Embed Size (px)
Citation preview

Proc. Nati. Acad. Sci. USAVol. 85, pp. 3797-3801, June 1988Biophysics
Determination of the aggregate size in detergent solution of thelight-harvesting chlorophyll a/b-protein complex fromchloroplast membranes
(membrane protein/sedimentation equilibrium/analytical ultracentrifugation/three-component system)
P. J. G. BUTLER*t AND W. KUHLBRANDTttMedical Research Council Laboratory of Molecular Biology, Hills Road, Cambridge CB2 2QH, England; and tBiophysics Section, The Blackett Laboratoryand the Agricultural and Food Research Council Photosynthesis Research Group, Department of Pure and Applied Biology, Imperial College of Science andTechnology, London SW7 2BZ, England
Communicated by M. F. Perutz, January 25, 1988
ABSTRACT The molecular mass of an oligomeric integralmembrane protein, the light-harvesting chlorophyll a/b-pro-tein complex from the photosynthetic membranes of chloro-plasts, has been determined in detergent solution by analyticalultracentrifugation and measurement of the density incrementat constant chemical potential of all diffusible solutes. Thetechnique used eliminates any problems resulting from deter-gent binding to the protein, is independent of the particulardetergent used (in this case the nonionic n-octyl fi-D-glucopyranoside), and gives the apparent weight-average mo-lecular mass at different protein concentrations, allowingextrapolation to zero concentration. It means that the solutionsof the complex must be brought to dialysis equilibrium with thesolvent detergent solution and also requires a reliable methodfor measuring the protein concentration, for which amino acidanalysis was used. The detergent-solubilized complex was atrimer that dissociated into monomers and dimers at lowprotein concentration. The accurate concentration determina-tions also allowed the molar chlorophyll-to-protein ratio to bemeasured as 15, corresponding to 8 chlorophyll a and 7chlorophyll b molecules.
Integral membrane proteins are released from the lipidbilayers of biological membranes by solubilization withdetergents. In the solubilized state, the hydrophobic, mem-brane-spanning part of the protein is thought to be sur-rounded by a detergent micelle. When mild, nonionic deter-gents, such as Triton X-100 or n-octyl ,B-D-glucopyranosideare used, solubilized membrane proteins frequently maintaintheir native configuration. While such detergent-solubilizedmembrane proteins may be isolated and purified by methodssimilar to those developed for soluble, globular proteins,subsequent physical characterization may be complicatedboth by the mass of bound detergent and also by the possiblepresence of residual lipid in the protein-containing micelles.
In their native, functional state, many membrane proteinsare multisubunit complexes that either may contain a numberof different polypeptides or may be oligomers with multiplecopies of the same subunit. The use of analytical ultracen-trifugation is well-established for the determination of themolecular mass, and hence aggregate size, for soluble pro-teins. For membrane proteins in detergent solution, thisanalysis is more complicated because the solution containsthree components: the buffer, the protein, and the detergent,ofwhich the latter two interact with each other with unknownstoichiometry. These components have different buoyantdensities, which prevents the use of estimated partial specificvolumes to calculate the molecular mass of the protein-de-
tergent complex from sedimentation measurements, let alonethat of the protein component, and various techniques havebeen tried to avoid these problems and to make this calcu-lation. We have applied the analysis of Casassa and Eisen-berg of such three-component systems in sedimentationequilibrium (1) to an integral membrane protein, the light-harvesting chlorophyll a/b-protein complex from chloro-plast membranes (LHC II) solubilized in n-octyl f3-D-gluco-pyranoside. We believe that this analysis has not previouslybeen applied in this situation and find that it allows astraightforward determination of the aggregate size.LHC II is the most abundant membrane protein in chlo-
