Embed Size (px)
Citation preview

Advances in Parasitology, Volume 81 ISSN 0065-308X, http://dx.doi.org/10.1016/B978-0-12-407826-0.00004-7 133
© 2013 Elsevier Ltd.All rights reserved.
CHAPTER FOUR
G6PD Deficiency: Global Distribution, Genetic Variants and Primaquine TherapyRosalind E. Howes*,1, Katherine E. Battle*, Ari W. Satyagraha†, J. Kevin Baird‡,§, Simon I. Hay**Department of Zoology, University of Oxford, Oxford, UK†Eijkman Institute for Molecular Biology, Jakarta, Indonesia‡Eijkman-Oxford Clinical Research Unit, Jakarta, Indonesia§Centre for Tropical Medicine, Nuffield Department of Clinical Medicine, University of Oxford, Oxford, UK1Corresponding author: E-mail: [email protected]
Contents
1. Introduction� 1352. Historical�Overview� 136
2.1. Favism� 1362.2. The�Path�to�Primaquine� 1372.3. Primaquine�Tolerability�and�Safety� 140
3. Glucose-6-Phosphate�Dehydrogenase�Deficiency:�The�Enzyme�and�Its�Gene� 1433.1. G6PD�Genetics�and�Inheritance� 1433.2. The�G6PD�Enzyme� 1463.3. The�Pentose�Phosphate�Pathway�as�an�Anti-Oxidative�Defence� 1473.4. Clinical�Manifestations�of�G6PD�Deficiency� 148
4. Diagnosing�G6PD�Deficiency� 1504.1. Phenotypic�Diagnostic�Tests� 1504.2. Molecular�Diagnostic�Tests� 1524.3. The�Case�for�a�New�Diagnostic�for�Safe�P. vivax�Radical�Cure� 153
5. Mapping�the�Spatial�Distribution�of�G6PD�Deficiency� 1555.1. G6PD�Deficiency�Prevalence�Mapping� 155
5.1.1. Generating a Map: the Evidence-Base 1565.1.2. Generating a Map: the G6PD Mapping Model 1565.1.3. G6PD Deficiency Prevalence Map: an Overview 157
5.2. G6PD�Deficient�Population�Estimates� 1615.3. G6PD�Deficiency�Mutation�Mapping� 163
6. Spatial�Co-occurrence�of�G6PD�Deficiency�with�P. vivax�Endemicity� 1646.1. G6PD�Deficiency�in�Asia� 1666.2. G6PD�Deficiency�in�Asia-Pacific� 1676.3. G6PD�Deficiency�in�the�Americas� 1686.4. G6PD�Deficiency�in�Africa,�Yemen�and�Saudi�Arabia�(Africa+)� 169

