-
PhycologyCourse handouts
ChlorophytesKalle Olli1
Abstract
Chlorophyta are commonly known as the green algae or
chlorophytes, because they appear bright grass green, as domost
plants. This is because the chlorophylls a and b of green algae are
usually not concealed by large amounts ofaccessory pigments.Indeed,
plants have evolved from green algae, and in addition so similar
pigmentation, there are other structuralsimilarities between
terrestrial plants and chlorophytes. Firstly, the architecture of
chloroplasts is very similar. Also,both use cellulose as the
structural material to build cell walls.The poor set of accessory
makes chlorophytes relatively inefficient light users, and in many
aquatic habitatschlorophytes grow well in the well illuminated
layers.Chlorophytes form starch with the chloroplast (as do
plants), usually in association with a pyrenoid (which plants donot
have). The Chlorophyta thus differ from the rest of the eukaryotic
algae in forming the storage product in thechloroplast instead of
in the cytoplasm.The Chlorophyta are primarily freshwater; only
about 10% of the algae are marine, whereas 90% are freshwater.Some
orders are predominantly marine (Caulerpales, Dasycladales,
Siphonocladales), whereas others are predominantlyfreshwater
(Ulotrichales, Coleochaetales) or exclusively freshwater
(Oedogoniales, Zygnematales).The freshwater species have a
cosmopolitan distribution, with few species endemic in a certain
area. In the marineenvironment, the green algae in the warmer
tropical and semitropical waters tend to be similar everywhere in
theworld. This is not true of the Chlorophyta in the colder marine
waters; the waters of the Northern and Southernhemispheres have
markedly different species. The warmer waters near the equator have
acted as a geographical barrierfor the evolution of new species and
genera.
1EMU
Contents
Introduction to green algae 2Morphological diversity . . . . . .
. . . . . . . . . . 2Calcifying chlorophytes . . . . . . . . . . .
. . . . . 3Oil algae chlorophytes . . . . . . . . . . . . . . . .
3Biotechnology . . . . . . . . . . . . . . . . . . . . . 3
Cell structure 3Cell wall . . . . . . . . . . . . . . . . . . .
. . . . . 3Pigmentation . . . . . . . . . . . . . . . . . . . . .
4Eyespot and phototaxis . . . . . . . . . . . . . . . . 4
Phototaxis by the secretion of mucilage . . . . 4
Reproduction 4Asexual reproduction . . . . . . . . . . . . . . .
. . 4Sexual reproduction . . . . . . . . . . . . . . . . . . 5
Where do green algae belong and classification 5Subdivision of
chlorophytes . . . . . . . . . . . . . 5
Streptophyta 5Charophyceae . . . . . . . . . . . . . . . . . . .
. . 5
Thallus . . . . . . . . . . . . . . . . . . . . . 5
Reproduction . . . . . . . . . . . . . . . . . . 7Habtiats . . .
. . . . . . . . . . . . . . . . . . 7
Zygnematophyceae . . . . . . . . . . . . . . . . . .
7Chloroplasts . . . . . . . . . . . . . . . . . . . 7Photomovement
of chloroplasts . . . . . . . . 8Practical use . . . . . . . . . .
. . . . . . . . . 9
Habitats . . . . . . . . . . . . . . . . . . . . . . . .
9Reproduction . . . . . . . . . . . . . . . . . . . . . 9
Chlorophyta 9Prasinophyceae . . . . . . . . . . . . . . . . . .
. . 9
Pyramimonas . . . . . . . . . . . . . . . . . .10Ostreococcus .
. . . . . . . . . . . . . . . . .10
Ulvophyceae . . . . . . . . . . . . . . . . . . . .
.10Ulotrichales . . . . . . . . . . . . . . . . . . .10Ulvales . .
. . . . . . . . . . . . . . . . . . .11Cladophorales . . . . . . .
. . . . . . . . . . .11Dasycladales . . . . . . . . . . . . . . . .
. .11Caulerpales . . . . . . . . . . . . . . . . . .
.11Siphonocladales . . . . . . . . . . . . . . . . .12
Acknowledgments 12
-
Starch
Two membrane envelope
Stacked thylakoids
Figure 1. Scheme of a green algal chloroplast. The
chloroplasthas a two membrane enevelope, thylakoids are stacked,
reservepolysacharide — starch — accumulates within the
chloroplasts.
