Embed Size (px)
Citation preview

CHARACTERIZATION OF SLP-2 FUNCTION IN T LYMPHOCYTES
(Spine title: Characterization of SLP-2 function in T lymphocytes)
(Thesis format: Integrated Article)
by
Darah A. Christie
Graduate Program in Microbiology and Immunology
A thesis submitted in partial fulfillment of the requirements for the degree of
Doctor of Philosophy
The School of Graduate and Postdoctoral Studies The University of Western Ontario
London, Ontario, Canada
© Darah Christie 2012

ii
THE UNIVERSITY OF WESTERN ONTARIO School of Graduate and Postdoctoral Studies
CERTIFICATE OF EXAMINATION
Supervisor ______________________________ Dr. Joaquín Madrenas Supervisory Committee ______________________________ Dr. Ewa Cairns ______________________________ Dr. Sean Cregan
Examiners ______________________________ Dr. John McCormick ______________________________ Dr. Lakshman Gunaratnam ______________________________ Dr. Peter Chidiac ______________________________ Dr. Nathalie Labrecque
The thesis by
Darah A. Christie
entitled:
Characterization of SLP-2 function in T lymphocytes
is accepted in partial fulfillment of the requirements for the degree of
Doctor of Philosophy
______________________ _______________________________ Date Chair of the Thesis Examination Board

iii
Abstract
Stomatin-like protein 2 (SLP-2) is a widely expressed mitochondrial-resident protein and a
member of the highly conserved stomatin and SPFH families, which includes Stomatins,
Prohibitins, Flotillins and bacterial HflC/K proteins. Studies on SPFH domain-containing
family members have pointed to a role in the organization of membranes into functional
domains as the mechanism underlying the control of a wide variety of cellular functions. The
Madrenas laboratory originally identified SLP-2 in the detergent-insoluble fraction of human
T cells activated through the T cell receptor and found that over-expression of SLP-2
increased T cell activation. Based on these results and on the conservation of function across
other SPFH family members, we hypothesized that SLP-2 acts to facilitate mitochondrial and
plasma membrane organization in response to T cell activation.
To investigate the role of SLP-2 during T cell activation, we generated stably transfected
Jurkat T cells expressing GFP-tagged SLP-2 and also produced recombinant human SLP-2
protein. These tools allowed us to demonstrate a direct interaction of SLP-2 with the
mitochondrial lipid cardiolipin and to show an association of SLP-2 with the prohibitin
complex. Furthermore, an increased recruitment of prohibitin to mitochondrial membranes
upon SLP-2 over-expression was also observed. At the cellular level, cells over-expressing
SLP-2 were found to have an increase in mitochondrial biogenesis, increased respiratory
chain activity and decreased susceptibility to apoptotic induction. Next, T cell-specific SLP-
2 conditional knockout mice were generated and found to have altered T cell mitochondrial
membrane organization in the absence of SLP-2. SLP-2-deficient T cells showed
significantly lower levels of cardiolipin in detergent-resistant mitochondrial fractions, along
with decreased complex I levels and activity. The absence of SLP-2 also led to a decrease in
T cell activation, which translated into a decreased rejection rate for non-matched tissue
transplants. Finally, for the first time it has been shown that SLP-2 is found in two
intracellular pools, one in mitochondria and the other associated with plasma membrane, with
the plasma membrane pool surrounding T cell receptors upon T cell activation. This implies
a fundamental role for SLP-2 in membrane organization.

iv
Taken together, the results presented here demonstrate a role for SLP-2 in the
compartmentalization of mitochondrial and plasma membranes into functional domains.
This, in turn, affects multi-domain receptor and multi-protein complex assembly and thus
affects such functions as mitochondrial biogenesis, respiratory chain activity, apoptotic
induction and T cell activation.
Keywords
Stomatin-like protein 2, mitochondria, membrane organization, cardiolipin, immunology, T
cell activation

v
Statement of Co-Authorship
In regards to the work presented in Chapter 2:
Christie, D., Lemke, C.D., Elias, I.M., Chau, L.A., Kirchhof, M.G., Li, B., Ball, E.H., Dunn, S.D., Hatch, G.M. and Madrenas, J. Stomatin-like protein 2 binds cardiolipin and regulates mitochondrial biogenesis and function. Mol. Cell. Biol 31 (18) 3845-56 (2011).
D. Christie contributed to experimental design, performed experiments for figures 2.1, 2.2,
2.3, 2.4, 2.5, 2.7, and 2.8 and wrote the manuscript. C.D. Lemke contributed to experimental
design and performed experiments for figure 2.1, 2.2, 2.7 and 2.8. I.M. Elias produced
recombinant human SLP-2, used in the vesicle precipitation assay in figure 2.6. L.A. Chau
performed the vesicle assay in figure 2.6. M.G. Kirchhof generated the SLP-2-GFP
expressing Jurkat cell line. B. Li performed apoptosis induction experiments for figure 2.8a
and b. E.H. Ball provided experimental design for the vesicle precipitation assay. S.D. Dunn
provided experimental design for recombinant human SLP-2 production. G.M. Hatch
provided experimental design and performed experiments for cardiolipin analysis and
enzyme activities for figures 2.3 and 2.7. J. Madrenas is the principle investigator of the lab
and contributed to experimental design and manuscript preparation.
In regards to the work presented in Chapter 3: Christie, D., Lian, D., Wang, H., Hatch, G. and Madrenas, J. SLP-2 deficiency in T cells is associated with abnormal mitochondrial compartmentalization of cardiolipin, decreased mitochondrial respiration and impaired CD4+ T cell responses. Manuscript submitted.
D. Christie contributed to experimental design and performed all experiments with the
exception of the heart transplant experiment and wrote the manuscript. D. Lian and H. Wang
performed the heart transplant surgery. G. Hatch performed cardiolipin measurements for
figure 3.2c and complex II+III complex activity for figure 3.2g. J. Madrenas is the principle
investigator of the lab and contributed to experimental design and manuscript preparation.

vi
In regards to the work presented in Chapter 4:
Christie, D., Kirchhof, M.G., Vardhana, S., Dustin, M.L., Madrenas, J. Plasma membrane and mitochondrial stomatin-like protein 2 pools coalesce at the immunological synapse during T cell activation. Manuscript in preparation.
D. Christie contributed to experimental design, performed experiments for figure 4.1, 4.2,
4.3, and 4.5 and wrote the manuscript. M.G. Kirchhof generated the SLP-2-GFP expressing
Jurkat cell line and performed the experiments for figure 4.4. S. Vardhana and M.L. Dustin
performed experiments for figure 4.6. J. Madrenas is the principle investigator of the lab and
contributed to experimental design and manuscript preparation.

vii
Acknowledgments
I would like to thank my supervisor, Dr. Joaquín Madrenas for his guidance and support
throughout my PhD studies, as well as my supervisory committee members Dr. Ewa Cairns
and Dr. Sean Cregan for helpful discussions and suggestions. I would also like to thank Dr.
Greg Dekeban for his role as acting co-supervisor during the last stages of my thesis studies.
I have had the pleasure to work with many excellent colleagues during my studies and I
thank Wendy Teft, Todd Fairhead, Cait Lemke, Mark Kirchhof, Michelle McCully, Samar
Sayedyahossein, Isaac Elias, Peter Mitsopoulos, Teresa Fernández, Holly Lemmon and
Cynthia Tang for many helpful discussions and insights. I would also like to thank Luan
Chau and Thu Chau for their endless technical expertise. As I have relied very heavily on
mouse work, I thank Melissa Pickering and Nikki Vannieuwenhuyze for taking excellent
care of my mouse colony. I would also like to thank Dr. Grant Hatch from the University of
Manitoba for sharing his cardiolipin expertise as well as Dr. Stan Dunn, Dr. Eric Ball and Dr.
Fred Possmayer from the Department of Biochemistry for help with experimental design.
Finally, I thank the department of Microbiology and Immunology. This has been an
excellent environment to complete my PhD studies and I thank all of the undergraduates,
graduate students, post-doctoral fellows, faculty members and staff for creating a supportive,
enthusiastic and creative environment.

viii
Table of Contents
CERTIFICATE OF EXAMINATION ................................................................................. ii Abstract .................................................................................................................................... iii Statement of Co-Authorship ..................................................................................................... v Table of Contents ................................................................................................................... viii List of Tables .......................................................................................................................... xii List of Figures ........................................................................................................................ xiii List of Appendices .................................................................................................................. xv List of Abbreviations ............................................................................................................. xvi Chapter 1 ................................................................................................................................... 1 1 Introduction ......................................................................................................................... 1
1.1 The SPFH family of proteins and membrane compartmentalization ........................... 4 1.1.1 The stomatin family ............................................................................................ 10 1.1.2 Stomatin-like Protein 2 ....................................................................................... 11 1.1.3 Prohibitins ........................................................................................................... 15
1.2 Mitochondrial functions ............................................................................................. 17 1.2.1 Bioenergetics and metabolism ............................................................................ 17 1.2.2 The metabolic regulation of T cells .................................................................... 21 1.2.3 Calcium signalling .............................................................................................. 23 1.2.4 Apoptosis ............................................................................................................ 25
1.3 Rationale and Hypothesis .......................................................................................... 30 1.3.1 Rationale ............................................................................................................. 30 1.3.2 Hypothesis .......................................................................................................... 31 1.3.3 Specific Aim 1: To characterize the function of the mitochondrial pool of SLP-2 ............................................................................................................................ 32 1.3.4 Specific Aim 2: To generate and characterize a T cell-specific SLP-2 conditional
knockout mouse .................................................................................................. 33 1.3.5 Specific Aim 3: To characterize the plasma membrane-associated pool of SLP-2 ............................................................................................................................ 35
1.4 References .................................................................................................................. 37 Chapter 2 ................................................................................................................................. 50 2 Stomatin-Like Protein 2 Binds Cardiolipin and Regulates Mitochondrial Biogenesis and
Function ........................................................................................................................... 50 2.1 Introduction ................................................................................................................ 50 2.2 Materials and Methods ............................................................................................... 51
2.2.1 Cells .................................................................................................................... 51 2.2.2 Plasmids, siRNA and T cell transfectants ........................................................... 52 2.2.3 Upregulation of SLP-2 expression ...................................................................... 52 2.2.4 Antibodies ........................................................................................................... 52 2.2.5 Reagents .............................................................................................................. 53 2.2.6 Confocal microscopy .......................................................................................... 53 2.2.7 Transmission electron microscopy ..................................................................... 53 2.2.8 Measurement of mitochondrial mass .................................................................. 54 2.2.9 Measurement of cardiolipin ................................................................................ 54 2.2.10 Measurement of cardiolipin mass in stable doxycycline-inducible SLP-2-GFP
transfected Jurkat cells ...................................................................................... 54

ix
2.2.11 Radiotracer studies of de novo cardiolipin synthesis ........................................ 55 2.2.12 Real time-PCR .................................................................................................. 55 2.2.13 Mitochondrial DNA quantification ................................................................... 55 2.2.14 Cell lysate preparation ...................................................................................... 56 2.2.15 Mitochondria isolation ...................................................................................... 56 2.2.16 Prohibitin and SLP-2 membrane association .................................................... 56 2.2.17 NADH dehydrogenase activity ......................................................................... 56 2.2.18 ATP quantitation ............................................................................................... 56 2.2.19 T cell stimulation .............................................................................................. 57 2.2.20 Oxygen consumption ........................................................................................ 57 2.2.21 Induction and detection of apoptosis ................................................................ 57 2.2.22 Generation of human recombinant SLP-2 (hrSLP-2). ...................................... 57 2.2.23 Phospholipid vesicle co-precipitation assay ..................................................... 59
2.3 Results ........................................................................................................................ 59 2.3.1 SLP-2 is localized mostly in mitochondria ......................................................... 59 2.3.2 Upregulation of SLP-2 expression increases mitochondrial biogenesis ............. 62 2.3.3 SLP-2 interacts with prohibitins and binds cardiolipin-enriched microdomains 71 2.3.4 Upregulation of SLP-2 expression is associated with increased mitochondrial
functions .............................................................................................................. 76 2.3.5 Upregulation of SLP-2 expression increases T cell responses and resistance to
apoptosis ............................................................................................................. 80 2.4 Discussion .................................................................................................................. 83 2.5 References .................................................................................................................. 87
Chapter 3 ................................................................................................................................. 91 3 SLP-2 deficiency in T cells is associated with abnormal mitochondrial membrane
compartmentalization of cardiolipin, decreased mitochondrial respiration and impaired CD4+ T cell responses ....................................................................................................... 91
3.1 Introduction ................................................................................................................ 91 3.2 Materials and Methods ............................................................................................... 93
3.2.1 Mice .................................................................................................................... 93 3.2.2 Confirmation of SLP-2 deletion ......................................................................... 93 3.2.3 Isolation of detergent-insoluble fraction of mitochondrial membranes .............. 94 3.2.4 Measurement of cardiolipin ................................................................................ 94 3.2.5 Enzyme activity assays ....................................................................................... 95 3.2.6 Thymocyte and T cell population analysis ......................................................... 95 3.2.7 Flow cytometry ................................................................................................... 95 3.2.8 Stimulation of T lymphocytes ............................................................................. 95 3.2.9 In vivo immunization and ex vivo recall response ............................................. 96 3.2.10 Intracellular staining cytokine analysis ............................................................. 96 3.2.11 Real time PCR analysis ..................................................................................... 96 3.2.12 Heart transplants ............................................................................................... 97 3.2.13 Activation-induced cell death assay ................................................................. 97 3.2.14 Statistical analysis ............................................................................................. 97
3.3 Results ........................................................................................................................ 97 3.3.1 Generation of T cell-specific SLP-2 knockout mice .......................................... 97 3.3.2 SLP-2 deficiency is associated with abnormal cardiolipin compartmentalization
in mitochondrial membranes and reduced mitochondrial respiration ................ 98 3.3.3 Impaired ex vivo responses of SLP-2-deficient T cells ..................................... 104

x
3.3.4 SLP-2 deficiency impairs T cell responses in vivo ........................................... 109 3.3.5 The IL-2 defect of SLP-2 deficient T cells occurs at the post-transcriptional level .......................................................................................................................... 112 3.3.6 Altered metabolic capacity of SLP-2-deficient T cells ..................................... 116
3.4 Discussion ................................................................................................................ 119 3.5 References ................................................................................................................ 123
Chapter 4 ............................................................................................................................... 129 4 Plasma membrane and mitochondrial Stomatin-like Protein 2 pools coalesce at the
immunological synapse during T cell activation ............................................................ 129 4.1 Introduction .............................................................................................................. 130 4.2 Material and Methods .............................................................................................. 131
4.2.1 Cells .................................................................................................................. 131 4.2.2 Plasmids, siRNA and T cell transfectants ......................................................... 131 4.2.3 Mice .................................................................................................................. 132 4.2.4 Antibodies ......................................................................................................... 132 4.2.5 Mitochondria isolation ...................................................................................... 133 4.2.6 Immunoprecipitations ....................................................................................... 133 4.2.7 Cell surface biotinylation .................................................................................. 133 4.2.8 Confocal microscopy ........................................................................................ 133
4.3 Results ...................................................................................................................... 135 4.3.1 The largest cellular pool of SLP-2 is associated with mitochondria ................ 135 4.3.2 Detection of a small pool of SLP-2 associated with the plasma membrane ..... 138 4.3.3 Homo-oligomerization of SLP-2 ...................................................................... 141 4.3.4 SLP-2 redistributes to the immunological synapse during T cell activation .... 141 4.3.5 The SLP-2 pools predominantly partition in the pSMAC of the immunological
synapse .............................................................................................................. 146 4.3.6 Mitochondrial recruitment to the IS does not require SLP-2 ............................ 152
4.4 Discussion ................................................................................................................ 155 4.5 References ................................................................................................................ 159
Chapter 5 ............................................................................................................................... 164 5 Discussion ....................................................................................................................... 164
5.1 Summary of results .................................................................................................. 164 5.1.1 Stomatin-like protein 2 binds cardiolipin and regulates mitochondrial biogenesis
and function ...................................................................................................... 164 5.1.2 SLP-2 deficiency in T cells is associated with abnormal mitochondrial
membrane compartmentalization of cardiolipin, decreased mitochondrial respiration and impaired CD4+ T cell responses ............................................... 166
5.1.3 Plasma membrane and mitochondrial stomatin-like protein 2 coalesce at the immunological synapse during T cell activation .............................................. 167
5.2 Model of function: SLP-2 organizes membrane domains ....................................... 169 5.2.1 Assembly of multi-domain receptors, including respiratory chain supercomplex
formation ........................................................................................................... 176 5.2.2 Calcium buffering ............................................................................................. 177 5.2.3 Mitochondria membrane associated protease regulation .................................. 179 5.2.4 Specialized membrane microdomain formation ............................................... 180 5.2.5 T cell activation ................................................................................................ 181
5.3 Future directions ...................................................................................................... 183 5.3.1 Characterization of the structure of SLP-2 ....................................................... 183

xi
5.3.2 Characterization of cardiolipin-enriched microdomains .................................. 184 5.3.3 Intracellular trafficking of SLP-2 ..................................................................... 186 5.3.4 Functional role of SLP-2 in cell physiology ..................................................... 187
5.4 SLP-2 and disease .................................................................................................... 188 5.4.1 SLP-2 and mitochondrial disorders .................................................................. 188 5.4.2 SLP-2 in cancer prognosis and metastasis ........................................................ 189
5.5 Conclusions .............................................................................................................. 190 5.6 References ................................................................................................................ 192
Appendix A: Ethics Approval ............................................................................................... 199 Appendix B: Copyright Permission ...................................................................................... 201 Curriculum Vitae .................................................................................................................. 202

xii
List of Tables
Table 3.1: T cell subsets in lymph nodes of SLP-2 T-K/O mice upon OVA immunization 113 Table 4.1: Segregation of mitochondrial SLP-2-GFP from the cSMAC in the mature IS, by
wide field fluorescence microscopy ................................................................... 150 Table 4.2: Percentage of Jurkat T cells on supported planar bilayers organizing SLP-2-GFP
close to pSMAC at the contact interface by wide field fluorescence microscopy ............................................................................................................................ 151

xiii
List of Figures
Figure 1.1: Alignment of the predicted SPFH domain of SPFH family members in human, mouse and E. coli proteins. .................................................................................... 6
Figure 1.2: Phylogenetic analysis of the SPFH domain of human, mouse and E. coli family members. ................................................................................................................ 9
Figure 1.3: Features of the SLP-2 amino acid sequence. ........................................................ 13
Figure 1.4: Schematic overview of the respiratory chain. ...................................................... 19 Figure 2.1: The major pool of SLP-2 in human T cells is located in mitochondria ............... 61
Figure 2.2: Induction of SLP-2 expression triggers mitochondrial biogenesis in human T cells. ..................................................................................................................... 64
Figure 2.3: Induction of SLP-2 expression increases de novo cardiolipin biosynthesis, nuclear transcription programs, and mitochondrial DNA replication. ............................. 67
Figure 2.4: SLP-2 upregulation precedes PGC-1α upregulation and mitochondrial biogenesis during T cell activation. ....................................................................................... 70
Figure 2.5: SLP-2 interacts with prohibitins. .......................................................................... 73 Figure 2.6: SLP-2 binds cardiolipin. ....................................................................................... 75
Figure 2.7: Induction of SLP-2 expression is associated with higher mitochondrial activity. 78 Figure 2.8: SLP-2 upregulation increases T cell responses and protects against apoptosis. .. 82
Figure 3.1: Generation of T cell-specific SLP-2 knockout mice. ......................................... 100 Figure 3.2: T cells deficient in SLP-2 have abnormal cardiolipin compartmentalization and
decreased activity of respiratory complexes I and II+III. .................................. 103 Figure 3.3: Grossly normal thymic T cell development and peripheral T cell pool in SLP-2 T-
K/O mice. ........................................................................................................... 106 Figure 3.4: T cells deficient in SLP-2 produce less IL-2 in response to T cell activation. ... 108
Figure 3.5: T cells deficient in SLP-2 have decreased responses in vivo. ............................ 111
Figure 3.6: SLP-2-deficiency in T cells induces a post-transcriptional defect in IL-2 production. ......................................................................................................... 115
Figure 3.7: Altered metabolic capacity of SLP-2-deficient T cells during activation. ......... 118
Figure 4.1: SLP-2 is a mitochondrial protein. ....................................................................... 137
Figure 4.2: A small pool of SLP-2 is associated with the plasma membrane. ..................... 140 Figure 4.3: Homo-oligomerization of SLP-2. ....................................................................... 143
Figure 4.4: SLP-2 polarizes to the immunological synapse during T cell activation. .......... 145

xiv
Figure 4.5: Redistribution of plasma membrane-associated SLP-2 and mitochondria-associated SLP-2 during immunological synapse formation. ............................ 148
Figure 4.6: SLP-2-deficient T cells show normal mitochondrial recruitment upon T cell stimulation. ......................................................................................................... 154
Figure 5.1: A step-wise model of the formation of cardiolipin-enriched microdomains in the mitochondrial membrane as facilitated by SLP-2. ............................................. 173
Figure 5.2: Model of supercomplex formation upon SLP-2 upregulation in response to T cell activation. ........................................................................................................... 175

xv
List of Appendices
Appendix A: Ethics Approval ............................................................................................... 199 Appendix B: Copyright Permission ...................................................................................... 201

xvi
List of Abbreviations
AAA ATPases associated with diverse cellular activities
ActD Actinomycin D
ADP Adenosine diphosphate
AMPK AMP-activated protein kinase
ANOVA Analysis of variance
AP-1 Activator protein 1
Apaf-1 Apoptotic protease activating factor 1
APC Antigen presenting cell
ASIC Acid sensing ion channel
ATP Adenosine triphosphate
BAK Bcl-2 homologous antagonist/killer
BAX Bcl-2 associated X protein
BCL-2 B cell lymphoma-2
BLAST Basic local alignment search tool
CCO Cytochrome c oxidase I
CD Cluster of differentiation
Cdk Cyclin dependent kinase
CL Cardiolipin
CoA Coenzyme A
CRAC Calcium release activated calcium
CS Citrate synthase
cSMAC Central supramolecular activation cluster
DAG Diacylglycerol

xvii
DEG/EnaC Degenerin/epithelial sodium channel
DI-GEM Detergent-insoluble glycolipid-enriched microdomain
DIABLO Direct IAP binding protein with low pI
DN Double negative
DP Double positive
Drp-1 Dynamin-related protein-1
ELISA Enzyme linked immunosorbent assay
ER Endoplasmic reticulum
ERK Extracellular signal-regulated kinase
EST Expressed sequence tag
FAD Flavin adenine dinucleotide
Gads GRB2-related adapter downstream of Shc
GAPDH Glyceraldehyde 3-phosphate dehydrogenase
GFP Green fluorescent protein
GRB2 Growth factor receptor-bound protein 2
hrSLP-2 Human recombinant SLP-2
IAP Inhibitor of apoptosis
ICAM-1 Intercellular adhesion molecule 1
IFN Interferon
IL Interleukin
IP Immunoprecipitation
IP3 Inositol triphosphate
IS Immunological synapse
ITAM Immunoreceptor tyrosine-based activation motif
Itk IL-2 inducible T cell kinase

xviii
LAT Linker of activated T cells
LCMV Lymphocytic choriomeningitis virus
LFA-1 Lymphocyte function-associated antigen 1
MAP Mitogen activated protein
MEF Mouse embryonic fibroblast
Mff Mitochondrial fission factor
Mfn Mitofusin
MHC Major histocompatibility complex
mTOR Mammalian target of rapamycin
mtRFP Mitochondrial red fluorescence protein
NAD Nicotinamide adenine dinucleotide
NAO Nonyl acridine orange
NFAT Nuclear factor of activated T cells
NFκB Nuclear factor κB
OPA-1 Optic atrophy 1
OVA Ovalbumin
OXPHOS Oxidative phosphorylation
P Pellet
PBMC Peripheral blood mononuclear cells
PC Phosphatidylcholine
PCC Pigeon cytochrome c
PE Phosphatidylethanolamine
PG Phosphatidylglycerol
PGC PPAR (peroxisome proliferator-activated receptor) - γ coactivator
PHB Prohibitin

xix
PI Phosphatidylinositol
PI3K Phosphoinositide 3-kinase
PKC Protein kinase C
PLC-γ1 Phospholipase C-γ1
PMA Phorbol myristate acetate
PS Phosphatidylserine
pSMAC Peripheral supramolecular activation cluster
RT-PCR Reverse transcriptase polymerase chain reaction
S Supernatant
SDH Succinate dehydrogenase
SDS-PAGE Sodium dodecyl polyacrylamide gel electrophoresis
SEE Staphylococcal enterotoxin E
SHP Src homology phosphatase
siRNA Small interfering RNA
SLP Stomatin-like protein
SLP-76 Src homology 2 domain containing leukocyte protein of 76kDa
SMAC Supramolecular activation cluster
SMAC Second mitochondria-derived activator of caspases
SOS Son of sevenless
SP Single positive
SPFH Stomatin, Prohibitin, Flotillin, HflC/K
STIM1 Stromal interaction molecule 1
T-K/O T cell knockout
tBID Truncated BID
TCA Tricarboxylic acid

xx
TCR T cell receptor
TH T helper
TIRFM Total internal reflection fluorescence microscopy
TMHMM Transmembrane hidden Markov model
TRAF TNF-receptor associated factor
TRAIL TNF-related apoptosis-inducing ligand
Treg T regulatory
UQ Ubiquinone
VDAC Voltage dependent anion channel
ZAP-70 Zeta-chain associated protein kinase 70
ΔΨm Mitochondrial transmembrane potential

1
Chapter 1
1 Introduction
The immune system has evolved to provide a defense against invading pathogens,
including bacteria, viruses and fungi that threaten the health and survival of the animal.
T cells function to directly lyse infected cells or to secrete cytokines, which activate other
immune cells to eliminate pathogens. The function of T cells requires the translation of
antigen recognition into gene expression to facilitate pathogen elimination. This occurs
via intracellular signalling cascades that translate T cell receptor (TCR) binding into the
activation of transcriptional programs. T cell signalling events are complex and require
the interaction of a large number of signalling and adapter proteins at both the
extracellular and cytoplasmic faces of the plasma membrane. The organization of the
plasma membrane into specialized micro-domains to sequester signalling components
away from other plasma membrane proteins could strengthen the association of signalling
components to promote T cell activation 1, 2.
The TCR is a dimer composed of two transmembrane proteins with large extracellular
domains forming a ligand recognition site. The TCR ligand is a specific antigenic
peptide bound to a major histocompatibility complex (MHC) molecule. The TCR is
found at the cell surface in association with CD3 complex composed of molecules of γε,
δε or ζζ pairs 3, 4. The intracellular domains of the CD3 proteins contain regions known
as immunoreceptor tyrosine-based activation motifs (ITAM), which consist of two
tyrosine residues separated by a defined amino acid sequence (YXXL/I-X6-8-YXXL/I) 5, 6.
These tyrosine residues can be phosphorylated and act as a docking site for proteins
involved in the T cell signalling cascade. Upon antigen recognition, CD4 or CD8
molecules cluster together with the TCR, by virtue of their binding to MHC of class II or
class I respectively, on antigen presenting cells. Both CD4 and CD8 molecules are
associated with the Src family kinase Lck, which phosphorylates the ζ chain ITAMs 7.
The Syk family kinase ZAP-70 then binds the phosphorylated ITAMs and, in turn,

2
phosphorylates the adapters LAT (linker of activated T cells) and SLP-76 (Src homology
2 domain containing leukocyte protein of 76 kDa). LAT is a transmembrane adapter
protein which upon phosphorylation by ZAP-70, provides docking sites for a number of
signalling and adapter molecules, including Gads (GRB2-related adapter downstream of
Shc), which bind to and recruit other adapters, such as SLP-76 8. The complex of
signalling and scaffold proteins recruited to the TCR upon antigen binding is known as
the TCR signalosome.
A major signalling event triggered at the LAT/SLP-76 scaffold is the recruitment and
activation of phospholipase C γ1 (PLCγ1). This enzyme is recruited upon
phosphorylation of LAT and is activated by another kinase recruited to the LAT/SLP-76
scaffold, the Tec family kinase Itk (IL-2 inducible T cell kinase). Activated PLCγ
cleaves the lipid phosphatidylinositol-4,5-bisphosphate into diacylglycerol (DAG) and
inositol-1,4,5-triphosphate (IP3), each of which initiate signalling pathways 5. DAG
activates the mitogen activated protein (MAP) kinase cascade, leading to the activation of
Elk1, a subunit of the dimeric AP-1 transcription factor complex. The MAP kinase
cascade is also activated by guanine nucleotide exchange factor son of sevenless (SOS)
which is recruited to the LAT/SLP-76 scaffold by the adapter GRB2. In addition to the
MAP kinase cascade, DAG also activates protein kinase Cθ (PKCθ), which leads to the
activation of the nuclear factor κB (NFκB) family of transcription factors. In parallel, IP3
triggers the release of intracellular calcium stores, which triggers the opening of cell
surface calcium channels. The surge of intracellular calcium then activates calcineurin,
which in turn, activates the nuclear factor of activated T cells (NFAT) family of
transcription factors 9. The transcription factors activated from these various signalling
cascades upregulate the expression of effector molecules needed for T cell responses.
TCR ligation and signalling also triggers the formation of the immunological synapse, a
unique organization of the T cell membrane into specialized domains involved in the
maintenance and regulation of T cell signalling 10, 11. The peripheral supramolecular
activation cluster (pSMAC) is a ring of high avidity lymphocyte function associated
antigen 1 (LFA-1) and intracellular adhesion molecule 1 (ICAM-1), which facilitate the
tight association with the antigen presenting cell. On the other hand, as the TCR:MHC

3
complexes form in the pSMAC, they migrate into and concentrate at the center of the
SMAC (cSMAC), driven by actin polymerization 12. The pSMAC excludes large
inhibitory molecules, including phosphatases such as CD45, which is segregated to the
distal SMAC, to prevent inactivation of the T cell signalosome. Although it was
originally thought that T cell signalling occurred at the cSMAC, blocking the formation
of new TCR:MHC complexes showed that as TCR:MHC complexes migrate into the
cSMAC they dissociate from ZAP-70, indicating a loss of T cell signalling 13. Thus, the
organization of the immune synapse allows for the termination of TCR signalling
followed by TCR degradation upon entering the cSMAC 14.
In addition to the membrane organization following antigen recognition, T cell signalling
is thought to involve membrane compartmentalization into lipid microdomains enriched
in cholesterol and glycolipids known as detergent-insoluble glycolipid-enriched
microdomains (DI-GEMs) or lipid rafts. These are highly structured assemblies of
tightly packed lipids and proteins found in the plasma membrane that are thought to exist
in nano-units that coalesce in response to cell stimulation 1, 2. Studies have shown that T
cell stimulation leads to the aggregation of lipid rafts at the T cell signalosome, which
may be important for T cell activation 15-17. In addition, a number of T cell signalosome
components are constitutively associated with lipid raft domains, including Lck and LAT 18. This localization appears to be essential for T cell activation as a mutant LAT unable
to associate with lipid rafts leads to the inhibition of T cell responses 19. The formation of
lipid raft domains from the nanoassemblies is likely facilitated by lipid-lipid, lipid-protein
and protein-protein interactions and as such, actin rearrangement during T cell activation
is thought to help raft organization 20. A number of studies have also shown that
cholesterol and sphingomyelin disruption leads to altered T cell responses, likely due to
disruption in raft formation 21, 22. This membrane organization is likely important for
sequestering signalling components while also excluding inhibitors such as CD45 or
SHP-1, which are normally found outside DI-GEMs 23, 24.
Within this framework and with the aim to identify novel proteins involved in the
regulation of T cell activation and membrane organization, our lab identified stomatin-
like protein 2 (SLP-2), a member of the highly conserved stomatin family. The

4
investigation of the functional role played by SLP-2 in T cell activation will be the focus
of this thesis.
1.1 The SPFH family of proteins and membrane compartmentalization
The organization of membrane components into defined functional domains plays an
important role in many cellular processes, including cell signalling, membrane
trafficking, ion channel activity and viral budding 1, 25. The SPFH family of proteins,
consisting of Stomatin, Prohibitin, Flotillin and the bacterial HflC/K proteins, has been
proposed to play a role in the organization of membrane domains by binding directly to
lipids, as well as to other proteins, to facilitate membrane compartmentalization 26, 27.
Members of this family are anchored to various cellular membranes, including the plasma
membrane, early endosomes, Golgi apparatus, mitochondria and endoplasmic reticulum.
SPFH family members are defined by the highly conserved, shared SPFH domain (Fig.
1.1, 1.2), which is proposed to facilitate lipid association as the SPFH family members
podocin and Mec-2 have been shown to bind directly to the lipid raft component
cholesterol and many SPFH family members are enriched in DI-GEMs, where they carry
out many diverse cellular functions 26, 27. Prohibitins are mitochondrial proteins proposed
to be involved in mitochondrial chaperone activity 28, 29, organization of mitochondrial
DNA nucleoids 30, regulation of lifespan 31, 32, proliferation and apoptosis 33,
compartmentalization of mitochondrial membranes 34, 35 and mitochondrial biogenesis 29,
36, 37. Flotillins are widely expressed and found at the plasma membrane as well as
intracellular membranes and have been proposed to organize lipid rafts, which may
regulate a number of functions, including insulin-stimulated glucose uptake, neuronal
regeneration, phagocytosis, cell proliferation, signal transduction and cytoskeletal
remodeling (reviewed in 38, 39). Finally, the bacterial proteins HflC/K are hypothesized to
be involved in the regulation of FtsH, a zinc metalloprotease with high similarity to the

5
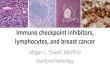
Figure 1.1: Alignment of the predicted SPFH domain of SPFH family members in
human, mouse and E. coli proteins.
The predicted SPFH domains from human (h) and mouse (m) SPFH family members
stomatin, SLP-1, SLP-2, SLP-3, podocin, flotillin 1, flotillin 2, PHB1 and PHB2 along
with E. coli HflC and HflK proteins were aligned using Clustal W and conserved residues
and the SPFH domain consensus sequence were predicted using Boxshade software.

6
Figure 1.1
hStomatin 52 MCIKIIKEYERAIIFRLGRILQGG-----AKGPGLFFILPCTDSFIKVD-MRTISFDIPP mStomatin 52 ICIKIVKEYERVIIFRLGRILQGG-----AKGPGLFFILPCTDSLIKVD-MRTISFDIPP hSLP-3 49 ---------ERAVVFRLGRIQADK-----AKGPGLILVLPCIDVFVKVD-LRTVTCNIPP mSLP-3 45 MCLKIIKEYERAVVFRLGRIQADK-----AKGPGLILVLPCIDVFVKVD-LRTVTCNIPP hPodocin 123 FCVKVVQEYERVIIFRLGHLLPGR-----AKGPGLFFFLPCLDTYHKVD-LRLQTLEIPF mPodocin 125 FCIKVVQEYERVIIFRLGHLLPGR-----AKGPGLFFFLPCLDTYHKVD-LRLQTLEIPF hSLP-1 77 FALKIVPTYERMIVFRLGRIRT-------PQGPGMVLLLPFIDSFQRVD-LRTRAFNVPP mSLP-1 77 FALKIVPTYERMIVFRLGRIRN-------PQGPGMVLLLPFIDSFQRVD-LRTRAFNVPP hSLP-2 36 TVVLFVPQQEAWVVERMGRFHR-------ILEPGLNILIPVLDRIRYVQSLKEIVINVPE mSLP-2 36 TVILFVPQQEAWVVERMGRFHR-------ILEPGLNVLIPVLDRIRYVQSLKEIVINVPE hFlotillin1 4 ---TCGPNEAMVVSG-FCRSP-PV-----MVAGGRVFVLPCIQQIQRIS-LNTLTLNVKS mFlotillin1 4 ---TCGPNEAMVVSG-FCRSP-PV-----MVAGGRVFVLPCIQQIQRIS-LNTLTLNVKS hFlotillin2 6 ---TVGPNEALVVSGGCCGSDYKQ-----YVFGGWAWAWWCISDTQRIS-LEIMTLQPRC mFlotillin2 6 ---TVGPNEALVVSGGCCGSDYKQ-----YVFGGWAWAWWCISDTQRIS-LEIMTLQPRC hPHB1 26 ALYNVDAGHRAVIFDRFRGVQ-DI-----VVGEGTHFLIPWVQKPIIFD-CRSRPRNVP- mPHB1 26 ALYNVDAGHRAVIFDRFRGVQ-DI-----VVGEGTHFLIPWVQKPIIFD-CRSRPRNVP- hPHB2 39 SVFTVEGGHRAIFFNRIGGVQQDT-----ILAEGLHFRIPWFQYPIIYD-IRARPRKIS- mPHB2 39 SVFTVEGGHRAIFFNRIGGVQQDT-----ILAEGLHFRIPWFQYPIIYD-IRARPRKIS- E.coliHflC 18 MSVFVVKEGERGITLRFGKVLRDDDNKPLVYEPGLHFKIPFIETVKMLD-ARIQTMDNQA E.coliHflK 95 SGFYTIKEAERGVVTRFGKFSH-------LVEPGLNWKPTFIDEVKPVN-VEAVRELAAS consensus 1 i ii ervii rlgri ilgpGl filpcid kvd lrtis nip hStomatin 106 QEILTKDSVTISVDGVVYYRVQN----------ATLAVANITNADSATRLLAQTTLRNVL mStomatin 106 QEVLTKDSVTISVDGVVYYRVQN----------ATLAVANITNADSATRLLAQTTLRNAL hSLP-3 94 QEILTRDSVTTQVDGVVYYRIYS----------AVSAVANVNDVHQATFLLAQTTLRNVL mSLP-3 99 QEILTRDSVTTQVDGVVYYRIYS----------AVSAVANVNDVHQATFLLAQTTLRNVL hPodocin 177 HEIVTKDMFIMEIDAICYYRMEN----------ASLLLSSLAHVSKAVQFLVQTTMKRLL mPodocin 179 HEVVTKDMFIMEIDAVCYYRMEN----------ASLLLSSLAHVSKAIQFLVQTTMKRLL hSLP-1 129 CKLASKDGAVLSVGADVQFRIWD----------PVLSVMTVKDLNTATRMTAQNAMTKAL mSLP-1 129 CKLASKDGAVLSVGADVQFRIWD----------PVLSVMAVKDLNTATRMTAHNAMTKAL hSLP-2 89 QSAVTLDNVTLQIDGVLYLRIMD----------PYKASYGVEDPEYAVTQLAQTTMRSEL mSLP-2 89 QSAVTLDNVTLQIDGVLYLRIMD----------PYKASYGVEDPEYAVTQLAQTTMRSEL hFlotillin1 53 EKVYTRHGVPISVTGIAQVKIQGQNKEMLAAACQMFLGKTEAEIAHIALETLEGHQRAIM mFlotillin1 53 EKVYTRHGVPISVTGIAQVKIQGQNKEMLAAACQMFLGKTEAEIAHIALETLEGHQRAIM hFlotillin2 57 EDVETAEGVALTVTGVAQVKIMT-EKELLAVACEQFLGKNVQDIKNVVLQTLEGHLRSIL mFlotillin2 57 EDVETAEGVALTVTGVAQVKIMT-EKELLAVACEQFLGKNVQDIKNVVLQTLEGHLRSIL hPHB1 78 VITGSKDLQNVNITLRILFRPVASQLP------RIFTSIGEDYDERVLPSITTEILKSVV mPHB1 78 VITGSKDLQNVNITLRILFRPVASQLP------RIYTSIGEDYDERVLPSITTEILKSVV hPHB2 92 SPTGSKDLQMVNISLRVLSRPNAQELP------SMYQRLGLDYEERVLPSIVNEVLKSVV mPHB2 92 SPTGSKDLQMVNISLRVLSRPNAQELP------SMYQRLGLDYEERVLPSIVNEVLKSVV E.coliHflC 77 DRFVTKEKKDLIVDSYIKWRISDFSR-----YYLATGGGDISQAEVLLKRKFSDRLRSEI E.coliHflK 147 GVMLTSDENVVRVEMNVQYRVTN----------PEKYLYSVTSPDDSLRQATDSALRGVI consensus 61 iltkd v i vdgvv yri v i dvd av l q lr vl

7
hStomatin 156 GTKNLSQILSD-REEIAHNMQSTLD----------------------------------- mStomatin 156 GTKNLSQILSD-REEIAHHMQSTLD----------------------------------- hSLP-3 144 GTQTLSQILAG-REEIAHSIQTLLD----------------------------------- mSLP-3 149 GTQTLSQILSG-REEIAHSIQTLLD----------------------------------- hPodocin 227 AHRSLTEILLE-RKSIAQDAKVALD----------------------------------- mPodocin 229 AHRSLTEILLE-RKSIAQDVKVALD----------------------------------- hSLP-1 179 LKRPLREIQME-KLKISDQLLLEIN----------------------------------- mSLP-1 179 LRRPLQEIQME-KLKIGDQLLLEIN----------------------------------- hSLP-2 139 GKLSLDKVFRE-RESLNASIVDAIN----------------------------------- mSLP-2 139 GKLSLDKVFRE-RESLNANIVDAIN----------------------------------- hFlotillin1 113 AHMTVEEIYKD-RQKFSEQVFKVAS----------------------------------- mFlotillin1 113 AHMTVEEIYKD-RQKFSEQVFKVAS----------------------------------- hFlotillin2 116 GTLTVEQIYQD-RDQFAKLVREVAA----------------------------------- mFlotillin2 116 GTLTVEQIYQD-RDQFAKLVREVAA----------------------------------- hPHB1 132 ARFDAGELITQ-RELVSRQVSDDLT----------------------------------- mPHB1 132 ARFDAGELITQ-RELVSRQVSDDLT----------------------------------- hPHB2 146 AKFNASQLITQ-RAQVSLLIRRELT----------------------------------- mPHB2 146 AKFNASQLITQ-RAQVSLLIRRELT----------------------------------- E.coliHflC 132 GRLDVKDIVTDSRGRLTLEVRDALNSGSAGTEDEVTTPAADNAIAEAAERVTAETKGKVP E.coliHflK 197 GKYTMDRILTEGRTVIRSDTQRELEET--------------------------------- consensus 121 gh tl il d re i m l hStomatin 180 ----DATDAWGIKVERVEIKDVKLPVQLQRAMAAEAEASREARAKVIAAEGE-------- mStomatin 180 ----DATDDWGIKVERVEIKDVKLPVQLQRAMAAEAEAAREARAKVIAAEGE-------- hSLP-3 168 ----DATELWGIRVARVEIKDVRIPVQLQRSMAAEAEATREARAKVLAAEGEMNASKSLK mSLP-3 173 ----DATELWGIRVARVEIKDVRIPVQLQRSMAAEAEATREARAKVLAAEGE-------- hPodocin 251 ----SVTCIWGIKVERIEIKDVRLPAGLQHSLAVEAEAQRQAKVRMIAAEAEK------- mPodocin 253 ----AVTCIWGIKVERTEIKDVRLPAGLQHSLAVEAEAQRQAKVRVIAAEGEK------- hSLP-1 203 ----DVTRAWGLEVDRVELAVEAVLQPPQDSPAG-------------------------- mSLP-1 203 ----DVTRAWGLEVDRVELAVEAVLQPPQDSLTVPSLDS--------------------- hSLP-2 163 ----QAADCWGIRCLRYEIKDIHVPPRVKESMQMQVEAERRKRATVLESEG--------- mSLP-2 163 ----QAADCWGIRCLRYEIKDIHVPPRVKESMQMQVEAERRKRATVLESEG--------- hFlotillin1 137 ----SDLVNMGISVVSYTLKDIHDDQDYLHSLGKARTAQVQKDARIGEAEAKRDAGIREA mFlotillin1 137 ----SDLVNMGISVVSYTLKDIHDDQDYLHSLGKARTAQVQKDARIGEAEAKRDAGIREA hFlotillin2 140 ----PDVGRMGIEILSFTIKDVYDKVDYLSSLGKTQTAVVQRDADIGVAEAERDAGIREA mFlotillin2 140 ----PDVGRMGIEILSFTIKDVYDKVDYLSSLGKTQTAVVQRDADIGVAEAERDAGIREA hPHB1 156 ----ERAATFGLILDDVSLTHLTFGKEFTEAVEAKQVAQQEAERARFVVEKAEQQ----- mPHB1 156 ----ERAATFGLILDDVSLTHLTFGKEFTEAVEAKQVAQQEAERARFVVEKAEQQ----- hPHB2 170 ----ERAKDFSLILDDVAITELSFSREYTAAVEAKQVAQQEAQRAQFLVEKAKQE----- mPHB2 170 ----ERAKDFSLILDDVAITELSFSREYTAAVEAKQVAQQEAQRAQFLVEKAKQE----- E.coliHflC 192 VINPNSMAALGIEVVDVRIKQINLPTEVSEAIYNRMRAEREAVARRHRSQGQ-------- E.coliHflK 224 ----IRPYDMGITLLDVNFQAARPPEEVKAAFDDAIAARENEQQYIREAEAY-------- consensus 181 d wgi verveikdvklp dl sma a a a v aeg

8
Figure 1.2: Phylogenetic analysis of the SPFH domain of human, mouse and E. coli
family members.
The predicted SPFH domains from human (h) and mouse (m) SPFH family members
stomatin, SLP-1, SLP-2, SLP-3, podocin, flotillin 1, flotillin 2, PHB1 and PHB2 along
with E. coli HflC and HflK proteins were aligned using Clustal W and a phylogenetic tree
was generated.

9
Figure 1.2

10
AAA (ATPases associated with diverse cellular activities) family of ATPases 40, a
function that may be conserved in prohibitin 37. While each member has unique
functions, many of which are currently unknown, all seem to be involved in regulating
the formation or organization of DI-GEMs to affect various cell processes, a function that
is likely conserved in the stomatin family 27, 38, 39.
1.1.1 The stomatin family
The stomatin family consists of 5 members, stomatin, stomatin-like protein 1 (SLP-1),
SLP-2, SLP-3 and podocin 41. This is a highly conserved family of proteins with
homologues found in primates, rodents, birds, amphibians, teleosts, insects, nematodes,
fungi, plants, prokaryotes and Archaea 41. Stomatin was originally identified in patients
with stomatocytosis, a hemolytic anemia resulting from increased membrane
permeability to sodium and potassium 42. Erythrocytes from these patients were found to
be deficient in stomatin 43, indicating a possible role in cation channel regulation.
Surprisingly, although there is high similarity between human and mouse stomatin, a
stomatin knockout mouse failed to show a stomatocytosis phenotype, indicating that
while stomatin may be involved in regulating cation transport, it is not responsible for the
phenotype of erythrocytes of patients with stomatocytosis 44. Further evidence of
stomatin-mediated ion channel regulation has been shown for the degenerin / epithelial
sodium channel (DEG/ENaC) family of ion channels, including the acid-sensing ion
channels (ASIC) 45-49. Initial studies into DEG/ENaC regulation was carried out in
Caenorhabditis elegans, which expresses three stomatin homologues, mec-2, unc-1 and
unc-24 50-52. Mec-2 was originally identified in touch-insensitive mutants and is thought
to regulate DEG/ENaC mechanosensory transduction channels as Mec-2 localizes to the
touch-receptor neurons in C. elegans and co-expression in oocytes with the DEG/ENaC
ion channel components Mec-4 and Mec-10 leads to increased current in cells 47. This
finding was corroborated with human and mouse stomatin, both of which associate with
ASIC when co-expressed in COS-7 cells and co-expression with ASIC3 leads to reduced
current in cells, indicating a regulatory role for stomatin 46. Furthermore, SLP-3 is also
expressed in mouse dorsal root ganglions and SLP-3 mutant mice have decreased

11
mechanosensitivity 53, similar to Mec-2 mutants and suggesting SLP-2 and its
homologues may be important in complex receptor assemblies.
Unc-1 was originally identified in C. elegans mutants with altered sensitivity to volatile
anesthetics, including an increase in sensitivity to the inhaled anesthetic diethyl ether 51.
Interestingly, this phenotype is conserved in the stomatin knockout mouse, as these mice
also show an increased sensitivity to diethyl ether 54. Taken together, studies of stomatin
family members in both C. elegans and mouse models have shown a role for this family
in ion channel regulation, yet the exact mechanisms remain elusive.
Stomatin family members have also been shown to localize to DI-GEMs and have been
proposed to organize these membranes into functional domains through associations with
actin microfilaments 55-57. Podocin is found in detergent-resistant membranes in
glomeruli 58, 59 and is thought to control intracellular distribution of proteins involved in
kidney ultrafiltration as HEK 293 cells co-expressing podocin and nephrin show co-
localization in DI-GEMs while cells lacking podocin fail to recruit nephrin to these
domains 60. Similarly, SLP-1 has also been shown to control the localization of stomatin 61, as cells over-expressing SLP-1 show an increase in recruitment of stomatin to the late
endosomal compartment, which is the subcellular localization of SLP-1. The localization
of these proteins to lipid domains is likely important for proper cellular function.
1.1.2 Stomatin-like Protein 2
SLP-2 was originally identified in a BLAST search of the EST database using the human
stomatin cDNA sequence 62. The 1071 nucleotide SLP-2 gene encodes a 357 amino acid
protein and is located on human chromosome 9p13. Sequence analysis indicates that
SLP-2 lacks a putative transmembrane domain and palmitoylation sites found in other
stomatin family members. However, SLP-2 does share sequence homology with the
other stomatin family members in the predicted C-terminal α-helical region and proximal
β-sheet domains. Similar to other stomatin family members, SLP-2 is also found in

12
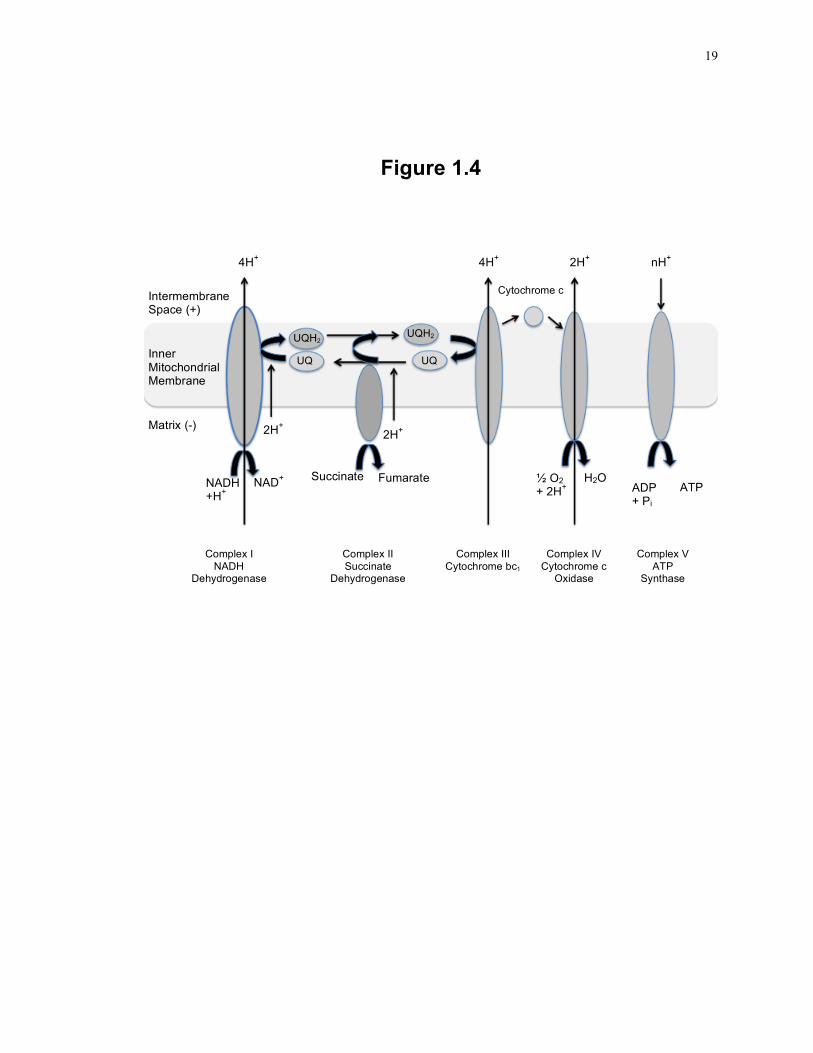
Figure 1.3: Features of the SLP-2 amino acid sequence.
Alignment of human and mouse SLP-2 amino acid sequence, showing conservation of
sequence as well as putative post-translational modifications. The mitochondrial
targeting sequence, predicted by the Mitoprot program, is shown in the yellow shaded
sequences. The SPFH domain is shown in the underlined sequence, as predicted by
InterPro scan. Putative post-translational modifications of the SLP-2 protein, as
predicted by Prosite, are indicated by bold text and arrows above the sequence to denote
the type of modification. The stomatin family consensus sequence is indicated by *
above the sequence alignment, where the consensus sequence is defined as
RX2(L/I/V)(S/A/N)X6(L/I/V)DX2TX2WG(L/I/V)X(K/R)(L/I/V)E(L/I/V)(K/R) 62. The
conservation of sequence between mouse and human SLP-2 is indicated below the
sequence alignment, where * indicates a conservation, : indicates highly similar residues
and . indicates weakly similar residues. Abbreviations: myr: myristoylation site,
PKC: protein kinase C phosphorylation site, cAMP: cAMP and cGMP – dependent
protein kinase phosphorylation site, CKII: casein kinase II phosphorylation site, N-gly:
N-linked glycosylation site, h – human modification only, m – mouse modification only.
13
Figure 1.3
cAMP myr PKC myr hSLP-2 MLARAARGTGALLLRGSLLASGRAPRRASSGLPRNTVVLFVPQQEAWVVERMGRFHRILE 60 mSLP-2 MLARAARGTGALLLRGSVQASGRVPRRASSGLPRNTVILFVPQQEAWVVERMGRFHRILE 60
*****************: ****.*************:********************** PKC/CKII N-Gly hSLP-2 PGLNILIPVLDRIRYVQSLKEIVINVPEQSAVTLDNVTLQIDGVLYLRIMDPYKASYGVE 120 mSLP-2 PGLNVLIPVLDRIRYVQSLKEIVINVPEQSAVTLDNVTLQIDGVLYLRIMDPYKASYGVE 120 ****:******************************************************* N-Gly(h) PKC CKII(h)
* ** ** * ***** ***** hSLP-2 DPEYAVTQLAQTTMRSELGKLSLDKVFRERESLNASIVDAINQAADCWGIRCLRYEIKDI 180 mSLP-2 DPEYAVTQLAQTTMRSELGKLSLDKVFRERESLNANIVDAINQAADCWGIRCLRYEIKDI 180
***********************************.************************ cAMP/CKII myr amidation CKII hSLP-2 HVPPRVKESMQMQVEAERRKRATVLESEGTRESAINVAEGKKQAQILASEAEKAEQINQA 240 mSLP-2 HVPPRVKESMQMQVEAERRKRATVLESEGTRESAINVAEGKKQAQILASEAEKAEQINQA 240
************************************************************ CKII hSLP-2 AGEASAVLAKAKAKAEAIRILAAALTQHNGDAAASLTVAEQYVSAFSKLAKDSNTILLPS 300 mSLP-2 AGEASAVLAKAKAKAEAIRILAGALTQHNGDAAASLTVAEQYVSAFSKLAKDSNTVLLPS 300
**********************.********************************:**** myr myr PKC/CKII CKII hSLP-2 NPGDVTSMVAQAMGVYGALTKAPVPGTPDSLSSGSSRDVQGTDASLDEELDRVKMS 356 mSLP-2 NPSDVTSMVAQAMGVYGALTKAPVPGAQN--SSQSRRDVQATDTSI-EELGRVKLS 353
**.***********************: : ** * ****.**:*: ***.***:* N-Gly(m) CKII (m)

14
association with DI-GEM 63, which may be mediated by protein myristoylation or may
occur through the SPFH domain, in a similar manner to Mec-2 and podocin, both of
which have been shown to associate with cholesterol through the SPFH domain 26. As
shown in Fig. 1.3 human and mouse SLP-2 are highly conserved and share a number of
putative post-translational modifications, including possible myristoylation sites to
mediate membrane association. Furthermore, identification of multiple putative
phosphorylation sites indicates a possible role in cell signalling cascades and a
mitochondrial targeting sequence at the amino terminus of both SLP-2 sequences
indicates mitochondrial localization.
Northern blot analysis demonstrated that SLP-2 was widely expressed, including heart,
brain, lung, liver, placenta, pancreas, skeletal muscle and kidney, with the highest
expression levels found in heart, liver and pancreas. Intracellularly, SLP-2 is found
predominantly in mitochondria 64, 65. This subcellular localization is dictated by a
mitochondrial targeting sequence at the amino terminus of SLP-2, which is cleaved upon
transport into the mitochondria 64, 65. The mitochondrial localization of SLP-2 has been
confirmed in several studies investigating the mitochondrial proteome in a variety of
tissues, including mouse liver, brain, heart and kidney, as well as human heart and T
cells66-69. Furthermore, confocal imaging studies have shown full length SLP-2 localized
within mitochondria and fusion of SLP-2 residues 1-50 to a green fluorescent protein
(GFP) tag resulted in mitochondrial localization of GFP, whereas deletion of SLP-2
residues 1-50 eliminated the mitochondrial localization64, 65. SLP-2 is tightly associated
with the inner mitochondrial membrane, facing the intermembrane space 64, 65.
In the initial report of SLP-2 identification, SLP-2 was found in a high molecular weight
complex 62. Subsequent studies have shown SLP-2 to complex with mitochondrial
resident proteins mitofusin 2 and prohibitins 1 and 2, although the function of these
associations is currently unknown 64, 65. Knockdown of SLP-2 leads to a decrease in
mitochondrial transmembrane potential as well as a decrease in protein levels of complex
I and IV of the respiratory chain and decreased prohibitin 1 and 2 64, 65. The function of
SLP-2 is currently unknown.

