Embed Size (px)
Citation preview

THE JOURNAL OF BIOLOGICAL CHEMISTRY Vol. 246, No. 1, Issue of January 10, pp. 152-159, 1971
Printed in U.S.A.
Chitin and Yeast Budding
LOCALIZATION OF CHITIN IN YEAST BUD SCARS
(Received for publication, June 4, 1970)
ENRICO CABIB* AND BLAIR BOWERS
From the National Institute of Arthritis and Metabolic Diseases and the National Heart and Lung Institute, National Institutes of Health, Bethesda, Maryland 20014
SUMMARY
Yeast cells were extracted with alkali and acetic acid according to the method of Bacon et al. (Biochem. J., 101, 36C (1966)). Examination of the extracted cell ghosts under the electron microscope revealed very thin cell envelopes with prominent bud scars in the shape of a shallow crater with a raised rim. The extracted cell ghosts, which had a high chitin content (15%), were treated with purified chi- tinase and glucanase. Electron micrographs showed that chitinase treatment did not destroy the integrity of the cell envelope, but largely eliminated the bud scar rims. On the other hand, after incubation with glucanase only fragments in the shape of bud scars, isolated or in groups, were ob- served. It is concluded that chitin is localized in a ring around the bud scar, sandwiched between two layers of glucan. The possible function of chitin during budding is discussed on the basis of this localization.
During extraction of the yeast cells, a glucan fraction was isolated, which is soluble above pH 10, but rapidly forms a gel upon neutralization. This glucan was used to adsorb and purify the glucanase.
Earlier studies of the budding process in yeast have shown (1) that after separation of the bud from the mother cell, there remains on the surface of the latter a characteristic, crater-like formation, which has been designated as the “bud scar.” 1 The presence of chitin in the bud scars (2) has been confirmed by Bacon et al. (3,4), who isolated from yeast a bud scar preparation containing a high proportion of hexosamine. In the present paper, the precise localization of chitin in the bud scars is studied by electron microscopy, before and after the action of specific enzymes.
EXPERIMENTAL PROCEDURE
Preparation of Chitin-rich Cell Wall Fragments
Saccharomyces carlsbergensis strain 74 (National Collection of Yeast Cultures, England) was grown overnight at 30” in Wicker-
* To whom reprint requests should be sent. 1 An electron micrograph of such a scar, as seen in section, is
shown below (Fig. 5a).
ham’s medium (5) to the stationary phase. After collecting the cells in a Sharples centrifuge, the yeast was washed twice by centrifugation with distilled water. The final pellet was extracted with a modification of the procedure described by Bacon et al. (3), as follows.
Xtep l-For each 10 g of yeast, wet weight, 8 ml of 6% NaOH were added and the mixture was stirred for 1 hour at 75-80”. Added were 30 ml of water, and the suspension was centrifuged for 10 min at 16,000 x g. The pellet was re-extracted at 75-80” three more times, with 16 ml of 3% NaOH each time.
Step !&The final pellet from Step 1 was suspended in 37 ml of water and brought to pH 5 with acetic acid. Glacial acetic acid was added to obtain a final concentration of 1 M. The suspension was heated for 3 hours at 75” with stirring and finally centrifuged at 16,000 x g for 10 min. The pellet was washed once with distilled water.
Step S-The washed pellet was suspended in 15 ml of 3% NaOH and stirred for 100 min at 75”. After centrifuging at 16,000 x g for 10 min, three layers were visible, i.e., a compact pellet, a loose layer of particles, and a clear supernatant fluid. The latter was saved for the preparation of “alkali-soluble glucan” (see below), and the loose layer was discarded. The pellet was submitted again to Steps 2 and 3, sedimented by centrifugation at 39,000 x g for 10 min, and finally washed three times with 6-ml portions of distilled water. The first two washings were turbid and revealed small particles upon exami- nation under the light microscope. The pellet was further washed twice with ethanol and twice with ether, and dried at room temperature.
Starting from 67 g of yeast, wet weight (corresponding to about 13.5 g, dry weight), a yield of 4.8 g of insoluble material, dry weight, was obtained after Step 1. This material is des- ignated “cell ghosts.” The final product after repeating Steps 2 and 3 weighed 115 mg and was labeled “extracted cell ghosts.”
The same treatment was applied to a sample of Saccharomyces cerevisiae strain A364A,2 with similar results.
Isolation of Alkali-soluble Glucan
To the supernatant fluid from Step 3 (see preceding section), 2 volumes of 95% ethanol were added. A flocculent precipitate formed which was recovered by centrifugation for 10 min at 20,000 x g and redissolved in 50 ml of water. Two volumes of 95% ethanol were added and flocculation induced by stepwise
a Strain A364A was kindly provided by Dr. L. H. Hartwell.
