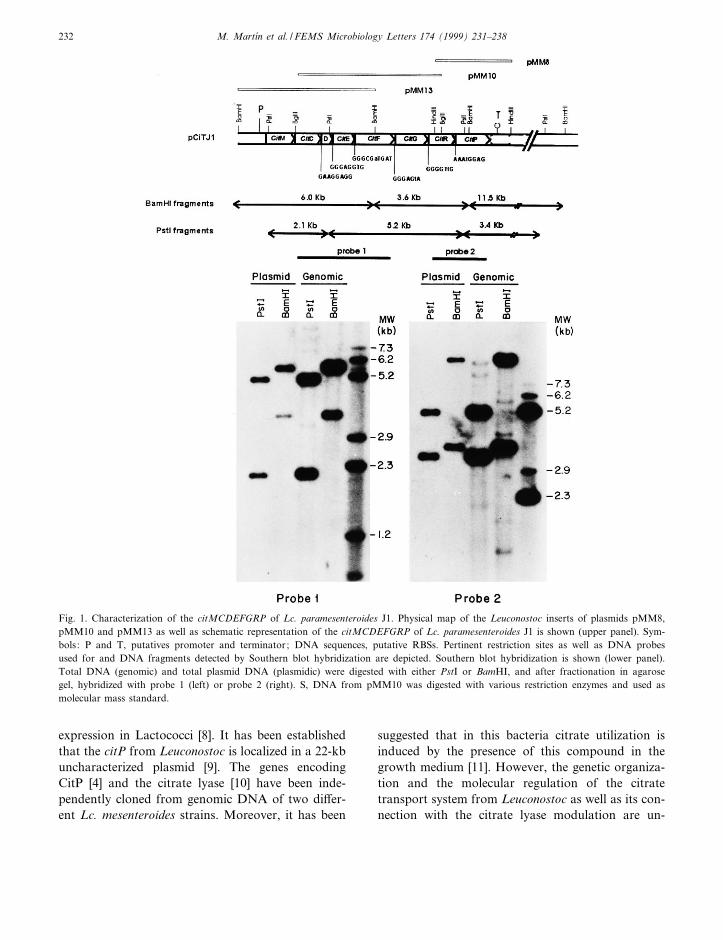
Fig. 1. Characterization of the citMCDEFGRP of Lc. paramesenteroides J1. Physical map of the Leuconostoc inserts of plasmids pMM8,pMM10 and pMM13 as well as schematic representation of the citMCDEFGRP of Lc. paramesenteroides J1 is shown (upper panel). Sym-bols : P and T, putatives promoter and terminator ; DNA sequences, putative RBSs. Pertinent restriction sites as well as DNA probesused for and DNA fragments detected by Southern blot hybridization are depicted. Southern blot hybridization is shown (lower panel).Total DNA (genomic) and total plasmid DNA (plasmidic) were digested with either PstI or BamHI, and after fractionation in agarosegel, hybridized with probe 1 (left) or probe 2 (right). S, DNA from pMM10 was digested with various restriction enzymes and used asmolecular mass standard.
M. Mart|èn et al. / FEMS Microbiology Letters 174 (1999) 231^238232

known. In this work, we have cloned and establishedthe organization of the Lc. paramesenteroides plas-midic citMCDEFGRP gene cluster, which seems toencode a citrate permease and a citrate lyase induci-ble by citrate.
2. Materials and methods
2.1. Bacterial strains, growth media and plasmidtransfer
The bacterial strains used in this work were: Leu-conostoc paramesenteroides J1[pCITJ1] and Escheri-chia coli DH5K [12]. Lc. paramesenteroides wasgrown at 30³C without shaking in MRS medium[9] lacking ammonium citrate and containing 2% glu-cose (MRSG) or MRSG supplemented with 1% so-dium citrate (MRSGC). E. coli was grown in Luria-Bertani broth [12] and transformed as previously de-scribed [8]. Transformants were selected in agar me-dium containing ampicillin at 50 Wg ml31. The me-dium used for screening citrate utilization by E. coliwas Christensen agar (Difco Laboratories).