Introduction to green algaeChlorophytes include a wide diversity
of unicellular flagel-lates (and some complex colonial forms),
non-flagellatedunicells and colonies, filamentous forms, and some
morecomplex macroalgae, including green seaweeds. Unicellu-lar and
filamentous green algae are significant componentsof freshwater
planktonic, and periphytic communities. Verytiny green algae such
as Ostreacoccus, which at less than1 μm in diameter is barely
visible with the light or fluores-cence microscope, are members of
the extremely abundantand productive marine picoplankton.Tropical
nearshore waters are frequently dominated by
green seaweeds having very unusual bodies composed of gi-ant,
multinucleate cells, known as siphonalean forms. Someof these,
notably species of the genus Caulerpa, form veryserious and
extensive nuisance growths in theMediterraneanand other parts of
the world.The best known subgroups include theChlorophyceae
(e.g.,
Chlamydomonas, Volvox),Ulvophyceae
(marinemacroalgae),andTrebouxiophyceae. There are several
additional distinctlineages, mostly of small scaly unicellular
flagellates, thatcollectively are referred to as
‘prasinophytes’.Chlorophytes contain at least one plastid, and most
of the
green algae are considered to be autotrophic. Some of
theprasinophycean green algae feed on particles and
thereforeexhibit phagotrophy and mixotrophy.Features that are
common to nearly all chlorophytes in-
clude:
1. Flagella, commonly occurring in pairs ormultiples of two,that
are of approximately equal length and without tri-partite, tubular
hairs. Fibrillar hairs (Chlamydomonas)and Golgi-produced scales
(Pyramimonas), are present insome genera.
2. Chloroplasts bound by a two-membrane envelope (withno
enclosing periplastidal endoplasmic reticulum).
3. Chlorophylls a + b.
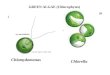
Figure 2. Two unicellular chlorophytes, representing
flagellatedmonad (Clamydomonas, left) and non-flagellated
coccoid(Chlorella, right; Source.).
4. Chloroplast thylakoids occurring singly or in stacks
ofvariable numbers.
5. Production and storage of starch (α-1,4-linked polyglu-cans)
inside the chloroplasts.
Production and storage of the photosynthetic reserve in-side the
plastid is unique to the green algae (Fig. 1). In otheralgae the
photosynthetic storage product, whether starch orsome other
material, is found primarily in the cytoplasm.Plastidal starch of
green algae is reminiscent of cyanophycean
glycogen storage within cyanobacterial cells.The presence and
plastidal location of green algal starch
can be visualised by treating cells with a solution of
I2KI,which stains starch a dark blue-black. Staining for starch
isone of the most helpful ways to distinguish green algae
fromsimilar-appearing forms belonging to other algal
groups.Chloroplasts of green algae may or may not contain eye-
spots and pyrenoids. If eyespots are present in green
algalcells, they are always located inside the chloroplast,
neveroutside it as in euglenoids, some dinoflagellates, and
eustig-matophyceans.Among the green algae, chloroplasts are very
variable in
shape and number per cell, but are typically uniform
withingenera. As a result, chloroplast shape and number are of-ten
useful taxonomic characters, more so than is typical forother
groups of eukaryotic algae.
Morphological diversityMorphological diversity of the green
algae ranges from tinyflagellates to multicellular macroscopic
organisms. I thinkall known cell types and life forms are present
in the greenalgae: unicellular flagellates, non-flagellate unicells
(Fig.2), motile colonies, nonmotile colonies (Fig. 3), coloniesof
regular size and shape known as coenobia (Fig. 4), un-branched
filaments, branched filaments (Fig. 5), tissue likecellular sheets,
and multinucleate coenocytes (Fig. 6).
https://toddcaldecott.com/herbs/chlorella/
-
Figure 3. Two colonial chlorophytes, representing
non-motilecolony (Palmella, left; Source.) and motile colony
(Eudorina,right; ). Palmella colony lives within a gelatinous bag;
the cellsareuniformly arranged at the peripheral matrix; 2-4 cells
form asmall subset. Eudorina colony is ellipsoidal (sometimes
nearlyspherical), 60-200 μm long, consisting of 32 or 16 or 64
cells,each 18-20 μm in diameter.
Figure 4. Coenobial colonies — Pediastrum (left), a
four-cellcoenobium of Scenedesmus (right). Coenobium is a
colonycontaining a fixed number of cells, with little or no
specialization.
Figure 5. Cladophora — a branched filamentous green algae(left)
Source., and Spirogyra— unbranced filaments. Source.
Figure 6. Multicellular green algae — Ulva (left)
andmulti-nucleate single cell coenotcytic macrophyte
—Acetabularia.
Figure 7. Calcifying green algae — Halimeda incrassata fromGulf
of Mexico (left; Source.). Calcium carbonate is deposited inits
tissues, making Halimeda inedible to most herbivores.Oil-algae —
Botryococcus braunii (right; Source.) — a greenalgae that has
hydrocarbons typically around 30–40% of their celldry weight,
making it a potential candidate for biotechnology.
Calcifying chlorophytesA variety of tropical green macroalgae,
in particular Hal-imeda, precipitate calcium carbonate onto their
bodies (Fig.7). When they die, these algae contribute substantially
tothe production of carbonate sand, and over geological timesuch
calcareous algae have generated important carbonatedeposits.