roplasts. It functions as the major collector of solar energy inplant photosynthesis (2), mediates cation-induced membraneappression (3), and has a role in the regulation of photosyn-thesis (4). The polypeptide binds chlorophyll (Chl) a and Chlb at a molar ratio of 1.15:1 (5) as well as apparently non-stoichiometric amounts of carotenoids (xanthophylls) (6).LHC II is coded for by a nuclear multigene family (7-9). Themature protein contains at least three different polypeptides(5), which have similar amino acid sequences and molecularmasses (10). Two genes encoding the major polypeptide ofpeaLHC II have been sequenced (7, 11), indicating molecularmasses of about 25,000. (In this work we have used an actualvalue of 25,045, calculated from the sequence in ref. 11). LHCII solubilized in nonionic detergents has been crystallized.Two-dimensional (12, 13) and three-dimensional crystals ofLHC 11 (5) have been obtained. The crystal structure ofLHCII, determined by electron microscopy and image analysis,indicated that the complex is a trimer and has 3-fold symmetry(13, 14).To investigate the aggregate size of LHC II in solution we
have measured the molecular mass of the detergent-solubilized complex by analytical ultracentrifugation. Fromthe work of Casassa and Eisenberg (1), the molecular mass ofthe protein component alone, even in a three-componentsystem, can be determined by sedimentation equilibrium inan analytical ultracentrifuge, if the apparent partial specificvolume (Qp') used for the calculation is derived from thedensity increment (Op/ac2),, for the protein at constantchemical potential of all other components. This can beobtained by measurement with a solution at dialysis equilib-rium with the same solvent as used for the sedimentationequilibrium experiment. If the concentration of the macro-molecule (c2) is measured solely as the protein, the mole-cular mass determined will be that of the protein componentof the aggregate alone, allowing the oligomer size of LHC IIto be determined. The total molecular mass can then be
Abbreviations: LHC II, light-harvesting chlorophyll a/b-proteincomplex; Chl, chlorophyll; Mw app, weight-average apparent molec-ular mass.*To whom reprint requests should be addressed.
3797
The publication costs of this article were defrayed in part by page chargepayment. This article must therefore be hereby marked "advertisement"in accordance with 18 U.S.C. §1734 solely to indicate this fact.

3798 Biophysics: Butler and Kuhlbrandt
calculated from this and the ratio of chlorophyll to protein,which can be measured on the sample employed for themolecular mass measurement.We have performed a number of experiments in which we
have taken solutions of LHC II at dialysis equilibrium inn-octyl ,8-D-glucopyranoside (hence inherently also at aggre-gation equilibrium) and measured the density increment, thechlorophyll-to-protein ratio, and the molecular mass for thecomplex. From these measurements we have determined theaggregate size and the molecular mass ofLHC II, and we findthat these are wholly consistent with the crystal structure (13,14). The method described here is both simple and general,and we believe that it could also be applied to othermembrane proteins solubilized with any dialyzable deter-gent.
METHODSIsolation and Purification of LHC II. LHC II was isolated
and purified by the method of Burke et al. (15) as modified byKuhlbrandt et al. (16). Briefly, broken chloroplasts wereisolated from 4-week-old pea seedlings. Destacked chloro-plast membranes were isolated and solubilized for 30 min at20'C by using Triton X-100 at a detergent/Chl ratio of 10:1(wt/wt) and a Chl concentration of 0.5 mg/ml. LHC II wasisolated by sucrose density gradient centrifugation and pu-rified by precipitation as for crystallization (5). The precip-itated, washed complex was resolubilized in 50 mM n-octylf3,D-glucopyranoside (Bachem, Bubendorf, Switzerland) at afinal Chl concentration of 3-5 mg/ml.The complex was further purified by ion-exchange column
chromatography at 20'C using Fractogel TSK DEAE-650 (S)(Merck). The approximate column dimensions were 10 cm x1 cm. The column was equilibrated with buffer A [10 mMsodium/potassium phosphate (pH 7.4) containing 30 mMn-octyl B-D-glucopyranosidel. An amount of the concen-trated LHC II stock in 50 mM n-octyl P-D-glucopyranosidecorresponding to 5 mg of Chl was applied to the column andwashed with 50 ml of buffer A to remove free chlorophyll.The complex was released with buffer B [10 mMsodium/potassium phosphate (pH 7.4) containing 200 mMNaCl and 50 mM n-octyl f-D-glucopyranoside]. Peak frac-tions containing roughly 75% of the Chl were pooled anddiluted with 5 volumes of deionized water. Upon addition ofKCI to a final concentration of 100 mM, the complexaggregated and was collected by centrifugation (25,000 x gfor 5 min). The precipitate was suspended in deionized waterand recentrifuged. Finally, the purified LHC II was dissolvedat a final Chl concentration of 2-3 mg/ml in 35 mM n-octylf3-D-glucopyranoside and dialyzed for several days at 15°Cagainst buffer C [2 mM sodium/potassium phosphate (pH7.4) containing 3 mM NaN3 and 35 mM n-octyl f-D-glucopyranoside].Measurement of Protein and Chi Concentration. The pro-