2.2. DNA analysis and manipulation
Plasmidic DNA for cloning and sequencing experi-ments was isolated from Lc. paramesenteroides andE. coli as described [4,12]. The DNA sequence ofboth strands of the cit cluster was determined fromthe pMM8, pMM10 and pMM13 plasmids with theautomated DNA sequencing instrumentation (ABIPRISM0, Perkin Elmer) at the Centro de Investiga-ciones Bioloègicas. For Southern blot analysis, ge-nomic DNA was prepared from cultures grown tothe stationary phase. Cells were sedimented and re-suspended in lysis bu¡er containing 50 mM Tris-HCl, 0.25% sucrose, 0.58% NaCl, lysozyme at 25mg ml31 and RNase A at 400 Wg ml31. After incu-bation for 30 min at 37³C, 1% SDS and proteinase Kat 20 Wg ml31 were added to the extract and incuba-tion proceeded for 30 min. Then, total extracts werephenol extracted and ethanol precipitated. PlasmidicDNA was separated from chromosomal DNA byfractionation in CsCl gradients following standardprocedures. DNAs were fractionated in a 0.7% agar-ose gel, transferred to a nylon membrane and South-
ern blot performed applying stringent hybridizationand washing conditions [12]. Restriction fragmentsof pMM10 were 32P-labeled with the multiprimeDNA labeling sequence system (Amersham) andused as a probe (see details in Fig. 1).
2.3. Determination of the citrate permease and citratelyase activities
To determine citrate permease activity, Leuconos-toc cultures were grown in either MRSG or MRSGCto an absorbance of 0.6 at 650 nm. Then cultureswere sedimented, resuspended in MRSG mediumsupplemented with 0.1% sodium citrate and treatedat 30³C with rifampicin at 200 Wg ml31 and chlor-amphenicol at 5 Wg ml31. At the time indicated inFig. 2 cultures were centrifuged, and the concentra-tion of citric acid present in the supernatants assayedenzymatically by using the test kit 139076 fromBoehringer Mannheim. To determine citrate lyaseactivity, Leuconostoc cultures were grown in eitherMRSG or MRSGC to an absorbance of 0.6 at 650nm, and total extracts prepared as previously de-scribed for L. lactis [5]. Citrate lyase activity wasdetermined from samples of these extracts containing
FEMSLE 8740 29-4-99
Fig. 2. Induction of citrate utilization by citrate in Lc. parame-senteroides J1[pCITJ1]. Exponential cultures grown in the pres-ence (b) or in the absence of citrate (a) were treated with rifam-picin and chloramphenicol and challenged with 0.1% citrate.Then at the indicated time cultures were centrifuged and concen-tration of citrate present in the supernatants determined. See de-tails in Section 2.
M. Mart|èn et al. / FEMS Microbiology Letters 174 (1999) 231^238 233

20 Wg of proteins at 25³C in a coupled spectrophoto-metric assay with malate and lactate dehydrogenasesas described by Bekal et al. [10]. One unit of enzymeactivity is de¢ned as 1 pmol of citrate converted toacetate and oxaloacetate per min.
3. Results
3.1. Cloning of the citrate utilization system of Lc.paramesenteroides J1 strain
The citrate permease P catalyses the transport ofcitrate in Lactococcus and Leuconostoc [2,3]. The twoCitP carriers have 99% identity of amino acids [4],however expression of this permease seems to beregulated di¡erentially in these two LAB [8,11].These results suggested to us the existence of a di¡er-ential gene organization for the citrate transport sys-tem of Lactococcus and Leuconostoc. Therefore, weattempted the isolation of Leuconostoc strains able toutilize citrate and harboring a plasmidic citratetransport operon lacking the lactococcal IS982 [5].To this end, total plasmid DNA from 30 bacterialstrains isolated from Argentinean cheeses werescreened by dot blot hybridization with two DNAprobes from plasmid pCIT264 of L. diacetylactis.These probes included DNA sequences from eitherthe citP gene or the IS982 (data not shown). A bac-terial strain named J1, which showed a citP� IS3
genotype, was isolated. This strain contains at leasttwo plasmids and was classi¢ed as Leuconostoc para-mesenteroides by its metabolic properties as de-scribed by Villani et al. [13]. The capability of Lc.paramesenteroides J1 to utilize citrate was establishedby its ability to produce diacetyl from citrate as de-scribed by Kin [14]. In order to clone the L. para-mesenteroides J1 citP gene, a plasmidic 2.4-kb Hin-dIII DNA fragment was identi¢ed by Southern blothybridization with the citP probe (data not shown).This DNA fragment was cloned into the HindIII siteof pBluescript SK� E. coli vector and the resultingplasmid was designated pMM8 (Fig. 1). E. coliDH5K, DH5K[ pBlueSK+] and DH5K[pMM8]strains were tested on Christensen citrate agar plates,on which colonies containing cells able to take upcitrate (Cit�) have a red halo around the colonies.Only strain DH5K[pMM8] showed a Cit� pheno-
type. This result indicated that a functional citPgene was present in the pMM8 plasmid. The regionlocated upstream of citP was detected by hybridiza-tion with the pMM8 insert. Two overlapping 5.8-kbBglII and 6.0-kb BamHI DNA fragments werecloned into the BamHI site of pBluescript SK� vec-tor and the resulting recombinant plasmids desig-nated pMM10 and pMM13 (Fig. 1).