15
1.1.3 Prohibitins
The SPFH-domain-containing proteins prohibitin (PHB)-1 and -2 are mitochondrial
proteins associated with the inner mitochondrial membrane 36, 70, 71. Both proteins contain
a hydrophobic region at the amino-terminus, which may facilitate the association with the
inner membrane 37. PHB-1 and PHB-2 hetero-oligomerize to form a large ring-like
complex and are dependent on co-expression for stability, as knockdown of PHB-1
results in the loss of PHB-2 and vice versa 29, 37, 70, 72. As such, PHB-1 and PHB-2 have
similar expression patterns and are widely expressed, although the expression levels vary
between tissues. Although studies have pointed to many roles for prohibitins, the
function of these proteins is currently unknown. While prohibitins have been implicated
in cellular proliferation, their expression levels do not correlate with the proliferative
state of the tissue 33,70. Prohibitins have also been linked to aging, as some senescent
cells show decreased PHB levels compared to young cells 70. Furthermore, prohibitin
knockdown during embryogenesis results in embryonic arrest in C. elegans 73. Prohibitin
knockdown after embryogenesis results in shortened life span in wild type animals, but
mutants unable to undergo diapause were protected by prohibitin deletion 32. Although
this result seems contradictory, it has been proposed that the loss of prohibitins initiates
signalling cascades that change cellular metabolism for fat utilization. In this way, wild
type animals that fail to undergo diapause cannot utilize fat stores for energy production,
resulting in decreased survival, while prohibitin loss in diapause mutants lead to fat store
utilization and increased survival.
Prohibitins associate with the mitochondrial AAA (m-AAA) protease and knockdown of
prohibitin leads to increased protein degradation by the protease, leading to the
suggestion that the prohibitin complex acts as a chaperone to prevent degradation of
functional proteins 29, 37. Moreover, prohibitins have also been shown to play a role in the
cleavage of the mitochondrial protein optic atrophy 1 (OPA-1), which is involved in
maintaining cristae structure 74. Deletion of PHB-2 in mouse embryonic fibroblasts led to
massive mitochondrial fragmentation along with loss of cristae or the formation of very
large cristae 33. These cells were found to have altered processing of OPA-1 with a loss

16
of the long forms. Over-expression of a long-form OPA-1 mutant that is not cleavable
led to recovery of cristae structure and less fragmentation of the mitochondrial network.
The prohibitin knockdown cells also showed increased susceptibility to apoptotic
induction, which was rescued by the mutant OPA-1. This is likely due to the role for
OPA-1 and cristae remodeling as an early step in cytochrome c release leading to the
induction of the intrinsic pathway of apoptosis 75. In this way, the prohibitin complex
plays a role in the sensitivity to apoptotic induction.
The mechanism of action for the prohibitin complex is still not understood. However, it
has recently been proposed that prohibitins may facilitate membrane
compartmentalization at the inner mitochondrial membrane 34. As this membrane has
extremely high protein content, proper functioning of these proteins may benefit from
segregation, in a way reminiscent of that described for T cell activation. Yeast cells
lacking prohibitins remain viable, but to further understand the function of these proteins,
synthetic lethal mutations were created to identify the genomic interactome of prohibitins 35. The loss of a number of non-essential genes were shown to be synthetic lethal in
combination with the loss of prohibitins, including genes involved in the assembly of
respiratory chain components and in the control of mitochondrial morphology. Two new
genes were identified in this screen, one required for phosphatidylethanolamine
biosynthesis and the other for cardiolipin biosynthesis, two lipids found at the inner
mitochondrial membrane. Cardiolipin is unique to mitochondrial membranes, and as
such, has been shown to be important for various mitochondrial functions 76. Cardiolipin
interacts with many mitochondrial proteins, including components of the electron
transport chain and mitochondrial translocases, and this interaction is required for optimal
activity of these complexes 76-78. In addition, there is evidence that cardiolipin is required
for the maintenance of mitochondrial membrane structure, which is essential for
oxidative phosphorylation 79, 80. The ring-like prohibitin complex has been proposed to
organize cardiolipin, and perhaps other proteins, into domains to facilitate various
mitochondrial functions. In the absence of prohibitins, these domains would not be
formed and increased cardiolipin biosynthesis may be required to maintain proper
mitochondrial functions 34, 35. The importance of mitochondrial membrane organization
will be discussed in the context of the major mitochondrial functions, including

17
bioenergetics, calcium signalling and apoptosis and in the importance of these functions
for T cell activation and function.
1.2 Mitochondrial functions
1.2.1 Bioenergetics and metabolism
Energy production is the predominant function associated with mitochondria as the
electron transport chain resides at the inner membrane and is responsible for producing
the majority of cellular ATP. Mitochondria have separate inner and outer membranes,
each with a unique lipid and protein composition. The inner membrane is enriched in the
lipid cardiolipin, which has been linked to the organization of the membrane into folded
cristae. This, in turn, gives a much larger membrane surface area in the small organelle,
which allows for a higher concentration of the protein complexes of the electron transport
chain. Glycolysis and the tricarboxylic acid (TCA) cycle break down glucose into the
substrates oxidized by the electron transport chain. Glucose is first metabolized to two
pyruvate molecules, two ATP and two NADH (nicotinamide dinucleotide) molecules
during glycolysis. Acetyl groups from pyruvate are then transferred to coenyzme A
(CoA), with the generation of one NADH molecule and this marks the entry into the
TCA. The oxidation of fatty acids by β-oxidation also generates acetyl-CoA, which can
enter the TCA cycle to generate ATP from the respiratory chain, which occurs under
conditions of limiting glucose. The enzymes of the TCA cycle are located within the
mitochondrial matrix and this cycle gives rise to three NADH molecules, one FADH2
(flavin adenine dinucleotide) molecule and one ATP. The NADH and FADH2 molecules
carry high-energy electrons, which are used to drive ATP production through the electron
transport chain. The principle behind this relies on the generation of energy as electrons
are transferred from carriers with low reduction potential to donors with high reduction
potential 81. The electron transport chain transfers electrons between a series of electron
acceptors with successively higher reduction potential. The energy released by these
transfers is harnessed to shuttle hydrogen ions across the inner membrane, resulting in a
net negative charge in the matrix. This gives rise to a transmembrane potential and

18
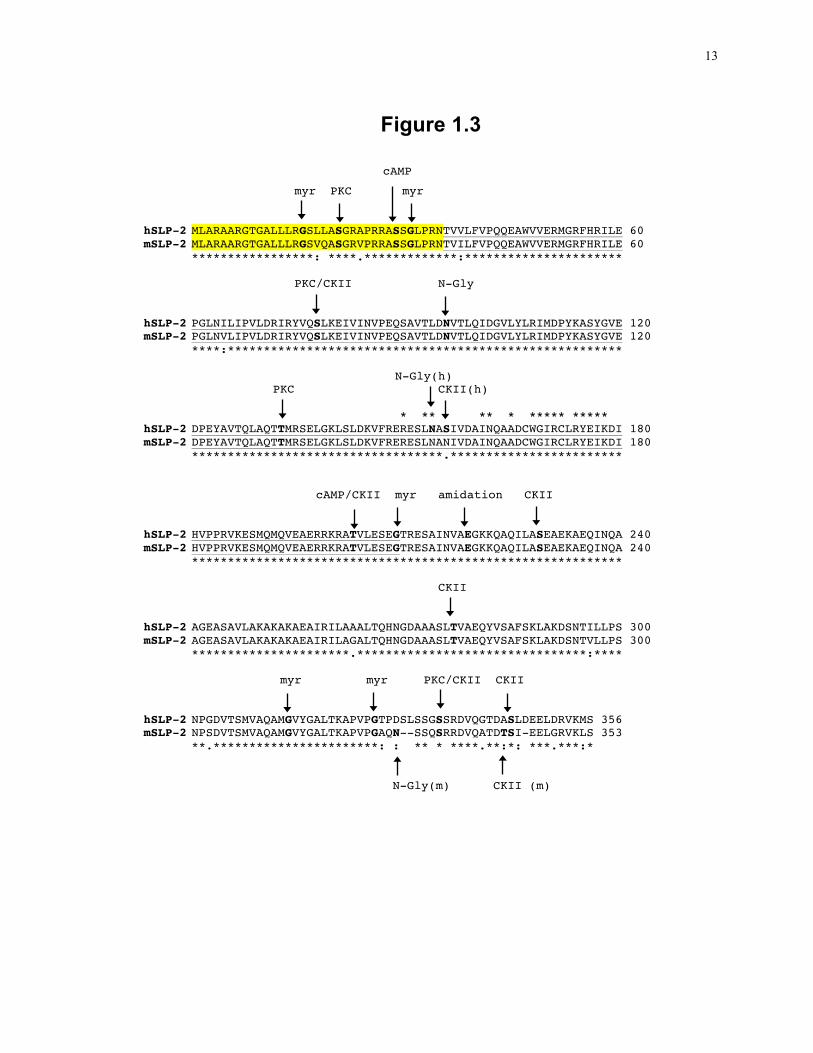
Figure 1.4: Schematic overview of the respiratory chain.
The components of the respiratory chain in association with the inner mitochondrial
membrane are shown along with the flow of electrons and hydrogen ions through the
complexes. NADH dehydrogenase and succinate dehydrogenase represent the entry
point for electrons into the respiratory chain at complex I and complex II, respectively.
Electrons are transferred through a series of electron acceptors associated with complex
III and IV and are ultimately transferred to oxygen to form water as an end product. At
complex I, III and IV, hydrogen ions are transported across the inner membrane into the
intermembrane space, resulting in a net negative charge in the mitochondrial matrix.
ATP synthase uses the energy released by hydrogen ions moving back into the matrix to
synthesis ATP from ADP. This figure is adapted from Rich and Marechal, 2010 81.
19
Figure 1.4
Intermembrane Space (+)
Inner Mitochondrial Membrane
Matrix (-)
NADH +H+
NAD+ Succinate Fumarate
UQ UQ
UQH2 UQH2
2H+ 2H+
Cytochrome c
½ O2 + 2H+
H2O ADP + Pi
ATP
4H+ 4H+ 2H+ nH+
Complex I NADH
Dehydrogenase
Complex II Succinate
Dehydrogenase
Complex III Cytochrome bc1
Complex IV Cytochrome c
Oxidase
Complex V ATP
Synthase

20
pH gradient across the inner membrane. The double membrane structure allows for two
distinct regions within the mitochondria, which allows this gradient to exist separately
from the cytosol. The transmembrane potential is exploited for ATP synthase, which
uses the energy released by transport of hydrogen ions along the potential gradient to
drive the phosphorylation of ADP to ATP.
The respiratory chain is composed of four molecular complexes located within the inner
mitochondrial membrane (Fig. 1.4). Complex I, NADH dehydrogenase, is a complex
formed by as many as 45 subunits and transfers electrons from NADH to a series of
cofactors resulting in the reduction of ubiquinone (UQ). Complex I is the largest of the
respiratory complexes and seven of the core hydrophobic subunits are encoded by the
mitochondrial DNA 81. Complex II, succinate dehydrogenase, is an enzyme of the TCA
cycle and converts succinate to fumarate with the transfer of two electrons to FAD to
form FADH2. FADH2 is then oxidized by the transfer of these electrons through a
number of cofactors to UQ. Complex I and II represent the entry point of electrons into
the respiratory chain. UQH2 is a mobile electron carrier that can freely diffuse in the
membrane to Complex III, cytochrome bc1, where the electrons are transferred from
UQH2 to a number of cofactors and then to cytochrome c. Cytochrome c is another
mobile electron carrier that diffuses through the inner membrane to Complex IV,
Cytochrome c oxidase, which again transfers electrons through a series of cofactors to the
final electron acceptor, O2 forming H2O, the final product of the respiratory chain. ATP
synthase forms a pore in the inner membrane through which H+ is transported back into
the matrix. The energy released drives the phosphorylation of ADP to ATP.
Complex I, III and IV span the inner membrane and use the energy released from electron
transfer to transport hydrogen ions across the inner membrane into the inter-membrane
space. Although these complexes are often described as separate complexes, recent work
has indicated that the complexes associate with one another, forming super complexes 82.
These complexes include I/III and II/III/IV and are quite prevalent as most complex I and
complex III in mouse liver mitochondria are found in super complexes. Some of these
complexes also contain the mobile electron carriers UQ and cytochrome c and together
this organization is thought to increase the efficiency of electron transfer between the

21
complexes, increasing the ATP production. To facilitate super complex formation and
efficient electron transfer, the components of the respiratory chain may be sequestered
into defined domains within the mitochondrial inner membrane. At the cell surface,
cholesterol is a major component involved in micro-domain formation but the cholesterol
content in the mitochondrial membrane is low. However, the mitochondrial lipid
cardiolipin may perform a similar role in membrane organization, as this lipid is
important for the proper function of the components of the electron transport chain and
super complexes are not stable in the absence of cardiolipin 77, 78, 83, 84. Furthermore,
cardiolipin has a unique structure that can alter membrane structure and curvature, which
may facilitate micro-domain formation and has been shown to form clusters in the
mitochondrial membrane 85, 86. Through the organization of the inner mitochondrial
membrane into cardiolipin-enriched domains, super complex formation may be favored,
leading to more efficient electron transport, giving rise to increased ATP production.
Studies altering the expression levels of SLP-2 indicate a role for SLP-2 in regulating the
respiratory chain. In cells treated with siRNA to knock down SLP-2 expression levels,
there is a slight decrease in the mitochondrial transmembrane potential and lower ATP
production 65, 87. Furthermore, SLP-2 expression is increased upon cellular stress, as is
ATP production, whereas knocking down SLP-2 expression before inducing cellular
stress inhibits this increase in ATP production 88. Thus, SLP-2 appears to play a role in
regulating the respiratory chain and ATP production. As SLP-2 has been shown to
interact with the prohibitin complex 34, 64, this regulation may be related to the role
proposed for prohibitins in the mitochondrial membrane compartmentalization and
respiratory chain function.
1.2.2 The metabolic regulation of T cells
The regulation of ATP production plays an important role in T cell activation as this is an
anabolic process requiring energy for cellular growth and function 89. As such, T cell
activation is tightly linked to metabolism, as co-stimulation of T cells through the CD28
receptor increases glycolysis while decreasing the requirement for ATP production

22
through the respiratory chain 90. Although there is much more ATP produced per glucose
molecule if the products of glycolysis enter the respiratory chain, these products can also
provide the building blocks for amino acid, nucleotide and lipid biosynthesis, all of which
are needed for cellular proliferation and function 91. Thus, under conditions of unlimited
glucose supply, T cells fulfill energy requirements as well as biosynthetic requirements
by relying on high throughput glycolysis.
The importance of metabolic regulation during T cell activation has recently been shown
in both CD4+ and CD8+ T cells. Upon CD28 stimulation, the PI3K/Akt pathway is
activated and leads to the activation of the mammalian target of rapamycin (mTOR), a
serine/threonine kinase involved in activating protein synthesis. The generation of T cell-
specific mTOR-deficient mice showed a failure to differentiate into TH1, TH2 or TH17
with increased differentiation into Tregs 92. Subsequently, an investigation into the
metabolic profiles of these cell types demonstrated that TH1, TH2 and TH17 cells all had
significantly higher glycolytic rates than Treg cells, while Treg cells had significantly
higher rates of β-oxidation, which relies on the respiratory chain for ATP production 93.
Treating cells with an inhibitor of β-oxidation led to a decrease in Treg cell counts, while
treatment with fatty acids to promote β-oxidation decreased TH1/2/17 cell numbers and
function and increased Treg function. Finally, treatment with the AMP-activated kinase
(AMPK) activator metformin led to a decrease in total CD4+ T cell populations while
increasing the Treg population. AMPK is activated in response to an increase in the ratio
of AMP to ATP and acts to inhibit protein synthesis through the inhibition of mTOR. It
also activates β-oxidation with the overall effect of decreasing ATP consumption 94.
The control of the mTOR pathway and β-oxidation has also been shown to play an
essential role in the development of CD8+ memory T cells 95, 96. Treatment of mice with
rapamycin, an inhibitor of mTOR, results in an increase in CD8+ T cell memory
responses to LCMV infection 96. Finally, TNF-receptor associated factor 6 (TRAF6)-
deficient mice with a defect in the expression of genes involved in β-oxidation were
found to have severely decreased CD8+ memory T cell formation 95. These mice also
showed low rates of β-oxidation and treatment with metformin or rapamycin to induce β-
oxidation led to a recovery of CD8+ memory T cells. Taken together, these results

23
demonstrate the importance of metabolic regulation in the generation of both CD4+ and
CD8+ T cell subsets, where glycolysis and β-oxidation play opposing roles in the T cell
subsets.
1.2.3 Calcium signalling
In addition to ATP synthesis, mitochondrial membrane potential also drives the uptake of
cytosolic calcium 97, 98. The mitochondrial outer membrane is relatively permeable to
ions, while the inner membrane is impermeable, an essential feature for the respiratory
chain. The inner membrane has a number of ion transporters, including a calcium
uniporter that transports calcium into the matrix 99. Calcium is exported from the matrix
by the sodium/calcium antiporter. The rate of Ca2+ uptake by the Ca2+ uniporter is
controlled by cytosolic calcium levels and transport is increased in response to high
cytosolic calcium, while the Na+/Ca2+ antiporter is regulated by free matrix Ca2+ levels.
In the matrix, calcium binds to phosphate and prevents the activation of the antiporter and
allows the mitochondria to sequester calcium from the cytosol in response to cytosolic
calcium spikes. Mitochondrial calcium activates a number of enzymes and leads to
increased ATP production, but prolonged elevated mitochondrial calcium levels lead to
apoptosis 100.
The ability of mitochondria to buffer calcium signals is of special importance in T cells
where stimulation triggers cytosolic Ca2+ elevations required for NFAT activation 101-103.
TCR signalling through the PLCγ pathway leads to the generation of IP3, which binds to
the IP3 receptor on the endoplasmic reticulum membrane to trigger the release of
intracellular Ca2+ stores. Upon depletion of the intracellular Ca2+ stores, the ER resident
protein stromal interaction molecule 1 (STIM1) then triggers the activation of the calcium
release-activated calcium (CRAC) channels at the cell surface, further increasing the
cytosolic Ca2+ levels 104, 105. Once activated, the CRAC channels are sensitive to
cytosolic calcium levels with elevated levels leading to channel inactivation 97,106. Thus,
in order to achieve elevated cytosolic Ca2+ levels, local Ca2+ levels must remain low.
Recent studies have demonstrated that mitochondrial calcium uptake plays an important

24
role in maintaining CRAC-dependent calcium uptake 107. Mitochondria were shown to
migrate to the plasma membrane upon thapsigargin-triggered ER calcium release and the
inhibition of this migration led to a rapid inactivation of CRAC channels. In addition, T
cell activation through the TCR triggers mitochondrial translocation to the
immunological synapse and the inhibition of this translocation led to decreased
intracellular Ca2+ levels 108. Thus, mitochondria are thought to act as a localized calcium
sink necessary to shuttle calcium away from CRAC channels to allow a sustained
calcium influx required for T cell activation. In this way, the organization of
mitochondrial membranes to facilitate supercomplex formation may also lead to
increased calcium buffering capacity as the increased transmembrane potential would
drive more calcium into the mitochondria, away from the CRAC channels, resulting in
prolonged calcium signalling.
To investigate the role of SLP-2 in mitochondrial calcium buffering Da Cruz et al treated
HeLa cells with shRNA to knock down SLP-2 levels 109. Cells expressing lower levels of
SLP-2 took up less calcium in response to histamine treatment and had a higher rate of
calcium efflux after stimulation. Treatment of these cells with an inhibitor of the
mitochondrial sodium/calcium exchanger normalized the calcium efflux rate between
control and knockdown cells, whereas the initial peak of calcium response remained
lower in the knockdown cells. This difference in calcium uptake was not due to a
difference in cellular calcium concentration as permeabilized cells also showed a
decreased mitochondrial calcium uptake and increased efflux rate in SLP-2 knockdown
cells. Likewise, in cells over-expressing SLP-2, the initial calcium uptake was slightly
higher compared to wild type cells and efflux rate was reduced, resulting in a prolonged
calcium signal. This role for SLP-2 in regulating mitochondrial calcium buffering
capacity is likely involved in the regulation of T cell responses, as Jurkat T cell activation
level is directly proportional to SLP-2 expression level 63.

25
1.2.4 Apoptosis
Mitochondria are not only involved in T cell activation but also in the regulation of T cell
responses by inducing cell death. Apoptosis is a form of controlled cell death induced in
response to extracellular signals or cellular stress, such as DNA damage or growth factor
withdrawal 110. The activation of the apoptotic pathway leads to chromatin condensation
and DNA fragmentation, cell shrinkage and membrane blebbing, ultimately reducing
cells to small apoptotic bodies that can be engulfed by macrophages to prevent
inflammatory responses. Apoptosis is essential for multi-cellular organisms to eliminate
damaged or unneeded cells. In T cell development, the failure of cells to succeed through
pre-TCR β selection, positive or negative selection leads to apoptosis and elimination of
TCR-defective or auto-reactive T cells 111. Moreover, apoptosis is also required after a
pathogen has been cleared by the immune response. In this case, T cells specific for the
pathogen have undergone massive proliferation and activation. While some will survive
as memory cells, most cells will undergo activation-induced cell death. In this way, the
body returns to homeostasis and reduces possible immunopathological effects of
activated T cells.
The main effectors of apoptosis are the caspases, a family of asparagine-proteases that
cleave a wide variety of structural and regulatory proteins 112. Capsases are composed of
two main subfamilies, in which capase 1 subfamily members are involved in pro-
inflammatory responses through the cleavage and activation of IL-1 and IL-18, while
caspase 3 subfamily members, including caspase 3 and 6-10 are involved in apoptosis 113,
114. Caspases exist as zymogens in healthy cells, but apoptotic stimuli induce signalling
cascades that result in procaspase homo-oligomerization, auto-cleavage and activation.
The activated caspases then cleave and activate additional caspases, leading to wide
spread protein degradation including complex I subunit NDUFS1, which in turn, leads to
the loss of the transmembrane potential. Caspases also activate a number of additional
apoptotic effectors, including DNA fragmentation factor 45 and pro-apoptotic kinases
involved in membrane blebbing and apoptotic body formation 110.
Mitochondria are important regulators of apoptosis and represent the convergence of both
extrinsic and intrinsic apoptosis pathways 110. The extrinsic pathway is activated by

26
extracellular signals, in response to ligation of cell surface death receptors, such as Fas or
TNF related apoptosis-inducing ligand (TRAIL), which induce a signal cascade resulting
in the activation of caspase 8 115. The intrinsic pathway is activated by intracellular
signals that result in the release of cytochrome c and direct IAP binding protein with low
pI (DIABLO) from the mitochondrial inner membrane space, allowing the formation of
the apoptosome, composed of Apaf-1, cytochrome c and procaspase 9 115.
Oligomerization of these components result in the activation of caspase 9, another
activator that initiates the caspase cascade 110. DIABLO inhibits the inhibitor of
apoptosis proteins (IAPs), which can bind to and inactivate various caspases 116.
The B cell lymphoma 2 (Bcl-2) family of proteins is responsible for sensing cellular
stress signals to induce the activation of the intrinsic pathway 116. This family consists of
both pro-apoptotic and anti-apoptotic family members, and cell signalling in normal
conditions or under cell stress tips the balance in favor of pro- or anti-apoptotic
responses. Under stress conditions, the pro-apoptotic members Bak and Bax oligomerize
and insert into the mitochondrial outer membrane, forming a pore that causes cytochrome
c release. There are currently two models explaining the control of Bax and Bak in the
transition from healthy to apoptotic conditions. In the activation model, Bax and Bak are
thought to be inactive in healthy cells, but apoptotic signals lead to the activation of pro-
apoptotic family members BIM, tBID and PUMA, which in turn, activate Bax and Bak
and lead to membrane insertion and cytochrome c release. In the neutralizing model, Bax
and Bak are thought to be constitutively active, but prevented from pore formation by the
association with the anti-apoptotic family members, including Bcl-2. Apoptotic signals
upregulate or release multiple pro-apoptotic family members from confined cellular
locations, and allow these proteins to bind the anti-apoptotic members, releasing Bax and
Bak from Bcl-2, to permit pore formation. Mitochondria link the extrinsic pathway to the
induction of the intrinsic pathway via caspase 8, as one target of caspase 8 is the pro-
apoptotic protein Bid, which upon cleavage, associates with the mitochondrial membrane
to activate Bax/Bak pore formation to trigger cytochrome c release 110, 117. In this way,
mitochondria play a role in both intrinsic and extrinsic apoptosis.

27
Apoptosis is also tightly coupled to changes in mitochondrial morphology. Although
mitochondria have traditionally been thought of as isolated organelles, they have been
shown to be a highly dynamic population undergoing fusion and fission events, which
affect a wide variety of cellular functions 118-120. Mitochondrial fusion is mediated by the
GTPases mitofusin (Mfn) 1 and 2 and OPA-1. Mfn1 and 2 facilitate outer membrane
fusion by undergoing homo- or hetero-oligomerizion between adjacent mitochondria.
The intermembrane space resident OPA-1 facilitates inner membrane fusion, although the
mechanism is currently unclear 121. Fission is carried out by the GTPase Drp1, which
oligomerizes into a ring-like structure around the mitochondrial outer membrane at the
site of the fission event. Drp1 is a cytosolic protein with a small pool localized to the
mitochondrial membrane in a punctate pattern. The recruitment of Drp1 is thought to be
facilitated by the outer membrane protein Fis1, although the outer membrane protein
mitochondrial fission factor (Mff) has also been proposed to recruit Drp1 to fission sites 122-124. The importance of mitochondrial dynamics has been shown in development as
Drp1, Mfn1, Mfn2 and OPA-1 deletions are all embryonic lethal 125-128. Furthermore,
fusion is important for content sharing of the mitochondrial network, including the
replacement of damaged mitochondrial DNA (mtDNA). Indeed, in fusion-deficient cells,
mtDNA is lost 129. Finally, fusion and fission events are also regulated during the cell
cycle, with hyperfusion occurring as cells transition from G1 to S phase, while the mitotic
kinase Cdk1/cyclin B phosphorylates Drp1 to promote fission 130, 131. In this way,
contents of the mitochondrial network are shared to replace damaged components before
splitting and dividing into the daughter cells.
Changes in the expression levels of the fusion or fission components have been shown to
play a role in apoptosis. Mitochondrial fission, facilitated by Drp1 recruitment, is an
early apoptotic event 132, 133. Inhibition of Fis1 or Drp1 or over-expression of OPA-1
inhibits fission, which is protective against apoptosis 133, 134. In addition, the loss of
transmembrane potential that accompanies apoptosis also results in elevated OPA-1
cleavage, which then inactivates OPA-1 and decreases mitochondrial fusion, leading to
an imbalance of mitochondrial dynamics in favor of mitochondrial fission 74. The
components of fusion and fission machinery also interact with members of the Bcl-2
family of apoptotic proteins to regulate apoptosis. The pro-apoptotic Bax co-localizes

28
with Drp1 and Mfn2 upon apoptotic induction and the knockdown of Fis1 inhibits the
recruitment, whereas knockdown of both Drp1 and Fis1 inhibit mitochondrial fission to
protect against apoptosis 135, 136.
SLP-2 may play a role in regulating mitochondrial fusion upon cellular stress to protect
against apoptotic induction 88. Mitochondrial stress induced by chloramphenicol
treatment led to the specific upregulation of SLP-2 64. Interestingly, mouse embryonic
fibroblasts (MEF) subjected to stress stimuli such as UV irradiation, actinomycin D or
cycloheximide show mitochondrial hyperfusion, resulting in elongated mitochondria 88.
Deletion of OPA-1 and Mfn1, both required for fusion, abrogated this response, as did
SLP-2 knockdown. This loss of hyperfusion appears to be due to altered OPA-1
processing as SLP-2 deficient cells lack the longer isoforms of OPA-1. Transfecting cells
with a non-cleavable mutant OPA-1 protein restored mitochondrial hyperfusion.
Treatment of SLP-2 knockdown cells with a metalloprotease inhibitor prevented the loss
of long form OPA-1, indicating a possible role for SLP-2 in protease regulation. This is
supported by an earlier study in which metalloprotease inhibition restored prohibitin and
respiratory chain component loss upon SLP-2 knockdown 64, 88. Although prohibitins
control OPA-1 processing 33 and are known to associate with SLP-2, cells lacking
prohibitin 1 and 2 show similar hyperfusion to wild type cells upon cycloheximide
treatment 88 indicating these hyperfusion events are independent of the prohibitin-SLP-2
complex. Upon hyperfusion, cells have increased ATP production compared to untreated
cells, while mutant cells unable to undergo hyperfusion (OPA-1-/- or shSLP-2) do not
show increased ATP production. Hyperfusion appears to be protective against apoptosis
as cells lacking Mfn1 or SLP-2 had increased apoptosis in response to actinomycin D
treatment or UV irradiation.
Membrane organization in the mitochondria may also play a role in apoptosis. As
discussed above, there are many players involved in the induction of apoptosis, including
the pro- and anti-apoptotic members of the Bcl family, the members of the caspase
cascade as well as the proteins involved in mitochondrial membrane dynamics 110, 116, 133.
The organization of the mitochondrial membrane into specialized microdomains would
facilitate the interaction of these components to induce apoptosis, leading to membrane

29
permeabilization, cytochrome c release and caspase activation in response to apoptotic
signals. Apoptotic induction via anti-Fas treatment led to a re-organization of pro-
apoptotic Bax and truncated Bid (tBid) from detergent-soluble to detergent-insoluble
domains isolated from whole cells 137. These results demonstrate the re-organization of
apoptotic components into membrane microdomains during apoptotic responses. Further
characterization of these mitochondrial detergent-insoluble membranes demonstrated an
enrichment in cardiolipin whereby all cardiolipin was found in the detergent-insoluble
domains and none in the detergent-soluble domains 138.
The enrichment of cardiolipin in these detergent-insoluble domains likely plays an
important functional role in the induction of apoptosis as cardiolipin can bind to many
components of the apoptotic pathway, including cytochrome c, tBid and caspase 8 139-142.
The pro-apoptotic protein tBid may also play a role in the organization of membranes
into cardiolipin-enriched detergent-insoluble domains as tBid binds to cardiolipin
monolayers and induces re-organization of the cardiolipin into regions of very tightly
packed lipid 143. This binding of tBid to cardiolipin is an essential step in apoptotic
induction as incubation of recombinant tBid with isolated mitochondria leads to
cytochrome c release, yet the addition of excess free cardiolipin prevents this binding and
leads to a decrease in cytochrome c release 141. Furthermore, apoptotic induction results
in the accumulation of many of the proteins known to associate with cardiolipin into
these cardiolipin-enriched domains, including activated caspase 8, Bak, tBid and DLP1 138, 144. Interestingly, the fission protein hFis1 and the fusion protein OPA-1 are
constitutively localized to the cardiolipin-enriched membrane domains, but DLP1 is
recruited upon apoptotic induction, indicating the coordination of apoptosis and
mitochondrial fission 144. The importance of these domains is demonstrated in cells
lacking cardiolipin, either by knockdown of cardiolipin synthase or tafazzin, the
remodeling enzyme involved in the final steps of cardiolipin synthesis, as these cells
exhibit an increased resistance to apoptotic induction 142, 145.

30
1.3 Rationale and Hypothesis
1.3.1 Rationale
In light of the important role played by cell membrane organization at both the plasma
and mitochondrial membranes, initial experiments were performed to identify novel
players involved in membrane organization during T cell activation. To this end, Jurkat
T cells were activated through their T cell receptor and DI-GEMs were isolated. Proteins
were extracted from these fractions and subjected to mass spectrometry 63. In addition to
proteins known to be involved in T cell activation, previous work in the Madrenas
laboratory also identified a novel protein, stomatin-like protein 2 (SLP-2), which is a
member of the highly conserved stomatin family. As discussed above, members of this
family have been proposed to facilitate membrane microdomain formation and
organization, 55-57 although the function of SLP-2 remains unclear.
Preliminary experiments were performed to investigate the role of SLP-2 in the immune
response in general and in T cell activation in particular. Immunohistochemistry and
immunoblot analysis demonstrated SLP-2 expression in a number of human lymphoid
tissues, including lymph nodes, thymus, and tonsils, as well as in peripheral blood
mononuclear cells, including T cells, B cells and monocytes 63. SLP-2 expression was
also detected in areas of lymphocyte activation, such as germinal centers. Although the
expression of SLP-2 was low in naïve T cells, stimulation of these cells with
Staphylococcal enterotoxin E (SEE) increased the expression of SLP-2, pointing to a role
during T cell activation. Furthermore, SLP-2 was also most highly expressed in activated
and memory B cells, with much lower expression in naïve B cells, demonstrating a
widespread role for SLP-2 during adaptive immune responses.
To investigate the role of SLP-2 in membrane organization, DI-GEM fractions were
collected from both Jurkat T cells and primed peripheral blood mononuclear cells at
various time points during T cell stimulation with SEE and analyzed for SLP-2 levels.
Under resting conditions, a small pool of SLP-2 was associated with the detergent-
insoluble fraction and SEE stimulation increased the recruitment of SLP-2 to these
domains in a time-dependent manner, peaking at five minutes after activation. These

31
microdomains are likely important for T cell activation as membrane organization has
long been hypothesized to play an important role in T cell signalling 1, 2 and as such, the
association of SLP-2 with T cell signalling components was investigated. SLP-2 was
found to be associated with a number of T cell signalosome components, including
CD3ε, Lck, ZAP-70, LAT and PLC-γ1. SLP-2 was also shown to associate with actin,
another important player in membrane domain organization 20, and the inhibition of actin
polymerization decreased T cell activation upon SEE stimulation. The functional
importance of these associations was demonstrated by SLP-2 siRNA knockdown studies,
in which SLP-2 depletion resulted in decreased duration of ERK and PLC-γ1
phosphorylation, indicative of a shortened duration of T cell signalling. This decrease in
T cell signalling upon SLP-2 depletion led to decreased T cell activation, shown by lower
IL-2 production in response to SEE stimulation. The loss of SLP-2 had a greater impact
on primed T cells, whereas knockdown of SLP-2 in naïve T cells had a negligible effect
on activation, as these cells have very low SLP-2 expression. However, in primed cells,
which have upregulated SLP-2, the loss of SLP-2 led to a significant decrease in T cell
activation. Furthermore, increasing SLP-2 expression by transfection of Jurkat T cells
with a doxycycline-inducible SLP-2-GFP construct resulted in increased T cell activation
in response to SEE stimulation. Taken together, these results point to an important role
for SLP-2 in T cell activation, potentially through the organization of membrane domains
into specialized regions for T cell signalling.
1.3.2 Hypothesis
These preliminary data document a role for SLP-2 in T cell signalling and activation as
both over-expression and knockdown of SLP-2 affects T cell responses. The importance
of SLP-2 during T cell activation is further supported by the upregulation of SLP-2
expression in response to T cell stimulation. However, this work focused primarily on
events at the plasma membrane, yet SLP-2 is found predominantly at the mitochondrial
inner membrane 64, 65. In line with the proposed roles of other stomatin and SPFH family
members in membrane organization, and given the mitochondrial localization of SLP-2, I
hypothesize that SLP-2 acts to facilitate mitochondrial and plasma membrane

32
organization in response to T cell activation. In order to address this hypothesis, I
investigated the function of the mitochondrial pool of SLP-2 by examining mitochondrial
membrane composition as well as mitochondrial biogenesis and function in response to
SLP-2 over-expression. I also generated T cell-specific SLP-2 conditional knockout mice
and examined the activation and effector functions of SLP-2-deficient T cells to further
characterize the role of SLP-2 in mitochondrial membrane organization and function as
well as T cell activation. Finally, although the majority of SLP-2 is localized to the
mitochondria, there appears to be a small pool at the plasma membrane, as indicated by
the association of SLP-2 with the T cell signalosome. This plasma membrane pool of
SLP-2 thus was further investigated in resting T cells and in response to T cell
stimulation and the re-organization of both surface and mitochondrial SLP-2 pools in
response to T cell stimulation was be examined.
1.3.3 Specific Aim 1: To characterize the function of the mitochondrial pool of SLP-2
To further investigate the function of the mitochondrial pool of SLP-2, Jurkat T cells
were stably transfected with a doxycycline-inducible SLP-2 construct, tagged with GFP
at the C-terminus. When SLP-2 expression was induced in these cells, there was an
increase in mitochondrial populations, as shown by increased staining with the
mitochondrial-specific dye MitoTracker Red measured by both flow cytometry and
confocal microscopy. Furthermore, these cells also had increased mitochondrial counts
per cell slice as shown by transmission electron microscopy. To further investigate this
increase, the pathways required for mitochondrial biogenesis, including nuclear
transcription of mitochondrial-targeted genes, replication of mitochondrial DNA and lipid
biosynthesis were examined. SLP-2 over-expressing cells were found to have increased
PGC-1α transcript levels, indicative of activated nuclear transcription programs. In
addition, mitochondrial DNA levels were also elevated in the SLP-2 over-expressing
cells, as was cardiolipin biosynthesis. Taken together, these results point to an increase
in mitochondrial biogenesis in response to SLP-2 over-expression.

33
The mechanism of SLP-2-induced mitochondrial biogenesis is currently unclear.
However, it was hypothesized that SLP-2 organizes the mitochondrial membrane into
specialized domains to control mitochondrial function and biogenesis. As SLP-2 was
shown to interact with both the prohibitin complex and with cardiolipin in a dose-
dependent manner, we propose that SLP-2 may act as a bridge to facilitate the association
of the prohibitin complex with the mitochondrial inner membrane. In support of this
model, over-expression of SLP-2 increased the recruitment of prohibitins to
mitochondrial membranes. SLP-2 over-expressing cells also demonstrated increased
enzymatic activities of components of the citric acid cycle and the electron transport
chain, including citrate synthase, NADH dehydrogenase and succinate dehydrogenase.
As a result, SLP-2 over-expressing cells had increased ATP production and oxygen
consumption, indicative of enhanced activity of the respiratory chain. Furthermore, SLP-
2 over-expression was protective against induction of the intrinsic apoptotic pathway.
Taken together, we propose that SLP-2 facilitates mitochondrial membrane organization
through association with the prohibitin complex, which in turn, increases mitochondrial
biogenesis and function.
1.3.4 Specific Aim 2: To generate and characterize a T cell-specific SLP-2 conditional knockout mouse
To further investigate the function of SLP-2 in vivo, conventional genome-wide SLP-2
knockout mice were generated (C. Lemke and J. Madrenas, unpublished results).
However, the loss of SLP-2 expression resulted in embryonic lethality at the pre-
implantation stage, indicating an essential role for SLP-2 in early embryonic
development. Therefore, a conditional SLP-2 knockout was created using the Cre-lox
system. As preliminary work focused on the role of SLP-2 in T cells, mice expressing
Cre-recombinase under the control of the CD4 promoter were utilized to initiate genomic
deletion at the double-positive stage of T cell development. This resulted in mice with
genomic SLP-2 deletion in all double positive αβ T cells. Thus, all mature peripheral
CD4+ and CD8+ T cells lack expression of SLP-2 and the mice are termed SLP-2 T-K/O.

34
SLP-2 T-K/O mice showed grossly normal T cell development compared to control mice.
The total number of thymocytes, splenocytes and lymph node cells were similar between
control and SLP-2 T-K/O mice, as was double negative, double positive and single
positive thymic populations, as well as non T cells and both CD4+ and CD8+ T cells in
the spleen and lymph nodes. Confirming earlier results with human T cells, murine SLP-
2 T-K/O cells also showed decreased T cell activation ex vivo in response to a number of
different T cell stimuli, with the defect being more pronounced in primed SLP-2 T-K/O
cells. This defect in T cell activation was shown to be a post-transcriptional block in IL-2
production. However, analysis of the hallmark cytokines showed normal differentiation
into TH1, TH2, and TH17 subsets upon SEE stimulation in both naïve and primed SLP-2
T-K/O cells. Analysis of FoxP3+ staining also showed similar Treg populations in both
control and SLP-2 T-K/O mice.
In demonstrating the biological significance of these ex vivo studies, SLP-2 T-K/O mice
exhibited impaired responses to in vivo T cell challenge. SLP-2 T-K/O mice showed
decreased recall responses to OVA immunization and were also slower to reject an
allograft from a non-matched donor. In agreement with the decrease in T cell responses,
the SLP-2 T-K/O mice also showed a small but significant reduction in CD4+ T cell
populations, which did not appear to be due to apoptosis. The decreased T cell activation
may lead to less T cell proliferation in vivo, resulting in lower T cell populations and
decreased T cell responses.
As previous work with SLP-2 over-expression in the Jurkat system indicated a role for
SLP-2 in membrane organization, detergent-resistant glycolipid-enriched microdomains
were investigated in SLP-2 T-K/O mice. Although there was no significant difference in
total cardiolipin levels in these mice, there was significantly less cardiolipin in the
detergent-insoluble mitochondrial membrane fractions, accompanied by a decrease in
associated prohibitin 1, although total prohibitin levels were similar to control cells. In
addition, a number of subunits of complex I of the respiratory chain were also found to be
decreased in SLP-2 T-K/O cells, including the NDU FA9, NDU FS3 and NDU FB8
subunits, while there was no change in the levels of the other complexes of the
respiratory chain. The SLP-2 T-K/O cells also had a significant reduction in the activity

35
of complex I, suggesting a functional outcome of the loss of complex I subunits. The
defect in respiratory chain function was extended to complex II + III, which also showed
decreased activity in SLP-2 T-K/O cells. Together, these results point to an altered
mitochondrial membrane organization, which may lead to decreased complex I assembly
and increased degradation of complex I subunits, ultimately resulting in decreased
complex I activity and decreased T cell responses in SLP-2-deficient T cells.
1.3.5 Specific Aim 3: To characterize the plasma membrane-associated pool of SLP-2
Although SLP-2 has been shown to localize predominantly to the mitochondria, it has
also been detected in detergent-insoluble glycosphingolipid-enriched membrane fractions
isolated from whole T cells, which include many surface proteins. To confirm the
existence of a second pool of SLP-2 at the plasma membrane and to further investigate
the function of this pool, Jurkat T cells stably transfected with doxycycline-inducible
SLP-2-GFP were utilized for SLP-2 over-expression and confocal imaging studies. In
addition to full length SLP-2-GFP, a deletion construct lacking the amino-terminal
region, which includes the mitochondrial targeting sequence and the putative
myristoylation sites, was also examined.
In agreement with the results found in specific aim 1, both endogenous SLP-2 and SLP-
2-GFP localized to the mitochondria. As expected, the truncation mutant lacking the
mitochondrial-targeting domain failed to localize to mitochondria and instead showed a
diffuse cytosolic localization. Analysis of the plasma membrane-associated pool
demonstrated the presence of both full length and the truncation mutant of SLP-2,
although higher expression levels of the truncation mutant were required for detection of
the plasma membrane-associated pool. This may indicate a role for the amino terminal
region in membrane association. In addition to human T cells, both the mitochondrial
and surface pool of SLP-2 were also found in primary mouse T cells, indicating a
conservation of two distinct localizations for SLP-2 across species.

36
In agreement with earlier results showing recruitment of SLP-2 to detergent-insoluble
membranes upon T cell activation, confocal microscopy demonstrated the recruitment of
SLP-2 to the immunological synapse upon T cell activation. Furthermore, this pool of
SLP-2 was actively maintained at the synapse, indicating a role for SLP-2 in T cell
activation. To better visualize the cell surface pool of SLP-2 during T cell activation,
total internal reflection fluorescence microscopy was used to limit the detection to a
depth of 200nm from the cell surface. These images showed the surface pool of SLP-2
surrounding TCR clusters at the immunological synapse, with SLP-2 being weakly
excluded from the TCR clusters. The ability of SLP-2 to self-associate may allow SLP-2
to organize the T cell membrane to facilitate TCR microcluster formation and to prevent
signal complex dissociation.
Taken together, the investigation of SLP-2 function using both human and mouse
systems, in both over-expression and knockout studies, indicates a role for SLP-2 in
membrane organization, predominantly of the mitochondrial membranes, but also for the
plasma membrane.

37
1.4 References
1. Simons, K. & Gerl, M.J. Revitalizing membrane rafts: new tools and insights. Nat Rev Mol Cell Biol 11, 688-699 (2010).
2. Lingwood, D. & Simons, K. Lipid rafts as a membrane-organizing principle. Science 327, 46-50 (2010).
3. Pitcher, L.A., Young, J.A., Mathis, M.A., Wrage, P.C., Bartok, B. & van Oers, N.S. The formation and functions of the 21- and 23-kDa tyrosine-phosphorylated TCR zeta subunits. Immunol Rev 191, 47-61 (2003).
4. Alarcon, B., Gil, D., Delgado, P. & Schamel, W.W. Initiation of TCR signaling: regulation within CD3 dimers. Immunol Rev 191, 38-46 (2003).
5. Smith-Garvin, J.E., Koretzky, G.A. & Jordan, M.S. T cell activation. Annu Rev Immunol 27, 591-619 (2009).
6. Love, P.E. & Hayes, S.M. ITAM-mediated signaling by the T-cell antigen receptor. Cold Spring Harb Perspect Biol 2, a002485 (2010).
7. Salmond, R.J., Filby, A., Qureshi, I., Caserta, S. & Zamoyska, R. T-cell receptor proximal signaling via the Src-family kinases, Lck and Fyn, influences T-cell activation, differentiation, and tolerance. Immunol Rev 228, 9-22 (2009).
8. Wange, R.L. LAT, the linker for activation of T cells: a bridge between T cell-specific and general signaling pathways. Sci STKE 2000, re1 (2000).
9. Hogan, P.G., Chen, L., Nardone, J. & Rao, A. Transcriptional regulation by calcium, calcineurin, and NFAT. Genes Dev 17, 2205-2232 (2003).
10. Lin, J., Miller, M.J. & Shaw, A.S. The c-SMAC: sorting it all out (or in). J Cell Biol 170, 177-182 (2005).
11. Dustin, M.L., Chakraborty, A.K. & Shaw, A.S. Understanding the structure and function of the immunological synapse. Cold Spring Harb Perspect Biol 2, a002311 (2010).
12. Monks, C.R., Freiberg, B.A., Kupfer, H., Sciaky, N. & Kupfer, A. Three-dimensional segregation of supramolecular activation clusters in T cells. Nature 395, 82-86 (1998).
13. Varma, R., Campi, G., Yokosuka, T., Saito, T. & Dustin, M.L. T cell receptor-proximal signals are sustained in peripheral microclusters and terminated in the central supramolecular activation cluster. Immunity 25, 117-127 (2006).

38
14. Fooksman, D.R., Vardhana, S., Vasiliver-Shamis, G., Liese, J., Blair, D.A., Waite, J., Sacristan, C., Victora, G.D., Zanin-Zhorov, A. & Dustin, M.L. Functional anatomy of T cell activation and synapse formation. Annu Rev Immunol 28, 79-105 (2010).
15. Xavier, R., Brennan, T., Li, Q., McCormack, C. & Seed, B. Membrane compartmentation is required for efficient T cell activation. Immunity 8, 723-732 (1998).
16. Janes, P.W., Ley, S.C. & Magee, A.I. Aggregation of lipid rafts accompanies signaling via the T cell antigen receptor. J Cell Biol 147, 447-461 (1999).
17. Viola, A., Schroeder, S., Sakakibara, Y. & Lanzavecchia, A. T lymphocyte costimulation mediated by reorganization of membrane microdomains. Science 283, 680-682 (1999).
18. Webb, Y., Hermida-Matsumoto, L. & Resh, M.D. Inhibition of protein palmitoylation, raft localization, and T cell signaling by 2-bromopalmitate and polyunsaturated fatty acids. J Biol Chem 275, 261-270 (2000).
19. Zhang, W., Irvin, B.J., Trible, R.P., Abraham, R.T. & Samelson, L.E. Functional analysis of LAT in TCR-mediated signaling pathways using a LAT-deficient Jurkat cell line. Int Immunol 11, 943-950 (1999).
20. Chichili, G.R. & Rodgers, W. Clustering of membrane raft proteins by the actin cytoskeleton. J Biol Chem 282, 36682-36691 (2007).
21. Kabouridis, P.S., Janzen, J., Magee, A.L. & Ley, S.C. Cholesterol depletion disrupts lipid rafts and modulates the activity of multiple signaling pathways in T lymphocytes. Eur J Immunol 30, 954-963 (2000).
22. Jin, Z.X., Huang, C.R., Dong, L., Goda, S., Kawanami, T., Sawaki, T., Sakai, T., Tong, X.P., Masaki, Y., Fukushima, T., Tanaka, M., Mimori, T., Tojo, H., Bloom, E.T., Okazaki, T. & Umehara, H. Impaired TCR signaling through dysfunction of lipid rafts in sphingomyelin synthase 1 (SMS1)-knockdown T cells. Int Immunol 20, 1427-1437 (2008).
23. Zhang, M., Moran, M., Round, J., Low, T.A., Patel, V.P., Tomassian, T., Hernandez, J.D. & Miceli, M.C. CD45 signals outside of lipid rafts to promote ERK activation, synaptic raft clustering, and IL-2 production. J Immunol 174, 1479-1490 (2005).
24. Su, M.W., Yu, C.L., Burakoff, S.J. & Jin, Y.J. Targeting Src homology 2 domain-containing tyrosine phosphatase (SHP-1) into lipid rafts inhibits CD3-induced T cell activation. J Immunol 166, 3975-3982 (2001).
25. Dart, C. Lipid microdomains and the regulation of ion channel function. J Physiol 588, 3169-3178 (2010).

39
26. Huber, T.B., Schermer, B., Muller, R.U., Hohne, M., Bartram, M., Calixto, A., Hagmann, H., Reinhardt, C., Koos, F., Kunzelmann, K., Shirokova, E., Krautwurst, D., Harteneck, C., Simons, M., Pavenstadt, H., Kerjaschki, D., Thiele, C., Walz, G., Chalfie, M. & Benzing, T. Podocin and MEC-2 bind cholesterol to regulate the activity of associated ion channels. Proc Natl Acad Sci U S A 103, 17079-17086 (2006).
27. Browman, D.T., Hoegg, M.B. & Robbins, S.M. The SPFH domain-containing proteins: more than lipid raft markers. Trends Cell Biol 17, 394-402 (2007).
28. Langer, T., Kaser, M., Klanner, C. & Leonhard, K. AAA proteases of mitochondria: quality control of membrane proteins and regulatory functions during mitochondrial biogenesis. Biochem Soc Trans 29, 431-436 (2001).
29. Nijtmans, L.G., de Jong, L., Artal Sanz, M., Coates, P.J., Berden, J.A., Back, J.W., Muijsers, A.O., van der Spek, H. & Grivell, L.A. Prohibitins act as a membrane-bound chaperone for the stabilization of mitochondrial proteins. EMBO J 19, 2444-2451 (2000).
30. Kasashima, K., Sumitani, M., Satoh, M. & Endo, H. Human prohibitin 1 maintains the organization and stability of the mitochondrial nucleoids. Exp Cell Res 314, 988-996 (2008).
31. Coates, P.J., Jamieson, D.J., Smart, K., Prescott, A.R. & Hall, P.A. The prohibitin family of mitochondrial proteins regulate replicative lifespan. Curr Biol 7, 607-610 (1997).
32. Artal-Sanz, M. & Tavernarakis, N. Prohibitin couples diapause signalling to mitochondrial metabolism during ageing in C. elegans. Nature 461, 793-797 (2009).
33. Merkwirth, C., Dargazanli, S., Tatsuta, T., Geimer, S., Lower, B., Wunderlich, F.T., von Kleist-Retzow, J.C., Waisman, A., Westermann, B. & Langer, T. Prohibitins control cell proliferation and apoptosis by regulating OPA1-dependent cristae morphogenesis in mitochondria. Genes Dev 22, 476-488 (2008).
34. Osman, C., Merkwirth, C. & Langer, T. Prohibitins and the functional compartmentalization of mitochondrial membranes. J Cell Sci 122, 3823-3830 (2009).
35. Osman, C., Haag, M., Potting, C., Rodenfels, J., Dip, P.V., Wieland, F.T., Brugger, B., Westermann, B. & Langer, T. The genetic interactome of prohibitins: coordinated control of cardiolipin and phosphatidylethanolamine by conserved regulators in mitochondria. J Cell Biol 184, 583-596 (2009).
36. Berger, K.H. & Yaffe, M.P. Prohibitin family members interact genetically with mitochondrial inheritance components in Saccharomyces cerevisiae. Mol Cell Biol 18, 4043-4052 (1998).