tein concentration was determined by standard amino acidanalysis (17, 18) of three different LHC II preparations. Thedialyzed stock solution was diluted 1:10 with deionizedwater. To 10-pAl aliquots ofthe diluted stock, 50 gl of6 M HCOcontaining 2 nmol of norleucine as an internal standard wasadded with a few crystals ofphenol to prevent the breakdownof tyrosine and phenylalanine. Duplicate samples were hy-drolyzed in evacuated sealed glass tubes at 110°C for 24, 48,72, 96, and 120 hr and analyzed on a Beckman 121 MBautomatic amino acid analyzer equipped with a Beckman 126data system integrator. Serine and threonine were deter-mined by extrapolation to zero hydrolysis time. Leucine,isoleucine, and valine were determined by extrapolation toinfinite hydrolysis time.The concentration of Chl a and Chl b in the solutions used
for amino acid analysis was measured spectroscopically after
extraction into 80% (vol/vol) acetone as described by Arnon(19). The absorbance of samples in buffer C was recorded at280 nm and 670 nm.
Analytical Ultracentrifugation. The densities of buffer Cand of the concentrated LHC II stock solution, dialyzedagainst buffer C, were measured with a densitometer (modelDMA06; Anton Paar, Graz, Austria) working at 15'C. Thedensity increment (ap/0c2) was calculated from these mea-sured values and used to calculate the apparent partialspecific volume (p') from the equation:
'= [1 - (aP/0C2),. lip0, [1]
where p is the density of the solution, pa is the density of thesolvent, and c2 is the concentration of the protein.Dialyzed LHC II stock was diluted 1:50 to 1:500 with buffer
C. Sedimentation equilibrium measurements were made witha Beckman An-G analytical rotor in a Beckman L8-70ultracentrifuge equipped with a Prep UV scanner interfacedto an Apple III microcomputer, which recorded the opticaldensity at 280 nm at approximately 400 equally spaced pointsalong the cell during each scan. The long column method wasused, and the samples were centrifuged to equilibrium at15,000 rpm and 15°C after overspeeding for about 4 hr at20,000 rpm. Scans were taken at intervals of about 24 hr and,when no difference could be seen between successive scans,the later one was taken to be at equilibrium. The baseline wasdetermined by overspeeding at 40,000 rpm until all thematerial had sedimented away from the meniscus and thenslowing down to 15,000 rpm before taking a further scan.Duplicate measurements were made for each oftwo differentLHC II preparations.Apparent weight average molecular masses (Mwa p) were
determined for successive sets of 11 data points from theequation:
Mw,app = [dln(c2)/dr2]p2RT/W2(1-_p), [2]
where r is the radius at each point, and the concentration isthat of the midpoint of each set of 11 data points. Plots ofMW app and (Mw,app)' against the concentration of the LHCII monomer (c2) were made for each set of data. Thedependence of Mwapp upon concentration departed from thesimple correction for nonideality:
1/Mw,app = 1/M + Bc2, [3]
where B is the second virial coefficient, at both low (<2 ZMmonomer) and high protein concentration. From the absolutevalues, the former appears to be due to dissociation of theLHC II aggregate at low concentration. We think that thedeparture from this equation at higher protein concentrationsis probably due to a greater nonideality in the protein-deter-gent solution than has usually been found with simple proteinsolutions, with higher-order virial coefficients being nonzero.In practice, a good fit to the data above the lowest proteinconcentration was obtained by using a least-squares Cheby-shev algorithm (Numerical Algorithms Group, Oxford, En-gland) to fit a second-order polynomial, with little furtherimprovement with higher polynomials. From this the valuefor Mwapp of the LHC II aggregate could be obtainedextrapolated to zero concentration, where the apparent valueis expected to equal the true molecular mass.