3.2. Molecular characterization of thecitMCDEFGRP cluster of Lc. paramesenteroidesJ1 strain
The nucleotide sequence of the DNA segments ofLc. paramesenteroides cloned in plasmids pMM8,pMM10 and pMM13 was determined and depositedin the EMBL Sequence Database with the accessionnumber AJ132782. Nucleotide sequence analysis ofthe 8800-bp DNA region revealed the presence ofeight open reading frames transcribed in the samedirection. The cluster was named citMCDEFGRPand its £anking putative transcriptional promoterand terminator were identi¢ed. Preceding citM wasobserved a canonical 310 hexamer TATAAT and a335 hexamer TTtACA which shears ¢ve residueswith the consensus sequence of c70 promoters. Aninverted repeat TAGGCAGCTGACTTCACATTG-TTGAGATCAGCTGCCTTTT, which is character-istic of the b-independent terminators, was detectedat 103 nucleotides (nt) downstream of citP. Preced-ing the ¢rst ATG of citE, citF, citG, citR and citPputative ribosomal binding sites (RBSs) complemen-tary to the 3P-end of the Leuconostoc 16S rRNA 5P-ATCACCTCCTTT-3P [15] were observed (see Fig.1). These RBSs have, respectively, a predicted freeenergy of binding (vG in kcal mol31) to the 3P-endof the 16S rRNA of 310.3, 39.5, 35.2, 34.1 and33.1 according to the rules of Schurr et al. [16]. ThecitM should encode a 40-kDa polypeptide. This poly-peptide showed homology with malic enzymes fromorganisms belonging to the three kingdoms: prokar-ya, archaea (Table 1) and eukarya (data not shown).The citC gene, which starts 12 nt downstream ofcitM, should encode a 39-kDa polypeptide.This poly-peptide showed a signi¢cant identity with the citratelyase ligase of Lc. mesenteroides, Haemophilus in£u-enzae, Klebsiella pneumoniae and E. coli (Table 2). Inaddition, the citDEF genes seem to encode the Q, L
FEMSLE 8740 29-4-99
M. Mart|èn et al. / FEMS Microbiology Letters 174 (1999) 231^238234

and K subunits of the citrate lyase, since their geneproducts are homologous to their counterparts ofLc. mesenteroides, H. in£uenzae, K. pneumoniae andE. coli (Table 2). The citD gene overlaps the citCcoding region. Alignment of the inferred polypeptidewith the citD gene product of four di¡erent bacteria(Table 2) predicted that the second ATG of the openreading frame is the start codon for translation ofcitD in L. paramesenteroides. Supporting this hy-pothesis, the second and not the ¢rst ATG of citDis preceded by a putative RBS (see Fig. 1), which hasa predicted free energy of 39.0 kcal mol31. On thesebases, it is expected that the Leuconostoc ribosomesshould be able to utilize this RBS. If the formerassumption is correct, citD encodes a 10-kDa poly-peptide. The putative start ATG codon of citE islocated 1 nt downstream citD, and citE should en-code a 34-kDa polypeptide. The deduced 42 aminoacids from the N-terminal region showed 100% iden-tity (data not shown) with the amino acid sequenceof the citrate lyase b subunit determined from Lc.mesenteroides subsp. dextranicus 18T [10]. The citF
gene should encode a 50-kDa polypeptide and it islocated 130 nt downstream and 43 nt upstream ofcitE and citG, respectively. The citG gene shouldencode a polypeptide of 51 kDa, which is homolo-gous to its counterparts of Lc. mesenteroides, H. in-£uenzae, K. pneumoniae and E. coli, with unknownfunction (Table 2). The citR gene is located 1 ntdownstream and 25 nt upstream of citG and citP,respectively. This gene should encode a polypeptideof 30 kDa containing 265 amino acids. The 197 ami-no acids located at the C-terminal region of thispolypeptide showed 97% identity (data not shown)with the CitR regulator of L. diacetylactis [6], whichlacks the N-terminal region of its counterpart fromLeuconostoc. The citP gene should encode a 47-kDapolypeptide. The inferred polypeptide showed as ex-pected 99.2% and 90% identity with the citrate per-mease P of L. diacetylactis and Lc. mesenteroides(data not shown). The aforementioned results indi-cated that the citrate transport system, the citratelyase complex and a malic enzyme are encoded bylinked genes in Lc. paramesenteroides.
