Embed Size (px)
Citation preview

Journal of Medical Virology 5:87-100 (1980)
Characterization of HBeAg by Physicochemical and lmmunochemical Methods
David Katz, Joseph L. Melnick, and F. Blaine Hollinger
Department of Virology and Epidemiology, Baylor College of Medicine, Houston
HBeAg, which is a hepatitis B-associated antigen, was purified and characterized. The end product from ion exchange, gel filtration and immunoadsorbent col- umns consisted mainly of immunoglobulin G (IgG) molecules, identified by immunoelectrophoresis and polyacrylamide gel electrophoresis. At least four polypeptides with molecular weights of 55,000, 38,000, 25,000 and 20,000 daltons were detected in four different preparations of HBeAg. Two of these have molecular weights similar to the heavy and light chains of IgG (55,000 and 25,000 daltons). It is postulated that HBeAg may be a small ligand of 20,000 daltons with an affinity for IgG. This affinity, especially if it involves the light chain of IgG, could account for the 38,000 or 42,000 molecular weight polypeptides seen by autoradiography.
Key words: HBeAg, purification, affinity to IgG, HBsAg, hepatitis virus
INTRODUCTION
The “e system” of antigens (HBeAg) and antibodies (anti-HBe) in hepatitis B viral infection was first described in 1972 by Magnius and Espmark [ 19721 . As the system evolved, it became apparent that HBeAg held a unique relationship to hepatitis B surface antigen (HBsAg) and the hepatitis B virus (HBV). Thus, a strong correlation was found be- tween high levels of HBsAg and the presence of Dane particles [Cappel et al, 1977; Hind- man et al, 1976; Nordenfelt et al, 1975; Takahash et al, 1976; Trepo et al, 1977; Werner et al, 19771, viral DNA [Werner et al, 19771, Dane particle-associated hepatitis B core antigen (HBcAg) [Murphy et al, 1976; Okada et al, 1976; Trepo et al, 19761, and specific endogenous DNA polymerase activity [Imai et al, 1976; Nordenfelt et al, 19751 ,
More direct experimental approaches undertaken to study the nature of HBeAg have led to two opposing conclusions: 1) HBeAg is part of the HBV and presumably is coded by
Accepted for publication August 30, 1979.
David Katz is now at Israel Institute for Biological Research, Department of Virology, P.O.B. 19, Ness- Ziona, Israel.
Address reprint requests to F. Blaine Hollinger, MD, Department of Virology and Epidemiology, Baylor College of Medicine, Houston, TX 77030.
0146-661 5/80/0501-0087$02.60 0 1980 Alan R. Liss, Inc.

88 Katz, Melnick, and Hollinger
the virus. This is supported by findings that anti-HBe precipitates Dane particles and tubular forms [Neurath et al, 19761, that HBeAg is released from Dane particles by detergents [Lam et al, 19771 and that HBeAg is found in infected hepatocytes by immunofluorescence [Trkpo et al, 19761 . 2) HBeAg is an altered host protein synthesized by the host as a re- sponse to the hepatitis B infection. Neurath and Strick reported that HBeAg has properties of an immunoglobulin [Neurath and Strick, 19761, while Vyas et a1 reported an apparent association with lactate dehydrogenase isoenzyme-5 (LDH-5) [Vyas et al, 19771 .
characterization and purification of HBeAg and anti-HBe. Results indicate a close associa- tion of HBeAg with immunoglobulin G (IgC) during all purification steps and a possible affinity of the HBeAg for additional human serum proteins.
In our study, immunochemical and plysicochemical methods were employed for the
MATERIALS AND METHODS
Description of Assays
similar to that described by Van Oss and Bronson [1969]. The method was modified so that three sets of wells (six peripheral and one middle well in each set) were arranged on one agarose-coated glass microslide. In this way a total of 12 unknown samples along with six known HBeAg-positive samples could be run in one test. Each slide was then placed in the middle of a tray whose walls are equidistant from each side of the gel. After the addi- tion of the rheophoresis buffer a plastic cover containing three holes was placed over each tray in such a way that the holes were centered above the middle wells containing the anti- body.
The slides were incubated on the bench for 24 hours and transferred for further in- cubation to a humid chamber. Washed gels were dried and stained for proteins by Amido black [Uriel, 19641.
A set of reference standards for HBeAg (containing subdeterminants e and e,) and anti-HBe were obtained by assaying several HBsAg-positive plasmas. Specificity and sensi- tivity of our reagents were substantiated by comparing them with known reference re- agents supplied by Mr. B. L. Murphy from the Center for Disease Control in Phoenix, Arizona and from Dr. L. Magnius of the State Bacteriology Laboratory in Stockholm, Swe- den. Further evaluation of 26 HBeAg and anti-HBe-coded sera showed close agreement be- tween the laboratories. Plasma containing the HBeAg reference material was clarified by low speed centrifugation, distributed into vials in 2-ml amounts and kept frozen at -20°C. The anti-HBe standard was prepared from plasma by ammonium sulfate precipita- tion (33% final concentration), resuspension to one-fourth of the original volume, and dialy- sis against 0.15 M saline containing 0.01 M total phosphates and 0.1% sodium azide, pH 7.4 (PBS). This preparation was stored at -20°C in 1 ml aliquots.
HBsAg was detected either by complement futation (CF) [US Department of Health, Education, and Welfare, 19691 or by a solid phase radioimmunoassay (HepRIA, North American Biologicals, Inc., Miami, Florida). Electrophoresis on Titan Ill cellulose acetate plates, staining, and scanning procedures were performed according to the manufacturer’s instructions (Helena Laboratories, Beaumont, Texas). Immunoelectrophoresis (IEP) for detection of human serum proteins was performed using reagents and plates obtained from Behring Diagnostic (Somerville, New Jersey). Immunoglobins were quantitated by radial immunodiffusion (North American Biologicals, Inc.) according to the manufacturer’s in- structions.