Oil algae chlorophytesThe microscopic green alga Botryococcus
(Fig. 7) producesvery large amounts of lipid and is also the source
of somepetroleums. This alga is a potential modern-day source
ofrenewable energy-rich compounds.
BiotechnologyDunaliella (Figs. 4-11, 20-37) and Haematococcus
(Fig.4-12, 4-13).are widely cultivated for production of
usefulorganic compounds, while Chlorella (Figs. 4-1, 19-2) isgrown
for use as a human food supplement. Selenastrumis a single-celled
green alga that is widely used in bioassaysof water quality (Fig.
4-6, 20-42).
Cell structure
Cell wallCell walls usually have cellulose as the main
structuralpolysaccharide, although xylans or mannans often
replacecellulose in the Caulerpales.The primitive algae in the
Prasinophyceae have extracel-
lular scales, or a wall derived from interlacing scales,
com-posed of acidic polysaccharides.
Volvocales have walls composed of glycoproteins.
3CELL STRUCTURE
http://protist.i.hosei.ac.jp/PDB/Images/Chlorophyta/Palmella/sp_5.htmlhttp://protist.i.hosei.ac.jp/PDB/Images/Chlorophyta/Cladophora/sp_3.htmlhttp://protist.i.hosei.ac.jp/PDB/Images/Chlorophyta/Spirogyra/group_D/sp_04.htmlhttp://cfb.unh.edu/phycokey/Choices/Chlorophyceae/siphonous_greens/Bryopsidales/HALIMEDA/Halimeda_Image_page.htmhttps://alchetron.com/Botryococcus-braunii
-
PigmentationChloroplast pigments are similar to those of higher
plants— chlorophyll a and b are present. The main carotenoid
islutein.However chlorophytes may not always have green col-
oration and therefore are sometimes difficult to recognise
asgreen algae. Widely encountered examples
includeTrentepohlia,which often forms dramatic orange-red growths
on cliff facesand other terrestrial substrates, the flagellate
Haematococ-cus, which colours bird baths and other such structures
red(Fig. 17-1), and Chlamydomonas nivalis, which can coloursnow red
(see Fig. 1-11, Text Box 20-1).InTrentepohlia andChlamydomonas
nivalis the red colour
is due toβ-carotene1, which accumulates between thylakoidsin the
chloroplast. InHaematococcus the red colour is due toastaxanthin,
which accumulates in lipid globules outsidethe chloroplast.
Hematochrome is a general term for these
carotenoids.Accumulation of hematochromes colours the cells orange
orred, with hematochrome accumulating up to 8–12% of thecellular
contents in Dunaliella.Accumulation of carotenoids occurs under
conditions of
nitrogen deficiency, high irradiance (Trentepohlia,
Chlamy-domonas nivalis) or high salinity (Dunaliella). The
largeamounts of carotenoid pigments obscure chlorophylls andserves
a photoprotective function.Animals can not synthesise these
carotenoids and they ac-
quire the pigments through the food chain from primary
pro-ducers. Hematochromes are responsible for the colouring infish,
crustaceans and birds (such as the pink in flamingos).
Eyespot and phototaxisMost of the flagellated cells that show
phototactic move-ment have an eyespot. In chlorophytes, the eyespot
is al-ways in the chloroplast, usually in the anterior portion
nearthe flagella bases. The eyespot consists of lipid
droplets,usually between the chloroplast envelope and the
outermostband of thylakoids. The eyespot is coloured orange-red
fromthe carotenoids in the lipid droplets.The photoreceptor in
Chlamydomonas is in the plasma
membrane above the eyespot and consists of a chromophore(colored
substance) linked to a protein — opsin, that isembedded in the
plasma membrane.The chromophore is 11-cis-retinal (the aldehyde
of
vitamin A). Light excitation causes isomerization of
11-cis-retinal into trans, triggering a conformational change
thatinitiates the signalling process.The chromophore 11-cis-retinal
and the protein opsin pro-
duce a rhodopsin, a general class of compounds that ab-sorb
light maximally around wavelengths of 500 nm.
1The same pigment is responsible for the orange colour of
carrots.
The eyespot filters light by reflecting blue and green lightback
onto the photoreceptor in the plasma membrane as thealga swims
through the medium. This results in changes inmembrane potential
involving rhodopsin. Entry of calciuminto the cell is affected by
the membrane potential of theplasmamembrane, and, in turn, the
concentration of calciumions in the cytoplasm affects the rate of
beating of the flag-ella.The swimming direction of the cell is
affected by the rate
of beating, because at one concentration of calcium ions,each
flagellum beats differently. Therefore, changing the cy-toplasmic
calcium concentration differentially changes thebeat of each
flagellum, causing the cell to swim in a differentdirection.