152
by guest on September 6, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Issue of January 10, 1971 E. Cabib and B. Bowers 153
addition of 5 M NaCl (several milliliters). After a washing with 66% ethanol, containing 0.1 M ammonium acetate, the pellet was washed twice with 95% ethanol and twice with ether, and dried at room temperature; yield, 540 mg. This material is labeled “crude glucan.”
Before use, 25 mg of crude glucan was suspended in 1 ml of distilled water. A homogeneous suspension was obtained by working the material into a paste with a minimal amount of water prior to final dilution. The resulting suspension was centrifuged for 10 min at 12,000 x g. The pellet was washed twice with l-ml portions of distilled water and finally resuspended in 1 ml of water. The final product is designated as “washed glucan.” The yield of the operation, as calculated from dry weight measurements, was 70%.
PuriJication of Glucanase
All operations were carried out at O-5”, unless specified other- wise. Snail intestinal juice (2 ml) (“glusulase,” Endo Labora- tories, Inc., Garden City, New York) was diluted with an equal volume of 20 mM KC1 and filtered through a Sephadex G-25 column (1.8 x 15 cm), previously equilibrated with 10 m&f KCl, containing 1 mM EDTA. The fraction corresponding to the void volume was discarded, and a volume equivalent to 4 times that of the original, undiluted snail juice was collected. To 1 ml of the filtered extract, 3 ml of 10 mM KCI, containing 1 mM EDTA, were added, followed by 0.8 ml of the washed glucan suspension. The mixture was kept for 10 min at 0” and then centrifuged for 5 min at 20,000 X g. To the supernatant fluid another 0.8 ml of the glucan suspension was added and the process repeated. Each one of the two pellets was washed with 1 ml of distilled water and resuspended in 1 ml of a mixture containing citrate-phosphate buffer (6), pH 6.4, at one-third of the normal strength, 2 mg per ml of bovine serum albumin, and 1 mM 2-mercaptoethanol. The two suspensions were combined and incubated at 30” for 45 min during which period the glucan dissolved almost completely. The solution was dialyzed during 4 hours against two l-liter changes of 10 InM KCl, containing 1 mM 2-mercaptoethanol. A slight remaining turbidity was eliminated by centrifuging for 10 min at 12,000 x g. The recovery of glucanase activity was between 15 and 20%. The glucanase to chitinase ratio was increased about 20.fold over that of the original extract; chitinase was barely detectable in the purified preparations.
Pur&ation of Chhitinase
All operations were carried out at (r5”, unless specified other- wise. Commercial chitinase (25 mg, Worthington) was dis- solved in 10 ml of 0.05 M potassium phosphate buffer, pH 6.3, and 2.5 ml of a colloidal chitin (7) suspension, containing 11.1 mg of solids per ml, was added. After 10 min at 0” the sus- pension was centrifuged for 5 min at 20,000 x g. The pellet was washed once with 5 ml of 0.05 M phosphate buffer, pH 6.3, and resuspended in 5 ml of the same buffer containing 2 mg per ml of bovine serum albumin. The suspension was incubated at 30” for 2 hours, after which only a small amount of insoluble material remained. The suspension was dialyzed for 4 hours against two l-liter changes of 0.05 M phosphate buffer, pH 6.3, and centrifuged for 10 min at 12,000 X g. The recovery of chitinase activity was 50%. No glucanase activity was de- tectable in the purified enzyme. The purified chitinase con-
tained no diacetylchitobiase activity, and the disaccharide of acetylglucosamine was the final product of the reaction.
Enzymatic Determination
For the glucanase assay the incubation mixture contained 10 ~1 of citrate-phosphate buffer, pH 6.4 (6), 0.19 mg of washed glucan, and enzyme, in a total volume of 50 ~1. After 15 min at 30”, the reaction was stopped by addition of the copper re- agent and reducing power was determined (8). A unit of enzyme is defined as that amount which will liberate 1 pmole of glucose per min.
For chitinase determination the incubation mixture contained 0.05 M phosphate buffer, pH 6.3,3.3 mg per ml of colloidal chitin (7), and enzyme, in a total volume of 0.1 ml. After 15 min of incubation at 30’ the reaction was stopped by freezing in acetone- Dry Ice. Each tube was individually treated in the following way. After thawing the incubation mixture, 0.3 ml of 0.05 M
phosphate buffer, pH 6.5, was added; the tube was heated for 1 min in a boiling water bath and rapidly cooled in ice. The contents were immediately filtered through a Millipore mem- brane (pore size 0.45 cl). After this operation was completed with all the samples, a 200+1 aliquot of each filtrate was mixed with 10 ~1 of Sephadex-filtered snail enzyme (see above) and incubated at 30” for 30 min. The tubes were heated for 1 min at loo”, and free acetylglucosamine was determined (9). Since chitinase is not completely destroyed by a brief heating at loo”, the filtration is necessary to separate unreacted substrate from products. The chitinase contained little diacetylchitobiose, even before purification. The snail extract served as a source of the latter enzyme to complete the transformation of the products into free acetylglucosamine. One unit of chitinase is defined as that amount which will cause the liberation of 1 pmole of diacetylchitobiose per min.