RESULTS AND DISCUSSIONConcentration of Protein and Chl. It follows from Eq. 2 that
the molecular mass determined by sedimentation equilibriumand the measurement of the density increment at dialysisequilibrium is a function of the protein concentration, c2. In
Proc. Nad. Acad. Sci. USA 85 (1988)
Proc. Natl. Acad. Sci. USA 85 (1988) 3799
the absence ofa reliable calibrated method for measuring thisconcentration, we measured the LHC II protein in solutionby amino acid analysis, based on the amino acid sequence ofthe major LHC II polypeptide (7, 10, 11). Although thecomplex contains several different polypeptides, the aminoacid sequences of major and minor components seem to bevery similar (10). Results shown in Table 1 indicate closeagreement between the amino acid composition of the majorpolypeptide and total LHC II.Measuring the Chl concentration of solutions subjected to
amino acid analysis, we found a total of 15.5 molecules ofChla and Chl b, on average, per polypeptide of 25,000 molecularmass (Table 2). The Chl a/Chl b ratio of crystalline LHC II(5) is consistent with 8 Chl a and 7 Chl b per polypeptide.These results are in agreement with the Chl/protein ratioreported by Burke et al. (15) but higher than the values foundby Thornber et al. (20) and by Mullet (21). However, LHC IIis apt to lose some Chl upon treatment with ionic detergentsor extended exposure to nonionic detergents at high deter-gent/Chl ratios, which may account for the difference. Thetotal molecular mass of protein and Chl in monomeric LHCII is 38,500. This figure excludes xanthophylls, which formpart of the complex, and also lipids, which may be present invariable amounts.The molar extinction of monomeric LHC II in 35 mM
n-octyl f3-D-glucopyranoside was 3.0 x 105 (SD = 0.22 x105) at 280 nm. At the absorption maximum of Chl a in thedetergent-solubilized complex in the blue region (670 nm), themolar extinction was 5.46 x 105 (SD = 0.14 x 105).
Analytical Ultracentrifugation. The density increment(ap/aC2)O, required to calculate the apparent partial specificvolume of protein, was measured as the difference in densityof solution and solvent. Two LHC II preparations gaveapparent partial specific volumes of 0.261 ml g1 and 0.312ml-g . These values are low when compared to the truepartial specific volume of simple globular proteins, but in factthey are apparent values for the protein component alone of
Table 1. Amino acid composition (mol %) of LHC IIAmino Amino acid DNA Peptide sequenceacid analysis sequence if different
Ala 9.71 9.44 9.87-10.30*Arg 2.83 3.00Asp 8.89 4.72 4.29Asn 8.913.86Cys 0.00 0.42Gly 11.92 12.44Glu 904 6.01 6.43GlJ 1.71 1.71-2.14*His 1.21 1.29Ile 2.79 3.00Leu 9.34 9.87Lys 4.54 5.15 4.7-5.15*Met 1.06 2.14Phe 5.80 6.01 6.01-6.44*Pro 8.79 6.87Ser 6.19 7.73 6.87-7.30*Thr 3.51 3.43Trp t 2.57Tyr 3.21 3.43 3.00-3.43*Val 7.31 6.87Amino acid composition (mol %) ofLHC II protein determined by
amino acid analysis (average of 30 measurements), from the DNAsequence of the major LHC II polypeptide (11) and from the partialpeptide sequence ofLHC II containing major and minor polypeptides(10).*Uncertainty because of sequence heterogeneity of major and minorpolypeptides (10).tNot determined.
Table 2. Chl/protein molar ratio of purified LHC II
MolarPreparation Protein, mM Chl, mM ratio
1 0.224 3.50 15.622 0.199 3.27 16.433 0.129 1.89 14.65
The protein concentration was measured in terms of the polypep-tide monomer of molecular mass 25,000.
an aggregate that contains also detergent, Chl, carotenoids,and presumably, variable amounts of lipid. A low value of (p'corresponds to a high value of the density increment (ap/ac2),, (see Eq. 1), and since Ac2 was taken for the proteinalone while Ap was measured for the whole complex, this isexactly what would be expected. Variations in the lipidcontent between LHC II preparations have been found evenafter extraction with a harsh detergent (sodium dodecylsulfate; see ref. 22) and will be even greater with the nonionicdetergent used here (as observed for Triton X-100; see ref.23), probably accounting for the relatively large difference inapparent partial specific volumes calculated for the twopreparations. However, as predicted from the analysis ofCasassa and Eisenberg (1), both preparations gave the sameresult for the molecular mass of the protein component whendata measured on a given preparation were consistently usedthroughout each calculation.
