Embed Size (px)
Citation preview

THE JOURNAL OF BIOLOGICAL CHEMISTRY Vol. 243, No. 21, Issue of November 10, pp. 5761-5769,196s
Printed in U.S.A.
Characterization of Two Species of Methionine Transfer
Ribonucleic Acid from Bakers’ Yeast
(Received for publication, June 24, 1968)
KEIICHI TAKEISHI AND TYUNOSIN UKITA
From the Faculty of Pharmaceutical Sciences, University of Tokyo, Tokyo, Japan
SUSUMU NISHIMURA
From the Virology Division, National Cancer Center Research Institute, Chuo-lcu, Tokyo, Japan
SUMMARY
The biochemical properties of two species of methionine transfer ribonucleic acid from bakers’ yeast, separated by diethylaminoethyl Sephadex column chromatography, have been studied. Each species of the two methionine tRNAs, designated methionine tRNA I and methionine tRNA II, respectively, has been purified by benzoylated DEAE- cellulose column chromatography.
Methionine tRNA I can be esterified with methionine by Escherichio coli aminoacyl-tRNA synthetase as efficiently as by yeast aminoacyl-tRNA synthetase, whereas methionine tRNA II can be charged only partially by the E. coli enzyme under the same conditions used in the charging of methionine tRNA I. Methionine tRNA I has been shown to be con- verted to N-formyhnethionyl-tRNA I by E. coli methionyl- tRNA transformylase in the presence of PO-formyltetra- hydrofolate, while methionine tRNA1Iis-not,- In-ribosomal binding studies, 14C-methionyl-tRNA I is recognized by the condons AUG, GUG, and UUG, while 14C-methionyl-tRNA II responds principally to AUG. In addition, methionine has been found to be incorporated from both methionyl- tRNAs I and II into polypeptide as directed by messenger RNA obtained from bacteriophage f2 in a cell-free system from E. coli. Our present report also provides evidence that yeast N-formyhnethionyl-tRNA I serves as an initiator of protein synthesis programmed with the natural messenger in an E. coli system in vitro.
group from NlO-formyltetrahydrofolate, while the other (methi- onyl-tRNA& was not (2, 3). The specificity of these two methionyl-tRNAs in codon recognition has also been established by ribosomal binding and amino acid incorporation studies (24). Furthermore, N-formylmethionyl-tRNAF has been shown to serve as an initiator in polypeptide synthesis by a number of experiments utilizing an E. coli cell-free system, in which viral RNA (5-9) or synthetic polynucleotides (2, 4, 9-12) were used as messengers. It has also been confirmed that the same initiator participates in the protein synthesis directed by each cistron of a polycistronic messenger (13, 14). Recently, it has been shown, with natural messenger, that this is also the case with a system in vitro from Euglena gracilis (15).
Little is known, however, about yeast methionine tRNA. It was previously shown that the methionine-accepting species of yeast tRNA was heterogeneous, since only a part of yeast methionine tRNA could be charged with E. coli aminoacyl-tRNA synthetase (16). However, further isolation and charac- terization of these tRNAs were not carried out. No definitive demonstration of methionyl-tRNA transformylase and of the involvement of N-formylmethionine in the chain initiation of protein synthesis in yeast has been encountered yet, although Marcker and Sanger (1) suggested the existence of N-formyl- methionyl-tRNA in tRNA isolated from yeast grown in a medium containing V-labeled sulfate.
5761
The discovery of N-formylmethionyl transfer ribonucleic acid in Escherichia coli by Marcker and Sanger (1) prompted the characterization of methionine tRNA from E. coli and studies on the mechanism of the initiation of protein biosynthesis. Countercurrent distribution of E. coli tRNA resulted in separa- tion of at least two methionine-accepting species. It was shown that one (methionyl-tRNAF) was capable of accepting a formyl
We previously reported that fractionation of yeast tRNA by DEAE-Sephadex column chromatography resulted in separation of at least two methionine-specific tRNAs, methionine tRNAs I wx!.~I~(17),. I~~.be.~~eselLt,.pi.ae~,.~ko.t~e.-~tb~~~~~~ccep~iag
species from yeast tRNA were investigated with respect to charging with methionine by E. coli aminoacyl-tRNA synthetase, the formylation of the methionyl-tRNA by E. coli trans- formylase, and the stimulation of binding to ribosome of the methionyl-tRNA in the presence of trinucleotides. We also tested whether or not yeast N-formylmethionyl-tRNA I, which was found to be formylated by the E. coli enzyme, serves as an initiator of protein synthesis programmed by the natural messen- ger RNA from bacteriophage f2 in the E. coli system in vitro. This paper also deals with the transfer of methionine from
by guest on February 9, 2020http://w
ww
.jbc.org/D
ownloaded from

Two Methionine tRNA Species from Yeast Vol. 243, No. 21
methionyl-tRNA I and II into polypeptide in the same E. coli system, as mentioned above.
During the course of this investigation, we learned that similar studies on yeast methionine tRNA have been undertaken by RajBhandary and Ghosh.’
EXPERIMENTAL PROCEDURE
Xuterials-Uniformly 14C-labeled n-methionine (specific activ- ity, 187 mC per mmole) and 14C-sodium formate (specific activity, 42.8 mC per mmole) were purchased from New England Nuclear and The Radiochemical Centre, respectively. Poly (II,, G) was a commercial product of Miles Chemical Company. GpUpG and GpUpA were enzymatically prepared with the use of RNase Tl from guanosine 2’,3’-cyclic phosphate and the dinucleoside monophosphates UpG and UpA, respectively (18). ApUpG and UpUpG were generously provided by Dr. Y. Inoue of Tohoku University, Sendai. Benzoylated DEAE-cellulose was prepared by complete benzoylation of DEAE-cellulose according to the method of Gillam et al. (19). Folic acid was obtained from Daiichi Pure Chemicals Company, Ltd. dl-l’etrahydrofo- late was prepared by a modification of the method of O’Dell et al. (20) as described by Hat& et al. (21).
Preparation of N’O-Formyltetrahydrofolic Acid--14C-Labeled N5~1’J-methenyltetrahydrofolate was prepared from X-sodium formate and dl-tetrahydrofolate with crystalline formyltetra- hydrofolate synthetase from Clostridium acidiurici (specific activity, 32,500 units per mg, a kind gift of Dr. J. C. Rabinowitz of the University of California, Berkeley) as described by Rabino- witz and Pricer (22)) with a modification in the concentrations of dl-tetrahydrofolate and formate. Under the conditions de- scribed below, about 70 y0 of the r4C-sodium formate added to the reaction mixture was converted to the I%-formyl group of 14C- formyltetrahydrofolate. The reaction mixture contained 0.6 ml of 1 M triethanolamine HCl (pH 8.0), 0.6 ml of 0.05 M ATP, 0.6 ml of 0.1 M MgC12,O.g ml of 0.1 M dl-tetrahydrofolate, 0.31 ml of 0.012 M r4C-sodium formate, 0.89 ml of 1 M mercaptoethanol, and 0.6 ml of the enzyme solution (80 units per ml). The reaction mixture was incubated at 37” for 2 min without the enzyme; the enzyme was then added, and the incubation was continued for 30 min. To stop the reaction, 1.5 ml of 1.5 N
HCT were added. 14C-Methenyltetrahydrofolate was purified by paper chromatography (the sample, placed on Whatman No. 3MM paper, was developed by a descending technique with the system 1 M formic acid-O.01 M mercaptoethanol). The ultra- violet absorption spectrum of 14C-methenyltetrahydrofolate obtained was practically identical with that described in the literature (23). The preparation thus obtained was dissolved in 0.01 N HCl-0.01 M mercaptoethanol and stored at -20”. Unlabeled methenyltetrahydrofolate was also prepared by the procedure described above, under the reaction conditions de- scribed by Rabinowitz and Pricer (22). NlO-Formyltetrahydro- folate could be generated in situ by adjusting solutions of the methenyl derivative to pH 7 or higher.