Analytical
Total hexoses were determined with anthrone, according to Trevelyan and Harrison (10). Reducing power was measured as described by Hestrin, Feingold, and Schramm (8). Glucose oxidase tests were performed with the Glucostat reagent (Worth- ington). Acetylglucosamine was determined according to Reissig, Strominger, and Leloir (9). Glycogen was measured enzymatically according to Bueding and Hawkins (1 I).
Electron Microscopy
Cell wall residues, after extraction and enzyme digestion, were pelleted and fixed in 3% glutaraldehyde in 0.1 M sodium phosphate buffer, pH 6.8, for 1 hour at room temperature, and postfixed in 1% osmium tetroxide in the same buffer for 1 hour at 4”. Embedment was in Epon (12). Thin sections stained with 1% aqueous uranyl acetate and lead citrate (13) were examined in a Siemens 101 electron microscope at 60 or 80 kv. Shadowed and unstained cell walls were deposited directly on a grid coated with a Formvar substrate film. The material was shadowed with a carbon-platinum pellet at an angle of approxi- mately 25”.
RESULTS
A&a&soluble Glucan-Intact cells of S. carlsbergensis were submitted to essentially the same procedure used by Bacon et al. (3), as outlined under “Experimental Procedure.” As observed by these authors, a large percentage of the yeast glucan
by guest on September 6, 2018
http://ww
w.jbc.org/
Dow
nloaded from

154 Chitin in Yeast Bud Scars Vol. 246, No. 1
TABLE I
Composition of alkali-soluble glucan
Incubation with purified glucanase was carried out as outlined under “Experimental Procedure,” except that limiting amounts of glucan were used. Incubation was carried out for 1 and 2 hours. The longer incubations resulted in only slightly higher values. Glycogen was measured enzymatically (II).
Determination Percentage of dry
weight calculated as glucose
Anthrone on intact polysaccharide. . . . . After glucanase, reducing power.. . . After glucanase, glucose oxidase.. . . Glycogen .
%
85 74 88
1.G
TABLE II
Composition of yeast cell wall residues
Determinations were carried out as described in the legend of Table I and under “Experimental Procedure.” In the determina- tion of chitin the same result was obtained whet,her the incubation was carried out with chitinase and snail intestinal extract added together, or sequentially (chitinase first). In these experiments the conditions were the same as indicated under “Experimental Procedure,” except that limiting amounts of ghosts were used. Incubation times in excess of 1 hour did not increase appreciably the liberation of acetylhexosamine. The fractions of S. carls- bergensis used for eIectron microscopy were those obtained in Experiment 1.
Anthrone on intact ma- teriala .
Reducing power after glucanase*. .
Glucose oxidase after glucanase@. .
Acetylglucosamine af - ter chitinase and di- acetylchitobiase”.
36 1 28
0.8 1 0.82
Cell ghosts Extracted cell ghosts _-
s -
: E
a Expressed as glucose. b Expressed as acetylglucosamine.
% dv wt
45 48 48
43 44 51
54
15 14.9 14.5
dissolved when the alkali extraction was repeated after treatment with acetic acid. The solubilized material was isolated by precipitation with 66% ethanol. The polysaccharide had the unusual property of being soluble at a pH greater than 10, while forming a gel under more acidic conditions. The process was readily reversible. This property was used to advantage for the purification of the glucan, particularly for elimination of the glycogen still present at this stage. The solubility properties of the purified material are similar to those described by Eddy and Woodhead (14) for a glucan obtained by prolonged extrac- tion of yeast with cold alkali. However, it is somewhat puzzling that those authors were able to extract the polysaccharide without previous acid treatment. On the basis of the procedure
employed for its preparation, our material would correspond to the branched p(1 ---f 3)-glucan studied by Bacon et al. (4) and by Manners and Masson (15).
Analytical data on the purified polysaccharide appear in Table I. At least 85% of the dry weight was anthrone-reacting material, calculated as glucose. Treatment with a purified
glucanase (see below) released essentially all of this material as glucose, as judged from reducing power and glucose oxidase determinations. The glycogen content, as measured with specific enzymes, was negligible. After 4 hours of heating in 2 N sulfuric acid at loo”, paper chromatography of the hydrolysate with butanol-pyridine-water (6:4:3) gave rise to a single silver nitrate-reducing spot, with the same Rp as glucose.
