Embed Size (px)
Citation preview

THE JOURNAL OF BIOLOGICAL CHEMISTRY C) 1991 by The American Society for Biochemistry and Molecular Biology, Inc.
Vol. 266, No 33, Issue of November ’ 25, pp. 22522-22528.1991 Printed in U. S. A.
Characterization of Rabbit Lung Lysosomes and Their Role in Surfactant Dipalmitoylphosphatidylcholine Catabolism*
(Received for publication, June 11, 1991)
Evelyn D. Rider$, Kent E. Pinkertong, and Alan H. Jobe$ From the $Harbor- UCLA Medical Center, Department of Pediatrics, Torrance, California, 90509, and the §University of California, School of Veterinary Medicine, Department of Anatomy, Davis, California 95616
Although alveolar surfactant is rapidly catabolized in adult rabbit lungs, the pathways have not been characterized. Pathways of surfactant secretion and recycling involve lamellar bodies and multivesicular bodies, organelles shown to be related to lysosomes by cytochemistry and autoradiography. Since lysosomes are central to intracellular catabolic events, it is pos- sible that lysosomes are involved in intrapulmonary surfactant catabolism. Lysosomes relatively free of contaminating organelles (as determined morphologi- cally and by marker enzymes for mitochondria, endo- plasmic reticulum, peroxisomes, and plasma mem- branes) were obtained from post-lavage lung homoge- nates of 1-kg rabbits by differential centrifugation in buffered sucrose and gradient separation in percoll (density, 1.075-1.165). The role of lung lysosomes in catabolism of dipalmitoylphosphatidylcholine (DPC) was then studied in rabbits killed 4, 12, and 24 h following intratracheal injection of [3H]DPC and [“C] dihexadecyl phosphatidylcholine (DPC-ether). While equal amounts of label were in the lamellar body con- taining fractions at 4 h, nearly 6-fold more DPC-ether label than DPC label was recovered in the lysosomal fractions. By 24 h, there was 15-fold more DPC-ether in the lysosomes. This is the first report of successful isolation of lysosomes relatively free of other organ- elles from rabbit lungs. The tracer studies indicate DPC and DPC-ether follow similar intracellular proc- essing after alveolar uptake. The subsequent accumu- lation of the ether analog in the lysosomal fractions supports a role for these organelles in surfactant DPC catabolism.
Dipalmitoylphosphatidylcholine (DPC)’ is the major phos- pholipid component of lung surfactant. As recovered by al- veolar lavage procedures, it represents about 60% of the surfactant lipid by weight (1). This predominance has made DPC a useful tool for in uiuo investigations of surfactant phospholipid metabolic pathways, particularly alveolar up- take, recycling, and secretion. Resolution of surfactant phos- pholipid catabolic pathways using DPC has been more diffi- cult because DPC is rapidly degraded in adult animal lungs such that about 75% of airspace surfactant DPC is lost from the lungs within 24 h of administration (2). Dihexadecyl
* This work was supported by National Institutes of Health Grants HD-11932 and HD-93301 (to E. D. R.). The costs of publication of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked “aduertisement” in accordance with 18 U.S.C. Section 1734 solely to indicate this fact.
’ The abbreviations used are: DPC, dipalmitoylphosphatidylcho- line; DPC-ether, dihexadecyl phosphatidylcholine; Hepes, 4-(2-hy- droxyethy1)-1-piperazineethanesulfonic acid.
phosphatidylcholine (DPC-ether), a poorly hydrolyzed diether analog of DPC resistant to degradation by phospholipases AI (3) and Az (4), was previously shown to have similar alveolar uptake and recycling kinetics as synthetic DPC and surfac- tant-derived DPC despite moderate but nontoxic accumula- tion in the lung tissue (6). The resistance to phospholipase Ai additionally makes DPC-ether insensitive to the pathways involved with platelet-activating factor metabolism ( 5 ) . This analog should be useful in defining intracellular processing of surfactant DPC within the lung.
The intracellular site of surfactant degradation, likewise, has not been defined. At the subcellular level in type I1 cells, surfactant DPC secretion and recycling involve lamellar bod- ies and multivesicular bodies, respectively. These organelles have been shown to be related to lysosomes by cytochemistry (7,8) and autoradiography (9). Since lysosomes are central to general catabolic activities within cells, it seems reasonable that intrapulmonary intracellular degradation of surfactant phospholipids may involve the lysosomal pathway. Focused investigations of lung lysosomes, however, have been ham- pered by technical difficulties in isolating lysosomes free of other contaminating organelles (10). This study reports iso- lation of relatively pure fractions of lung lysosomes utilizing differential and density gradient centrifugation techniques. Intracellular surfactant phospholipid catabolism was then investigated using radiolabeled synthetic DPC and DPC-ether to determine whether lung lysosomes have a role in intrapul- monary surfactant DPC degradation.
EXPERIMENTAL PROCEDURES
Materials
All substrates and chemicals for the enzyme assays were obtained from Sigma. Percoll was from Pharmacia LKB Biotechnology Inc. Synthetic DPC and DPC-ether were prepared as before (6, 11) from the respective precursors, dipalmitoyl dimethyl phosphatidylethanol- amine and dihexadecyl dimethyl phosphatidylethanolamine, using a methylation process in the presence of potassium carbonate and 18- Crown-6 (Aldrich Chemical Co.) in benzene. Dipalmitoyl dimethyl phosphatidylethanolamine was methylated with [methyl-”Hliodide (14.4 Ci/mmol, Amersham) to yield DPC (700 Ci/mol) while dihex- adecyl dimethyl phosphatidylethanolamine was methylated with [mett~yl-’~C]iodide (56.6 Ci/mol, ICN Radiochemicals, Irvine, CA) to yield DPC-ether (20 Ci/mol). Efficiencies of radiolabel conversions were 25 and 83% for “H and “C, respectively. Pure synthetic DPC or DPC-ether was separated by thin-layer chromatography using chlo- roform/methanol/acetic acid/water (65/25/8/4, v/v). Pure unlabeled lipids were run in a parallel lane as standards of identity. All precursor lipids and pure standards were from Serdary Research Laboratories (Port Huron, MI). Each synthetic lipid contained one radiolabeled methyl group on the choline head group. Prior to preparation of liposome suspensions for intratracheal injections the purity of the labeled lipids was confirmed by repeat thin layer chromatography.