Preparation of tRNA and Phage RNA-Unfractionated tRNAs were prepared from E. coli strains B and S26RIE carrying sup- pressor gene Su-1, by the method of Zubay (24). The tRNA from the latter strain was a gift from Dr. H. Ishikura of the Tokyo Medical and Dental University. Methionine tRNAF, a formylatable species with a purity of 90% or better, was pre-
1 U. L. RajBhandary and H. P. Ghosh, unpublished data.
pared from E. coli strain B by successive application of DEAE- Sephadex column chromatography (25) and reversed phase column chromatography (26). Details will be published later. The method for preparation of unfractionated tRNA from bakers’ yeast (Saccharomyces cerevisiae) was described in a previous paper (17). Bacteriophage f2 SUS~,~ a mutant of f2 with a nonsense mutation in the gene specifying the coat protein, was prepared by infecting E. coli S26RIE in a medium containing polypeptone (1%)) NaCl (0.8 yO), glucose (0.1%)) yeast extract (0.1%)) a nd CaClz (0.0227$, the pH of which was adjusted to 7.2 with NaOH. The phage was purified by a pro- cedure including ammonium sulfate fractionation and banding in a CsCl density gradient, as described by Loeb and Zinder (27). RNA was prepared by shaking suspensions of the purified phage with phenol. RNA concentration was determined by assuming 24 Azeo units3 per mg (28).
Fractionation of Yeast tRNA--DEAE-Sephadex A-50 column chromatography of the unfractionated tRNA from bakers’ yeast was carried out according to the method reported previously (17). After the tRNA solution (103,000 A260 units in 600 ml of the init,ial buffer) had been applied to the column (6 x 150 cm), elution was performed by a linear gradient of both NaCl (0.375 to 0.525 M) and MgClz (0.008 to 0.016 M) in the presence of 0.02 M Tris-HCl (pH 7.5). (Further details are described in Refer- ence 17.) Each methionine tRNA fraction separated by DEAE- Sephadex column chromatography was further purified by chromatography on beneoylated DEAE-cellulose according to the procedure of Gillam et al. (19). The methionine tRNA in the various fractions was precipitated by addition of 2.5 volumes of ethanol directly to the effluent and storage at -20” overnight. The tRNA was isolated by centrifugation, dissolved in deionized water, and stored at -20”.
Preparation of Cell Extracts-The S-30 fraction (30,000 x g supernatant of cell extracts) was prepared from E. coli S26RIE harvested in the logarithmic phase and stored in liquid nitrogen after dialysis against the standard buffer according to the method of Nirenberg and Matthaei (29). It was not incubated before use. The method described by Nishimura et al. (30) was used for preparation of the ribosomes from E. coli B. Crude yeast. aminoacyl-tRNA synthetase was prepared from frozen cells harvested in the logarithmic phase as described previously (17j. Transformylase activity could not be detected under the condi- tions described below for the enzyme preparation thus obtained. Crude E. coli aminoacyl-tRNA synthetase was prepared by the procedure described previously (25). It was also used as the enzyme preparation for the transformylation reaction (31).
Assay for Amino Acid and Formate Acceptor Activity of tRNA- The assay procedure detailed in the previous paper (17) was used to determine the amino acid acceptor activity of tRNA. The reaction mixture consisted of 10 pmoles of sodium cacodylate (pH 7.0), 1 pmole of magnesium acetate, 1 pmole of KCl, 0.2 pmole of ATP, 1.5 to 3.0 mpmoles of 14C-amino acid (specific activity, 10 mC per mmole), 10 to 50 ~1 of effluent or 0 to 2 A2e0 units of tRNA preparation, and 50 to 150 pg of crude aminoacyl- tRNA synthetase from yeast or E. coli in a total volume of 0.1 ml. Incubation was performed at 37” for 10 min.
The reaction mixture for the assay of transformylation of
z The phage was kindly donated from Dr. T. Andoh of the Institute of Medical Science, University of Tokyo.
3 A?,, unit refers-to absorbance per ml as measured in a l-cm light path at 260 rnp and at neutral pH.
by guest on February 9, 2020http://w
ww
.jbc.org/D
ownloaded from

Issue of November 10, 1968 K. Takeishi, T. Ukita, and X. Nishimura 5763
methionyl-tRNA contained 10 pmoles of sodium cacodylate (pH 7.2), 1 pmole of magnesium acetate, 1 pmole of KCl, 0.2 pmole of ATP, 5 mpmoles of l*C-n-methionine, 0.4 mpmole of ~4C-forn~yltetrahydrofolate (specific activity, 42.8 mC per mmole), 10 to 50 ~1 of effluent or 0 to 2 A260 units of tRNA preparation, and 100 to 200 pg of crude aminoacyl-tRNA synthe- tase from E. coli or yeast in a total volume of 0.1 ml. Incubation was carried out at 37” for 10 min. The radioactivity incorpo- rated was counted in a liquid scintillation spectrometer by the paper disc method described previously (25).
Preparation of Y-Methionyl-tRN.4 and 14C-Formylmethionyl- tRNrl-14C-Methionyl-tRNA was prepared from yeast as described by Nishimura and Novelli (32), with a slight modifi- cation. The react,ion mixture for charging tRNA with 14C- methionine contained the same components described in the preceding paragraph, except for the use of 14C-methionine with a specific activity of 187 mC per mmole. After 10 min of incu- bation at 37”, 25 ~1 of 2 M sodium acetate buffer (pH 4.35) and 0.5 ml of phenol saturated with deionized water were added successively to 0.5 ml of the reaction mixture, followed by vigorous shaking for 5 min at 4”. The phenol layer isolated after centrifugation was subjected to back-extraction with 0.2 ml of Buffer A, which contained 0.05 M sodium acetate (pH 5.0) and 0.005 M magnesium acetate, in the manner described above. The combined supernatant fluid was dialyzed against Buffer A in a cold room for 15 hours.
XJ-Formylmethionyl-tRNA was prepared as follows. The incubation mixture consisted of the same components described in the preceding section. The enzyme preparation from E. coli was used. The incubation mixture was allowed to react at 37” for 10 min, followed by the same procedures used in the prepara- tion of 14C-methionyl-tRNA, except that the extraction of the reaction mixture (5.0 ml) with phenol was performed twice and the combined phenol layer was back-extracted with 5 ml of Buffer A. After dialysis against Buffer A, 0.1 volume of 2 M
sodium aeetate {pH 5.0) and 2 volumesof .&ha& -wereadded to the inner solution, and the precipit,ate formed was collected, washed once with a mixture of 2 volumes of ethanol and 1 volume of Buffer A, and dissolved in 0.5 ml of Buffer A. The amount of methionine and formate bound to each tRNA preparation is shown in Table I.