HBeAg and anti-HBe were detected by a bi-dimensional immunorheophoresis method

Characterization of HBeAg 89
Extraction of HBeAg From Plasma by Polyethylene Glycol Precipitation PEG-HBeAg
Preliminary experiments were designed to determine the most convenient way to ex- tract large quantities of HBeAg from plasma while simultaneously reducing the overall pro- tein concentration. Based on our previous studies with polyethylene glycol (PEG-6000) and low pH on the differential precipitation of HBsAg from plasma [de Kizzo et al, 19721 , we determined the optimal conditions for extracting large quantities of HBe Ag from plasma. An HBsAg-positive plasma (CF titer, 1 :640) which contained HBeAg, was clarified by low speed centrifugation (3500g for 20 min) and divided into four equal portions. The pH level was adjusted to either 4.5, 6,0,7.5, or 9.0 by adding 10 N HC1 or NaOH to the magneti- cally-stirred samples. Each sample was further subdivided into five 10 ml aliquots. Poly- ethylene glycol PEG-6000) was added to yield the following final concentration (w/w): 5.0%,7.5%, 10.0%, and 12.5%. One fraction from each pH group received no polyethylene glycol. The mixtures were stirred for one hour at room temperature, centrifuged at 3500g for 20 minutes and the supernatant removed. Precipitates were dissolved in PBS to the origi- nal 10-ml volume, and suitable portions were electrophoresed on cellulose acetate strips and tested for HBeAg.
Chromatography
Rad Laboratories, Richmond, California), packed with approximately 450 ml of Bio-gel A-5 m in PBS. Fractions were collected with a LKB fraction collector (LKB Instruments, Inc., Bromma, Sweden) and monitored continuously for proteins in a Uvicord 11 (LKB) at a 280 nm wavelength. The protein content of single fractions or preparations was also estimated in a W spectrophotometer at 280 nm using El2 = 1.40 for protein mixtures and 1.35 for IgG [Layne, 19511 . Fractions were tested for HBsAg, immunoglobulins and HBeAg. When indicated, selected fractions were pooled and concentrated by vacuum dialy- sis [Peterson, 19701 prior to testing for HBeAg.
Bio-Gel A equilibrated with 0.003 M sodium citrate (pH 7.4) buffer as described by Neurath [1976]. Samples of 4-5 ml were dialyzed against the starting buffer before being applied to the column. Stepwise elution of proteins was accomplished with 0.03 M and 0.3 M so- dium citrate, pH 7.4. The effluent was continuously monitored for protein at ODzso to achieve complete elution of each peak. Fractions were examined for the presence of HBsAg, HBe Ag, and various immunoglobulins as previously described.
Biospecific adsorbents were prepared for the purification of HBeAg by two different methods: 1) cross-linking of pooled normal human serum (NHS) or anti-HBe with glutaralde- hyde to form an insoluble immunoadsorbent and 2) coupling of anti-HBe to sepharose 4B cyanogen bromide-activated beads. Care was taken to exclude any plasmas which might also contain 19 S rheumatoid factor, as determined by the latex agglutination test [Win- chester, 19761.
The NHS glutaraldehyde immunoadsorbent (Glut-NHS) was prepared according to the method of Avrameas and Ternynck [1969]. The insolubilized preparation was sequen- tially washed with 0.2 M NaC1, pH 10.9; and PBS. The anti-HBe immunoadsorbent (Glut- anti-HBe) was prepared from an ammonium sulfate precipitated anti-HBe globulin fraction which had been adsorbed against NHS (Glut-NHS) to remove any anti-NHS activity prior to cross-linking with glutaraldehyde. Both a batch-type procedure and column chroma- tography were evaluated. Cyanogen bromide-activated sepharose-4B immunoadsorbents were prepared as follows: Sixty milligrams of an anti-HBe globulin fraction or 40 mg of a
Gel filtration experiments were performed on a 2.5 X 100 cm Bio-Rad column (Bio-
Anion exchange chromatography was carried out in 1.5 X 30 cm column of DEAE

90 Katz, Melnick, and Hollinger
partially purified HBeAg preparation (see Results) were added to 10 ml of coupling buffer (0.1 M NaHC03 in 0.5 M NaC1, pH 8.0). The solutions were then added to 7 ml of the CNBr-activated sepharose 4B gel (Pharmacia Fine Chemicals, Uppsala, Sweden) which had been swollen and washed according to the manufacturer’s instructions. After coupling for 2 hours at room temperature, the gel was washed with coupling buffer and treated with 1.0 M ethanolamine, pH 8.0, for an additional 2 hours at room temperature. Four alternate wash steps were carried out with 0.1 M acetate buffer in 1.0 M NaC1, pH 4.0, and coupling buffer. Columns were prepared (6 ml bed volume) and the gel was washed further with elution buffer (0.05 M sodium borate, pH 10.9) and with 0.2 M Tris in 0.5 M NaC1, pH 7.5 (Tris-NaC1) or PBS.
eluted from the biospecific adsorbents with 0.05 M sodium borate, pH 10.9.
Polyacrylamide Gel Electrophoresis (PAGE) and Autoradiography
by Hoefer Scientific Instruments (San Francisco, California). Sodium dodecyl sulphate (SDS), mercaptoethanol and urea were used for denaturation of proteins. Samples were solubilized and boiled for 2 minutes prior to application to the gels. After the completion of the electrophoresis, the gels were fixed with 15% trichloroacetic acid, stained with Coo- massie blue for proteins, and dried in a drier from Hoefer Scientific Instruments. Other de- tails for the PAGE procedure were as previously described [Fields et al, 19771 .
T method [Greenwood et al, 19631 . Desalting after labelling was done on 0.9 X 25 cm Sephadex G75 columns (Pharmacia). Analyses of labelled proteins (precipitated in rheo- phoresis) and of labelled polypeptides (in PAGE) were done by autoradiography . Dried gels were exposed to Kodak X-ray films (NS-2T) for the appropriate period and then develop- ed. Autoradiograms from PAGE gels were scanned in a Helena Quick-scan instrument. Molec- ular weights on PAGE were determined with the aid of known protein standards as previously described [Fields et al, 19771.
Adsorption of proteins was carried out at room temperature for 16-18 hours, then
Polypeptides were analyzed by PAGE on slab gels, using the equipment manufactured
Iodination of HBeAg-positive preparations with 12’1 was performed by the chloramine-
RESULTS
PEG-6000 treatment at various concentrations and pH levels and its effect on the pre- cipitation of HBsAg, HBeAg and plasma proteins were studied and the following conclusions were drawn: Reducing the plasma pH level to 4.5 appeared to be deleterious to the HBeAg. Dialyzing the specimen against PBS before testing did not alter the results.