40
37. Steglich, G., Neupert, W. & Langer, T. Prohibitins regulate membrane protein degradation by the m-AAA protease in mitochondria. Mol Cell Biol 19, 3435-3442 (1999).
38. Morrow, I.C. & Parton, R.G. Flotillins and the PHB domain protein family: rafts, worms and anaesthetics. Traffic 6, 725-740 (2005).
39. Langhorst, M.F., Reuter, A. & Stuermer, C.A. Scaffolding microdomains and beyond: the function of reggie/flotillin proteins. Cell Mol Life Sci 62, 2228-2240 (2005).
40. Kihara, A., Akiyama, Y. & Ito, K. A protease complex in the Escherichia coli plasma membrane: HflKC (HflA) forms a complex with FtsH (HflB), regulating its proteolytic activity against SecY. EMBO J 15, 6122-6131 (1996).
41. Green, J.B. & Young, J.P. Slipins: ancient origin, duplication and diversification of the stomatin protein family. BMC Evol Biol 8, 44 (2008).
42. Lande, W.M. & Mentzer, W.C. Haemolytic anaemia associated with increased cation permeability. Clin Haematol 14, 89-103 (1985).
43. Stewart, G.W., Hepworth-Jones, B.E., Keen, J.N., Dash, B.C., Argent, A.C. & Casimir, C.M. Isolation of cDNA coding for an ubiquitous membrane protein deficient in high Na+, low K+ stomatocytic erythrocytes. Blood 79, 1593-1601 (1992).
44. Zhu, Y., Paszty, C., Turetsky, T., Tsai, S., Kuypers, F.A., Lee, G., Cooper, P., Gallagher, P.G., Stevens, M.E., Rubin, E., Mohandas, N. & Mentzer, W.C. Stomatocytosis is absent in "stomatin"-deficient murine red blood cells. Blood 93, 2404-2410 (1999).
45. Rajaram, S., Spangler, T.L., Sedensky, M.M. & Morgan, P.G. A stomatin and a degenerin interact to control anesthetic sensitivity in Caenorhabditis elegans. Genetics 153, 1673-1682 (1999).
46. Price, M.P., Thompson, R.J., Eshcol, J.O., Wemmie, J.A. & Benson, C.J. Stomatin modulates gating of acid-sensing ion channels. J Biol Chem 279, 53886-53891 (2004).
47. Goodman, M.B., Ernstrom, G.G., Chelur, D.S., O'Hagan, R., Yao, C.A. & Chalfie, M. MEC-2 regulates C. elegans DEG/ENaC channels needed for mechanosensation. Nature 415, 1039-1042 (2002).
48. Sedensky, M.M., Siefker, J.M., Koh, J.Y., Miller, D.M., 3rd & Morgan, P.G. A stomatin and a degenerin interact in lipid rafts of the nervous system of Caenorhabditis elegans. Am J Physiol Cell Physiol 287, C468-474 (2004).

41
49. Tavernarakis, N., Shreffler, W., Wang, S. & Driscoll, M. unc-8, a DEG/ENaC family member, encodes a subunit of a candidate mechanically gated channel that modulates C. elegans locomotion. Neuron 18, 107-119 (1997).
50. Huang, M., Gu, G., Ferguson, E.L. & Chalfie, M. A stomatin-like protein necessary for mechanosensation in C. elegans. Nature 378, 292-295 (1995).
51. Rajaram, S., Sedensky, M.M. & Morgan, P.G. Unc-1: a stomatin homologue controls sensitivity to volatile anesthetics in Caenorhabditis elegans. Proc Natl Acad Sci U S A 95, 8761-8766 (1998).
52. Barnes, T.M., Jin, Y., Horvitz, H.R., Ruvkun, G. & Hekimi, S. The Caenorhabditis elegans behavioral gene unc-24 encodes a novel bipartite protein similar to both erythrocyte band 7.2 (stomatin) and nonspecific lipid transfer protein. J Neurochem 67, 46-57 (1996).
53. Wetzel, C., Hu, J., Riethmacher, D., Benckendorff, A., Harder, L., Eilers, A., Moshourab, R., Kozlenkov, A., Labuz, D., Caspani, O., Erdmann, B., Machelska, H., Heppenstall, P.A. & Lewin, G.R. A stomatin-domain protein essential for touch sensation in the mouse. Nature 445, 206-209 (2007).
54. Sedensky, M.M., Pujazon, M.A. & Morgan, P.G. Tail clamp responses in stomatin knockout mice compared with mobility assays in Caenorhabditis elegans during exposure to diethyl ether, halothane, and isoflurane. Anesthesiology 105, 498-502 (2006).
55. Snyers, L., Umlauf, E. & Prohaska, R. Association of stomatin with lipid-protein complexes in the plasma membrane and the endocytic compartment. Eur J Cell Biol 78, 802-812 (1999).
56. Salzer, U. & Prohaska, R. Stomatin, flotillin-1, and flotillin-2 are major integral proteins of erythrocyte lipid rafts. Blood 97, 1141-1143 (2001).
57. Snyers, L., Thines-Sempoux, D. & Prohaska, R. Colocalization of stomatin (band 7.2b) and actin microfilaments in UAC epithelial cells. Eur J Cell Biol 73, 281-285 (1997).
58. Boute, N., Gribouval, O., Roselli, S., Benessy, F., Lee, H., Fuchshuber, A., Dahan, K., Gubler, M.C., Niaudet, P. & Antignac, C. NPHS2, encoding the glomerular protein podocin, is mutated in autosomal recessive steroid-resistant nephrotic syndrome. Nat Genet 24, 349-354 (2000).
59. Schwarz, K., Simons, M., Reiser, J., Saleem, M.A., Faul, C., Kriz, W., Shaw, A.S., Holzman, L.B. & Mundel, P. Podocin, a raft-associated component of the glomerular slit diaphragm, interacts with CD2AP and nephrin. J Clin Invest 108, 1621-1629 (2001).

42
60. Huber, T.B., Simons, M., Hartleben, B., Sernetz, L., Schmidts, M., Gundlach, E., Saleem, M.A., Walz, G. & Benzing, T. Molecular basis of the functional podocin-nephrin complex: mutations in the NPHS2 gene disrupt nephrin targeting to lipid raft microdomains. Hum Mol Genet 12, 3397-3405 (2003).
61. Mairhofer, M., Steiner, M., Salzer, U. & Prohaska, R. Stomatin-like protein-1 interacts with stomatin and is targeted to late endosomes. J Biol Chem 284, 29218-29229 (2009).
62. Wang, Y. & Morrow, J.S. Identification and characterization of human SLP-2, a novel homologue of stomatin (band 7.2b) present in erythrocytes and other tissues. J Biol Chem 275, 8062-8071 (2000).
63. Kirchhof, M.G., Chau, L.A., Lemke, C.D., Vardhana, S., Darlington, P.J., Marquez, M.E., Taylor, R., Rizkalla, K., Blanca, I., Dustin, M.L. & Madrenas, J. Modulation of T cell activation by stomatin-like protein 2. J Immunol 181, 1927-1936 (2008).
64. Da Cruz, S., Parone, P.A., Gonzalo, P., Bienvenut, W.V., Tondera, D., Jourdain, A., Quadroni, M. & Martinou, J.C. SLP-2 interacts with prohibitins in the mitochondrial inner membrane and contributes to their stability. Biochim Biophys Acta 1783, 904-911 (2008).
65. Hajek, P., Chomyn, A. & Attardi, G. Identification of a novel mitochondrial complex containing mitofusin 2 and stomatin-like protein 2. J Biol Chem 282, 5670-5681 (2007).
66. Da Cruz, S., Xenarios, I., Langridge, J., Vilbois, F., Parone, P.A. & Martinou, J.C. Proteomic analysis of the mouse liver mitochondrial inner membrane. J Biol Chem 278, 41566-41571 (2003).
67. Mootha, V.K., Bunkenborg, J., Olsen, J.V., Hjerrild, M., Wisniewski, J.R., Stahl, E., Bolouri, M.S., Ray, H.N., Sihag, S., Kamal, M., Patterson, N., Lander, E.S. & Mann, M. Integrated analysis of protein composition, tissue diversity, and gene regulation in mouse mitochondria. Cell 115, 629-640 (2003).
68. Rezaul, K., Wu, L., Mayya, V., Hwang, S.I. & Han, D. A systematic characterization of mitochondrial proteome from human T leukemia cells. Mol Cell Proteomics 4, 169-181 (2005).
69. Taylor, S.W., Fahy, E., Zhang, B., Glenn, G.M., Warnock, D.E., Wiley, S., Murphy, A.N., Gaucher, S.P., Capaldi, R.A., Gibson, B.W. & Ghosh, S.S. Characterization of the human heart mitochondrial proteome. Nat Biotechnol 21, 281-286 (2003).
70. Coates, P.J., Nenutil, R., McGregor, A., Picksley, S.M., Crouch, D.H., Hall, P.A. & Wright, E.G. Mammalian prohibitin proteins respond to mitochondrial stress and decrease during cellular senescence. Exp Cell Res 265, 262-273 (2001).

43
71. Ikonen, E., Fiedler, K., Parton, R.G. & Simons, K. Prohibitin, an antiproliferative protein, is localized to mitochondria. FEBS Lett 358, 273-277 (1995).
72. Tatsuta, T., Model, K. & Langer, T. Formation of membrane-bound ring complexes by prohibitins in mitochondria. Mol Biol Cell 16, 248-259 (2005).
73. Artal-Sanz, M., Tsang, W.Y., Willems, E.M., Grivell, L.A., Lemire, B.D., van der Spek, H. & Nijtmans, L.G. The mitochondrial prohibitin complex is essential for embryonic viability and germline function in Caenorhabditis elegans. J Biol Chem 278, 32091-32099 (2003).
74. Ishihara, N., Fujita, Y., Oka, T. & Mihara, K. Regulation of mitochondrial morphology through proteolytic cleavage of OPA1. EMBO J 25, 2966-2977 (2006).
75. Arnoult, D., Grodet, A., Lee, Y.J., Estaquier, J. & Blackstone, C. Release of OPA1 during apoptosis participates in the rapid and complete release of cytochrome c and subsequent mitochondrial fragmentation. J Biol Chem 280, 35742-35750 (2005).
76. Schlame, M., Rua, D. & Greenberg, M.L. The biosynthesis and functional role of cardiolipin. Prog Lipid Res 39, 257-288 (2000).
77. Fry, M. & Green, D.E. Cardiolipin requirement for electron transfer in complex I and III of the mitochondrial respiratory chain. J Biol Chem 256, 1874-1880 (1981).
78. Joshi, A.S., Zhou, J., Gohil, V.M., Chen, S. & Greenberg, M.L. Cellular functions of cardiolipin in yeast. Biochim Biophys Acta 1793, 212-218 (2008).
79. Choi, S.Y., Gonzalvez, F., Jenkins, G.M., Slomianny, C., Chretien, D., Arnoult, D., Petit, P.X. & Frohman, M.A. Cardiolipin deficiency releases cytochrome c from the inner mitochondrial membrane and accelerates stimuli-elicited apoptosis. Cell Death Differ 14, 597-606 (2007).
80. Benard, G., Bellance, N., James, D., Parrone, P., Fernandez, H., Letellier, T. & Rossignol, R. Mitochondrial bioenergetics and structural network organization. J Cell Sci 120, 838-848 (2007).
81. Rich, P.R. & Marechal, A. The mitochondrial respiratory chain. Essays Biochem 47, 1-23 (2010).
82. Acin-Perez, R., Fernandez-Silva, P., Peleato, M.L., Perez-Martos, A. & Enriquez, J.A. Respiratory active mitochondrial supercomplexes. Mol Cell 32, 529-539 (2008).

44
83. Zhang, M., Mileykovskaya, E. & Dowhan, W. Cardiolipin is essential for organization of complexes III and IV into a supercomplex in intact yeast mitochondria. J Biol Chem 280, 29403-29408 (2005).
84. Wenz, T., Hielscher, R., Hellwig, P., Schagger, H., Richers, S. & Hunte, C. Role of phospholipids in respiratory cytochrome bc(1) complex catalysis and supercomplex formation. Biochim Biophys Acta 1787, 609-616 (2009).
85. Epand, R.F., Tokarska-Schlattner, M., Schlattner, U., Wallimann, T. & Epand, R.M. Cardiolipin clusters and membrane domain formation induced by mitochondrial proteins. J Mol Biol 365, 968-980 (2007).
86. Maniti, O., Lecompte, M.F., Marcillat, O., Desbat, B., Buchet, R., Vial, C. & Granjon, T. Mitochondrial creatine kinase binding to phospholipid monolayers induces cardiolipin segregation. Biophys J 96, 2428-2438 (2009).
87. Wang, Y., Cao, W., Yu, Z. & Liu, Z. Downregulation of a mitochondria associated protein SLP-2 inhibits tumor cell motility, proliferation and enhances cell sensitivity to chemotherapeutic reagents. Cancer Biol Ther 8, 1651-1658 (2009).
88. Tondera, D., Grandemange, S., Jourdain, A., Karbowski, M., Mattenberger, Y., Herzig, S., Da Cruz, S., Clerc, P., Raschke, I., Merkwirth, C., Ehses, S., Krause, F., Chan, D.C., Alexander, C., Bauer, C., Youle, R., Langer, T. & Martinou, J.C. SLP-2 is required for stress-induced mitochondrial hyperfusion. EMBO J 28, 1589-1600 (2009).
89. Frauwirth, K.A. & Thompson, C.B. Regulation of T lymphocyte metabolism. J Immunol 172, 4661-4665 (2004).
90. Frauwirth, K.A., Riley, J.L., Harris, M.H., Parry, R.V., Rathmell, J.C., Plas, D.R., Elstrom, R.L., June, C.H. & Thompson, C.B. The CD28 signaling pathway regulates glucose metabolism. Immunity 16, 769-777 (2002).
91. Vander Heiden, M.G., Cantley, L.C. & Thompson, C.B. Understanding the Warburg effect: the metabolic requirements of cell proliferation. Science 324, 1029-1033 (2009).
92. Delgoffe, G.M., Kole, T.P., Zheng, Y., Zarek, P.E., Matthews, K.L., Xiao, B., Worley, P.F., Kozma, S.C. & Powell, J.D. The mTOR kinase differentially regulates effector and regulatory T cell lineage commitment. Immunity 30, 832-844 (2009).
93. Michalek, R.D., Gerriets, V.A., Jacobs, S.R., Macintyre, A.N., MacIver, N.J., Mason, E.F., Sullivan, S.A., Nichols, A.G. & Rathmell, J.C. Cutting edge: distinct glycolytic and lipid oxidative metabolic programs are essential for effector and regulatory CD4+ T cell subsets. J Immunol 186, 3299-3303 (2011).

45
94. Hue, L. & Rider, M.H. The AMP-activated protein kinase: more than an energy sensor. Essays Biochem 43, 121-137 (2007).
95. Pearce, E.L., Walsh, M.C., Cejas, P.J., Harms, G.M., Shen, H., Wang, L.S., Jones, R.G. & Choi, Y. Enhancing CD8 T-cell memory by modulating fatty acid metabolism. Nature 460, 103-107 (2009).
96. Araki, K., Turner, A.P., Shaffer, V.O., Gangappa, S., Keller, S.A., Bachmann, M.F., Larsen, C.P. & Ahmed, R. mTOR regulates memory CD8 T-cell differentiation. Nature 460, 108-112 (2009).
97. Gunter, T.E., Yule, D.I., Gunter, K.K., Eliseev, R.A. & Salter, J.D. Calcium and mitochondria. FEBS Lett 567, 96-102 (2004).
98. Rimessi, A., Giorgi, C., Pinton, P. & Rizzuto, R. The versatility of mitochondrial calcium signals: from stimulation of cell metabolism to induction of cell death. Biochim Biophys Acta 1777, 808-816 (2008).
99. Nicholls, D.G. Mitochondrial ion circuits. Essays Biochem 47, 25-35 (2010).
100. Santo-Domingo, J. & Demaurex, N. Calcium uptake mechanisms of mitochondria. Biochim Biophys Acta 1797, 907-912 (2010).
101. Razzaq, T.M., Ozegbe, P., Jury, E.C., Sembi, P., Blackwell, N.M. & Kabouridis, P.S. Regulation of T-cell receptor signalling by membrane microdomains. Immunology 113, 413-426 (2004).
102. Goodridge, H.S. & Harnett, M.M. Introduction to immune cell signalling. Parasitology 130 Suppl, S3-9 (2005).
103. Oh-hora, M. & Rao, A. Calcium signaling in lymphocytes. Curr Opin Immunol 20, 250-258 (2008).
104. Gallo, E.M., Cante-Barrett, K. & Crabtree, G.R. Lymphocyte calcium signaling from membrane to nucleus. Nat Immunol 7, 25-32 (2006).
105. Baba, Y. & Kurosaki, T. Physiological function and molecular basis of STIM1-mediated calcium entry in immune cells. Immunol Rev 231, 174-188 (2009).
106. Hoth, M., Button, D.C. & Lewis, R.S. Mitochondrial control of calcium-channel gating: a mechanism for sustained signaling and transcriptional activation in T lymphocytes. Proc Natl Acad Sci U S A 97, 10607-10612 (2000).
107. Quintana, A., Schwarz, E.C., Schwindling, C., Lipp, P., Kaestner, L. & Hoth, M. Sustained activity of calcium release-activated calcium channels requires translocation of mitochondria to the plasma membrane. J Biol Chem 281, 40302-40309 (2006).

46
108. Quintana, A., Schwindling, C., Wenning, A.S., Becherer, U., Rettig, J., Schwarz, E.C. & Hoth, M. T cell activation requires mitochondrial translocation to the immunological synapse. Proc Natl Acad Sci U S A 104, 14418-14423 (2007).
109. Da Cruz, S., De Marchi, U., Frieden, M., Parone, P.A., Martinou, J.C. & Demaurex, N. SLP-2 negatively modulates mitochondrial sodium-calcium exchange. Cell Calcium 47, 11-18 (2009).
110. Jin, Z. & El-Deiry, W.S. Overview of cell death signaling pathways. Cancer Biol Ther 4, 139-163 (2005).
111. Bouillet, P. & O'Reilly, L.A. CD95, BIM and T cell homeostasis. Nat Rev Immunol 9, 514-519 (2009).
112. Parsons, M.J. & Green, D.R. Mitochondria in cell death. Essays Biochem 47, 99-114 (2010).
113. Earnshaw, W.C., Martins, L.M. & Kaufmann, S.H. Mammalian caspases: structure, activation, substrates, and functions during apoptosis. Annual review of biochemistry 68, 383-424 (1999).
114. Green, D.R., Galluzzi, L. & Kroemer, G. Mitochondria and the autophagy-inflammation-cell death axis in organismal aging. Science 333, 1109-1112 (2011).
115. Lawen, A. Apoptosis-an introduction. Bioessays 25, 888-896 (2003).
116. Youle, R.J. & Strasser, A. The BCL-2 protein family: opposing activities that mediate cell death. Nat Rev Mol Cell Biol 9, 47-59 (2008).
117. Luo, X., Budihardjo, I., Zou, H., Slaughter, C. & Wang, X. Bid, a Bcl2 interacting protein, mediates cytochrome c release from mitochondria in response to activation of cell surface death receptors. Cell 94, 481-490 (1998).
118. McBride, H.M., Neuspiel, M. & Wasiak, S. Mitochondria: more than just a powerhouse. Curr Biol 16, R551-560 (2006).
119. Scott, I. & Youle, R.J. Mitochondrial fission and fusion. Essays Biochem 47, 85-98 (2010).
120. Chan, D.C. Mitochondrial fusion and fission in mammals. Annu Rev Cell Dev Biol 22, 79-99 (2006).
121. Song, Z., Ghochani, M., McCaffery, J.M., Frey, T.G. & Chan, D.C. Mitofusins and OPA1 mediate sequential steps in mitochondrial membrane fusion. Mol Biol Cell 20, 3525-3532 (2009).
122. Yoon, Y., Krueger, E.W., Oswald, B.J. & McNiven, M.A. The mitochondrial protein hFis1 regulates mitochondrial fission in mammalian cells through an

47
interaction with the dynamin-like protein DLP1. Mol Cell Biol 23, 5409-5420 (2003).
123. Mozdy, A.D., McCaffery, J.M. & Shaw, J.M. Dnm1p GTPase-mediated mitochondrial fission is a multi-step process requiring the novel integral membrane component Fis1p. J Cell Biol 151, 367-380 (2000).
124. Otera, H., Wang, C., Cleland, M.M., Setoguchi, K., Yokota, S., Youle, R.J. & Mihara, K. Mff is an essential factor for mitochondrial recruitment of Drp1 during mitochondrial fission in mammalian cells. J Cell Biol 191, 1141-1158 (2010).
125. Chen, H., Detmer, S.A., Ewald, A.J., Griffin, E.E., Fraser, S.E. & Chan, D.C. Mitofusins Mfn1 and Mfn2 coordinately regulate mitochondrial fusion and are essential for embryonic development. J Cell Biol 160, 189-200 (2003).
126. Davies, V.J., Hollins, A.J., Piechota, M.J., Yip, W., Davies, J.R., White, K.E., Nicols, P.P., Boulton, M.E. & Votruba, M. Opa1 deficiency in a mouse model of autosomal dominant optic atrophy impairs mitochondrial morphology, optic nerve structure and visual function. Hum Mol Genet 16, 1307-1318 (2007).
127. Ishihara, N., Nomura, M., Jofuku, A., Kato, H., Suzuki, S.O., Masuda, K., Otera, H., Nakanishi, Y., Nonaka, I., Goto, Y., Taguchi, N., Morinaga, H., Maeda, M., Takayanagi, R., Yokota, S. & Mihara, K. Mitochondrial fission factor Drp1 is essential for embryonic development and synapse formation in mice. Nat Cell Biol 11, 958-966 (2009).
128. Wakabayashi, J., Zhang, Z., Wakabayashi, N., Tamura, Y., Fukaya, M., Kensler, T.W., Iijima, M. & Sesaki, H. The dynamin-related GTPase Drp1 is required for embryonic and brain development in mice. J Cell Biol 186, 805-816 (2009).
129. Chen, H., McCaffery, J.M. & Chan, D.C. Mitochondrial fusion protects against neurodegeneration in the cerebellum. Cell 130, 548-562 (2007).
130. Mitra, K., Wunder, C., Roysam, B., Lin, G. & Lippincott-Schwartz, J. A hyperfused mitochondrial state achieved at G1-S regulates cyclin E buildup and entry into S phase. Proc Natl Acad Sci U S A 106, 11960-11965 (2009).
131. Taguchi, N., Ishihara, N., Jofuku, A., Oka, T. & Mihara, K. Mitotic phosphorylation of dynamin-related GTPase Drp1 participates in mitochondrial fission. J Biol Chem 282, 11521-11529 (2007).
132. Jeong, S.Y. & Seol, D.W. The role of mitochondria in apoptosis. BMB Rep 41, 11-22 (2008).
133. Suen, D.F., Norris, K.L. & Youle, R.J. Mitochondrial dynamics and apoptosis. Genes Dev 22, 1577-1590 (2008).

48
134. Frezza, C., Cipolat, S., Martins de Brito, O., Micaroni, M., Beznoussenko, G.V., Rudka, T., Bartoli, D., Polishuck, R.S., Danial, N.N., De Strooper, B. & Scorrano, L. OPA1 controls apoptotic cristae remodeling independently from mitochondrial fusion. Cell 126, 177-189 (2006).
135. Karbowski, M., Lee, Y.J., Gaume, B., Jeong, S.Y., Frank, S., Nechushtan, A., Santel, A., Fuller, M., Smith, C.L. & Youle, R.J. Spatial and temporal association of Bax with mitochondrial fission sites, Drp1, and Mfn2 during apoptosis. J Cell Biol 159, 931-938 (2002).
136. Lee, Y.J., Jeong, S.Y., Karbowski, M., Smith, C.L. & Youle, R.J. Roles of the mammalian mitochondrial fission and fusion mediators Fis1, Drp1, and Opa1 in apoptosis. Mol Biol Cell 15, 5001-5011 (2004).
137. Garofalo, T., Giammarioli, A.M., Misasi, R., Tinari, A., Manganelli, V., Gambardella, L., Pavan, A., Malorni, W. & Sorice, M. Lipid microdomains contribute to apoptosis-associated modifications of mitochondria in T cells. Cell Death Differ 12, 1378-1389 (2005).
138. Sorice, M., Manganelli, V., Matarrese, P., Tinari, A., Misasi, R., Malorni, W. & Garofalo, T. Cardiolipin-enriched raft-like microdomains are essential activating platforms for apoptotic signals on mitochondria. FEBS Lett 583, 2447-2450 (2009).
139. Kagan, V.E., Tyurin, V.A., Jiang, J., Tyurina, Y.Y., Ritov, V.B., Amoscato, A.A., Osipov, A.N., Belikova, N.A., Kapralov, A.A., Kini, V., Vlasova, II, Zhao, Q., Zou, M., Di, P., Svistunenko, D.A., Kurnikov, I.V. & Borisenko, G.G. Cytochrome c acts as a cardiolipin oxygenase required for release of proapoptotic factors. Nat Chem Biol 1, 223-232 (2005).
140. Kim, T.H., Zhao, Y., Ding, W.X., Shin, J.N., He, X., Seo, Y.W., Chen, J., Rabinowich, H., Amoscato, A.A. & Yin, X.M. Bid-cardiolipin interaction at mitochondrial contact site contributes to mitochondrial cristae reorganization and cytochrome C release. Mol Biol Cell 15, 3061-3072 (2004).
141. Liu, J., Weiss, A., Durrant, D., Chi, N.W. & Lee, R.M. The cardiolipin-binding domain of Bid affects mitochondrial respiration and enhances cytochrome c release. Apoptosis 9, 533-541 (2004).
142. Gonzalvez, F., Schug, Z.T., Houtkooper, R.H., Mackenzie, E.D., Brooks, D.G., Wanders, R.J., Petit, P.X., Vaz, F.M. & Gottlieb, E. Cardiolipin provides an essential activating platform for caspase-8 on mitochondria. J Cell Biol 183, 681-696 (2008).
143. Gonzalvez, F., Pariselli, F., Dupaigne, P., Budihardjo, I., Lutter, M., Antonsson, B., Diolez, P., Manon, S., Martinou, J.C., Goubern, M., Wang, X., Bernard, S. & Petit, P.X. tBid interaction with cardiolipin primarily orchestrates mitochondrial

49
dysfunctions and subsequently activates Bax and Bak. Cell Death Differ 12, 614-626 (2005).
144. Ciarlo, L., Manganelli, V., Garofalo, T., Matarrese, P., Tinari, A., Misasi, R., Malorni, W. & Sorice, M. Association of fission proteins with mitochondrial raft-like domains. Cell Death Differ 17, 1047-1058 (2010).
145. Huang, Z., Jiang, J., Tyurin, V.A., Zhao, Q., Mnuskin, A., Ren, J., Belikova, N.A., Feng, W., Kurnikov, I.V. & Kagan, V.E. Cardiolipin deficiency leads to decreased cardiolipin peroxidation and increased resistance of cells to apoptosis. Free Radic Biol Med 44, 1935-1944 (2008).

50
Chapter 2
2 Stomatin-Like Protein 2 Binds Cardiolipin and Regulates Mitochondrial Biogenesis and Function 1
Stomatin-like protein 2 (SLP-2) is a widely expressed mitochondrial inner membrane
protein of unknown function. Here we show that human SLP-2 interacts with prohibitin-
1 and -2 and binds to the mitochondrial membrane phospholipid cardiolipin.
Upregulation of SLP-2 expression increases cardiolipin content and the formation of
metabolically active mitochondrial membranes and induces mitochondrial biogenesis. In
human T lymphocytes, these events correlate with increased complex I and II activities,
increased intracellular ATP stores, and increased resistance to apoptosis through the
intrinsic pathway, ultimately enhancing cellular responses. We propose that the function
of SLP-2 is to recruit prohibitins to cardiolipin to form cardiolipin-enriched
microdomains in which electron transport complexes are optimally assembled. Likely
through the prohibitin functional interactome, SLP-2 then regulates mitochondrial
biogenesis and function.
2.1 Introduction
The demands for energy-generating substrates in a eukaryotic cell rise as the cell
transitions from a resting state to an activated state. This process is well documented in
lymphocytes undergoing mitogenic responses. In these cells, the increasing energy
requirements are met by increased production of ATP either by glycolysis or by oxidative
1 The material contained in this chapter was published in: Christie, D., Lemke, C.D., Elias, I.M.,
Chau, L.A., Kirchhof, M.G., Li, B., Ball, E.H., Dunn, S.D., Hatch, G.M. and Madrenas, J. Stomatin-like protein 2 binds cardiolipin and regulates mitochondrial biogenesis and function. Mol. Cell. Biol. 31 (18): 3845-3856 (2011).
Copyright 2011. American Society for Microbiology

51
phosphorylation (OXPHOS); both processes being regulated by signalling from antigen
receptors and costimulatory molecules 1-3. Scattered evidence suggests that these
demands on cellular bioenergetics correlate with augmentation of mitochondrial
membrane surface and of mitochondrial numbers 4-6, a finding consistent with the key
role played by these organelles in supplying most cellular ATP 3. However, the
molecules that regulate mitochondrial biogenesis in response to cell activation remain
mostly unknown.
Recent evidence suggests that the biogenesis of mitochondrial membranes may be
regulated by the functional interactome of prohibitins (PHB) -1 and -2 7, but it is not
known how prohibitins sense the need to increase mitochondrial membrane biogenesis.
We have previously reported that SLP-2, an evolutionarily conserved mitochondrial
protein belonging to the stomatin family 8-10 and enriched in detergent-insoluble
microdomains of T lymphocytes, is upregulated during activation of these cells and
enhances T cell responses in vivo and in vitro 11. The function of SLP-2 is unknown.
Since SLP-2 is mostly located in mitochondrial membranes and has been shown to
interact with PHB-1 and PHB-2 12 we hypothesized that upregulation of SLP-2
expression may act as a linker between PHBs and mitochondrial biogenesis. Here, we
show that SLP-2 binds cardiolipin (CL) and facilitates the assembly of respiratory chain
components. Upregulation of SLP-2 expression then translates into enhanced
mitochondrial biogenesis and function.
2.2 Materials and Methods
2.2.1 Cells
Jurkat T cells were obtained from American Type Culture Collection (Manassas, VA).
The LG2 B lymphoblastoid cell line used as antigen-presenting cells was provided by Dr.
Eric Long (NIAID, NIH, Rockville, MD). Peripheral blood mononuclear cells (PBMCs)
were isolated from normal donors using Ficoll-Hypaque (Amersham Pharmacia Biotech,
Uppsala, Sweden). Cells were washed and resuspended at 1 x 106 cells/ml. PBMC blasts

52
were generated with Phorbol Myristate Acetate (PMA; 1 ng/ml) and Ionomycin (100
ng/ml) at 37oC for 72 h.
2.2.2 Plasmids, siRNA and T cell transfectants
Full length human SLP-2 cDNA was subcloned into the pEGFPN1 expression vector
(Clontech Inc., Palo Alto, CA) to create an in-frame translational fusion of SLP-2 with
GFP at the 3’ end (SLP-2-GFP) as described 11. The full length SLP-2-GFP was placed
into the doxycycline-inducible pBig2i vector 13. A doxycycline-inducible mutant of
human SLP-2 lacking the amino-terminal domain tagged with GFP was also constructed.
Stable transfectants were generated by nucleofection.
2.2.3 Upregulation of SLP-2 expression
For induction of high levels of SLP-2 expression, doxycycline (Sigma, St. Louis, MO)
was added in culture at 1 µg/mL for 18-24 h. Parental cells, which express low levels of
SLP-2, are referred to as low expressors (SLP-2lo) whereas SLP-2-GFP-transfected T
cells cultured in the presence of doxycycline are referred as high expressors (SLP-2hi).
Transfection of pBig2iGFP, pBig2iSLP2GFP, pmaxGFP® (Amaxa, Gaithersburg, MD)
or mock transfection into human PBMC or isolated primary CD4+ T helper cells were
conducted using Human T Cell Nucleofector Kit (Amaxa). After transfection, cells were
cultured for 24 h before stimulation.
2.2.4 Antibodies
The following monoclonal antibodies (MAb) were used in these studies: OPA-1 and
DLP1(Drp1) (BD Bisosciences, Mississauga, ON); cytochrome c , complex I subunit
NDUFS3, the 70kDa subunit of Complex II, Complex III core II, subunit II of Complex
IV, the alpha subunit of Complex V (MitoSciences, Eugene, OR), β-actin (Santa Cruz
Biotechnology, Santa Cruz, CA), and glyceraldehyde-3-phosphate dehydrogenase
(GAPDH) (Chemicon International, Temecula, CA). Rabbit polyclonal antibodies
against Mfn2 (Sigma, St. Louis, MO), PGC-1α (Santa Cruz Biotechnology, Santa Cruz,
CA), and SLP-2 (ProteinTech Group, Inc., Chicago, IL) were also used.

53
2.2.5 Reagents
The following working concentrations were used: 4 µM oligomycin, 1 µg/mL ionomycin
(both from Sigma). [3H]Acetate and [1-14C]linoleic acid were obtained from Amersham
(Oakville, Ontario, Canada). Lipid standards were obtained from Serdary Research
Laboratories (Englewood Cliffs, NJ). Thin layer chromatographic plates (silica gel G;
0.25 mm thickness) were obtained from Fisher Scientific (Winnipeg, Manitoba, Canada).
Ecolite scintillant was obtained from ICN Biochemicals (Montreal, Quebec, Canada).
Primers for real time-PCR were obtained from Invitrogen. All other chemicals were
certified ACS grade or better and obtained from Sigma Chemical Company or Fisher
Scientific.
2.2.6 Confocal microscopy
Confocal microscopy was performed with a Zeiss LSM 510 microscope (Carl Zeiss, Inc.) 14. Photobleaching experiments to test for mitochondrial fusion were carried out by
targeting an area of MitoTracker Red CMXRos labeled mitochondria within a cell for
repeated exposure to high power HeNe1 laser. Loss of fluorescence at the targeted point
versus an unrelated point within the same cell or another cell were then compared.
Similar loss in fluorescent intensity from the unrelated point within the same cell
indicated mitochondrial fusion between the two points.
2.2.7 Transmission electron microscopy
Cells were fixed in 2.5% gluteraldehyde in 0.1M sodium cacodylate buffer for 2 h,
washed in the buffer, fixed in 1% osmium tetroxide in 0.1M sodium cacodylate buffer for
1 h, washed in the buffer again and enrobed in Noble agar. Following a wash in distilled
water, cells were stained with 2% uranyl acetate for 2 h, then dehydrated through a
graded series of ethanol, cleared in propylene oxide, infiltrated with Epon resin and
embedded. Thin sections were stained with 2% uranyl acetate and lead citrate and were
viewed with a Philips 410 transmission electron microscope.

54
2.2.8 Measurement of mitochondrial mass
Human PBMCs were labeled with 100 nM MitoTracker Red® CMXRos (InvitrogenTM)
for 20 min at 37°C in complete RPMI 1640. Fluorescence was detected by flow
cytometry and analyzed with FlowJo flow cytometry analysis software (Treestar, Inc.,
Ashland, OR). To inhibit cardiolipin synthase during in vitro activation, human PBMCs
were stimulated with 1 ng/mL PMA and 100 ng/mL ionomycin in the presence of 4µM
triacsin C at 37°C for 24 hours 15.
2.2.9 Measurement of cardiolipin
Human PBMCs, resting or blasted were labeled with 10nM nonyl-acridine orange
(Sigma) for 15 min at 37°C. Fluorescence was detected by flow cytometry.
2.2.10 Measurement of cardiolipin mass in stable doxycycline-inducible SLP-2-GFP transfected Jurkat cells
Jurkat cells stably transfected with doxycycline-inducible SLP-2-GFP were incubated
without (SLP-2lo) or with (SLP-2hi) 1 µg/ml doxycycline for 72 h. After incubation, the
medium was aspirated, and the dishes were washed twice with 2 ml of ice-cold
phosphate-buffered saline. Cells were harvested in 2 ml of methanol:water (1:1, by
volume), and 25 µL aliquots were taken for protein determination 16. Subsequently, 0.5
ml of water and 2 ml chloroform were added to initiate phase separation. Samples were
centrifuged at 2000 rpm for 10 min, and the upper phase was aspirated. Two milliliters of
theoretical upper phase (methanol-0.9% NaCl-chloroform) (48:47:3, by volume) was
added, and centrifugation was repeated for 5 min. The organic phase was removed, dried
under nitrogen gas, and resuspended in chloroform:methanol (2:1, by volume). A 50-µl
aliquot was spotted onto thin-layer plates for separation of cardiolipin from other
phospholipids as described elsewhere 35. Phospholipids were removed from the plate, and
the phospholipid phosphorus content of cardiolipin was determined as described
previously 17.

55
2.2.11 Radiotracer studies of de novo cardiolipin synthesis
SLP-2lo or SLP-2hi cells were grown in Dulbecco’s modified Eagle medium (DMEM)
with 10% fetal bovine serum (FBS) and 100 U Penicillin, and 100 µg Streptomycin in 60
mm-diameter dishes and were incubated at 37°C under a humidified atmosphere of 5%
CO2. Cells were then incubated immediately after doxycycline treatment with 0.1 µM
[3H]acetate (10 µCi/dish) for 16 h or 0.1 mM [1-14C]linoleic acid (3 µCi/dish) bound to
albumin (molar ratio, 1:1) for 8 h. Cells were harvested, and the amount of radioactivity
incorporated into cardiolipin determined as described previously 18.
2.2.12 Real time-PCR
The cycler protocol for real-time PCR consisted of a 30-min reverse transcription (RT)
step at 50oC, followed by a 15 min Taq activation step at 95oC, followed by a 1-min
separation at 95oC and a melting curve that increased in temperature incrementally from
60oC to 95oC over the course of 20 min. The primers used for real time-PCR were
Invitrogen’s D-LUX 6-carboxyfluorescein (FAM)-labeled primer sets: for cardiolipin
synthase, reverse primer CGAACCGTGGTGTTGGAAGAGTT-FAM-G, forward primer
CGAGAGATGTAATGTTGATTGCTG; for phosphatidylglycerol phosphate synthase,
reverse primer CGGTGAGTCACTCAGGTTTGCACC-FAM-G, forward primer
TCGGCCTCCAGCACATTAAG; for PGC-1α, reverse primer TGC TTC GTC GTC
AAA AAC AG, forward primer TCA GTC CTC ACT GGT GGA CA; and for 18srRNA
reverse primer CGGGTGCTCTTAGCTGAGTGTCC-FAM-G, forward primer
CTCGGGCCTGCTTTGAACAC. Changes in mRNAs were analyzed on an Eppendorf
Mastercycler ep Realplex, software version 1.5.474, and data are presented as the mean
fold change (2-ΔΔCT) in mRNA expression relative to 18s rRNA expression 19.
2.2.13 Mitochondrial DNA quantification
Mitochondrial cytochrome c oxidase I (CCO) and nuclear polymerase γ accessory subunit
were amplified from total cellular DNA 20. Relative levels of mitochondrial DNA were
determined by comparing the threshold cycle (CT) detection values of CCO in SLP-2hi
and SLP-2lo cells and were normalized to nuclear DNA levels by the ΔΔCT method 19.

56
2.2.14 Cell lysate preparation
Cells were lysed in a 1% Triton X-100-containing buffer (1% Triton X-100, 150mM
NaCl, 10mM Tris [pH7.6], 5mM EDTA, 1mM sodium orthovanadate, 10µg/mL
leupeptin, 10µg/mL aprotinin, 25µM p-nitrophenyl-p’-guanidinobenzoate) at 4ºC for 30
min. Lysates were cleared by centrifugation (10,700 x g, 4ºC, 10 min), mixed with
sample buffer containing β-mercaptoethanol, boiled and blotted 21.
2.2.15 Mitochondria isolation
Intact mitochondria were isolated from 5 x 106 to 10 x 106 Jurkat T cells using the
Qproteome Mitochondria Isolation Kit (Qiagen). Mitochondrial preparations were mixed
with sample buffer containing β-mercaptoethanol, boiled, and analyzed by sodium
dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE).
2.2.16 Prohibitin and SLP-2 membrane association
Mitochondria were isolated and fractionated with 0.1M Na2CO3 as described previously 22. Fractions treated with H2O were used as an input control. Soluble and membrane
fractions were analyzed by immunoblotting for PHB-1 and SLP-2.
2.2.17 NADH dehydrogenase activity
NADH dehydrogenase activity from whole cell lysates was measured using the complex I
enzyme activity microplate assay kit (Mitosciences).
2.2.18 ATP quantitation
Mitochondrial and total-cell-lysate ATP levels were measured using the ATP
Determination Kit (Molecular ProbesTM, Eugene, OR). Briefly, 10 µL of ATP standards
(ranging from 1.95 to 1000 nM), mitochondrial fractions or cell lysates were added in
triplicate to 90 µL of a standard reaction solution in a 96-well Chromalux luminescent
assay microplate (Dynex Technologies, Chantilly, VA). Sample luminescence was
measured using an MLX microtiter plate luminometer (Dynex Technologies). Sample
ATP concentrations were calculated from the standard curve and were normalized to cell
equivalents.

57
2.2.19 T cell stimulation
Jurkat T cells were stimulated as described previously 14. A CD4+ T cell isolation kit and
MACS (magnetic cell sorting) columns (Miltenyi Biotec Inc., Auburn, CA) were used to
isolate primary CD4+ T helper cells from PBMCs. pBig2iGFP, pBig2iSLP2GFP,
pmaxGFP® or mock transfected PBMCs (0.2x106 cells/group) in the presence or absence
of 1µg/mL doxycycline were plated in triplicate on 96 well plates with various
concentrations of staphylococcal enterotoxin E (SEE) at 37°C and 5% CO2 for 24 h.
Supernatants were collected, and interleukin-2 (IL-2) was measured by an enzyme-linked
immunosorbent assay (ELISA) 23.
2.2.20 Oxygen consumption
Cells were incubated in a 96-well oxygen biosensor system (Becton Dickinson [BD], San
Jose, CA). Background fluorescence was measured before plating cells, and specific
fluorescence due to oxygen consumption was measured after 24 h of incubation at 37°C.
The fluorescent signal was measured on a Safire fluorescent plate reader (Tecan,
Switzerland). To calculate [O2] from fluorescence intensity, we followed the protocol
supplied by BD: [O2] = [(DR/NRF)-1]/KSV, where NRF is normalized relative
fluorescence, calculated as the fluorescence intensity of cells (I) divided by the
fluorescence intensity of a blank well (IB); DR is calculated as the fluorescence intensity
of 100mM sodium sulfite (IS) divided by IB; and KSV is calculated as (DR-1)/[O2]A,
where [O2]A equals 195µM at 37°C with 5% CO2.
2.2.21 Induction and detection of apoptosis
T cells were treated with 5 µg/mL of actinomycin D (Sigma) for 3 h to induce apoptosis,
which was measured by flow cytometry as ΔΨm (mitochondrial transmembrane potential)
loss using the cell-permeate probe MitoTracker Red CMXRos (InvitrogenTM), or as an
increase in Annexin V staining.
2.2.22 Generation of human recombinant SLP-2 (hrSLP-2).
We generated a hrSLP-22 cDNA coding for amino acids 36 to 356 of human SLP-2
downstream of a sequence coding for 6xHIS tag, a thrombin cleavage site, Thioredoxin,

58
and a tobacco etch virus (TEV) cleavage site in a pET28 plasmid. Rosetta strain E. coli
containing plasmids were streaked and single colonies were picked and grown to a A600
of 0.5 to 0.6. The culture was cooled to 15°C and induced with 0.1 mM isopropyl-β-D-
thiogalactopyranoside (IPTG) overnight. Bacteria were pelleted and scraped into pre-
weighed 50 ml centrifuge tubes, resuspended in 25 ml 50/10 buffer (50 mM Tris pH 8.0,
10 mM MgCl2), and centrifuged at 10,000 rpm for 10 min. The supernatant was decanted,
and the pellet was weighed and resuspended in 10x (wet weight) 50/10 buffer. The buffer
was fortified with 1 mM PMSF (Sigma), 5 mM p-aminobenzamidine (Sigma) and
Complete protease inhibitor cocktail (Roche). The bacteria were lysed by two passes at
10,000 PSI in a French pressure cell, and the lysates were centrifuged for 10 min at
10,000 rpm at 4°C. The supernatant was decanted into 60 ml ultracentrifuge tubes, spun
at 38,000 rpm at 4°C for 90 min, decanted again, and stored at -80°C.
Chromatography was run using Chelating Sepharose Fast Flow resin (GE BioScience).
Two column volumes of NiSO4 were applied to the column in order to bind the nickel.
The column was equilibrated with 10 column volumes of Buffer A (20 mM Tris-HCl [pH
8.0], 100 mM NaCl, 5 mM 2-mercaptoethanol, 10% glycerol, 20 mM imidazole). The
sample was added to the column and was allowed to flow through the resin at a rate of
approximately 0.5 ml/min. The column was washed with 10 column volumes of Buffer
A, followed by 2 column volumes of Buffer B (20 mM Tris-HCl [pH 8.0], 1 M NaCl, 5
mM 2-mercaptoethanol, 10% glycerol) and a further 2 volumes of Buffer A. Bound
proteins were eluted by competitive binding with Buffer C (20 mM Tris-HCl [pH 8.0],
100 mM NaCl, 5 mM 2-mercaptoethanol, 10% glycerol, 0-500 mM imidazole) and were
collected in 1.5 ml or 5 ml fractions. Fractions were stained with Coomassie Brilliant
Blue and were analysed on SDS-PAGE.
Protein samples were dialysed into 10 mM Tris (pH 8.0) overnight at 4°C. The protein
solution was then brought up to 2 mM EDTA and 1 mM dithiothreitol (DTT) and was
quantitated by the bicinchoninic acid (BCA) method (BCA protein assay kit; Pierce). The
SLP-2 construct was then incubated with TEV protease (ratio, 1:10) at room temperature
for 2 h. After cleavage, the sample was dialyzed into 10 mM Tris (pH 8.0) overnight at
4°C to remove EDTA and DTT.

59
2.2.23 Phospholipid vesicle co-precipitation assay
The phospholipid vesicle co-precipitation assay was performed as described previously 24. The required amounts of phospholipids were transferred into glass vials and were
dried under a gentle stream of Nitrogen gas. The required amount of binding buffer
(20mM Tris [pH7.5], 150mM NaCl, 1mM EDTA) was added, and samples incubated for
3 h at 52ºC. After this incubation, hrSLP-2 protein was added to the lipid vesicles
solution at a 5:1 ratio of lipid to protein and was incubated for 30 min at 37ºC. Samples
were centrifuged at 40,000 rpm for 10 min at room temperature; the supernatant was
removed; and pellets were resuspended in binding buffer. Laemmli sample buffer was
added to supernatant and pellet fractions, boiled for 7 min, and run on an SDS-PAGE.
Bovine heart cardiolipin was purchased from Sigma; chicken egg L-α-
phosphatidylcholine (PC), L-α-phosphadidylethanolamine (PE) and L-α-
phosphadidylglycerol, porcine brain L-α-phosphatidylserine, and soy L-α-
phosphadidylinositol were purchased from Avanti Polar Lipids, Inc. (Alabaster AB).
2.3 Results
2.3.1 SLP-2 is localized mostly in mitochondria
Studies by different groups, including our own, have shown that SLP-2 in human cells is
expressed in two pools 11, 12, 22. The major pool of SLP-2 is associated with the
mitochondrial inner membrane, whereas a quantitatively minor pool is associated with
the plasma membrane. This compartmentalization was corroborated biochemically by
subcellular fractionation of human T cells (Fig. 2.1a). SLP-2 was detected in membrane
and organelle/cytoskeleton fractions but not in nuclear or cytosolic compartments. Porin
(voltage-dependent anion channel [VDAC]), a protein detected in mitochondrial and
plasma membranes, showed partitioning pattern similar to that of SLP-2.
To study the role of SLP-2, we developed stable transfected cells in which SLP-2
expression can be tightly regulated using a doxycycline-inducible promoter coding for a
C-terminally GFP-tagged SLP-2 protein (SLP-2-GFP). This system allowed us to control

60
Figure 2.1: The major pool of SLP-2 in human T cells is located in mitochondria
(a) Subcellular fractions of Jurkat T cells were isolated by differential centrifugation and
were immunoblotted for SLP-2. Serial blotting for GAPDH, Histone H1, CD45, and
talin was performed to control for the quality of the fractions. Blotting of porin (VDAC),
another protein with a major subcellular pool in mitochondrial and a smaller pool in the
plasma membrane, is also shown. (b) Mitochondrial and cytosolic fractions of stable T
cell SLP-2-GFP transfectants expressing low (SLP-2lo) and high (SLP-2hi) levels of SLP-
2-GFP were isolated by differential centrifugation and immunoblotted for SLP-2,
electron transport complex III, and β-actin. Note that in addition to endogenous SLP-2
and SLP-2-GFP forms, we detected bands likely reflecting intermediate SLP-2
degradation fragments. (c) Stably expressed SLP-2-GFP colocalizes with MitoTracker
Red CMXRos by confocal microscopy. Bars, 5 µm (d) Three-dimensional reconstitution
image analysis. A lateral view of a T cell attached to a glass slide is shown. The yellow
units represent mitochondria and the mitochondrial network. Images are representative
of more than 100 cells from more than 5 independent experiments.
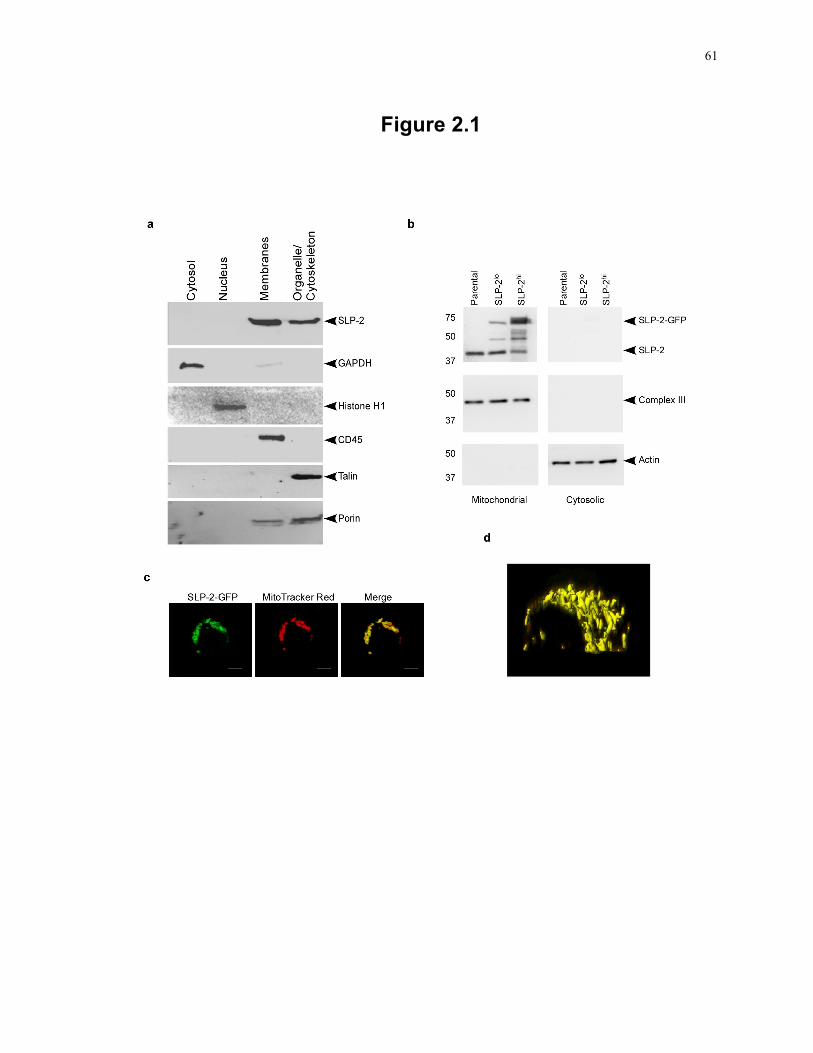
61
Figure 2.1

62
the levels of SLP-2 in human cells (T cells for these experiments) from very low (basal
levels of expression by Jurkat T cells stably transfected with this cDNA in the absence of
doxycycline; referred to below as SLP-2lo) to high (10-100 times higher expression upon
addition of doxycycline; referred to below as SLP-2hi). With these cells, we corroborated
the predominant localization of SLP-2 in mitochondria both by biochemical fractionation
(Fig. 2.1b) and by confocal microscopy, in which SLP-2-GFP colocalized with the
mitochondrial dye MitoTracker Red (Fig. 2.1c and 2.1d). Such an intracellular
distribution was determined by an N-terminal mitochondrial targeting sequence 12, 22, as
evidenced by the fact that SLP-2-GFP deletion mutants lacking the N-terminus abrogated
the mitochondrial localization, whereas expression of the SLP-2 N-terminal domain fused
to GFP was sufficient to determine mitochondrial localization of this protein (data not
shown 12).
2.3.2 Upregulation of SLP-2 expression increases mitochondrial biogenesis
To learn about the function of SLP-2, given its mitochondrial localization, we first
examined the effect of induction of SLP-2 expression on mitochondrial mass by using the
system mentioned above. We found that upregulation of SLP-2 expression, but not of a
control protein, induced a significant increase in the mass of metabolically-active
mitochondria (P < 0.01), as determined with the intact mitochondria label MitoTracker
Red® (Fig. 2.2a and b), and a significant increase in the number of mitochondria per T
cell (P < 0.001) (Fig. 2.2c and 2d). Such mitochondrial biogenesis required appropriate
targeting of SLP-2 to mitochondria, as evidenced by the fact that overexpression of a
SLP-2-GFP mutant lacking the N terminal mitochondrial targeting sequence (ΔNSLP-2)
did not induce a significant increase in the number of mitochondria (Fig. 2.2c and 2.2d),
although a slight increase over baseline was observed, likely due to limited transport of
the mutant as a dimer with endogenous full length SLP-2. Furthermore, the increased
mitochondrial mass observed after upregulation of SLP-2 expression was corroborated by
flow cytometry for primary human leukocytes from peripheral blood in the transition

63
Figure 2.2: Induction of SLP-2 expression triggers mitochondrial biogenesis in
human T cells.
(a and b) T cells expressing high levels of SLP-2 (SLP-2hi) and GFP-expressing control
cells, which contain lower levels of SLP-2 (SLP-2lo), were stained with MitoTracker Red,
and the signal was quantified in each cell. Bars, 5 µm. (c) Mitochondria visualized by
transmission electron microscopy were counted for 30 cell slices of SLP-2hi T cells, SLP-
2lo T cells, or T cells expressing a SLP-2 mutant in which the N-terminal mitochondrial
targeting sequence was deleted (ΔNSLP-2). Asterisks indicate significant differences
(***, P<0.001; **, P<0.01) as determined by Student’s t test. (d) Representative
transmission electron microscopy images of SLP-2lo, SLP-2hi and ΔNSLP-2 T cells.
Mitochondria are indicated by white stars. Bars, 2 µm. (e) Mitochondrial mass was
measured by flow cytometry in resting (shaded histogram) and activated (filled
histogram) human peripheral blood mononuclear cells after labeling with MitoTracker
Red. The result for the unstained control is shown by a dotted line. (f) Immunoblotting
of PBMC whole-cell lysates for SLP-2 demonstrated that SLP-2 levels were upregulated
after blasting. β−Actin blotting was used as a loading control. Densitometric units of
SLP-2 for each group normalized to actin levels are shown.
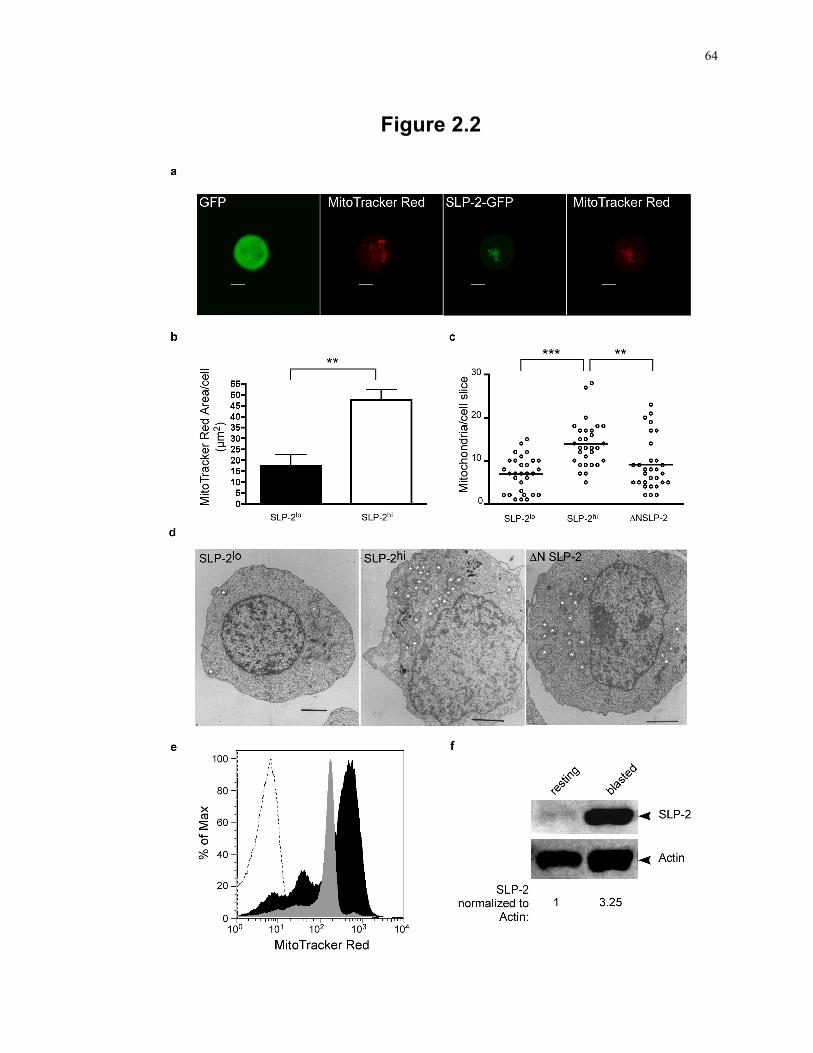
64
Figure 2.2

65
from resting state (in which they express low levels of SLP-2) to the primed blasted state
(in which they express high levels of SLP-2) (Fig. 2.2e and 2f).
Mitochondrial biogenesis requires de novo synthesis of cardiolipin, a phospholipid in the
mitochondrial membrane. Since upregulation of SLP-2 expression was associated with
increased mitochondrial biogenesis, we next tested whether SLP-2 overexpression
increased cardiolipin synthesis. A 37% increase in cardiolipin content (as a percentage of
total phospholipid phosphorus mass) was detected in SLP-2hi expressing cells (Fig. 2.3a).
This finding was corroborated in human PBMCs transitioning from resting state (i.e.,
expressing low levels of SLP-2) to the activated state (i.e., expressing high levels of SLP-
2) by flow cytometry with the cardiolipin dye nonyl-acridine orange (Fig. 2.3b). Since
the loading and retention of this dye are sensitive to changes in mitochondrial potential 25,
we verified that the increase in cardiolipin content was due to de novo synthesis of this
phospholipid by using radiolabeling assays incubating SLP-2lo and SLP-2hi cells with 3H-
acetate and 14C-linoleic acid, both of which are incorporated into newly synthesized
cardiolipin. SLP-2hi cells showed higher levels of radiolabeled cardiolipin under both
incubation conditions than did SLP-2lo cells (P < 0.05) (Fig. 2.3c and 2.3d). Finally, we
used real time RT-PCR to look at the levels of phosphatidylglycerol phosphate synthase
mRNA and cardiolipin synthase mRNA, since these resulting enzymes are involved in
the final steps of cardiolipin biosynthesis. As expected, the SLP-2hi cells had consistently
higher transcript levels of both enzymes (Fig. 2.3e and 2.3f), consistent with a state of
activation of lipid biosynthesis at the transcriptional level. Together, these data led us to
conclude that SLP-2 regulates cardiolipin synthesis, which is required for the formation
of mitochondrial membranes. To examine the requirement of cardiolipin synthase
upregulation for mitochondrial biogenesis in SLP-2hi cells, PBMCs were activated with 1
ng/mL PMA and 100 ng/mL ionomycin in the presence of 4 µM triacsin C, an inhibitor
of cardiolipin synthase. Under these conditions, the increase in mitochondrial mass upon
T cell activation was blocked (Fig. 2.3g), indicating a requirement for cardiolipin
synthesis in mitochondrial biogenesis induced by SLP-2 upregulation.