Assay of Yeast Methionyl-fRNA Binding to E. coli Ribosomes- The assay was carried out by the procedure of Nirenberg and Leder (33). The assay conditions used were: Condition A, the incubation mixture (0.05 ml) contained 0.1 M Tris-HCl (pH 7.5), 0.05 M KCl, 0.01 M magnesium acetate, 1.0 Azeo unit of ribosomes, I%-methionyl-tRNA and poly (UI,G) as indicated in Table II, with incubation at 37” for 8 min; Condition B, the incubation mixture (0.05 ml) contained 0.1 M Tris-HCl (pH 7.5), 0.05 M KCl, 0.02 M magnesium acetate, 1.5 ASGO units of ribosomes, 14C-methionvl-tRNA, and trinucleotides as specified in Fig. 5, with incubation at 25” for 15 min.
Assay for Amino Acid Incorporation into Protein-The incuba- tion mixture for assay of 14C-methionine incorporation from 1%.methionvl-tRNA into f2 protein contained the following components in a final volume of O.Imi: 5-pmoies of ‘Tris-HCi (pH 7.8), 2.4 Mmole of KCl, 0.6 pmole of mercaptoethanol, 0.3 Fmole of ATP, 0.01 pmole of GTP, 2.4 pmoles of phosphoenol- pyruvate, 5 pg of pyruvate kinase, 1.6 pmoles of magnesium acetate, 20 mpmoles of each of 20 unlabeled amino acids, 0.5 AzeO unit of E. coli S26RIE tRNA, 10 ~1 of an S-30 fraction from
TABLE I Properties of methionine tRNA fractions used in present study
tRNA fraction
Unfractionated tRNA
Methionine-tRNA I (tubes 541-580 in Fig. 1).
Methionine-tRNA II (tubes 661-750 in Fig. 1). .
Methionine-tRNA I (tubes 38-45 in Fig. 3). . .
Methionine-tRNA II (tubes loo-103 in Fig. 4).
Methionine.acceptor
-
Assay Aa
,.qmo1es/Amo unit
2g Met I, 19 i Met II, 10
88
51
455
168 !-
4.6 72
5.1
24
17
1
493
0
Formate acceptor activity
4.2
0.07
29
0
a The yeast enzyme, the E. coli enzyme, and the E. coli enzyme in the presence of the yeast enzyme were used in Assays A, B, and C, respectively.
b Calculated from formate acceptor activity, which was ob- tained by Assay B.
c Calculation of these values was based on the fact that an unfractionated methionine tRNA consisted of 65% methionine tRNA I (Met I), and the residual 35y0, of methionine tRNA II (Met II) (see the text).
E. coli S26RIE, I%-methionyl-tRNA as indicated, 6 mpmoles of unlabeled N’O-formyltetrahydrofolate, and 187 pg of f2 sus3 RNA, where indicated. The last three components were added after incubation of the other components at 37” for 7 min and readjustment of the pH to 7.8~ withmKOH, if necessary. After the additions, the incubation was continued at 37”. At the time designated, 1.5 ml of 5% trichloracetic acid were added to the incubation mixture, which then was heated at 90’ for 15 min. The acid-insoluble material was collected on a glass fiber filter (Whatman GF-83) and washed with about 20 ml of 5 y0 trichlor- acetic acid. After drying under an infrared lamp, the filter was counted for radioactivity in a liquid scintillation spectrometer.
Incorporation of 14C-formyl groups from 14C-formylmethionyl- tRNA into f2 proteinwas assayed as described for 14C-methionine incorporation, except for the addition of 100 mpmoles of un- labeled sodium formate per 0.1 ml (final volume) to the prelimi- nary incubation mixture, the omission of unlabeled N1O-formyl- tetrahydrofolate, and the substit,ution of 14C-formylmethionyl- tRNA for I%-methionyl-tRNA. Incubation was also performed at 37”. At the time specified, the reaction mixture (0.1 ml) was diluted with 5 ml of water and incubated at 37” for 15 min with 100 pg of pancreatic ribonuclease (Worthington). Then cold 100% trichloracetic acid was added to the chilled incubation mixture to make a final concentration of 5%, and the acid- insoluble radioactivity was determined as for 14C-methionine incorporation.
RESULTS
Separation and Properties of Two Methionine tRNA Species from Yeast-Bakers’ yeast tRNA was fractionated by DEAE-
by guest on February 9, 2020http://w
ww
.jbc.org/D
ownloaded from
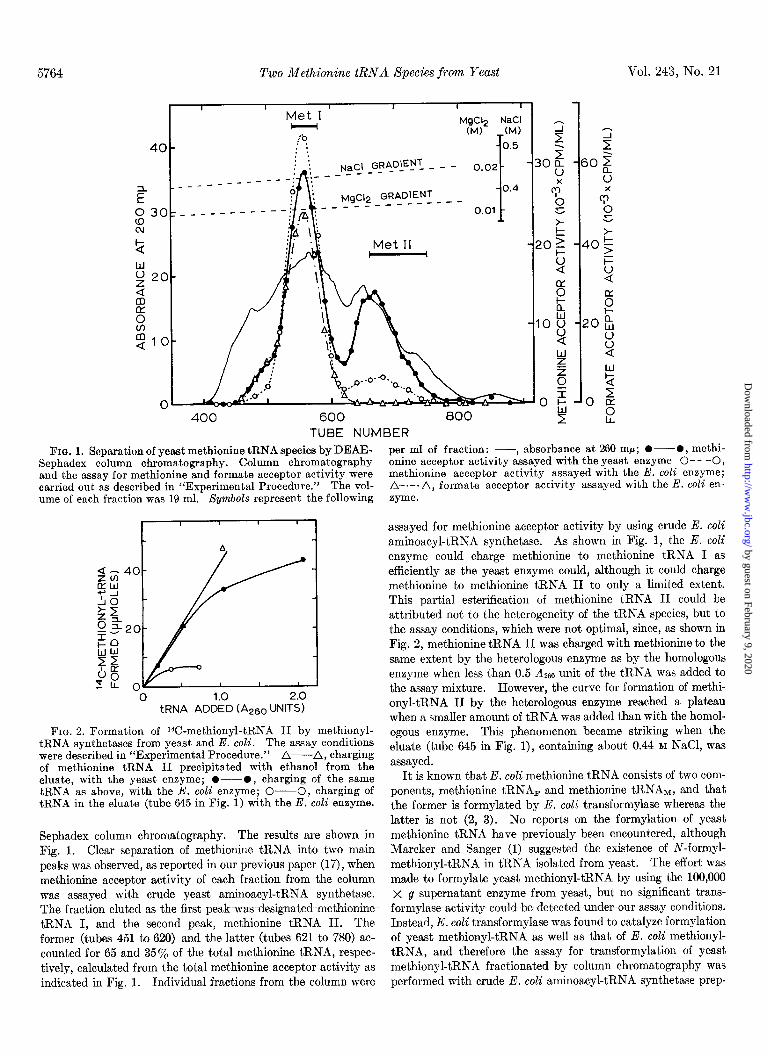
5764 Two Methionine tRNA Species from Yeast Vol. 243, No. 21
I I I I I I
Met I w 3
:‘o MCI2 “7;;
(Ml
40- : :
: : 0.02
400 600 800 5 TUBE NUMBER
FIG. 1. Separation of yeast methionine tRNA species by DEAE- Sephadex column chromatography. Column chromatography and the assay for methionine and formate acceptor activity were carried out as described in “Experimental Procedure.” The vol- ume of each fraction was 19 ml. Symbols represent the following
0 1.0 tRNA ADDED (AzeO “NIT%
FIG. 2. Formation of 14C-methionyl-tRNA II by methionyl- tRNA synthetases from yeast and E. coli. The assay conditions were described in “Experimental Procedure.” A-A, charging of methionine tRNA II precipitated with ethanol from the eluate, with the yeast enzyme; 0-0, charging of the same tRNA as above, with the E. coli enzyme; O-0, charging of tRNA in the eluate (tube 645 in Fig. 1) with the E. coli enzyme.