The optimal conditions for HBeAg enrichment were at pH 7.5 and 10% (w/w) PEG concentration. This preparation contained only 1 gram % of protein out of 6.0 gram %in the original plasma, while retaining most of the HBeAg and HBsAg activity. Albumin was reduced to 5.1% of that found in the non-PEG treated sample, while the other protein frac- tions ranged from 10% to 48.3% of their initial values. Subsequent studies showed that fibrinogen levels could be significantly reduced by recalcifying the plasma and removing the fibrin clot before PEG precipitation.
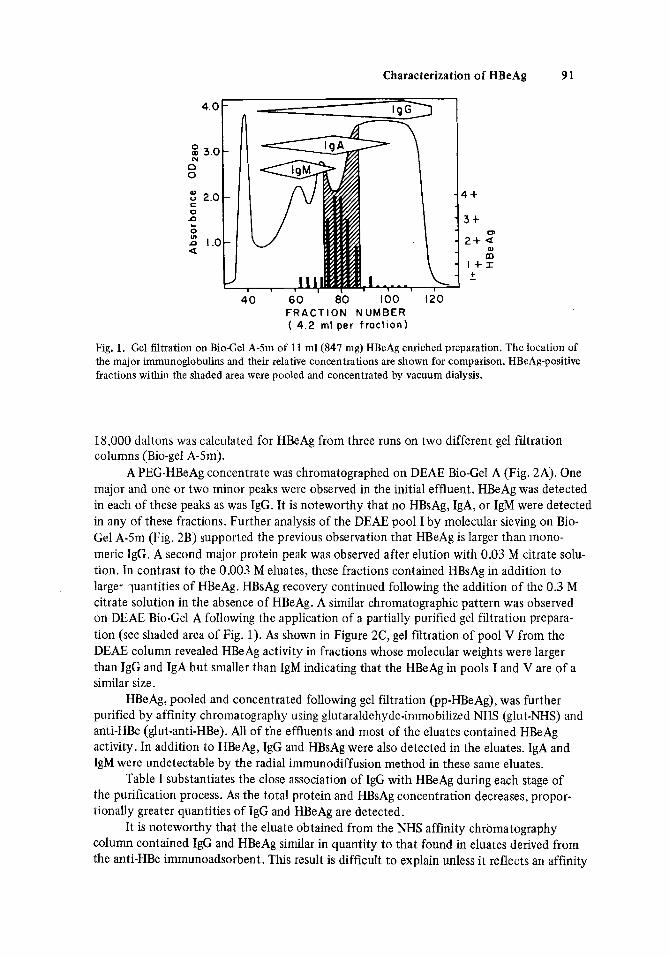
the results illustrated in Figure 1. HBeAg was distributed between the IgG and IgM peaks and approximated IgA in size. The void volume contained a substantial proportion of the HBsAg (not shown). However, detection of HBsAg was observed in all fractions suggesting considerable size heterogeneity of this molecule. A mean molecular weight of 339,000 f
Gel filtration on Bio-Gel A-5m of a PEG-concentrated preparation of HBeAg yielded
Characterization of HBeAg 91
4 +
Q) m
I + I
F R A C T I O N NUMBER ( 4 .2 ml per f ract ion)
Fig. 1. Gel filtration on BioCel A-5m of 11 ml(847 mg) HBeAg enriched preparation. The location of the major immunoglobulins and their relative concentrations are shown for comparison. HBeAg-positive fractions within the shaded area were pooled and concentrated by vacuum dialysis.
18,000 daltons was calculated for HBeAg from three runs on two different gel fdtration columns (Bio-gel A-5m).
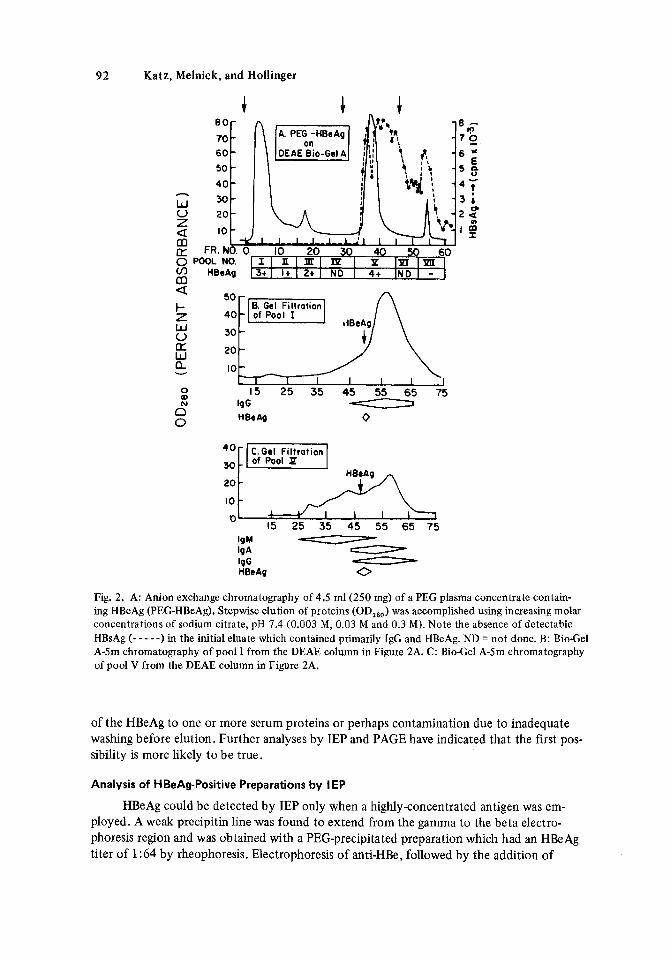
major and one or two minor peaks were observed in the initial effluent. HBeAg was detected in each of these peaks as was IgG. It is noteworthy that no HBsAg, IgA, or IgM were detected in any of these fractions. Further analysis of the DEAE pool I by molecular sieving on Bio- Gel A-5m (Fig. 2B) supported the previous observation that HBeAg is larger than mono- meric IgG. A second major protein peak was observed after elution with 0.03 M citrate solu- tion. In contrast to the 0.003 M eluates, these fractions contained HBsAg in addition to large- yantities of HBeAg. HBsAg recovery continued following the addition of the 0.3 M citrate solution in the absence of HBeAg. A similar chromatographic pattern was observed on DEAE Bio-Gel A following the application of a partially purified gel filtration prepara- tion (see shaded area of Fig. 1). As shown in Figure 2C, gel filtration of pool V from the DEAE column revealed HBeAg activity in fractions whose molecular weights were larger than IgG and IgA but smaller than IgM indicating that the HBeAg in pools I and V are of a similar size.