R.�E.�Howes et al.134
Abstract
Glucose-6-phosphate� dehydrogenase� (G6PD)� is� a� potentially� pathogenic� inherited�enzyme� abnormality� and,� similar� to� other� human� red� blood� cell� polymorphisms,� is�particularly� prevalent� in� historically� malaria� endemic� countries.� The� spatial� extent�of� Plasmodium vivax� malaria� overlaps� widely� with� that� of� G6PD� deficiency;� unfortu-nately� the� only� drug� licensed� for� the� radical� cure� and� relapse� prevention� of� P. vivax,�primaquine,�can�trigger�severe�haemolytic�anaemia�in�G6PD�deficient�individuals.�This�chapter� reviews�the�past�and�current�data�on�this�unique�pharmacogenetic�associa-tion,�which�is�becoming�increasingly�important�as�several�nations�now�consider�strate-gies� to� eliminate� malaria� transmission� rather� than� control� its� clinical� burden.� G6PD�deficiency� is� a� highly� variable� disorder,� in� terms� of� spatial� heterogeneity� in� preva-lence�and�molecular�variants,�as�well�as� its� interactions�with�P. vivax�and�primaquine.�Consideration�of�factors� including�aspects�of�basic�physiology,�diagnosis,�and�clinical�triggers�of�primaquine-induced�haemolysis�is�required�to�assess�the�risks�and�benefits�of�applying�primaquine� in�various�geographic�and�demographic�settings.�Given�that�haemolytically�toxic�antirelapse�drugs�will�likely�be�the�only�therapeutic�options�for�the�coming�decade,�it�is�clear�that�we�need�to�understand�in�depth�G6PD�deficiency�and�
7. Evolutionary�Drivers�of�the�Distribution�of�G6PD�Deficiency� 1707.1. Evidence�of�a�Selective�Advantage� 171
7.1.1. Epidemiological Evidence 1717.1.2. In vitro Evidence 1717.1.3. Case-Control In vivo Evidence 172
7.2. Neglect�of�the�Selective�Role�of�P. vivax�as�a�Driver�of�G6PD�Deficiency� 1738. Primaquine,�P. vivax�and�G6PD�Deficiency� 174
8.1. Mechanism�of�Primaquine-Induced�Haemolysis� 1758.1.1. Primaquine and its Metabolites 1758.1.2. A Role for Oxidative Stress 1768.1.3. A Role for Methaemoglobin 1778.1.4. A Role for Altered Redox Equilibrium 1778.1.5. Significance of Primaquine-Induced Haemolysis 178
8.2. Factors�Affecting�Haemolytic�Risk� 1798.2.1. Dose Dependency 1798.2.2. Variant Dependency 1808.2.3. Red Blood Cell Age Dependency 1838.2.4. Sex Dependency 184
8.3. Predicting�Haemolytic�Risk� 1849. Towards�a�Risk�Framework�for�P. vivax�Relapse�Treatment� 185
9.1. Assessing�National-Level�Haemolytic�Risk�of�Primaquine�Therapy� 1859.1.1. Proposed Framework for Ranking National-Level Risk from G6PD Deficiency 1859.1.2. Important Limitations to Predicting National-Level Haemolytic Risk 186
9.2. Assessing�Haemolytic�Risk�at�the�Level�of�the�Individual� 18710. Conclusions� 190Acknowledgements 191References 192

G6PD�Deficiency:�Global�Distribution,�Genetic�Variants�and�Primaquine�Therapy 135
primaquine-induced�haemolysis�to�determine�safe�and�effective�therapeutic�strategies�to�overcome�this�hurdle�and�achieve�malaria�elimination.
1. INTRODUCTION
Glucose-6-phosphate dehydrogenase (G6PD) is a ubiquitously expre-ssed enzyme that has a housekeeping role in all cells, and is particularly critical to the integrity and functioning of red blood cells (RBCs). The G6PD gene has many mutant alleles which entail a decrease in enzyme activity, expressing the G6PD deficient phenotype. This trait is widespread in many human populations in whom several of the underlying mutant alleles are present at variable polymorphic frequencies.
G6PD deficiency selectively affects RBCs for two reasons. First, most known mutations cause a decreased stability of the enzyme, and since these cells do not have the ability to synthesise proteins, the enzyme level decreases as cells age during their 120 days lifespan in circulation. Second, RBCs are exquisitely susceptible to oxidative stress from exogenous oxidizing agents in the blood as well as the oxygen radicals continuously generated as hae-moglobin cycles between its deoxygenated and oxygenated forms. When G6PD activity is deficient, they have a diminished ability to withstand stress, and therefore risk destruction (haemolysis).
Fortunately, the large majority of G6PD deficient subjects have no clini-cal manifestations and the condition remains asymptomatic until they are exposed to a haemolytic trigger. For centuries, the most common known trigger of haemolysis has been fava beans, and favism remains a public health problem in areas where these are a common food item and G6PD deficiency is prevalent. However, a haemolysing trigger of great contemporary public health significance is the antimalarial primaquine, a key drug for malaria control as the only licenced treatment against (i) the relapsing liver stages of Plasmodium vivax – hypnozoites – which become dormant in infected hepa-tocytes and subsequently reactivate blood-stage infections, and (ii) the sexual blood stages of all species of Plasmodium. Since its introduction, primaquine has emerged as a major drug trigger of haemolysis in G6PD deficient indi-viduals, making this a paradigm of pharmacogenetics. This chapter focuses on the use of primaquine as a hypnozoitocide in P. vivax malaria. The com-plex problem of its use as a gametocytocide in Plasmodium falciparum malaria is not further considered here: detailed reviews of the effectiveness and safety of single-dose transmission-blocking primaquine have recently been carried out for the WHO Primaquine Evidence Review group (Recht et al., 2012) and in a Cochrane Review (Graves et al., 2012).