Sephadex column chromatography. The results are shown in Fig. 1. Clear separation of methionine tRNA into two main peaks was observed, as reported in our previous paper (17)) when methionine acceptor activity of each fraction from the column was assayed with crude yeast aminoacyl-tRNA synthetase. The fraction eluted as the first peak-was -designated methionine tRNA I, and the second peak, methionine tRNA II. The former (tubes 451 to 620) and the latter (tubes 621 to 780) ac- counted for 65 and 35% of the total methionine tRNA, respec- tively, calculated from the total methionine acceptor activity as indicated in Fig. 1. Individual fractions from the column were
per ml of fraction: --, absorbance at 260 rnp; O---O, methi- onine acceptor activity assayed with the yeast enzyme 0- - -0, methionine acceptor activity assayed with the E. coli enzyme; A-.-.A, formate acceptor activity assayed with the E. coli en- zyme.
assayed for methionine acceptor activity by using crude E. coli aminoacyl-tRNA synthetase. As shown in Fig. 1, the E. coli enzyme could charge methionine to methionine tRNA I as efficiently as the yeast enzyme could, although it could charge methionine to methionine tRNA II to only a limited extent. This partial esterification of methionine tRNA II could be attributed not to the heterogeneity of the tRNA species, but to the assay conditions, which were not optimal, since, as shown in Fig. 2, methionine tRNA II was charged with methionine to the same extent by the heterologous enzyme as by the homologous enzyme when less than 0.5 A260 unit of the tRNA was added to the assay mixture. However, the curve for formation of methi- onyl-tRNA II by the heterologous enzyme reached a plateau when a smaller amount of tRNA was added than with the homol- ogous enzyme. This phenomenon became striking when the eluate (tube 645 in Fig. l), containing about 0.44 M NaCl, was assayed.
It is known that E. coli methionine tRNA consists of two com- ponents, methionine tRNAF and methionine tRNAM, and that the former is formylated by E. toll transformylase whereas the latter is not (2, 3). No reports on the formylation of yeast methionine tRNA have previously been encountered, although Marcker and Sanger (1) suggested the existence of N-formyl- methionyl-tRNA in tRNA isolated from yeast. The effort was made to formylate yeast methionyl-tRNA by using the 100,000 x g supernatant enzyme from yeast, but no significant trans- formylase activity could be detected under mr assay conditions. Instead, E. coli transformylase was found to catalyze formylation of yeast methionyl-tRNA as well as that of E. coli methionyl- tRNA, and therefore the assay for transformylation of yeast methionyl-tRNA fractionated by column chromatography was performed with crude E. coli aminoacyl-tRNA synthetase prep-
by guest on February 9, 2020http://w
ww
.jbc.org/D
ownloaded from
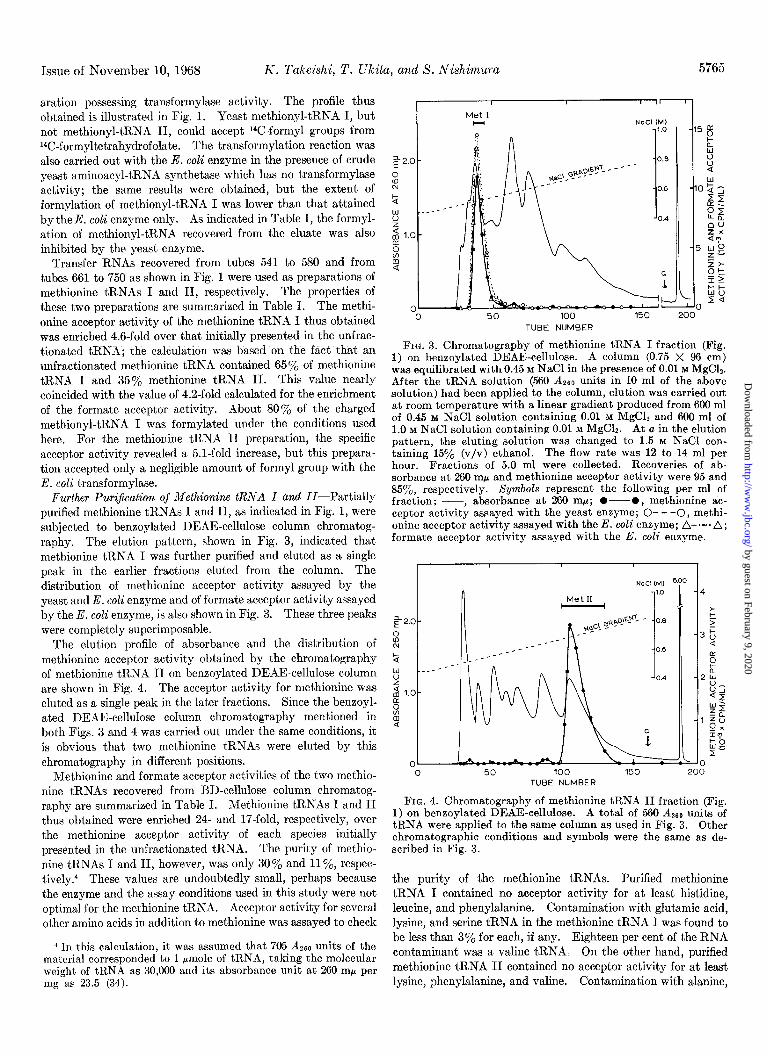
Issue of November 10, 1968 K. l’alceishi, T. Ulcita, and S. Nishimura 5765
aration possessing transformylase activity. The profile thus obtained is illustrated in Fig. 1. Yeast methionyl-tRNA I, but not methionyl-tRNA II, could accept 14C-formyl groups from 14Cformyltetrahydrofolate. The transformylation reaction was also carried out with the E. coli enzyme in the presence of crude yeast aminoacyl-tRNA synthetase which has no transformylase activity; the same results were obtained, but the extent of formylation of methionyl-tRNA I was lower than that attained by the E. coli enzyme only. As indicated in Table I, the formyl- ation of methionyl-tRNA recovered from the eluate was also inhibited by the yeast enzyme.
Transfer RNAs recovered from tubes 541 to 580 and from tubes 661 to 750 as shown in Fig. 1 were used as preparations of methionine tRNAs I and II, respectively. The properties of these two preparations are summarized in Table I. The methi- onine acceptor activity of the methionine tRNA I thus obtained was enriched 4.6-fold over that initially presented in the unfrac- tionated tRNA; the caicuiation was based on the fact that an unfractionated methionine tRNA contained 65% of methionine tRNA I and 35% methionine tRNA II. This value nearly coincided with the value of 4.2-fold calculated for the enrichment of the formate acceptor activity. About 80% of the charged methionyl-tRNA I was formylated under the conditions used here. For the methionine tRNA II preparation, the specific acceptor activity revealed a 5.1-fold increase, but this prepara- tion accepted only a negligible amount of formyl group with the E. coli transformylase.
Further Puri$cation of Methionine tRNA I c~nd II-Partially purified methionine tRNAs I and II, as indicated in Fig. 1, were subjected to benzoylated DEAE-cellulose column chromatog- raphy. The elution pattern, shown in Fig. 3, indicated that methionine tRNA I was further purified and eluted as a single peak in the earlier fractions eluted from the column. The distribution of methionine acceptor activity assayed by the yeast and E. co& enzyme and of formate acceptor activity assayed by the E. coli enzyme, is also shown in Fig. 3. These three peaks were completely superimposable.