22522
Lung Lysosomes and Surfactant Catabolism 22523
Lung Lysosome Isolation and Characterization Isolation of Lung Lysosomes-Young male rabbits weighing 1.0 ?
0.1 kg were killed using intravenous pentobarbital (200 mg/kg) con- taining heparin (1 unit heparin/mg pentobarbital) followed by exsan- guination. Tracheotomy was performed, and a tracheal tube was secured for alveolar lavage. The chest was then opened, the inferior vena cava clamped, and the lungs perfused via the main pulmonary artery with cold Tris-buffered sucrose solution (0.25 M sucrose con- taining 1 mM EDTA and pH adjusted to 7.0 with Tris base). A total of 400 ml was perfused while the lungs were simultaneously gently inflated through the tracheal tube to full distension with air at a rate of 10-15 inflations per min. This maneuver greatly enhanced the clearing of blood from the lung. The lungs were then lavaged with five aliquots of cold 0.9% NaCl as previously described to recover alveolar surfactant (12). The total recovered volume from the five aliquots was, on average, 344 +- 21 ml for each animal. After the lavage, the lungs were excised and the large conducting airways stripped from the lung parenchyma.
A cell and nuclei free organelle suspension was prepared by a series of differential centrifugations as modified from the method described by Symons and Jonas for rat liver (13). The lung parenchyma was chopped to 0.5-1-mm pieces with scissors and homogenized in a Ten Broeck tissue homogenizer (10 strokes with a Dounce homogenizer “B” pestle followed by six strokes with the Ten Broeck pestle) in Tris-buffered sucrose (0.25 M sucrose, 1 mM EDTA, pH 7.0). The homogenate, suspended to an average protein concentration of 20 mg/ml, was centrifuged for 10 min at 750 X g. All centrifugations were carried out at 4 “C in a Sorvall SS-34 rotor (Du Pont-New England Nuclear). The nuclear pellet was discarded, the centrifuga- tion repeated, and the resulting supernatant was centrifuged at 20,000 X g for 10 min. The supernatant was discarded, the pellet resuspended to 35 ml in Hepes-buffered sucrose (0.25 M sucrose, 20 mM Hepes, pH 7.0), and again centrifuged for 10 min at 20,000 X g. The resulting organelle pellet was resuspended in the Hepes-buffered sucrose to an average protein concentration of 10-15 mg/ml. The organelle suspen- sion was mixed with isotonic percoll (1 ml of 2.5 M sucrose, 200 mM Hepes, pH 7.0 added to 9 ml of percoll) at a proportion of 45/55 (v/v, organelle suspension/isotonic percoll). A shallow gradient of this mixture was generated by centrifugation at 35,000 X g for 90 min. The resulting gradient was divided into 10 equal volume frac- tions. The density of each fraction was measured prior to dilution with 10 volumes of Hepes-buffered sucrose and centrifugation at 27,000 X g to remove excess percoll. The resulting pellet from each fraction was resuspended in 1 ml of Hepes-buffered sucrose and stored at -70 “C for further analysis. Each gradient fraction assayed was subjected to the same series of biochemical and enzymatic char- acterizations. In the radiolabel tracer studies aliquots of pellets and supernatants at each centrifugation process and of the gradient fractions were also obtained for measurement of radiolabel recovery.
Biochemical Characterization-Lipid was extracted from aliquots of alveolar washes, post-lavage lung homogenates, organelle pellets, and each of the gradient fractions using chloroform/methanol (2/1, v/v) (14). Aliquots of all pellets and supernatants from each centrif- ugation step were similarly extracted to determine disposition of the lung associated labeled lipid. Lipid phosphorous was quantitated from these extracts using the method of Bartlett (15). In samples from animals receiving radiolabeled phospholipids, an aliquot was also taken for measurement of radioactivity in scintillation fluid. Protein was measured spectrophotometrically using bovine serum albumin as standard (16).
Subcellular Marker Enzyme Analyses-Lysosomal arylsulfatase B (EC 3.1.6.1) activity was measured spectrophotometricallyusingpara- nitrocatecholsulfate as the substrate (17). Mitochondrial succinate dehydrogenase (EC 1.3.99.1) was assayed with 2-(para-iodophenyl)- 3-(para-nitrophenyl)-5-phenyltetrazolium chloride as the acceptor (18). Plasma membrane-associated 5’-nucleotidase (EC 3.1.3.5) activ- ity was determined by the release of inorganic phosphate from aden- osine monophosphate (19). Endoplasmic reticulum-associated glu- cose-6-phosphatase (EC 3.1.3.9) activity was measured using glucose 6-phosphate as the substrate (20). For both the 5”nucleotidase and glucose-6-phosphatase assays, sodium-potassium tartrate (12 mM) was added in the assay medium to inhibit nonspecific acid phospha- tases (21). Inorganic phosphate release was measured by the method of Chen et al. (22). Catalase (EC 1.11.1.6) was used as the peroxisome marker and assayed as previously described (23, 24). All assays were performed in substrate excess.
Electron Microscopy-Separately pooled aliquots of fractions 3 to
5 and of fractions 8 to 9 from one gradient preparation were pelleted by centrifugation, fixed in 2% glutaraldehyde/2% tannic acid in 0.085 M sodium cacodylate buffer containing 3 mM EDTA and post-fixed in 1% osmium tetroxide and 2% uranyl acetate according to Young et al. (25). The specimens were then dehydrated in a graded series of acetone and propylene oxide solutions at 4 “C prior to embedding in araldite. Thin sections (60-70 nm) were then stained in 4% aqueous uranyl acetate and lead citrate.