R.�E.�Howes et al.136
The widespread prevalence of G6PD deficiency across populations in malaria endemic areas has hindered the use of this drug, despite its uniquely useful range of therapeutic properties. One aim of studying G6PD defi-ciency is to increase access to primaquine. Optimising primaquine use involves delivery of the drug in such a way as to maintain its therapeutic activity against parasites whilst reducing risk for G6PD deficient individuals. Here, we review the knowledge base and interactions between the different component parts of this relationship (the human G6PD gene, the parasite and the drug). All of these elements must be considered when weighing the risks and benefits of putting this valuable drug to work. We examine these factors in the historical context of their development – the discovery of G6PD deficiency having been tied to the early research into primaquine, and consider the important knowledge gaps which remain despite six decades of continuous use of this drug. Extensive work was conducted in the mid-twentieth century to improve the chemotherapeutic options for treating P. vivax, particularly against its relapsing form. This chapter opens by revisit-ing those early experiments, asserting why G6PD deficiency seems to be an unavoidably major hurdle in hypnozoitocidal therapeutics. We examine what is known about G6PD – the enzyme, its mutations and population genetics. We then consider how this problem is being or could be managed in the context of contemporary targets for malaria elimination. This leads us to suggest steps to be taken to move forward into a new era when G6PD deficiency no longer seriously impedes treatment of P. vivax infection in individual patients and when primaquine can be used to its full potential as an essential tool for eliminating reservoirs of P. vivax.
2. HISTORICAL OVERVIEW
2.1. FavismAwareness of the symptoms associated with G6PD deficiency was well established long before the underlying mechanisms were understood (Beutler, 2008). The earliest suspected reports of G6PD deficiency are from Pythagoras forbidding his students to eat fava beans (Vicia faba). His strong aversion to these commonly eaten beans must mean that favism had already been recognised as a dangerous disease; and since G6PD deficiency is com-mon in Greece, it is possible that he or some of his followers may have suffered from favism, the haemolytic condition triggered by ingesting these beans (Simoons, 1998). In more recent times there has been a vast literature on favism (Fermi and Martinetti, 1905; Luisada, 1940; Meloni et al., 1983). Fava

G6PD�Deficiency:�Global�Distribution,�Genetic�Variants�and�Primaquine�Therapy 137
beans are unique among other beans because they contain high concentra-tions of two glucosides, vicine and divicine; and their respective aglycones, convicine and isouramil, are powerful triggers of oxidative stress that causes the characteristic haemolytic attacks (Chevion et al., 1982; Luzzatto, 2009).
2.2. The Path to PrimaquineAlthough methylene blue was the first synthetic compound used to suc-cessfully treat acute malaria, it was never developed or distributed. The first such drug, structurally derived from methylene blue, was the 8-ami-noquinoline pamaquine (Fig. 4.1) which became widely used, and quickly feared. While Mühlens (1926) was correct in asserting in 1926 that his laboratory’s production of pamaquine (marketed as Plasmochin) was of “huge importance” and would have “immeasurable effect on malarial countries” (trans.), his clinical trials (conducted among malaria-infected individuals of European origin) did not support his assurances about its safety. The only side-effects reported in his initial paper were some cases of cyanosis in lips, gums, tongues and fingernails, which he did not consider prohibitive to the drug’s widespread use (in retrospect, this was probably cyanosis caused by methaemoglobinaemia). At least 250 case reports of toxic reac-tions to pamaquine were subsequently published; these were cases of acute haemolytic anaemia (AHA), some resulting in death (Beutler, 1959; Hardgrove and Applebaum, 1946). In 1938, the League of Nations recom-mended against use of this drug.
In 1941, no synthetic drug effectively competed with quinine for the treatment of malaria, and quinine could not protect against relapsing malaria. The Dutch operated a global cartel on the trade of quinine, with 95% of production coming from cinchona tree plantations on Java in the
Figure 4.1 Chemical structure of key 8-aminoquinolines: pamaquine, primaquine and tafenoquine.