FEMSLE 8740 29-4-99
Table 2Homology of the citCDEFG gene products of Lc. paramesenteroides, Lc. mesenteroides, H. in£uenzae, E. coli and K. pneumoniae
Gene Lc. paramesenteroides Lc. mesenteroides H. in£uenzae E. coli K. pneumoniae
CitC 100 47.8 34.6 30.9 32.5CitD 100 69.1 49.5 44.9 41.2CitE 100 83.4 51.5 49.0 54.3CitF 100 82.8 56.1 55.5 54.8CitG 100 48.9 32.9 35.3 30.1
Amino acid sequences were obtained from GenBank accession numbers Y10621 (Lc. mesenteroides), U32687, U32688 (H. in£uenzae), X79817(K. pneumoniae) and AE000166 (E. coli). Homology is given as percentage identity to Lc. paramesenteroides genes.
Table 1Homology of citM gene product to malic enzymes from di¡erent micro-organisms
Organism GenBank accession number Function Homology to CitMa
Leuconostoc mesenteroides Y10621 Malic enzyme (putative) 66.7Bacillus subtilis P16468 Malate oxidoreductase, oxaloacetate decarboxylase 46.8Synechocystis sp. D1017396 Malic enzyme 43.7Streptococcus bovis Q59826 Malate oxidoreductase, oxaloacetate decarboxylase 43.7Archaeoglobus fulgidus O28547 Malate oxidoreductasa 42.9Pyrococcus horikoshii D1028412 Malate oxidoreductasa 41.3Escherichia coli P78201 Malic enzyme (putative) 38.4Haemophilus in£uenza P43837 Malic enzyme (putative) 38.4Sinorhizobium meliloti O30808 Malic enzyme 36.6
aHomology was determined with the MegAlign program (DNASTAR) and it is expressed as percentage of identity to CitM.
M. Mart|èn et al. / FEMS Microbiology Letters 174 (1999) 231^238 235

3.3. Plasmidic location of the Lc. paramesenteroidescitMCDEFGRP cluster
The cloning strategy utilized in this work indicatedthat the cit cluster is carried by a plasmid. However,as far as we know these results constitute the ¢rstreport of a plasmidic location of the citrate lyasegenes in LAB. To con¢rm this assumption, total ge-nomic DNA and plasmidic DNA puri¢ed by CsClgradient were digested with either PstI or BamHI,fractionated in agarose gels and hybridized withtwo probes (Fig. 1). Probe 1 contains citCDEF,whereas probe 2 includes the 3P-end of citR andthe citP. Both DNA preparations showed the samehybridization pattern with each probe, revealing thatindeed the citMCDEFGRP cluster is present in aplasmid. The DNA fragments detected in the BamHIdigest indicate that the size of the Cit� plasmid (des-ignated pCITJ1) is approximately 21.1 kb. This re-sult correlates to the previous ascription of the Leu-conostoc citP to a 22-kb plasmid [9], and tohybridization of both probes with an undigestedplasmid of this size (result not shown). Moreover,detection of the 5.2-kb PstI and the 3.6-kb BamHIDNA fragments with both probes (Fig. 1) clearlyshows that the citrate lyase and the citrate permeasegenes are linked in the pCITJ1 plasmid.