The elution profile of absorbance and the distribution of methionine acceptor activity obtained by the chromatography of methionine tRNA II on benzoylated DEAE-cellulose column are shown in Fig. 4. The acceptor activity for methionine was eluted as a single peak in the later fractions. Since the benzoyl- ated DEAE-cellulose column chromatography mentioned in both Figs. 3 and 4 was carried out under the same conditions, it is obvious that two methionine tRNAs were eluted by this chromatography in different positions.
Methionine and formate acceptor activities of the two methio- nine tRNAs recovered from BD-cellulose column chromatog- raphy are summarized in Table I. Methionine tRNAs I and II thus obtained were enriched 24- and 17-fold, respectively, over the methionine acceptor activity of each species initially presented in the unfractionated tRNA. The purity of methio- nine tRNAs I and II, however, was only 30% and 11 ‘%, respec- tively.4 These values are undoubtedly small, perhaps because the enzyme and the assay conditions used in this study were not optimal for the methionine tRNA. Acceptor activity for several other amino acids in addition to methionine was assayed to check
4 In this calculation, it was assumed that 705 AUK, units of the material corresponded to 1 pmole of tRNA, taking the molecular weight, of tRNA as 30,000 and its absorbance unit at 260 rnF per mg as 23.5 (34).
TUBE NUMBER
FIG. 3. Chromatography of methionine tRNA I fraction (Fig. 1) on benzoylated DEAE-cellulose. A column (0.75 X 96 cm) was equilibrated with 0.45 M NaCl in the presence of 0.01 M MgC12. After the tRNA solution (560 A 260 units in 10 ml of the above solution) had been applied to the column, elution was carried out at room temperature with a linear gradient produced from 600 ml of 0.45 M NaCl solution containing 0.01 M MgCls and 600 ml of 1.0 M NaCl solution containing 0.01 M MgCh. At a in the elution pattern, the eluting solution was changed to 1.5 M NaCl con- taining 15y0 (v/v) ethanol. The flow rate was 12 to 14 ml per hour. Fractions of 5.0 ml were collected. Recoveries of ab- sorbance at 260 rnp and methionine acceptor activity were 95 and 85oj,, respectively. Symbols represent the following per ml of fraction: p, absorbance at 260 rnp; O---O, methionine ric- ceptor activity assayed with the yeast enzyme; 0- - -0, methi- onine acceptor activity assayed with the E. coli enzyme; A-.-. A; formate acceptor activity assayed with the E. coli enzyme.
I
50 100 150 200 TUBE NUMBER
FIG. 4. Chromatography of methionine tRNA II fraction (Fig. 1) on benzoylated DEAE-cellulose. A total of 560 A260 units of tRNA were applied to the same column as used in Fig. 3. Other chromatographic conditions and symbols were the same as de- scribed in Fig. 3.
the purity of the methionine tRNAs. Purified methionine tRNA I contained no acceptor activity for at least histidine, leucine, and phenylalanine. Contamination with glutamic acid, lysine, and serine tRNA in the methionine tRNA I was found to be less than 3% for each, if any. Eighteen per cent of the RNA contaminant was a valine tRNA. On the other hand, purified methionine tRNA II contained no acceptor activity for at least lysine, phenylalanine, and valine. Contamination with alanine,
by guest on February 9, 2020http://w
ww
.jbc.org/D
ownloaded from
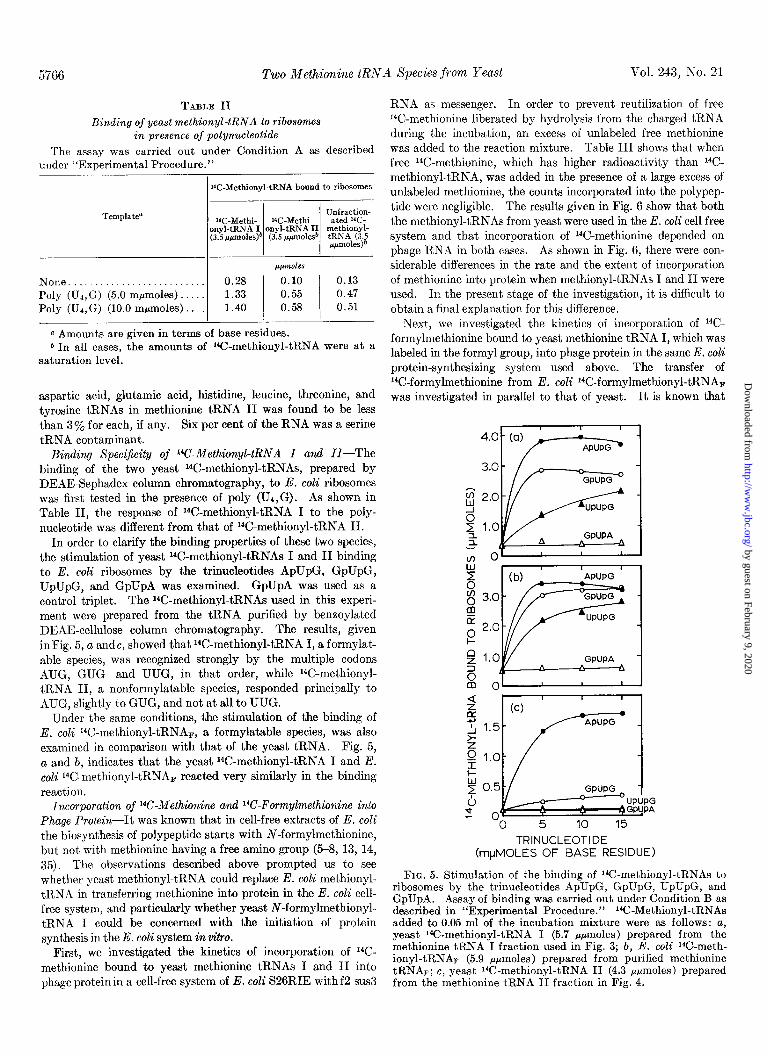
5766 Two Methionine tRNA Species from Yeast Vol. 243, No. 21
TABLE II Binding of yeast methionyl-tRNA to ribosomes
in presence of polynucleolide
The assay was carried out under Condition A &s described under “Experimental Procedure.”
W-Methionyl-tRNA bound to ribosomes
Templatea Unfraction- IGMethi- l4C-Methi- ated W-
onyl-tRNA I onyl-tRNA II methionyl- (3.5p~0les)~ (3.5 p,moksb tRNA (3.5
,iqnoks)b -
~pmzes
None.......................... 0.28 0.10 Poly (U,, G) (5.0 mrmoles) 1.33 Poly (U,, G) (10.0 mpmoles) . 1.40
I I
0.13 0.55 0.47 0.58 0.51
a Amounts are given in terms of base residues. b In all cases, the amounts of W-methionyl-tRNA were at a
saturation level.
aspartic acid, glutamic acid, histidine, leucine, threonine, and tyrosine tRNAs in methionine tRNA II was found to be less than 3% for each, if any. Six per cent of the RNA was a serine tRNA contaminant.
Binding Specificity of 14C-Methionyl-tRNA I and II-The binding of the two yeast K-methionyl-tRNAs, prepared by DEAE-Sephadex column chromatography, to E. coli ribosomes was first tested in the presence of poly (U,,G). As shown in Table II, the response of W-methionyl-tRNA I to the poly- nucleotide was different from that of 14C-methionyl-tRNA II.