Recovery of Radiolabeled Tracer Lipids from Gradient Fractions
Preparation of Intratracheal Injection Solutions-Liposome sus- pensions containing [3H]DPC and [“CIDPC-ether were prepared as previously described and associated with trace amounts of unlabeled natural surfactant in 0.45% NaCl (6). The unlabeled surfactant was purified from pooled lung washes of adult rabbit lungs by a series of centrifugations through sucrose (26). The liposomes prepared in this manner were shown to behave in a fashion similar to radiolabeled surfactant (27). The final liposome suspension contained 1.5 pCi of [3H]DPC, 0.15 pCi of [14C]DPC-ether, and 0.3 pmol of total phospho- lipid in each 3-ml injection volume.
Animal Procedures-Male New Zealand White rabbits each weigh- ing 1.15 ? 0.05 kg were anesthetized with diethyl ether and injected intratracheally with the liposome suspension containing the radiola- beled lipid tracers using an 8-Fr flexible bronchoscope (2). The animals rapidly recovered, and animals were killed in groups of four to six with intravenous pentobarbital at each of four time points. The lungs were processed, and resulting gradients were characterized as described above.
Data Analysis-All enzyme activity data were expressed as specific enzyme activity per mg of protein in the assay sample. For the initial gradient characterization, values were given as group means ? S.E. for measurements made in fractions from three separate gradient preparations. Radiolabel recovery data, given as means ? S.E., were expressed as radioactivity per pmol of lipid phosphorous. Differences in label recovery among the groups were evaluated by analysis of variance followed by the Student Newman Keuls test. Significance was defined at p < 0.05.
RESULTS
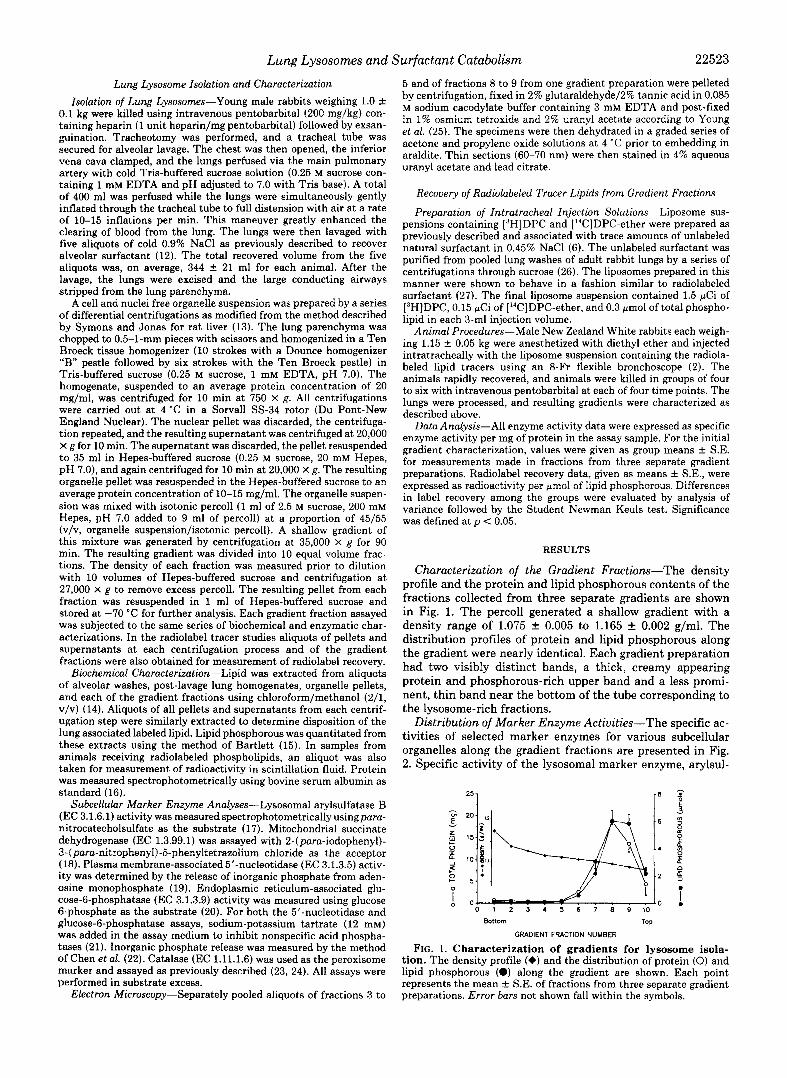
Characterization of the Gradient Fractions-The density profile and the protein and lipid phosphorous contents of the fractions collected from three separate gradients are shown in Fig. 1. The percoll generated a shallow gradient with a density range of 1.075 f 0.005 to 1.165 & 0.002 g/ml. The distribution profiles of protein and lipid phosphorous along the gradient were nearly identical. Each gradient preparation had two visibly distinct bands, a thick, creamy appearing protein and phosphorous-rich upper band and a less promi- nent, thin band near the bottom of the tube corresponding to the lysosome-rich fractions.