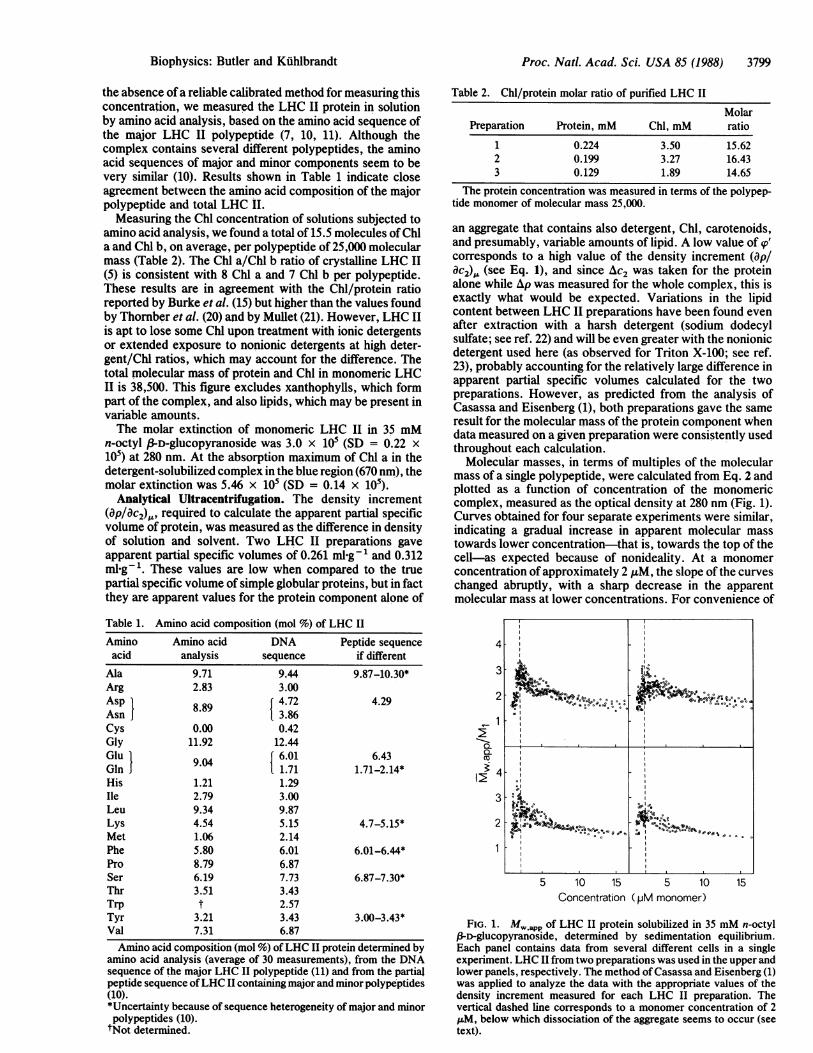
Molecular masses, in terms of multiples of the molecularmass of a single polypeptide, were calculated from Eq. 2 andplotted as a function of concentration of the monomericcomplex, measured as the optical density at 280 nm (Fig. 1).Curves obtained for four separate experiments were similar,indicating a gradual increase in apparent molecular masstowards lower concentration-that is, towards the top of thecell-as expected because of nonideality. At a monomerconcentration of approximately 2 ,uM, the slope of the curveschanged abruptly, with a sharp decrease in the apparentmolecular mass at lower concentrations. For convenience of
4'
2 ,~% toSe0°°°o ;>oSOs O
i4
2
2 -
SI I
5 10 15 5 10 15Concentration (pM monomer)
FIG. 1. MW,app of LHC II protein solubilized in 35 mM n-octyl,3-D-glucopyranoside, determined by sedimentation equilibrium.Each panel contains data from several different cells in a singleexperiment. LHC II from two preparations was used in the upper andlower panels, respectively. The method of Casassa and Eisenberg (1)was applied to analyze the data with the appropriate values of thedensity increment measured for each LHC II preparation. Thevertical dashed line corresponds to a monomer concentration of 2zM, below which dissociation of the aggregate seems to occur (seetext).