HBeAg, pooled and concentrated following gel filtration (pp-HBeAg), was further purified by affinity chromatography using glutaraldehyde-immobilized NHS (glut-NHS) and anti-HBe (glut-anti-HBe). All of the effluents and most of the eluates contained HBeAg activity. In addition to HBeAg, IgG and HBsAg were also detected in the eluates. IgA and IgM were undetectable by the radial immunodiffusion method in these same eluates.
the purification process. As the total protein and HBsAg concentration decreases, propor- tionally greater quantities of IgG and HBeAg are detected.
It is noteworthy that the eluate obtained from the NHS affinity chromatography column contained IgG and HBeAg similar in quantity to that found in eluates derived from the anti-HBe immunoadsorbent. This result is difficult to explain unless it reflects an affinity
A PEG-HBeAg concentrate was chromatographed on DEAE Bio-Gel A (Fig. 2A). One
Table 1 substantiates the close association of IgG with HBeAg during each stage of
92 Katz, Melnick, and Hollinger
8 0 -
70 60 50
40
30 20
10
+ - - - - - - -
t
h
w V Z
(r
a m
m g F a t- z W 0 LL: W a 0 OD N n 0
0 10 20 30 40 5 0 6 0 I l n l m l m I P IXIIrn 3t1 I + I 2 t 1 ND I 4 t IND I -
8. Gel Filtrotion
20
I I I I 10
55 65 75 I
Fig. 2. A: Anion exchange chromatography of 4.5 ml(250 mg) of a PEG plasma concentrate contain- ing HBeAg (PEG-HBeAg). Stepwise elution of proteins (OD,*,) was accomplished using increasing molar concentrations of sodium citrate, pH 7.4 (0.003 M, 0.03 M and 0.3 M). Note the absence of detectable HBsAg (- - - - -1 in the initial eluate which contained primarily I& and HBeAg. ND = not done. B: BioCel A-5m chromatography of pool I from the DEAE column in Figure 2A. C: BioCel A-5m chromatography of pool V from the DEAE column in Figure 2A.
of the HBeAg to one o r more serum proteins or perhaps contamination due to inadequate washing before elution. Further analyses by IEP and PAGE have indicated that the first pos- sibility is more likely to be true.
Analysis of HBeAgPositive Preparations by I EP
HBeAg could be detected by IEP only when a highly-concentrated antigen was em- ployed. A weak precipitin line was found to extend from the gamma to the beta electro- phoresis region and was obtained with a PEG-precipitated preparation which had an HBeAg titer of 1:64 by rheophoresis. Electrophoresis of anti-HBe, followed by the addition of

TABL
E I.
Eval
uatio
n of
Protein C
ompo
nent
s Ass
ocia
ted
With
Var
ious
Sta
ges o
f Pu
rifi
catio
n*
Prot
ein
conc
entr
atio
n H
BsA
g ac
tivity
a Ig
G
HB
eAg
activ
itya
Stag
e of
pur
ific
atio
n (m
dml)
(u
nits
per
mg
prot
ein)
(%
of
tota
l pro
tein
) (u
nits
per
mg
prot
ein)
Orig
inal
pla
sma
43.4
9,
000
12
2 PE
G p
reci
pita
tion
154
6,80
0 45
8
Gel
filt
ratio
n (p
p-H
BeA
g)
25
1,80
0 59
12
(poo
led
elua
tes
B to
E)
0.9
800
101
40
Aff
inity
chr
omat
ogra
phy
*Pro
tein
det
erm
inat
ions
wer
e m
ade
spcc
tros
copi
cally
at O
D,,,.
H
BsA
g w
as ti
trat
ed b
y R
IA; I
gG w
as d
eter
min
ed b
y ra
dial
-im
mun
odif
fusi
on;
HB
eAg
was
titr
ated
by
an im
mun
orhe
opho
resi
s tc
chni
quc.
aH
BsA
g an
d H
BeA
g ac
tivity
per
mg
of p
rote
in w
as c
ompu
ted
by d
eter
min
ing
the
num
ber
of u
nits
(ob
tain
ed b
y en
dpoi
nt d
ilutio
n) o
f an
tigen
pre
sent
in
1 m
l of
the
sam
ple
test
ed a
nd d
ivid
ing
this
num
ber
by t
he p
rote
in c
once
ntra
tion.
For
exa
mpl
e, t
he g
el f
iltra
tion
sam
ple
yiel
ded
an H
BsA
g ti
ter
of
1:9,
000
in 0
.2 m
l tes
t vol
ume.
Thus,45,000 u
nits
of
antig
en w
ere
cont
aine
d in
1 m
l. D
ivid
ing this v
olum
e by
25
mg/
ml p
rote
in c
once
ntra
tion
yiel
ds a
n H
ssA
g ac
tivity
of
1,80
0 un
its p
er m
g pr
otei
n.
94 Katz, Melnick, and Hollinger
HBeAg to the trough formed a precipitin line in the fast gamma region. Pools I, I11 and V from the DEAE column shown in Figure 2 and HBeAg-positive eluates from the NHS and anti-HBe immunoadsorbent column were electrophoresed and analyzed against gamma chain specific anti-IgG and an anti-NHS reagent. Of the three ion-exchange chromatography (DEAE) pools, pool I contained only IgG (and HBeAg) while pools I11 and V contained ad- ditional precipitin lines. Charge heterogeneity of the IgG molecules was observed in the various pools. This coincides with the elution characteristics of the pools on DEAE, especi- ally in relation to changes in the dissociating buffers. This implies that charge differences might also exist for HBeAg.