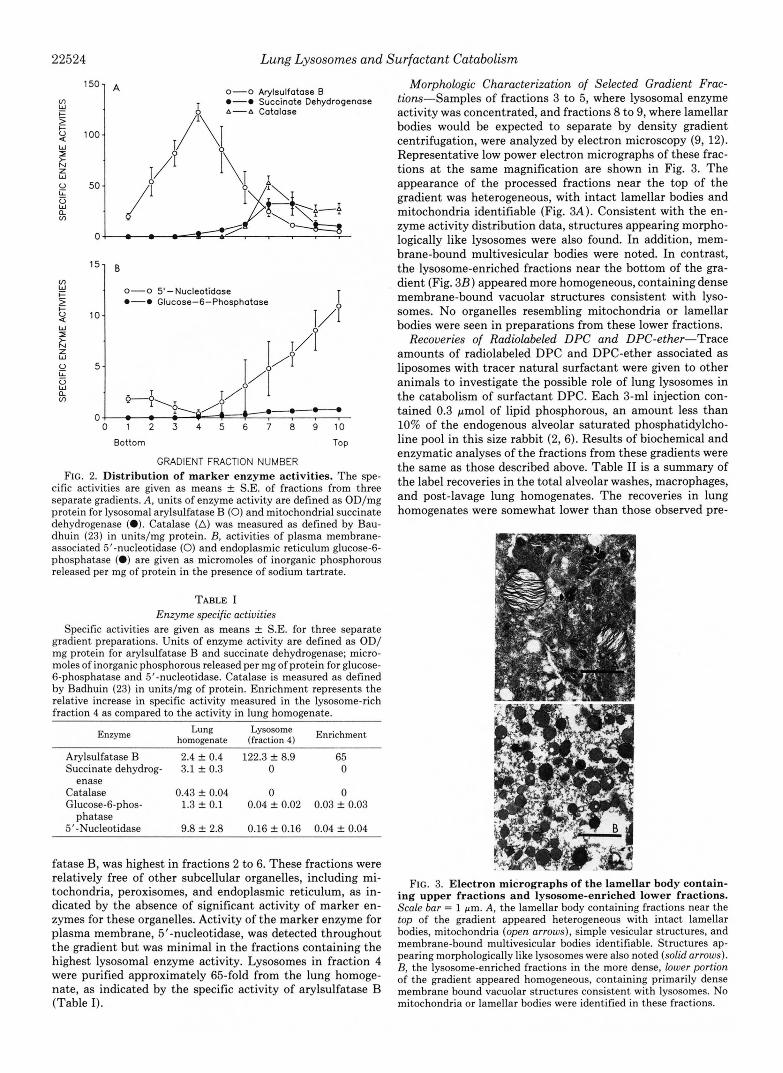
Distribution of Marker Enzyme Activities-The specific ac- tivities of selected marker enzymes for various subcellular organelles along the gradient fractions are presented in Fig. 2. Specific activity of the lysosomal marker enzyme, arylsul-
251
0
Bottom TOP
FIG. 1. Characterization of gradients for lysosome isola- tion. The density profile (e) and the distribution of protein (0) and lipid phosphorous (a) along the gradient are shown. Each point represents the mean ? S.E. of fractions from three separate gradient preparations. Error bars not shown fall within the symbols.
GRADIENT FWICTION NUMBER
22524 Lung Lysosomes and Surfactant Catabolism
1 0-0 Glucose-6-Phosohatase 0-0 5'-Nucleotidase
0 1 2 3 4 5 6 7 8 9 1 0
Bottom TOP
GRADIENT FRACTION NUMBER FIG. 2. Distribution of marker enzyme activities. The spe-
cific activities are given as means f S.E. of fractions from three separate gradients. A, units of enzyme activity are defined as OD/mg protein for lysosomal arylsulfatase B (0) and mitochondrial succinate dehydrogenase (0). Catalase (A) was measured as defined by Bau- dhuin (23) in units/mg protein. B, activities of plasma membrane- associated 5'-nucleotidase (0) and endoplasmic reticulum glucose-6- phosphatase (0) are given as micromoles of inorganic phosphorous released per mg of protein in the presence of sodium tartrate.
TABLE I Enzyme specific activities
Specific activities are given as means f S.E. for three separate gradient preparations. Units of enzyme activity are defined as OD/ mg protein for arylsulfatase B and succinate dehydrogenase; micro- moles of inorganic phosphorous released per mg of protein for glucose- 6-phosphatase and 5'-nucleotidase. Catalase is measured as defined by Badhuin (23) in units/mg of protein. Enrichment represents the relative increase in specific activity measured in the lysosome-rich fraction 4 as compared to the activity in lung homogenate.
Enzyme Lung homogenate (fraction 4) Enrichment
Lysosome
Arylsulfatase B 2.4 f 0.4 122.3 f 8.9 65 Succinate dehydrog- 3.1 f 0.3 0 0
Catalase enase
0.43 I 0.04 0 0 Glucose-6-phos- 1.3 f 0.1 0.04 f 0.02 0.03 f 0.03
5'-Nucleotidase 9.8 & 2.8 0.16 & 0.16 0.04 f 0.04 phatase
fatase B, was highest in fractions 2 to 6. These fractions were relatively free of other subcellular organelles, including mi- tochondria, peroxisomes, and endoplasmic reticulum, as in- dicated by the absence of significant activity of marker en- zymes for these organelles. Activity of the marker enzyme for plasma membrane, 5'-nucleotidase, was detected throughout the gradient but was minimal in the fractions containing the highest lysosomal enzyme activity. Lysosomes in fraction 4 were purified approximately 65-fold from the lung homoge- nate, as indicated by the specific activity of arylsulfatase B (Table I).
Morphologic Characterization of Selected Gradient Frac- tiom-Samples of fractions 3 to 5, where lysosomal enzyme activity was concentrated, and fractions 8 to 9, where lamellar bodies would be expected to separate by density gradient centrifugation, were analyzed by electron microscopy (9, 12). Representative low power electron micrographs of these frac- tions at the same magnification are shown in Fig. 3. The appearance of the processed fractions near the top of the gradient was heterogeneous, with intact lamellar bodies and mitochondria identifiable (Fig. 3A). Consistent with the en- zyme activity distribution data, structures appearing morpho- logically like lysosomes were also found. In addition, mem- brane-bound multivesicular bodies were noted. In contrast, the lysosome-enriched fractions near the bottom of the gra- dient (Fig. 323) appeared more homogeneous, containing dense membrane-bound vacuolar structures consistent with lyso- somes. No organelles resembling mitochondria or lamellar bodies were seen in preparations from these lower fractions.