66
Figure 2.3: Induction of SLP-2 expression increases de novo cardiolipin
biosynthesis, nuclear transcription programs, and mitochondrial DNA replication.
(a) Upregulation of SLP-2 expression in Jurkat T cells induces cardiolipin synthesis.
Upregulation of SLP-2 expression in T cells (SLP-2hi) (open bar) was induced with
doxycycline and was compared with baseline expression of SLP-2 (SLP-2lo) (filled bar).
The pool size of cardiolipin was determined and was expressed as a percent of the total
phospholipid phosphorus mass. (b) Cardiolipin levels in naïve (shaded histogram) and in
vitro-activated (filled histogram) PBMCs were measured by flow cytometry after staining
with nonyl-acridine orange. The result for the unstained control is shown by a dotted
line. (c and d) Incorporation of [3H]-acetate (c) and [14C]-linoleic acid (d) into
cardiolipin in SLP-2lo and SLP-2hi T cells. Data are shown as dpm/mg total cellular
protein. (e and f) Real-time PCR analysis of the transcript levels of phosphatidylglycerol
phosphate synthase (e) and cardiolipin synthase (f) in SLP-2lo and SLP-2hi T cells. (g)
The mitochondrial mass in naïve (shaded histograms) and in vitro-activated PBMC in the
absence (thin line) or presence (thick line) of triacsin C was measured by flow cytometry
after staining with MitoTracker Red. The result for the unstained control is shown by a
dotted line. (h and i) Real-time PCR analysis of the transcript levels of PGC-1α (h) and
mitochondrial DNA content (i) in SLP-2lo and SLP-2hi T cells. The results shown here
are representative of at least 3 independent experiments. *: P < 0.05.
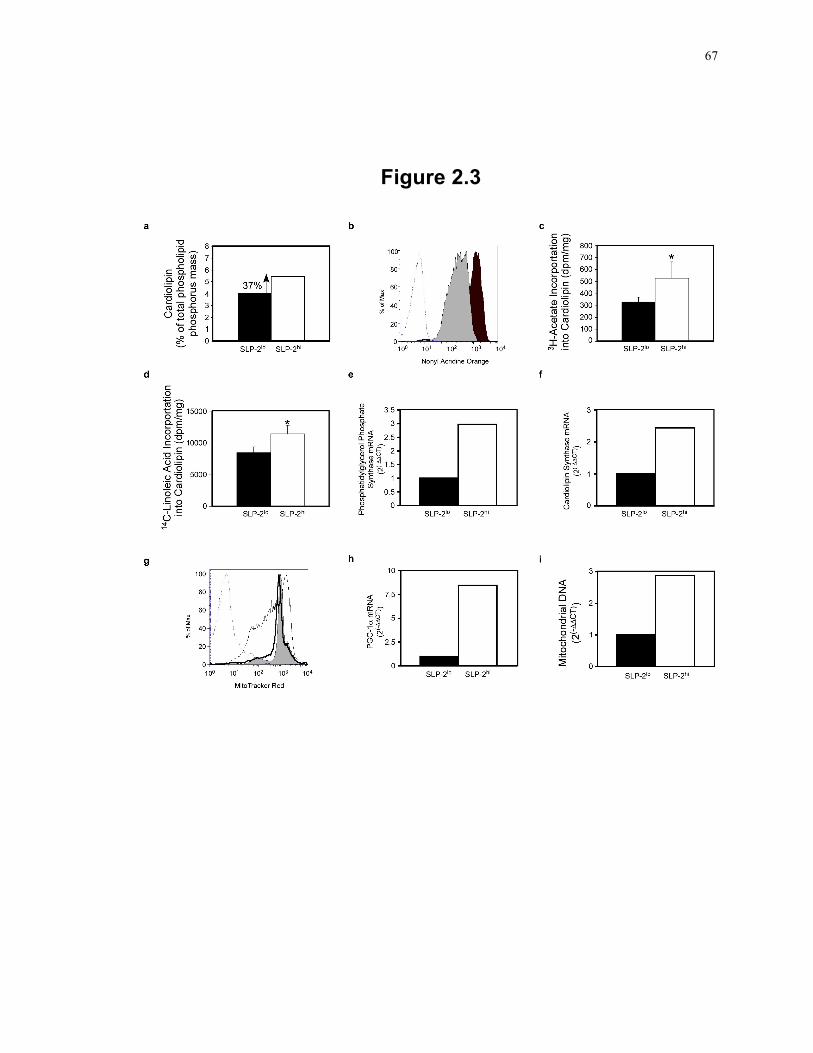
67
Figure 2.3

68
In addition to membrane formation, mitochondrial biogenesis also requires activation of a
nuclear transcription program that encodes the majority of mitochondrial proteins. This
nuclear transcriptional program is dependent on PGC-1α, a transcriptional co-activator
and master regulator of mitochondrial biogenesis 26. Thus, we performed real-time RT-
PCR to measure the levels of PGC-1α mRNA and found that these were increased in
SLP-2hi cells (Fig. 2.3h), supporting the idea that upregulation of SLP-2 expression is
associated with an increase in the expression of mitochondrially targeted genes. Of
interest, induction of SLP-2 expression also resulted in significant increases in the levels
of OPA-1 and mitofusin-2 (P < 0.05), both integral mitochondrial membrane proteins
associated with mitochondrial fusion, but not in the levels of Drp1 (involved in
mitochondrial fission), GAPDH, or cytochrome c (data not shown). However, the
increase in OPA-1 and mitofusin-2 did not translate functionally into enhanced
mitochondrial fusion.
Mitochondrial biogenesis requires replication of mitochondrial DNA. As expected from
the data presented above, we found that upregulation of SLP-2 correlated with a 70%
increase in the level of mitochondrial DNA (Fig. 2.3i). Taken together, these findings
lead us to conclude that higher levels of SLP-2 stimulate cardiolipin synthesis and
mitochondrial biogenesis.
To determine the kinetics of mitochondrial biogenesis in response to SLP-2 upregulation,
MitoTracker Red was used to measure mitochondrial mass in PBMCs stimulated with
PMA and ionomycin for 48 h. During the first 18 h of stimulation, the cells had a slight
decrease in mitochondrial mass, which was recovered by 24 h, and mitochondrial mass
was elevated by 48 h of stimulation (Fig. 2.4a). During this time, SLP-2 and PHB-1 were
both found to be upregulated as early as 1 h after stimulation, while PGC-1α was elevated
by 18 h poststimulation (Fig. 2.4b). This preceded the increase in mitochondrial
biogenesis detected by MitoTracker Red at 24 h poststimulation, implying that SLP-2
upregulation in response to T cell activation precedes PGC-1α upregulation and the
subsequent increase in mitochondrial biogenesis.

69
Figure 2.4: SLP-2 upregulation precedes PGC-1α upregulation and mitochondrial
biogenesis during T cell activation.
(a) PBMCs were stimulated with 1ng/mL PMA and 100ng/mL ionomycin for as long as
48 h. At the indicated time points, cells were stained with Mitotracker Red and were
analyzed for mitochondrial mass by flow cytometry. Shaded histograms represent
unstimulated cells while filled histograms represent stimulated cells. (b) Whole-cell
lysates were made at each indicated time point during stimulation with PMA and
ionomycin. The lysates were separated by SDS-PAGE and were serially blotted for
PGC-1α, PHB-1, SLP-2, and actin.
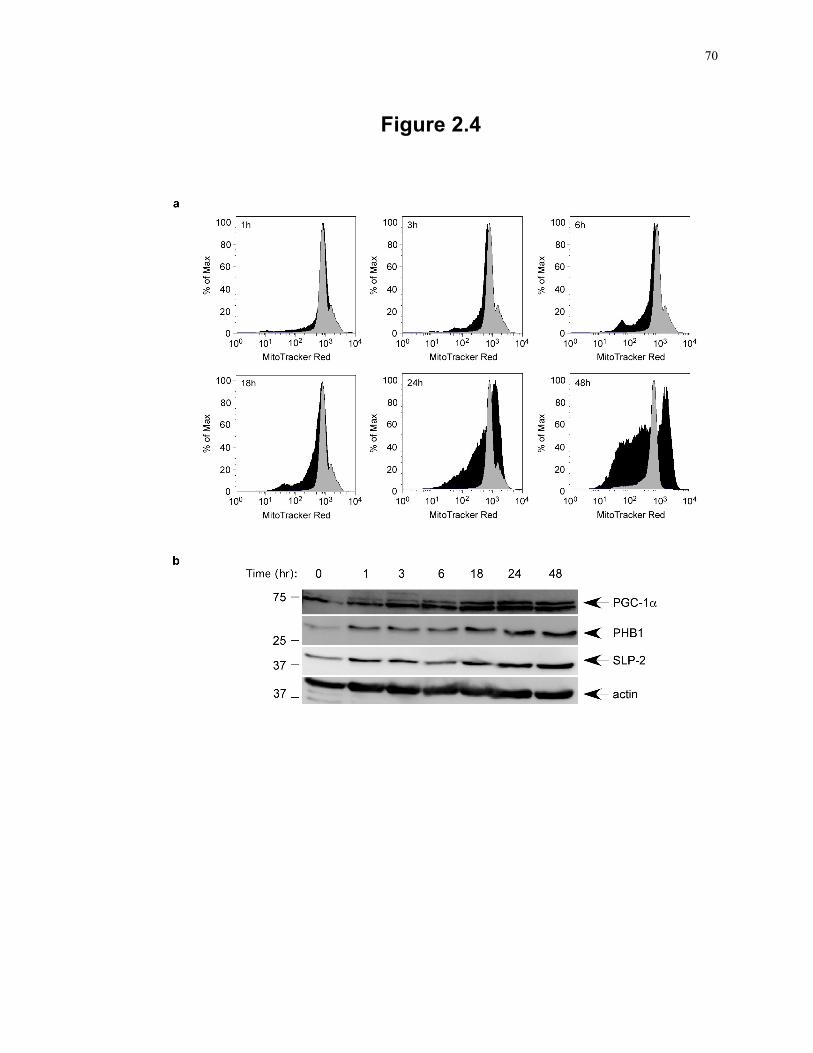
70
Figure 2.4

71
2.3.3 SLP-2 interacts with prohibitins and binds cardiolipin-enriched microdomains
Next, we examined how upregulation of SLP-2 expression may induce mitochondrial
biogenesis. Emerging data suggest that mitochondrial biogenesis is coordinated by a
mechanism involving PHB-1 and -2 7, 27. These proteins localize within phospholipid-
enriched, detergent-insoluble microdomains in mitochondrial membranes, and their
functional interactome includes genes such as Ups1 and Gep1 that are linked to the
synthesis of mitochondrial membrane phospholipids cardiolipin and
phosphatidylethanolamine, respectively 7, 27. We noticed that Saccharomyces cerevisiae
PHB-2 has a putative transmembrane domain (as predicted by the TMHMM
[transmembrane hidden Markov model] algorithm 28), but neither mouse nor human
PHBs have such a domain. Of interest, S. cerevisiae does not have a homolog of SLP-2,
the closest being PHB-1, with a clustal W score of 15 29. This suggested that, in
mammals, SLP-2 may act to link PHBs and phospholipid-enriched detergent-insoluble
microdomains in mitochondrial membranes. Such a possibility is consistent with the
observations that PHBs are upregulated in parallel with SLP-2 during activation 30 (Fig.
2.4b) and that SLP-2 interacts with PHBs 12.
We first confirmed that human SLP-2 interacted with PHB-1 and PHB-2 in T cells using
standard coprecipitation experiments (Fig. 2.5a). We found that immunoprecipitates of
SLP-2 contained both PHB-1 and PHB-2, as previously reported 12. Furthermore,
coprecipitation of PHB-1, PHB-2 and SLP-2 increased as the expression of SLP-2
increased with increasing levels of doxycycline in the culture (Fig. 2.5b). To demonstrate
that SLP-2 plays a role in linking prohibitins to phospholipid-enriched detergent-
insoluble microdomains, we isolated mitochondria from SLP-2lo and SLP-2hi cells and
extracted mitochondrial membranes with Na2CO3. We found a higher association of
PHB-1 with the mitochondrial membrane fraction in the SLP-2hi cells, although the total
level of PHB-1 was similar in the SLP-2lo and SLP-2hi cells (Fig. 2.5c).
Next, we generated human recombinant SLP-2 (hrSLP-2) and examined the profile of

72
Figure 2.5: SLP-2 interacts with prohibitins.
(a) Immunoprecipitates with an antiserum against human SLP-2 or with a pre-immune
serum from SLP-2-expressing Jurkat T cells were serially blotted for prohibitin 1 (PHB-
1), PHB-2 or SLP-2. WCL, whole cell lysate; IP, immunoprecipitating antiserum; Beads,
no cell lysate control; Pre-Imm, control immunoprecipitation with pre-immune serum.
(b) Immunoprecipitates and whole-cell lysates from SLP-2-GFP cells induced to express
increasing amounts of SLP-2 by increasing concentration of doxycycline were collected
and blotted as for Fig. 2.4b. (c) Mitochondria were isolated from SLP-2hi and SLP-2lo
cells and treated with Na2CO3 in order to examine the membrane association of
prohibitins in the presence of elevated SLP-2 expression. The fractions were serially
blotted for PHB-1 and SLP-2.

73
Figure 2.5

74
Figure 2.6: SLP-2 binds cardiolipin.
(a) Coprecipitation of hrSLP-2 with cardiolipin (CL) vesicles but not with other
phopholipid vesicles as shown by Western blotting of pellet (P) and supernatant (S)
fractions. PC, phosphatidylcholine; PE, phosphatidylethanolamine; PI,
phosphatidylinositol; PS, phosphatidylserine; PG, phosphatidylglycerol; Total protein,
hrSLP-2 aliquot as used for coprecipitation. (b) Titration of hrSLP-2 coprecipitation with
CL in mixed CL-PC-PE vesicles at the indicated ratios. An additional group containing
twice the amount of hrSLP-2 (cardiolipin + 2xSLP-2) was added.

75
Figure 2.6

76
interaction of this protein with different phospholipids using a phospholipid vesicle co-
precipitation assay 24. In this assay, molecules that interact with phospholipids
cosediment with the vesicles whereas non-interacting proteins remain in the supernatant.
We found that hrSLP-2 coprecipitated with cardiolipin vesicles (Fig. 2.6a, P [pellet
fraction]), whereas most of the hrSLP-2 remained in solution when vesicles made of
other phospholipids found in mitochondria (phosphatidylcholine and
phosphatidylethanolamine) or vesicles made of mostly non-mitochondrial phospholipids
(phosphatidylinositol and phosphatidylserine) were used (Fig. 2.6a; S [supernatant
fraction]). Of interest, hrSLP-2 bound to phosphatidylglycerol vesicles in lower amounts,
a finding likely due to the fact that cardiolipin is a composite of two phosphatidylglycerol
molecules. To further investigate the association of hrSLP-2 with cardiolipin, we
prepared phospholipid vesicles containing cardiolipin along with the two other major
phospholipids in mitochondria, phosphatidylcholine and phosphatidylethanolamine at
ratios of 1:1:1 (33% CL, 33% PC, 33% PE), 2.5:0.25:0.25 (83% CL, 8% PC, 8% PE) and
2.75:0.12:0.12 (92% CL, 4% PC, 4% PE). The interaction between hrSLP-2 and
cardiolipin was selective and titrated with the enrichment of this phospholipid in vesicles
also containing phosphatidylcholine and phosphatidylethanolamine (Fig. 2.6b). These
data demonstrate that SLP-2 binds to the prohibitin complex as well as to cardiolipin-
enriched membrane microdomains, potentially mediating the association between
prohibitins and mitochondrial membranes.
2.3.4 Upregulation of SLP-2 expression is associated with increased mitochondrial functions
To examine the functional effects of upregulation of SLP-2 expression, we first
determined the activity of citrate synthase, an enzyme of the citric acid cycle, which is
localized to the mitochondrial matrix. We found that SLP-2hi cells showed significantly
increased citrate synthase activity (P < 0.01), (Fig. 2.7a). Next, we measured the activity
of the electron transport complexes in the respiratory chain. We found that SLP-2hi cells
had significantly higher activities of NADH dehydrogenase and succinate dehydrogenase
(P < 0.05), complex I and II of the electron transport chain (Fig. 2.7b and 2.7c). In

77
Figure 2.7: Induction of SLP-2 expression is associated with higher mitochondrial
activity.
(a) Citrate synthase (CS) activity was measured in SLP-2lo (filled bar) and SLP-2hi (open
bar) Jurkat T cells. Data are means and standard deviations (SD) for triplicate samples;
**P < 0.01. Data are expressed as nmol/min/mg mitochondrial protein. (b) NADH
dehydrogenase was measured in SLP-2lo (filled bar) and SLP-2hi (open bar) Jurkat T
cells, plotted as in panel a, *P < 0.05. (c) Succinate dehydrogenase (SDH) was measured
in SLP-2lo and SLP-2hi Jurkat T cells, plotted as in panel a; *P < 0.05. Data are expressed
as nmol/min/mg mitochondrial protein. (d) Mitochondrial and total cellular ATP levels
were measured in SLP-2lo and SLP-2hi Jurkat T cells. Means and standard deviations
(SDs) for triplicate samples are plotted. *P < 0.05, **P < 0.01. (e and f) IL-2 levels in
culture supernatants of SLP-2lo (e) and SLP-2hi (f) Jurkat T cells pre-treated or not with
oligomycin (4 µM) and stimulated with antigen-presenting cells and SEE for 24 h.
Means and standard deviations for triplicate samples are plotted. Asterisks indicate
significant differences (*P < 0.05; ***P < 0.001) as determined by analysis of variance
(ANOVA) with a Bonferroni posttest. (g) Oxygen consumption was measured in SLP-2lo
and SLP-2hi T cells in media containing high (11mM) or low (0.4mM) glucose. Means
and standard deviations for triplicate samples are plotted. Asterisks indicate significant
differences (*P < 0.05; ***P < 0.001) as determined by analysis of variance (ANOVA)
with a Bonferroni posttest. (h) Mitochondria were isolated from SLP-2lo and SLP-2hi
cells, separated by SDS-PAGE, and serially blotted for subunits of each complex of the
respiratory chain as well as SLP-2.
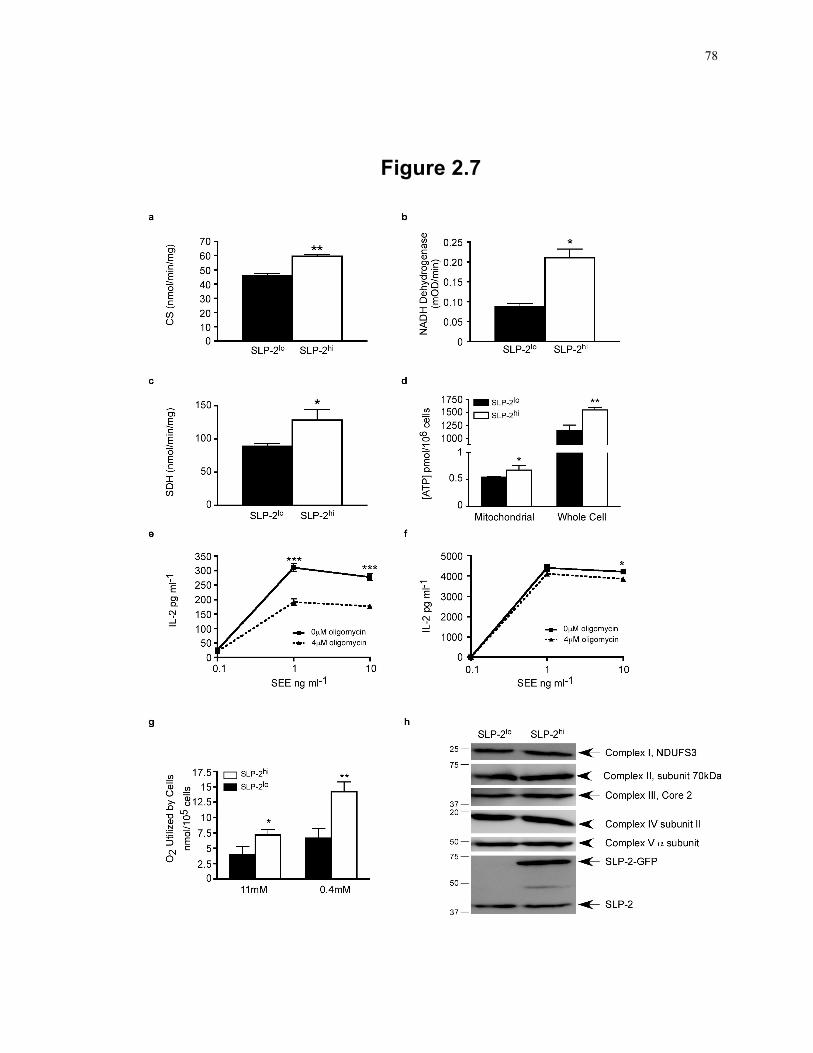
78
Figure 2.7

79
addition, mitochondrial and whole-cell ATP levels in resting SLP-2hi T cells were
significantly higher than those in SLP-2lo T cells (P < 0.05) (Fig. 2.7d). Such an increase
in ATP levels in SLP-2hi T cells correlated with increased resistance to depletion of
mitochondrial ATP with oligomycin, as evidenced by increased T cell responses,
indicated by IL-2 production. In these experiments, oligomycin-treated SLP-2lo T cells
and SLP-2hi T cells were stimulated with antigen-presenting cells and staphylococcal
enterotoxin E (SEE), and the production of interleukin-2 (IL-2) was determined after 24 h
of stimulation. Oligomycin treatment significantly reduced the IL-2 response to SEE of
SLP-2lo T cells (P < 0.001) (Fig. 2.7e) but did not affect the response of SLP-2hi T cells
(Fig. 2.7f). Consistent with our previous findings, SLP-2hi T cells had a higher IL-2
response than SLP-2lo T cells 11.
Given that the generation of ATP by OXPHOS is linked to the respiratory chain in the
mitochondrial inner membrane, we examined the effect of modulating SLP-2 expression
on oxygen consumption by T cells. We found that SLP-2hi T cells had slightly but
significantly higher oxygen consumption than SLP-2lo T cells (P < 0.05). In a low-
glucose medium, SLP-2hi T cells had much higher oxygen consumption than SLP-2lo T
cells (P < 0.01) (Fig. 2.7g). This is expected, since cells rely more heavily on OXPHOS
under low-glucose conditions. Therefore, we concluded that upregulation of SLP-2
expression is associated with increased oxygen consumption and OXPHOS, leading to
increased ATP stores in the cell.
Cardiolipin plays an essential role in the mitochondrial protein import machinery 31, and
as such, the increase in cardiolipin seen in response to SLP-2 upregulation could lead to
an increased rate of protein transport into the mitochondria. If so, the increased
enzymatic rates we detected in the SLP-2hi cells could be due to elevated protein levels in
the mitochondria. To test this, mitochondria were isolated from SLP-2lo and SLP-2hi
cells, and lysates from these mitochondria were blotted for representative subunits of
each complex of the respiratory chain. SLP-2lo and SLP-2hi cells had similar protein
levels for each complex of the respiratory chain (Fig. 2.7h), indicating that the increase in

80
enzymatic activity seen in the SLP-2-overexpressing cells was not due to increased levels
or increased translocation of electron transport complexes into mitochondria.
2.3.5 Upregulation of SLP-2 expression increases T cell responses and resistance to apoptosis
In light of the significantly enhanced mitochondrial biogenesis and function associated
with SLP-2 upregulation, we examined the physiological effects of these changes by
looking at the functional response of primary human T cells in which SLP-2 expression
was upregulated. As shown in Fig. 2.8a, peripheral blood T cells from normal volunteers
transiently nucleofected with doxycycline-inducible SLP-2-GFP cDNA and expressing
high levels of SLP-2 produced significantly higher levels of IL-2 (P < 0.05) upon
stimulation with SEE and antigen-presenting cells. In addition, these cells showed
increased resistance to apoptosis through the intrinsic pathway (Fig. 2.8b). Such
resistance to apoptosis was also observed in response to actinomycin D in Jurkat SLP-2hi
T cells (Fig. 2.8c). As an additional marker of apoptosis, actinomycin D-induced cell
death was also measured by an increase in annexin V labeling. In agreement with the
loss of transmembrane potential as measured by MitoTracker Red, SLP-2hi cells also
showed lower annexin V staining, which is indicative of a protection against the
induction of the intrinsic apoptosis pathway (Fig. 2.8d and e). Thus, consistent with the
increased mitochondrial biogenesis and function, upregulation of SLP-2 expression
enhances cell function.

81
Figure 2.8: SLP-2 upregulation increases T cell responses and protects against
apoptosis.
(a) Human peripheral blood T cells nucleofected with doxycycline-inducible SLP-2-GFP
and expressing high levels of SLP-2 (SLP-2hi) upon doxycycline-induction produce
significantly more IL-2 in response to antigen-presenting cells and SEE stimulation than
control (noninduced) SLP-2-GFP transfected T cells. **P < 0.01; * P < 0.05. (b) SLP-
2hi primary T cells are less susceptible to actinomycin D-induced apoptosis. Human
PBMCs were transiently transfected with doxycycline-inducible SLP-2-GFP and cultured
with (SLP-2hi) (shaded histogram) or without (SLP-2lo) (open histogram) doxycycline
and with actinomycin D (5µg/mL) for 3 h at 37°C. Cell death was measured by
MitoTracker Red staining, which was analyzed by flow cytometry. (c) Upregulation of
SLP-2 expression in doxycycline-inducible SLP-2-GFP transfected Jurkat T cells results
in increased resistance to actinomycin D-induced apoptosis. SLP-2hi (shaded histogram)
and SLP-2lo (open histogram) T cells were treated with 5µg/mL actinomycin D for 3 h at
37°C. (d) SLP-2hi (open bars) and SLP-2lo (filled bars) T cells were treated with
actinomycin D and stained for annexin V, an alternative marker of apoptosis.
***P<0.001. (e) One representative flow cytometry plot of annexin V staining of SLP-
2hi (shaded histogram) and SLP-2lo (open histogram) cells after induction of apoptosis
with actinomcyin D. The result for the unstained control is shown by a dotted line. The
data are representative of three independent experiments.
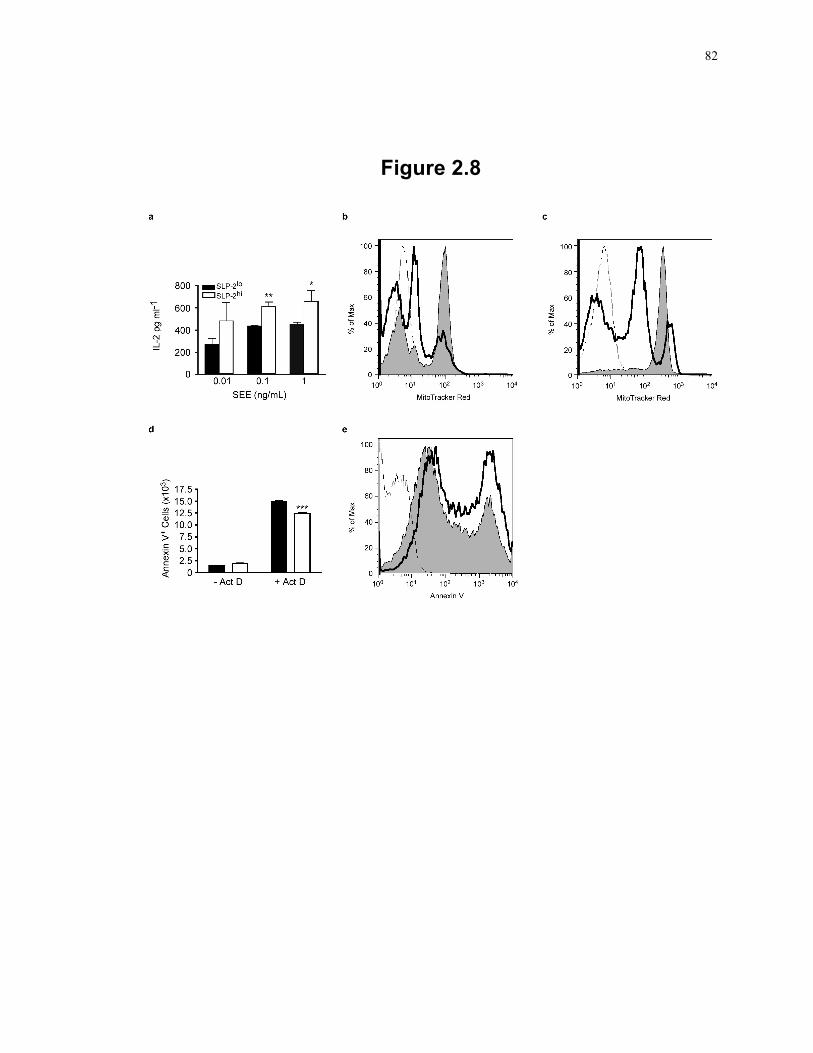
82
Figure 2.8

83
2.4 Discussion The functions of stomatins and other proteins sharing the
stomatin/prohibitin/flotillin/Hflk (SPFH) domain remain enigmatic. It has been proposed
that through this domain, stomatins, PHBs and flotillins interact with different cell
membranes and facilitate signalling. We report here that SLP-2, a mitochondrial member
of the stomatin family, binds cardiolipin and may help to recruit PHBs to cardiolipin-
enriched microdomains. Furthermore, upregulation of SLP-2 expression is associated
with increased mitochondrial biogenesis and function, ultimately resulting in enhanced
effector cell function.
Our data show that SLP-2 binds cardiolipin-enriched microdomains in a way that titrates
with the content of cardiolipin in phospholipid membranes. The mitochondrial
membrane is composed of 40% phosphatidylcholine, 30% phosphatidylethanolamine and
10-15% phosphatidylinositol and cardiolipin 32. But the cardiolipin content in the
mitochondrial membrane is not homogenously distributed. Our data suggest that SLP-2
can bind to clusters of cardiolipin, i.e., microdomains with a much higher localized
cardiolipin concentration. This selective binding of SLP-2 to cardiolipin is reminiscent
of the observation that two other members of the stomatin/PHB/flotillin/Hflk superfamily
(MEC-2 and podocin) bind cholesterol 33 and leads us to propose that SPFH domain-
containing proteins bind to selective phospholipid-enriched membrane microdomains and
may act as markers that recruit other proteins to the lipid microdomains. How these
proteins bind lipids is still unknown. The binding of SLP-2 to cardiolipin in the
mitochondrial inner membrane is likely independent of transmembrane insertion, since
SLP-2 lacks a putative transmembrane domain 8.
Upregulation of SLP-2 expression increases cardiolipin synthesis and mitochondrial
biogenesis. Recent evidence indicates that these two processes are tightly coordinated 27,
likely through the PHB functional interactome, which includes genes involved in
phospholipid synthesis, such as Ups1, whose deletion, in yeast, translates into a marked
decreased in cardiolipin content 7. Although we have not shown a direct interaction
among PHBs, SLP-2 and cardiolipin, our data suggest that SLP-2 may bring PHBs to

84
cardiolipin and may contribute to the organization of cardiolipin-enriched microdomains
in the mitochondrial inner membrane 34. Data from our laboratory using T cell-specific
SLP-2 knockout mice seem to corroborate this model (Chapter 3). This possible
mechanism accommodates the upregulation of both SLP-2 and PHBs during T cell
activation 30 and the detection of both proteins in detergent-insoluble microdomains of
cell membranes (i.e., cardiolipin-enriched microdomains in the mitochondrial inner
membrane and cholesterol-enriched microdomains in the plasma membrane) 11, 35-37. In
this context, an increase in SLP-2 levels as a result of cell activation would increase the
interaction of PHBs with cardiolipin in the mitochondrial inner membrane and activate
the PHB functional interactome, leading to de novo cardiolipin synthesis.
Using human T cell activation as a model, we have shown that upregulation of SLP-2
expression is a molecular step between mitochondrial biogenesis and cell activation and
function. SLP-2 does not seem to be critical under resting conditions, as suggested by the
absence or very low level of SLP-2 expression in resting naïve T cells. However, during
T cell activation, SLP-2 expression is dramatically upregulated, and this increases T cell
function, leading to increased IL-2 production and increased resistance to apoptosis
through the intrinsic pathway. PHBs are also upregulated during T cell activation 30,
whereas PHB-2 deletion decreases the ability of cells to proliferate and increases their
sensitivity to apoptosis through the intrinsic pathway 38. These observations corroborate
the model proposed here by which SLP-2 - PHB complexes would act in concert to
regulate mitochondrial biogenesis and function during T cell activation. Of interest,
modulation of PHB-2 levels also alters OPA-1 levels (e.g., cells lacking PHB-2 have
impaired processing of OPA-1 34), and in our experiments, T cells overexpressing SLP-2
have increased OPA-1 levels.
The increased mitochondrial function observed in SLP-2 overexpressing cells is in line
with the observation that SLP-2 controls the stability and the assembly of electron
transport complexes 39, 40. This is further corroborated by emerging data from our
laboratory using T cell-specific SLP-2-deficient mice, showing that a lack of SLP-2 is
associated with decreased recruitment of PHB-1 to cardiolipin-enriched mitochondrial
membranes, as well as a loss of complex I subunits (Chapter 3). Optimal assembly of

85
these complexes may require the formation of specialized membrane microdomains, and
this would correlate with increased respiratory chain function. Of interest, SLP-2 is
required for mitochondrial hyperfusion 41, a phenomenon linked with increased metabolic
demands and cellular stress. Levels of both OPA-1 and mitofusin-2, proteins required
for this process, were increased upon SLP-2 upregulation, but cytochrome c levels were
not. It is possible that SLP-2 expression specifically increases the expression of proteins
involved in mitochondrial fusion, or prevents these proteins from degradation, without
altering expression of other mitochondrial proteins.
Taken together, the evidence presented here leads us to propose that binding of SLP-2 to
cardiolipin recruits PHBs, helping to form cardiolipin-enriched membrane microdomains
in which the respiratory chain components are optimally assembled. The formation of
these microdomains is likely the result of the self-oligomerizing properties of SLP-2 8.
The increase in respiratory chain function may then induce further cardiolipin synthesis
through its effect on the pH gradient across the mitochondrial inner membrane 42,
enhancing mitochondrial biogenesis. The result of this proposed mechanism is the
provision of the enhanced mitochondrial function required for an optimal T cell response.
How the increased mitochondrial membrane formation increases nuclear transcription
and mitochondrial DNA replication, both required to increase mitochondrial numbers,
remains to be determined. A retrograde nuclear signalling pathway in yeast is starting to
emerge, but little is known about this pathway in mammalian cells. Specific signals may
involve mitochondrial stress, since it has been shown that mitochondrial dysfunction
increases nuclear transcription 26. In this context, SLP-2 may play a role by regulating
mitochondrial membrane biogenesis and its subsequent effect of calcium-dependent
signalling, which has been linked to the activation of retrograde signalling 43.
In summary, we have shown that increased levels of SLP-2, seen, for example, during
lymphocyte activation, stimulate mitochondrial biogenesis and function, providing for the
energetic requirements of activation. Such a function involves the binding of SLP-2 and
its interacting partners (PHBs) to cardiolipin to form membrane microdomains in which

86
respiratory complexes are optimally assembled. The ultimate result of this function is the
net enhancement of effector cell functions.

87
2.5 References
1. Frauwirth, K.A., Riley, J.L., Harris, M.H., Parry, R.V., Rathmell, J.C., Plas, D.R., Elstrom, R.L., June, C.H. & Thompson, C.B. The CD28 signaling pathway regulates glucose metabolism. Immunity 16, 769-777 (2002).
2. Frauwirth, K.A. & Thompson, C.B. Regulation of T lymphocyte metabolism. J Immunol 172, 4661-4665 (2004).
3. Fox, C.J., Hammerman, P.S. & Thompson, C.B. Fuel feeds function: energy metabolism and the T-cell response. Nat Rev Immunol 5, 844-852 (2005).
4. Carpentieri, U. & Sordahl, L.A. Respiratory and calcium transport functions of mitochondria isolated from normal and transformed human lymphocytes. Cancer Res 40, 221-224 (1980).
5. Nagy, G., Barcza, M., Gonchoroff, N., Phillips, P.E. & Perl, A. Nitric oxide-dependent mitochondrial biogenesis generates Ca2+ signaling profile of lupus T cells. J Immunol 173, 3676-3683 (2004).
6. D'Souza, A.D., Parikh, N., Kaech, S.M. & Shadel, G.S. Convergence of multiple signaling pathways is required to coordinately up-regulate mtDNA and mitochondrial biogenesis during T cell activation. Mitochondrion 7, 374-385 (2007).
7. Osman, C., Haag, M., Potting, C., Rodenfels, J., Dip, P.V., Wieland, F.T., Brugger, B., Westermann, B. & Langer, T. The genetic interactome of prohibitins: coordinated control of cardiolipin and phosphatidylethanolamine by conserved regulators in mitochondria. J Cell Biol 184, 583-596 (2009).
8. Wang, Y. & Morrow, J.S. Identification and characterization of human SLP-2, a novel homologue of stomatin (band 7.2b) present in erythrocytes and other tissues. J Biol Chem 275, 8062-8071 (2000).
9. Owczarek, C.M., Treutlein, H.R., Portbury, K.J., Gulluyan, L.M., Kola, I. & Hertzog, P.J. A novel member of the STOMATIN/EPB72/mec-2 family, stomatin-like 2 (STOML2), is ubiquitously expressed and localizes to HSA chromosome 9p13.1. Cytogenet Cell Genet 92, 196-203 (2001).
10. Green, J.B. & Young, J.P. Slipins: ancient origin, duplication and diversification of the stomatin protein family. BMC evolutionary biology 8, 44 (2008).
11. Kirchhof, M.G., Chau, L.A., Lemke, C.D., Vardhana, S., Darlington, P.J., Marquez, M.E., Taylor, R., Rizkalla, K., Blanca, I., Dustin, M.L. & Madrenas, J. Modulation of T cell activation by stomatin-like protein 2. J Immunol 181, 1927-1936 (2008).

88
12. Da Cruz, S., Parone, P.A., Gonzalo, P., Bienvenut, W.V., Tondera, D., Jourdain, A., Quadroni, M. & Martinou, J.C. SLP-2 interacts with prohibitins in the mitochondrial inner membrane and contributes to their stability. Biochim Biophys Acta 1783, 904-911 (2008).
13. Baroja, M.L., Luxenberg, D., Chau, T., Ling, V., Strathdee, C.A., Carreno, B.M. & Madrenas, J. The inhibitory function of CTLA-4 does not require its tyrosine phosphorylation. J Immunol 164, 49-55 (2000).
14. Arp, J., Kirchhof, M.G., Baroja, M.L., Nazarian, S.H., Chau, T.A., Strathdee, C.A., Ball, E.H. & Madrenas, J. Regulation of T-cell activation by phosphodiesterase 4B2 requires its dynamic redistribution during immunological synapse formation. Mol Cell Biol 23, 8042-8057 (2003).
15. Hauff, K., Linda, D. & Hatch, G.M. Mechanism of the elevation in cardiolipin during HeLa cell entry into the S-phase of the human cell cycle. Biochem J 417, 573-582 (2009).
16. Lowry, O.H., Rosebrough, N.J., Farr, A.L. & Randall, R.J. Protein measurement with the Folin phenol reagent. J Biol Chem 193, 265-275 (1951).
17. Rouser, G., Fkeischer, S. & Yamamoto, A. Two dimensional thin layer chromatographic separation of polar lipids and determination of phospholipids by phosphorus analysis of spots. Lipids 5, 494-496 (1970).
18. Hatch, G.M. & McClarty, G. Regulation of cardiolipin biosynthesis in H9c2 cardiac myoblasts by cytidine 5'-triphosphate. J Biol Chem 271, 25810-25816 (1996).
19. Livak, K.J. & Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 25, 402-408 (2001).
20. Cote, H.C., Brumme, Z.L., Craib, K.J., Alexander, C.S., Wynhoven, B., Ting, L., Wong, H., Harris, M., Harrigan, P.R., O'Shaughnessy, M.V. & Montaner, J.S. Changes in mitochondrial DNA as a marker of nucleoside toxicity in HIV-infected patients. N Engl J Med 346, 811-820 (2002).
21. Bueno, C., Lemke, C.D., Criado, G., Baroja, M.L., Ferguson, S.S., Rahman, A.K., Tsoukas, C.D., McCormick, J.K. & Madrenas, J. Bacterial superantigens bypass Lck-dependent T cell receptor signaling by activating a Galpha11-dependent, PLC-beta-mediated pathway. Immunity 25, 67-78 (2006).
22. Hajek, P., Chomyn, A. & Attardi, G. Identification of a novel mitochondrial complex containing mitofusin 2 and stomatin-like protein 2. J Biol Chem 282, 5670-5681 (2007).

89
23. Chau, T.A., McCully, M.L., Brintnell, W., An, G., Kasper, K.J., Vines, E.D., Kubes, P., Haeryfar, S.M., McCormick, J.K., Cairns, E., Heinrichs, D.E. & Madrenas, J. Toll-like receptor 2 ligands on the staphylococcal cell wall downregulate superantigen-induced T cell activation and prevent toxic shock syndrome. Nat Med 15, 641-648 (2009).
24. Bakolitsa, C., de Pereda, J.M., Bagshaw, C.R., Critchley, D.R. & Liddington, R.C. Crystal structure of the vinculin tail suggests a pathway for activation. Cell 99, 603-613 (1999).
25. Jacobson, J., Duchen, M.R. & Heales, S.J. Intracellular distribution of the fluorescent dye nonyl acridine orange responds to the mitochondrial membrane potential: implications for assays of cardiolipin and mitochondrial mass. J Neurochem 82, 224-233 (2002).
26. Scarpulla, R.C. Transcriptional paradigms in mammalian mitochondrial biogenesis and function. Physiol Rev 88, 611-638 (2008).
27. Gohil, V.M. & Greenberg, M.L. Mitochondrial membrane biogenesis: phospholipids and proteins go hand in hand. J Cell Biol 184, 469-472 (2009).
28. Krogh, A., Larsson, B., von Heijne, G. & Sonnhammer, E.L. Predicting transmembrane protein topology with a hidden Markov model: application to complete genomes. J Mol Biol 305, 567-580 (2001).
29. Thompson, J.D., Higgins, D.G. & Gibson, T.J. CLUSTAL W: improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res 22, 4673-4680 (1994).
30. Ross, J.A., Nagy, Z.S. & Kirken, R.A. The PHB1/2 phosphocomplex is required for mitochondrial homeostasis and survival of human T cells. J Biol Chem 283, 4699-4713 (2008).
31. Joshi, A.S., Zhou, J., Gohil, V.M., Chen, S. & Greenberg, M.L. Cellular functions of cardiolipin in yeast. Biochim Biophys Acta 1793, 212-218 (2008).
32. Osman, C., Voelker, D.R. & Langer, T. Making heads or tails of phospholipids in mitochondria. J Cell Biol 192, 7-16 (2011).
33. Huber, T.B., Schermer, B., Muller, R.U., Hohne, M., Bartram, M., Calixto, A., Hagmann, H., Reinhardt, C., Koos, F., Kunzelmann, K., Shirokova, E., Krautwurst, D., Harteneck, C., Simons, M., Pavenstadt, H., Kerjaschki, D., Thiele, C., Walz, G., Chalfie, M. & Benzing, T. Podocin and MEC-2 bind cholesterol to regulate the activity of associated ion channels. Proc Natl Acad Sci U S A 103, 17079-17086 (2006).

90
34. Artal-Sanz, M. & Tavernarakis, N. Prohibitin and mitochondrial biology. Trends Endocrinol Metab 20, 394-401 (2009).
35. Mielenz, D., Vettermann, C., Hampel, M., Lang, C., Avramidou, A., Karas, M. & Jack, H.M. Lipid rafts associate with intracellular B cell receptors and exhibit a B cell stage-specific protein composition. J Immunol 174, 3508-3517 (2005).
36. Kim, K.B., Lee, J.W., Lee, C.S., Kim, B.W., Choo, H.J., Jung, S.Y., Chi, S.G., Yoon, Y.S., Yoon, G. & Ko, Y.G. Oxidation-reduction respiratory chains and ATP synthase complex are localized in detergent-resistant lipid rafts. Proteomics 6, 2444-2453 (2006).
37. Foster, L.J., De Hoog, C.L. & Mann, M. Unbiased quantitative proteomics of lipid rafts reveals high specificity for signaling factors. Proc Natl Acad Sci U S A 100, 5813-5818 (2003).
38. Merkwirth, C., Dargazanli, S., Tatsuta, T., Geimer, S., Lower, B., Wunderlich, F.T., von Kleist-Retzow, J.C., Waisman, A., Westermann, B. & Langer, T. Prohibitins control cell proliferation and apoptosis by regulating OPA1-dependent cristae morphogenesis in mitochondria. Genes Dev 22, 476-488 (2008).
39. Houtkooper, R.H. & Vaz, F.M. Cardiolipin, the heart of mitochondrial metabolism. Cell Mol Life Sci 65, 2493-2506 (2008).
40. Acin-Perez, R., Fernandez-Silva, P., Peleato, M.L., Perez-Martos, A. & Enriquez, J.A. Respiratory active mitochondrial supercomplexes. Mol Cell 32, 529-539 (2008).
41. Tondera, D., Grandemange, S., Jourdain, A., Karbowski, M., Mattenberger, Y., Herzig, S., Da Cruz, S., Clerc, P., Raschke, I., Merkwirth, C., Ehses, S., Krause, F., Chan, D.C., Alexander, C., Bauer, C., Youle, R., Langer, T. & Martinou, J.C. SLP-2 is required for stress-induced mitochondrial hyperfusion. EMBO J 28, 1589-1600 (2009).
42. Gohil, V.M., Hayes, P., Matsuyama, S., Schagger, H., Schlame, M. & Greenberg, M.L. Cardiolipin biosynthesis and mitochondrial respiratory chain function are interdependent. J Biol Chem 279, 42612-42618 (2004).
43. Biswas, G., Adebanjo, O.A., Freedman, B.D., Anandatheerthavarada, H.K., Vijayasarathy, C., Zaidi, M., Kotlikoff, M. & Avadhani, N.G. Retrograde Ca2+ signaling in C2C12 skeletal myocytes in response to mitochondrial genetic and metabolic stress: a novel mode of inter-organelle crosstalk. EMBO J 18, 522-533 (1999).

91
Chapter 3 2
3 SLP-2 deficiency in T cells is associated with abnormal mitochondrial membrane compartmentalization of cardiolipin, decreased mitochondrial respiration and impaired CD4+ T cell responses
Stomatin-like protein 2 (SLP-2) is a mostly mitochondrial protein that regulates
mitochondrial biogenesis and function and modulates T cell activation. To determine the
mechanism of action of SLP-2, we generated T cell-specific SLP-2-deficient mice. These
mice had normal numbers of thymocytes and T cells in the periphery. However,
conventional SLP-2-deficient T cells had a post-transcriptional defect in IL-2 production
in response to TCR ligation, even under conditions of non-limiting glycolysis, and this
translated into reduced CD4+ T cell responses. SLP-2 deficiency was associated with
impaired cardiolipin compartmentalization in mitochondrial membranes, decreased levels
of the NDUFS3, NDUFB8 and NDUFA9 subunits of complex I of the respiratory chain
and subsequent decreased activity of this complex as well as of complex II + III of this
chain. Based on these results, we propose that SLP-2 organizes mitochondrial membrane
compartmentalization of cardiolipin, which is required for optimal assembly and function
of respiratory chain complexes. This function, in T cells, helps to ensure proper
metabolic response during activation.
3.1 Introduction
Stomatin-like protein 2 (SLP-2) is an abundant protein in the proteome of detergent-
insoluble microdomains of mitochondrial membranes 1. It is a member of the highly
conserved stomatin family, which is composed of stomatin, SLP-1, SLP-2 and SLP-3 2-7,
2 The material contained in this chapter is based on a submitted manuscript: Christie, D., Lian, D., Wang,
H., Hatch, G. and Madrenas, J. SLP-2 deficiency in T cells is associated with abnormal mitochondrial compartmentalization of cardiolipin, decreased mitochondrial respiration and impaired CD4+ T cell responses. 2012

92
and as such, is part of the SPFH superfamily that also includes prohibitins (PHB),
flotillins and the bacterial proteins HflC and HflK 8, 9. The function of these proteins has
been linked to the regulation of assembly and function of complex multi-chain receptors 2, 10-12. In line with this proposed function, we have recently shown that SLP-2 binds
cardiolipin and promotes the formation of specialized cardiolipin-enriched microdomains
that could facilitate the assembly of protein complexes within mitochondrial membranes 12.
SLP-2 can be detected in many tissues including lymph nodes, thymus and tonsils. In
addition, it is expressed at low levels in human peripheral blood mononuclear cells
(PBMC) including T cells, B cells and monocytes and its expression is upregulated upon
activation in vitro 1. Consistent with this distribution, it has been shown that SLP-2 plays
a role in T cell activation 1: upregulation of SLP-2 correlates with enhanced activation
and functional responses, whereas downregulation of SLP-2 expression is associated with
decreased duration of TCR signaling as well as a decrease in T cell activation.
To study the mechanism of action of SLP-2 in vivo, we initially generated a SLP-2
knockout mouse. However, this mouse was embryonic lethal at the pre-implantation
stage (C.D. Lemke and J. Madrenas, unpublished observation). To circumvent this
limitation, we generated a T cell-specific SLP-2 knockout mouse model. Using this
model, we report here that SLP-2-deficient T cells show abnormal cardiolipin
compartmentalization in mitochondrial membranes, decreased levels of the NDUFS3,
NDUFB8 and NDUFA9 subunits of complex I of the electron transport chain and
decreased activity of this complex, as well as decreased activity of complexes II + III of
this chain. This defect is associated with a selective post-transcriptional reduction in IL-2
production and decreased responses to stimulation through the TCR in vivo and ex vivo.
We propose that SLP-2 contributes to T cell activation primarily by regulating the
compartmentalization of cardiolipin in mitochondrial membranes and ensuring proper
assembly and function of the respiratory chain components in these membranes. This, in
turn, ensures optimal metabolic response during T cell activation.