In order to clarify the binding properties of these two species, the stimulation of yeast 14C-methionyl-tRNAs I and II binding to E. coli ribosomes by the trinucleotides ApUpG, GpUpG, UpUpG, and GpUpA was examined. GpUpA was used as a control triplet. The W-methionyl-tRNAs used in this experi- ment were prepared from the tRNA purified by beneoylated DEAE-cellulose column chromatography. The results, given inFig. 5, a andc, showed that “C-methionyl-tRNA I, a formylat- able species, was recognized strongly by the multiple codons AUG, GUG and UUG, in that order, while I%-methionyl- tRNA II, a nonformylatable species, responded principally to AUG, slightly to GUG, and not at all to UUG.
Under the same conditions, the stimulation of the binding of E. coli W-methionyl-tRNAF, a formylatable species, was also examined in comparison with that of the yeast tRNA. Fig. 5, a and b, indicates that the yeast 14C-methionyl-tRNA I and E. coli ‘4C-methionyl-tRNAF reacted very similarly in the binding reaction.
Incorporation of 14C-Methionine and 14C-Fomnylmethionine into Phage Protein-It was known that in cell-free extracts of E. coli the biosynthesis of polypeptide starts with N-formylmethionine, but not with methionine having a free amino group (5-8, 13, 14, 35). The observations described above prompted us to see whether yeast methionyl-tRNA could replace E. coli methionyl- tRNA in transferring methionine into protein in the E. coli cell- free system, and parttiularly whetheryxast N-formylmethionyl- tRNA I could be concerned with the initiation of protein synthesis in the E. coli system in vitro.
First, we investigated the kinetics of incorporation of 14C- methionine bound to yeast methionine tRNAs I and II into phage protein in a cell-free system of E. coli S26RIE with f2 sus3
RNA as messenger. In order to prevent reutilization of free 14C-methionine liberated by hydrolysis from the charged tRNA during the incubation, an excess of unlabeled free methionine was added to the reaction mixture. Table III shows that when free W-methionine, which has higher radioactivity than 14C- methionyl-tRNA, was added in the presence of a large excess of unlabeled methionine, the counts incorporated into the polypep- tide were negligible. The results given in Fig. 6 show that both t.he methionyl-tRNAs from yeast were used in the E. coli cell-free system and that incorporation of 14C-methionine depended on phage RNA in both cases. As shown in Fig. 6, there were con- siderable differences in the rate and the extent of incorporation of methionine into protein when methionyl-tRNAs I and II were used. In the present stage of the investigation, it is difficult to obtain a final explanation for this difference.
Next, we investigated the kinetics of incorporation of 14C- formylmethionine bound to yeast methionine tRNA I, which was labeled in the formyl group, into phage protein in the same E. coli protein-synthesizing system used above. The transfer of 14C-formylmethionine from E. coli 14C-formylmethionyl-tRNAF was investigated in parallel to that of yeast. It is known that
,
2 5 (cl ’ APUPG
Fi
1.5
p 1.0
I-
! j 0.5 GPUPG
(I, t - 0 E k%FAG
0 5 10 15
TRINUCLEOTIDE (mpMOLES OF BASE RESIDUE)
FIG. 5. Stimulation of the binding of 14C-methionyl-tRNAs to ribosomes by the trinucleotides ApUpG, GpUpG, UpUpG, and GpUpA. Assay of binding was carried out under Condition B as described in “Experimental Procedure.” 14C-Methionyl-tRNAs added to 0.05 ml of the incubation mixture were as follows: a, yeast 1’C-methionyl-tRNA I (5.7 rpmoles) prepared from the methionine tRNA I fraction used in Fig. 3; b, E. coli *4C-met,h- ionyl-tRNAF (5.9 ppmoles) prepared from purified methionine tRNAr; c, yeast W-methionyl-tRNA II (4.3 pfimoles) prepared from the methionine tRNA II fraction in Fig. 4.
by guest on February 9, 2020http://w
ww
.jbc.org/D
ownloaded from
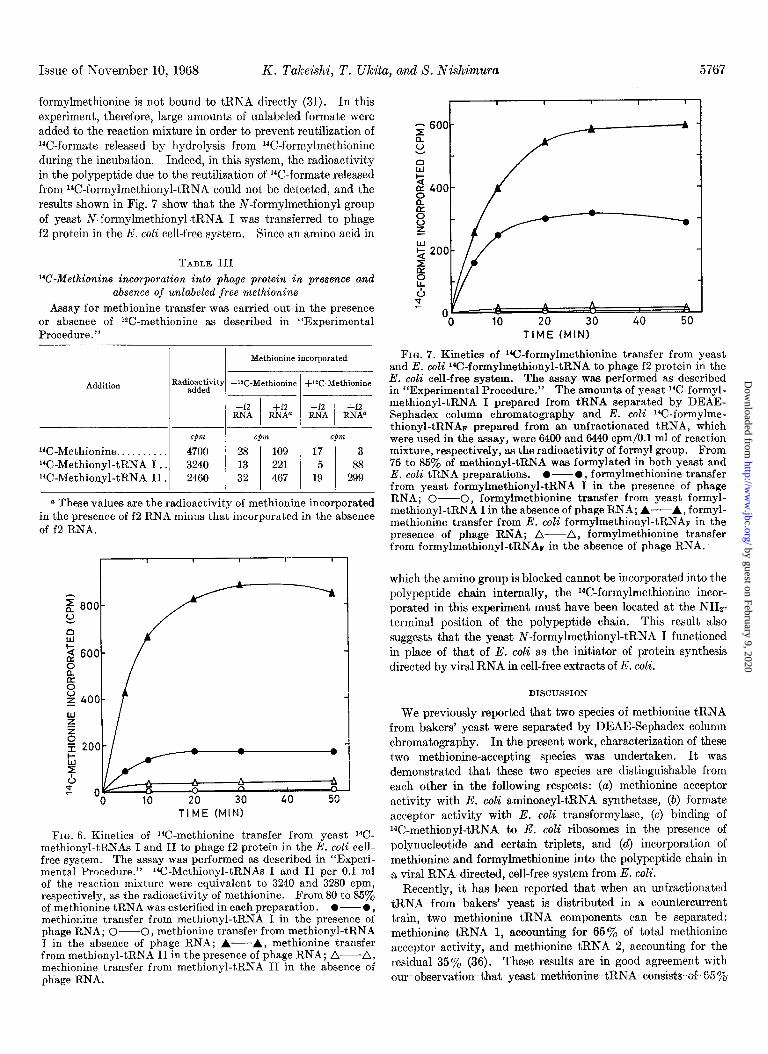
Issue of November 10, 1968 K. Takeishi, T. Ukita, and 8. Nishimura 5767
formylmethionine is not bound to tRNA directly (31). In this experiment, therefore, large amounts of unlabeled formate were added to the reaction mixture in order to prevent reutilization of 14C-formate released by hydrolysis from 14C-formylmethionine during the incubation. Indeed, in this system, the radioactivity in the polypeptide due to the reutilization of 14C-formate released from 14C-formylmethionyl-tRNA could not be detected, and the results shown in Fig. 7 show that the N-formylmethionyl group of yeast N-formylmethionyl-tRNA I was transferred to phage f2 protein in the E. coli cell-free system. Since an amino acid in
TABLE III
14C-Methionine incorporation into phage protein in presence and absence of unlabeled free methionine
Assay for methionine transfer was carried out in the presence or absence of WLmethionine as described in “Experimental Procedure.”