Biophysics: Butler and Kiihlbrandt
3800 Biophysics: Butler and Kuhlbrandt
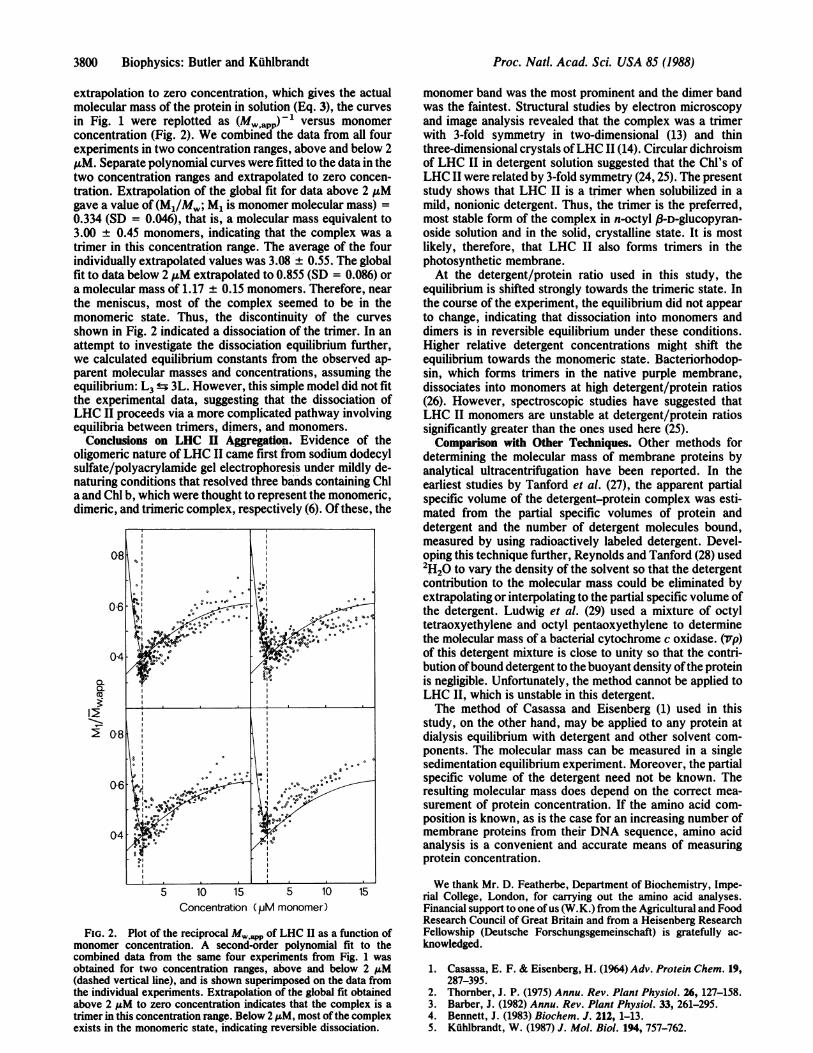
extrapolation to zero concentration, which gives the actualmolecular mass of the protein in solution (Eq. 3), the curvesin Fig. 1 were replotted as (Mw app)-' versus monomerconcentration (Fig. 2). We combined the data from all fourexperiments in two concentration ranges, above and below 2AuM. Separate polynomial curves were fitted to the data in thetwo concentration ranges and extrapolated to zero concen-tration. Extrapolation of the global fit for data above 2 ,.Mgave a value of (M1/Mw; M1 is monomer molecular mass) =0.334 (SD = 0.046), that is, a molecular mass equivalent to3.00 ± 0.45 monomers, indicating that the complex was atrimer in this concentration range. The average of the fourindividually extrapolated values was 3.08 ± 0.55. The globalfit to data below 2 ,uM extrapolated to 0.855 (SD = 0.086) ora molecular mass of 1.17 ± 0.15 monomers. Therefore, nearthe meniscus, most of the complex seemed to be in themonomeric state. Thus, the discontinuity of the curvesshown in Fig. 2 indicated a dissociation of the trimer. In anattempt to investigate the dissociation equilibrium further,we calculated equilibrium constants from the observed ap-parent molecular masses and concentrations, assuming theequilibrium: L3 ± 3L. However, this simple model did not fitthe experimental data, suggesting that the dissociation ofLHC II proceeds via a more complicated pathway involvingequilibria between trimers, dimers, and monomers.