The IgG patterns associated with the eluates from the affinity columns were similar to that shown for the HBeAg-positive DEAE pool I. No precipitin line other than IgG was found in the eluate from the NHS immunoadsorbent, which supports the hypothesis that the accompanying HBeAg activity may reflect an affinity of the antigen for one or more serum proteins.
Polyacrylarnide Gel Electrophoresis (PAGE)
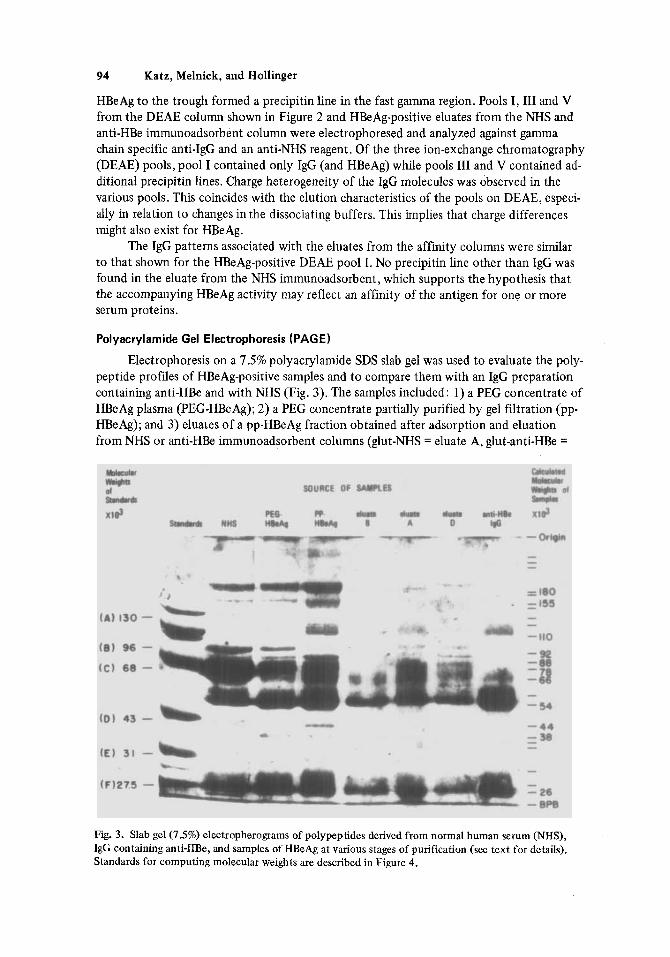
Electrophoresis on a 7.5% polyacrylamide SDS slab gel was used to evaluate the poly- peptide profiles of HBeAg-positive samples and to compare them with an IgG preparation containing anti-HBe and with NHS (Fig. 3). The samples included: 1) a PEG concentrate of HBeAg plasma (PEG-HBeAg); 2) a PEG concentrate partially purified by gel filtration (pp- HBeAg); and 3) eluates of a pp-HBeAg fraction obtained after adsorption and eluation from NHS or anti-HEk immunoadsorbent columns (glut-NHS = eluate A, glut-anti-HBe =
Fig. 3. Slab gel (7.5%) electropherograms of polypeptides derived from normal human serum (NHS), IgG containing anti-HBe, and samples of HBeAg at various stages of purification (see text for details). Standards for computing molecular weights are described in Figure 4.
Characterization of HBeAg 95
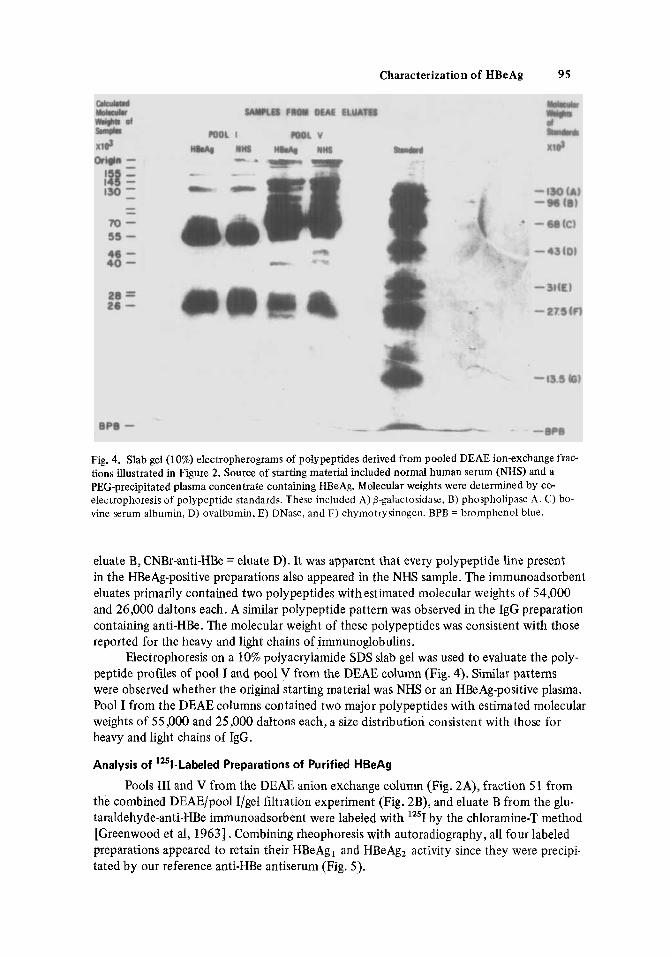
Fig. 4. Slab gel (10%) electropherograms of polypeptides derived from pooled DEAE ion-exchange frac- tions illustrated in Figure 2. Source of starting material included normal human serum (NHS) and a PEG-precipitated plasma concentrate containing HBeAg. Molecular weights were determined by co- electrophoresis of polypeptide standards. These included A) o-galactosidase, B) phospholipase A . C) bo- vine serum albumin, D) ovalbumin, E) DNase, and F) chymotrysinogen. BPB = bromphenol blue.
eluate B, CNBr-anti-HBe = eluate D). It was apparent that every polypeptide line present in the HBeAg-positive preparations also appeared in the NHS sample. The immunoadsorbent eluates primarily contained two polypeptides with estimated molecular weights o f 54,000 and 26,000 daltons each. A similar polypeptide pattern was observed in the IgG preparation containing anti-HBe. The molecular weight of these polypeptides was consistent with those reported for the heavy and light chains of immunoglobulins.