R.�E.�Howes et al.138
Netherlands East Indies. The fall of those holdings to the Imperial Japa-nese armed forces in early 1942 forced the Allies to use the few inferior synthetic drugs available, principally atabrine (also called mepacrine or quinacrine) and pamaquine (Elyazar et al., 2011). The embattled Ameri-cans holding out on the Bataan peninsula and Corregidor Island near Manila ( January to April 1942) suffered terribly from malaria. Condon-Rall (1992) encapsulated the significance of this, stating that the medical disaster that developed among the US troops in the Philippines was “a symptom as much as a cause of the American general military defeat”. In the region of New Guinea, 1598 American soldiers died of wounds sustained in battle, whereas 6292 perished with a diagnosis of malaria ( Joy, 1999). Similar figures were reported among Australian forces, who suffered 21,600 malaria casualties during the same campaigns. At Guadalcanal in the Solo-mon Islands during 1942, the US Army Americal division suffered malaria attack rates of 1.3/person–year (despite atabrine prophylaxis). More tell-ing, however, was what happened to this division when they were evacu-ated to nonmalarious Fiji for rest and recuperation: the malaria attack rate was 3.7/person–year, virtually all of it relapses of P. vivax (Downs et al., 1947). The US Navy estimated that 79% of the 113,774 recorded cases of malaria were relapses ( Joy, 1999).
Despite the great demand for antirelapse therapy, in 1943 the US Sur-geon General withdrew pamaquine for prevention of relapses (Office of the Surgeon General, 1943) due to its toxicity, which was highly significant in some individuals. Acute haemolytic attacks could be triggered by the use of daily pamaquine dosing (30 mg1), with associated jaundice, dark urine and weakness due to severe anaemia (Earle et al., 1948). Furthermore, when used with atabrine, its plasma levels increased 10-fold causing serious toxic-ity problems (Baird, 2011). This unexpected drug–drug interaction effec-tively removed the only therapeutic option to the serious threat of malaria relapse. The US government responded to this problem by launching one of the largest biomedical research endeavours up to that time. Created in 1943, the Board for the Coordination of Malaria Studies oversaw basic and clinical research on over 14,000 compounds for antimalarial activity (Condon-Rall, 1994). Beginning in 1944, academic clinical investigators and US armed forces clinicians set up the capacity to study induced malaria in volunteers at a number of prisons in the United States (Coatney et al., 1948), and the penitentiary at Stateville, Illinois specialised in the
1 All doses in this review are subscribed for adults weighing 40–70 kg, mean of 60 kg.

G6PD�Deficiency:�Global�Distribution,�Genetic�Variants�and�Primaquine�Therapy 139
evaluation of therapies against relapse (Comfort, 2009). They leveraged a strain of P. vivax taken from an American soldier infected in New Guinea in 1944, called the Chesson strain (Alving et al., 1948; Craige et al., 1947). In the context of the clinical trials, this strain offered the advantage of rela-tively frequent, rapid, and multiple relapses compared with Korean or North American strains, though also required higher drug dosages to achieve wholly efficacious radical cure.
A Stateville Penitentiary study protocol from 1948 states that approxi-mately 500 volunteers were involved (Alving et al., 1948) although a recent review of the overall project suggested that thousands of inmates were ultimately inoculated and included in the experiments (Comfort, 2009). The very modern and controlled prison environment provided a con-venient setting for running multiple complex protocols involving many different compounds (Alving et al., 1948), especially 8-aminoquinolines (Craige et al., 1948; Earle et al., 1948; Jones et al., 1948). The stage was thus set for careful clinical observation of the AHA induced by this class of compounds. The vetting of 24 candidate 8-aminoquinolines, a family of drugs whose antirelapse efficacy had been demonstrated by pamaquine, for safety, tolerability, and efficacy in subjects not sensitive to haemolysis had been completed by about 1949. Subsequently, the best candidate, pri-maquine (Edgcomb et al., 1950; Elderfield et al., 1955), was widely used in American soldiers in the Korean War (1950–1953). It is thus important to understand that the 8-aminoquinolines were not evaluated for optimum safety in G6PD deficient subjects. Instead, the US Army programme later strived to evaluate safety of primaquine alone in vulnerable subjects.
Studies were conducted with pamaquine to investigate the predisposing conditions which made some individuals particularly vulnerable to hae-molysis. These suggested that pamaquine sensitivity was racially correlated, being more common in subjects of African origin (6 of 76) than Caucasians (1 of 87) (Earle et al., 1948). They also found that haemolysis was unlikely to be associated with the plasma pamaquine levels. These observations led investigators to suspect a “predisposing factor” in certain individuals, triggered by the drug acting as a “precipitating factor” (Earle et al., 1948); although they found race to be the most significant predictor of haemolysis when considered alongside a range of haematological and exogenous factors, its genetic basis remained only a possibility. Indeed, it required 10 more years of investigation to zero in on that cause. Carson et al. (1956), working at the Stateville Penitentiary, described G6PD deficiency as the basis of “pri-maquine sensitivity” in 1956.