93
3.2 Materials and Methods
3.2.1 Mice
A full length genomic fragment containing the mouse SLP-2 gene was cloned within the
lox sites of the 3LoxP3NwCD vector. Upon sequence confirmation, the plasmid was
electroporated into C57BL/6 embryonic stem cells and clones were selected for
neomycin resistance. The resistant clones were screened by Southern blot and PCR
analysis to confirm homologous recombination of the SLP-2-floxed DNA sequence.
Clones containing the SLP-2-floxed sequence were injected into B6/Try blastocysts and
implanted into a pseudo-pregnant mouse. The offspring was selected by chimeric coat
colour and backcrossed with C57BL/6 mice to produce SLP-2lox/wt mice in the C57BL/6
background. SLP-2 floxed mice were crossed with C57BL/6 mice transgenic for Cre
recombinase under the control of the CD4 promoter, purchased from Taconic (Hudson,
NY) 13, to generate a T cell-specific knockout of SLP-2 strain (SLP-2 T-K/O). SLP-2
floxed mice lacking Cre were used as control mice. Breeding colonies were derived from
the same original SLP-2 floxed breeders and kept in parallel. C3H/HeJ mice were
obtained from Jackson Labs. Mice were maintained in the animal facility at the
University of Western Ontario with approval from the Animal Use Subcommittee in
accordance with the Canadian Council on Animal Care Guidelines.
3.2.2 Confirmation of SLP-2 deletion
T cells were isolated by magnetic separation using the MACS pan T cell isolation kit
(Miltenyi Biotec, Auburn, CA). To confirm deletion at the genetic level, total genomic
DNA was extracted using the Qiagen DNeasy blood and tissue kit (Qiagen, Mississauga,
ON) and full length SLP-2 PCR was performed using the following primers: forward
primer 5’ ACTTCCACCCTTCAGTCCAGGTCG 3’, reverse primer 5’
ACTTGGATTCTGTGAAAGCAGACAC 3’. Samples were separated by DNA
electrophoresis in a 1% agarose gel and visualized by ethidium bromide staining. To
confirm deletion at the protein level, thymocytes and T cells were pelleted in PBS
containing sodium orthovanadate (400 μM) and EDTA (400 μM) and resuspended in
lysis buffer (1% Triton X-100, 150 mM NaCl, 10 mM Tris (pH7.6), 5 mM EDTA, 1 mM

94
sodium orthovanadate, 10 μg/mL leupeptin, 10 μg/mL aprotinin, 25 μM p-nitrophenyl-p’
guanidinobenzoate) at 4ºC for 30 min. These lysis conditions are sufficient to disrupt all
cellular membranes, excluding the nuclear membrane. Lysates were then cleared of
debris by centrifugation (10,700 x g, 4ºC, 10 min), mixed with sample buffer containing
β-mercaptoethanol and boiled. Cell lysates were resolved by standard SDS-PAGE and
western blotting with rabbit polyclonal anti-SLP-2 antisera (Proteintech Group, Chicago,
IL) and mouse monoclonal anti-GAPDH antibody (Chemicon International, Temecula,
CA). Detection of proteins after western blotting was performed using the BM
Chemiluminescence Substrate POD system (Roche, Laval, QC, Canada).
3.2.3 Isolation of detergent-insoluble fraction of mitochondrial membranes
Intact mitochondria were isolated from primed T cells using the QproteomeTM
Mitochondria Isolation Kit (Qiagen). Detergent-insoluble fractions were isolated by
incubation on ice in 25 mM HEPES buffer with 0.1% triton-X, as previously described 14.
Fractions were analyzed by western blotting or thin layer chromatography.
3.2.4 Measurement of cardiolipin
Soluble and insoluble mitochondrial fractions were washed in 2 ml of methanol:water
(1:1, by vol) and 25 µL aliquots were taken for protein determination 15. Subsequently,
0.5 ml of water and 2 ml chloroform were added to initiate phase separation. Samples
were centrifuged at 2000 rpm for 10 min and the upper phase was aspirated. Two ml of
theoretical upper phase (methanol:0.9% NaCl:chloroform) (48:47:3, by vol) was added
and centrifugation was repeated for 5 min. The organic phase was removed and dried
under nitrogen gas and resuspended in chloroform:methanol (2:1, by vol). A 50 µl aliquot
was spotted onto thin layer plates for separation of cardiolipin from other phospholipids
as described 16. Phospholipids were removed from the plate and phospholipid phosphorus
content of cardiolipin was determined as described 17.

95
3.2.5 Enzyme activity assays
NADH dehydrogenase activity was measured from whole cell lysates using the complex I
enzyme activity microplate assay kit (Mitosciences). The activity of Complex II + III
(succinate cytochrome c reductase), measured as reduction of cytochrome c after addition
of succinate, was determined as described previously 18.
3.2.6 Thymocyte and T cell population analysis
Thymocytes, splenocytes and lymph node cells were isolated from control and SLP-2 T-
knockout mice and stained with the following antibodies: PE-conjugated or APC-
conjugated rat anti-CD4 antibody, FITC-conjugated or PerCP-Cy5.5-conjugated rat anti-
CD8 antibody, APC-conjugated Armenian hamster anti-CD3 antibody, PE-conjugated rat
anti-CD25 antibody, FITC-conjugated rat anti-CD44 antibody or with isotype control
antibodies (all from Becton Dickinson, San Jose, CA).
3.2.7 Flow cytometry
Fluorescence was detected by flow cytometry, using a BD FACSCaliburTM flow
cytometer and CellQuest computer software (Becton Dickinson, San Jose, CA) and
analyzed with FlowJo flow cytometry analysis software (Treestar, Inc., Ashland, OR).
3.2.8 Stimulation of T lymphocytes
Splenocytes isolated from control and SLP-2 T-K/O mice were stimulated with
staphylococcal enterotoxin E (SEE) superantigen (Toxin Technologies, Sarasota, FL) or
0.01µg/mL to 1µg/mL of anti-CD3 antibody (Cedarlane, Burlington, ON) and 0.2 µg/mL
anti-CD28 antibody (eBioscience, San Diego, CA) in 96 well round bottom or flat bottom
plates for 24h at 37°C 19. After 24 h stimulation, supernatants were collected for IL-2
and IFNγ measurement by ELISA 20. Cultures were then supplemented with 3H-
thymidine and incubated for 24 h to measure cell proliferation. For glucose experiments,
splenocytes were stimulated with SEE for 24 h in 10% FBS supplemented RPMI media
containing either high (11 mM) or low (4 µM) glucose concentrations.

96
3.2.9 In vivo immunization and ex vivo recall response
Mice were immunized subcutaneously with 100µg ovalbumin in emulsion with Freund’s
complete adjuvant (Difco, Becton Dickinson, San Jose, CA). Eight to ten days later mice
were sacrificed, lymph nodes collected and single cell suspensions prepared for in vitro
stimulation with ovalbumin or pigeon cytochrome c. Culture supernatants were collected
after 48 h for cytokine assays, and cultures supplemented with 3H-thymidine for
proliferation assay 24 h later. Antigen presenting cells were treated with 50µg/mL
mitomycin C for 30 min at 37°C followed by washing, and used at a 1:4 ratio with CD4+
T cells.
3.2.10 Intracellular staining cytokine analysis
Splenocytes were stimulated with 1µg/mL anti-CD3 antibody and 0.2µg/mL anti-CD28
antibody in flat bottom plates for 24 to 36 h at 37°C, in the presence of brefeldin A
(eBioscience, San Diego, CA). Cells were surface stained for CD4-APC and CD8-
PerCP-Cy5.5 followed by intracellular staining for IL-2-PE and IFNγ-FITC using
Invitrogen fix and permeabilization reagents (Life Technologies, Grand Island, NY).
3.2.11 Real time PCR analysis
Splenocytes were stimulated with 1µg/mL anti-CD3 antibody and 0.2µg/mL anti-CD28
antibody in flat bottom plates for 6 h at 37°C. Total RNA was isolated from stimulated
cells or an equal number of un-stimulated cells using the Qiagen RNeasy kit and RNA
was quantified using a NanoDrop (Thermo Scientific, Wilmington, DE). cDNA was
synthesized from 500 ng RNA using the iScript cDNA synthesis kit (BioRad,
Mississauga, Ont) and transcript levels were quantified on the BioRad CFX96 real time
PCR detection system, using iTaq SYBR green supermix with ROX (BioRad). IL-2
primer sequences: Fwd CCTGAGCAGGATGGAGAATTACA, Rev
TCCAGAACATGCCGCAGAG 21, mouse GAPDH real time primers were purchased
from SABiosciences (Qiagen). Relative levels of IL-2 transcript were determined by
comparing the threshold cycle (CT) detection values of IL-2 in WT and SLP-2 T-K/O
cells in stimulated and un-stimulated cells and were normalized to GAPDH levels by the
ΔΔCT method 22.

97
3.2.12 Heart transplants
Abdominal heterotopic transplants were performed as described previously 23. After
transplant, graft viability was assessed daily by abdominal palpation of the transplant
heartbeat. Graft rejection was determined by lack of heartbeat.
3.2.13 Activation-induced cell death assay
Splenocytes were stimulated with 250 ng/mL ionomycin and 1ng/mL PMA (Sigma, St
Louis, MO) for 12, 24, 36, 48 and 60 h in complete RPMI media. At each time point
cells were collected and stained with FITC-conjugated annexin V and propidium iodide
(Clontech Inc, Palo Alto, CA). Fluorescence was detected by flow cytometry.
3.2.14 Statistical analysis
Data analysis was performed with GraphPad Prism software (GraphPad Software, La
Jolla, CA). Data are presented at mean ± SEM. Two tailed Student’s t test or ANOVA
with Bonferroni post-hoc testing were used to determine p values. P values lower than
0.05 were considered significant.
3.3 Results
3.3.1 Generation of T cell-specific SLP-2 knockout mice
To study the effect of SLP-2 in vivo, we first tried to generate conventional SLP-2
knockout mice but no viable embryos were obtained after more than 300 term
pregnancies (C. D. Lemke and J Madrenas, unpublished observations), suggesting that
SLP-2 deficiency is embryonic lethal at the pre-implantation stage. To circumvent this
problem, conditional SLP-2 knockout mice were generated in which the whole SLP-2
gene was replaced by a lox flanked genomic SLP-2 gene. The resulting SLP-2lox mouse
was bred together with a transgenic mouse containing the Cre-recombinase gene under
the control of the CD4 promoter 13 to generate mice in which T cells lacked SLP-2
expression. These resulting knockout mice are called SLP-2 T-K/O throughout the paper.

98
SLP-2 knockout in the T cell lineage was confirmed at the genomic level by PCR
analysis and at the protein level by western blot analysis. Using genomic PCR with
primers flanking the entire SLP-2 sequence to give a 4kb fragment for full length SLP-2
and a 500bp fragment for SLP-2 deletion, we detected the full length SLP-2 fragment in
CD4+ thymocytes, naïve and primed T cells and splenic non-T cells from control mice.
In contrast, we failed to detect the full length SLP-2 in SLP-2 T-K/O CD4+ thymocytes
and naïve and blast T cells. In these mice, only the truncated product, indicative of
genomic deletion of SLP-2, was observed (Fig. 3.1a). In the non-T cell population from
spleen, we still detected the truncated SLP-2 fragment in addition to the full length SLP-2
fragment, likely due to contamination of the prep by T cells (up to 2% of total cell
numbers in the prep).
The deletion of genomic SLP-2 translated into significantly lower SLP-2 protein
expression in SLP-2 T-K/O CD4+ thymocytes, and complete loss of SLP-2 expression in
peripheral T cells but not in spleen CD4-CD8- cells (Fig. 3.1b), confirming the restriction
of SLP-2 deletion to conventional T cells. Moreover, and as reported with primary
human T cells 1, these experiments showed the upregulation of SLP-2 protein levels upon
activation of wild type T cells (see blast vs. naïve, in Fig. 3.1b).
3.3.2 SLP-2 deficiency is associated with abnormal cardiolipin compartmentalization in mitochondrial membranes and reduced mitochondrial respiration
Proteins of the SPFH superfamily have been linked to the organization of membrane
microdomains 24-29. Based on this assumption, and since SLP-2 is mostly a mitochondrial
protein localized in the intermembrane space 30, 31, we examined if SLP-2 deficiency
would alter mitochondrial membrane compartmentalization. We first looked at the effect
of SLP-2 deficiency on PHBs since these proteins interact with each other and contribute
to the formation of detergent-insoluble microdomains in human mitochondria 14, 32.
Triton-X insoluble domains from T cell mitochondria from control and SLP-2 T-K/O
mice were prepared and blotted for PHB1 and SLP-2. Although total cellular levels of

99
Figure 3.1: Generation of T cell-specific SLP-2 knockout mice.
(a) Genomic DNA was extracted from control and SLP-2 T-K/O CD4+ thymocytes, naïve
and blast T cells and non T cells and genomic SLP-2 deletion was analyzed by PCR using
primers flanking the entire SLP-2 gene. (b) Whole cell lysates from CD4+ thymocytes,
naïve and blast T cells and non T cells were sequentially immunoblotted for SLP-2 and
for GAPDH (as a loading control). CD4+ thymocytes were isolated from single cell
suspension of control (WT) and SLP-2 T-K/O thymocytes; splenocyte single cell
suspensions were separated into T cell and non T cell fractions. Untreated T cells are
referred to as naïve T cells while T cells stimulated for 72 hours with ionomycin and
PMA are referred to as blast T cells. Data are representative of 3 independent
experiments using multiple mice with material from a representative mouse per group ran
per lane.
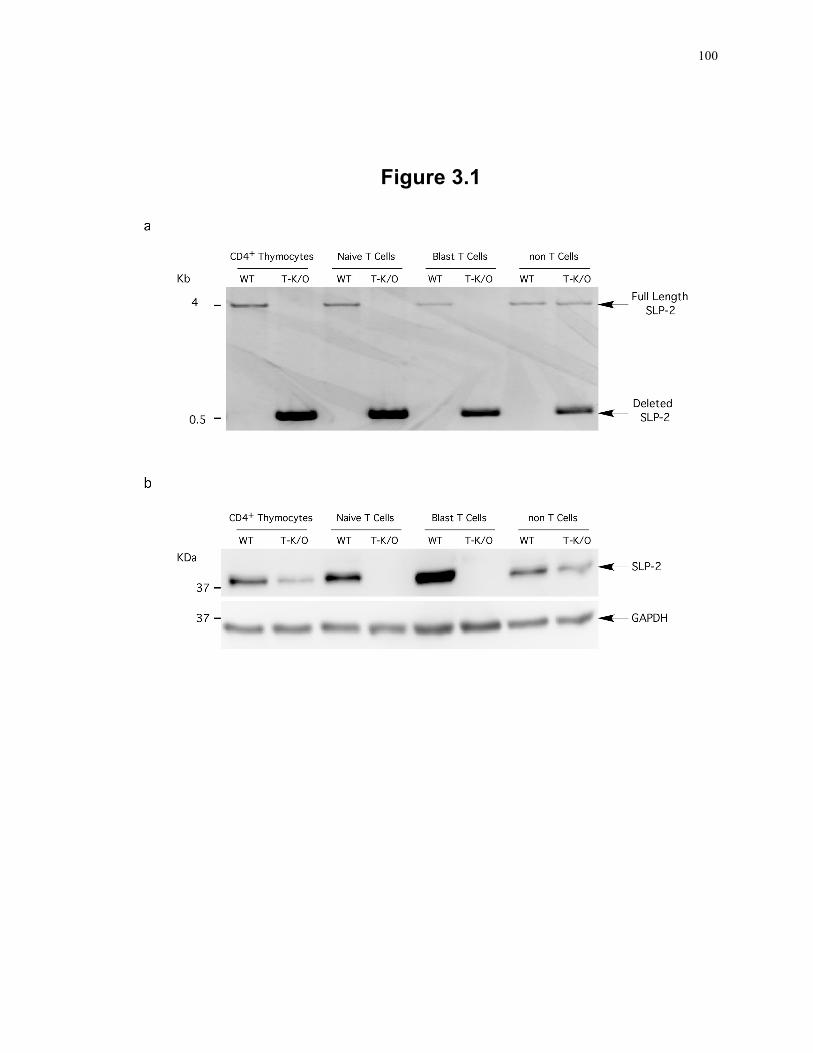
100
Figure 3.1

101
PHB1 were not affected in SLP-2-deficient T cells (see below), the level of PHB1 in the
mitochondrial detergent-insoluble microdomain fraction was reduced, although the levels
in the detergent-soluble fraction were similar between control and SLP-2 T-K/O cells
(Fig. 3.2a and e). As the amount of PHB1 in detergent-soluble fractions of control and
SLP-2 T-K/O cells was similar, the loss of PHB1 from the insoluble fraction suggested
an abnormal compartmentalization in the SLP-2 T-K/O mitochondrial membrane.
We have recently shown that SLP-2 not only interacts with PHBs but also binds
cardiolipin, the signature phospholipid linked to detergent-insoluble microdomains of
mitochondrial membranes 12. Therefore, the defect in mitochondrial membrane
compartmentalization of PHB1 associated with SLP-2 deficiency prompted us to test if
the content or distribution of cardiolipin was also impaired. Total cardiolipin content in
T cells from control and SLP-2 T-K/O mice as detected with nonyl-acridine orange
staining was not significantly different (Fig. 3.2b). However, the cardiolipin content in
mitochondrial detergent-insoluble microdomains isolated from primed SLP-2 T-K/O T
cells was significantly (p<0.05) lower than that of control samples (Fig. 3.2c). Altogether,
these results imply that SLP-2 deficiency in T cells is associated with impaired
mitochondrial membrane compartmentalization of cardiolipin.
Proper compartmentalization of mitochondrial membranes into cardiolipin-enriched
microdomains is required for optimal assembly and function of the components of the
respiratory chain 33, 34. Therefore, we examined the effects of SLP-2 deficiency in
assembly and function of the complexes of the respiratory chain. First, we analyzed the
levels of mitochondrial respiratory chain components in SLP-2-deleted T cells using a
cocktail of antibodies against different subunits of complexes I to V of the respiratory
chain. SLP-2 T-K/O cells had lower levels of the NDUFB8 subunit of complex I and
slight decrease of the 30 kDa subunit of complex II compared to control cells (Fig. 3.2d).
We corroborated this finding with two other subunits of the 45 described subunits of
complex I, including the NDUFS3 and NDUFA9 subunits (Fig. 3.2e). Note that the total
cellular levels of PHB1 were not affected by SLP-2 deficiency. SLP-2 deficiency

102
Figure 3.2: T cells deficient in SLP-2 have abnormal cardiolipin
compartmentalization and decreased activity of respiratory complexes I and II+III.
(a) Mitochondrial detergent-soluble and insoluble fractions from WT and T-K/O cells
were separated by SDS-PAGE and analyzed by serial blotting for SLP-2 and PHB1. Blot
is representative of 3 independent experiments. (b) Control and SLP-2 T-K/O T cells
were stained with 10 nM nonyl-acridine orange to assess total cellular cardiolipin content
by flow cytometry. The plot represents one control (heavy line) and one knockout mouse
(fine line), representing 3 independent experiments with 3 mice per group. (c)
Cardiolipin was measured in mitochondrial detergent-insoluble fractions collected from
control (black bars) and SLP-2 T-K/O (white bars) T cells. (d) Whole cell lysates from
control (WT) and SLP-2 T-K/O T cells were sequentially immunoblotted for components
of each complex of the respiratory chain and SLP-2, or (e) blotted for individual subunits
of complex I, PHB1 and actin (as a loading control). Blots are representative of 3
independent experiments. (f) Complex I activity was measured in control (black bars)
and SLP-2 T-K/O T cells (white bars). (g) Complex II+III activity was measured in
control (black bars) and SLP-2 T-K/O T cells (white bars). *: p<0.05, **: p<0.01.
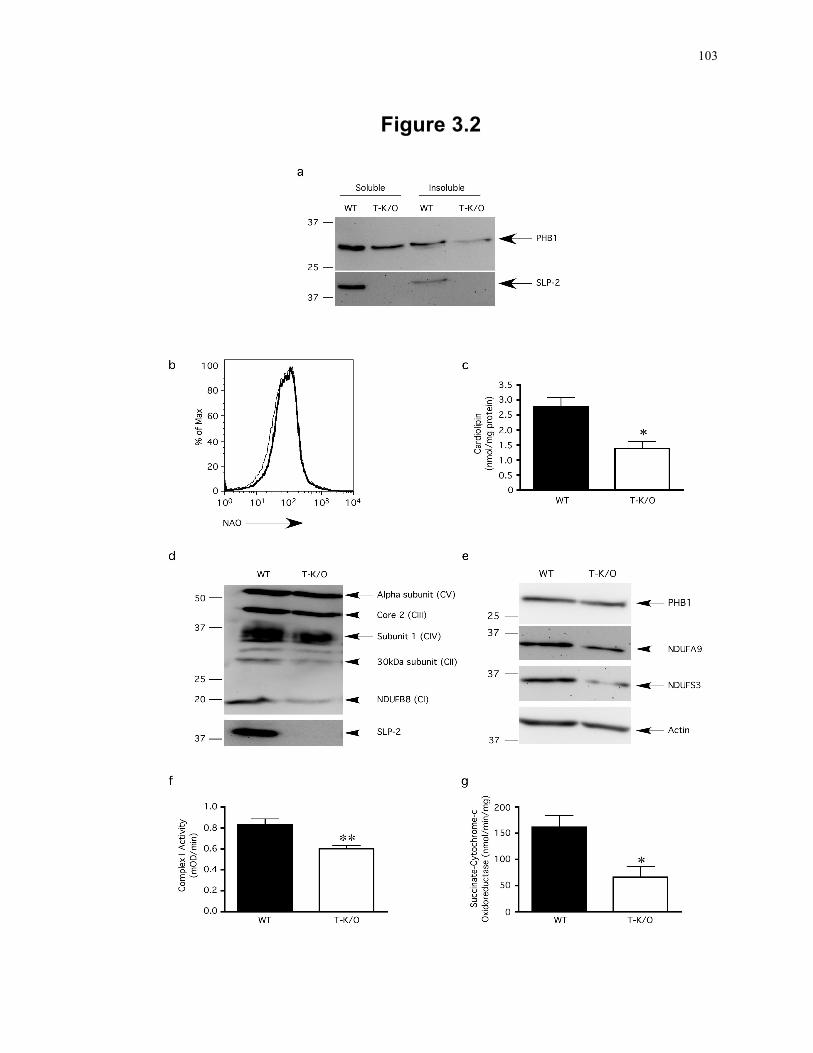
103
Figure 3.2

104
did not grossly affect the levels of complexes III to V, including ATP synthase, which
were similar in control and SLP-2 T K/O T cells.
The observed decrease in levels of complex I subunits translated into a significant
(p<0.01) decrease in the functional activity of complex I NADH dehydrogenase in SLP-2
T-K/O T cells (Fig. 3.2f). In addition, an assay of the combined activity of succinate
dehydrogenase and cytochrome c oxidoreductase (complex II + III activity) also showed
a significant decrease in SLP-2-deficient T cells (p<0.05 Fig. 3.2g).
3.3.3 Impaired ex vivo responses of SLP-2-deficient T cells
Next we examined the effect of SLP-2 deletion and the altered mitochondrial membrane
compartmentalization of cardiolipin on T cell function. First, we looked at the cellularity
of thymus, spleen and lymph nodes. Organ sizes were similar between control and SLP-2
T-K/O mice. The total thymocyte and splenocyte cell counts (Fig. 3.3a) as well as the
number of double negative, double positive, CD4+ single positive and CD8+ single
positive thymocytes and splenic cells (Fig. 3.3b and c) were similar in unmanipulated
SLP-2 T-K/O and control mice, implying a grossly normal T cell development in these
mice. Although the total cell count in lymph nodes was similar in control and SLP-2 T-
K/O mice (Fig. 3.3a), there was a slight but consistent decrease in CD4+ T cells in the
lymph nodes of SLP-2 T-K/O mice (p = 0.09, Fig. 3.3d).
Next, we tested the functional responses of SLP-2-deficient T cells. Naïve splenocytes
from control and SLP-2 T-K/O mice were stimulated ex vivo with increasing
concentrations of staphylococcal enterotoxin (SEE) superantigen. We found that SLP-2
T-K/O mice produced significantly less IL-2 upon superantigen stimulation compared to
control mice (p<0.01) (Fig. 3.4a). The decrease in IL-2 production, however, did not
translate into significantly less T cell proliferation (Fig. 3.4b), and SLP-2 T-K/O
splenocytes produced similar levels of IFNγ and IL-17 in response to superantigen
stimulation (data not shown).

105
Figure 3.3: Grossly normal thymic T cell development and peripheral T cell pool in
SLP-2 T-K/O mice.
(a) Eight-week-old wild type (black bars) and SLP-2 T-K/O (white bars) mice were
sacrificed and single cell suspensions were made from whole thymus, spleens and lymph
nodes and live cells were counted. To analyze CD4+ and CD8+ T cell populations, single
cell suspension of (b) thymocytes, (c) splenocytes and (d) lymph nodes were stained with
anti-CD4-PE and anti-CD8-FITC, or isotype-matched control antibodies and surface
expression detected by FACS. T cell populations were calculated by multiplying the total
thymocyte count or total splenocyte count by the population percentage obtained by
FACS. Splenocytes and lymph nodes were also stained with anti-CD3-APC antibodies
and total CD3+ T cell populations were calculated as described above. These plots
represent an average of 6 individual mice for each genotype, performed in 4 separate
experiments. DN: double negative thymocytes; DP: double positive thymocytes; SP:
single positive.
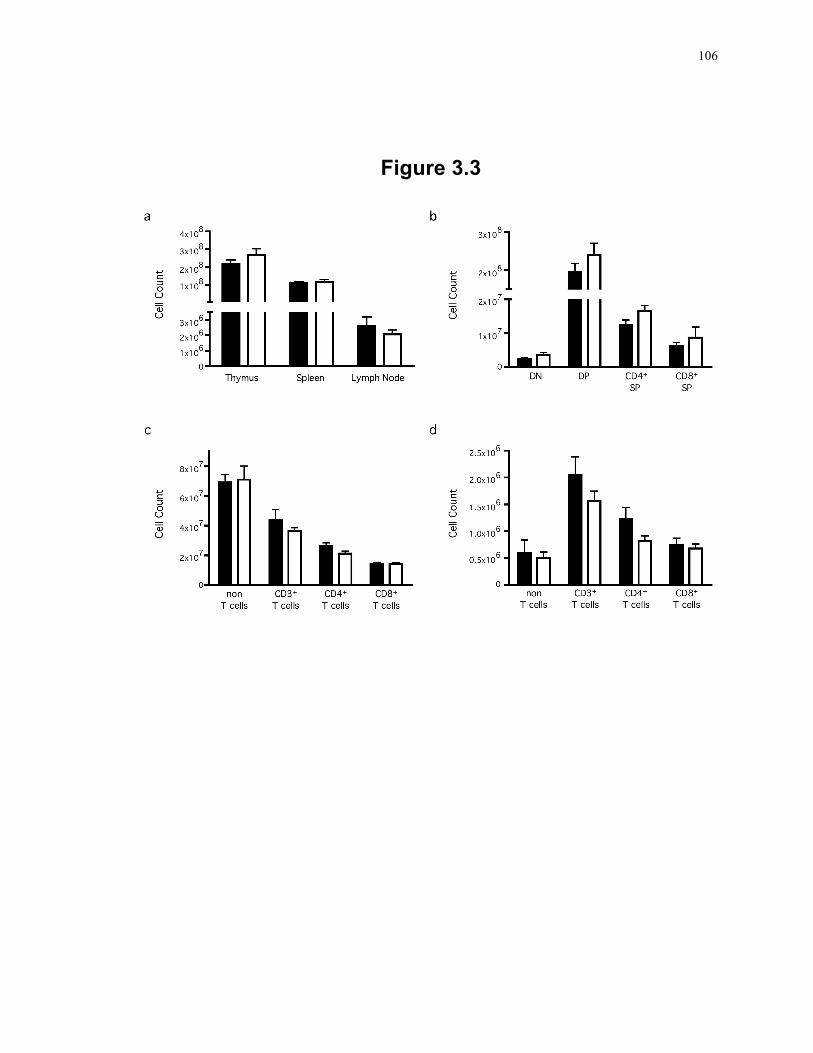
106
Figure 3.3

107
Figure 3.4: T cells deficient in SLP-2 produce less IL-2 in response to T cell activation.
Naive or ex vivo primed splenocytes from WT (black bars) or SLP-2 T-K/O (white bars)
mice were analyzed for IL-2 production and proliferation in response to superantigen
stimulation. Naïve splenocytes were incubated with increasing concentrations of SEE
superantigen at 37°C. After 24 hours, supernatants were collected and IL-2 secretion was
measured by ELISA (a). Cultures were supplemented with additional media and 3H-
thymidine and incubated for an additional 24 hours after which time the cells were
harvested and 3H-thymidine incorporation was measured (b). In C and D, splenocytes
from WT and SLP-2 T-K/O mice were stimulated with 250 ng/mL ionomycin and 1
ng/mL PMA for 72 hours and rested for 48 hours. The primed cells were then analyzed
for IL-2 secretion (c) and proliferation (d) in response to superantigen stimulation in the
same manner as described for naïve cells. Plots in this figure represent an average of 4
mice per group and are representative of at least 3 independent experiments, *: p<0.05,
**: p<0.01, ***: p<0.001.
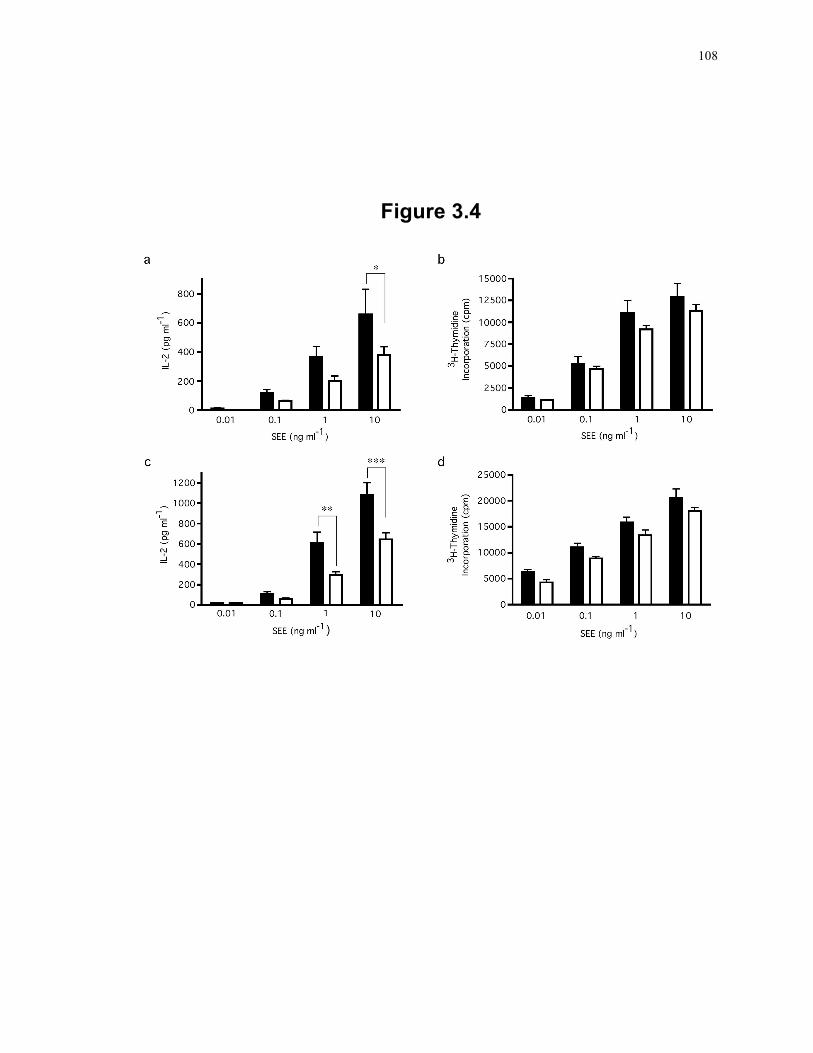
108
Figure 3.4

109
We have previously reported that SLP-2 levels increase upon priming 1 implying that the
role of this protein is greater upon T cell priming. To test the secondary response of SLP-
2-deficient T cells, control and SLP-2 T-K/O splenocytes were stimulated in vitro with
PMA and ionomycin for 72 hours followed by 48 hours resting, before being stimulated
with SEE superantigen. Again, primed SLP-2 T-K/O cells produced significantly less IL-
2 compared to control cells (p<0.001, Fig. 3.4c). We observed a trend towards less
proliferation in SLP-2 T-K/O cells but it did not reach statistical significance (Fig. 3.4d).
The significant decrease in IL-2 responses seen in SLP-2 T-K/O T cells was also
observed when naïve and in vitro primed cells were stimulated with anti-CD3 and anti-
CD28 antibodies (data not shown). Together, these results demonstrate that SLP-2-
deficient T cells have decreased activation leading to decreased IL-2 responses.
3.3.4 SLP-2 deficiency impairs T cell responses in vivo
To assess how the impairment in T cell responses observed ex vivo translated into T cell
responses in vivo, we first assessed the capacity of SLP-2-deficient T cells to respond to
immunization. Mice were immunized with ovalbumin (OVA) and ex vivo recall
responses tested 10 days after immunization. The response to pigeon cytochrome c
(PCC) was used as a control. In these experiments, SLP-2-deficient cells had
significantly decreased responses (p<0.05), measured by both IL-2 production and
proliferation (Fig. 3.5a and b).
As an additional model for in vivo T cell activation, we examined the response to
vascularized allografts in SLP-2 T-K/O mice compared to that in SLP-2 competent mice.
Heterotopic heart transplants from C3H mice (H-2k) into C57BL/6 (H-2b) wild type and
SLP-2 T-K/O mice were performed and graft survival monitored. In these experiments,
we observed a slight but consistent and significant (p<0.05) delay in the rejection of
allografts in SLP-2 T-K/O mice (Fig. 3.5c). Together, these results corroborated that

110
Figure 3.5: T cells deficient in SLP-2 have decreased responses in vivo.
Mice were immunized subcutaneously with 100 µg ovalbumin in emulsion with Freund’s
complete adjuvant. Ten days post-immunization lymph nodes were isolated and single
cell suspension from WT (black bars) or SLP-2 T-K/O (white bars) mice were stimulated
in vitro with various concentrations of ovalbumin as well as the control peptide pigeon
cytochrome c (PCC). After 48 hours, supernatants were collected and IL-2 levels were
measured by ELISA (a). Cultures were supplemented with additional media and 3H-
thymidine and incubated for an additional 24 hours, after which time the cells were
harvested and the 3H-thymidine incorporation was measured (b). These plots are
representative of 4 mice for each group and are representative of 3 independent
experiments. (c) Wild type and SLP-2 knockout mice were grafted with hearts isolated
from C3H mice. Rejection of the transplant tissue was measured by the beating of the
transplant heart, with the endpoint of rejection being the loss of heartbeat. These plots
report graft survival for 6 mice per group, *: p<0.05, **: p<0.01, ***: p<0.001.
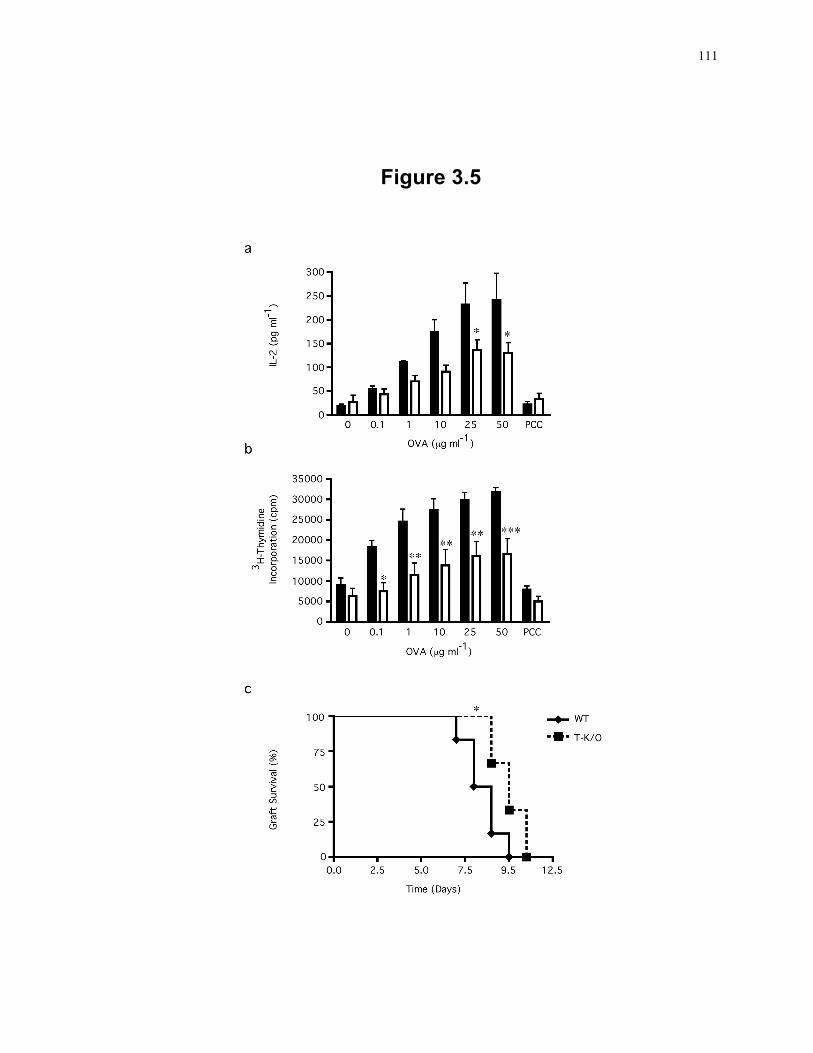
111
Figure 3.5

112
SLP-2 deficiency in T cells is associated with impaired responses to TCR-dependent
stimulation in vivo.
The lymph nodes from control and SLP-2 T-K/O mice had comparable numbers of
CD19+ B cells and CD3+ T cells both before and after immunization (data not shown).
Although there was no difference in the total number of T cells, SLP-2 T-K/O mice had a
consistent and significant decrease in the percentage of CD4+ T cells but not in the
percentage of CD8+ T cells after immunization (Table 3.1). The decrease in the
percentage of CD4+ T cells was primarily due to decreased proportion of activated T
cells, as demonstrated by the decrease in CD4+CD25+ and CD4+CD44hi T cell
populations. The percentage and function of regulatory CD4+CD25+FoxP3+ T cells were
comparable between control and SLP-2 T-K/O mice (Table 1 and data not shown).
Furthermore, the decrease in activated CD4+ T cells was not due to an increased
susceptibility to activation-induced cell death (data not shown).
3.3.5 The IL-2 defect of SLP-2 deficient T cells occurs at the post-transcriptional level
To further investigate the low IL-2 responses in SLP-2 T-K/O mice, we examined IL-2
production at the single cell level by performing intracellular staining and analyzing cell
populations by flow cytometry. Naïve splenocytes were isolated from WT and SLP-2 T-
K/O mice and stimulated for 24 hours in the presence of anti-CD3 and anti-CD28
antibodies. In agreement with the IL-2 data presented above, significantly fewer CD4+
and CD8+ SLP-2 T-K/O cells produced IL-2 in response to stimulation, compared to WT
T cells (p<0.001 and p<0.01 respectively, Fig. 3.6a and b). In addition, the geometric
mean of the IL-2 fluorescence staining was significantly lower in the SLP-2 T-K/O
CD4+IL-2+ and CD8+IL-2+ cell population compared to WT populations (p<0.001 and
p<0.01 respectively, Fig. 3.6c and d). Together, these data indicate that fewer SLP-2-
deficient T cells became activated to express IL-2, and those T cells that were activated
produced less IL-2 compared to WT T cells. Surprisingly, when we investigated the
transcript levels of IL-2 in stimulated T cells, we found no significant difference in IL-2
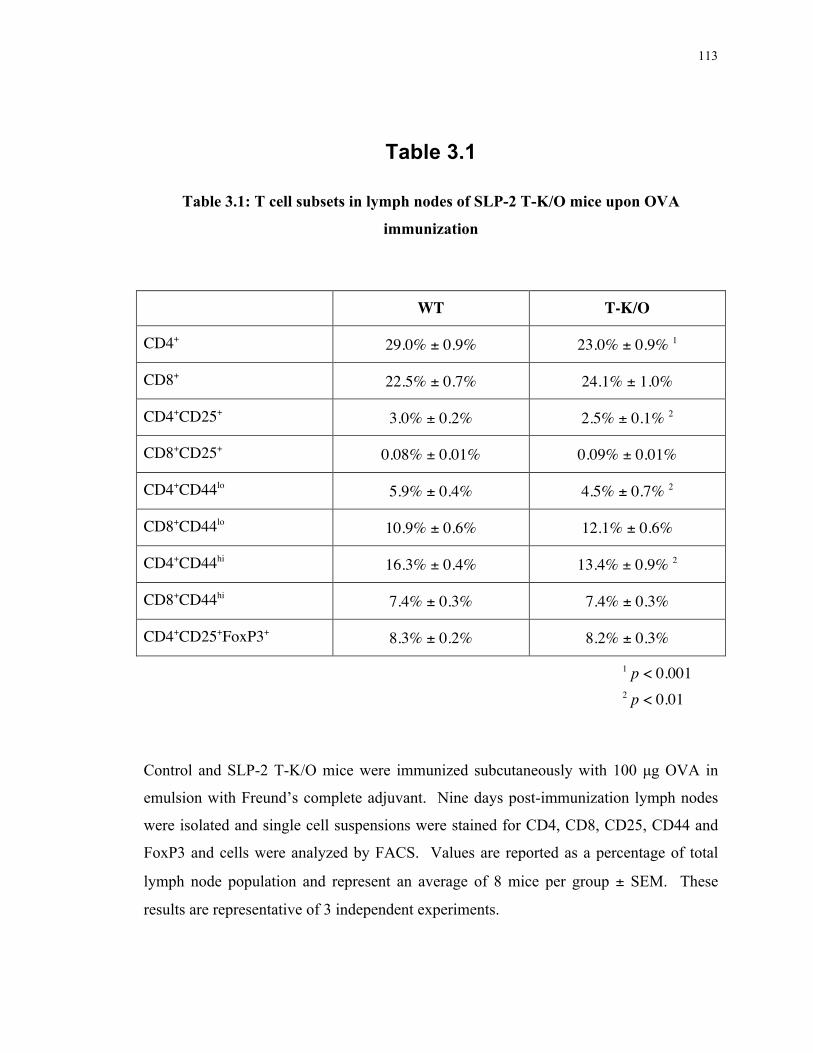
113
Table 3.1
Table 3.1: T cell subsets in lymph nodes of SLP-2 T-K/O mice upon OVA
immunization
WT T-K/O
CD4+ 29.0% ± 0.9% 23.0% ± 0.9% 1
CD8+ 22.5% ± 0.7% 24.1% ± 1.0%
CD4+CD25+ 3.0% ± 0.2% 2.5% ± 0.1% 2
CD8+CD25+ 0.08% ± 0.01% 0.09% ± 0.01%
CD4+CD44lo 5.9% ± 0.4% 4.5% ± 0.7% 2
CD8+CD44lo 10.9% ± 0.6% 12.1% ± 0.6%
CD4+CD44hi 16.3% ± 0.4% 13.4% ± 0.9% 2
CD8+CD44hi 7.4% ± 0.3% 7.4% ± 0.3%
CD4+CD25+FoxP3+ 8.3% ± 0.2% 8.2% ± 0.3%
1 p < 0.001 2 p < 0.01
Control and SLP-2 T-K/O mice were immunized subcutaneously with 100 µg OVA in
emulsion with Freund’s complete adjuvant. Nine days post-immunization lymph nodes
were isolated and single cell suspensions were stained for CD4, CD8, CD25, CD44 and
FoxP3 and cells were analyzed by FACS. Values are reported as a percentage of total
lymph node population and represent an average of 8 mice per group ± SEM. These
results are representative of 3 independent experiments.

114
Figure 3.6: SLP-2-deficiency in T cells induces a post-transcriptional defect in IL-2 production.
(a-d) Naive splenocytes from WT (black bars) or SLP-2 T-K/O (white bars) mice were
analyzed for intracellular IL-2 staining in response to anti-CD3 and anti-CD28
stimulation using flow cytometry for CD4, CD8 and IL-2. Plots show the percentage of
CD4 or CD8 cells expressing IL-2 (a and b respectively) or the geometric mean of the
IL-2-PE stain in CD4+IL-2+ or CD8+IL-2+ populations (c and d respectively). (e) Real-
time PCR analysis of IL-2 mRNA in WT (black bars) and SLP-2 T-K/O (white bars)
cells from same samples as in a to d. (f-i) Naive splenocytes from WT (black bars) or
SLP-2 T-K/O (white bars) mice were analyzed for intracellular IFNγ staining in response
to anti-CD3 and anti-CD28 stimulation using flow cytometry for CD4, CD8 and IFNγ.
Plots show the percentage of CD4+ or CD8+ cells expressing IFNγ (f and g respectively)
or the geometric mean of the IFNγ-FITC stain in CD4+IFNγ+ or CD8+IFNγ+ populations
(h and i respectively). All plots represent an average of 4 mice and are representative of 3
independent experiments. *:p<0.05, **: p<0.01, ***: p<0.001.
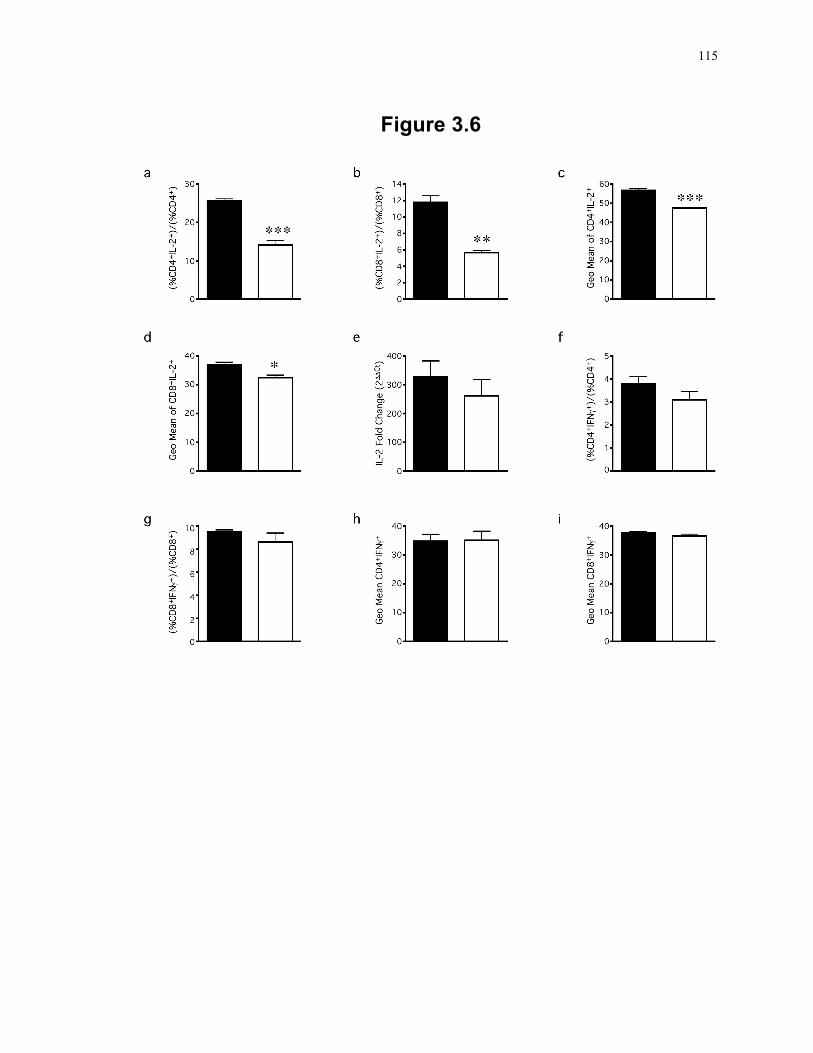
115
Figure 3.6

116
transcript levels between SLP-2 T-K/O and WT cells (Fig. 3.6e), which may point to a
possible role for SLP-2 in the processing of IL-2 mRNA.
Interestingly, the defect in cytokine production seen in SLP-2-deficient T cells seemed to
be limited to IL-2. In addition to normal IFNγ and IL-17 secretion from stimulated SLP-
2 T-K/O cells, we also found that a similar proportion of WT and SLP-2 T-K/O cells
become IFNγ expressing cells in response to T cell stimulation (Fig. 3.6f and g).
Furthermore, quantification of fluorescent signal from IFNγ+ populations indicate a
similar expression level in both WT and SLP-2 T-K/O cells (Fig. 3.6h and i). These
findings support the idea of a unique role for SLP-2 in the regulation of IL-2 expression.
3.3.6 Altered metabolic capacity of SLP-2-deficient T cells
The metabolic requirements of conventional T cell activation are mostly dependent on
glycolysis if glucose is not limiting 35, whereas the contribution of mitochondrial
oxidative phosphorylation is relatively secondary. Given the involvement of SLP-2 in
mitochondrial function, we next tested the effect of limiting glucose supply on the
impairment of T cell activation due to SLP-2 deficiency. Compared to WT T cells, SLP-
2 T-K/O cells showed significantly less IL-2 production in response to stimulation with
SEE superantigen under both high glucose (11 mM) and low glucose (4 µM) conditions
(Fig. 3.7). Under non-limiting glucose, significantly less IL-2 was produced by SLP-2 T-
K/O cells in response to 10 ng/mL SEE stimulation (p<0.001) compared to WT cells,
whereas limiting glucose (to low levels in media and fetal bovine serum) led to a broader
reduction of IL-2 production, i.e. seen in response to 1ng/mL SEE (p<0.05) and to 10
ng/mL SEE (p<0.001). However, limiting glucose supply did not decrease further the
already lower IL-2 response of SLP-2 T-K/O cells, implying an altered metabolic
capacity of SLP-2-deficient T cells that is independent of and insensitive to glycolysis.

117
Figure 3.7: Altered metabolic capacity of SLP-2-deficient T cells during activation.
Splenocytes from WT (black bars) and SLP-2 T-K/O (white bars) mice were analyzed for
IL-2 production in response to indicated concentrations of SEE superantigen stimulation
under conditions of high glucose (11 mM) or low glucose (4 µM), both in the presence of
10% fetal bovine serum in the culture medium. Results represent mean and standard
deviation of triplicate samples. **: p<0.01, ***: p<0.001.

118
Figure 3.7

119
3.4 Discussion
We report here the characterization of SLP-2 deficiency in T cells in vivo. To do so, and
given that systemic SLP-2-deficiency is embryonically lethal, we had to generate a
conditional knockout mouse strain utilizing the Cre-Lox system with Cre recombinase
under the control of the CD4 promoter. This led to the deletion of the SLP-2 gene during
the double positive stage of thymocyte development. The SLP-2 T-K/O mice were viable
and bred well, giving sufficient numbers for biochemical and functional studies of the
effects of SLP-2-deficiency in T lymphocytes.
We found that SLP-2 deficiency in T cells is associated with abnormal cardiolipin
compartmentalization in mitochondrial membranes. SLP-2 is tightly associated with the
mitochondrial inner membrane 30, 31, and other members of the SPFH family have been
linked to the organization of various membrane microdomains 24-29. Therefore, it is
plausible to suggest that SLP-2 promotes the compartmentalization of the mitochondrial
inner membrane into cardiolipin-enriched microdomains. This proposed role is
consistent with the effects of SLP-2 deficiency on mitochondrial function. Formation of
cardiolipin microdomains is required for optimal assembly of the electron transport
complexes in the mitochondrial inner membrane, likely into supercomplexes 36, to allow
for rapid and efficient electron transport through the respiratory chain. Thus, in the
absence of SLP-2, mitochondrial inner membrane compartmentalization is altered and
this interferes with the optimal assembly of the respiratory chain, ultimately resulting in
defective respiration.
How does SLP-2 contribute to the formation of cardiolipin-enriched microdomains? We
have recently shown that SLP-2 binds cardiolipin and interacts with the mitochondrial
resident proteins PHB 1 and 2 12, 31. Thus, SLP-2 may couple PHBs with cardiolipin-
enriched microdomains. This model supports the proposed involvement of PHBs in the
compartmentalization of the mitochondrial inner membrane 37. Also, by facilitating
mitochondrial membrane organization, SLP-2 enhances mitochondrial function
increasing respiration and resistance to apoptosis as we have recently shown under

120
conditions of SLP-2 upregulation in vitro 12. In addition, this model helps to explain the
observations that cardiolipin is required for proper function of the respiratory chain
complexes 38 and that cardiolipin-enriched microdomains play a role in the induction of
apoptosis 14, 32, 39.
Deficiency in SLP-2 is associated with a decrease in mitochondrial respiration as
measured by the activities of complexes I and II+III. The physiological relevance of a
reduction of complex I activity is likely greater than what is measured since the assay
commonly used to determine the activity of complex I only detects partial reaction of
complex I 40-42. Defective complex I activity is considered the most common defect of
the respiratory chain in humans 43 and causes a diverse set of diseases including Leigh
syndrome and renal tubular acidosis. Complex I represents the major entry point into the
respiratory chain through the oxidation of NADH to NAD+ with the transfer of two
electrons into the respiratory chain 44. We show that, in the absence of SLP-2, there is
inefficient assembly of complex I subunits and confirm the suggestion of a previous in
vitro study that downregulation of SLP-2 levels may cause a reduction in the level of
components of the respiratory chain 31. Cardiolipin-enriched microdomains may be
especially required for optimal assembly of complex I because of the large, multichain
nature of this complex, which includes at least 45 subunits 44. Such an inefficient
assembly of this multichain complex may result in increased degradation of the subunits
by mitochondrial proteases, explaining the decrease in complex I activity and protein
levels in SLP-2 T-K/O T cells 31, 45. The defect in mitochondrial membrane
compartmentalization of cardiolipin in the absence of SLP-2 not only affects the function
of complex I but also of complex II+III and likely of other protein supercomplexes in the
mitochondrial membrane as suggested by the data presented here and by preliminary
proteome characterization of detergent-insoluble microdomains (S. D. Dunn and J.
Madrenas, unpublished observations).
In T cells, the impairment in mitochondrial membrane compartmentalization and
complexes I and II+III activities associated with SLP-2 deficiency results in a minimal
defect under resting conditions. Such a paucity of immune manifestations is also
observed in humans with complex I deficiency. Although these patients have a reduction

121
of complex I activity in peripheral blood leukocytes, they do not show overt immune
deficiency 46, 47. This may be due to a compensatory reliance of T cells on glycolysis for
ATP production inasmuch as access to glucose is not limited 35.
However, the phenotype of SLP-2 deficiency in T cells becomes apparent upon TCR-
mediated activation and results in lower IL-2 production and decreased T cell responses.
It is likely that this reflects the increased metabolic demands on T cells during activation.
The impairment in electron uptake by complex I and decreased complex II+III activity
will result in less protons shuttled across the inner membrane, leading to lower ATP
production and decreased mitochondrial function. The activation-dependent phenotype is
consistent with the recent observation that complex I plays an important role in the
activation of T cells 48, 49. Also, the decrease in CD4+ T cell expansion is expected
because low NAD+ levels, seen under conditions of decreased complex I activity, are
associated with mild lymphopenia 46, 50-52.
A surprising finding in SLP-2-deficient T cells is the selective post-transcriptional defect
in IL-2 production in response to TCR stimulation, documented by the decrease in both
intracellular and secreted levels of IL-2 protein with abundant IL-2 transcripts. A similar
phenotype has been reported in anergic T cells in which IL-2 transcript levels were
similar in T cells stimulated under both activating and anergic conditions, but IL-2
protein was only detected in activated T cells 53, 54. This post-transcriptional block in
cytokine production affected other cytokines, such as IFNγ, IL-4 and IL-17 and was
dependent on AU-rich regions in the 3’ UTR of cytokine mRNA 54, 55. In contrast to
anergic conditions, the post-transcriptional block in SLP-2-deficient T cells appears to be
limited to IL-2 expression as IFNγ and IL-17 were expressed normally. The precise
nature of the SLP-2-dependent metabolic checkpoint for the processing of IL-2 mRNA is
currently unknown.
Why the metabolic demands upon activation would be higher in CD4+ T cells than in
CD8+ T cells as implied by the selective effect of SLP-2 deficiency on CD4+ T cell
expansion remains to be determined. Recent work has started to unravel the role of
metabolic pathways in the differentiation of various T cell subsets 49, 56, 57. Memory CD8

122
T cells seem to rely on fatty acid oxidation, wherein mice with a block in fatty acid
metabolism showed significantly decreased memory CD8 T cells, and upregulation of
fatty acid oxidation with metformin, an AMPK activator, increased the generation of
CD8+ T cells 57. Furthermore, rapamycin treatment increased CD8 memory generation,
likely through the upregulation of fatty acid oxidation 56, 57. Treg cells also require high
levels of fatty acid oxidation 49. Indeed, precursor cells can be driven into the Treg
lineage by the addition of excess fatty acid, metformin or rapamycin. In contrast, effector
CD4+ T cells (TH1, TH2 and TH17 cells) require high levels of glycolysis 35. Thus, the
difference in the numbers CD4+ and CD8+ T cells in SLP-2 T-K/O mice may be a
reflection of differences in the metabolic requirements of these cells such as CD4+ T cells
are more sensitive to mitochondrial defects than CD8+ T cells.
In summary, we show that the loss of SLP-2 in T cells alters cardiolipin
compartmentalization in mitochondrial membranes and prevents proper function of
complexes I and II+III of the respiratory chain. Such a decreased respiratory activity
provides a straightforward explanation for the impaired activation and defective
downstream T cell responses in SLP-2-deficient mice. Availability of the SLP-2 T-K/O
mouse strain may thus be helpful to dissect different aspects of metabolic regulation of T
cell responses.