3.4. In£uence of citrate in the levels of citratepermease and citrate lyase ofLc. paramesenteroides J1
The presence of citrate in the growth medium in-duces the citrate transport system of Lc. mesenter-oides [11] but does not a¡ect that of L. lactis [2]. Onthese basis, we analyzed the ability of Lc. parame-senteroides J1[pCITJ1] to utilize citrate after growthin the presence or in the absence of this compound.The ability of this strain to take up citrate was testedby means of citrate consumption (Fig. 2). For thisanalysis, exponential cultures grown either in MRSGor MRSGC were centrifuged and resuspended inMRSG medium containing 0.1% citrate and treatedwith rifampicin and chloramphenicol to inhibit denovo synthesis of proteins as described in Section2. At the time indicated in Fig. 2 samples were with-drawn, cells were removed by centrifugation and theconcentration of citrate present in the supernatants
determined. Cultures previously grown in the pres-ence of citrate were able to take up more than 99.9%of the citrate present in the medium after an incuba-tion period of 60 min at 30³C. By contrast, signi¢-cant incorporation of citrate was not detected duringthe sampling period with cultures previously grownin the absence of citrate. These results indicate thatsynthesis of the citrate permease P is induced bycitrate. In addition, the in£uence of growth mediumin levels of citrate lyase of Lc. paramesenteroidesJ1[pCITJ1] was tested. For this analysis, total ex-tracts were prepared from exponential culturesgrown either in MRSG or MRSGC and citrate lyaseactivity determined as described in Section 2.Growth in medium containing citrate resulted in athree fold increase of the citrate lyase activity, since0.51 þ 0.02 units mg31 and 1.71 þ 0.04 units mg31
were detected, respectively, in cultures grown inMRSG and MRSGC. The overall results show thatboth citrate transport and its conversion to acetateand oxaloacetate are induced by the presence of cit-rate in the growth medium. Furthermore, dot blothybridization analysis indicates that this inductiontakes place at the transcriptional level (results notshown).
4. Discussion
In this work we have shown that the cit-MCDEFGRP cluster of Lc. paramesenteroides J1 islocated in a 21-kb plasmid. This is the ¢rst reportshowing that genes involved in the transport andmetabolism of citrate are linked in a Gram-positivebacterium. Homology of the gene products with pre-viously characterized enzymes indicates that they en-code enzymes that are speci¢cally required for citratefermentation. The CitP is the citrate permease, whichcatalyses the transport of citrate inside the cells. ThecitCDEFG gene products should be involved in theconversion of citrate to acetate and oxaloacetate.The expected encoded enzymes are a ligase (CitC)catalyzing the ATP-dependent acetylation of thephosphoribosyl dephospho-coenzyme A group of cit-rate lyase, the three di¡erent subunits of citrate lyase(CitDEF), and a protein of unknown function(CitG) which has been proposed to be involved inthe conversion of apo-citrate to holocitrate lyase
FEMSLE 8740 29-4-99
M. Mart|èn et al. / FEMS Microbiology Letters 174 (1999) 231^238236

[18]. This cluster of the citCDEFG genes is alsopresent in the chromosome of K. pneumoniae andH. in£uenzae (see Fig. 3) and in Lc. mesenteroides195 strain [10] at an unknown location (not shown).In addition, in the E. coli chromosome a cit-CDEFXG cluster of similar con¢guration is present,citX being a gene of unknown function. However,the genes encoding the citrate carriers, althoughthey are proximal to citCDEFG in the Gram-nega-tive bacteria and in Lc. paramesenteroides, have dif-ferent arrangements in the various hosts (Fig. 2). ThecitS gene of K. pneumoniae is included within thechromosomal citrate regulon, which is induced bycitrate [17], but citS is located divergently to cit-CDEFG. This gene is cotranscribed with the oadGABgenes, which encode the subunits of the oxaloacetatedecarboxylase responsible for conversion of oxaloa-cetate to pyruvate [17]. The citS gene encodes theCitS carrier, which has 33.8% identity with theCitP of Lc. paramesenteroides, and citP and citSseem to be evolved from a common ancestor gene.The citT and Ybh1 of E. coli and H. in£uenzae en-code citrate carriers with 38% sequence identity,which belong to a family of permeases that doesnot include CitP [18]. These citrate carrier genesare located downstream of citG like citP in Lc. para-mesenteroides, but the citR gene is not present in theGram-negative genomes. The presence of citR up-