Purilfied Chit&use and Glucanase-In order to identify the poly- saccharides present in the various yeast residues, it was necessary to employ enzyme preparations which would attack specifically either glucan or chitin. A purified preparation of chitinase was obtained by adsorption onto its substrate, essentially as outlined by Jeuniaux (16). The availability of the glucan described in the preceding section, and its insolubility in water at neutral pH, made it possible to apply a similar method to the purification of glucanase. The purified chitinase was essentially free from glucanase and vice versa. However, it is likely that these preparations contained other lytic enzymes. In fact, the glucanase preparation caused a slow liberation of reducing power from glycogen (less than 10% of that obtained with glucan as a substrate). It is uncertain at the present time whether this activity is intrinsic to the glucanase or caused by contamination with an amylase. Likewise, the free glucose liberated by
glucanase action (Tables I and II), may be the product of more than one enzyme acting on the different linkages of glucan.
Treatment of Yeast Cell Ghosts with Chit&use and Glucanase- The preparations used for treatment with enzymes were the insoluble residue after the first alkali extractions (cell ghosts) and that remaining after acetic acid and further alkali digestion (extracted cell ghosts). As shown in Table II, the chitin content of cell ghosts, as measured by exhaustive digestion with chitinase, was less than 1% of the dry weight, but was enriched almost 20-fold in the extracted cell ghosts. The total recovery of chitin from the ghosts to the extracted ghosts was about 45%. The amount of acetylglucosamine released from cell ghosts was not increased by using a mixture of crude chitinase plus snail enzyme, which brought about an almost complete dissolution of this material.
The remaining carbohydrate in the extracted cell ghosts appeared to be glucan, as indicated by the approximate equiv- alence between the anthrone-reacting substances and the glucose liberated by incubation with the glucanase preparation (Table I13).
About 35% of the dry weight remains unaccounted for. In this regard it should be mentioned that particles were observed inside the envelopes of the extracted cell ghosts (see Fig. 5~). After treatment with chitinase plus glucanase, a small insoluble residue remained. No defined structures were visible by electron microscopy. Thus, it seems probable that the material not accounted for corresponds to particles entrapped within the cell envelopes and liberated but not solubilized by enzymatic deg- radation of the wall.
s The reason for the relatively high value obtained with glucose oxidase, as compared to that from reducing power determinations, is not known.
by guest on September 6, 2018
http://ww
w.jbc.org/
Dow
nloaded from
Issue of January 10, 1971 E. Cabib mad B. Bowsrs
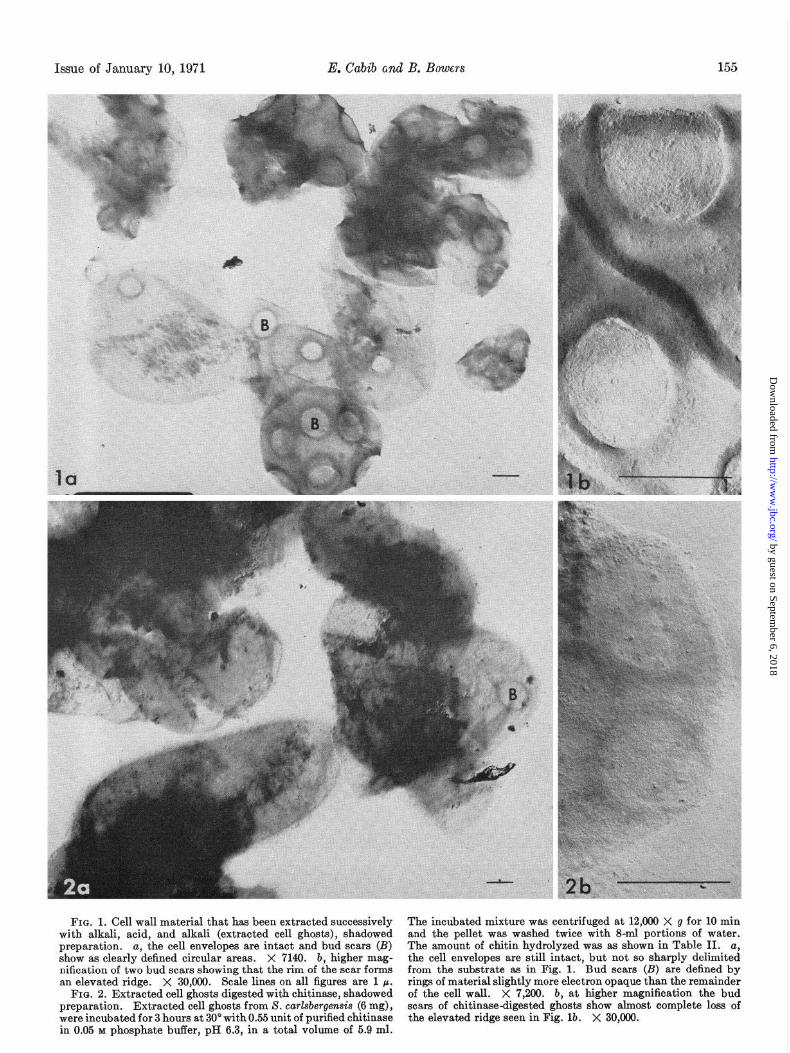
FIG. 1. Cell wall material that has been extracted successively with alkali. acid. and alkali (extracted cell ghosts). shadowed preparation. a, the cell envelopes are intact and bud scars (B) show as clearly defined circular areas. X 7149. b, higher mag- nification of two bud scars showing that the rim of the scar forms an elevated ridge. X 30,090. Scale lines on all figures are 1 p.