Recoveries of Radiolabeled DPC and DPC-ether-Trace amounts of radiolabeled DPC and DPC-ether associated as liposomes with tracer natural surfactant were given to other animals to investigate the possible role of lung lysosomes in the catabolism of surfactant DPC. Each 3-ml injection con- tained 0.3 pmol of lipid phosphorous, an amount less than 10% of the endogenous alveolar saturated phosphatidylcho- line pool in this size rabbit (2,6). Results of biochemical and enzymatic analyses of the fractions from these gradients were the same as those described above. Table I1 is a summary of the label recoveries in the total alveolar washes, macrophages, and post-lavage lung homogenates. The recoveries in lung homogenates were somewhat lower than those observed pre-
FIG. 3. Electron micrographs of the lamellar body contain- ing upper fractions and lysosome-enriched lower fractions. Scale bar = 1 pm. A, the lamellar body containing fractions near the top of the gradient appeared heterogeneous with intact lamellar bodies, mitochondria (open arrows), simple vesicular structures, and membrane-bound multivesicular bodies identifiable. Structures ap- pearing morphologically like lysosomes were also noted (solid arrows). B, the lysosome-enriched fractions in the more dense, lower portion of the gradient appeared homogeneous, containing primarily dense membrane bound vacuolar structures consistent with lysosomes. No mitochondria or lamellar bodies were identified in these fractions.

Lung Lysosomes and Surfactant Catabolism 22525
TABLE I1 Radiolabel recovery in lipid extracts
Radiolabel recovered in the specimen as a percentage of the total amount given intratracheally, expressed as means f S.E. Each 3-ml injection suspension contained 1.5 pCi of [3H]DPC and 0.15 pCi of [14C]DPC-ether.
Time analyzed Gradients ['HIDPC ['TIDPC-ether
h 4 3 Alveolar wash" 23.1 k 3.7 35.1 f 2.0
Macrophageb 1.7 7.2 Lung homogenate 34.5 f 4.0 40.4 f 5.2
12 3 Alveolar wash 7.9 f 0.9 16.9 f 1.4 Macrophage 1.7 9.1 Lung homogenate 27.7 f 1.5 43.4 f 2.4
24 4 Alveolar wash 6.7 f 1.7 20.7 -C 2.2 Macrophage 1.5 13.7 Lung homogenate 19.1 & 3.3 36.3 +- 6.1
" Alveolar wash values include macrophage contribution. Macrophage values given as the average of two observations at
each time point.
TABLE I11 Disposition of lung-associated labeled lipids
Data are given as means f S.E., expressed as a percentage of the
Time ['HIDPC ["CIDPC-ether label recovered in the lung homogenate.
h 4 Lung homogenate 100 100
Cellular debris" 33.7 f 2.4 33.1 It 3.2 Discarded supernatantsb 41.7 f 4.0 40.2 f 5.3 Organelle pellet 14.8 f 2.1 16.0 f 3.5
12 Lung homogenate 100 100 Cellular debris 54.1 f 8.6 54.6 f 4.6 Discarded supernatants 30.0 f 6.5 15.5 f 2.7 Organelle pellet 14.0 f 1.5 16.3 f 0.7
24 Lung homogenate 100 100 Cellular debris 51.8 k 5.5 68.9 f 10.6 Discarded supernatants 34.5 & 2.9 11.6 f 0.6 Organelle pellet 14.6 k 1.3 15.4 k 4.0
" Cellular debris represent the discarded pellets from the two 750 X g centrifugation.
Discarded supernatants resulted from the two 20,000 X g centrif- ugations performed prior to obtaining the organelle pellet which was loaded on the percoll gradient.
viously (6); however, these lung homogenates were obtained by a distinctly different process. The apparent accumulation of ether-derived label in the total alveolar compartment rep- resented retention within the macrophages due to lack of degradation. When the label recoveries in the alveolar washes were corrected for macrophage contribution, no significant differences between the ester derived and ether derived labels were seen.
Table I11 is a summary of the disposition of the lung- associated label. To achieve our initial intent of recovering relatively pure lysosome preparations with minimal contam- ination from other organelles, yield was compromised, as evident by the greater than 70-85% losses during processing. The total amount of each label recovered in the final organelle pellets used for gradient separation were similar at approxi- mately 15% of the amount recovered in the corresponding lung homogenates. In pilot experiments, we found that this percentage could be increased by nearly 3-fold when the cellular debris pellet obtained from the initial 750 X g cen- trifugation was rehomogenized two more times (data not shown). This indicated that cell lysis and release of intact intracellular organelles were incomplete following the first
homogenization. The residual cellular debris from the rehomogenizations became progressively more gelatinous and difficult to separate from the solubilized organelles. This process lengthened the isolation considerably and was not used in the rest of the study as we wished to minimize the total processing time from animal sacrifice to organelle sep- aration in order to minimize ongoing degradation in isolated organelles. However, consistency of the isolation procedure was evident in the similar percentage recoveries in organelle pellets over the 24-h study period.
The patterns of label recoveries in lipid extracts of the gradient fractions from each of the various time points studied are summarized in Fig. 4. At 0 time, essentially no label was found in any of the gradient fractions (results not shown). While similar amounts of label were found in the upper fractions containing the lamellar bodies and other cellular organelles at 4 h (Fig. 4A), a nearly 6-fold accumulation of the ether label was noted in the lysosome-enriched fractions. Progressive accumulation of the [14C]DPC-ether label oc- curred in the lysosomal fractions with time as presented in Fig. 4, B and C, such that by 24 h, there was a 15-fold accumulation relative to the ['HIDPC. The ratios of the ["C] DPC-ether label to the [3H]DPC label in each gradient frac- tion at the different time points studied are shown in Fig. 40. Despite the striking retention of the ether analog within the lysosomal fractions, the ratio of the two lipids remained at unity near the top of the gradient where lamellar bodies and other organelles were noted on morphologic evaluation.