I I Methionine incorporated
Addition I I
CM cpm em
W-Methionine.. 4700 28 W-Methionyl-tRNA I . . 3240 13 I%-Methionyl-tRNA II. 2460 32
Q These values are the radioactivity of methionine incorporated in the presence of f2 RNA minus that incorporated in the absence of f2 RNA.
52 E 800
::
z 8 600
B 0
2 400
ki f
g 200
Li T Y - 0
0 10 20 30 40 50 TIME (MINI
FIG. 6. Kinetics of 14C-methionine transfer from yeast 14C- methionyl-tRNAs I and II to phage f2 protein in the E. coEi cell- free system. The assay was performed as described in “Experi- mental Procedure.” 1%.Methionyl-tRNAs I and II per 0.1 ml of the reaction mixture were equivalent to 3240 and 3280 cpm, respectively, as the radioactivity of methionine. From 80 to 85yo of methionine tRNA was esterified in each preparation. O-0, methionine transfer from methionyl-tRNA I in the presence of phage RNA; O-O, methionine transfer from methionyl-tRN.4 I in the absence of phage RNA; A---A, methionine transfer from methionyl-tRNA II in the presence of phage RNA; A-A, methionine transfer from methionyl-tRNA II in the absence of phage RNA.
I I I I I
p” 0 0 10 20 30 40 50
TIME (MINI
FIG. 7. Kinetics of ‘Gformylmethionine transfer from yeast and E. coli “C-formylmethionyl-tRNA to phage f2 protein in the E. coli cell-free system. The assay was performed as described in “Experimental Procedure.” The amounts of yeast “C-formyl- methionyl-tRNA I prepared from tRNA separated by DEAE- Sephadex column chromatography and E. coli 14C-formylme- thionyl-tRNAv prepared from an unfractionated tRNA, which were used in the assay, were 6400 and 6440 cpm/O.l ml of reaction mixture, respectively, as the radioactivity of formyl group. From 75 to 85yo of methionyl-tRNA was formylated in both yeast and E. coli tRNA preparations. O-O, formylmethionine transfer from yeast formylmethionyl-tRNA I in the presence of phage RNA; O---O, formylmethionine transfer from yeast formyl- methionyl-tRNA I in the absence of phage RNA; A-A, formyl- methionine transfer from E. coli formylmethionyl-tRNAr in the presence of phage RNA; A-A, formylmethionine transfer from formylmethionyl-tRNAF in the absence of phage RNA.
which the amino group is blocked cannot be incorporated into the polypeptide chain internally, the 14C-formylmethionine incor- porated in this experiment must have been located at the NHt- terminal position of the polypeptide chain. This result also suggests that the yeast N-formylmethionyl-tRNA I functioned in place of that of E. coli as the initiator of protein synthesis directed by viral RNA in cell-free extracts of E. coli.
DISCUSSION
We previously reported that two species of methionine tRNA from bakers’ yeast were separated by DEAE-Sephadex column chromatography. In the present work, characterization of these two methionine-accepting species was undertaken. It was demonstrated that these two species are distinguishable from each other in the following respects: (a) methionine acceptor activity with E. coli aminoacyl-tRNA synthetase, (a) formate acceptor activity with E. coli transformylase, (c) binding of I%-methionyl-tRNA to E. coli ribosomes in the presence of polynucleotide and certain triplets, and (d) incorporation of methionine and formylmethionine into the polypeptide chain in a viral RNA-directed, cell-free system from E. coli.
Recently, it has been reported that when an unfractionated tRNA from bakers’ yeast is distributed in a countercurrent train, two methionine tRNA components can be separated: methionine tRNA 1, accounting for 65% of total methionine acceptor activity, and methionine tRNA 2, accounting for the residual 350/, (36). These results are in good agreement with our observation that yeast methionine tRNA consists--of 65%
by guest on February 9, 2020http://w
ww
.jbc.org/D
ownloaded from

5768 Two Methionine tRNA Species from Yeast Vol. 243, No. 21
methionine tRNA I and 35yo methionine tRNA II. As the biological functions of methionine tRNAs 1 and 2 designated by Lindahl, Adams, and Fresco (36) have not been characterized in detail, it is not certain whether these methionine tRNAs correspond to our methionine tRNAs I and II, respectively. The methionine tRNAs prepared by Lindahl et al. seem to be different from our methionine tRNAs at least in the following respect. Methionine tRNA 1 reported by Lindahl et al. con- t,ained a methionine-accepting species which is interconvertible between the denatured and the renatured forms, whereas our two tRNA preparations showed neither increased nor decreased methionine acceptor activity upon either heating at 60” for 5 min in the presence of 0.01 M Mg ++ (36) or dialysis against 0.1 M EDTA solution at 4” for 72 hours.5
Each of the methionine tRNAs I and II, which were separated and partially purified by DEAE-Sephadex column chroma- tography, was eluted as a single peak from the benzoylated DEAE-cellulose column. Thus, it is very probabie that each of the tRNAs is a single species. It is obvious that the methio- nine tRNA I and II molecules are essentially different, because only the methionine tRNA I species was formylated specifically by E. coli transformylase and the two tRNA species showed different coding specificity in the trinucleotide-stimulated ribosomal binding reaction. As for t.he binding studies on E. coli met’hionyl-tRNA, it has been shown that methionyl-tRNAF responds primarily t,o the triplets ApUpG, GpUpG, and UpUpG, while methionyl-tRNAM is recognized principally by ApUpG and slightly by GpUpG (4). These results and those obtained in the present binding studies indicate that the recognition of triplet codons by yeast methionyl-tRNAs I and II is the same as that by E. coli methionyl-tRNAF and methionyl-tRNA,, respectively, and that a correlation exists between the formate acceptor activity of two methionine tRNA species and their codon recognition by the ribosomal binding technique.
It has been known that if a species specificity exists in the reaction between tRNA and aminoacyl-tRNA synthetases, the previous!y aminoacylated tRNA can be utilized in transfer reaction in cell-free extracts, the origin of which differs from that of the tRNA (37-40). It is noteworthy that N-formyl- methionine was also incorporated into a polypeptide chain via yeast tRNA in the E. coli protein-synthesizing system. This result indicates that yeast N-formylmethionyl-tRNA I as well as E. coli N-formylmethionyl-tRNAF serves as an initiator of protein synthesis in an E. coli system in vitro. From this, it seems likely that N-formylmethionine is also involved in the chain initiation of protein synthesis in yeast. We could not detect any significant transformylase activity in the yeast extracts. As Marcker and Sanger (1) suggested the existence of N-formylmethionyl-tRNA in tRNA isolated from yeast, the negative result obtained here could be attributed to the presence of an enzyme capable of hydrolyzing the ester linkage between N-formylmethionine and tRNA (41), or to other factors, and not to the absence of methionyl-tRNA transformylase. Our finding that yeast extracts inhibit the formylation of yeast methionyl-tRNA by E. coli transformylase could be explained by the- presence- of them enzyme mentioned above, whi& hy- drolyzes N-formylmethionyl-tRNA in cell-free extracts of yeast.
It is assumed that there are three sites on the tRNA molecule with respect to its biological functions: (a) to recognize a correct aminoacyl-tRN,4 synthetase, (b) to recognize the binding sites
5 J. R. Fresco, personal communication.
on ribosomes, and (c) to rec‘ognize the correct codon in messenger RNA. In addition to these sites, the formylatable methionine tRNA molecule must have a site that recognizes the trans- formylase, which seems to be specific for this tRNA molecule only. From the results described above, the similar behavior of yeast methionine tRNA I and E. coli methionine tRNA, to E. coli aminoacyl-tRNA synthetase, E. coli transformylase, the binding reaction to E. coli ribosomes, and the transfer reaction in the E. coli cell-free system suggests that both the formylatable methionine tRNA species have very similar recognition sites corresponding to the above four functions. From this point of view, it will be of considerable interest to compare the detailed structures of these two methionine tRNA molecules. Further investigations on these problems are in progress.