stream of citP seems to be a gene organization char-acteristic of LAB, since we have previously shownthat in L. lactis a citQRP operon carried by thepCIT264 plasmid exists ([6] and Fig. 3). However,the citQ, which encodes a leader peptide in Lacto-cocci, is absent in Leuconostoc. Moreover, CitR is aregulator in L. lactis [6] and its counterpart fromLeuconostoc of unknown function seems to be com-posed of two domains: a C-domain which has 97%identity with CitR of Lactococci and a N-domainwhich has homology to esterases and peroxidases(data not shown). In addition, in L. diacetylactisthe genes encoding for the citrate lyase are locatedin the chromosome [19]. The presence of the citMgene in a citMCDEFG cluster seems to be character-istic of Leuconostoc, because it has been detected inLc. paramesenteroides J1 (this report) and in Lc. mes-enteroides 195 (accession number Y10621, unpub-lished results of Bekal et al.). The citM gene encodesa polypeptide which has signi¢cant homology withmalic enzymes (Table 1). Some of these enzymes areable to catalyze conversion of oxaloacetate to pyru-vate (Table 1). Therefore, it is feasible that in Lc.paramesenteroides the cit operon encodes enzymesthat support transport and conversion of citrate topyruvate. We have shown that in Lc. paramesenter-oides the transport of citrate and citrate lyase activityare induced by the presence of citrate in the growth
FEMSLE 8740 29-4-99
Fig. 3. Genetic organization of genes involved in citrate utilization in LAB and Gram-negative bacteria. Characterized and putative pro-moters (P) are indicated. Homologous genes are shaded or dashed in the same color. See details in the text.
M. Mart|èn et al. / FEMS Microbiology Letters 174 (1999) 231^238 237

medium (Fig. 2 and data in the text), whereas in L.diacetylactis transcription of the citQRP is inducedat acidic pH [7]. Moreover, Leuconostoc is able togrow with citrate as a sole carbon source ([9] andresults not shown), whereas Lactococci requires co-metabolism of sugar and citrate for e¤cient growth[7]. Therefore, the di¡erential gene organization inLactococcus and Leuconostoc is likely to re£ect dif-ferent physiological functions of citrate metabolismin these two genus.
Acknowledgments
We thank M. Espinosa for helpful discussions andF. Sesma for providing LAB strains for screening.This work was partially supported by exchangegrants of Consejo superior de Investigaciones Cien-ti¢cas (CSIC), Spain and Consejo Nacional de Inves-tigaciones Cient|è¢cas (CONICET), Argentina. Re-search at the Centro de Investigaciones Bioloègicaswas under the auspices of CSIC and was supportedby the Comisioèn Interministerial de Ciencia y Tec-nolog|èa Grant BIO97-0347 and Comunidad de Ma-drid Grant 06G/002/96. Research at PROMUBIEwas supported by CONICET, Fundacioèn Antorchasand Agencia Nacional de Promocioèn Cient|è¢ca yTecnoloègica.
References
[1] Cocaign-Bousquet, M., Garrigues, C., Loubiere, P. and Lind-ley, N.D. (1996) Physiology metabolism of pyruvate in Lacto-coccus lactis. Antonie van Leeuwenhoek 70, 253^267.
[2] Magni, C., Loèpez de Felipe, F., Sesma, F., Loèpez, P. and deMendoza, D. (1994) Citrate transport in Lactococcus lactissubsp. lactis biovar diacetylactis. Expression of the citrate per-mease P. FEMS Microbiol. Lett. 118, 75^82.
[3] Bandell, M., Lhotte, M.E., Marty-Teyssset, C., Veyrat, A.,Prevoèst, H., Dartois, V., Divieés, C., Konings, W.N. and Lol-kema, J.S. (1998) Mechanism of the citrate transporters incarbohydrate and citrate cometabolism in Lactococcus andLeuconostoc species. Appl. Environ. Microbiol. 64, 1594^1600.
[4] Vaughan, E.E., David, S., Harrington, A., Daly, C., Fitzger-ald, C.F. and de Vos, W.M. (1995) Characterization of plas-mid-encoded citrate permease (citP) genes from Leuconostocspecies reveals high sequence conservation with the Lactococ-cus lactis citP gene. Appl. Environ. Microbiol. 61, 3172^3176.