Phototaxis by the secretion of mucilageA second type of
phototactic movement in the chlorophytesuses secretion ofmucilage
in desmids. Already in1848 desmidmovement on a surface of mud
brought to the lab was de-scribed, and presumed to be due to the
stimulus of light.Penium (a desmid chlorophyte) aligned their long
axis
and moved toward the light, accumulating on the lightedside of
the culture vessel they were growing in. The move-ment is brought
about by the extrusion of slime through cellwall pores in the
apical part of the cell.
ReproductionThe high diversity within chlorophytes also
translates into ahigh diversity of reproductive strategies
Asexual reproductionMost common in unicellular organisms is
simple cell divi-sion.For colonial forms the simplest is
fragmentation of colonies
into two or more parts, each part becoming a new colony.Further,
zoosporogenesis commonly occurs, usually
induced by a change in the environment of the alga. Inthe
chlorophytes, zoospores are normally produced in veg-etative cells,
and only in a few cases are they formed inspecialised
sporangia.Next, aplanospores are non-flagellated and have a
wall
distinct from the parent cell wall. Aplanospores are consid-ered
to be abortive zoospores and have the ability to form anew plant on
germination.Next, autospores are aplanospores that have the
same
shape as the parent cell, and are common in the
Chlorellales(e.g. Chlorella).Next, coenobia are colonies with a
definite number of
cells arranged in a specific manner (e.g., Volvox, Pedias-trum,
Scenedesmus). Generawith colonies arranged in coeno-
4REPRODUCTION
-
Figure 8. Isogamous, anisogamous, and oogamous
sexualreproduction.
bia form daughter colonies with a certain number of cells.
Inmaturation of the daughter coenobia, there is enlargementbut no
division of vegetative cells in the coenobia.
Sexual reproductionSexual reproduction in theChlorophyceaemay be
isogamous,anisogamous, or oogamous, with the general line of
evolu-tion occurring in the same direction (Fig. 8).If the species
is isogamous or anisogamous, the gametes
are usually not formed in specialized cells although in
theoogamous species, gametes are normally formed in special-ized
gametangia (e.g. Coleochaete). Whereas most chloro-phytes form
motile flagellated gametes (zoogametes), inthe Zygnematales
aplanogametes or amoeboid gametesare formed.
Where do green algae belong andclassificationThe green algae and
land plants, collectively known asChloro-bionta or Viridiplantae,
form a monophyletic group withinArchaeplastida2.
Subdivision of chlorophytesThe green algae are divided into two
major clades, the strep-tophytes and chlorophytes sensu stricto
(Fig. 9). The formerincludes land plants, as well as many green
algae. Strepto-phyte green algae are often referred to as
‘charophytes’, andthe best studied groups are the Zygnematophyceae,
whichare unicellular or filamentous freshwater algae, and
theCharo-phyceae, which are truly multicellular freshwater
algae.Despite the similarity in complexity betweenCharophyceae
and land plants, phylogenetic evidence indicates that landplants
are more closely related to Zygnematophyta.
2The Archaeplastida (meaning ‘ancient plastids’; sometimes
alsocalled Plantae) is an eukaryotic supergroup whose
plastids/chloroplastswere acquired directly through a symbiosis
with a cyanobacterium.
StreptophytaThe Streptophyta include land plants (embryophytes)
andtheir closest green algal relatives.The evolutionary
relationship of charophytes to embryophytes
remains unresolved. There is strong support for the hypoth-esis
that conjugating green algae (Zygnematohyceae) con-stitute the
sister group to embryophytes, but it can also
beColeochaetophyceae.
CharophyceaeThe charophytes, or stoneworts, are a group of
large, parenchy-matous green algae with six extant genera in one
family.They are distributedworldwide in freshwater ponds and
lakesand occasionally in brackish water, including the Baltic
Sea.The genus Chara was erected by Vaillant in 1719 for sev-
eral living species of this genus and formally recognised
byLinnaeus (1753) as one of several genera of algae.
Thallus
The charophyte thallus is composed of basal rhizoids, withan
upright main axis consisting of alternating internodes andnodes
(Figs. 10, 11). The rhizoids grow downward, an-choring the thallus
axis in the sediment, and the axes growupward.Charophytes are
relatively large for green algae and can
grow up to a half meter or more in height. Some generaaccumulate
calcium carbonate externally.The charophyte axis has a distinctive
node-internode struc-
ture (Fig. 11).Internodes consist of giant cells, which are
multinucleate,
and with numerous ellipsoidal plastids distributed in the
cy-toplasm surrounding a large central vacuole. The
cytoplasmstreams actively lengthwise around the cell
periphery.Nodes comprise several, smaller, uninucleate cells
that
give rise towhorls of leaflike organs of limited growth
called‘branchlets’, and secondary axes (branches of unlimited
growth),which also exhibit the node-internode construction. A
singleapical meristematic cell occurs on each axis tip.Growth
occurs through division of an apical cell at the tips
of the main axes or secondary branches. A single cuttingface of
the apical cell produces an alternation of internodalcells and
nodal initials.Due to their large size and apparent complexity,
charo-
phytes may be mistaken for bryophytes or certain
aquaticangiosperms (e.g., Ceratophyllum) in the field.The plantlike
structures of charophytes, complex asym-
metric sperm, and large, protected egg cells led earlier
work-ers to see them as intermediate in complexity between
greenalgae and embryophytes.