R.�E.�Howes et al.140
The newly qualified clinician, Ernest Beutler (1928–2008), went to Stat-eville and participated in the ground-breaking work characterising G6PD deficiency. The remainder of his prolific and distinguished professional life substantially advanced understanding of this important disorder (Beutler, 2009).
A Chicago University database archives approximately 150 publications arising from the Stateville Penitentiary Malaria Treatment Trials (http://www.lib.uchicago.edu/e/collections/sci/malaria.html). In this next sec-tion, we review those studies relating to G6PD deficiency and tolerance to effective primaquine dosing. While these studies have been held as a prime example of unethical human experimentation (Harcourt, 2011), their legacy still forms the foundation of P. vivax radical cure today.
2.3. Primaquine Tolerability and SafetyThe early experimental clinical studies in nondeficient individuals estab-lished that a total primaquine dose of approximately 200 mg was needed to achieve P. vivax radical cure (Alving et al., 1953; Coatney et al., 1953; Edgcomb et al., 1950). The next step was establishing an effica-cious dosing regimen with acceptable safety profiles. Most et al. (1946) described a 14-day regimen of quinine and pamaquine very early in the 8- aminoquinoline clinical development programme, which they consid-ered to be the optimum compromise between safety (3- to 7-day dosing came with high risk of intolerability or toxicity) and practicality (dos-ing up to 21 days risked poor compliance). The 14 days of daily 15 mg dosing was subsequently applied to all candidate 8-aminoquinolines for the simple reason that their therapeutic indexes were essentially similar to pamaquine. The course of haemolysis in G6PD deficient subjects, with the onset of symptoms after the third dose, permitted withdrawal of the treat-ment after relatively little exposure to the drug. Higher daily doses over a shorter duration, although equally efficacious, were considered too risky for use in unscreened patients. The 14-day regimen emerged before G6PD deficiency was known as the basis of the 8-aminoquinolines’ most severe toxicity. The compromise thus effectively included unscreened G6PD defi-cient patients. Indeed, the US Army would not screen its troops for G6PD deficiency until 2005. They considered the recommended dosing regimens not threatening, largely because withdrawal of therapy after low exposures to drug was at least possible, and even 14 days of treatment did not appear seriously harmful among the African American troops exposed to that dosage.