[5] Loèpez, P., Drider, D., Garc|èa-Quintaèns, N., Corrales, M.A.,Magni, C., Mart|èn, M. and de Mendoza, D. (1998) Regulation
of expression of the Lactococcus lactis biovar diacetylactiscitrate transport system. Lait 78, 11^16.
[6] Loèpez de Felipe, F., Magni, C., de Mendoza, D. and Loèpez,P. (1995) Citrate utilization gene cluster of the Lactococcuslactis biovar diacetylactis : organization and regulation of ex-pression. Mol. Gen. Genet. 246, 590^599.
[7] Garc|èa-Quintaèns, N., Magni, C., de Mendoza, D. and Loèpez,P. (1998) The citrate transport system of Lactococcus lactissubsp. Lactis biovar diacetylactis is induced by acid stress.Appl. Environ. Microbiol. 64, 850^857.
[8] Loèpez de Felipe, F., Magni, C., de Mendoza, D. and Loèpez,P. (1996) Transcriptional activation of the citrate permease Pgene of Lactococcus lactis biovar diacetylactis. Mol. Gen.Genet. 250, 428^436.
[9] Lin, J., Schmitt, P. and Divieès, C. (1991) Characterization of acitrate negative mutant of Leuconostoc ssp. mesenteroides :metabolic and plasmidic properties. Appl. Microbiol. Biotech-nol. 34, 628^631.
[10] Bekal, S., Van Beeumen, J., Samyn, B., Garmyn, D., Heinini,S., Divieés, C. and Prevoèst, H. (1998) Puri¢cation of Leuconos-toc mesenteroides citrate lyase and cloning and characteriza-tion of the citCDEFG gene cluster. J. Bacteriol. 180, 647^654.
[11] Marty-Teysset, C., Lolkema, J.S., Schmitt, P., Divieés, C. andKonings, W.N. (1996) Proton motive force generation by cit-rolactic fermentation in Leuconostoc mesenteroides. J. Bacter-iol. 178, 2178^2185.
[12] Sambrook, J., Fritsch, E.F. and Maniatis, T. (1989) MolecularCloning: A Laboratory Manual. Cold Spring Harbor Labo-ratory, Cold Spring Harbor, NY.
[13] Villani, F., Moschetti, G., Blaiotta, G. and Coppola, S. (1997)Characterization of strains of Leuconostoc mesenteroides byanalysis of soluble whole-cell protein pattern, DNA ¢nger-printing and restriction of ribosomal DNA. J. Appl. Micro-biol. 82, 578^588.
[14] Kin, N. (1948) Modi¢cation of the Voges-Proskauer test forrapid colorimetric determination of acetylmethylcarbinol plusdiacetyl in butter cultures. Dairy Ind. 13, 860^866.
[15] Mart|ènez-Murcia, A.J. and Collins, M.D. (1990) A phyloge-netic analysis of the genus Leuconostoc based on reverse tran-scriptase sequencing of 16 S rRNA. FEMS Microbiol. Lett.70, 73^84.
[16] Schurr, T., Nadir, E. and Margalit, H. (1993) Identi¢cationand characterization of the Escherichia coli ribosomal bindingsites by free energy computation. Nucleic Acids Res. 21, 4019^4023.
[17] Meyer, M., Dimroth, P. and Bott, M. (1997) In vitro bindingof the response regulator CitB and its carboxy-terminal do-main to A+T-rich DNA target sequences in the control regionof the divergent citC and citS operons of Klebsiella pneumo-niae. J. Mol. Biol. 269, 719^731.
[18] Pos, K.M., Dimroth, P. and Bott, M. (1998) The Escherichiacoli citrate carrier CitT: a member of a novel eubacterial trans-porter family related to the 2-oxoglutarate/malate translocatorfrom spinach chloroplasts. J. Bacteriol. 180, 4160^4165.
[19] Hill, C., Daly, C. and Fitzgerald, G.F. (1991) Isolation ofchromosomal mutations of Lactococcus lactis biovar diacety-lactis 18-16 after introduction of Tn919. FEMS Microbiol.Lett. 81, 135^140.
FEMSLE 8740 29-4-99
M. Mart|èn et al. / FEMS Microbiology Letters 174 (1999) 231^238238