5STREPTOPHYTA
-
Figure 9. Overview phylogeny of the green lineage. Source:
[?]
Figure 10. Charophyte thallus morphology. (a) Charadrummondii;
(b) Nitella haagenii; (c) Lamprothamniummacropogon; (d) Tolypella
polygyra. Source: [?]
Figure 11. Main external features of Chara. Source.
6STREPTOPHYTA
http://www.biologydiscussion.com/algae/life-cycle-algae/chara-occurrence-structure-and-reproduction-algae/21135
-
Figure 12. General thallus structure of Chara. showing
thelocation of gametangia — oogonium and antheridium. Source.
Reproduction
Asexual reproduction. occurs through growth of erect axesfrom
nodes on the rhizoids, and through contracted starch-filled
branches, and tubercular, starch-filled outgrowths ofthe rhizoids
called bulbils, which may fall away and germi-nate separate from
the thallus.Sexual reproduction. is oogamous. Oogonia and
antheridiaare the female and male gametangia, respectively,
whichinclude gamete-producing cells and associated vegetativecells
(Fig. 12).Each oogonium contains a single large egg cell,
whereas
sperm are produced in filaments with numerous antheridialcells,
packed inside a spherical antheridium (Fig. 12).Oogonia and
antheridia occur on the branchlets at nodes
and may be associated with small sterile cells. The oogoniaare
oblong, 200–1000 μm long by 200–600 μm wide. Maleantheridia are
spherical and range from 200 to 1500 μm indiameter, often bright
orange in colour. Sexual structuresare easily visible with a hand
lens or even with the nakedeye.Sperms have two flagella attached
slightly below the apex
of an asymmetric, helically twisted cell reminiscent of
spermcells in mosses and liverworts.
Habtiats
Charophytes are primarily freshwater organisms, but are
oc-casionally abundant in brackish waters. They occur in quietor
gently flowing waters, from very shallow (several cm) todeep
(>10 m), as long as light levels are adequate. Habitats
are typically alkaline (hard water).Stands of charophytes
provide habitat for invertebrates
and structural refuges for juvenile vertebrates (fish and
frogs).Charophytes are often early colonisers and water
clarifiers.Practical applications for charophytes include
managementofwater quality through encouragement of charophyte
coloni-sation. Nutrients are absorbed by charophytes through
theirrhizoids and photosynthetic thallus, and charophyte
commu-nities can be a significant store of nitrogen in small
waterbodies. Uptake by charophytes removes nutrients from thewater
column that would otherwise be available for growthof other algae.
The decline of charophytes following eu-trophication can be
explained largely by decreases in waterclarity and competition with
angiosperms.
ZygnematophyceaeThe Zygnematophyceae are among the most diverse
greenalgae, with a variety of thallus types (filaments,
unicells,colonies; Fig. 13), and approximately 4,000 described
species.The group lacks flagella at all stages of the life cycle.
Sex-ual reproduction, when present, involves conjugation orthe
union of two haploid vegetative protoplasts (individualcells of
filaments or unicells) to form a zygospore, whichundergoes meiosis
to produce a new haploid thallus upongermination.
Zygnematophyceae contains some of the most beautifulmicroscopic
organisms known (Fig. 15).TheZygnematophyceae is usually divided
into two groups
Desmidiales and Zygnematales.Zygnematales are generally oblong,
rod shaped, or cylin-
drical, and the smooth cell wall lacks pores; the primarywall is
a homogeneous piece, lacking a median constriction.The family
Zygnemataceae (14 genera, over 800 species)included filamentous
algae.TheDesmidiales (41 genera, 3,500+ species) contains the
desmids, which are divided into four families, the
Closte-riaceae, Gonatozygaceae, Peniaceae, and Desmidiaceae,the
latter being the largest of the four families (36 genera,3,000
species, 12,000 subspecific taxa). Most are unicells.Each cell
consists of two mirrorimage parts called semicellsthat are joined
at a narrow midregion or isthmus wherethe nucleus is located.
Chloroplasts and other nonnuclearcell contents are divided equally
between semicells. Thestructure of semicells is often complex, with
two, three, ormore planes of symmetry.
Chloroplasts
Chloroplast shapes range from asteroid (Cylindrocystis
andZygnema) to laminate (Gonatozygon, Mesotaenium,Mougeo-tia) to
ribbon-like (Spirogyra and Spirotaenia, Fig. 16). Anaxile, ridged
chloroplast (stellate) is found in many desmids
7STREPTOPHYTA
https://www.carlsonstockart.com/photo/stonewort-chara-charophyte-green-algae-characeae/
-
Figure 13. Structural diversity in the Zygnematophyceae.