The partitioning of label recovery into water-soluble me- tabolites along the gradient is shown in Fig. 5. Label in the aqueous phase as a percentage of the total label (lipid phase + aqueous phase) contained in each fraction are presented for the lysosome-enriched and the more heterogeneous upper areas of the gradient. A greater percentage of total label was recovered as water-soluble metabolites in the denser fractions, near the bottom of the gradient as compared to the lighter fractions containing other organelles toward the top of the gradient. The pattern of label partitioning did not signifi- cantly change over the 24-h study period for either the 3H or 14C-derived label. Consistent with relatively greater resistance to phospholipase and nonenzymatic acid hydrolysis of the ether analog, at 12 and 24 h, significantly less of the I 4 C label was recovered in the aqueous phase of the lysosome-enriched fractions as compared to the 'H label derived from DPC. In contrast, less than 3% of the total label in the fractions near the top of the gradient were recovered in water-soluble me- tabolites for both 3H and I4C. While the specific identities of the labeled aqueous metabolites were not determined in this study, the presence of label in the choline headgroup of the phospholipids would make these choline-containing water- soluble metabolites such as glycerophosphocholine, phospho- rylcholine, and free choline.
DISCUSSION
There is considerable knowledge of surfactant composi- tional and functional complexity. However, an understanding of the basic mechanisms involved in surfactant catabolism remains limited. In the era of exogenous surfactant replace- ment therapy, understanding of the catabolic phenomena is important to the optimal application of this therapeutic regi- men. The cellular heterogeneity of the lung and the very rapid degradation of DPC have hampered attempts to resolve the catabolic process while investigation of intracellular events were made difficult by technical limitations in separation of organelles, such as the lysosomes, which may have a role in catabolism. The present study was designed primarily to

22526 Lung Lysosomes and Surfactant Catabolism
FIG. 4. Recoveries of DPC and DPC-ether in gradient fractions. Radioactivity measurements are given as dpm/pmol lipid phosphorous in the re- spective gradient fractions expressed as means & S.E. Error bars not shown fall within the symbols. To correct for the different activities of the label lipids in the initial suspension given to the animal (1.5 pCi of ["HIDPC (0) and 0.15 pCi of ["CIDPC-ether (O), the measured ether label recoveries were multiplied by the activity ratio of the two labels in the injection suspension. The striking obser- vation is that despite significant reten- tion of the ether within the lysosomal fractions, 5-fold a t 4 h ( A ) , 10-fold at 12 h ( B ) , and 16-fold at 24 h (C), the ratio of the lipids remained at unity in the upper fractions containing lamellar bod- ies and other cellular organelles. These ratios are shown in D.
75] A 4 hr
1 0".
' IC-DPCElhsr
0-0 3H-DPC
v x 50 a -
\ $ 1 25
751 c 24 hr
12 hr
D 0-0 4 hr 0-0 12 hr a-A 24 hr I
1 2 5 4 5 6 7 8 9 1 0 Bottom TOP Bottom
GRADIENT FRACTION NUMBER GRADIENT FRACTION NUMBER TOP
develop a technique of isolating lung lysosomes relatively free of other organelles. Using radiolabeled phospholipids associ- ated with trace amounts of natural surfactant, we also inves- tigated the possible role of lysosomes in intracellular surfac- tant phospholipid processing and degradation. Rabbits were used because of the knowledge about surfactant metabolism available in the literature for this species. Young adult rabbits weighing about 1 kg were used because this size rabbit has surfactant metabolism similar to larger adult animals and the lungs of these young animals are not obviously infected, a problem often encountered in older rabbits.
The fractionation scheme utilized in the present study generated the very shallow density gradient required in order to adequately separate lysosomes from other subcellular or- ganelles of very similar densities. An inherent problem with fractionation schemes for organelle separation is relative pu- rity. Results of the marker enzyme analyses indicated that the fractions with the peak activity for the lysosomal marker enzyme, arylsulfatase B, were relatively free of other contam- inating organelles. The mid-peak density of 1.12 was consist- ent with what had been reported for liver lysosomes (13) and fibroblast lysosomes (28). The integrity of the lysosome iso- lations were further supported by the findings on electron microscopic examination of selected fractions. No similar isolation process in lung tissue had been attempted to our knowledge. Previous attempts using other separation tech- niques to generate shallow enough gradients for adequate separation of lysosomes have not been attempted in the lung (13,28-31). Sucrose gradients yielded lysosome-enriched frac- tions with significant contamination by other organelles, in- cluding mitochondria, endoplasmic reticulum, plasma mem- branes, and peroxisomes (10). The density of the upper gra- dient fractions (1.075-1.090) where lamellar bodies were found on morphologic examination was somewhat higher than what had been observed when isolation was carried out in buffered sucrose gradients (10).
DPC-ether was used as a probe in this system to help localize the intracellular processing and degradation of DPC. Radiolabeled DPC served as a marker for movement of the naturally occurring surfactant phospholipid through the sub- cellular compartments. Previous observations (6, 34) indi- cated that these two lipids were virtually indistinguishable in
the alveolar space in that both had similar alveolar clearance kinetics, turnover times, and reutilization efficiencies despite accumulation of the ether analog within the lung tissue. This accumulation was nontoxic to the animal and presumably occurred due to inadequate degradation, primarily as a result of resistance to phospholipases AI and AS imparted by the ether bonds in place of the ester bonds of DPC. It was additionally shown that, similar to DPC, the lung was the primary site of degradation for the DPC-ether (6). These characteristics made this analog an extremely useful tool for the possible localization of intracellular surfactant DPC cat- abolic processes.
The tracer studies indicated that both DPC and DPC-ether followed similar initial intracellular routing. However, as early as 4 h following intratracheal injection, moderate accumula- tion of the ether label relative to the DPC label was found in the presumably degradative lysosome-enriched fractions. The relative accumulation was progressive over time, consistent with ineffective degradation of the ether analog with ongoing breakdown of the DPC. The lack of impact of this relative accumulation on the ratio of DPC to DPC-ether label in the upper fractions containing the lamellar bodies indicated that phospholipid within the catabolic lysosomal compartment was unable to contribute directly to the recycling process. These results were consistent with earlier findings in whole animal studies whereby accumulation of the ether analog within the lung tissue did not alter recoveries of the material from the alveolar space when macrophage contribution was accounted for (6).