Conclusions on LHC II Aggregation. Evidence of theoligomeric nature of LHC II came first from sodium dodecylsulfate/polyacrylamide gel electrophoresis under mildly de-naturing conditions that resolved three bands containing Chla and Chl b, which were thought to represent the monomeric,dimeric, and trimeric complex, respectively (6). Of these, the
0-6
d2l .
0.4.
lb
5 10 15 5 10 15Concentration (pM monomer)
FIG. 2. Plot of the reciprocal M pp of LHC II as a function ofmonomer concentration. A second-order polynomial fit to thecombined data from the same four experiments from Fig. 1 wasobtained for two concentration ranges, above and below 2 ,uM(dashed vertical line), and is shown superimposed on the data fromthe individual experiments. Extrapolation of the global fit obtainedabove 2 u.M to zero concentration indicates that the complex is atrimer in this concentration range. Below 2 1M, most ofthe complexexists in the monomeric state, indicating reversible dissociation.
monomer band was the most prominent and the dimer bandwas the faintest. Structural studies by electron microscopyand image analysis revealed that the complex was a trimerwith 3-fold symmetry in two-dimensional (13) and thinthree-dimensional crystals ofLHC 11(14). Circular dichroismof LHC II in detergent solution suggested that the Chl's ofLHC II were related by 3-fold symmetry (24,25). The presentstudy shows that LHC II is a trimer when solubilized in amild, nonionic detergent. Thus, the trimer is the preferred,most stable form of the complex in n-octyl P-D-glucopyran-oside solution and in the solid, crystalline state. It is mostlikely, therefore, that LHC II also forms trimers in thephotosynthetic membrane.At the detergent/protein ratio used in this study, the
equilibrium is shifted strongly towards the trimeric state. Inthe course of the experiment, the equilibrium did not appearto change, indicating that dissociation into monomers anddimers is in reversible equilibrium under these conditions.Higher relative detergent concentrations might shift theequilibrium towards the monomeric state. Bacteriorhodop-sin, which forms trimers in the native purple membrane,dissociates into monomers at high detergent/protein ratios(26). However, spectroscopic studies have suggested thatLHC II monomers are unstable at detergent/protein ratiossignificantly greater than the ones used here (25).Comparison with Other Techniques. Other methods for
determining the molecular mass of membrane proteins byanalytical ultracentrifugation have been reported. In theearliest studies by Tanford et al. (27), the apparent partialspecific volume of the detergent-protein complex was esti-mated from the partial specific volumes of protein anddetergent and the number of detergent molecules bound,measured by using radioactively labeled detergent. Devel-oping this technique further, Reynolds and Tanford (28) used2H20 to vary the density of the solvent so that the detergentcontribution to the molecular mass could be eliminated byextrapolating or interpolating to the partial specific volume ofthe detergent. Ludwig et al. (29) used a mixture of octyltetraoxyethylene and octyl pentaoxyethylene to determinethe molecular mass of a bacterial cytochrome c oxidase. (lvp)of this detergent mixture is close to unity so that the contri-bution of bound detergent to the buoyant density of the proteinis negligible. Unfortunately, the method cannot be applied toLHC II, which is unstable in this detergent.The method of Casassa and Eisenberg (1) used in this
study, on the other hand, may be applied to any protein atdialysis equilibrium with detergent and other solvent com-ponents. The molecular mass can be measured in a singlesedimentation equilibrium experiment. Moreover, the partialspecific volume of the detergent need not be known. Theresulting molecular mass does depend on the correct mea-surement of protein concentration. If the amino acid com-position is known, as is the case for an increasing number ofmembrane proteins from their DNA sequence, amino acidanalysis is a convenient and accurate means of measuringprotein concentration.