G6PD�Deficiency:�Global�Distribution,�Genetic�Variants�and�Primaquine�Therapy 141
Primaquine sensitivity was nonetheless a major concern during the devel-opment of primaquine (Editorial, 1952, 1955) (though did not preclude its licencing by the US Food and Drug Association [FDA] in 1952) and exten-sive studies were undertaken at Stateville to understand this problem. Prima-quine toxicity was investigated among both African American (Hockwald et al., 1952) and Caucasian–American prisoner volunteers (Clayman et al., 1952), comparing the toxicity of primaquine with pamaquine, and assessing the influence of co-administration with quinine. Follow-on experiments were carried out on sensitive individuals who had undergone haemoly-sis to investigate the severity and toxicity of multiple-drug treatments and regimens on the same individuals. Although all cases of severe haemolysis recovered without transfusion after cessation of treatment, the higher dose of 30 mg primaquine daily was deemed too dangerous for administration without close supervision: of 110 African American volunteers given 30 mg primaquine daily over 14 days, five developed severe anaemia with severity comparable with that following equivalent dosage of pamaquine, and there were 17 cases of mild anaemia. Reducing the schedule to 15 mg daily doses did not trigger any cases of severe anaemia, thus this was deemed a safe daily dose; nevertheless, 12 patients still developed mild anaemia. The relatively safe 15 mg dose among this population was corroborated by other large-scale studies (Alving et al., 1952; Hockwald et al., 1952). Parallel toxicity studies were conducted among Caucasian volunteers. As would be expected, abdominal complaints among others were also reported for high-dose regi-mens (60, 120, 240 mg daily); no symptoms were reported with 15 mg daily regimens (n = 699 Caucasian volunteers at Stateville Penitentiary), and only mild side-effects noted with the 30 mg dose. Severe haemolysis was not encountered among this group of patients, even at doses as high as 240 mg daily – this clearly contrasts with the threshold of haemolytic susceptibility in the African American volunteers.
Haemolysis in primaquine sensitive African American subjects was noticed to be self-limiting (Dern et al., 1954a). Radiochromium labelling (51Cr) used to mark sensitive and nonsensitive RBCs showed that cells maintained the same haemolytic predisposition to risk regardless of the sta-tus of the host they were transfused into (Dern et al., 1954b). Time-series data then characterised the course of the haemolysis and found a self-limit-ing pattern: in spite of continued drug administration throughout the initial acute haemolysis (which lasted about a week, at which point up to half the original cell population had haemolysed with the 30 mg/day dosage), a marked recovery and then re-establishment of equilibrium of haemoglobin

R.�E.�Howes et al.142
concentrations was observed. In one experiment, G6PD deficient subjects were given 30 mg primaquine daily for 4 months (Kellermeyer et al., 1961). Following trough haematocrits around day 7–10, these levels returned to normal within a week or so despite continued daily dosing with prima-quine. This cycle of haemoglobin levels mirrored the percentage of reticu-locytes in the blood (Dern et al., 1954a). 59Fe-labelling studies of RBCs demonstrated that only older RBCs (63–76 days old vs. 8–21 days old) were susceptible – this age-dependent phenomenon led the investigators to astutely surmise the involvement of an enzyme deficiency (Beutler et al., 1954).
The self-limiting nature of primaquine-induced haemolysis in Afri-can American volunteers, however, cannot be generalised to all G6PD deficient patients globally. Differences were noted between individuals originating from different areas. Studies of a severe G6PD variant (Medi-terranean variant) in the 1960s showed even the youngest reticulocytes to be vulnerable to primaquine (Piomelli et al., 1968). In other words, con-tinued daily dosing would result in progressively steeper losses of RBC and, presumably, death if not discontinued. The existence of these highly vulnerable variants, in contrast to the relatively nonthreatening African type (A- variant), imposes risk of fatal outcomes with the unbridled appli-cation of primaquine among populations where the character of locally prevalent G6PD variants is not known. The safety of primaquine hinges on G6PD status and primaquine sensitivity phenotype of the variant involved.
The impediments imposed by G6PD deficiency and primaquine toxic-ity in these patients still severely limit the effectiveness of this singularly important drug. Growing acknowledgement of this problem today, espe-cially in the context of emergent malaria elimination strategies, has recently activated research endeavours on this old and persistent problem. The disap-pointing search for superior alternative drugs, both in scope and outcome, that essentially ended around 1980 is detailed in Chapter 4 of Volume 80. Only one plausible successor to primaquine exists today, tafenoquine (Fig. 4.1), a GlaxoSmithKline (GSK) - Medicines for Malaria Venture (MMV) partnership drug currently in Phase IIb/III trials originally discovered and developed by the US Army (Crockett and Kain, 2007; Shanks et al., 2001). However, also being an 8-aminoquinoline, tafenoquine presents similar haemolytic challenges as primaquine to G6PD deficient individuals, and this complicates the path to licencing: tafenoquine has already been in development since the 1980s. Chapter 4, Volume 80 also lays out alternative