Electrophoresis on a 10% polyacrylamide SDS slab gel was used to evaluate the poly- peptide profiles of pool I and pool V from the DEAE column (Fig. 4). Similar patterns were observed whether the original starting material was NHS or an HBeAg-positive plasma. Pool I from the DEAE columns contained two major polypeptides with estimated molecular weights of 55,000 and 25,000 daltons each, a size distribution consistent with those for heavy and light chains of IgG.
Analysis of '251-Labeled Preparations of Purified HBeAg
Pools 111 and V from the DEAE anion exchange column (Fig. 2A), fraction 51 from the combined DEAE/pool I/gel filtration experiment (Fig. 2B), and eluate B from the glu- taraldehyde-anti-HBe immunoadsorbent were labeled with '''1 by the chloramine-T method [Greenwood et al, 19631. Combining rheophoresis with autoradiography, all four labeled preparations appeared to retain their HBeAg, and HBeAg, activity since they were precipi- tated by our reference anti-HBe antiserum (Fig. 5).
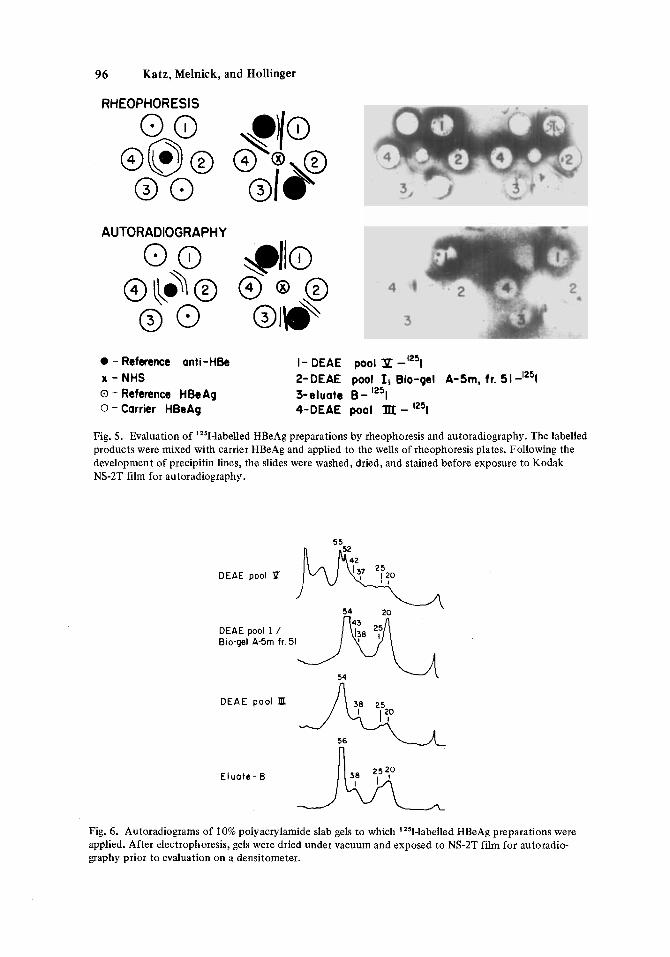
96 Katz, Melnick, and Hollinger
RHEOPHORESIS
AUTORADIOGRAPHY
0 - Reference anti-HBe I- DEAE pool P x - NHS 2-DEAE pool Ii Bio-gel A-5m, fr. 51-'251 0 - Reference HBeAg 0 - Carrier HBeAg
3-eluate B- 125t 4-DEAE pool IIX - 1251
Fig. 5. Evaluation of '251-labelled HBeAg preparations by rheophoresis and autoradiography. The labelled products were mixed with carrier HBeAg and applied to the wells of rheophoresis plates. Following the development of precipitin lines, the slides were washed, dried, and stained before exposure to Kodak NS-2T film for autoradiography.
DEAE pool I?
DEAE pool I / Bio-gel A-5m fr.
D E A E pool IE
Eluate- B
Fig. 6. Autoradiograms of 10% polyacrylamide slab gels to which '251-labelled HBeAg preparations were applied. After electrophoresis, gels were dried under vacuum and exposed to NS-2T film for autoradio- graphy prior to evaluation on a densitometer.

Characterization of HBeAg 97
The "'1-labeled preparations were analyzed on 10% PAGE under dissociating condi- tions as described in Materials and Methods. The dried gels were covered with X-ray films for one week and results were scanned with a Helena Quick-scan instrument. Molecular weights of the labeled polypeptides were estimated from unlabeled standards electro- phoresed at the same time (Fig. 6) . As expected, DEAE pool V had the largest number of polypeptides (eight) while 4 to 5 polypeptides could be seen in the other preparations. Polypeptides which were common to all four preparations had estimated molecular weights of 55,000 (p55), 38,000 (p38), 25,000 (p25), and 20,000 (p20). The p55 component is similar in size to the heavy chain of immunoglobulins. Correspondingly, the p25 and p20 polypeptides presumably represent the lambda and kappa light chains [Virella and Coelho, 19741. The association of HBeAg with an immunoglobulin is also suggested by tlus experi- ment, since 70-80% of the polypeptides from the purified HBeAg preparations have molecu- lar weight consistent with heavy and light chains.
The nature of the p38 component is not known (see Discussion). However polypep- tides of similar molecular weights (40,000) have been observed in unlabeled NHS prepara- tions (Fig. 3).
DISCUSSION
HBeAg was purified and characterized by physiocochemical and immunochemical methods. A concentrated stock was prepared by PEG-6000 precipitation of a pool of five plasmas which were positive for HBsAg and HBeAg. Gel filtration of this material revealed peak HBeAg activity in a region between the IgG and IgM peaks with a mean molecular weight of 339,000 ? 18,000 daltons. This result is in agreement with the previous findings of other authors [Fields et al, 1978; Magnius, 1975; Neurath and Strick, 19771.