We thank Mr. D. Featherbe, Department of Biochemistry, Impe-rial College, London, for carrying out the amino acid analyses.Financial support to one ofus (W.K.) from the Agricultural and FoodResearch Council of Great Britain and from a Heisenberg ResearchFellowship (Deutsche Forschungsgemeinschaft) is gratefully ac-knowledged.
1. Casassa, E. F. & Eisenberg, H. (1964) Adv. Protein Chem. 19,287-395.
2. Thornber, J. P. (1975) Annu. Rev. Plant Physiol. 26, 127-158.3. Barber, J. (1982) Annu. Rev. Plant Physiol. 33, 261-295.4. Bennett, J. (1983) Biochem. J. 212, 1-13.5. Kuhlbrandt, W. (1987) J. Mol. Biol. 194, 757-762.
Proc. Natl. Acad. Sci. USA 85 (1988)

Biophysics: Butler and Kuhlbrandt
6. Ryrie, I. J., Anderson, J. M. & Goodchild, D. J. (1980) Eur. J.Biochem. 107, 345-354.
7. Coruzzi, G., Broglie, R., Cashmore, A. & Chua, N. H. (1983)J. Biol. Chem. 258, 1399-1402.
8. Dunsmuir, P., Smith, S. M. & Bedbrook, J. (1983) J. Mol.Appl. Genet. 2, 285-300.
9. Karlin-Neumann, G. A., Kohorn, B. D., Thornber, J. P. &Tobin, E. M. (1985) J. Mol. Appl. Genet. 3, 45-61.
10. Burgi, R., Suter, F. & Zuber, H. (1987) Biochim. Biophys. Acta890, 346-351.
11. Cashmore, A. R. (1984) Proc. NatI. Acad. Sci. USA 81,2960-2964.
12. Li, J. (1985) Proc. Natl. Acad. Sci. USA 82, 386-390.13. Kuhlbrandt, W. (1984) Nature (London) 307, 478-480.14. Kuhlbrandt, W. (1987) J. Mol. Biol., in press.15. Burke, J. J., Ditto, C. L. & Arntzen, C. J. (1978) Arch.
Biochem. Biophys. 187, 252-263.16. Kuhlbrandt, W., Thaler, T. & Wehrli, E. (1983) J. Cell Biol. 96,
1414-1424.17. Spackman, D. H., Stein, W. & Moore, S. (1958) Anal. Chem.
30, 1190-1206.
Proc. Nati. Acad. Sci. USA 85 (1988) 3801
18. Hamilton, P. B. (1963) Anal. Chem. 35, 2055-2064.19. Amnon, D. I. (1949) Plant Physiol. 24, 1-10.20. Thornber, J. P., Alberte, R. S., Hunter, F. S., Shiozawa, J. A.
& Kan, K. S. (1977) Brookhaven Symp. Biol. 28, 132-148.21. Mullet, J. E. (1983) J. Biol. Chem. 258, 9941-9948.22. Thomber, J. P., Steward, J. C., Hatton, M. W. C. & Bailey,
J. L. (1967) Biochemistry 6, 2006-2014.23. Heinz, E. & Siefermann-Harms, D. (1981) FEBS Lett. 124,
105-111.24. van Metter, R. L. (1977) Biochim. Biophys. Acta 462, 642-658.25. Ide, J. P., Klug, D. R., Kuhlbrandt, W., Giorgi, L. B. &
Porter, G. (1987) Biochim. Biophys. Acta 893, 349-364.26. Reynolds, J. A. & Stoeckenius, W. (1977) Proc. Natl. Acad.
Sci. USA 74, 2803-2804.27. Tanford, C., Nozaki, Y., Reynolds, J. A. & Makino, S. (1974)
Biochemistry 13, 2369-2376.28. Reynolds, J. A. & Tanford, C. (1976) Proc. Natl. Acad. Sci.
USA 73, 4467-4470.29. Ludwig, B., Grabo, M., Gregor, I., Lustig, A., Regenass, M.
& Rosenbusch, J. P. (1982) J. Biol. Chem. 257, 5576-5578.





