FIG. 2. Extracted cell ghosts digested with chitinase, shadowed preparation. Extracted cell ghosts from S. carlsbergensis (6 mg), were incubated for 3 hours at 30” with 0.55 unit of purified chitinase in 0.05 M phosphate buffer, pH 6.3, in a total volume of 5.9 ml.
The incubated mixture was centrifuged at 12,000 X g for 10 min and the pellet was washed twice with S-ml portions of water. The amount of chitin hydrolyzed was as shown in Table II. a, the cell envelopes are still intact, but not so sharply delimited from the substrate as in Fig. 1. Bud scars (B) are defined by rings of material slightly more electron opaque than the remainder of the cell wall. X 7,200. b, at higher magnification the bud scars of chitinase-digested ghosts show almost complete loss of the elevated ridge seen in Fig. lb. X 30,000.
by guest on September 6, 2018
http://ww
w.jbc.org/
Dow
nloaded from
156 Chitin in Yeast Bud Scars Vol. 246, No. 1
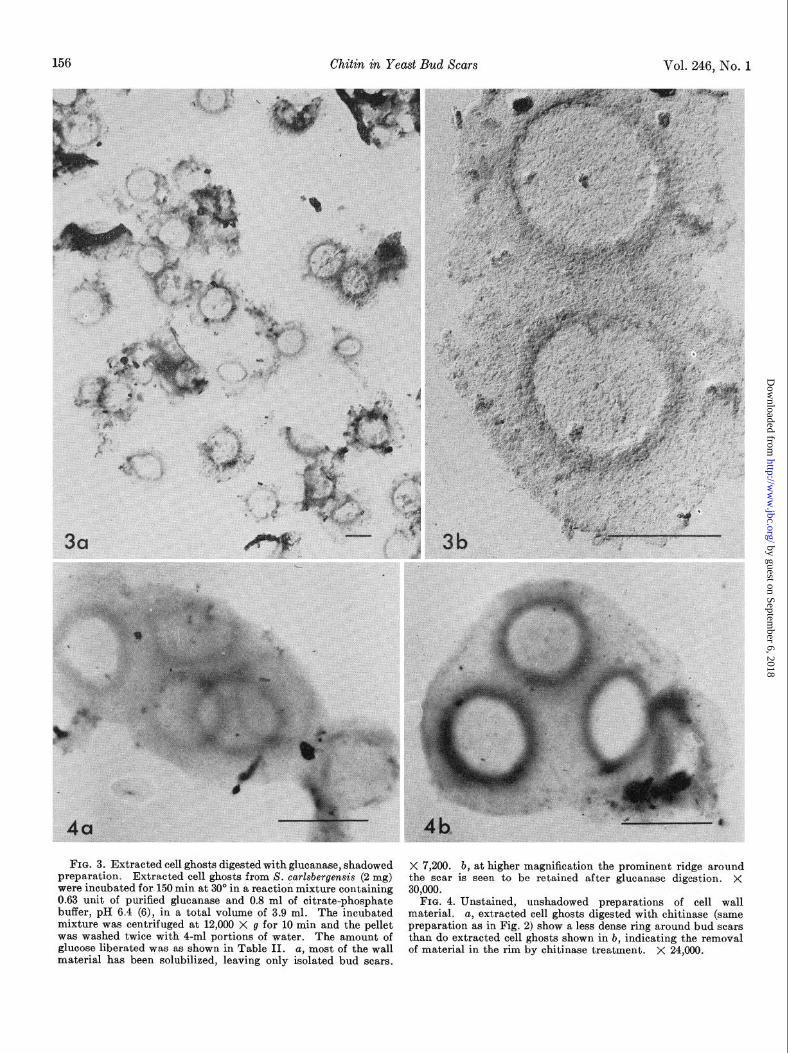
FIG. 3. Extracted cell ghosts digested with glucanase, shadowed X 7,200. b, at higher magnification the prominent ridge around the scar is seen to be retained after glucanase digestion. X preparation. Extracted cell ghosts from S. carkbergensis (2 mg)
were incubated for 150 min at 30’ in a reaction mixture containing 0.63 unit of purified glucanase and 0.8 ml of citrate-phosphate
30,000.