Chromatography of PEG concentrates of HBeAg on DEAE Bio-gel A demonstrated considerable charge heterogeneity among HBeAg populations. The majority of antigen eluted with the 0.03 M sodium citrate, in accordance with the finding of Neurath et al[Neurath and Strick, 19761 ;however, HBeAg was also detected in the effluent containing the start- ing buffer (0.003 M citrate). It is noteworthy that this eluate of HBeAg was free of HBsAg as determined by radioimmunoassay. Net charge is one of the factors which determines elution of serum proteins from anionexchange columns, Therefore proteins which elute earlier generally migrate more slowly in an electrical field. Such a condition prevailed for IgG for the various D E W pools. Another factor in determining the elution pattern of a protein is molecular size. Larger molecules of the same net charge density per unit surface area are often eluted later. In the case of HBeAg, molecular weight determinations of the various antigenic populations were similar, and the majority of the molecules migrated with the fast moving IgG molecules. However, because of the significant charge heterogeneity of the HBeAg population, it is postulated that the molecules are sparsely distributed through- out the agarose during IEP, making detection difficult.
HBeAg was adsorbed to anti-HBe immunoadsorbents and eluted with sodium borate at pH 10.9. The eluates, whch were positive for HBeAg, contained IgG as the only other identifiable human protein. Contamination of the immunoabsorbents with IgM rheumatoid factor (RF) could conceivably produce results such as those obtained. To prevent this, all reagents were ascertained to be free of RF prior to use and then were passed over a glutar- aldehyde-immobilized NHS immunoadsorbent . Since no human proteins other than IgG were found in our purified preparations, and the amount of IgG was equal to the total amount of protein, it is believed that this result reflects an association of HBeAg with IgG.

98 Katz, Melnick, and Hollinger
The affinity of the HBeAg to glutaraldehyde polymerized NHS was demonstrated. Neurath [ 19781 found receptors of HBsAg in HBeAg-positive sera. It would be interesting to see if the HBeAg (or a small fragment of it) could be the receptor.
Purified HBeAg, eluted from immunoadsorbent or DEAE columns and analyzed by SDS-urea PAGE revealed four to eight polypeptides. The major polypeptides had molecular weights of 55,000 and 25,000 daltons, similar to those observed for the heavy and light chains of IgG. 1251-labeled preparations of HBeAg retained their immunologic activity for the HBeAg, and HBeAgz subdeterminants, as shown by rheophoresis and autoradiography. Further analysis of these labeled preparations by PAGE followed by autoradiography also showed a polypeptide composition resembling that of IgC. Polypeptides with molecular weights of 20,000 and 25,000 presumably represented kappa and lambda chains.
By totaling the molecular weights of the major polypeptides (55,000 and 25,000 dal- tons) and multiplying by 2, a value of 160,000 is obtained, which is close to the expected value for monomeric IgC. On the basis of the close association of HBeAg with IgC through a variety of purification steps, it is postulated that the HBeAg is structurally associated with a dimeric form of IgG. Since our calculated molecular weight for HBeAg is apparently 340,000 daltons rather than 320,000 expected for an IgG dimer, it is suggested that the HBeAg determinant may be in the form of a ligand of about 20,000 daltons. Such a small polypeptide would electrophorese in the region of the light chain polypeptides and could be masqueraded by them. It is also possible that some of the HBeAg determinants might remain associated to the light chains of IgG and form the 38,000 or 42,000 polypeptides seen by autoradiography . Conversely, those polypeptides may be degradation products of heavy chains. Our results, indicating an association of HBeAg with IgG, are in accordance with the finding of Neurath and Strick [1977] . Similar results were recently reported for HBeAg derived from chimpanzees [Fields et al, 19781. The discovery that HBeAg is associ- ated with lactic dehydrogenase-5 (LDH-5) [Vyas et al, 19771 and more recent reports at the viral hepatitis symposium in San Francisco [McAuiiffe and Purcell, 1978; Millman and Magnius, 19781 indicated that the HBeAg determinant is in the form of a low molecular weight ligand with an affinity for several different serum components. During the prepara- tion of this manuscript evidence was presented [Takahashi et al, 19793 indicating that the HBeAg is part of the core of Dane particles. HBeAg activity was associated with molecules of the size of 19,000 and 45,000 daltons.
Using a sensitive radioimmunoassay, Neurath [personal communication, 19781 de- termined the molecular weight of the HBeAg in a dissociating environment, as 17,000 dal- tons. These findings are supportive of our molecular weight estimation of the HBeAg de- terminant (20,000 daltons). The biological significance of the affinity of the HBeAg to IgG and presumably to other normal serum proteins is yet unknown.
ACKNOWLEDGMENTS
This work was supported, in part, by Public Health Service Grant 5 R01 A1 12,714 from the National Institute of Allergy and Infectious Diseases, National Institutes of Health.
for help in preparing the manuscript. We thank Elizabeth Constantian for excellent technical assistance and Karen Merrill

Characterization of HBeAg 99
REFERENCES
Avrameas S, Ternynck T (1969): The cross-linking of proteins with glutaraldehyde and its use for the preparation of immunoadsorbents. Immunochemistry 6:53-66.
Cappel R, de Cuyper F, Van Beers D (1977): e-Antigen and antibody, DNA polymerase, and inhibitors of DNA polymerase in acute and chronic hepatitis. Journal of Infectious Diseases 136:617-622.
de Rizzo E, Pandey R, Wallis C, Melnick JL (1972): Concentration and purification of hepatitis B anti- gen with polyethylene glycol and polyelectrolyte 60, a cross-linked copolymer of isobutylene maleic anhydride. Infection and Immunity 6:335-338.
Fields HA, Bradley DW, Davis CL, Maynard JE (1978): Purification and partial characterization of hepatitis e antigen (HBeAg). Infection and Immunity 20:792-803.
Fields HA, Hollinger FB, Desmyter J, et a1 (1977): Biochemical and biophysical properties of hepatitis B core particles derived from Dane particles and infected hepatocytes. Intervirology 8:336-350.
Greenwood FC, Hunter WM, Glover JS (1963): The preparation of I3’I-labeled human growth hormone of high specific radioactivity. Biochemistry Journal 89: 114-1 23.