(a)Spirogyra sp.; (b) Zygnema sp.; (c) Spirotaenia condensata;
(d)Roya obtusa var. montana; (e) Netrium digitus; (f)
Gonatozygonaculeatum; (g) Micrasterias rotata; (h) Euastrum
evolutum var.glaziovii; (i) Xanthidium cristatum var. hipparquii.
Structures: c– chloroplast, n – nuclear region at site of isthmus
betweensemicells, p – pyrenoid, v – apical vacuole. Scale bar = 10
μm ineach micrograph
Figure 14. Micrasterias melitensis (left) and Euastrumapiculata
(right) from the marvellous collection by Ernst Haeckel.
Figure 15. Desmidiales: Cosmarium (left) and Closterium(right).
Cosmarium is a very species rich genus. It is a desmid,with two
mirror-image like half cells, joint by an isthmus.Closterium has a
characteristic half-lunar shape.
Figure 16. Sprirogyra:has one of the most conspicuous
ribbonshaped chloroplasts, arranged in a spiral configuration in
theperiphery of the cell.
includingNetrium, Closterium, andPenium. Species of
Desmidi-aceae contain some of the largest and most elaborate
chloro-plasts known among the green algae. Their chloroplasts
areoften ridged, lobed, and highly dissected.Pigments include those
typical of green algae and em-
bryophytes, i.e., the descendants of a common ancestor
thatincludes all green algae and embryophytic plants: chloro-phylls
a and b, β and γ-carotenes, and several xanthophylls.Chloroplasts
usually contain one or more pyrenoids aroundwhich starch is
stored.
Photomovement of chloroplasts
The laminate chloroplast of many taxa, e.g. Mougeotia
andMesotaenium are able to moves within the cell. The chloro-plasts
display maximum surface area or face toward low-intensity light. In
high-intensity light, the chloroplast alignsitself with the edge
profile toward the light. Presumably
8STREPTOPHYTA
-
thesemotions optimise photosynthetic performance andmin-imise
damage to the photosynthetic apparatus.
Practical use
Members of the Zygnematophyta have not been exploitedfor
economic use in any major way. A few species havebeen used in fish
aquaculture. Some studies suggest thatgreen algae in general and
Spirogyra in particular may beuseful for the detection and recovery
of certain metals fromcontaminated waterways. Some conjugating
green algae,including Spirogyra, Mougeotia, and the Desmidiales,
areused as indicators of trophic status and water quality.
HabitatsMostly freshwater. These algae are common in ponds,
ephemeralpools, marshes, and bogs, lakes, and streams. They
readilycolonise artificial habitats, reservoirs, cattle tanks,
roadsideditches, irrigation canals.They habit surfacemats, benthos
aswell as plankton. Most
conjugating green algae are benthic or periphytic and growon
surfaces or occasionally attached to substrates by meansof rhizoids
or mucilage. Rhizoids that attach to substratemay be present in all
of the filamentous Zygnematales (e.g.,Mougeotia, Spirogyra, and
Zygnema).Many, but not all, are found in oligotrophic
tomesotrophic
waters of moderate to low pH. The diversity of habitats
oc-cupied spans a wide range and may be quite specific for
indi-vidual species. Species show distinct preferences for
certainhabitats characterised by water chemistry and
productivity.This makes the group as a good indicators of habitat
typesand water quality.
ReproductionAsexual reproduction is by fragmentation and cell
division.A fundamental feature distinguishing theZygnematophyta
from other chlorophytes is sexual reproduction by
conjugationinvolving the fusion of non-flagellate gametes (Fig.
17).Sexual cycles consist of:
1. conjugation (the physical joining of cells or filamentsand
subsequent union of gametes to form a zygote)
2. formation of a thick-walled zygospore
3. a period of zygospore dormancy
4. and germination of the zygospore to produce
vegetativecells.
Zygnematophyta display zygoticmeiosis—growing cellsare haploid,
andmeiosis occurs in the zygote, the only diploidcell in the sexual
cycle. Strains of speciesmay be homothallic
Figure 17. Sprirogyra:reproduction by conjugation.
— conjugation is intraclonal, or heterothallic — conju-gation is
interclonal between plus and minus mating types.Optimal conditions
for conjugation vary from species to
species. Filamentous Zygnematales often conjugate whenfilaments
are transferred to nutrient-poor conditions.
ChlorophytaThe chlorophyte clade is composed of four algal
classes (Fig.9):
Prasinophyceae — paraphyletic class of scaly naked uni-cellular
flagellates, mostly marine.
Ulvophyceae — predominantly marine, but a number offreshwater
species. All filamentous marine green algaeor larger green seaweeds
belong here.