123
3.5 References
1. Kirchhof, M.G., Chau, L.A., Lemke, C.D., Vardhana, S., Darlington, P.J., Marquez, M.E., Taylor, R., Rizkalla, K., Blanca, I., Dustin, M.L. & Madrenas, J. Modulation of T cell activation by stomatin-like protein 2. J Immunol 181, 1927-1936 (2008).
2. Stewart, G.W., Hepworth-Jones, B.E., Keen, J.N., Dash, B.C., Argent, A.C. & Casimir, C.M. Isolation of cDNA coding for an ubiquitous membrane protein deficient in high Na+, low K+ stomatocytic erythrocytes. Blood 79, 1593-1601 (1992).
3. Seidel, G. & Prohaska, R. Molecular cloning of hSLP-1, a novel human brain-specific member of the band 7/MEC-2 family similar to Caenorhabditis elegans UNC-24. Gene 225, 23-29 (1998).
4. Wang, Y. & Morrow, J.S. Identification and characterization of human SLP-2, a novel homologue of stomatin (band 7.2b) present in erythrocytes and other tissues. J Biol Chem 275, 8062-8071 (2000).
5. Green, J.B., Fricke, B., Chetty, M.C., von During, M., Preston, G.F. & Stewart, G.W. Eukaryotic and prokaryotic stomatins: the proteolytic link. Blood Cells Mol Dis 32, 411-422 (2004).
6. Green, J.B. & Young, J.P. Slipins: ancient origin, duplication and diversification of the stomatin protein family. BMC Evol Biol 8, 44 (2008).
7. Goldstein, B.J., Kulaga, H.M. & Reed, R.R. Cloning and characterization of SLP3: a novel member of the stomatin family expressed by olfactory receptor neurons. J Assoc Res Otolaryngol 4, 74-82 (2003).
8. Browman, D.T., Hoegg, M.B. & Robbins, S.M. The SPFH domain-containing proteins: more than lipid raft markers. Trends Cell Biol 17, 394-402 (2007).
9. Tavernarakis, N., Driscoll, M. & Kyrpides, N.C. The SPFH domain: implicated in regulating targeted protein turnover in stomatins and other membrane-associated proteins. Trends Biochem Sci 24, 425-427 (1999).
10. Huang, M., Gu, G., Ferguson, E.L. & Chalfie, M. A stomatin-like protein necessary for mechanosensation in C. elegans. Nature 378, 292-295 (1995).
11. Wetzel, C., Hu, J., Riethmacher, D., Benckendorff, A., Harder, L., Eilers, A., Moshourab, R., Kozlenkov, A., Labuz, D., Caspani, O., Erdmann, B., Machelska, H., Heppenstall, P.A. & Lewin, G.R. A stomatin-domain protein essential for touch sensation in the mouse. Nature 445, 206-209 (2007).

124
12. Christie, D.A., Lemke, C.D., Elias, I.M., Chau, L.A., Kirchhof, M.G., Li, B., Ball, E.H., Dunn, S.D., Hatch, G.M. & Madrenas, J. Stomatin-like protein 2 binds cardiolipin and regulates mitochondrial biogenesis and function. Mol Cell Biol 31, 3845-3856 (2011).
13. Lee, P.P., Fitzpatrick, D.R., Beard, C., Jessup, H.K., Lehar, S., Makar, K.W., Perez-Melgosa, M., Sweetser, M.T., Schlissel, M.S., Nguyen, S., Cherry, S.R., Tsai, J.H., Tucker, S.M., Weaver, W.M., Kelso, A., Jaenisch, R. & Wilson, C.B. A critical role for Dnmt1 and DNA methylation in T cell development, function, and survival. Immunity 15, 763-774 (2001).
14. Ciarlo, L., Manganelli, V., Garofalo, T., Matarrese, P., Tinari, A., Misasi, R., Malorni, W. & Sorice, M. Association of fission proteins with mitochondrial raft-like domains. Cell Death Differ 17, 1047-1058 (2010).
15. Lowry, O.H., Rosebrough, N.J., Farr, A.L. & Randall, R.J. Protein measurement with the Folin phenol reagent. J Biol Chem 193, 265-275 (1951).
16. Hatch, G.M. Regulation of cardiolipin biosynthesis in the heart. Mol Cell Biochem 159, 139-148 (1996).
17. Rouser, G., Fkeischer, S. & Yamamoto, A. Two dimensional thin layer chromatographic separation of polar lipids and determination of phospholipids by phosphorus analysis of spots. Lipids 5, 494-496 (1970).
18. Sangle, G.V., Chowdhury, S.K., Xie, X., Stelmack, G.L., Halayko, A.J. & Shen, G.X. Impairment of mitochondrial respiratory chain activity in aortic endothelial cells induced by glycated low-density lipoprotein. Free Radic Biol Med 48, 781-790 (2010).
19. Chau, T.A., McCully, M.L., Brintnell, W., An, G., Kasper, K.J., Vines, E.D., Kubes, P., Haeryfar, S.M., McCormick, J.K., Cairns, E., Heinrichs, D.E. & Madrenas, J. Toll-like receptor 2 ligands on the staphylococcal cell wall downregulate superantigen-induced T cell activation and prevent toxic shock syndrome. Nat Med 15, 641-648 (2009).
20. Madrenas, J., Wange, R.L., Wang, J.L., Isakov, N., Samelson, L.E. & Germain, R.N. Zeta phosphorylation without ZAP-70 activation induced by TCR antagonists or partial agonists. Science 267, 515-518 (1995).
21. Fairhead, T., Lian, D., McCully, M.L., Garcia, B., Zhong, R. & Madrenas, J. RIP2 is required for NOD signaling but not for Th1 cell differentiation and cellular allograft rejection. Am J Transplant 8, 1143-1150 (2008).
22. Livak, K.J. & Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 25, 402-408 (2001).

125
23. Zhang, Z., Zhu, L., Quan, D., Garcia, B., Ozcay, N., Duff, J., Stiller, C., Lazarovits, A., Grant, D. & Zhong, R. Pattern of liver, kidney, heart, and intestine allograft rejection in different mouse strain combinations. Transplantation 62, 1267-1272 (1996).
24. Snyers, L., Umlauf, E. & Prohaska, R. Association of stomatin with lipid-protein complexes in the plasma membrane and the endocytic compartment. Eur J Cell Biol 78, 802-812 (1999).
25. Snyers, L., Thines-Sempoux, D. & Prohaska, R. Colocalization of stomatin (band 7.2b) and actin microfilaments in UAC epithelial cells. Eur J Cell Biol 73, 281-285 (1997).
26. Salzer, U. & Prohaska, R. Stomatin, flotillin-1, and flotillin-2 are major integral proteins of erythrocyte lipid rafts. Blood 97, 1141-1143 (2001).
27. Mairhofer, M., Steiner, M., Salzer, U. & Prohaska, R. Stomatin-like protein-1 interacts with stomatin and is targeted to late endosomes. J Biol Chem 284, 29218-29229 (2009).
28. Morrow, I.C. & Parton, R.G. Flotillins and the PHB domain protein family: rafts, worms and anaesthetics. Traffic 6, 725-740 (2005).
29. Langhorst, M.F., Reuter, A. & Stuermer, C.A. Scaffolding microdomains and beyond: the function of reggie/flotillin proteins. Cell Mol Life Sci 62, 2228-2240 (2005).
30. Hajek, P., Chomyn, A. & Attardi, G. Identification of a novel mitochondrial complex containing mitofusin 2 and stomatin-like protein 2. J Biol Chem 282, 5670-5681 (2007).
31. Da Cruz, S., Parone, P.A., Gonzalo, P., Bienvenut, W.V., Tondera, D., Jourdain, A., Quadroni, M. & Martinou, J.C. SLP-2 interacts with prohibitins in the mitochondrial inner membrane and contributes to their stability. Biochim Biophys Acta 1783, 904-911 (2008).
32. Sorice, M., Manganelli, V., Matarrese, P., Tinari, A., Misasi, R., Malorni, W. & Garofalo, T. Cardiolipin-enriched raft-like microdomains are essential activating platforms for apoptotic signals on mitochondria. FEBS Lett 583, 2447-2450 (2009).
33. Osman, C., Haag, M., Potting, C., Rodenfels, J., Dip, P.V., Wieland, F.T., Brugger, B., Westermann, B. & Langer, T. The genetic interactome of prohibitins: coordinated control of cardiolipin and phosphatidylethanolamine by conserved regulators in mitochondria. J Cell Biol 184, 583-596 (2009).
34. Osman, C., Voelker, D.R. & Langer, T. Making heads or tails of phospholipids in mitochondria. J Cell Biol 192, 7-16 (2011).

126
35. Frauwirth, K.A., Riley, J.L., Harris, M.H., Parry, R.V., Rathmell, J.C., Plas, D.R., Elstrom, R.L., June, C.H. & Thompson, C.B. The CD28 signaling pathway regulates glucose metabolism. Immunity 16, 769-777 (2002).
36. Acin-Perez, R., Fernandez-Silva, P., Peleato, M.L., Perez-Martos, A. & Enriquez, J.A. Respiratory active mitochondrial supercomplexes. Mol Cell 32, 529-539 (2008).
37. Osman, C., Merkwirth, C. & Langer, T. Prohibitins and the functional compartmentalization of mitochondrial membranes. J Cell Sci 122, 3823-3830 (2009).
38. Fry, M. & Green, D.E. Cardiolipin requirement for electron transfer in complex I and III of the mitochondrial respiratory chain. J Biol Chem 256, 1874-1880 (1981).
39. Garofalo, T., Giammarioli, A.M., Misasi, R., Tinari, A., Manganelli, V., Gambardella, L., Pavan, A., Malorni, W. & Sorice, M. Lipid microdomains contribute to apoptosis-associated modifications of mitochondria in T cells. Cell Death Differ 12, 1378-1389 (2005).
40. Li, Q., Vande Velde, C., Israelson, A., Xie, J., Bailey, A.O., Dong, M.Q., Chun, S.J., Roy, T., Winer, L., Yates, J.R., Capaldi, R.A., Cleveland, D.W. & Miller, T.M. ALS-linked mutant superoxide dismutase 1 (SOD1) alters mitochondrial protein composition and decreases protein import. Proc Natl Acad Sci U S A 107, 21146-21151 (2010).
41. Favaro, E., Ramachandran, A., McCormick, R., Gee, H., Blancher, C., Crosby, M., Devlin, C., Blick, C., Buffa, F., Li, J.L., Vojnovic, B., Pires das Neves, R., Glazer, P., Iborra, F., Ivan, M., Ragoussis, J. & Harris, A.L. MicroRNA-210 regulates mitochondrial free radical response to hypoxia and krebs cycle in cancer cells by targeting iron sulfur cluster protein ISCU. PLoS One 5, e10345 (2010).
42. McIntosh, R., Lee, S., Ghio, A.J., Xi, J., Zhu, M., Shen, X., Chanoit, G., Zvara, D.A. & Xu, Z. The critical role of intracellular zinc in adenosine A(2) receptor activation induced cardioprotection against reperfusion injury. J Mol Cell Cardiol 49, 41-47 (2010).
43. Finsterer, J. Leigh and Leigh-like syndrome in children and adults. Pediatr Neurol 39, 223-235 (2008).
44. Rich, P.R. & Marechal, A. The mitochondrial respiratory chain. Essays Biochem 47, 1-23 (2010).
45. Steglich, G., Neupert, W. & Langer, T. Prohibitins regulate membrane protein degradation by the m-AAA protease in mitochondria. Mol Cell Biol 19, 3435-3442 (1999).

127
46. Distelmaier, F., Koopman, W.J., van den Heuvel, L.P., Rodenburg, R.J., Mayatepek, E., Willems, P.H. & Smeitink, J.A. Mitochondrial complex I deficiency: from organelle dysfunction to clinical disease. Brain 132, 833-842 (2009).
47. Ma, Y.Y., Zhang, X.L., Wu, T.F., Liu, Y.P., Wang, Q., Zhang, Y., Song, J.Q., Wang, Y.J. & Yang, Y.L. Analysis of the Mitochondrial Complex I-V Enzyme Activities of Peripheral Leukocytes in Oxidative Phosphorylation Disorders. J Child Neurol Epub ahead of print (2011).
48. Kaminski, M.M., Sauer, S.W., Klemke, C.D., Suss, D., Okun, J.G., Krammer, P.H. & Gulow, K. Mitochondrial reactive oxygen species control T cell activation by regulating IL-2 and IL-4 expression: mechanism of ciprofloxacin-mediated immunosuppression. J Immunol 184, 4827-4841 (2010).
49. Michalek, R.D., Gerriets, V.A., Jacobs, S.R., Macintyre, A.N., MacIver, N.J., Mason, E.F., Sullivan, S.A., Nichols, A.G. & Rathmell, J.C. Cutting edge: distinct glycolytic and lipid oxidative metabolic programs are essential for effector and regulatory CD4+ T cell subsets. J Immunol 186, 3299-3303 (2011).
50. Bruzzone, S., Fruscione, F., Morando, S., Ferrando, T., Poggi, A., Garuti, A., D'Urso, A., Selmo, M., Benvenuto, F., Cea, M., Zoppoli, G., Moran, E., Soncini, D., Ballestrero, A., Sordat, B., Patrone, F., Mostoslavsky, R., Uccelli, A. & Nencioni, A. Catastrophic NAD+ depletion in activated T lymphocytes through Nampt inhibition reduces demyelination and disability in EAE. PLoS One 4, e7897 (2009).
51. Triepels, R.H., Van Den Heuvel, L.P., Trijbels, J.M. & Smeitink, J.A. Respiratory chain complex I deficiency. Am J Med Genet 106, 37-45 (2001).
52. Holen, K., Saltz, L.B., Hollywood, E., Burk, K. & Hanauske, A.R. The pharmacokinetics, toxicities, and biologic effects of FK866, a nicotinamide adenine dinucleotide biosynthesis inhibitor. Invest New Drugs 26, 45-51 (2008).
53. Garcia-Sanz, J.A. & Lenig, D. Translational control of interleukin 2 messenger RNA as a molecular mechanism of T cell anergy. The Journal of experimental medicine 184, 159-164 (1996).
54. Villarino, A.V., Katzman, S.D., Gallo, E., Miller, O., Jiang, S., McManus, M.T. & Abbas, A.K. Posttranscriptional silencing of effector cytokine mRNA underlies the anergic phenotype of self-reactive T cells. Immunity 34, 50-60 (2011).
55. Shaw, G. & Kamen, R. A conserved AU sequence from the 3' untranslated region of GM-CSF mRNA mediates selective mRNA degradation. Cell 46, 659-667 (1986).

128
56. Araki, K., Turner, A.P., Shaffer, V.O., Gangappa, S., Keller, S.A., Bachmann, M.F., Larsen, C.P. & Ahmed, R. mTOR regulates memory CD8 T-cell differentiation. Nature 460, 108-112 (2009).
57. Pearce, E.L., Walsh, M.C., Cejas, P.J., Harms, G.M., Shen, H., Wang, L.S., Jones, R.G. & Choi, Y. Enhancing CD8 T-cell memory by modulating fatty acid metabolism. Nature 460, 103-107 (2009).

129
Chapter 4 3
4 Plasma membrane and mitochondrial Stomatin-like Protein 2 pools coalesce at the immunological synapse during T cell activation
Stomatin-like protein 2 (SLP-2) is a member of the stomatin – prohibitin – flotillin –
HflC/K (SPFH) superfamily. Recent evidence indicates that SLP-2 is involved in the
organization of cardiolipin-enriched microdomains in mitochondrial membranes and the
regulation of mitochondrial biogenesis and function. In T cells, this role translates into
enhanced T cell activation. Although the major pool of SLP-2 is associated with
mitochondria, we show here that there is an additional pool of SLP-2 associated with the
plasma membrane of T cells. Both plasma membrane-associated and mitochondria-
associated pools of SLP-2 coalesce at the immunological synapse (IS) upon T cell
activation. SLP-2 is not required for formation of IS nor for the re-localization of
mitochondria to the IS because SLP-2-deficient T cells showed normal re-localization of
these organelles in response to T cell activation. Interestingly, upon T cell activation, we
found the surface pool of SLP-2 mostly excluded from the central supramolecular
activation complex, and enriched in the peripheral area of the IS where signalling TCR
microclusters are located. Based on these results, we propose that SLP-2 facilitates the
compartmentalization not only of mitochondrial membranes but also of the plasma
membrane into functional microdomains. In this latter location, SLP-2 may facilitate the
optimal assembly of TCR signalosome components. Our data also suggest that there may
be a net exchange of membrane material between mitochondria and plasma membrane,
explaining the presence of some mitochondrial proteins in the plasma membrane.
3 The material contained in this chapter is based on a submitted manuscript: Christie, D., Kirchhof, M.G.,
Vardhana, S., Dustin, M.L., and Madrenas, J. Mitochondrial and plasma membrane pools of Stomatin-like protein 2 coalesce at the immunological synapse during T cell activation. 2012

130
4.1 Introduction
Stomatin-like protein 2 (SLP-2), a widely expressed mitochondrial protein identified in
proteome analyses from several tissues and species, is a member of the stomatin family
as well as the stomatin – prohibitin – flotillin – HflC/K (SPFH) superfamily 1-4. In
mitochondria, SLP-2 associates tightly with the mitochondrial inner membrane. How
this association occurs is unclear because, unlike other stomatin family members, SLP-2
lacks a putative transmembrane domain. It contains though a SPFH domain, a highly
conserved region found in SPFH superfamily members 5-8, and this domain has been
linked to protein : lipid membrane interactions 9-11.
Recent studies have begun to shed light on the function of SLP-2. Similar to other SPFH
family members, SLP-2 has been found in large protein complexes and has been shown
to interact with mitochondrial proteins, including prohibitin (PHB) 1 and 2 as well as
mitofusin 2 5, 6, 8. We also have shown that SLP-2 binds directly to cardiolipin and can
regulate the localization of the prohibitin complex to cardiolipin-enriched domains in the
mitochondrial membrane 12. Furthermore, over-expression of SLP-2 leads to increased
mitochondrial biogenesis and function 5, 6, 8 whereas SLP-2 depletion results in decreased
mitochondrial transmembrane potential, reduced mitochondrial calcium uptake in
response to cell stimulation, enhanced apoptotic responses to cell stress and increased
degradation of mitochondrial proteins 5, 8, 12-14 (Christie et al, manuscript submitted). All
this evidence led us to propose that SLP-2 regulates all these mitochondrial functions (i.e.
energy production, calcium buffering and apoptosis) by organizing mitochondrial
membranes into defined cardiolipin-enriched microdomains, which then facilitate the
optimal assembly of membrane-associated molecular complexes.
Although most recent work on SLP-2 has focused on the function in mitochondria, we
originally identified SLP-2 in the glycolipid-enriched, detergent-insoluble microdomains
of human T cells activated through the T cell receptor 12. In these cells, we demonstrated
that SLP-2 was recruited to glycolipid-enriched detergent-insoluble microdomains and
interacted with numerous components of the TCR signalosome upon T cell stimulation,
including CD3, ZAP-70, LAT, lck and PLCγ. Furthermore, the level of SLP-2
expression correlated with the response of T cells to stimulation. Under conditions of

131
SLP-2 over-expression, T cells had significantly increased responses while knockdown of
SLP-2 resulted in significantly decreased T cell signalling. Together, these results
suggested that SLP-2 plays an important role in T cell activation, either by increasing
mitochondrial biogenesis 6 or by regulating T cell signalling at the plasma membrane 12.
To investigate the role of SLP-2 during T cell activation, we utilized a Jurkat T cell line
stably transfected with an inducible construct encoding either full-length SLP-2 tagged
with gfp or a gfp-tagged mutant construct lacking the amino-terminal region. As
expected, we found that most SLP-2 was associated with mitochondria. This primary
location was determined by an amino-terminal mitochondrial-targeting sequence of SLP-
2. Interestingly, a small pool of SLP-2 was also detected in association with the plasma
membrane. Upon T cell stimulation both pools of SLP-2 coalesced at the immunological
synapse (IS), and localized mostly in the peripheral supramolecular activation complex
(pSMAC). SLP-2 was not required for mitochondrial trafficking to the IS. Our data
indicate that there is a small pool of SLP-2 in the plasma membrane that, by analogy with
the mitochondrial pool of this protein, may play a role in the compartmentalization of the
plasma membrane, ensuring optimal assembly of multimolecular complexes.
4.2 Material and Methods
4.2.1 Cells
Jurkat T cells (E6.1) were obtained from American Type Culture Collection (Manassas,
VA) and cultured in supplemented RPMI 1640 medium. The B lymphoblastoid cell line,
LG2, was kindly provided by Dr. Eric Long (NIAID, NIH, Rockville, MD) and cultured
in standard supplemented RPMI 1640 media.
4.2.2 Plasmids, siRNA and T cell transfectants
Human SLP-2 cDNA was subcloned into the pEGFPN1 expression vector (Clontech Inc.
Palo Alto, CA) to create an in-frame translational fusion of the 3’ end of SLP-2 to gfp, as
previously described 15, 16. Subsequently, the SLP-2-gfp fusion was cloned into the
doxycycline inducible pBIG2i vector 15. A mutant construct lacking the amino-terminal

132
region (deletion of amino acid residues 1-77) was generated and cloned into pBIG2i.
Stable transfectants were generated by electroporating linearized plasmid into Jurkat E6.1
T cells and screening for stable expression of SLP-2-gfp. Doxycycline (Sigma, St. Louis,
MO) was added to overnight cultures at 1 µg/mL to induce SLP-2-gfp expression, which
was monitored by direct flow cytometry (Becton Dickinson [BD], San Jose, CA).
4.2.3 Mice
A full-length genomic fragment containing the mouse SLP-2 gene was cloned within the
lox sites of the 3LoxP3NwCD vector. Upon sequence confirmation, the plasmid was
electroporated into C57BL/6 embryonic stem cells and clones were selected for
neomycin resistance. The resistant clones were screened by Southern blot and PCR
analysis to confirm homologous recombination of the SLP-2-floxed DNA sequence.
Clones containing the SLP-2-floxed sequence were injected into B6/Try blastocysts and
implanted into a pseudo-pregnant mouse. The offspring was selected by chimeric coat
colour and backcrossed with C57BL/6 mice to produce SLP-2lox/wt mice in the C57BL/6
background. SLP-2 floxed mice were crossed with C57BL/6 mice transgenic for Cre
recombinase under the control of the CD4 promoter, purchased from Taconic (Hudson,
NY) 17, to generate a T cell specific SLP-2 knockout strain (SLP-2 T-K/O, Christie et al,
manuscript submitted). SLP-2 floxed mice lacking Cre were used as control mice.
Breeding colonies were derived from the same original SLP-2 floxed breeders and kept in
parallel. Mice were maintained in the animal facility at the University of Western
Ontario with approval from the Animal Use Subcommittee in accordance with the
Canadian Council on Animal Care Guidelines.
4.2.4 Antibodies
A rabbit polyclonal antiserum against human SLP-2 was generated by immunization with
a peptide spanning amino acids 343 to 356 (ProSci Inc., Poway, CA). Polyclonal
antibodies against SLP-2 (Protein Tech Group, Chicago, IL) and gfp (BD Bioscience,
San Jose, CA) were used in these studies. Pre-immune rabbit serum was used as an
isotype control (ProSci Inc., Poway, CA). Monoclonal antibodies against β-actin (Santa
Cruz Biotechnology, Santa Cruz, CA), the α-subunit of Complex V (MitoSciences,

133
Eugene, OR), biotin (Jackson ImmunoResearch, West Grove, PA), Caspase 3 (Cell
Signaling, Danvers, MA), GAPDH (Chemicon International, Temecula, CA) and PE-
labelled anti-CD3 (UCHT-I) were used (BD Bioscience, San Jose, CA) were used.
4.2.5 Mitochondria isolation
Intact mitochondria were isolated from 20 x 106 Jurkat T cells, SLP-2 T-K/O or wild type
control cells using the Qproteome Mitochondria Isolation Kit (Qiagen). Cytosolic
fractions were concentrated in Amicon Ultra-0.5 centrifugal filter units with Ultracel-30
membrane (Millipore, Billerica, MA). Mitochondrial and cytosolic preparations were
mixed with sample buffer containing β-mercaptoethanol, boiled, and analyzed by sodium
dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE).
4.2.6 Immunoprecipitations
Protein A or protein G agarose beads were coated with either SLP-2 antisera, pre-immune
sera, anti-gfp or anti-biotin antibodies overnight at 4oC 16. Beads were washed to remove
excess antibody and cell lysates were incubated with the antibody-coated beads overnight
at 4oC. Following immunoprecipitation, unbound proteins were removed and antibody-
bound protein complexes were eluted in sample buffer with β-mercaptoethanol.
Immunoprecipitated samples were separated by SDS-PAGE and immunoblotted with the
appropriate antibodies.
4.2.7 Cell surface biotinylation
Jurkat or mouse T cells were incubated with EZ-link sulfo-NHS-biotin (Pierce, Rockford,
IL) in PBS pH 8.0, for 30 minutes at room temperature. Cells were then washed three
times with cold PBS, lysed and immunoprecipitated for biotin 15, 18. Lysates and
immunoprecipitates were separated by SDS-PAGE and blotted with antibodies against
SLP-2.
4.2.8 Confocal microscopy
Confocal microscopy was performed with a Zeiss LSM 510 microscope. Jurkat T cells or
murine T cells (1x106/ml) were incubated on poly-L-lysine-coated (0.01%, Sigma) glass

134
bottom microwell dishes (MatTek Corp., MA) for 10 minutes to promote cell adherence
at 37ºC. SLP-2-gfp localization and mitochondrial movement was monitored by staining
cells with 100 nM MitoTracker Red CMXRos (Invitrogen, Burlington, ON, Canada) for
15 min at 37°C in complete RPMI 1640. SLP-2-gfp distribution during IS formation was
assessed by culturing doxycycline-induced SLP-2-gfp stably transfected T cells with LG2
antigen-presenting cells (APC) pre-incubated with 1 µg/ml staphylococcal enterotoxin E
(SEE) for either 10 or 30 minutes. Following the allotted time of co-incubation, the T
cell-APC conjugates were rapidly fixed with 4% paraformaldehyde, washed with
PBS+1% FCS and stained with PE conjugated anti-CD3 for 30 minutes on ice. For
experiments imaging mitochondrial localization in primary mouse cells, T cells were
isolated by magnetic separation using the MACS pan T cell isolation kit (Miltenyi Biotec,
Auburn, CA). Cells were stained with MitoTracker Red, stimulated with dynabeads
mouse T activator CD3/CD28 (Invitrogen, Burlington, ON, Canada) for 30 minutes and
cells were fixed and imaged 19. For experiments using planar membranes, glass-
supported dioleoylphosphatidylcholine bilayers incorporating Cy5-ICAM-1 (300
molecules/µm2) and 0.1% cap-biotin were prepared in a Bioptechs flow cell. Unlabelled
Streptavidin (8 µg/mL) and Cy3-conjugated anti-human CD3 (OKT3 clone, 10 µg/mL),
were loaded sequentially in HBS/HSA buffer (Hepes buffered saline supplemented with
5 mM glucose, 2 mM MgCl2, 1 mM CaCl2, and 1% human serum albumin). Jurkat T
cells were also suspended in HBS/HAS buffer. All microscopy was performed on an
automated microscope with a Hamamatsu USA Orca-ER cooled CCD camera. The
hardware on the microscope was controlled using IPLAB software (Scanalytics) on a
PowerMac G4 Macintosh computer. Images were exposed in wide-field for 1-2 s at a
resolution of 0.11 µm per pixel using the 60x 1.45 N.A. objective. Interference reflection
microscopy (IRM) is based on destructive interference in green light reflected from the
bilayer-cell interface leading to a dark area where cells are in close contact with the
bilayer. Images of cell interaction with the planar membrane were collected by IRM and
fluorescent images to examine re-localization of T cell proteins were collected by total
internal reflection fluorescence microscopy (TIRFM). Images were inspected using
Metamorph (Molecular Devices).

135
4.3 Results
4.3.1 The largest cellular pool of SLP-2 is associated with mitochondria
In order to assess the subcellular localization and movement of SLP-2 in T cells, we
constructed stably transfected Jurkat T cells with a doxycycline inducible construct
encoding SLP-2 tagged with gfp at the C-terminus. In addition, we also constructed a
truncated version of SLP-2 lacking the N-terminus of the protein (ΔN-SLP-2-gfp), as this
region contains a predicted mitochondrial targeting sequence 8. Transfected T cells were
visualized using confocal microscopy (Fig. 4.1a). As expected, we found that most SLP-
2-gfp was distributed in aggregates within the cytosol, which were shown to be
mitochondria by staining with the mitochondrial dye MitoTracker Red. In the absence of
the predicted mitochondrial targeting sequence (ΔN-SLP-2-gfp), the protein failed to
partition selectively in mitochondria and was instead found in a diffuse cytosolic pattern,
confirming the functionality of the mitochondrial-targeting domain in the N-terminus of
SLP-2.
The mitochondrial localization of SLP-2 was further supported by subcellular
fractionation and western blotting. As shown in Fig. 4.1b, both endogenous SLP-2 and
SLP-2-gfp were detected in the mitochondrial but not the cytosolic fractions of
transfected Jurkat T cells. The SLP-2-gfp blots showed intermediate degradation
products from the full length chimeric construct. The ΔN-SLP-2-gfp was absent in the
mitochondrial fraction and detected only in the cytosolic fraction, further confirming the
presence of a mitochondrial targeting domain at the amino terminus of SLP-2.
The previous results were corroborated using T cells from wildtype and T-cell-specific
SLP-2 knockout mice (Fig. 4.1c). Similar to human SLP-2, murine SLP-2 also partitioned
in mitochondria. As expected, the SLP-2 T-K/O cells had no detectable SLP-2, verifying
the identity of the mitochondrial SLP-2 band in the wild type mice. Altogether, these
results confirmed the predominant mitochondrial location of SLP-2 and validated the use
of the inducible SLP-2-gfp transfected T cells.

136
Figure 4.1: SLP-2 is a mitochondrial protein.
(a) Full-length SLP-2-gfp and ΔN-SLP-2-gfp were subcloned into a doxycycline-
inducible vector and transfected into Jurkat T cells. These stable transfectants were
imaged by confocal microscopy for SLP-2-gfp (first column of micrographs – green), or
after staining with MitoTracker Red (second column of micrographs – red). The third
colum of photomicrographs show overlapping of red and green signals as yellow signal.
(b) Mitochondrial and cytosolic fractions of parental, SLP-2-gfp and ΔN-SLP-2-gfp T
cell transfectants were isolated by differential centrifugation and immunoblotted for SLP-
2, the α-subunit of ATP synthase and actin. (c) Mitochondrial and cytosolic fractions
were isolated from T cells of wild type mice and T-cell specific SLP-2 conditional
knockout mice and blotted as in B. These results are representative of at least 3
independent experiments, and of more than 100 imaged cells.
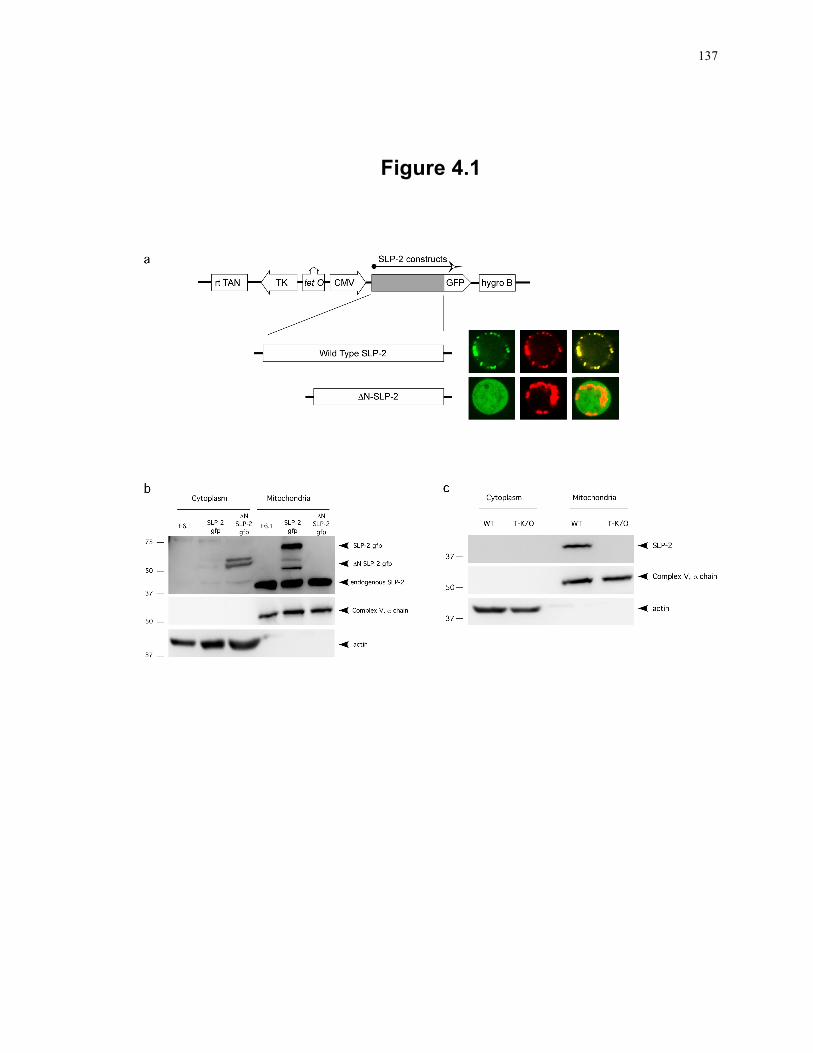
137
Figure 4.1

138
4.3.2 Detection of a small pool of SLP-2 associated with the plasma membrane
Although initial studies of SLP-2 have focused on the mitochondrial localization 5, 6, 8, we
originally identified SLP-2 in total glycolipid-enriched, detergent-insoluble
microdomains from activated T cells 12. This suggested to us that there may be SLP-2
associated with other membranes, potentially plasma membranes. In order to test the
presence of SLP-2 associated with the plasma membrane, T cells were biotinylated and a
a biotin immunoprecipitation performed. In these experiments, untransfected Jurkat T
cells as well as SLP-2-gfp and ΔN-SLP-2-gfp stably transfected Jurkat T cells were used
to determine the association of full length SLP-2 and SLP-2 lacking the mitochondrial
targeting domain with T cell surface molecules (Fig. 4.2a). Both the endogenous and
SLP-2-gfp were found in association with cell surface molecules. In addition, the ΔN-
SLP-2-gfp was also found in association with the plasma membrane, indicating that the
mitochondrial targeting sequence is not required for association with surface molecules.
However, the interaction of ΔN-SLP-2-gfp with membrane proteins was much less
efficient because when expression of full length SLP-2-gfp and ΔN-SLP-2-gfp were
titrated the full-length SLP-2 associated with the plasma membrane at lower levels of
expression compared to ΔN-SLP-2-gfp (Fig. 4.2b). Even the very low level of leaky
expression of SLP-2-gfp in uninduced cells was sufficient for some SLP-2-gfp to interact
with plasma membrane proteins while ΔN-SLP-2-gfp surface pool was only detected
when cells expressed high levels of this mutant (by induction with 0.2 µg/mL of
doxycycline or higher).
The small surface-associated pool of SLP-2 was also found in mouse T cells. Biotin
immunoprecipitation was performed with wild type and SLP-2 T-K/O T cells and SLP-2
was found to co-precipitate with surface proteins in the wild type T cells but not in cells
lacking SLP-2 (Fig. 4.2c). This result confirms the existence of a plasma membrane-
associated pool of SLP-2 in association with surface proteins with an extracellular
domain (i.e. biotinylated).

139
Figure 4.2: A small pool of SLP-2 is associated with the plasma membrane.
(a) Intact parental Jurkat T cells, as well as Jurkat T cells stably transfected with full
length SLP-2-gfp or ΔN-SLP-2-gfp were biotinylated. Next, cells were lysed and surface
proteins were immunoprecipitated with an antibody against biotin. The
immunoprecipitate samples (Biotin ip) were immunoblotted for SLP-2 to detect the pool
of SLP-2 associated with surface molecules. Whole cell lysates (WCL) from the
transfectants were blotted to show expression levels of the transgenes. Samples were also
blotted for caspase 3 as a negative control for surface protein pull-down. (b) Increasing
levels of SLP-2-gfp or ΔN-SLP-2-gfp were induced with increasing concentrations of
doxycycline and biotin immunoprecipitation was performed. Whole cell lysates (WCL)
and biotin immunoprecipitates (biotin ip) were blotted as in a. (c) T cells isolated from
wild type and SLP-2 T-K/O mice were analyzed by biotin immunoprecipitation and
blotted as in a. Samples were also blotted for actin as a loading control for SLP-2 T-K/O
samples and also for GAPDH as a negative control for surface protein pull-down. These
results are representative of at least 3 independent experiments.
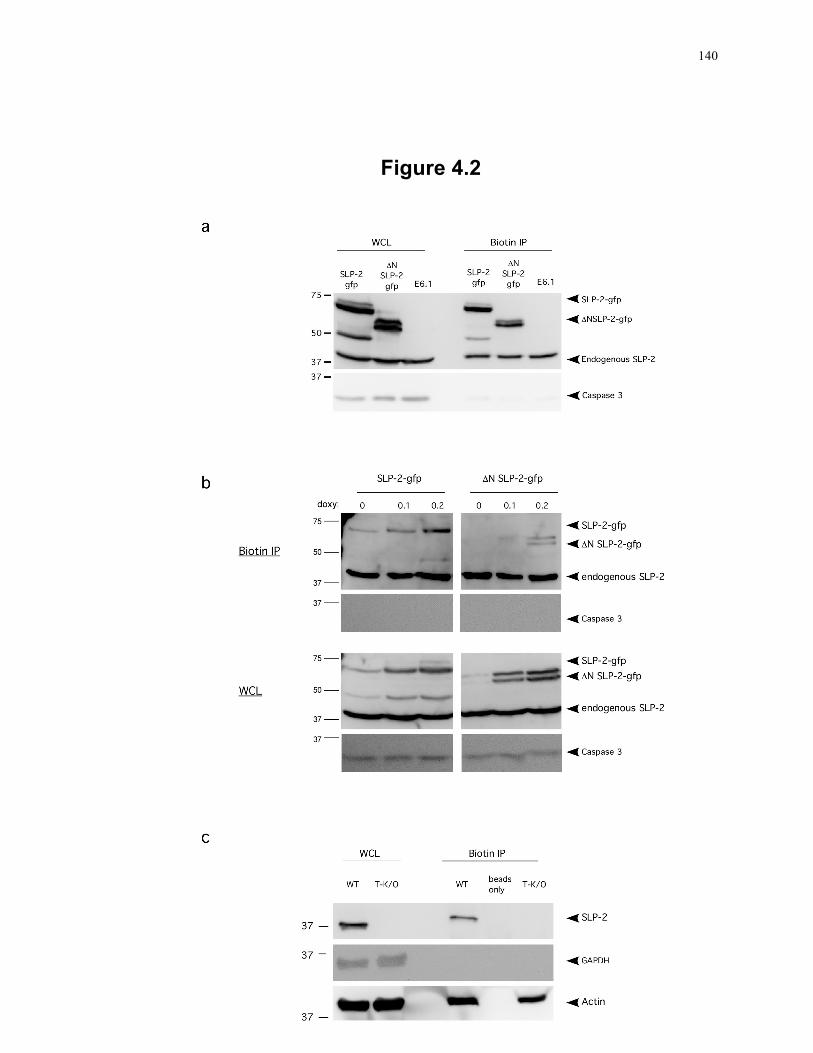
140
Figure 4.2

141
4.3.3 Homo-oligomerization of SLP-2
We have previously shown that SLP-2 is capable of interacting with numerous proteins,
including multiple components of the TCR signalosome and prohibitins 6, 12. However, it
is currently unclear if SLP-2 is capable of homo-oligomerization in vivo, without
exogenous cross-linkers. This may be an important feature of SLP-2 required to organize
specialized membrane microdomains. To test for homo-oligomerization of SLP-2, we
utilized the gfp-tagged constructs to examine co-precipitation of endogenous SLP-2 with
gfp-tagged SLP-2. In these experiments, parental Jurkat cells, SLP-2-gfp or ΔN-SLP-2-
gfp expressing Jurkat cells were immunoprecipitated with anti-gfp antibodies and
samples were blotted sequentially for SLP-2 and gfp (Fig. 4.3). As expected, gfp
immunoprecipitation of the parental cells failed to bring down any proteins,
demonstrating the specificity of this system. GFP-immunoprecipitation of SLP-2-gfp co-
precipitated endogenous SLP-2, documenting the ability of full-length SLP-2 to homo-
oligomerize. Of interest, ΔN-SLP-2-gfp did not co-precipitate with endogenous SLP-2 in
significant amounts. This suggests that the N-terminal region of SLP-2, by itself or
through its effect on mitochondrial targeting, is important for effective homo-
oligomerization of SLP-2.
4.3.4 SLP-2 redistributes to the immunological synapse during T cell activation
Next, we investigated the intracellular movement of SLP-2 during T cell activation.
Upon activation of SLP-2-gfp stably transfected T cells with APC and SEE, both the
plasma membrane-associated pool and the intracellular pool of SLP-2 polarized towards
putative immunological synapses, coalescing into major clusters close to the plasma
membrane, in the periphery of the IS and underneath the T cell-APC interface (Fig. 4.4a).
At the cell population level, such polarization was apparent in 60% of T cells forming
putative immunological synapses (Fig. 4.4b). Video capture of the redistribution of SLP-
2-gfp during T cell activation showed that as activation proceeded, greater than 80% of
the polarized SLP-2-gfp partitioned in the periphery of the synapse and underneath

142
Figure 4.3: Homo-oligomerization of SLP-2.
Parental Jurkat T cells and Jurkat T cells stably transfected with SLP-2-gfp or ΔN-SLP-2-
gfp were lysed. Lysates were used for immunoprecipitation with anti-GFP antibodies
(GFP ip), and immunoblotted serially for SLP-2 and gfp. Whole cell lysates (WCL) were
also blotted to show expression levels of endogenous SLP-2 and the SLP-2 transgenes.
This result is representative of at least 3 independent experiments.
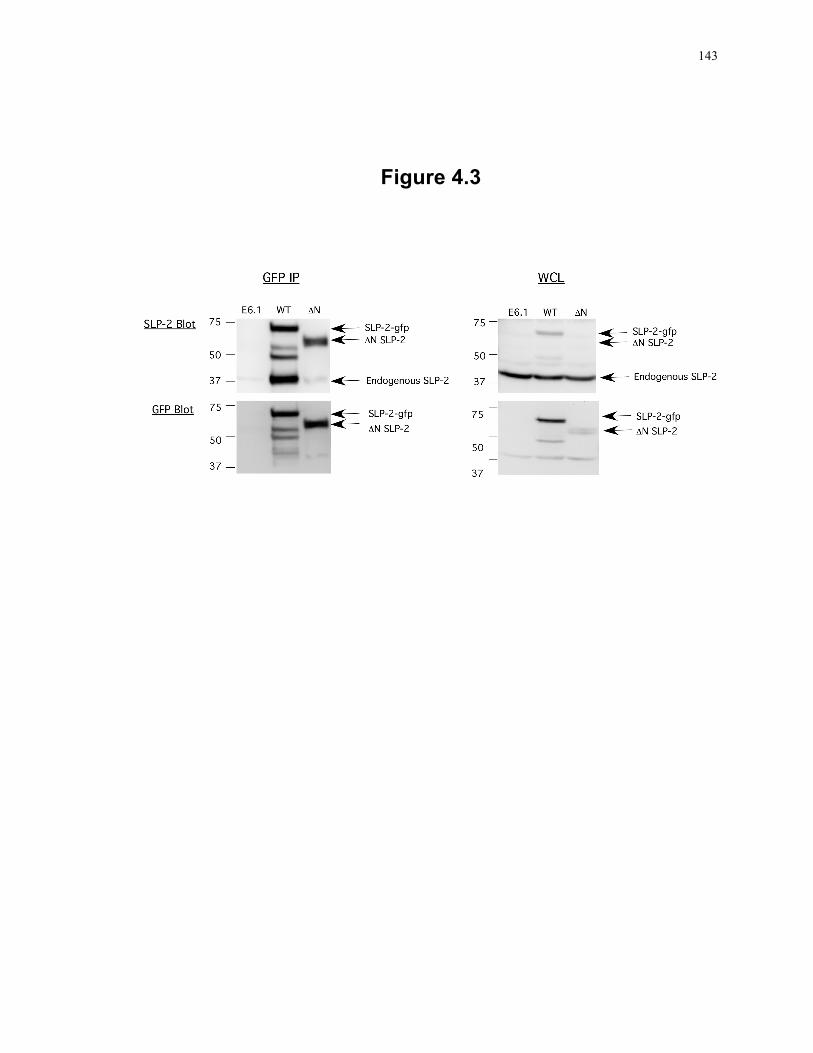
143
Figure 4.3

144
Figure 4.4: SLP-2 polarizes to the immunological synapse during T cell activation.
(a) Stable SLP-2-gfp-transfected E6.1 Jurkat T cells were cultured with APC in the
presence (+SEE) or absence (-SEE) of SEE and examined by confocal microscopy for the
formation of putative synapses (identified as CD3-positive red clusters at the interface
between T cell and APC) and for the location of SLP-2-gfp signal (green). Images are
representative of at least 50 putative IS. Concomitant studies done with control
transfected T cells demonstrated that expression of SLP-2-gfp did not interfere with IS
formation. (b) Confocal images collected from 50 putative IS images (in A) were
analyzed for SLP-2-gfp intracellular localization. SLP-2-gfp signal in un-stimulated (0
min) or stimulated cells (30 min) was classified as predominantly proximal to the IS
(white bars), predominantly distal to the IS (black bars), or diffuse throughout the cell
(grey bars). (c) Redistribution of SLP-2 during T cell activation is shown by a series of
videomicroscopy capture images during IS formation. The dotted circle outlines the APC
interacting with the Jurkat T cell and the arrow indicates the mature IS with the SLP-2
localization. (d) SLP-2-gfp-transfected Jurkat T cells were stimulated with antibodies
against CD3 and examined by confocal microscopy. Photobleaching was induced at the
arrow-indicated site and regaining of the signal at that site was monitored for 6 minutes at
20 second intervals. Non-specific distribution is documented by progressive regaining of
signal with non-bleached SLP-2, while active distribution correlates with lack of
regaining of signal.
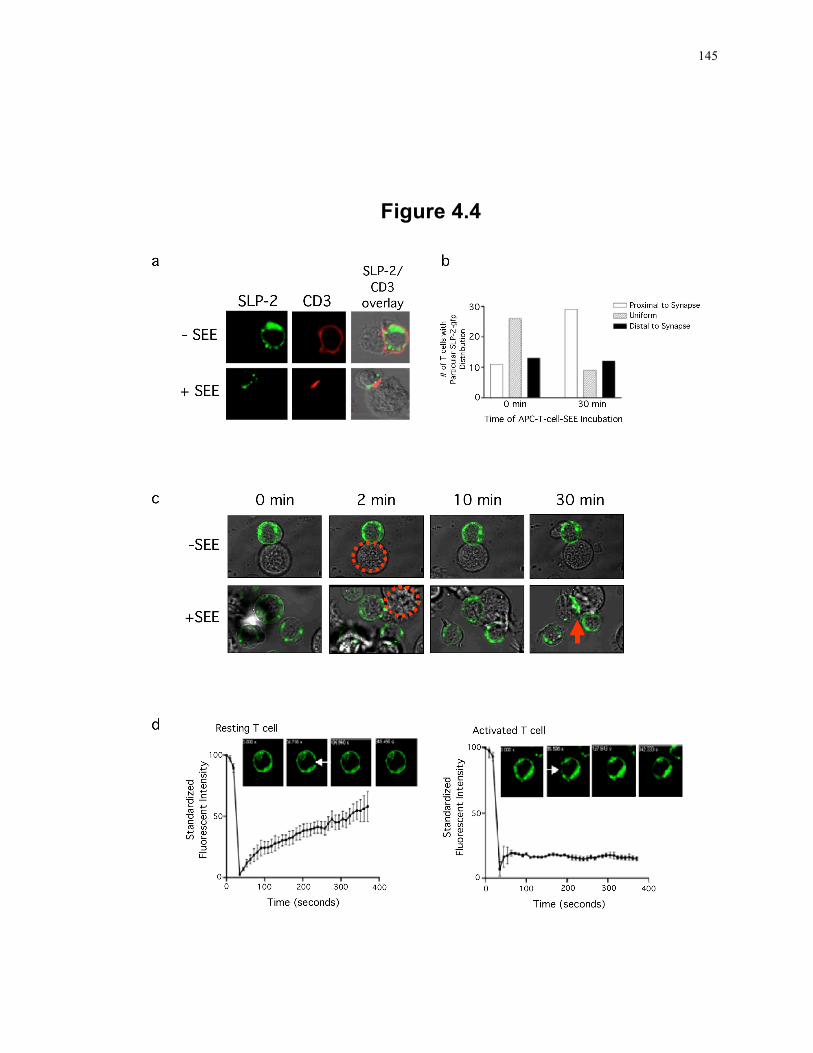
145
Figure 4.4

146
the putative IS at late time points (Fig. 4.4c). Only a small fraction of SLP-2 was
detectable in the centre of the synapse. Accumulation of SLP-2 in the periphery of the IS
mirrored the distribution of pSMACs, known to be enriched in LFA-1. The SLP-2
clusters during T cell activation were actively retained and stable as shown by
photobleaching experiments (Fig. 4.4d). In these experiments, the SLP-2-gfp signal was
restored after 400 seconds of bleaching the plasma membrane of non-stimulated T cells
but was not restored when SLP-2-gfp clusters in OKT3-stimulated T cells were bleached
within the same timeline, indicating that T cell activation was associated with stable SLP-
2-gfp clusters unable to freely move within the cell.
4.3.5 The SLP-2 pools predominantly partition in the pSMAC of the immunological synapse
To further refine our data concerning SLP-2-gfp redistribution to the IS, we used
supported planar bilayers containing anti-CD3 and ICAM-1 to which T cells form IS-like
structures. This system has excellent optics for resolution of components in the IS. With
this system, we visualized the formation of IS by Jurkat cells expressing SLP-2-gfp (Fig.
4.5). Under conditions of doxycycline-induced SLP-2-gfp overexpression, we confirmed
the presence of two pools of this molecule. The plasma membrane-associated pool of
SLP-2 was visualized by total internal reflection fluorescence microscopy (TIRFM) as
clusters located at less than 200nm from the glass plane (i.e., sites of TCR and LFA-1
engagement) (Fig. 4.5a). The small plasma membrane pool of SLP-2 distributed
uniformly in a granular appearance interspersed with TCR microclusters in the nascent IS
(Fig. 4.5a). As activation proceeded, the TCR microclusters were surrounded by densely
packed SLP-2 clusters. These TCR microclusters weakly excluded the SLP-2 clusters
resulting in up to 50% reduction of SLP-2 fluorescence in the cSMACs.