Hindman SH, Gravelee CR, Murphy BL, et al (1976): “e” antigen, Dane particles, and serum DNA poly- merase activity in HBsAg carriers. Annals of Internal Medicine 85:458-460.
Imai M, Tachibana FC, Moritsugu Y, et a1 (1 976): Hepatitis B antigen-associated deoxyribonucleic acid polymerase activity and e antigen/anti-e system. Infection and Immunity 14:631-635.
Lam KC, Tong MJ, Rakela J (1977): Release of e antigen from a Dane particle-rich preparation of hepa- titis B virus. Infection and Immunity 16:403-404.
Layne E (1957): Spectrophotometric and turbidimetric methods for measuring proteins. In Colowick SP, Kaplan NO (eds): “Methods in Enzymology,” Vol3. New York: Academic Press, Inc., pp 447- 454.
Magnius LO (1975): Characterization of a new antigen antibody system associated with hepatitis B. Clinical Experimental Immunology 20: 209-2 16.
Magnius LO, Espmark JA (1972): New specificities in Australia antigen positive sera distinct from Le Bouvier determinants. Journal of Immunology 109: 1017-1021.
McAuliffe VJ, Purcell RH (1978):Current status for tests for HBeAg and anti-HBe. In Vyas GN, Cohen SN, Schmid R (eds): “Viral Hepatitis.” Philadelphia: The Franklin Institute Press, pp 161-171.
Millman I, Magnius LO (1978): Summary of workshop A d : HBeAg and anti-HBe. In Vyas GN, Cohen SN, Schmid R (eds): “Viral Hepatitis.” Philadelphia: The Franklin Institute Press, pp 665-671.
Murphy BL, Peterson JM, Smith ML, et al(1976): Correlation between fluorescent antibody detection of hepatitis B core antigen in liver biopsies and the presence of e antigen in serum. Infection and Immunity 1 3 : 296 - 297.
Hepatitis.” Philadelphia: The Franklin Institute Press, pp 183-187.
antigen has the properties of an immunoglobulin. Proceedings of the National Academy of Sci- ence 74:1702-1706.
merase associated with the hepatitis B candidate virus. Intervirology 7:356-359.
particles and the tubular forms of hepatitis B surface antigen. Journal of General Virology 30:
Neurath AR (1978): Discussion: Structure of HBeAg. In Vyas GN, Cohcn SN, Schmid R (eds): “Viral
Neurath AR, Strick N (1977): Host specificity of a serum marker for hepatitis B:Evidence that “e”
Neurath AR, Strick N (1976): Evidence against the postulated identity of e antigen with DNA poly-
Neurath AR, Trkpo C , Chen M, Prince AM (1 976): Identification of additional antigenic sites on Dane
211-285. Nordenfelt E, Kjellen L (1975): Dane particles, DNA polymerase, and e-antigen in two different cate-
Okada K, Kamiyama I, Inomata M, et al(1976): e antigen and anti-e in the serum of asymptomatic car- gories of hepatitis B antigen carriers. Intervirology 5:225-232.
rier mothers as indicators of positivc and negative transmission of hepatitis B virus to their infants. New England Journal of Medicine 294:746-749.
Biochemistry and Molecular Biology.” Amsterdam: North-Holland Publishing Co., Vol 2, pp Peterson EA (1970): Cellulosic ion exchangers. In Work TS, Work E (eds): “Laboratory Techniques in
336-368. Takahashi K, Imai M, Tsuda F, et al(1976): Association of Dane particles with e antigen in the serum
of asymptomatic carriers of hepatitis B surface antigen. Journal of Immunolow 117:102-105. Takahashi K, Akahane Y, Gotanda T, et a1 (1979): Demonstration of hepatitis B e-antigen in the core
of Dane particles. The Journal of Immunology 122:275-279.

100 Katz, Melnick, and Hollinger
Trkyo C, Bird RG, Zuckerman AJ (1 977): Correlations between the detection of e antigen or antibody and electron microscopic pattern of hepatitis B surface antigen (HBsAg) associated particles in the serum of HBsAg carriers. Journal of Clinical Pathology 30:216-220.
tions with hepatitis B surface and hepatitis core antigens, liver disease, and outcome in hepatitis B infections. Gastroenterology 71 :804-808.
Trkpo CG, Vitvitski L, Neurath R, et a1 (1976): Letter: Detection of e antigen by immunoflumescence in cyto- plasm of hepatocytes of HBsAg carriers. Lancet 1:486.
U.S. Department of Health, Education, and Welfare (1969): Palmer DF, Casey HL, Olsen JR, et a1 (eds): “A Guide to the Performance of the Standardized Diagnostic Complement Fixation Method and Adaptation to Micro Test,” 1st ed. Atlanta: USDHEW Public Health Service, National Communi- cable Disease Center.
immuno-electrophoresis in agar. In Grabar P, Burtin P (eds): “Immunoelectrophoretic Analysis.” Amsterdam, London, New York: Elsevier Publishing Co., pp 30-57.
Trkpo CG, Magnius LO, Schaefer RA, Prince AM (1975): Detection of e antigen and antibody: Correla-
Uriel J (1964): The characterization reactions of the protein constituents following electrophoresis or
Van Oss CJ, Bronson PM (1969): Immunorheophoresis. Immunochemistry 6:775-778. Virella G , Coelho IM (1974): Communications to the editors: Unexpected mobility of human lambda
chains in sodium dodecyl sulphate-polyacrylamide gel electrophoresis. Immunochemistry 11: 157-160.
Vyas GN, Peterson DL, Townsend RM, et al(1977): Hepatitis B “e” antigen: An apparent association with lactate dehydrogenase isozymed. Science 198: 1068-1070.
Werner BG, O’Connell AP, Summers J (1977): Association of e antigen with Dane particle DNA in sera from asymptomatic carriers of hepatitis B surface antigen. Proceedings of the National Academy of Science 74:2149-2151.
of Clinical Immunology.” Washington D.C.: American Society for Microbiology, pp 665-668. Winchester R (1976): Test for detection of rheumatoid factors. In Rose NR, Friedman H (eds): “Manual





