Chlorophyeae
Trebouxiophyceae
PrasinophyceaeHere belong primarily marine green flagellates
with scalescomposed of acidic polysaccharides. Prasinophytes are
re-garded as the modern representatives of the earliest
greenalgae.Within the group, the flagellar number varies from one
in
Pedinomonas to 16 in Pyramimonas cyrtoptera.Prasinophycean
flagella typically emerge from an apical
depression or pit. The cell membrane of most forms is cov-ered
with one or more layers of often extremely
elaboratescales.Prasinophyceans generally also have a single
plastid (though
it may be highly lobed), and usually possess at least
onestarch-sheathed pyrenoid.The cells of most prasinophyceans are
enclosed by one to
five layers of scales attached to the cell membrane, with
thescales of each layer characteristic for the species.
9CHLOROPHYTA
-
Figure 18. Pyramimonas light microscopy (left) and SEM(right).
Note the coverage of cell and flagella with scales.
Figure 19. EM image of cell scales in Pyramimonas (left).
Notethat different types of scales are in many layers. Scheme
ofPyramimonas flagellum, showing the coverage with flagellascales
(right).
Pyramimonas
Pyramimonas is a flagellate unicell found in marine, brack-ish
(incl. Baltic Sea), or freshwaters. Flagella, mostly 4, butcan be
also 8 or 16, depending on species, emerge from adeep, narrow pit
in the middle of four lobbed cell anterior.The cells are somewhat
heart-shaped (cordate) (Fig. 19).There are several layers of body
and flagellar scales (Fig.19).
Ostreococcus
Ostreococcus is a genus of unicellular coccoid or
sphericallyshaped green algae. It includes prominent members of
theglobal picoplankton community. Ostreococcus tauri has anaverage
size of 0.8 μm in diameter and is the smallest eu-karyotic cell
known (Fig. 20). The alga has a relativelylarge nucleus, a single
chloroplast with a starch granule, amitochondrion, and a Golgi
apparatus.Due to its small size, the genus was discovered as late
as
in 1994.
UlvophyceaeMostlymulticellular thalloidmarine benthic green
algae. Thelife cycle usually involves the alternation of a haploid
thallus
Figure 20. Ostreococcus tauri. Left: The general
organisationSource: [?] . Right: EM image, showing the the nucleus,
themitochondria and the chloroplast. Source.
with a diploid thallus. Here belong the conspicuous coeno-cytic
benthic algae of tropical marine waters.Six orders, first three are
common algae in temperate wa-
ters, others are predominantly tropical:
Ulotrichales — uninculeate filamentous algae with a pari-etal
chloroplast.
Ulvales —uninucleate cells with a parietal chloroplast; thal-lus
is a hollow cylinder or a sheet, one or two cells thick.
Cladophorales — multinucleate filamentous algae with aparietal
perforate or reticulate chloroplast.
Dasycladales — thallus has radial symmetry composed ofan erect
axis bearing branches; thallus uninucleate butmultinucleate just
before reproduction.
Caulerpales — coenocytic algae lacking cellulose in
thewalls.
Siphonocladales — algae with segregative cell division.
UlotrichalesUlothrix (Fig. ) is found in quiet or running
freshwater andoccasionally on wet rocks or soil. The thallus
consists ofunbranched filaments of indefinite length that are
attachedto the substratum by a special basal cell. All of the
cellsexcept the basal one are capable of cell division and
formingzoospores or gametes.Species with narrow cells form 1, 2, or
4 quadriflagellate
zoospores per cell, whereas those with broad cells form 2,4, 8,
16, or 32 zoospores per cell. The zoospores have aconspicuous
eyespot and are liberated through a pore in theside of the parent
wall. Zoospores that are not dischargedfrom the parent may secrete
a wall and become thin-walledaplanospores. These later germinate to
form a new filament.Gametes ofUlothrix are formed in the sameway as
zoospores
but are biflagellate. The gametes are of the same size,
withfusion occurring only between gametes from different
fila-ments. The zygote remains for a while, settles, secretes
athick wall, and undergoes a resting period during which it
10CHLOROPHYTA
https://genome.jgi.doe.gov/OstRCC809_1/OstRCC809_1.home.html
-
Figure 21. Ulothrix zonata. Filaments consist of many
cells;rhizoid formed at one end. Cells 30–60 μm in diam., 15-30
μmlong; Source.
accumulates a large amount of storage material. The
firstdivision of the zygote is meiotic, with the zygote forming 4to
16 zoospores or aplanospores.Inmany northern lakes,Ulothrix zonata
grows abundantly
in early spring in shallow waters along shorelines.
Ulothrixzonata is dominant until thewater temperature reaches
10°C,when it disappears owing to massive conversion of the thal-lus
to zoospores.