Consistent with the lysosome-enriched fractions containing the degradative compartment, the recovery of labeled water- soluble choline metabolites as a fraction of total label (in lipid and aqueous phases of the extracts) was highest in the lower gradient fractions which contained the lysosomes. We did not further characterize the water-soluble metabolites in the pres- ent study. Work by other investigators using rat type I1 cells in primary culture and from isolated perfused whole lung models have shown that the water-soluble choline metabolites include glycerophosphocholine, phosphorylcholine, and free choline (32, 33). Contribution from newly biosynthesized cy- tidine 5'-diphosphate-choline, another water-soluble choline intermediate was not likely to be significant in the lower

Lung Lysosomes and Surfactant Catabolism 22527
40] A 13 4 h r
30 ffl 24 hr
20
10
0 Fractions 3-5 Fractions 8- 10
30 -1
10
0 Fractions 3-5 Fractions 8- 10 (Lysosomes)
FIG. 5 . Recoveries of [3H]DPC ( A ) and [I4C]DPC-ether ( B ) - derived water-soluble choline metabolites in the indicated gradient fractions over time. Values, given in means k S.E., are expressed as label in the aqueous phase as a percentage of the total lahel (lipid phase + aqueous phase) contained in each gradient frac- tion. A significantly greater percentage of total label was recovered in the aqueous phase in the denser lysosome-enriched fractions (num- h(>rs 9-5) as compared to the lighter fractions (numbers 8-10) near the top of the gradient containing a mixture of organelles including lamellar bodies, endoplasmic reticulum, and mitochondria ( p < 0.01 fractions 3-5 us. fractions 8-10 at all time points for both “H and “C). The pat.terns of label recoveries over time were not different for either .‘H or “C. At later times, significantly more of the [“HIDPC label partitioned into the aqueous phase in the lysosome-enriched fractions as compared to the [“CIDPC-ether label, consistent with ineffective degradation of the ether analogue ( p < 0.01, ‘’H us. ’ T at 12 and 24 h).
fractions enriched for lysosomes because these fractions con- tained little, if any, endoplasmic reticulum activity. The per- centages of total label that partitioned into the aqueous phase were very similar at the three time points studied, suggesting that the degradative capacity in these fractions remained relatively constant. Less of the ether-derived label partitioned into the aqueous phase presumably due to the relative resist- ance to degradation. Less than 3% of the total label was in the aqueous phase in the upper fractions, indicating that no significant degradation of DPC occurred in the organelles contained within these heterogenous fractions, which in- cluded endoplasmic reticulum and mitochondria.
Fig. 6 is a schematic diagram of possible intracellular traf- ficking of surfactant phospholipids following uptake from the alveolar space which would be consistent with previous obser- vations in whole animal studies and with the results of these experiments. This diagram is a modification of a previously proposed model which was adapted from the concepts of several investigators (7, 8, 34-36). Alveolar surfactant phos-
pholipids are internalized via endocytosis and form multives- icular endosomes. A fixed fraction of these multivesicular bodies then are directed to enter either the recycling pathway leading to resecretion via the lamellar bodies or the catabolic pathway involving the lysosomes, which eventually lead to degradation.
There was no direct communication of the catabolic com- partment with the recycling pathways since there were no differences in the amount of ether-associated label in the upper fractions containing the lamellar bodies despite accu- mulations in the lysosomal fractions. Although these lamellar body-containing fractions do not represent a traditional la- mellar body preparation, this further supported the earlier concept that once within the catabolic compartment, phos- pholipids re-entered the secretory pathway primarily through reincorporation of degradation products into newly synthe- sized DPC via the endoplasmic reticulum and Golgi apparatus. Moreover, the incorporation of labeled degradation products (choline and lysophosphatidylcholine) into newly synthesized DPC was relatively low since there was no relative increase in the amount of DPC label in the upper fractions containing the greatest concentration of endoplasmic reticulum as indi- cated by marker enzyme analyses. The very small recovery of aqueous degrative metabolites in the upper fractions enriched with endoplasmic reticulum and mitochondria suggest that relatively little degradative processing of DPC to yield these products occurred within these organelles. Alternatively, it is possible that the water-soluble metabolites may be cleared rapidly from these compartments, a possibility not addressed in this study.
This schema proposes the type I1 cell as the primary cellular compartment involved in the metabolic process. However, it is possible that other cells, including type I cells, may have significant roles. Because the lysosomal preparations were obtained from whole lung homogenates, the origin of the lysosomes cannot be specified. Additionally, evidence exists supporting the heterogenous nature of lysosomal bodies (7, 28). While we cannot directly address this issue in this study, the enzymatic and morphologic observations indicate such heterogeneity. Lysosomal activities found in the upper frac- tions of the gradient were consistent with the presence of morphologically identifiable lysosomal organelles in these fractions. The nature of these “lighter” lysosomes, as well as their role in the intracellular processing of the surfactant phospholipids are likely to be different from the “heavier” lysosomes obtained in the lower fractions of the gradient.
The anatomic observations of Kalina and Socher (8) sup- port the intracellular trafficking of surfactant DPC outlined above. At the ultrastructural level, using gold-labeled surfac- tant, these authors demonstrate internalization of surfactant into isolated rat type I1 cells via clathrium-coated pits, fol- lowed by deposition into electron-lucent multivesicular bod- ies. The localization of the labeled surfactant within different intracellular organelles occurred in a time-dependent fashion. At later time points, label deposition within lamellar bodies and acid phosphatase-containing organelles, such as dense multivesicular bodies and homogenous vesicles consistent with lysosomes, was observed. These observations indicate that lysosomal organelles have a role in intracellular process- ing of surfactant within the isolated type I1 cells and are consistent with the findings of the present study.