Acknowledgments-We are grateful to The Central Research Laboratory of Sankyo Company, Ltd., Tokyo, for preparation of large quantities of crude tRNA~ from yeast, and to Dr. T. Andoh of the Institute of Medical Science, University of Tokyo, for the preparation of phage RNA and his valuable discussions. We also thank Mr. T. Sekiya for performing the benzoglated DEAE-cellulose column chromatographic experiments.
REFERENCES
1. 2.
3.
4.
5.
6.
7.
8. 9.
10. 11.
12.
13.
14.
15.
16.
17.
18.
19.
MARCXER, K., AND SANGER, F., J. Mol. Biol., 8, 835 (1964). CLARK. B. F. C.. AND MARCKER, K. A., J. Mol. Biol., 17, 394
(1966). ’ KELLOGG, D. A., DOCTOR, B. P., LOEBEL, J. E., AND NIREN-
BERG, M. W., Proc. Nat. Acad. Sci. U. S. A., 66, 912 (1966). GHOSH, H. P., SILL, D., KHORANA, II. G., J. Mol. Biol., 26,
275 (1967). ADAMS, J. M., AND CAPECCHI, M. R., Proc. ib’at. Acad. Sci.
U. S. A.. 66, 147 (1966). WEBSTER, ‘R. E., E~GE~HARDT, D. L., AND ZINDER, N. D.,
Proc. Nut. Acad. Sci. U. S. A., 66, 155 (1966). KOLAKOB’SKY, D., AND NAKAMOTO, T., Proc. Nat. Acad. Sci.
U. X. A., 66, 1786 (1966). CLARK. B. F. C.. AND MARCKER. K. A., Nature, 211,378 (1966) STANLEY, W. M:, JR., SALAS, G., WA&DA, A.‘J., AND OCHOA,
S., Proc. Nut. Acad. Sci. U. S. A., 66, 290 (1966). CLARK, B. F. C., AND MARCKER, K. A., Nature, 207,1038 (1965). NAKAMOTO, T., AND KOLAKOFSKY, D., Proc. Nat. Acad. Sci.
U. X. A., 66, 606 (1966). THACH, R. E., DEWEY, K. F., BROWN, J. C., AND DOTY, P.,
Science, 163, 416 (1966). CAPECCHI, M. R., Proc. Nat. Acad. Sci. U. S. A., 66, 1517
(1966). VIRUELA, E., SILAS, M., AND OCI~OA, S., Proc. Nat. Acad.
Sci. U. S. 4., 67, 729 (1967). SCHWARTZ, J. II., MEYER, R., EISENSTADT, J. M., AND BRAN-
ERMAN, G., J. Mol. Biol., 26, 571 (1967). YAMANE, T., AND SUEOKA, N., Proc. Nat. Acad. Sci. U. S. A.,
60, 1093 (1963). TAKEISHI. K.. NISHIMURA, S., AND UKITA, T., Biochim. Bio-
phys. A&, i46, 605 (196j). ’ SEKIYA, T., FURUICHI, Y., YOSHIDA, M., AND UKITA, T., J.
Biochem. (Tokyo), 63, 514 (1968) GILLAM, I., MILLWARD, S., BLEW, D., VON TIGERSTROM, M.,
WIMMER, E., AND TENER, G. M., Biochemistry, 6, 3043 (1967).
20. O'DELL, B. L., VANDENBELT, J. M., BLOOM, E. S., AND PFIF- FNER,‘J. J., j. Amer. Chem. Sot., 69,250 (1947).
21. HATEFI. Y.. TALBERT. P. T., OSBORN. M. J.,~ AND BJEN- NEKE&S, I?. M., Biockemical’Prep., 7, 89 (1959).
22. RAL~INOWITZ, J. C., AND PRICER, W. E., JR., J. Biol. Chem., 237, 2898 (1962).
23. RABINOWITZ, J. C., in P. D. BOYER, H. LARDY, AND K. MYR- IJBCK (Editors), The enzymes, Vol. 2, Academic Press, New York, 1960, p. 201.
24. ZUI~AY, G., J. Mol. Biol., 4, 347 (1962).
by guest on February 9, 2020http://w
ww
.jbc.org/D
ownloaded from

Issue of November 10, 1968 K. Talceishi, 1’. Ukita, and X. Nishimura 5769
25. NISHIMURA, S., HARADA, F., NAIEUSHIMA, U., AND SENO, T., Biochim. Biophys. Beta, 142, 133 (1967).
26. WEISS, J. F., AND KELMERS, A. D., Biochemistry, 6, 2507 (1967).
27. LOEB, T., AND ZINDER, N. I)., Proc. Nal. Acad. Sci. U. S. A., 47, 282 (1961).
28. NATHANS, D., NOTANI, G., SCHWAIZTZ, J. H., AND ZINDER, N. D., Proc. Nat. Acad. Sci. U. S. A., 48, 1424 (1962).
29. NIRENBERG, M. W., AND MATTHAEI, J. H., Proc. Nat. Acad. Sci. U. S. A., 47, 1588 (1961).
30. NISHIMURA, S., JONES, D. S., OHTSUKA, E., HAYATSU, H., JACOB, T. M., AND KHORANA, H. G., J. Mol. Biol., 13, 283 (1965).
31. MARCEER, K., J. Mol. Biol., 14, 63 (1965). 32. NISHIYURA, S., AND NOVELLI, D., Proc. Nat. Acad. Sci. U. S.
A., 63, 178 (1965).
33. NIRENBERG, M., AND LEDER, P., Science, 146, 1399 (1964). 34. APGAR, J., HOLLEY, R. W., AND MERRILL, S. H., J. BioZ. Chem.,
237, 796 (1962). 35. NOLL, H., Science, 161. 1241 (1966). 36. LINDAHL, T., ADAMS, A., AND FRESCO, J. R., J. Biol. Chem.,
242, 3129 (1967). 37. NATHANS, D., AND LIPMANN, F., Biochim. Biophys. Ada. 43.
126 (1960). 38. VON EHRENSTEIN, G., AND LIPMANN, F., Proc. Nat. Acad.
Sci. U. S. A., 47, 941 (1961). 39. WEISBLUM, B., CHERAYIL, J. D., BOCK, R. M., AND SBLL,
D.. J. Mol. Biol., 28. 275 (1967). 40. PARI&, B., MILAN&I, G., VAN E&TEN, J. L., PERANI, A., AND
CIFERRI, O., J. Mol. Biol., 28, 295 (1967). 41. K&SEL, H., AND RAJBHANDARY, U. L., J. Mol. Biol., in press.
by guest on February 9, 2020http://w
ww
.jbc.org/D
ownloaded from

Keiichi Takeishi, Tyunosin Ukita and Susumu Nishimurafrom Bakers' Yeast
Characterization of Two Species of Methionine Transfer Ribonucleic Acid
1968, 243:5761-5769.J. Biol. Chem.
http://www.jbc.org/content/243/21/5761Access the most updated version of this article at
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
http://www.jbc.org/content/243/21/5761.full.html#ref-list-1
This article cites 0 references, 0 of which can be accessed free at
by guest on February 9, 2020http://w
ww
.jbc.org/D
ownloaded from





