147
Figure 4.5: Redistribution of plasma membrane-associated SLP-2 and
mitochondria-associated SLP-2 during immunological synapse formation.
Jurkat T cells stably expressing SLP-2-gfp were incubated on planar membranes
containing ICAM-1 and anti-CD3 to induce immunosynapse formation. Cells were
imaged at early (<5 min) and late (15-30 min) stages of immunosynapse formation to
demonstrate SLP-2 redistribution upon TCR ligation. (a) Plasma membrane-associated
SLP-2 redistribution during immunosynapse formation was imaged by TIRFM, to
eliminate signal from the mitochondrial pool of SLP-2. Cell contact with the planar
membrane was imaged by IRM, TCR images were obtained by wide-field fluorescence
microscopy and the image overlay represents SLP-2-gfp as green and TCR as red. (b)
Total SLP-2-gfp, including both plasma membrane-associated and mitochondrial pools
was imaged at early and late stages of immunosynapse formation. The bright field image
shows cells being imaged, IRM images show contact with the bilayer as dark areas, and
the TCR, ICAM-1 and SLP-2-gfp fluorescence channels are shown in gray scale and two
red-green merges (green is always SLP-2). The dotted lines in the ICAM-1 picture
represent, from the periphery to the centre of the picture, the outer boundaries of the
distal SMAC, of the pSMAC, and of the cSMAC. Images are representative 3 separate
experiments. (c) SLP-2-gfp expressing Jurkat cells were transfected with mitochondria-
targeted RFP to verify mitochondrial localization of intracellular SLP-2-gfp. Images were
obtained by wide-field fluorescence microscopy and are representative of three separate
experiments. The dotted lines in the ICAM-1 image are as described in b. The
percentage of cells showing segregation of the mitochondrial pool of SLP-2 away from
the cSMAC is shown in Table 4.1 and this organization of SLP-2 into the pSMAC
requires anti-CD3 ligation (Table 4.2).
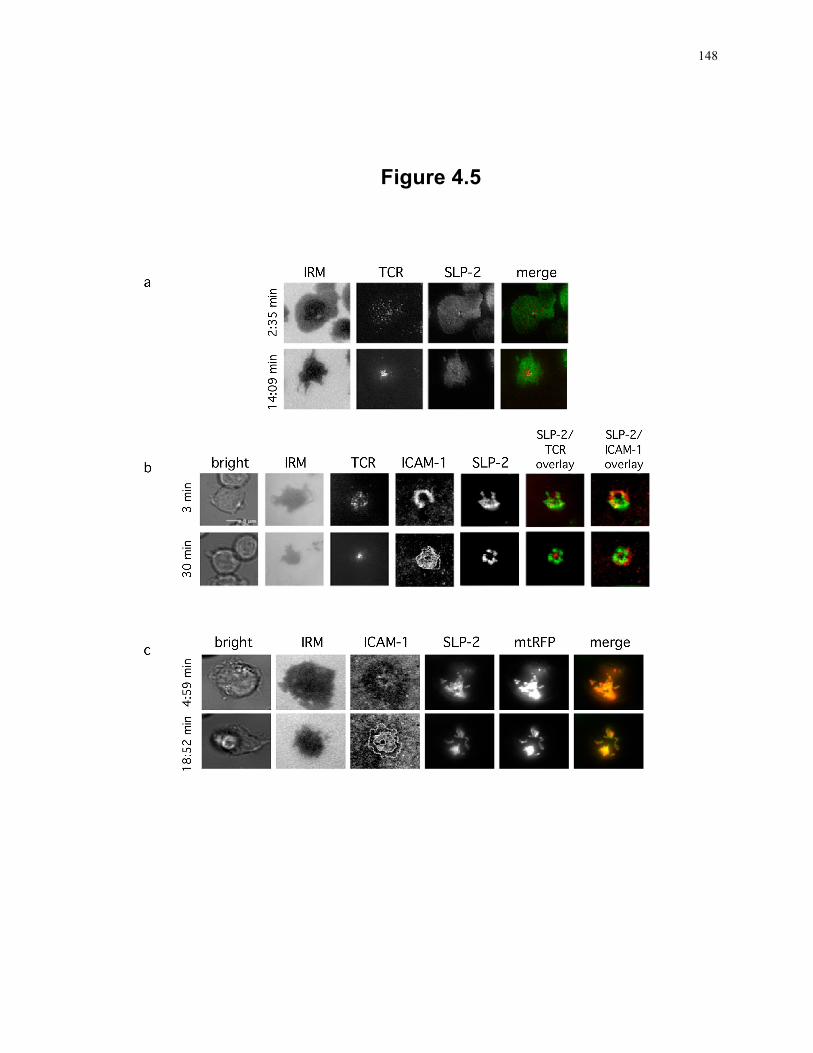
148
Figure 4.5

149
In addition, the major mitochondria-associated pool of SLP-2-gfp located at more than
200 nm from the plasma membrane, and also polarized to the IS and segregated from the
cSMACs to distribute close to the pSMACs as activation proceeded (Fig. 4.5b and Table
4.1). The mitochondrial origin of this pool of SLP-2 was confirmed by co-localization of
the gfp signal with a mitochondria-targeted red fluorescence protein (mtRFP) (Fig. 4.5c).
The polarization and partitioning of SLP-2 to the synapse required TCR engagement. In
experiments using planar membranes containing only ICAM-1 without anti-CD3, ICAM-
1 ligated LFA-1 on the Jurkat T cell to induce immunosynapse formation. Under these
conditions, SLP-2 failed to redistribute to the pSMAC and was instead distributed
throughout the membrane (Table 4.2). However, in the presence of both ICAM-1 and
anti-CD3 in the planar membrane, immunosynapse formation was accompanied by
redistribution of SLP-2-gfp to the pSMAC, with 86% of cells showing exclusion of SLP-
2 from the cSMAC (Table 4.1), indicating the requirement of TCR ligation for SLP-2
redistribution. Together, the results presented here confirm the presence of both a
mitochondrial and plasma membrane pool of SLP-2 and demonstrates the redistribution
of both SLP-2 pools to surround TCR microclusters in the pSMAC during T cell
activation. These data support a general model of SLP-2 function involving the
organization of membranes into functional microdomains to support multiple cell
functions and specifically T cell activation, as shown here.
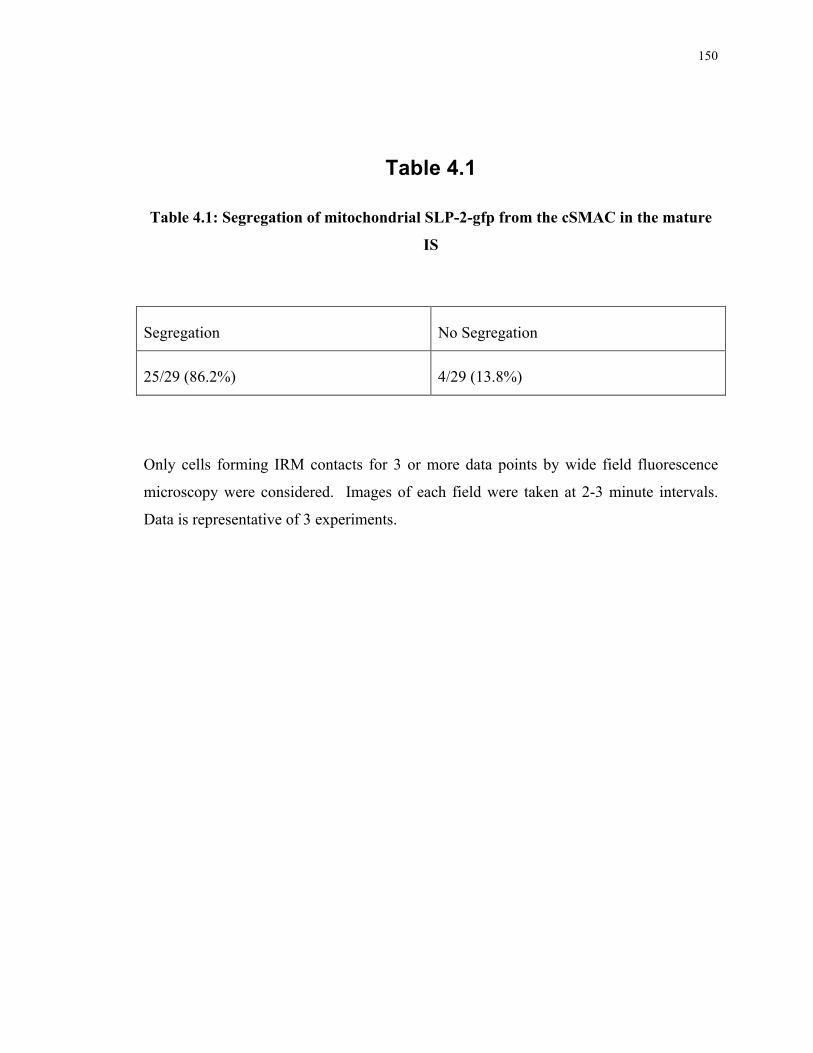
150
Table 4.1
Table 4.1: Segregation of mitochondrial SLP-2-gfp from the cSMAC in the mature
IS
Segregation No Segregation
25/29 (86.2%) 4/29 (13.8%)
Only cells forming IRM contacts for 3 or more data points by wide field fluorescence
microscopy were considered. Images of each field were taken at 2-3 minute intervals.
Data is representative of 3 experiments.

151
Table 4.2
Table 4.2: Percentage of Jurkat T cells on supported planar bilayers organizing
SLP-2-gfp close to pSMAC at the contact interface
Peripheral ring No peripheral ring
10µg/mL OKT3 + 300 mol/µm2 ICAM-1 291 8
300 mol/µm2 ICAM-1 only 0 29
1P<0.0001 vs. ICAM-1 only

152
4.3.6 Mitochondrial recruitment to the IS does not require SLP-2
It has been shown previously that mitochondria relocate to the IS upon T cell stimulation
and that this migration is important for T cell activation 20. We have previously shown
that SLP-2 interacts with the actin cytoskeleton and treatment of Jurkat cells with
cytochalasin D inhibits T cell responses 12. Interestingly, the migration of mitochondria
to the IS during T cell stimulation is dependent on microtubules as well as the actin
cytoskeleton, and inhibition of actin polymerization inhibits mitochondrial movement and
T cell activation 20. To determine if SLP-2 was involved in mitochondrial translocation
to the IS, we imaged mitochondrial movement in response to TCR stimulation in wild
type and SLP-2-deficient primary T cells using confocal microscopy (Fig. 4.6a). For
these experiments, we used cells from conditional SLP-2 knockout mice we have recently
generated (Chapter 3), and stimulated these T cells with anti-CD3 + anti-CD28 antibody
coated beads for 30 minutes at 37°C 15. We found that, in unstimulated cells, the
mitochondrial population was located primarily around the large nucleus (representative
confocal images shown in Fig. 4.6a, quantification of mitochondrial localization shown in
Fig. 4.6b). Upon T cell stimulation, the mitochondrial population re-distributed and was
generally found proximal to the stimulating bead, under the IS (Fig. 4.5c). In addition,
many cells also showed mitochondrial movement to the uropod, classified here as distal
localization (Fig. 4.6a and 4.6c), which is characteristic of migrating lymphocytes 21.
Importantly, there was no apparent defect in mitochondrial migration in T cells lacking
SLP-2, indicating that mitochondrial movement in T cells is independent of SLP-2.

153
Figure 4.6: SLP-2-deficient T cells show normal mitochondrial recruitment upon T
cell stimulation.
T cells were isolated from wild type and SLP-2 T-K/O (T-K/O) mice and stained with
MitoTracker Red. Stained cells were plated on poly-L-lysine coated confocal dishes and
incubated for 10 minutes to promote adherence. Cells were stimulated for 30 minutes
with anti-CD3 and anti-CD28 antibody coated beads or left unstimulated. Cells were
then fixed and imaged by confocal microscopy. These images are representative of at
least 100 individual cells, imaged in three independent experiments. (a) Representative
confocal images are shown for wild type and SLP-2-deficient T cells in the absence (0
min) and presence (30 min) of T cell stimulation (2 different cells for each).
Mitochondria are shown in red and stimulating beads can be seen in the light image
overlay. (b) The location of the mitochondria was quantified in un-stimulated wild type
(black bars) and SLP-2-deficient (white bars) T cells was quantified as either a polar
distribution with mitochondria being clustered together at one end of the cell or a uniform
distribution, with mitochondria located throughout the entire cell. (c) The location of the
mitochondria in stimulated wild type and SLP-2-deficient T cells was quantified as
proximal to the stimulating bead, distal to the bead or uniformly distributed throughout
the cytoplasm, in a manner similar to that in figure 4.4. These plots represent an average
of 3 independent experiments, in which 30 cells for each group were counted.

154
Figure 4.6

155
4.4 Discussion
SLP-2 is a member of the highly conserved stomatin family as well as the SPFH
superfamily, consisting of stomatins, prohibitins, flotillins and HflC/K proteins 9.
Although the function of the stomatin family is currently unknown, these proteins are
found in glycolipid-enriched, detergent-insoluble microdomains and may be involved in
the interaction between protein and lipids in cellular membranes 22-24. Specifically, SLP-
2 is a mostly mitochondrial protein that has been linked to the formation of cardiolipin-
enriched microdomains, thus regulating mitochondrial functions, including energy
production and apoptotic responses 6. SLP-2 has also been identified in detergent-
resistant, glycolipid-enriched microdomains of activated T cells and shown to regulate T
cell activation through the TCR. The mechanism of action by which SLP-2 does it is
unknown 12. The work presented here investigates the link between T cell activation and
the role of SLP-2 in membrane organization. We show that there are two intracellular
pools of SLP-2, one major pool associated to mitochondria and another much less
abundant pool associated to the plasma membrane. Both pools coalesce at the IS upon
TCR ligation, and partition in the peripheral areas of the synapse and are actively
maintained in this location during activation. Together, these results suggest that SLP-2
regulates T cell activation by organizing functional microdomains in mitochondrial and
plasma membranes. These microdomains can then facilitate the optimal assembly and
function of multi-chain complexes or receptors.
The compartmentalization of cellular membranes into specialized microdomains is
thought to facilitate the assembly of multi-molecular complexes. This role has been
shown in the mitochondrial inner membrane for individual complexes of the respiratory
chain and supercomplexes of these individual units, together leading to increased electron
transport efficiency and increased ATP production. This role correlates with SLP-2
directly binding to cardiolipin and interacting with prohibitins 6, 12. Other SPFH family
members, such as prohibitin and flotillin are also proposed to organize membrane
domains 10, 11. Alternatively, SLP-2-deficient T cells have less cardiolipin-enriched
domains and decreased mitochondrial respiration (Chapter 3), and loss of transmembrane

156
potential 5, 25. Altogether, the net effect of the increase in SLP-2 levels upon T cell
activation would be an increase in cardiolipin-enriched microdomains and enhanced
mitochondrial function resulting in increased T cell function.
The data presented here extends the role of SLP-2 as an organizer of membrane
compartmentalization to the plasma membrane. Upon TCR ligation, there is recruitment
of a large number of adapter and signalling molecules to form the TCR signalosome 26.
Assembly of this multi-molecular complex is also associated with re-organization of
plasma membranes, through the aggregation of detergent-resistant, glycolipid-enriched
microdomains previously referred to as lipid rafts 27-30, and the formation of an IS 31.
Within the IS, stable signalling TCR microclusters containing kinases and adapters are
detected in the outer ring of the synapse together with LFA-1 and other adhesion
molecules forming the pSMAC 32. As activation proceeds, TCR microclusters move
from the periphery to the centre of the synapse (cSMACs), destined for internalization
and degradation 33-35, a process that ensures signal down-regulation 36. The results
presented here are compatible with the idea that SLP-2 facilitates the assembly of the
TCR signalosome during T cell activation. In a manner similar to Mec-2 and podocin,
members of the SPFH superfamily, SLP-2 may associate with cholesterol at the plasma
membrane and with components of the TCR signalosome the stability of this
multimolecular complex 7, 12. The ability of SLP-2 to homo-oligomerize may allow SLP-
2 to form a ring-like structure around the signalosome to separate components from other
proteins at the plasma membrane and to prevent dissociation of the complex, in a manner
similar to what SLP-2:prohibitin complexes may do in mitochondria 6, 11.
How SLP-2 specifically interacts with cardiolipin in the mitochondrial membrane and
other lipids in the plasma membrane is an issue for further investigation. SLP-2 lacks a
putative transmembrane domain and palmitoylation sites found in other stomatin family
members 5. We have previously indicated that there may be putative myristoylation sites
at the amino-terminus of SLP-2, which may facilitate membrane association 12.
Alternatively, the SPFH domain may be directly involved in this association as it has
been shown for the superfamily members Mec-2 and podocin and its binding to
cholesterol and for SLP-2 and its binding to cardiolipin 6, 7.

157
Using video capture microscopy we have shown that the mitochondrial-associated pool
of SLP-2 redistributed to the IS upon T cell activation. This result corroborates the
observation of mitochondrial relocation to the IS upon T cell stimulation 20. In this work,
the authors suggest a model in which mitochondria act as a localized calcium sink to
maintain Calcium Release Activated Calcium (CRAC) channel activation, strengthening
T cell signalling and activation. Studies looking at knockdown of SLP-2 with siRNA
have shown that the loss of SLP-2 leads to decreased calcium uptake by the mitochondria 13. Similarly, we have found that mitochondrial calcium uptake is increased in cells over-
expressing full-length SLP-2 (C. Lemke and J. Madrenas, unpublished observations).
However, it is currently unclear how SLP-2 modulates calcium uptake by the
mitochondria. While an inhibitor of mitochondrial sodium/calcium exchange channel
equalized the rate of calcium extrusion from the mitochondria in control and SLP-2-
depleted cells, it did not restore full calcium uptake into the mitochondria 13, indicating
that calcium exchange rates alone cannot account for the role of SLP-2 in mitochondrial
calcium uptake.
Since we had previously shown that SLP-2 interacts with actin and it has been shown that
the mitochondrial translocation to the IS requires actin polymerization 12, 20, it was
plausible that SLP-2 may link the mitochondria to the actin cytoskeleton and facilitate the
movement of mitochondria to the IS. However, our data indicate that this is not the case
because we found that mitochondrial translocation to the IS proceeded normally in SLP-
2-deficient T cells. Thus, SLP-2 is not required for mitochondrial translocation to the IS
during T cell activation.
The identification of SLP-2 and other mitochondrial proteins at the plasma membrane
may reflect membrane exchange between these organelles and other cellular
compartments and organelles, and point to a novel pathway of intracellular trafficking.
We have confirmed the presence of a mitochondrial targeting sequence in the amino-
terminus of SLP-2. This raised the question of the intracellular trafficking pathway
leading to the presence of SLP-2 in the cell surface. It is possible that the plasma
membrane pool of SLP-2 arises by default when a small proportion of newly synthesized
SLP-2 fails to be transported to the mitochondria. However, it is also possible that a

158
small amount of membrane exchange occurs between the mitochondrial and plasma
membranes, allowing for the exchange of membrane-associated proteins, which is
supported by the existence of mitochondria-derived vesicles and its fusion with other
organelles such as lysosomes 37-40. In addition to SLP-2, a small surface pool has also
been shown for other mitochondrial proteins such as prohibitin, porin, NADH
dehydrogenase, ubiquinol-cytochrome c reducatase and ATP synthase, indicating that the
surface association of mitochondrial proteins is not unique to SLP-2 41-44. The purpose of
mitochondrial proteins at the plasma membrane is unclear, although the association of
SLP-2 with components of the T cell signalosome suggest that both pools of SLP-2 are
likely functional.
In summary, the work presented here provides the first evidence of two pools of SLP-2 in
human and mouse T cells and it suggests to a conservation of function at both
membranes. In the mitochondria, SLP-2 organizes the membrane into cardiolipin-
enriched domains to facilitate respiratory chain function. At the plasma membrane, SLP-
2 may facilitate immunosynapse organization, leading to increased T cell signalling and
activation. It is likely that both of these functions have the same mechanistic basis, i.e.
organization of specialized microdomains that facilitate the assembly of multi-molecular
complexes and ultimately ensure their optimal function.

159
4.5 References
1. Da Cruz, S., Xenarios, I., Langridge, J., Vilbois, F., Parone, P.A. & Martinou, J.C. Proteomic analysis of the mouse liver mitochondrial inner membrane. J Biol Chem 278, 41566-41571 (2003).
2. Chevallet, M., Lescuyer, P., Diemer, H., van Dorsselaer, A., Leize-Wagner, E. & Rabilloud, T. Alterations of the mitochondrial proteome caused by the absence of mitochondrial DNA: A proteomic view. Electrophoresis 27, 1574-1583 (2006).
3. Rezaul, K., Wu, L., Mayya, V., Hwang, S.I. & Han, D. A systematic characterization of mitochondrial proteome from human T leukemia cells. Mol Cell Proteomics 4, 169-181 (2005).
4. Taylor, S.W., Fahy, E., Zhang, B., Glenn, G.M., Warnock, D.E., Wiley, S., Murphy, A.N., Gaucher, S.P., Capaldi, R.A., Gibson, B.W. & Ghosh, S.S. Characterization of the human heart mitochondrial proteome. Nat Biotechnol 21, 281-286 (2003).
5. Hajek, P., Chomyn, A. & Attardi, G. Identification of a novel mitochondrial complex containing mitofusin 2 and stomatin-like protein 2. J Biol Chem 282, 5670-5681 (2007).
6. Christie, D.A., Lemke, C.D., Elias, I.M., Chau, L.A., Kirchhof, M.G., Li, B., Ball, E.H., Dunn, S.D., Hatch, G.M. & Madrenas, J. Stomatin-like protein 2 binds cardiolipin and regulates mitochondrial biogenesis and function. Mol Cell Biol 31, 3845-3856 (2011).
7. Huber, T.B., Schermer, B., Muller, R.U., Hohne, M., Bartram, M., Calixto, A., Hagmann, H., Reinhardt, C., Koos, F., Kunzelmann, K., Shirokova, E., Krautwurst, D., Harteneck, C., Simons, M., Pavenstadt, H., Kerjaschki, D., Thiele, C., Walz, G., Chalfie, M. & Benzing, T. Podocin and MEC-2 bind cholesterol to regulate the activity of associated ion channels. Proc Natl Acad Sci U S A 103, 17079-17086 (2006).
8. B, W.A., G, I.H., L, M.I., M, L.J., V, P.M., H, I.N., V, A.M., P, J.S., H, M.T. & A, I.Z. [Part VI. Antiparasitic treatment for Chagas disease]. Rev Chilena Infectol 25, 384-389, discussion 387 (2008).
9. Browman, D.T., Hoegg, M.B. & Robbins, S.M. The SPFH domain-containing proteins: more than lipid raft markers. Trends Cell Biol 17, 394-402 (2007).
10. Morrow, I.C. & Parton, R.G. Flotillins and the PHB domain protein family: rafts, worms and anaesthetics. Traffic 6, 725-740 (2005).

160
11. Osman, C., Merkwirth, C. & Langer, T. Prohibitins and the functional compartmentalization of mitochondrial membranes. J Cell Sci 122, 3823-3830 (2009).
12. Kirchhof, M.G., Chau, L.A., Lemke, C.D., Vardhana, S., Darlington, P.J., Marquez, M.E., Taylor, R., Rizkalla, K., Blanca, I., Dustin, M.L. & Madrenas, J. Modulation of T cell activation by stomatin-like protein 2. J Immunol 181, 1927-1936 (2008).
13. Da Cruz, S., De Marchi, U., Frieden, M., Parone, P.A., Martinou, J.C. & Demaurex, N. SLP-2 negatively modulates mitochondrial sodium-calcium exchange. Cell Calcium 47, 11-18 (2009).
14. Tondera, D., Grandemange, S., Jourdain, A., Karbowski, M., Mattenberger, Y., Herzig, S., Da Cruz, S., Clerc, P., Raschke, I., Merkwirth, C., Ehses, S., Krause, F., Chan, D.C., Alexander, C., Bauer, C., Youle, R., Langer, T. & Martinou, J.C. SLP-2 is required for stress-induced mitochondrial hyperfusion. EMBO J 28, 1589-1600 (2009).
15. Arp, J., Kirchhof, M.G., Baroja, M.L., Nazarian, S.H., Chau, T.A., Strathdee, C.A., Ball, E.H. & Madrenas, J. Regulation of T-cell activation by phosphodiesterase 4B2 requires its dynamic redistribution during immunological synapse formation. Mol Cell Biol 23, 8042-8057 (2003).
16. Baroja, M.L., Cieslinski, L.B., Torphy, T.J., Wange, R.L. & Madrenas, J. Specific CD3 epsilon association of a phosphodiesterase 4B isoform determines its selective tyrosine phosphorylation after CD3 ligation. J Immunol 162, 2016-2023 (1999).
17. Lee, P.P., Fitzpatrick, D.R., Beard, C., Jessup, H.K., Lehar, S., Makar, K.W., Perez-Melgosa, M., Sweetser, M.T., Schlissel, M.S., Nguyen, S., Cherry, S.R., Tsai, J.H., Tucker, S.M., Weaver, W.M., Kelso, A., Jaenisch, R. & Wilson, C.B. A critical role for Dnmt1 and DNA methylation in T cell development, function, and survival. Immunity 15, 763-774 (2001).
18. Darlington, P.J., Baroja, M.L., Chau, T.A., Siu, E., Ling, V., Carreno, B.M. & Madrenas, J. Surface cytotoxic T lymphocyte-associated antigen 4 partitions within lipid rafts and relocates to the immunological synapse under conditions of inhibition of T cell activation. The Journal of experimental medicine 195, 1337-1347 (2002).
19. Tomas, E.M., Chau, T.A. & Madrenas, J. Clustering of a lipid-raft associated pool of ERM proteins at the immunological synapse upon T cell receptor or CD28 ligation. Immunol Lett 83, 143-147 (2002).
20. Quintana, A., Schwindling, C., Wenning, A.S., Becherer, U., Rettig, J., Schwarz, E.C. & Hoth, M. T cell activation requires mitochondrial translocation to the immunological synapse. Proc Natl Acad Sci U S A 104, 14418-14423 (2007).

161
21. Campello, S., Lacalle, R.A., Bettella, M., Manes, S., Scorrano, L. & Viola, A. Orchestration of lymphocyte chemotaxis by mitochondrial dynamics. J Exp Med 203, 2879-2886 (2006).
22. Snyers, L., Umlauf, E. & Prohaska, R. Association of stomatin with lipid-protein complexes in the plasma membrane and the endocytic compartment. Eur J Cell Biol 78, 802-812 (1999).
23. Snyers, L., Thines-Sempoux, D. & Prohaska, R. Colocalization of stomatin (band 7.2b) and actin microfilaments in UAC epithelial cells. Eur J Cell Biol 73, 281-285 (1997).
24. Mairhofer, M., Steiner, M., Salzer, U. & Prohaska, R. Stomatin-like protein-1 interacts with stomatin and is targeted to late endosomes. J Biol Chem 284, 29218-29229 (2009).
25. Wang, Y., Cao, W., Yu, Z. & Liu, Z. Downregulation of a mitochondria associated protein SLP-2 inhibits tumor cell motility, proliferation and enhances cell sensitivity to chemotherapeutic reagents. Cancer Biol Ther 8, 1651-1658 (2009).
26. Smith-Garvin, J.E., Koretzky, G.A. & Jordan, M.S. T cell activation. Annu Rev Immunol 27, 591-619 (2009).
27. Xavier, R., Brennan, T., Li, Q., McCormack, C. & Seed, B. Membrane compartmentation is required for efficient T cell activation. Immunity 8, 723-732 (1998).
28. Janes, P.W., Ley, S.C. & Magee, A.I. Aggregation of lipid rafts accompanies signaling via the T cell antigen receptor. J Cell Biol 147, 447-461 (1999).
29. Janes, P.W., Ley, S.C., Magee, A.I. & Kabouridis, P.S. The role of lipid rafts in T cell antigen receptor (TCR) signalling. Semin Immunol 12, 23-34 (2000).
30. Viola, A., Schroeder, S., Sakakibara, Y. & Lanzavecchia, A. T lymphocyte costimulation mediated by reorganization of membrane microdomains. Science 283, 680-682 (1999).
31. Fooksman, D.R., Vardhana, S., Vasiliver-Shamis, G., Liese, J., Blair, D.A., Waite, J., Sacristan, C., Victora, G.D., Zanin-Zhorov, A. & Dustin, M.L. Functional anatomy of T cell activation and synapse formation. Annu Rev Immunol 28, 79-105 (2010).
32. Gascoigne, N.R. & Zal, T. Molecular interactions at the T cell-antigen-presenting cell interface. Curr Opin Immunol 16, 114-119 (2004).
33. Lee, K.H., Dinner, A.R., Tu, C., Campi, G., Raychaudhuri, S., Varma, R., Sims, T.N., Burack, W.R., Wu, H., Wang, J., Kanagawa, O., Markiewicz, M., Allen,

162
P.M., Dustin, M.L., Chakraborty, A.K. & Shaw, A.S. The immunological synapse balances T cell receptor signaling and degradation. Science 302, 1218-1222 (2003).
34. Lee, K.H., Holdorf, A.D., Dustin, M.L., Chan, A.C., Allen, P.M. & Shaw, A.S. T cell receptor signaling precedes immunological synapse formation. Science 295, 1539-1542 (2002).
35. Varma, R., Campi, G., Yokosuka, T., Saito, T. & Dustin, M.L. T cell receptor-proximal signals are sustained in peripheral microclusters and terminated in the central supramolecular activation cluster. Immunity 25, 117-127 (2006).
36. Mossman, K.D., Campi, G., Groves, J.T. & Dustin, M.L. Altered TCR signaling from geometrically repatterned immunological synapses. Science 310, 1191-1193 (2005).
37. Braschi, E., Goyon, V., Zunino, R., Mohanty, A., Xu, L. & McBride, H.M. Vps35 mediates vesicle transport between the mitochondria and peroxisomes. Curr Biol 20, 1310-1315 (2010).
38. Neuspiel, M., Schauss, A.C., Braschi, E., Zunino, R., Rippstein, P., Rachubinski, R.A., Andrade-Navarro, M.A. & McBride, H.M. Cargo-selected transport from the mitochondria to peroxisomes is mediated by vesicular carriers. Curr Biol 18, 102-108 (2008).
39. Soubannier, V., McLelland, G.L., Zunino, R., Braschi, E., Rippstein, P., Fon, E.A. & McBride, H.M. A Vesicular Transport Pathway Shuttles Cargo from Mitochondria to Lysosomes. Curr Biol (2012).
40. Andrade-Navarro, M.A., Sanchez-Pulido, L. & McBride, H.M. Mitochondrial vesicles: an ancient process providing new links to peroxisomes. Curr Opin Cell Biol 21, 560-567 (2009).
41. Bathori, G., Parolini, I., Tombola, F., Szabo, I., Messina, A., Oliva, M., De Pinto, V., Lisanti, M., Sargiacomo, M. & Zoratti, M. Porin is present in the plasma membrane where it is concentrated in caveolae and caveolae-related domains. The Journal of biological chemistry 274, 29607-29612 (1999).
42. Bae, T.J., Kim, M.S., Kim, J.W., Kim, B.W., Choo, H.J., Lee, J.W., Kim, K.B., Lee, C.S., Kim, J.H., Chang, S.Y., Kang, C.Y., Lee, S.W. & Ko, Y.G. Lipid raft proteome reveals ATP synthase complex in the cell surface. Proteomics 4, 3536-3548 (2004).
43. Kim, K.B., Lee, J.W., Lee, C.S., Kim, B.W., Choo, H.J., Jung, S.Y., Chi, S.G., Yoon, Y.S., Yoon, G. & Ko, Y.G. Oxidation-reduction respiratory chains and ATP synthase complex are localized in detergent-resistant lipid rafts. Proteomics 6, 2444-2453 (2006).

163
44. Kim, B.W., Lee, C.S., Yi, J.S., Lee, J.H., Lee, J.W., Choo, H.J., Jung, S.Y., Kim, M.S., Lee, S.W., Lee, M.S., Yoon, G. & Ko, Y.G. Lipid raft proteome reveals that oxidative phosphorylation system is associated with the plasma membrane. Expert Rev Proteomics 7, 849-866 (2010).

164
Chapter 5
5 Discussion
5.1 Summary of results
5.1.1 Stomatin-like protein 2 binds cardiolipin and regulates mitochondrial biogenesis and function
To investigate the role of SLP-2 in T cells, a doxycycline-inducible SLP-2 construct
tagged with GFP at the C-terminus was stably transfected into Jurkat T cells, a human T
cell line. Sub-cellular fractionation demonstrated that SLP-2 was localized to the
mitochondria and this was confirmed by confocal imaging showing co-localization of
SLP-2-GFP with the mitochondrial-specific dye MitoTracker Red. SLP-2-GFP over-
expressing cells had an increase in mitochondrial mass and an increase in the number of
mitochondria per cell slice in transmission electron microscopy images. This was
confirmed in primary human peripheral blood mononuclear cells (PBMCs), in which
ionomycin and PMA treatment led to increased SLP-2 expression and increased
mitochondrial mass.
To investigate the mechanism of increased mitochondrial populations upon SLP-2 over-
expression, the cellular programs required for mitochondrial biogenesis, including lipid
synthesis, nuclear transcription and mitochondrial DNA replication, were examined. In
both SLP-2 over-expressing Jurkat T cells and stimulated PBMCs, the level of the
mitochondrial-specific lipid cardiolipin was increased. An increase in radiolabelled
precursor incorporation demonstrated that the increase in cardiolipin was due to increased
biosynthesis. Supporting this finding, expression of enzymes required for the final two
steps of cardiolipin biosynthesis were also elevated in SLP-2 over-expressing cells. This
increased cardiolipin biosynthesis was required for mitochondrial biogenesis as inhibition
of cardiolipin synthase blocked mitochondrial biogenesis in response to T cell
stimulation. In addition, SLP-2 over-expressing cells also had increased expression
levels of PGC-1α, a transcriptional co-activator and master regulator of nuclear

165
transcription programs for mitochondrial targeted genes. Finally, SLP-2 over-expressing
cells showed increased levels of mitochondrial DNA, which is essential for functional
mitochondrial populations. The timeline of mitochondrial biogenesis upon T cell
stimulation demonstrated that expression of SLP-2 and prohibitins increased at early time
points, followed by an increase in PGC-1α expression. These events preceded increased
mitochondrial mass, which was seen by 24 hours post-stimulation.
As a possible mechanism for the increase in mitochondrial biogenesis upon SLP-2 over-
expression, the interaction of SLP-2 with the prohibitin complex was examined, as the
functional interactome of prohibitins has been shown to contain components of the
cardiolipin biosynthetic machinery 1. SLP-2 was found to associate with both PHB-1 and
PHB-2 and it was hypothesized that SLP-2 may recruit the prohibitin complex to the
mitochondrial membrane. Indeed, upon SLP-2 over-expression, there was a specific
increase in the association of PHB-1 with the mitochondrial membrane, while total
cellular expression levels of PHB-1 remained unchanged. Furthermore, it was found that
recombinant human SLP-2 bound to cardiolipin-enriched vesicles in a titratable fashion,
in which vesicles with higher cardiolipin content bound more SLP-2. Based on these
findings, it is proposed that SLP-2 may act as a bridge between cardiolipin in the
mitochondrial membrane and the prohibitin complex, recruiting prohibitins to facilitate
mitochondrial membrane compartmentalization.
The major function associated with the mitochondria is the energy production capacity of
the respiratory chain. As such, the effect of increased mitochondrial biogenesis upon
SLP-2 over-expression on respiratory chain function was examined. As expected, these
cells had higher enzymatic activity for both complex I and complex II as well as
increased ATP levels and oxygen consumption. This increased respiratory chain function
was due to an increase in activity but not expression levels of the respiratory chain
subunits. Finally, SLP-2 over-expressing cells were also less susceptible to the induction
of the intrinsic apoptotic pathway. Together, these results demonstrate that over-
expression of SLP-2 increases mitochondrial biogenesis, energy production and apoptosis
resistance, likely through the formation of cardiolipin-enriched microdomains.

166
5.1.2 SLP-2 deficiency in T cells is associated with abnormal mitochondrial membrane compartmentalization of cardiolipin, decreased mitochondrial respiration and impaired CD4+ T cell responses
To further investigate the role of SLP-2 in T cell activation, SLP-2 knockout mice were
generated and found to be embryonic lethal at the pre-implantation stage. To circumvent
this problem, conditional knockout mice were generated using the Cre-lox system
wherein SLP-2 was flanked by lox sites and bred with mice expressing Cre recombinase
under the control of the CD4 promoter. This resulted in T cell-specific conditional SLP-2
deletion at the double positive stage of T cell development with total loss of SLP-2
expression in mature T cells.
T cell-specific SLP-2 conditional knockout mice (SLP-2 T-K/O) showed normal T cell
development, with cell numbers in thymus, spleen and lymph nodes comparable to those
found in control mice. In addition, thymic subsets were also similar between control and
SLP-2 T-K/O mice, including double negative, double positive and CD4+ or CD8+ single
positive populations. As well, CD3+, CD4+, CD8+ and non-T cell populations were
similar between control and SLP-2 T-K/O mice in both the spleen and lymph nodes.
These results indicate that T cell development proceeds independently of SLP-2
expression.
It has been shown previously that SLP-2 expression levels correlate with T cell activation 2. Similarly, T cells from SLP-2 T-K/O mice had decreased activation in response to ex
vivo stimulation. To demonstrate the importance of this result in vivo, T cell recall
responses to primary immunization were examined in SLP-2 T-K/O mice. As expected
from ex vivo data, SLP-2 T-K/O mice showed significantly lower activation during
secondary exposure to antigen. As an additional in vivo model of SLP-2 function, a
transplant model was examined and control mice were found to reject non-matched
allografts an average of two days earlier than SLP-2 T-K/O mice, indicating that
decreased T cell activation upon SLP-2 deletion delays allograft rejection. This decrease

167
in T cell activation in SLP-2 T-K/O mice was shown to be a post-transcriptional block in
IL-2 production.
Examination of the effect of SLP-2 deletion on mitochondrial function showed that SLP-
2 T-K/O T cells had decreased protein levels of multiple subunits of complex I of the
respiratory chain while the remaining complexes were found in normal levels.
Furthermore, SLP-2 T-K/O T cells also had decreased activity of complex I as well as
complex II + III. Investigation of cardiolipin indicated that while SLP-2 T-K/O T cells
had normal total cellular levels of cardiolipin, there were decreased levels of cardiolipin
in detergent-insoluble microdomains and less recruitment of prohibitins to these domains.
To examine the effect of glucose starvation on SLP-2 T-K/O cells, T cells were
stimulated under conditions of excess or limiting glucose and responses were measured.
Under both excess and limiting glucose, SLP-2 T-K/O T cells had significantly lower
activation than control cells. Interestingly, under conditions of limiting glucose, control
cells had partial inhibition of T cell activation while SLP-2 T-K/O T cells were activated
to a similar extent under both glucose conditions. These results further demonstrate the
altered metabolic capacity of T cells lacking SLP-2 and point to the importance of
membrane organization for proper cellular functions.
5.1.3 Plasma membrane and mitochondrial stomatin-like protein 2 coalesce at the immunological synapse during T cell activation
To further investigate the subcellular localization and movement of SLP-2 in T cells,
SLP-2-GFP expressing Jurkat T cells were utilized. In addition, a Jurkat T cell line
stably expressing a doxycycline-inducible truncation mutant of SLP-2, lacking the amino
terminus and tagged at the carboxy terminus with GFP (ΔN-SLP-2-GFP) was also
generated. In comparison with full length SLP-2-GFP, which showed co-localization
with MitoTracker Red, ΔN-SLP-2-GFP showed diffuse cytosolic staining and no longer
co-localized with Mitotracker Red, confirming the presence of a mitochondrial targeting
domain at the amino-terminus of SLP-2 3. This divergent localization of full length and

168
truncated SLP-2 was confirmed by subcellular fractionation, in which SLP-2-GFP was
found in the mitochondrial fraction while ΔN-SLP-2-GFP was found only in the cytosol.
Surprisingly, a small pool of SLP-2 was also found in association with the plasma
membrane. While both SLP-2-GFP and ΔN-SLP-2-GFP were shown to associate with
the plasma membrane, full length SLP-2-GFP retained plasma membrane association at
lower expression levels compared to ΔN-SLP-2-GFP. This result indicates that while the
amino terminal region may play a role in membrane association, it is not essential.
It has been shown that mitochondria translocate to the immunological synapse upon T
cell activation and this translocation is required for optimal activation 4. In light of this
and given that SLP-2 is predominantly mitochondrial, the location of SLP-2-GFP during
T cell activation by superantigen stimulation was examined. In resting Jurkat T cells,
SLP-2-GFP distributed uniformly throughout the cell, surrounding the large nucleus.
Upon T cell activation, SLP-2-GFP re-localized to the immunological synapse and this
localization was actively maintained during T cell activation. Since the actin
cytoskeleton is required for mitochondrial translocation and SLP-2 has been shown
previously to associate with actin 2, 4, mitochondrial translocation in cells lacking SLP-2
was examined. The mitochondria in SLP-2 T-K/O T cells were found to redistribute to
the immunological synapse in a manner similar to that in wild type T cells, indicating that
mitochondrial trafficking during T cell activation proceeds independently of SLP-2.
To further investigate the surface pool of SLP-2, total internal reflection fluorescence
microscopy (TIRFM) was used to image 200 nm from the support plain, eliminating in
this way the mitochondrial pool of SLP-2. With this technique, the surface pool of SLP-2
was shown to be preferentially excluded from the cSMAC during T cell activation and
was found to surround TCR microclusters in the pSMAC. SLP-2 has been shown to
homo-oligomerize and it is possible that SLP-2 may form a large complex surrounding
the T cell signalosome, leading to increased association and activation of T cell signal
complexes. Furthermore, the presence of a small pool of the mitochondrial resident
protein SLP-2 at the plasma membrane may be indicative of membrane exchange
between the mitochondria and the cell surface, representing a novel mechanism for
protein trafficking in the cell.

169
5.2 Model of function: SLP-2 organizes membrane domains
SLP-2 is a member of the highly conserved stomatin family and therefore of the SPFH
superfamily, consisting of stomatins, prohibitins, flotillins and bacterial HflC/K proteins 5-7. This family of proteins is defined by the shared SPFH domain, a globular domain
composed of both a hydrophobic region and a hydrophilic region containing conserved
charged residues on the surface of the domain 8. These proteins are found in association
with either the plasma membrane or various intracellular membranes, either as integral or
peripheral membrane proteins, many of which are proposed to associate through
palmitoylation modifications 5, 9. The function of the SPFH domain is currently unclear,
although it has been linked to protein-lipid interactions such as those seen in association
with the plasma membrane 10. It is likely that a highly conserved domain such as the
SPFH domain would have a conservation of function. Indeed, studies of SPFH family
members have pointed to a general mechanism of action involving the organization of
membranes into defined domains to affect a wide variety of cellular functions depending
on the subcellular localization of these proteins 11-15. Plasma membrane-associated
flotillins have been shown to play a role in insulin signalling, phagocytosis, axon growth
and lymphocyte activation, while mitochondrial-associated prohibitins are involved in
cellular lifespan, mitochondrial morphology and assembly of the respiratory chain and
plasma membrane-associated stomatin has been proposed to be involved in ion channel
regulation 13, 16, 17. Based on the conserved mechanism of action across SPFH family
members and on the results presented here, we propose that SLP-2 organizes membranes
into specialized microdomains, thus affecting many mitochondrial and cellular functions.
The organization of the plasma membrane into specialized domains has long been
proposed to play an important role in various cellular processes, including signal
transduction and membrane trafficking 18. These domains, previously termed lipid rafts
but referred to here as detergent-insoluble glycolipid-enriched microdomains (DI-GEMs),
are highly ordered membrane domains enriched in sphingolipids, cholesterol and GPI-
anchored proteins 18-20. DI-GEMs are thought to exist in resting cells as nanoscale
assemblies, which coalesce in response to stimulation 18, 19. The coalescence of the

170
smaller assemblies into larger domains has been proposed to occur through lipid-lipid,
lipid-protein and protein-protein interactions. Although most studies of DI-GEMs
involve the plasma membrane, SLP-2 is associated predominantly with the mitochondrial
membrane, where there is a very limited level of cholesterol, the major component of
plasma membrane DI-GEMs. However, recent studies have indicated the possibility of
DI-GEM regions in the mitochondrial membrane 21-25. These studies have pointed to a
role for the mitochondrial-specific lipid cardiolipin in the formation of specialized
cardiolipin-enriched microdomains involved in induction of apoptosis, mitochondrial
fission and cristae formation 23, 25, 26. Our finding that SLP-2 binds cardiolipin may reveal
a key step in the formation of these microdomains in the mitochondria. Cardiolipin is a
lipid of unique structure that contains four acyl chains and has been found to bind directly
to a number of proteins, and this association is required for proper function of
components of the respiratory chain as well as the protein import machinery 27-29. As
such, the formation of cardiolipin-enriched microdomains not only segregates physically
mitochondrial proteins but also plays an important role in proper protein function.
Although previous work has pointed to the existence of cardiolipin-enriched
microdomains in the mitochondrial membrane, it is unclear how these regions arise. We
propose a model in which SLP-2 contributes to the formation of cardiolipin-enriched
microdomains through a direct association with cardiolipin and through interactions with
various proteins. We have presented here a number of findings to support a role for SLP-
2 in membrane organization. Similar to other SPFH family members, SLP-2 has been
found in DI-GEM fractions, specifically in T cells undergoing activation and as T cell
activation proceeds, SLP-2 is increasingly recruited to DI-GEM fractions, supporting a
role in membrane organization 2. In addition, we have found that recombinant SLP-2
binds directly to cardiolipin, in such a way that increased concentrations of cardiolipin
leads to increased SLP-2 binding. This is reminiscent of work showing the plasma
membrane-associated SPFH domain containing proteins Mec-2 and podocin binding
directly to cholesterol 10. Furthermore, we have found that SLP-2 alters the recruitment
of proteins to DI-GEM regions, as the over-expression of SLP-2 leads to an increased
recruitment of prohibitins to the mitochondrial membrane. This is similar to results
looking at stomatin-like protein 1 and podocin, both of which have been shown to control

171
localization of other proteins 15, 30, 31. Finally, the formation of DI-GEMs is altered in the
absence of SLP-2, as these fractions have significantly less cardiolipin content in mice
lacking SLP-2.
We also propose that the oligomerization of SLP-2 may play a role in membrane
organization. Similar to other SFPH family members, we have shown here that SLP-2 is
capable of undergoing homo-oligomerization 10, 32, 33. Oligomerization of SPFH proteins
has been proposed to play a role in protein stabilization, as loss of either prohibitin 1 or
prohibitin 2 prevents the formation of the large heteroligomeric prohibitin complex and
results in the loss of both prohibitin proteins 5, 34. However, we found that while the
truncation mutant of SLP-2 lacking the amino terminal region failed to oligomerize with
full length endogenous SLP-2, its expression was not lost. This indicates that
oligomerization of SLP-2 is not required for protein stability and instead we propose that
the oligomerization of SLP-2 into large complexes may facilitate the concentration of
cardiolipin into cardiolipin-enriched microdomains. Although cardiolipin only
constitutes approximately 10-15% of the total lipid content of the mitochondrial
membrane 14, the oligomerization of cardiolipin-bound SLP-2 proteins could alter the
cardiolipin distribution in the mitochondrial membrane and concentrate it into regions in
which the localized cardiolipin content is much higher than 15%. Furthermore, those
proteins bound to cardiolipin or with SLP-2, either directly or indirectly, would also be
concentrated into these regions, leading to the formation of large protein complexes (Fig.
5.1). The proteins recruited to these regions would in turn determine the function of the
domain. In this way, treatment with the inducer of apoptosis, anti-Fas could trigger the
formation of domains enriched in pro-apoptotic factors and fission proteins 23, 35.

172
Figure 5.1: A step-wise model of the formation of cardiolipin-enriched
microdomains in the mitochondrial membrane as facilitated by SLP-2.
(a) The mitochondrial membrane is composed of a mixture of lipids, including the
mitochondrial-specific lipid cardiolipin, which compromises approximately 10-15% of
the total mitochondrial lipids 36. (b) SLP-2 binds directly to cardiolipin and undergoes
homo-oligomerization (c) to concentrate cardiolipin into cardiolipin-enriched
microdomains. (d) Additional proteins are recruited to these domains through direct or
indirect association with either cardiolipin (respiratory chains components) or SLP-2
(prohibitin complex). Abreviations: CL – cardiolipin, SLP-2 – Stomatin-like protein 2,
PHB – prohibitin, C – complex of the respiratory chain, CI – NADH dehydrogenase, CII
– succinate dehydrogenase, CIII – cytochrome bc1 complex, CIV – cytochrome c
oxidase, CV – ATP synthase.
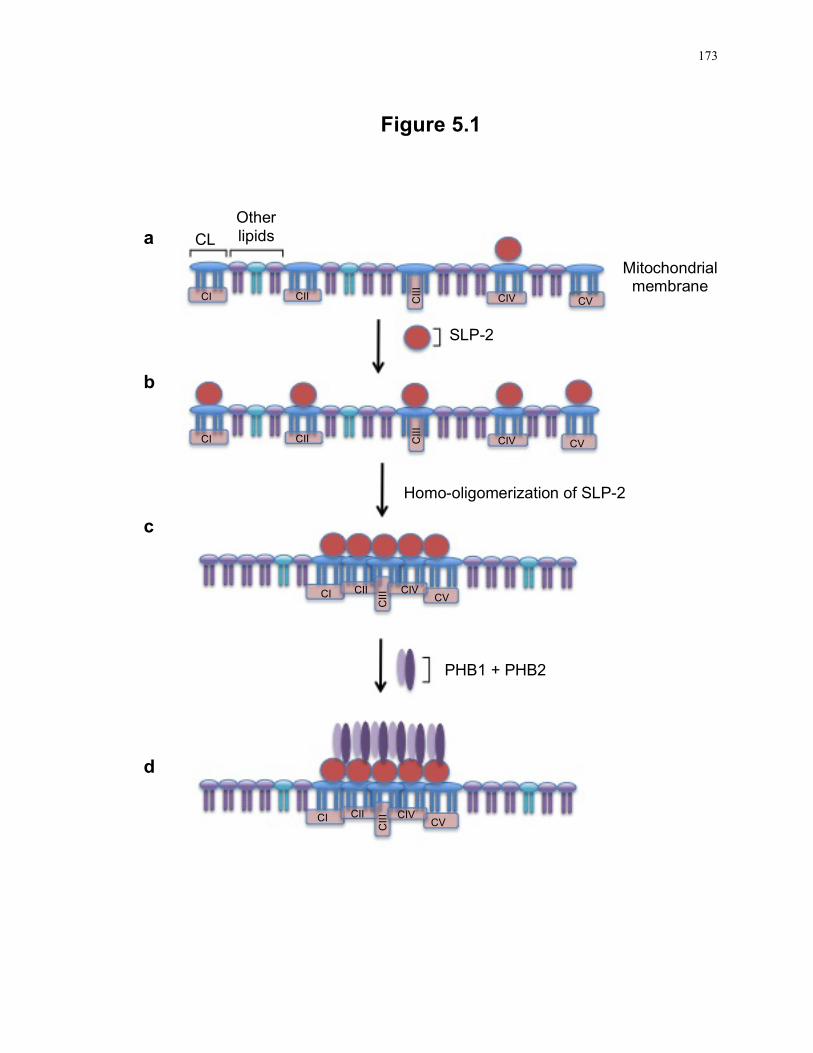
173
Figure 5.1
CL Other lipids
SLP-2
Mitochondrial membrane
Homo-oligomerization of SLP-2
PHB1 + PHB2
CI CII CII
I
CIV CV
CI CII CII
I
CIV CV
CI CII
CII
I CIV CV
CI CII
CII
I CIV CV
a
b
c
d

174
Figure 5.2: Model of supercomplex formation upon SLP-2 upregulation in response
to T cell activation.
SLP-2 is expressed at low levels in naive T cells but both SLP-2 and prohibitin 1 and 2
are upregulated upon T cell activation. Through self-oligomerization, prohibitin
association and the interaction with cardiolipin, SLP-2 organizes the mitochondrial
membrane into cardiolipin-enriched domains, which facilitates supercomplex formation,
leading to more efficient respiratory chain function and increased mitochondrial
biogenesis. Abreviations: CL – cardiolipin, SLP-2 – Stomatin-like protein 2, PHB –
prohibitin, C – complex of the respiratory chain, CI – NADH dehydrogenase, CII –
succinate dehydrogenase, CIII – cytochrome bc1 complex, CIV – cytochrome c oxidase,
CV – ATP synthase.
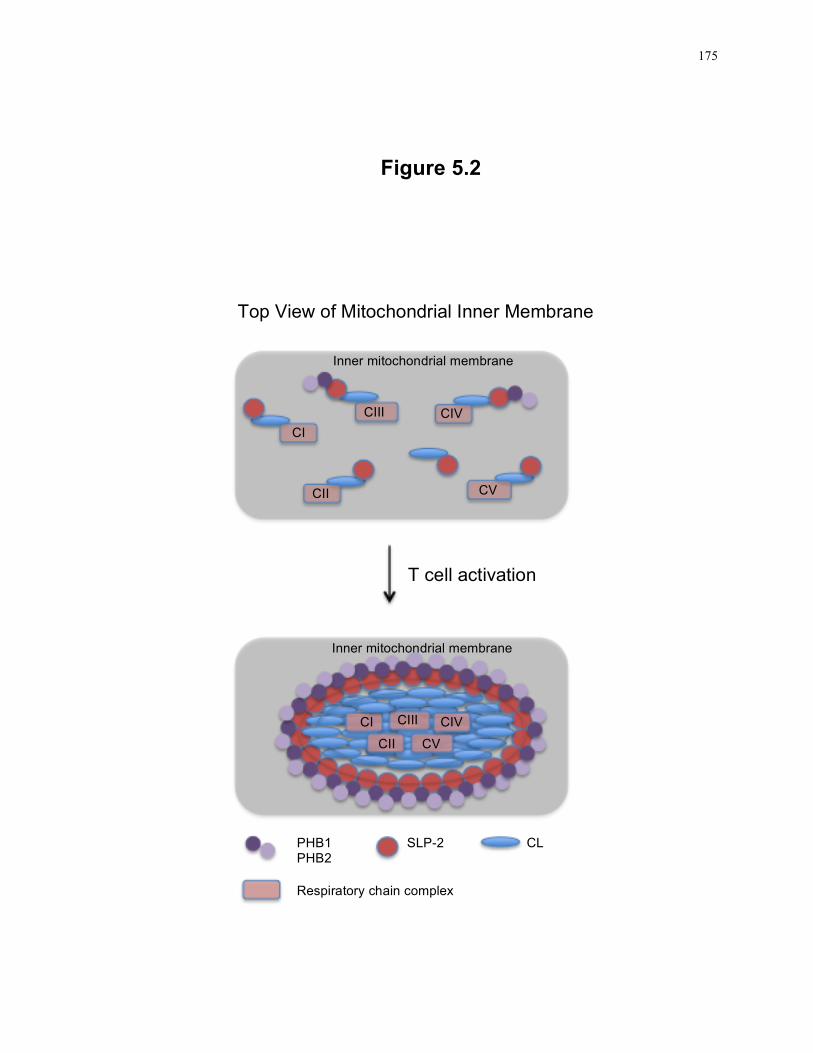
175
Figure 5.2
Top View of Mitochondrial Inner Membrane
Inner mitochondrial membrane
T cell activation
Inner mitochondrial membrane
CI
CIII
CII
CIV
CV
CI CIII
CII
CIV
CV
PHB1 PHB2
SLP-2 CL
Respiratory chain complex

176
5.2.1 Assembly of multi-domain receptors, including respiratory chain supercomplex formation
A major function of the mitochondria is the energy production capacity of the respiratory
chain. Given that the respiratory chain is composed of a large number of subunits
organized into five separate complexes, it is attractive to speculate a role for cardiolipin-
enriched domains to enhance the efficiency of the respiratory chain. This is supported by
the detection of respiratory chain components in the cardiolipin-enriched DI-GEMs of
mouse T cells. As electrons are transported through the respiratory chain, closer packing
of the complexes in the mitochondrial membrane would facilitate more efficient transfer
of electrons and reduce the possibility of energy loss. Indeed, it has been shown that
multiple components of the respiratory chain, including complexes I, III and IV are found
in association with each other, leading to the formation of supercomplexes 37. Given that
cardiolipin is known to be required for proper functioning of various components of the
respiratory chain and that supercomplex formation is inhibited in cells lacking cardiolipin 27, 28, 38-40, we propose that supercomplexes may be formed in cardiolipin-enriched
domains organized by SLP-2 and cardiolipin interactions (Fig. 5.2). In the absence of
SLP-2, cardiolipin-enriched DI-GEM domains may be less stable, causing supercomplex
dissociation and decreased respiratory chain efficiency. Supporting this model, studies
have shown a small decrease in transmembrane potential upon SLP-2 knockdown and we
have found a decrease in cardiolipin content in DI-GEMs along with decreased complex I
activity in SLP-2-deficient T cells 41, 42. As complex I is the largest of the respiratory
chain complexes, consisting of 45 subunits, and cardiolipin is known to be required for
complex I activity, it is likely that cardiolipin-enriched DI-GEMs play a role in the
assembly and/or the activity of complex I 29, 43. In addition, we and others have shown an
association between SLP-2 and the prohibitin complex and we have found increased
recruitment of prohibitins upon SLP-2 over-expression 3. This recruitment of the
prohibitin complex to cardiolipin-enriched microdomains may also play a role in the
assembly of complex I, as suggested by studies looking at the assembly of complex I 44,
45. We have also shown that cells over-expressing SLP-2 have increased ATP production
and oxygen consumption, although these cells do not express higher levels of the
respiratory chain components. Together, these results support a model in which

177
increased SLP-2 expression would lead to increased supercomplex formation, giving rise
to a better functioning respiratory chain.
In addition to increased ATP production, we propose that increased supercomplex
formation in response to SLP-2 over-expression also triggers an increase in mitochondrial
biogenesis. This response appears to rely on increased cardiolipin biosynthesis as
inhibition of cardiolipin synthase blocks mitochondrial biogenesis. It has been shown
that increases in matrix pH leads to increased cardiolipin biosynthesis 46. We propose
that enhanced cardiolipin-enriched domain formation upon SLP-2 over-expression
increases supercomplex formation, leading to more efficient respiratory chain function,
thus increasing the matrix pH and inducing cardiolipin biosynthesis. This, in turn, may
trigger nuclear transcription programs by an undefined mitochondria to nucleus signalling
pathway 47.
5.2.2 Calcium buffering
In addition to energy production, the mitochondria also play a major role in calcium
signalling as the transmembrane potential is a critical driving force to take calcium up
from the cytosol. SLP-2 has been shown to play a role in the calcium buffering capacity
of the mitochondria as cells treated with siRNA to deplete SLP-2 were found to have
decreased calcium uptake into the mitochondria and also to have an increased rate of
calcium efflux through the sodium/calcium exchanger 48. Treatment of these cells with
an inhibitor of the sodium/calcium exchanger equalized the rate of calcium efflux to that
of control cells, but SLP-2-depleted cells still had lower calcium uptake. This result
indicates that the loss of SLP-2 affects both the net calcium uptake as well as the loss of
calcium through the ion exchanger.
Our model of SLP-2 as an organizer of mitochondrial membranes could explain both the
ion channel-dependent and independent effects of SLP-2 depletion. As discussed above,
cardiolipin-enriched microdomains formed by association with SLP-2 would lead to more
efficient electron transport and a higher transmembrane potential, thus allowing an

178
increase in calcium uptake, which we have seen upon SLP-2 over-expression in Jurkat T
cells (C. Lemke and J. Madrenas, unpublished results). Furthermore, work by Da Cruz et
al has found a decrease in mitochondrial calcium efflux when SLP-2 is over-expressed in
HeLa cells, but no difference in the initial uptake 48. In the absence of SLP-2, the
formation of cardiolipin-enriched domains would be inhibited, leading to decreased
complex I activity, which could lead to a decrease in transmembrane potential. This, in
turn, would inhibit the net uptake of calcium into the mitochondria. Surprisingly, our
SLP-2-defecient T cells did not show a deficiency in mitochondrial calcium uptake, as
was expected from the work of Da Cruz et al, demonstrating decreased calcium uptake in
SLP-2-depleted HeLa cells 48. The discrepancy concerning the effect of SLP-2 on
mitochondrial calcium uptake could be due to differences in SLP-2 expression and over-
expression levels in mouse T cells, Jurkat T cells and HeLa cells. Both HeLa cells and
Jurkat T cells express high levels of endogenous SLP-2 and since the level of SLP-2
over-expression in the HeLa system was not reported, it is difficult to compare this
system to our Jurkat system. Perhaps our Jurkat T cells were over-expressing much
higher levels of SLP-2, leading to increased membrane compartmentalization and
respiratory chain activity, resulting in a higher transmembrane potential to drive in higher
levels of calcium. Primary mouse T cells express very low levels of SLP-2 and may rely
less heavily on SLP-2 membrane compartmentalization for calcium signalling compared
to HeLa cells. In this way, the loss of SLP-2 from the high expressing HeLa cells may
have a more prominent and detectable effect on mitochondrial calcium uptake compared
to the low SLP-2 expressing primary mouse T cells. These discrepancies also point to the
importance of examining the role of SLP-2 across different cell types, as it is likely that
different expression levels across tissues may alter the sensitivity to the loss of SLP-2 on
cell function.
Altered membrane organization could also explain the increased activity of the
sodium/calcium exchanger in Hela cells as DI-GEMs have been shown to play a role in
the regulation of a large number of ion channels 49-52. In this light, cardiolipin-enriched
microdomains formed by SLP-2 may play a role in the inhibition of calcium efflux from
the mitochondria, possibly through a direct inhibition of the ion channel by SLP-2 or

179
through an indirect mechanism in which an inhibitor of the channel is also recruited to
the cardiolipin-enriched microdomain.
5.2.3 Mitochondria membrane associated protease regulation
Early studies of the SPFH family members pointed to a role in the regulation of
proteases, as the loss of prohibitins was found to increase proteolysis of mitochondrial
proteins, reminiscent of the role of the bacterial protein HflC/K in the regulation of the
FtsH protease 7, 53. Subsequent work demonstrated that prohibitins bind directly to the
mitochondrial AAA-protease to negatively regulate its function and prevent proteolysis
of target proteins 53. Supporting this model of protease regulation, cells deficient in SLP-
2 due to treatment with targeted siRNA have decreased protein levels of prohibitins,
subunits of complex I and IV as well as altered processing of OPA-1 3, 54. Pointing to a
role for proteolysis over altered expression, treatment of SLP-2-deficient cells with a
metalloprotease inhibitor led to a partial recovery of protein levels, although the levels
did not return to those found in control cells 3. This result could indicate that both altered
expression and proteolysis may contribute to the decrease in protein levels or it could
point to wider variety of proteases acting to degrade these mitochondrial proteins in the
absence of SLP-2. As an alternative explanation for this degradation pattern, it is likely
that the organization of the mitochondrial membrane into defined domains could protect
proteins from proteolysis by excluding proteases, similar to the exclusion of the CD45
phosphatase from the immunological synapse to prevent premature termination of T cell
signalling 55. In this model, and as we have shown here, the loss of SLP-2 would lead to
altered distribution of cardiolipin into DI-GEMs. This in turn could prevent the physical
separation of the proteases from the target proteins, leading to the increase in protein
turnover. Furthermore, the partial recovery of protein levels upon metalloprotease
inhibition points to a role for a wider variety of proteases in protein turnover and the
physical segregation due to cardiolipin-enriched microdomains could protect proteins
from multiple proteases.