In summary, these results indicate that lysosomes relatively free of other contaminating organelles can be successfully obtained from rabbit lungs. The pattern of radiolabeled tracer phospholipid recoveries indicated that DPC-ether followed similar initial intracellular routing as DPC. The accumulation

22528 Lung Lysosomes and Surfactant Catabolism
T Y P E II CELL \ ALVEOLUS n
FIG. 6. Possible pathways of in- tracellular surfactant phosphatidyl- choline processing following alveo- lar uptake. See text for details. lyso- PC, lysophosphatidylcholine.
RETICULUM
I GOLGI
I f -
LYSO- PC CHOLINE FATTY ACIDS
Exhausted Lysosomes
D PC DPC-Ether Multiveticular Bodies Multivesiculor
" ENDOSOME
ENDOCYTOSIS
Multivesiculor
D PC A DPC- Ether retion
of the ether label in the lysosomal fractions and the recovery of water-soluble metabolites in these fractions indicate that these catabolic organelles have a role in intrapulmonary in- tracellular surfactant DPC degradation. The primary objec- tive of this study to develop a method of isolating pure isolates of lung lysosomes resulted in significant compromise of yield. The magnitude of the role of lysosomes in surfactant DPC degradation needs to be studied further in preparations where losses to processing can be minimized. The label studies further support the distinct compartmentalization and sepa- ration of the recycling and catabolic processes for surfactant DPC within the lung.
Acknowledgments-We thank Dr. Adam Jonas, with whom we had many helpful discussions, Dr. Machiko Ikegami for critical review of the manuscript, Judith Purtell for technical assistance, and Melissa Chechowitz for preparation of the electron micrographs.
REFERENCES 1. King, R. J., and Clements, J. A. (1972) Am. J. Physiol. 223,715-
2. Pettenazzo, A., Ikegami, M., Seidner, S., and Jobe, A. (1988) J.
3. Kucera, G. L., Sisson, P. J., Thomas, M. J., and Waite, M. (1988)
4. Jacobs, H., Jobe, A., Ikegami, M., Miller, D., and Jones, S. (1984)
5. Wilcox, R. W., Wykle, R. L., Schmitt, J. D., and Daniel, L. W.
6. Rider, E. D., Ikegami, M., and Jobe, A. H. (1990) J. Appl. Physiol.
7. Williams, M. C. (1984) Proc. Natl. Acacl. Sci. U. S. A. 81, 6383-
8. Kalina, M., and Socher, R. (1990). J. Histochem. Cytochem. 18,
9. Chevalier, G., and Collet, A. J . (1972) Anat. Rec. 174, 289-310 10. Hook, G. E. R., and Gilmore, L. B. (1982) J. Biol. Chem. 257,
11. Patel, K. M., and Sparrow, J. T. (1979) Lipids 14, 596-597
726
Appl. Physiol. 64, 120-127
J. Biol. Chem. 263, 1920-1928
Biochim. Biophys. Acta 793, 300-309
(1987) Lipids 22, 800-807
69, 1856-1862
6387
483-492
9211-9220
12. Jacobs, H. A,, Jobe, A., Ikegami, M., and Jones, S. (1982) J. Biol.
13. Symons, L. J., and Jonas, A. J. (1986) Anal. Biochem. 164,382-
14. Bligh, E. G., and Dyer, W. J. (1959) Can. J. Biochem. Physiol.
15. Bartlett, G. R. (1959) J. Biol. Chem. 234,466-468 16. Bradford, M. M. (1976) Anal. Biochem. 72, 248-254 17. Baum, H., Dodgson, K. S., and Spencer, B. (1959) Clin. Chim.
18. Pennington, R. J . (1961) Biochem. J. 80, 649-654 19. Hinton, R. H., Dobrota, M., Fitzsimons, J. T. R., and Reid, E.
20. Morri, D. J . (1971) Methods Enzymol. 22, 130-141 21. Dobrota, M., and Hinton, R. H. (1980) Anal. Biochem. 103,97-
22. Chen. P. S.. Jr.. Toribara. T. Y.. and Warner. H. (1956) Anal.
Chem. 257,1805-1810
390
37,911-917
Acta 4,453-455
(1970) Eur. J. Biochem. 12,349-361
102
Chem. 28; 1756-1758 '
I , .
23. Baudhuin. P.. Beaufav. H.. Rahman-Li. Y.. Sellintrer. 0. 2.. " I I
Wattiaux, R., Jacques, P., and de Duve; C. (1964) giochem. J:
24. Peters, T. J., Muller, M., and DeDuve, C. (1972) J. Exp. Med.
25. Young, S. L., Kremers, S. A., Apple, J. S., Crapo, J. D., and
26. Jobe, A., Ikegami, M., Glatz, T., Yoshida, Y., Diakomanolis, E.,
27. Ikegami, M., Jobe, A., and Duane, G. (1985) Biochim. Biophys.
28. Rome, L. H., Garvin, A. J., Allietta, M. M., and Neufeld, E. F.
29. Wattiaux, R., Wattiaux-de Coninck, S., Ronveaux-Dupal, M. F.,
30. Wolff, D. A., and Pertoft, H. (1972) Biochem. Biophys. Acta 286,
31. Pertoft, H., Warmegird, B., and Hook, M. (1978) Biochem. J .
32. Chander, A., Reicherter, J., and Fisher, A. B. (1987) J. Clin.
92,179-184
136,1117-1139
Brumley, G. W. (1981) Am. Physiol. SOC. 51, 248-253
and Padbury, J. (1981) J. Clin. Invest. 67, 370-375
Acta 835,352-359
(1979) Cell 17, 143-153
and Dubois, F. (1978) J. Cell Biol. 78, 349-368
197-204
174,309-317
Znuest. 79.1133-1138 33. Fisher, A. B., Chander, A., and Reicherter, J. (1987) Am. J .
34. Jobe. A. (1988) Sem. Perinatol. 12, 231-244 Physiol. 253, 792-796
35. Williams; M. C. (1987) Am. Reu. Respir. Dis. 135, 744-746 36. Wright, J. R., and Clements, J. A. (1987) Am. Reu. Respir. Dis.
135,426-444





