buffer, pH 6.4 (6), in a total volume of 3.9 ml. The incubated FIG. 4. Unstained, unshadowed preparations of cell wall
material. mixture was centrifuged at 12,000 X g for 10 min and the pellet
a, extracted cell ghosts digested with chitinase (same
was washed twice with 4-ml portions of water. preparation as in Fig. 2) show a less dense ring around bud scars
The amount of glucose liberated was as shown in Table II.
than do extracted cell ghosts shown in b, indicating the removal a, most of the wall
material has been solubilized, leaving only isolated bud scars. of material in the rim by chitinase treatment. X 24,000.
by guest on September 6, 2018
http://ww
w.jbc.org/
Dow
nloaded from
Issue of January 10, 1971 E. Cabib and B. Bowers
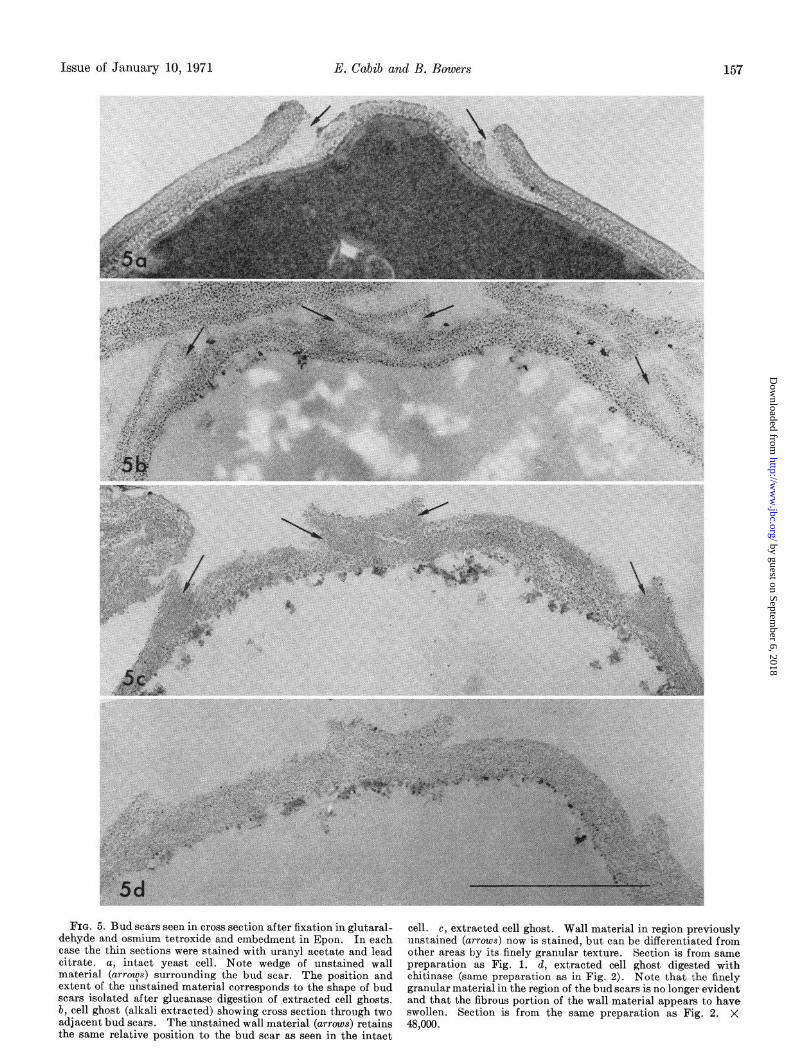
FIG. 5. Bud scars seen in cross section after fixation in glutaral- cell. dehyde and osmium tetroxide and embedment in Epon. In each
c, extracted cell ghost. Wall material in region previously
case the thin sections were stained with uranyl acetate and lead unstained (arrows) now is stained, but can be differentiated from
citrate. other areas by its finely granular texture. Section is from same
a, intact yeast cell. Note wedge of unstained wall preparation as Fig. 1. material (arrys) surrounding the bud scar. The position and
d, extracted cell ghost digested with
extent of the unstained material corresponds to the shape of bud chitinase (same preparation as in Fig. 2). Note that the finely
scars isolated after glucanase digestion of extracted cell ghosts. granular material in the region of the bud scars is no longer evident
b, cell ghost (alkali extracted) showing cross section through two and that the fibrous portion of the wall material appears to have swollen.
adjacent bud scars. The unstained wall material (arrows) retains Section is from the same preparation as Fig. 2. X
the same relative position to the bud scar as seen in the intact 48,000.
by guest on September 6, 2018
http://ww
w.jbc.org/
Dow
nloaded from

158 Chitin in Yeast Bud Scars Vol. 246, No. 1
The analytical data for preparations of cell ghosts and ex- tracted cell ghosts from X. cerevisiae strain A364A were identical with those obtained with S. carlsbergensis.