Ulvales
Ulvales have a thallus that is either an expanded sheet
one(Monostroma) or two cells (Ulva; Fig. 5.33) thick. Thethallus of
Enteromorpha is a hollow cylinder.Ulva thallus is composed of two
layers of cells, with each
cell having a large cup-shaped chloroplast toward the exte-rior
of the cell (Fig. 5.33). The holdfast is formed by thecells of the
thallus, sending down long slender filamentsthat coalesce to form
the holdfast. The holdfast portion isperennial and proliferates new
blades each spring.Cell division may occur anywhere in the thallus,
but all
divisions are in a plane perpendicular to the thallus
surface.Ulva (Fig. 5.33) has an isomorphic alternation of
gener-
ations, with the gametophyte forming biflagellate gametesand the
sporophyte producing quadriflagellate zoospores.Ulva is normally a
marine genus although it can be found
in brackish waters, particularly in estuaries, and also in
theBaltic Sea. It normally grows on rocks in the intertidal
zone,Ulva is an opportunistic alga, capable of rapid colonisa-
tion and growth when conditions are favourable. This oc-curs
because of a rapid growth rate and the ability to takeup and store
nutrients available in pulsed supply. Becauseof this Ulva has
proliferated in many eutrophied areas. Inenclosed and semienclosed
waters Ulva comprises a largeproportion of drift plants, which may
smother other benthiccommunities or be cast ashore where they
decompose, caus-ing considerable aesthetic nuisance.
Ulva is commonly known as the sea lettuce or green laver,and has
been eaten as a salad or used in soups.
Cladophorales
The filamentous genera in this order havemultinucleate
cells,usually with a parietal or reticulate chloroplast. The
fila-ments may be branched or unbranched.Cladophora (Fig. 5.35(b))
andChaetomorpha (Figs. 5.35(a),
5.36), each with an isomorphic alternation of generations,are
common members.Cladophora. is found in freshwater and marine
habitats. Itmay be the most ubiquitous macroalga in freshwaters
world-wide. This filamentous alga can reach nuisance levels as
aresult of eutrophication.Cladophora is predominantly benthic, and
is often found
in the region of unidirectional flow or in periodic wave
ac-tion. Cladophora is colonised by a wide variety of
epiphytesbecause it offers a substrate that is anchored against
flowdisturbance.
Dasycladales
Here belong tropical and subtropical marine algae, most ofthem
calcified.Due to calcification they fossilise readily.
TheDasycladales
has a paleontological record that extends back to the
Precambrian–Cambrian boundary (ca. 570 million years ago). Of the
175known fossil genera, only 11 are extant. The Dasycladalesare in
fact living fossils3
Acetabularia (Figs. 5.38, 5.40, 5.41), (mermaid’s wine-glass) is
the best known. At maturity Acetabularia has anaked axis with a
single gametangial disc at the apex.Acetabularia is a warm-water
alga found in shallow pro-
tected lagoons and on the borders ofmangrove swamps, grow-ing on
shells, coral fragments, and other algae. The thallusis
calcified.
Caulerpales
Caulerpales contains the coenocytic or siphonaceous greenalgae.
The non-septate thallus resembles a garden hose with-out any cross
walls separating the usually large thallus, ex-cept during
reproduction.Cellulose is usually not a wall component and is
replaced
by a β-1,3 linked xylan or a β-1,4 linked mannan.Caulerpales are
marine algae and occur as sea-weeds in
the warmer oceans.The coenocytic algae in the Dasycladales and
Caulerpales
respond to injury by rapidly forming gel-like wound plugs,
3Living fossils are organisms that include extant clades that
havesurvived for long intervals of geological time at low numerical
diversityand exhibit primitive morphological characteristics that
have undergonelittle evolutionary change.
11CHLOROPHYTA
http://protist.i.hosei.ac.jp/PDB/Images/Chlorophyta/Ulothrix/sp_7.html
-
thereby preventing loss of cytoplasm. The wound plugsare formed
from extruded cytoplasm that forms the plugthrough interaction of
carbohydrates and lectins (Ross et al.,2005). A new cell wall is
formed under the gelatinous plug.
SiphonocladalesThese algae havemulticellular thalli, are wholly
marine, andare usually tropical. The cells are multinucleate, with
retic-ulate chloroplasts.
Acknowledgments
12CHLOROPHYTA
Introduction to green algaeMorphological diversityCalcifying
chlorophytesOil algae chlorophytesBiotechnology
Cell structureCell wallPigmentationEyespot and
phototaxisPhototaxis by the secretion of mucilage
ReproductionAsexual reproductionSexual reproduction
Where do green algae belong and classificationSubdivision of
chlorophytes
StreptophytaCharophyceaeThallusReproductionHabtiats
ZygnematophyceaeChloroplastsPhotomovement of
chloroplastsPractical use
HabitatsReproduction
ChlorophytaPrasinophyceaePyramimonasOstreococcus
UlvophyceaeUlotrichalesUlvalesCladophoralesDasycladalesCaulerpalesSiphonocladales
Acknowledgments