180
This regulation of protein turnover by physical segregation of target proteins from
proteases has implications for multiple mitochondrial functions, including energy
production as well as mitochondrial dynamics. Indeed, recently, SLP-2-deficient cells
have been shown to have an altered pattern of OPA-1 degradation, which in turn, inhibits
mitochondrial fusion in response to stress induction 54. In this study, cells were found to
undergo mitochondrial hyperfusion in response to stress induction by the treatment with
actinomycin D, cyclohexamide or UV and this response was protective against apoptotic
cell death. Knockdown of OPA-1, mitofusin1 or SLP-2 prevented hyperfusion and cells
showed increased cell death in response to stress induction. The induction of hyperfusion
also led to an increase in ATP production, indicating increased respiratory chain activity,
which may be partially explained by an increased formation of cardiolipin-enriched
domains, leading to enhanced supercomplex formation, as cells lacking SLP-2 failed to
increase ATP production. In agreement, we have found that SLP-2 over-expressing cells
are protected against apoptotic cell death, possibly through enhanced hyperfusion events
and increased ATP production.
5.2.4 Specialized membrane microdomain formation
In addition to the protection of proteins from proteolysis, individual cardiolipin-enriched
domains may recruit specific sets of proteins to alter various mitochondrial functions. As
discussed above, formation of domains enriched in respiratory chain components would
facilitate supercomplex assembly and lead to increased ATP production. Mitochondrial
fusion and hyperfusion may be facilitated by the formation of domains enriched in OPA-
1 and the recruitment of the mitofusins, as OPA-1 is found to constitutively associate
with cardiolipin-enriched domains and mitofusin 2 could be recruited through association
with SLP-2 23, 41. Similarly, these domains could also regulate mitochondrial fission as
hFis is constitutively associated with cardiolipin-enriched domains and DLP1 is recruited
in response to various treatments 23. In this case, the balance between fusion and fission
may be regulated by recruitment of factors to cardiolipin-enriched domains in response to
cellular signals. This has been demonstrated during induction of apoptosis, which
induces recruitment of pro-apoptotic factors tBid and Bax to cardiolipin-enriched

181
domains, indicating a role for cellular signalling in recruitment of proteins to these
domains 25.
5.2.5 T cell activation
Although the main pool of SLP-2 is found in association with mitochondrial membranes,
a small pool is also detected at the cell surface in T cells. While we initially hypothesized
that SLP-2 may be involved in mitochondrial translocation and maintenance at the
immunological synapse in response to stimulation, we found normal localization in SLP-
2-deficient cells. Based on this result, we propose that while the mechanism of the
surface pool and mitochondrial pool is likely conserved, it is probable that they act
independently to control unique cellular functions.
Immunosynapse formation plays an important role in regulating T cell activation upon
ligand binding. At early time points, up to 15 minutes after ligand binding, LFA-1 is
concentrated at the center of the contact site after which time TCR microclusters migrate
into the center of the synapse, surrounded by LFA-1, forming the cSMAC and pSMAC
regions respectively. By 60 minutes after binding, the TCR is no longer detected in the
cSMAC, which is likely required for down-regulation of T cell responsiveness 56. The
importance of the immunosynapse structure for T cell signalling has been show in mice
lacking CD2AP, which fail to form pSMAC and cSMAC structures 57. These cells show
altered responses to stimulation, with delayed ZAP-70 and ζ chain phosphorylation. This
delay is thought to occur because the formation of the immunosynapse is required to
optimize interactions between the TCR and MHC, which essential for proper TCR
signalling. For optimal TCR triggering, the TCR must be serially engaged by MHC
complexes. If ligand-binding affinity is too high, dissociation is prevented and there is no
serial triggering of TCR complexes, resulting in decreased TCR signalling. Likewise,
weak ligand binding also prevents TCR triggering. The immunological synapse is
proposed to promote TCR concentration at the interface with the APC, allowing
intermediate affinity ligands to activate T cells through rapid rebinding upon MHC
dissociation to strengthen T cell signalling responses.

182
Given the localization of the surface pool of SLP-2 surrounding TCR microclusters at the
interface with LFA-1, we propose that SLP-2 may strengthen the formation of the
immunological synapse by stabilizing the assembly of the TCR signalosome. As DI-
GEM domains coalesce during T cell activation, the plasma membrane-associated pool of
SLP-2 may be involved in facilitating and stabilizing these interactions, by self-
oligomerization as well as interactions with components of the T cell signalosome and
actin 2. This may result in the assembly of larger signalling complexes or may stabilize
the signalling complexes to allow stronger signalling responses. In addition, although not
shown experimentally, SLP-2 may interact with cholesterol to facilitate DI-GEM
clustering, as stomatin family members MEC-2 and podocin have both been shown to
interact with cholesterol through the shared SPFH domain 10. As SLP-2 levels increase,
the strength and duration of T cell signalosome association would also increase, leading
to increased T cell signalling and activation. Likewise, a loss of SLP-2 would result in
decreased stability of the DI-GEM domains in activated T cells, causing a dissociation of
signalling platforms, ultimately leading to impaired T cell responses. This model is
supported by results showing a decrease in T cell signalling upon SLP-2 depletion in
human T cells 2. Surprisingly, we did not see a loss of T cell signalling in SLP-2-
deficient murine T cells in response to a variety of stimulation conditions. As we have
not yet shown the plasma membrane-associated pool of SLP-2 to localize with TCR
microclusters in murine T cells, it is possible that surface-associated SLP-2 is not
involved in TCR signalosome formation in mice. Alternatively, this result could also
indicate that there is a redundancy in the function of SLP-2 in the formation or
stabilization of TCR microclusters in mice. We performed microarray analysis of both
naïve and stimulated SLP-2-deficient T cell but found no upregulation of any SLP-2-
related genes. However, it is possible that endogenous levels of other SPFH family
members could rescue TCR signalosome formation/stability in the absence of SLP-2.
Indeed, flotillins have been detected at the plasma membrane in T cells and previous
work in our lab has shown both stomatin and SLP-1 expression in Jurkat T cells (M.
Kirchhof and J. Madrenas, unpublished results) 11. It is possible that these family
members could facilitate TCR signalosome formation/stability in the absence of SLP-2

183
but since none are targeted to the mitochondria, these proteins are unable to rescue the
mitochondrial phenotype.
Although it is currently unclear how SLP-2 is localized to two cellular membranes or
why a small pool of a mitochondrial protein would be localized to the plasma membrane,
the localization of SLP-2 to the pSMAC of the immunological synapse indicates a role
for SLP-2 in T cell signalling and demonstrates that both pools of SLP-2 are likely
functional. Thus, at both the mitochondrial and plasma membrane, SLP-2 facilitates
membrane organization to regulate various cellular functions.
5.3 Future directions
5.3.1 Characterization of the structure of SLP-2
We have shown in this work that the deletion of the amino terminal region of SLP-2
prevents association with full length endogenous SLP-2. However, deletion mutants of
stomatin indicate that a short region at the C-terminus is required for homo-
oligomerization 32. Given the high degree of similarity between SLP-2 and the other
stomatin family members, it is likely that the region involved in oligomerization would
be conserved. An explanation for this discrepancy may involve the subcellular
localization of the SLP-2 truncation mutant. Since the mitochondrial target sequence is
located at the amino terminus and we have shown that loss of this region prevents
mitochondrial localization, it is likely that the loss of oligomerization between the
truncation mutant and endogenous full length SLP-2 is due to different subcellular
localization. Since endogenous SLP-2 is localized to the mitochondria and the truncation
mutant is cytosolic, it is unlikely that these proteins would form oligomers. In this way,
the work presented here has only touched on the role of the amino terminal region in
mitochondrial localization and the requirement for this localization to control
mitochondrial functions.
If our model of SLP-2 function is correct, proper functioning of SLP-2 will require
homo-oligomerization, as well as association with cardiolipin and with various

184
mitochondrial proteins. Each of these associations likely require distinct regions of the
SLP-2 protein, allowing SLP-2 to form homo-oligomers while also binding to cardiolipin
and other proteins, which would be required for the formation of cardiolipin-enriched
microdomains. A series of SLP-2 deletion constructs retaining mitochondrial targeting,
expressed in Jurkat T cells, could identify the regions required for homo-oligomerization
and for the association with known binding partners, such as the prohibitin complex.
Furthermore, since we know that SLP-2 over-expression increases mitochondrial
biogenesis and function, the link between loss of physical associations and SLP-2
function could be measured. The association with cardiolipin could be investigated by
generating recombinant proteins with point mutations to identify residues or regions
required for cardiolipin binding.
Information regarding the structure of the SPFH domain is essential to better understand
its functional role in membrane compartmentalization across all family members.
Currently, there is no crystal structure available for any member of the SPFH family.
However, we have generated recombinant human SLP-2, which could be used to generate
the crystal structure of SLP-2. This would provide a three-dimentional model of SLP-2
and could provide insight into the physical interaction with its binding partners as well as
information concerning the formation of microdomains in mitochondrial membranes.
This would represent the first crystal structure for a full length SFPH family member, as
only the SPFH domain has been crystalized thus far 8, 58.
5.3.2 Characterization of cardiolipin-enriched microdomains
Cardiolipin-enriched microdomains in the mitochondrial membrane have been isolated as
detergent-insoluble fractions and are thought to play a role in apoptosis induction 23, 25.
We have proposed here, that similar to other SPFH family members, SLP-2 plays a role
in membrane organization by facilitating the formation of these cardiolipin-enriched
microdomains 11, 14. However, while we have shown an interaction between SLP-2 and
cardiolipin, we have not directly shown the formation of cardiolipin domains and have
instead relied on detergent-insolubility to define these regions. As such, more work is
required to directly visualize cardiolipin-enriched regions within mitochondrial

185
membranes and to visualize the distribution of cardiolipin in the presence and absence of
SLP-2. This has proved to be difficult thus far due to the limitations of confocal
microscopy as it is difficult to image regions within the mitochondrial membrane.
Electron microscopy would provide excellent resolution of the mitochondrial membrane
structure and immunogold labeling could provide information on the distribution of SLP-
2 within the mitochondrial membrane, as well as the localization of SLP-2 binding
partners in the presence and absence of SLP-2. However, using this method to image
cardiolipin would be more complex and would require the use of a purified, specific
antibody against cardiolipin, which is not commercially available at this time.
Alternatively, cryo-electron microscopy and tomography could be used to obtain high-
resolution structures of the mitochondrial membrane 26, 59, 60. In this way, mitochondrial
membranes could be isolated and imaged to identify cardiolipin-enriched and protein-
enriched domains.
Although we have found prohibitins and components of the respiratory chain in
association with cardiolipin-enriched microdomains and others have reported the
association of pro-apoptotic proteins and proteins involved in mitochondrial dynamics 22,
25, there has not been a large-scale analysis of proteins localized to cardiolipin-enriched
microdomains. To this end, these regions could be isolated on the basis of detergent-
insolubility and subjected to mass spectrometry analysis. The proteins identified could
be organized into functional groups, such as respiratory chain components, components
of the protein import machinery, mitochondrial dynamics, etc. In this way, we could
identify those mitochondrial functions that might rely on cardiolipin-enriched
microdomain formation. This could also provide more insight into the function of SLP-2
in the formation of cardiolipin-enriched microdomains, as it is possible that some
mitochondrial functions may rely on SLP-2 for microdomain formation while others may
rely on microdomains formed independently of SLP-2. To investigate the requirement of
SLP-2, we could look at the targeting of the various proteins to cardiolipin-enriched
microdomains in the presence and absence of SLP-2.

186
5.3.3 Intracellular trafficking of SLP-2
While we have shown here that SLP-2 is found in two intracellular pools, at both the
mitochondrial and plasma membranes, it remains to be shown how this pattern of
localization arises. As demonstrated here, SLP-2 contains an amino-terminal
mitochondrial targeting sequence, which is required for mitochondrial localization 3, 41.
How SLP-2 traffics to the plasma membrane is currently unclear. It is possible that a
small portion of newly synthesized SLP-2 escapes mitochondrial trafficking and is
diverted to the plasma membrane by the default pathway of protein segregation 61.
However, an alternative explanation arises from the discovery of mitochondrial vesicles,
which may play a role in membrane exchange between the mitochondrial membrane and
other cellular membranes 62. In this way, it is possible that all newly synthesized SLP-2
is transported into the mitochondria, but the association with the mitochondrial
membrane allows SLP-2 to be transported to the plasma membrane in mitochondrial
vesicles, either by the passive loading of a mitochondrial protein or by an unidentified
pathway to target mitochondrial proteins for exchange. Given that membrane
organization is involved in vesicular transport, it is plausible that the organization of the
mitochondrial membrane into cardiolipin-enriched microdomains could facilitate
mitochondrial vesicle formation 18. Since work has shown that newly synthesized SLP-2
is cleaved upon transport into mitochondria 41, the amino acid sequence of plasma
membrane SLP-2 could provide insight into the cellular trafficking pathway taken by
SLP-2. To perform this analysis, SLP-2 could be isolated from both the mitochondria
and plasma membrane and subjected to mass spectrometry analysis to determine protein
sequence. If the plasma membrane pool is found to contain the full SLP-2 sequence,
while the mitochondrial pool is lacking the amino-terminal region, it is likely that the
plasma membrane pool traffics directly. However, if both mitochondrial- and plasma
membrane-associated SLP-2 are lacking the amino-terminal region, it is likely that the
plasma membrane pool arises from membrane exchange with the mitochondria. If SLP-2
does traffic through the mitochondria, this could point to a new paradigm in protein
trafficking, one that does not necessarily depend on a single targeting domain resulting in
a single intracellular localization. Instead, membrane exchange between various

187
organelles and the plasma membrane could result in protein trafficking independent of
targeting signalling domains encoded in the protein.
5.3.4 Functional role of SLP-2 in cell physiology
Although we have focused primarily on the role played by SLP-2 in T cells, expression of
SLP-2 is not restricted to T cells, but is widely expressed, being detected in such tissues
as brain, muscle and heart, among others 63. Indeed, as SLP-2 is a mitochondrial protein,
it is expressed across many tissues, especially those with high energy demand and the
loss of SLP-2 could lead to a wide variety of potential phenotypes, depending on the
affected tissue and its dependence on the mitochondria, as discussed below. In addition,
although mitochondria play an important role in T cell differentiation, activation of these
cells relies primarily on glycolysis 64-66. As such, it is not surprising that the deletion of
SLP-2 in T cells leads to a very subtle phenotypic effect. However, it is likely that
tissues relying more heavily on the energy production capacity of the mitochondria would
show a more prominent phenotype upon deletion of SLP-2. To this end, tamoxifen-
inducible SLP-2 knockout mice have been generated, in which treatment of mice with
tamoxifen results in widespread deletion of SLP-2 across all tissues. This model will
allow for the study of the effects of SLP-2 deletion on multiple tissues, including high
energy demanding tissues such as muscle and brain. It is likely that the loss of SLP-2 and
the subsequent depression of respiratory chain activity will impact the function of these
tissues, leading to a more prominent phenotype than that seen with T cell-specific
deletion. This model could point out tissues where SLP-2 plays a more essential
functional role, allowing more thorough investigation of SLP-2 function in these tissues.
A tamoxifen-inducible SLP-2 knockout model will also allow for the deletion of SLP-2 at
various time points during development, for further investigation of the role of SLP-2
during embryogenesis. As SLP-2 has been proposed to play a role in mitochondrial
dynamics and given that the deletion of multiple components of the fusion and fission
machinery are also embryonic lethal, the tamoxifen-inducible model could provide a
system to further study the role of mitochondrial dynamics during embryogenesis 54, 67.

188
5.4 SLP-2 and disease
5.4.1 SLP-2 and mitochondrial disorders
Mitochondria carry out many essential functions, primarily the generation of energy
required to power cellular functions. Given the requirement for mitochondria across all
tissues, it follows that dysfunctions of the mitochondria would have wide reaching
effects. Indeed, mitochondrial dysfunction has been implicated in a wide variety of
diseases, including cardiovascular disease, diabetes, gastrointestinal disease, neurological
diseases such as Alzheimer’s disease and Parkinson’s disease as well as cancer 68, 69.
Mitochondrial dysfunction is also the leading cause of inborn metabolic disorders 70.
This group of disorders is widely variable across patients, with differing symptoms and
levels of severity 71. This variability arises due to the complexity of the respiratory chain
and the large number of genes required for proper function. A large number of studies
have been performed on patients with a variety of mitochondrial disorders and many
inherited mutations have been identified, in both nuclear and mitochondrial DNA 71, 72.
The most common form of mitochondrial disorder in infants is Leigh syndrome, in which
patients tend to have normal pre-natal development, but develop symptoms during the
first year, with most dying before 3 years of age 73, 74. These patients generally exhibit
central and peripheral nervous system abnormalities, including optic atrophy,
opthalmoparesis, hypotonia, ataxia and dystonia 71, 74. While causative mutations have
been identified in all respiratory chain complexes, the largest number of mutations have
been identified within complex I genes, which is to be expected, as this complex requires
the correct expression and assembly of 45 subunits 73.
Complex I disorders also present as a spectrum of disorders with varying severity, in
some cases affecting isolated tissues while in other cases affecting multiple systems 70.
As expected, a number of genetic mutations have been identified as causative for this
group of disorders, including mutations in all of the subunits composing the core of
complex I 74. However, the essential role of complex I assembly has also been found to
play a role in mitochondrial disorders, as mutations in accessory subunits have also been

189
shown to be causative for complex I disorders 70, 73. Given the proposed role for SLP-2 in
the assembly of complex I, further study of SLP-2 and cardiolipin-enriched domains may
provide insight into regulation of complex I assembly, which would in turn provide
insight into the role of complex I assembly and mitochondrial disorders.
Another group of mitochondrial disorders involves mutations in proteins involved in
mitochondrial dynamics. The mutation of mitofusin 2 and OPA-1 both cause neural
disorders, characterized by impairment of motor and sensory neurons or the optic nerve,
respectively 75. Furthermore, altered regulation of mitochondrial fusion and fission have
been found in a number of neurological disorders, including Alzheimer’s disease,
Parkinson’s disease and Huntington’s disease 76. Given that SLP-2 has been shown to
play a role in mitochondrial hyperfusion in response to cellular stress, possibly by
regulating the cleavage of OPA-1, a further understanding of the role for SLP-2 in
mitochondrial dynamics could provide insight into these neurological diseases.
5.4.2 SLP-2 in cancer prognosis and metastasis
The role of SLP-2 in membrane organization has effects on multiple cellular functions,
which when deregulated may have health implications. Indeed, a number of studies have
found an upregulation of SLP-2 expression in multiple types of cancer, including
esophageal squamous cell carcinoma, lung cancer, laryngeal cancer, endometrial
adenocarcinoma, breast cancer, laryngeal squamous cell carcinoma, osteosarcoma and
pulmonary squamous cell carcinoma 77-81. Furthermore, SLP-2 expression has been
shown to positively correlate with tumor size, lymph node metastasis, clinical stage,
distant metastasis and decreased overall survival and disease-free survival 78, 79, 81. To
investigate possible mechanisms of SLP-2 in cancer progression, SLP-2 was knocked
down in esophageal squamous cell carcinoma cells, which led to decreased cell growth
due to S-phase arrest with no change in apoptosis 77. When these cells were injected into
nude mice, they showed slower tumor progression and smaller tumors compared to mice
injected with the parental cells. Upon treatment with chemotherapeutic agents, the SLP-
2-depleted cells showed increased apoptosis compared to parental cells 42. Finally, SLP-

190
2-depleted cells were also reported to have slightly decreased transmembrane potential
and ATP levels. The reported changes were very small and likely did not influence the
energy requirements of the cells as cancer cells are known to rely more heavily on
glycolysis for ATP production 82. However, these changes may reflect the ability of SLP-
2 to induce hyperfusion upon stress induction, which may decrease the apoptotic
response of the SLP-2 expressing cancer cells in response to chemotherapeutics. The loss
of SLP-2 expression by siRNA depletion also led to decreased cell motility, both in the
migration of cells in a wound healing model and in cell migration across a transwell 42.
Given that the expression of SLP-2 is positively correlated with lymph node and distant
metastasis in multiple types of cancer 78, 79, 81, it is likely that SLP-2 may play a direct role
in metastasis by regulating the migration of cancer cells. Indeed, we have found SLP-2
in association with the adhesion factor LFA-1, which may point to a role in the regulation
of both cell adhesion and migration. Together, these results indicate that proper
regulation of SLP-2 expression may have an important role in cancer progression and
treatment.
5.5 Conclusions
Although SLP-2 is a widely expressed mitochondrial protein belonging to a highly
conserved protein family, very little work has been done to study this protein and the
mechanism of action has remained unclear. The work presented in this thesis provides
novel insight into the role of SLP-2 in T cells and points to a mechanism for SLP-2 in the
organization of membranes into functional domains.
We initially confirmed the mitochondrial localization of SLP-2 in Chapter 2 by
constructing stably transfected Jurkat T cell expressing SLP-2-GFP. The co-localization
of SLP-2-GFP with MitoTracker Red demonstrated the mitochondrial localization of
SLP-2. In addition, the over-expression of this SLP-2-GFP construct led to the finding
that increased expression of SLP-2 caused an increase in mitochondrial biogenesis, along
with an increase in activity of the respiratory chain and protection against apoptosis.
Providing evidence for the mechanism of action, we also showed that SLP-2 can bind

191
directly to cardiolipin as well as to the prohibitin complex and over-expression of SLP-2
increased the recruitment of the prohibitin complex to the mitochondrial membrane. We
next constructed T cell-specific SLP-2 conditional knockout mice, as described in
Chapter 3, and found that while SLP-2 was not required for T cell development, it was
required for optimal T cell activation. Importantly, we have shown that the decreased T
cell activation upon in vitro stimulation is biologically relevant, as our SLP-2 T-K/O
mice show a decreased rate of non-matched transplant rejection. Furthermore, this model
also provided additional evidence for the mechanism of action for SLP-2 in that T cells
lacking SLP-2 showed decreased levels of complex I subunits as well as lower activity of
complex I. Additionally, these cells also had lower levels of cardiolipin recruited to
detergent-insoluble microdomains within the mitochondrial membrane. Finally, in
Chapter 4 we have identified a second subcellular localization of SLP-2. In addition to
the mitochondrial localization, a small pool of SLP-2 was found at the plasma membrane.
We have shown that this pool was recruited to the pSMAC of the immunological synapse
in response to T cell stimulation, pointing to a role for the surface pool in T cell
activation. Furthermore, we have also shown the ability of SLP-2 to homo-oligomerize,
providing further evidence for our proposed model of SLP-2 function.
Based on these results, we propose that SLP-2 exerts effects on mitochondrial and
cellular functions by facilitating the organization of membranes into functional
microdomains. At the mitochondrial membrane, this is likely carried out in the following
steps: i) SLP-2 binds directly to cardiolipin, ii) SLP-2 undergoes homo-oligomerization
to concentrate cardiolipin into cardiolipin-enriched region, iii) SLP-2 and cardiolipin bind
to other proteins, recruiting them to cardiolipin-enriched domains to define domain
function. This model may be extended to the plasma membrane pool of SLP-2, if SLP-2
is capable of binding to cholesterol in a manner similar to its binding to cardiolipin. In
this way, SLP-2 facilitates membrane microdomain formation to effect such
mitochondrial and cellular functions as energy production, apoptotic induction and T cell
activation.

192
5.6 References
1. Osman, C., Haag, M., Potting, C., Rodenfels, J., Dip, P.V., Wieland, F.T., Brugger, B., Westermann, B. & Langer, T. The genetic interactome of prohibitins: coordinated control of cardiolipin and phosphatidylethanolamine by conserved regulators in mitochondria. J Cell Biol 184, 583-596 (2009).
2. Kirchhof, M.G., Chau, L.A., Lemke, C.D., Vardhana, S., Darlington, P.J., Marquez, M.E., Taylor, R., Rizkalla, K., Blanca, I., Dustin, M.L. & Madrenas, J. Modulation of T cell activation by stomatin-like protein 2. J Immunol 181, 1927-1936 (2008).
3. Da Cruz, S., Parone, P.A., Gonzalo, P., Bienvenut, W.V., Tondera, D., Jourdain, A., Quadroni, M. & Martinou, J.C. SLP-2 interacts with prohibitins in the mitochondrial inner membrane and contributes to their stability. Biochim Biophys Acta 1783, 904-911 (2008).
4. Quintana, A., Schwindling, C., Wenning, A.S., Becherer, U., Rettig, J., Schwarz, E.C. & Hoth, M. T cell activation requires mitochondrial translocation to the immunological synapse. Proc Natl Acad Sci U S A 104, 14418-14423 (2007).
5. Browman, D.T., Hoegg, M.B. & Robbins, S.M. The SPFH domain-containing proteins: more than lipid raft markers. Trends Cell Biol 17, 394-402 (2007).
6. Green, J.B. & Young, J.P. Slipins: ancient origin, duplication and diversification of the stomatin protein family. BMC Evol Biol 8, 44 (2008).
7. Tavernarakis, N., Driscoll, M. & Kyrpides, N.C. The SPFH domain: implicated in regulating targeted protein turnover in stomatins and other membrane-associated proteins. Trends Biochem Sci 24, 425-427 (1999).
8. Kuwahara, Y., Unzai, S., Nagata, T., Hiroaki, Y., Yokoyama, H., Matsui, I., Ikegami, T., Fujiyoshi, Y. & Hiroaki, H. Unusual thermal disassembly of the SPFH domain oligomer from Pyrococcus horikoshii. Biophys J 97, 2034-2043 (2009).
9. Snyers, L., Umlauf, E. & Prohaska, R. Cysteine 29 is the major palmitoylation site on stomatin. FEBS Lett 449, 101-104 (1999).
10. Huber, T.B., Schermer, B., Muller, R.U., Hohne, M., Bartram, M., Calixto, A., Hagmann, H., Reinhardt, C., Koos, F., Kunzelmann, K., Shirokova, E., Krautwurst, D., Harteneck, C., Simons, M., Pavenstadt, H., Kerjaschki, D., Thiele, C., Walz, G., Chalfie, M. & Benzing, T. Podocin and MEC-2 bind cholesterol to regulate the activity of associated ion channels. Proc Natl Acad Sci U S A 103, 17079-17086 (2006).

193
11. Langhorst, M.F., Reuter, A. & Stuermer, C.A. Scaffolding microdomains and beyond: the function of reggie/flotillin proteins. Cell Mol Life Sci 62, 2228-2240 (2005).
12. Morrow, I.C. & Parton, R.G. Flotillins and the PHB domain protein family: rafts, worms and anaesthetics. Traffic 6, 725-740 (2005).
13. Stuermer, C.A. Microdomain-forming proteins and the role of the reggies/flotillins during axon regeneration in zebrafish. Biochim Biophys Acta 1812, 415-422 (2011).
14. Osman, C., Merkwirth, C. & Langer, T. Prohibitins and the functional compartmentalization of mitochondrial membranes. J Cell Sci 122, 3823-3830 (2009).
15. Huber, T.B., Simons, M., Hartleben, B., Sernetz, L., Schmidts, M., Gundlach, E., Saleem, M.A., Walz, G. & Benzing, T. Molecular basis of the functional podocin-nephrin complex: mutations in the NPHS2 gene disrupt nephrin targeting to lipid raft microdomains. Hum Mol Genet 12, 3397-3405 (2003).
16. Artal-Sanz, M. & Tavernarakis, N. Prohibitin and mitochondrial biology. Trends Endocrinol Metab 20, 394-401 (2009).
17. Goodman, M.B., Ernstrom, G.G., Chelur, D.S., O'Hagan, R., Yao, C.A. & Chalfie, M. MEC-2 regulates C. elegans DEG/ENaC channels needed for mechanosensation. Nature 415, 1039-1042 (2002).
18. Simons, K. & Gerl, M.J. Revitalizing membrane rafts: new tools and insights. Nat Rev Mol Cell Biol 11, 688-699 (2010).
19. Lingwood, D. & Simons, K. Lipid rafts as a membrane-organizing principle. Science 327, 46-50 (2010).
20. Leslie, M. Mysteries of the cell. Do lipid rafts exist? Science 334, 1046-1047 (2011).
21. Epand, R.F., Tokarska-Schlattner, M., Schlattner, U., Wallimann, T. & Epand, R.M. Cardiolipin clusters and membrane domain formation induced by mitochondrial proteins. J Mol Biol 365, 968-980 (2007).
22. Sorice, M., Manganelli, V., Matarrese, P., Tinari, A., Misasi, R., Malorni, W. & Garofalo, T. Cardiolipin-enriched raft-like microdomains are essential activating platforms for apoptotic signals on mitochondria. FEBS Lett 583, 2447-2450 (2009).
23. Ciarlo, L., Manganelli, V., Garofalo, T., Matarrese, P., Tinari, A., Misasi, R., Malorni, W. & Sorice, M. Association of fission proteins with mitochondrial raft-like domains. Cell Death Differ 17, 1047-1058 (2010).

194
24. Garofalo, T., Tinari, A., Matarrese, P., Giammarioli, A.M., Manganelli, V., Ciarlo, L., Misasi, R., Sorice, M. & Malorni, W. Do mitochondria act as "cargo boats" in the journey of GD3 to the nucleus during apoptosis? FEBS Lett 581, 3899-3903 (2007).
25. Garofalo, T., Giammarioli, A.M., Misasi, R., Tinari, A., Manganelli, V., Gambardella, L., Pavan, A., Malorni, W. & Sorice, M. Lipid microdomains contribute to apoptosis-associated modifications of mitochondria in T cells. Cell Death Differ 12, 1378-1389 (2005).
26. Acehan, D., Malhotra, A., Xu, Y., Ren, M., Stokes, D.L. & Schlame, M. Cardiolipin affects the supramolecular organization of ATP synthase in mitochondria. Biophys J 100, 2184-2192 (2011).
27. Schlame, M., Rua, D. & Greenberg, M.L. The biosynthesis and functional role of cardiolipin. Prog Lipid Res 39, 257-288 (2000).
28. Joshi, A.S., Zhou, J., Gohil, V.M., Chen, S. & Greenberg, M.L. Cellular functions of cardiolipin in yeast. Biochim Biophys Acta 1793, 212-218 (2008).
29. Fry, M. & Green, D.E. Cardiolipin requirement for electron transfer in complex I and III of the mitochondrial respiratory chain. J Biol Chem 256, 1874-1880 (1981).
30. Mairhofer, M., Steiner, M., Salzer, U. & Prohaska, R. Stomatin-like protein-1 interacts with stomatin and is targeted to late endosomes. J Biol Chem 284, 29218-29229 (2009).
31. Sedensky, M.M., Siefker, J.M., Koh, J.Y., Miller, D.M., 3rd & Morgan, P.G. A stomatin and a degenerin interact in lipid rafts of the nervous system of Caenorhabditis elegans. Am J Physiol Cell Physiol 287, C468-474 (2004).
32. Umlauf, E., Mairhofer, M. & Prohaska, R. Characterization of the stomatin domain involved in homo-oligomerization and lipid raft association. J Biol Chem 281, 23349-23356 (2006).
33. Tatsuta, T., Model, K. & Langer, T. Formation of membrane-bound ring complexes by prohibitins in mitochondria. Mol Biol Cell 16, 248-259 (2005).
34. Berger, K.H. & Yaffe, M.P. Prohibitin family members interact genetically with mitochondrial inheritance components in Saccharomyces cerevisiae. Mol Cell Biol 18, 4043-4052 (1998).
35. Sorice, M., Matarrese, P., Tinari, A., Giammarioli, A.M., Garofalo, T., Manganelli, V., Ciarlo, L., Gambardella, L., Maccari, G., Botta, M., Misasi, R. & Malorni, W. Raft component GD3 associates with tubulin following CD95/Fas ligation. FASEB J 23, 3298-3308 (2009).

195
36. Osman, C., Voelker, D.R. & Langer, T. Making heads or tails of phospholipids in mitochondria. J Cell Biol 192, 7-16 (2011).
37. Acin-Perez, R., Fernandez-Silva, P., Peleato, M.L., Perez-Martos, A. & Enriquez, J.A. Respiratory active mitochondrial supercomplexes. Mol Cell 32, 529-539 (2008).
38. Choi, S.Y., Gonzalvez, F., Jenkins, G.M., Slomianny, C., Chretien, D., Arnoult, D., Petit, P.X. & Frohman, M.A. Cardiolipin deficiency releases cytochrome c from the inner mitochondrial membrane and accelerates stimuli-elicited apoptosis. Cell Death Differ 14, 597-606 (2007).
39. Zhang, M., Mileykovskaya, E. & Dowhan, W. Gluing the respiratory chain together. Cardiolipin is required for supercomplex formation in the inner mitochondrial membrane. J Biol Chem 277, 43553-43556 (2002).
40. Zhang, M., Mileykovskaya, E. & Dowhan, W. Cardiolipin is essential for organization of complexes III and IV into a supercomplex in intact yeast mitochondria. J Biol Chem 280, 29403-29408 (2005).
41. Hajek, P., Chomyn, A. & Attardi, G. Identification of a novel mitochondrial complex containing mitofusin 2 and stomatin-like protein 2. J Biol Chem 282, 5670-5681 (2007).
42. Wang, Y., Cao, W., Yu, Z. & Liu, Z. Downregulation of a mitochondria associated protein SLP-2 inhibits tumor cell motility, proliferation and enhances cell sensitivity to chemotherapeutic reagents. Cancer Biol Ther 8, 1651-1658 (2009).
43. McKenzie, M. & Ryan, M.T. Assembly factors of human mitochondrial complex I and their defects in disease. IUBMB Life 62, 497-502 (2010).
44. Marques, I., Dencher, N.A., Videira, A. & Krause, F. Supramolecular organization of the respiratory chain in Neurospora crassa mitochondria. Eukaryot Cell 6, 2391-2405 (2007).
45. Bourges, I., Ramus, C., Mousson de Camaret, B., Beugnot, R., Remacle, C., Cardol, P., Hofhaus, G. & Issartel, J.P. Structural organization of mitochondrial human complex I: role of the ND4 and ND5 mitochondria-encoded subunits and interaction with prohibitin. Biochem J 383, 491-499 (2004).
46. Gohil, V.M., Hayes, P., Matsuyama, S., Schagger, H., Schlame, M. & Greenberg, M.L. Cardiolipin biosynthesis and mitochondrial respiratory chain function are interdependent. J Biol Chem 279, 42612-42618 (2004).
47. Scarpulla, R.C. Transcriptional paradigms in mammalian mitochondrial biogenesis and function. Physiol Rev 88, 611-638 (2008).

196
48. Da Cruz, S., De Marchi, U., Frieden, M., Parone, P.A., Martinou, J.C. & Demaurex, N. SLP-2 negatively modulates mitochondrial sodium-calcium exchange. Cell Calcium 47, 11-18 (2010).
49. Dart, C. Lipid microdomains and the regulation of ion channel function. J Physiol 588, 3169-3178 (2010).
50. Levitan, I., Fang, Y., Rosenhouse-Dantsker, A. & Romanenko, V. Cholesterol and ion channels. Subcell Biochem 51, 509-549 (2010).
51. Pani, B. & Singh, B.B. Lipid rafts/caveolae as microdomains of calcium signaling. Cell Calcium 45, 625-633 (2009).
52. Pristera, A. & Okuse, K. Building Excitable Membranes: Lipid Rafts and Multiple Controls on Trafficking of Electrogenic Molecules. Neuroscientist Epub ahead of print (2011).
53. Steglich, G., Neupert, W. & Langer, T. Prohibitins regulate membrane protein degradation by the m-AAA protease in mitochondria. Mol Cell Biol 19, 3435-3442 (1999).
54. Tondera, D., Grandemange, S., Jourdain, A., Karbowski, M., Mattenberger, Y., Herzig, S., Da Cruz, S., Clerc, P., Raschke, I., Merkwirth, C., Ehses, S., Krause, F., Chan, D.C., Alexander, C., Bauer, C., Youle, R., Langer, T. & Martinou, J.C. SLP-2 is required for stress-induced mitochondrial hyperfusion. EMBO J 28, 1589-1600 (2009).
55. Fooksman, D.R., Vardhana, S., Vasiliver-Shamis, G., Liese, J., Blair, D.A., Waite, J., Sacristan, C., Victora, G.D., Zanin-Zhorov, A. & Dustin, M.L. Functional anatomy of T cell activation and synapse formation. Annu Rev Immunol 28, 79-105 (2010).
56. Lee, K.H., Holdorf, A.D., Dustin, M.L., Chan, A.C., Allen, P.M. & Shaw, A.S. T cell receptor signaling precedes immunological synapse formation. Science 295, 1539-1542 (2002).
57. Lee, K.H., Dinner, A.R., Tu, C., Campi, G., Raychaudhuri, S., Varma, R., Sims, T.N., Burack, W.R., Wu, H., Wang, J., Kanagawa, O., Markiewicz, M., Allen, P.M., Dustin, M.L., Chakraborty, A.K. & Shaw, A.S. The immunological synapse balances T cell receptor signaling and degradation. Science 302, 1218-1222 (2003).
58. Yokoyama, H., Fujii, S. & Matsui, I. Crystal structure of a core domain of stomatin from Pyrococcus horikoshii Illustrates a novel trimeric and coiled-coil fold. J Mol Biol 376, 868-878 (2008).

197
59. Dudkina, N.V., Kouril, R., Bultema, J.B. & Boekema, E.J. Imaging of organelles by electron microscopy reveals protein-protein interactions in mitochondria and chloroplasts. FEBS letters 584, 2510-2515 (2010).
60. Fujiyoshi, Y. & Unwin, N. Electron crystallography of proteins in membranes. Curr Opin Struct Biol 18, 587-592 (2008).
61. Burgess, T.L. & Kelly, R.B. Constitutive and regulated secretion of proteins. Annu Rev Cell Biol 3, 243-293 (1987).
62. Andrade-Navarro, M.A., Sanchez-Pulido, L. & McBride, H.M. Mitochondrial vesicles: an ancient process providing new links to peroxisomes. Curr Opin Cell Biol 21, 560-567 (2009).
63. Wang, Y. & Morrow, J.S. Identification and characterization of human SLP-2, a novel homologue of stomatin (band 7.2b) present in erythrocytes and other tissues. J Biol Chem 275, 8062-8071 (2000).
64. Pearce, E.L., Walsh, M.C., Cejas, P.J., Harms, G.M., Shen, H., Wang, L.S., Jones, R.G. & Choi, Y. Enhancing CD8 T-cell memory by modulating fatty acid metabolism. Nature 460, 103-107 (2009).
65. Michalek, R.D., Gerriets, V.A., Jacobs, S.R., Macintyre, A.N., MacIver, N.J., Mason, E.F., Sullivan, S.A., Nichols, A.G. & Rathmell, J.C. Cutting edge: distinct glycolytic and lipid oxidative metabolic programs are essential for effector and regulatory CD4+ T cell subsets. J Immunol 186, 3299-3303 (2011).
66. Frauwirth, K.A., Riley, J.L., Harris, M.H., Parry, R.V., Rathmell, J.C., Plas, D.R., Elstrom, R.L., June, C.H. & Thompson, C.B. The CD28 signaling pathway regulates glucose metabolism. Immunity 16, 769-777 (2002).
67. Chen, H. & Chan, D.C. Emerging functions of mammalian mitochondrial fusion and fission. Hum Mol Genet 14 Spec No. 2, R283-289 (2005).
68. Fosslien, E. Mitochondrial medicine--molecular pathology of defective oxidative phosphorylation. Ann Clin Lab Sci 31, 25-67 (2001).
69. Brandon, M., Baldi, P. & Wallace, D.C. Mitochondrial mutations in cancer. Oncogene 25, 4647-4662 (2006).
70. Lazarou, M., Thorburn, D.R., Ryan, M.T. & McKenzie, M. Assembly of mitochondrial complex I and defects in disease. Biochim Biophys Acta 1793, 78-88 (2009).
71. Smeitink, J., van den Heuvel, L. & DiMauro, S. The genetics and pathology of oxidative phosphorylation. Nat Rev Genet 2, 342-352 (2001).

198
72. Calvo, S., Jain, M., Xie, X., Sheth, S.A., Chang, B., Goldberger, O.A., Spinazzola, A., Zeviani, M., Carr, S.A. & Mootha, V.K. Systematic identification of human mitochondrial disease genes through integrative genomics. Nat Genet 38, 576-582 (2006).
73. Koene, S., Willems, P.H., Roestenberg, P., Koopman, W.J. & Smeitink, J.A. Mouse models for nuclear DNA-encoded mitochondrial complex I deficiency. J Inherit Metab Dis 34, 293-307 (2011).
74. Finsterer, J. Leigh and Leigh-like syndrome in children and adults. Pediatr Neurol 39, 223-235 (2008).
75. Chen, H. & Chan, D.C. Physiological functions of mitochondrial fusion. Ann N Y Acad Sci 1201, 21-25 (2010).
76. Cho, D.H., Nakamura, T. & Lipton, S.A. Mitochondrial dynamics in cell death and neurodegeneration. Cellular and molecular life sciences : CMLS 67, 3435-3447 (2010).
77. Zhang, L., Ding, F., Cao, W., Liu, Z., Liu, W., Yu, Z., Wu, Y., Li, W. & Li, Y. Stomatin-like protein 2 is overexpressed in cancer and involved in regulating cell growth and cell adhesion in human esophageal squamous cell carcinoma. Clin Cancer Res 12, 1639-1646 (2006).
78. Cao, W., Zhang, B., Liu, Y., Li, H., Zhang, S., Fu, L., Niu, Y., Ning, L., Cao, X., Liu, Z. & Sun, B. High-level SLP-2 expression and HER-2/neu protein expression are associated with decreased breast cancer patient survival. Am J Clin Pathol 128, 430-436 (2007).
79. Cao, W.F., Zhang, L.Y., Liu, M.B., Tang, P.Z., Liu, Z.H. & Sun, B.C. Prognostic significance of stomatin-like protein 2 overexpression in laryngeal squamous cell carcinoma: clinical, histologic, and immunohistochemistry analyses with tissue microarray. Hum Pathol 38, 747-752 (2007).
80. Guo, Q.C., Shen, J.N., Jin, S., Wang, J., Huang, G., Zhang, L.J., Yin, J.Q., Zou, C.Y. & Li, M.T. Comparative proteomic analysis of human osteosarcoma and SV40-immortalized normal osteoblastic cell lines. Acta Pharmacol Sin 28, 850-858 (2007).
81. Chang, D., Ma, K., Gong, M., Cui, Y., Liu, Z.H., Zhou, X.G., Zhou, C.N. & Wang, T.Y. SLP-2 overexpression is associated with tumour distant metastasis and poor prognosis in pulmonary squamous cell carcinoma. Biomarkers 15, 104-110 (2009).
82. Warburg, O. On respiratory impairment in cancer cells. Science 124, 269-270 (1956).

199
Appendix A: Ethics Approval

200

201
Appendix B: Copyright Permission
AMERICAN SOCIETY FOR MICROBIOLOGY LICENSE TERMS AND CONDITIONS
Sep 19, 2011 This is a License Agreement between Darah A Christie ("You") and American Society for Microbiology ("American Society for Microbiology") provided by Copyright Clearance Center ("CCC"). The license consists of your order details, the terms and conditions provided by American Society for Microbiology, and the payment terms and conditions. License Number 2752590906268 License date Sep 19, 2011 Licensed content publisher American Society for Microbiology Licensed content publication Molecular and Cellular Biology Licensed content title Stomatin-Like Protein 2 Binds Cardiolipin and
Regulates Mitochondrial Biogenesis and Function
Licensed content author Darah A. Christie, Joaquin Madrenas Licensed content date Sep 15, 2011 Volume 31 Issue 18 Start page 3845 End page 3856 Type of Use Dissertation/Thesis Format Electronic Portion Full article Order reference number Title of your thesis / dissertation
Characterization of SLP-2 function in T lymphocytes
Expected completion date Jan 2012 Estimated size(pages) 200 Billing Type Invoice Billing address Robarts Research Institute Room 2278, 100 Perth Drive London, ON N6A 5K8 Canada Customer reference info Total 0.00 USD

202
Curriculum Vitae
Name Darah Christie
Education
2004 B.Sc. University of Waterloo, Waterloo, Ontario, Canada Honours Biochemistry
2007 M.Sc. University of Waterloo, Waterloo, Ontario, Canada
Department of Biology Supervisor: Dr. B. Dixon
2008-12 Ph.D. University of Western Ontario, London, Ontario Canada (Expected) Department of Microbiology and Immunology Supervisor: Dr. J. Madrenas
Honours and Awards
2004 NSERC – Undergraduate Student Research Award Entrance Scholarship – University of Waterloo
2005 CIHR Canada Graduate Scholarship – Master’s Award (Declined) Ontario Graduate Scholarship President’s Graduate Scholarship – University of Waterloo Entrance Scholarship – University of Waterloo 2007 Graduate Studies Research Travel Assistantship – University of Waterloo 2008 CIHR Doctoral Research Award (2008-2011)
Microbiology and Immunology Graduate Entrance Award Western Graduate Research Scholarship – Microbiology Schulich Scholarship for Medical Research
2009 eBioscience Award for Excellence in Graduate Poster Presentation
Western Graduate Research Scholarship – Microbiology Schulich Scholarship for Medical Research
2011 Microbiology and Immunology Graduate Travel Award Ontario Graduate Scholarship

203
Research Publications
1. Christie, D., Wei, G., Fujiki, K. and Dixon, B. Cloning and characterization of a cDNA encoding walleye (Sander vitreum) beta-2 microglobulin. Fish Shellfish Immun. 22:727-733 (2007).
2. Yue, D., Brintnell, W., Mannik, L.A., Christie, D., Haeryfar, S. M., Madrenas, J., Chakrabarti, S., Bell, D.A. and Cairns, E. CTLA4-Ig blocks the development and progression of citrullinated human fibrinogen-induced arthritis in DR4 tg mice. Arthritis Rheum 62(10) 2941-52 (2010).
3. Christie, D., Lemke, C.D., Elias, I.M., Chau, L.A., Kirchhof, M.G., Li, B., Ball, E.H., Dunn, S.D., Hatch, G.M. and Madrenas, J. Stomatin-like protein 2 binds cardiolipin and regulates mitochondrial biogenesis and function. Mol. Cell. Biol 31 (18) 3845-56 (2011).
4. Christie, D., Lian, D., Wang, H., Hatch, G. and Madrenas, J. SLP-2 deficiency in T cells is associated with abnormal mitochondrial compartmentalization of cardiolipin, decreased mitochondrial respiration and impaired CD4+ T cell responses. Manuscript in revision.
5. Christie, D., Kirchhof, M.G., Vardhana, S., Dustin, M.L., Madrenas, J. Mitochondrial and plasma membrane pools of stomatin-like protein 2 coalesce at the immunological synapse during T cell activation. Manuscript submitted.
Abstracts and Presentations
1. Graduate Student Research Conference. Waterloo, Ont. May 2007. Abstract and
oral presentation. “Regulation of Major Histocompatibility Class II Associated Invariant Chain Genes in Rainbow Trout”
2. Canadian Society of Zoology Annual Meeting. Montreal, Que. May 2007. Abstract and poster presentation. “Regulation of Major Histocompatibility Class II Associated Invariant Chain Genes in Rainbow Trout”
3. Robarts Research Day. London, Ont. March 2008. Abstract and poster
presentation. “Generation of a T Cell-Specific Knockout Mouse for Stomatin-like Protein 2 (SLP-2)”

204
4. Margaret Moffat Research Day. London, Ont. March 2008. Abstract and poster presentation. “Generation of a T Cell-Specific Knockout Mouse for Stomatin-like Protein 2 (SLP-2)”
5. Infection and Immunity Research Forum. London, Ont. November 2008.
Abstract and poster presentation. “Generation of a T Cell-Specific Knockout Mouse for Stomatin-like Protein 2 (SLP-2)”
6. American Association of Immunology Annual Meeting. Seattle, WA. May 2009. Abstract, oral and poster presentation. “Stomatin-like protein 2: a link between T cell activation and mitochondrial function”
7. Infection and Immunity Research Forum. London, Ont. November 2009.
Abstract and poster presentation. “The role of stomatin-like protein 2 in mitochondrial biogenesis”
8. Cold Spring Harbor, Gene Expression & Signaling in the Immune System. Cold
Spring Harbor, NY. April 2010. Abstract and poster presentation. “Stomatin-like protein 2: a link between T cell activation and mitochondrial function”
9. American Association of Immunology Annual Meeting. San Francisco, CA.
May 2011. Abstract and poster presentation. “SLP-2 deficiency in T cells is associated with decreased respiratory complex I levels and impaired TCR-dependent responses”
Teaching Experience
2004 Fundamentals of Microbiology - Laboratory Techniques
2005 Techniques in Molecular Biology Virology - Laboratory Techniques
2006 Techniques in Molecular Biology
Cellular and Molecular Immunology - Laboratory Techniques
2007 Cellular and Molecular Immunology - Laboratory Techniques Introduction to Applied Microbiology
2008 - 11 Laboratory Techniques in Microbiology and Immunology
Relevant Technical Experience
Bacterial expression of recombinant protein Polyclonal antibody production and purification

205
Southern, northern and western blotting One-dimensional and two-dimensional SDS-PAGE Reverse Transcriptase (RT) PCR Real Time PCR Mitochondrial isolation Chemical cell transformation Enzyme-Linked Immunosorbent Assay (ELISA) In vitro cell culture – primary culture and established cell lines Flow cytometry Confocal microscopy Animal handling
Training and Workshops
2008 Mitochondrial Molecular Biology and Pathology, May
National Institutes of Health, Bethesda, Maryland
Voluntary Services
2005-07 Grand River Regional Cancer Center – Present the New Patient
Orientation Session weekly
2006 TA Workshop – help train new teaching assistants 2009 Fall Preview Day – Discuss Microbiology and Immunology undergraduate
program with prospective students 2010 Fall Graduate Student Recruitment Fair – Discuss graduate work in the
Schulich School of Medicine and Dentistry 2010-11 Immunity and Infection Research Forum Committee Member – V.P. of
Finance and V.P. of Travel