Electron Microscopy of Cell Ghost Preparations Before and After Enzymatic Treatments--At variance with the results of Bacon et al. (3), our extracted cell ghosts did not consist mainly of isolated bud scars. Rather, complete cell envelopes, albeit very thin and collapsed, predominated (see Fig. 14). The craters of the bud scars were quite prominent, as seen in Fig. lb. Upon treatment with chitinase, the intactness of the cell en- velopes was not affected (see Fig. 2a), but the thick ridge of the bud scars disappeared almost completely, as observed in Fig. 2, a and b. Conversely, after glucanase action most of the cell envelope disappeared, and only fragments in the shape of bud scars, isolated or in small groups, remained (Fig. 3~). In these scars the ridge was still prominent, as can be seen in Fig. 3b. In the photographs of Fig. 4, a and b, which were taken without shadowing, it can be appreciated that the electron density of the annular area around the scar decreased strikingly after chitinase treatment, thus confirming that material had been removed from that region.
Fig. 5 depicts the appearance of sections of the bud scar region, before and after the various treatments. It is proposed that the electron-transparent material present in the borders of the scar (Fig. 5a) is preserved in Fig. 5, b and c, at the sites indicated with arrows. The difference in aspect between the different photo- graphs may be ascribed to variations in staining of this material, which takes up the stain poorly. Most of the external layer of the cell walls, presumably composed of mannan-protein (17), is lost in the first alkali extractions, i.e. between the stages shown in Fig. 5, a and b. At the same time the glucan swells, which explains why no apparent reduction in total wall thickness is seen. From electron micrographs of the cell ghosts (not shown) it appears that the swollen cell wall has often taken up a large proportion of the intracellular space. The swelling is much more pronounced in regions away from the bud scars. Most of this glucan is later extracted by alkali after the preliminary treatment with acetic acid. The difference between intact and chitinase-treated extracted cell ghosts (Fig. 5, c and d) is not as striking as that seen in Figs. 1 and 2, since the crater ridge is still apparent in the section shown. Nevertheless, it can be observed that the apparently amorphous material, present in Fig. 5, a to c (arrows), has disappeared in Fig. 5d, and that only the somewhat fibrous glucan remained. The disappearance of the chitin “filling” would allow the scar to flatten out when dried on a grid, thus imparting to it the aspect shown in Fig. 2b.5
DISCUSSION
In the cell wall of filamentous fungi, chitin appears to be distributed more or less uniformly over the entire wall structure (18). On the other hand, the chitin of budding yeast is localized at the site of the bud scar, as reported by others (2,3) and shown in more detail in the present paper. Bacon et al. (4) have
4 In other preparations the cell wall was more extensively broken down and fragments, containing several bud scars, were more abundant. Cells of S. cerevisiae strain A364A gave rise to wall residues very similar to those of S. carlsbergensis, both before and after enzymatic attack.
5 Glucanase-treated material (shown to consist only of bud scars in shadowed preparations, Fig. 3) stains so poorly that it was not possible to resolve clearly the structures in thin sections.
reported difficulties and lack of reproducibility in trying to ascertain the localization of chitin with specific enzymes. The explanation for these difficulties may be that they used the preparation herein termed cell ghosts. The large amount of glucan and other substances contained in this material probably masked the effect of chitinase.
Our results indicate that chitin is concentrated in a ring which surrounds the bud scar. Although the dimensions of the ring are difficult to determine, it may be concluded from Figs. 3 and 5 that the polysaccharide is mainly accumulated in the crater ridge and also extends for some distance into the surrounding wall. It is probable that the “plug” in the center of the bud scar, which is missing in some of the cell ghosts, also contains a certain amount of chitin (cf. Fig. 3b). The precise location of chitin at the bud scar site suggests that this polymer is involved in the budding process. The recent study of yeast budding by electron microscopy carried out by Marehant and Smith (19) may well be relevant to this problem. These authors showed that from the incipient stage of bud formation an electron- transparent material accumulates around the neck or channel which connects parent and daughter cell. In later stages, the material appears to occupy increasingly more of the periplasmic space, thus squeezing the sides of the cytoplasmic membrane together. Finally, a septum is formed, the bud scar plug, and most of the electron-transparent material is seen in an annular space around the septum. Comparing the final location of the material described by Marchant and Smith (19) (see also Fig. 5~) with that of chitin, it appears probable that we are dealing with the same substance. If this is so, one may perhaps assume that the role of chitin in budding is to provide a rigid ring to protect the channel between mother and daughter cell during the passage of the nucleus and other materials into the bud. The subsequent increased production of chitin would assist in constricting the channel to the point wherein the membranes are pinched off and the cytoplasms of the two cells become separated. The next step would be the formation of the septa, mainly con- stituted by glucan (and perhaps mannan), interspersed with some of the previously existing chitin. After separation between mother and daughter cell, this region constitutes the central plug of the scar.
Another possibility, which does not exclude that mentioned above, is that the chitin ring prevents growth of the new wall backwards, i.e. into the mother cell.
AcknowledgmentsWe are greatly indebted to Dr. L. W. Labaw for taking several electron micrographs, including those shown in Fig. 4. We also wish to express our gratitude to Doctors F. A. Keller and L. B. Rothman-Denes for useful dis- cussions and to Doctors G. Ashwell and W. B. Jakoby for a critical reading of the manuscript. Dr. L. B. Rothman-Denes also carried out some determinations of glycogen.
REFERENCES
1. BERAN, K., in A. H. ROSE AND J. F. WILKINSON (Editors), Advances in microbial physiology, Vol. 2, Academic Press, New York, 1968, p. 143.
2. HOUWINK, A. L., AND KREGER, D. R., Antonie Van Leeuwen- hoek J. Microbial. Serol.. 19. 1 (1953).
3. BACON, J. S. D., DAVIDSON, g. D.‘, JONES, D., AND TAYLOR, I. F.; Biochem. J., 101, 36C (1966).
4. BACON. J. S. D.. FARMER. V. C.. JONES. D.. AND TAYLOR. I. F., Biochem. j., 114, 55j (1969j. ’ ’
5. WICKERHAM, L. J., U. S. Dep. Agr. Tech. Bull., 1029 (1951).
by guest on September 6, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Issue of January 10, 1971 E. Cabib and B. Bowers 159
6. GOMORI, G., in S. P. COLOWICE AND N. 0. KAPLAN (Editors), Methods in enzymology, Vol. 1, Academic Press, New York, 1955, p. 138.
7. BERGER, L. R., AND REYNOLDS, D. M., Biochim. Biophys. Acta, 29, 522 (1958).
8. HESTRIN, S., FEINGOLD, D. S., AND SCHRAMM, M., in S. P. COLOWICK AND N. 0. KAPLAN (Editors), Methods in enzy- mology, Vol. 1, Academic Press, New York, 1955, p. 231.
9. REISSIG, J. L., STROMINGER, J. L., AND LELOIR, L. F., J. Biol. Chem.; 217,959 (1955).
10. TREVELYAN. W. E.. AND HARRISON. J. S.. Biochem. J.. 60. 298 (1952).
’ , .
11. BUEDING, E., AND HAWKINS, J., Anal. Biochem., 7, 26 (1964). 12. LUFT, J. H., J. Biophys. Biochem. Cytol., 9, 409 (1961).
13. REYNOLDS, E. S., J. Cell Biol., 17,208 (1963). 14. EDDY. A. A., AND WOODHEAD, J. S., Fed. Eur. Biochem. Sot.
Let& 1, 67 (1968). 15. MANNERS. D. J.. AND MASSON. A. J.. Fed. Eur. Biochem. Sot.
Lett., 4,‘122 (l.969). ’ ’ 16. JEUNIAUX, C., in E. F. NEUFELD AND V. GINSBURG (Editors),
Methods in enzymology, VoZ. 8, Academic Press, New York, 1966, p. 644.
17. MCLELLAN, W. L., JR., MCDANIEL, L. E., AND LAMPEN, J. O., J. Bacterial., 102, 261 (1970).
18. MAHADEVAN, P. R., AND TATUM, E. L., J. Cell Biol., 36, 295 (1967).
19. MARCHANT, R., AND SMITE, D. G., J. Gen. Microbial., 63, 163 (1968).
by guest on September 6, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Enrico Cabib and Blair BowersSCARS
Chitin and Yeast Budding: LOCALIZATION OF CHITIN IN YEAST BUD
1971, 246:152-159.J. Biol. Chem.
http://www.jbc.org/content/246/1/152Access the most updated version of this article at
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
http://www.jbc.org/content/246/1/152.full.html#ref-list-1
This article cites 0 references, 0 of which can be accessed free at
by guest on September 6, 2018
http://ww
w.jbc.org/
Dow
nloaded from





