G6PD�Deficiency:�Global�Distribution,�Genetic�Variants�and�Primaquine�Therapy 143
and unexplored approaches to mitigating 8-aminoquinoline toxicity. The safe exclusion of patients from harm caused by this drug by the diagnosis of G6PD deficiency currently requires technical capacities nearly wholly absent where most malaria patients live, although work is ongoing to develop a practical point-of-care kit. Greater understanding of the geo-graphic distribution of G6PD deficiency in relation to endemic malaria will inform decisions on primaquine therapeutics policy and practice. The remainder of this chapter details the science and technology underpinning all of these important endeavours.
3. GLUCOSE-6-PHOSPHATE DEHYDROGENASE DEFICIENCY: THE ENZYME AND ITS GENE
The G6PD enzyme plays a critical role in maintaining RBC integrity through catalysing a key step in the cell’s metabolic production of reducing equivalents that maintain reduction–oxidation (redox) equilibrium of the cytoplasm. This protects the cell from oxidative attack by radicals derived from oxygen and organic compounds such as drugs and their metabolites. In spite of its vital function, the G6PD enzyme is highly variable, both biochemically and genetically. Detailed reviews of G6PD genetics, bio-chemistry and clinical characteristics have been previously published (Beu-tler, 1994, 1996; Cappellini and Fiorelli, 2008; Luzzatto, 2006, 2009, 2010; Mason et al., 2007; Mehta et al., 2000; WHO Working Group, 1989).
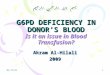
3.1. G6PD Genetics and InheritanceThe advent of molecular diagnostics following the successful mapping of the G6PD gene’s 13 exons (Martini et al., 1986) which span 18.5 kb, and the gene’s cloning and sequencing in 1986 (Persico et al., 1986; Takizawa et al., 1986) started to uncover the genetic basis to the enzyme’s great vari-ability (Vulliamy et al., 1988) (Fig. 4.2). This Mendelian X-linked gene is one of the most highly polymorphic of the human genome with at least 186 mutations having been described (Minucci et al., 2012). That said, not all mutations are polymorphic and of public health significance, but many instead appear only sporadically within populations: almost half (66 of 140 mutations reviewed in 2005 by Mason and Vulliamy) are associated with the most severe clinical phenotypes and are very rare.
Most mutations are single point substitutions (121 of 140 (Beutler and Vulliamy, 2002; Mason and Vulliamy, 2005)) leading to amino acid sub-stitutions. The absence of more severe mutations reflects the enzyme’s

R.�E.�Howes et al.144
Figure 4.2 Diversity of mutations in the G6PD gene and enzyme. Panel A shows the distribution of common mutations along the G6PD gene coding sequence. Exons are shown as open numbered boxes. Open circles are mutations causing Class II and III vari-ants; filled circles are Class I variants; filled squares are small deletions; the cross rep-resents a nonsense mutation; “f” shows a splice site mutation. (Figure from Cappellini and Fiorelli (2008), reprinted with permission from Elsevier; figure originally modified from Luzzatto and Notaro (2001)). Panel B shows the distribution of amino acid substitutions across the enzyme’s tetrameric structure (each identical monomer subunit is labelled A–D), numbered according to the affected amino acids. The diamonds indicate poly-morphic or sporadic mutations, and their colour shows the associated clinical pheno-type. The grey shadowed areas cover the two dimer interfaces. Across this region, a molecule of structural NADP per monomer is buried which stabilises the monomers and the associations between them. Each mutation is shown in only one monomer, but would be present in all four. (Figure from Mason et al. (2007), reprinted with permission from Elsevier). The positions of a few common mutations (A-, Mediterranean, Seattle, Union) are shown both in the gene (Panel A) and the enzyme (Panel B). (For a colour version of this figure, the reader is referred to the online version of this book).

G6PD�Deficiency:�Global�Distribution,�Genetic�Variants�and�Primaquine�Therapy 145
housekeeping function which requires some residual activity for cell sur-vival. Knockout studies in mice found G6PD-null mutations to be lethal (Longo et al., 2002) and a high degree of evolutionary conservation of certain regions of the gene was identified by comparing the position of mutations across 42 different organisms, pinpointing certain regions of the gene as highly conserved, and hence essential for enzyme function and cell survival (Notaro et al., 2000). All known mutations have been found to affect the coding regions of the gene and none described in the regula-tory regions (Beutler and Vulliamy, 2002; Fig. 4.2), suggesting that reduced enzyme activity levels are associated with enzyme instability, rather than deficiencies in gene expression.
The G6PD gene’s position on the X chromosome has important impli-cations for its population genetics. Unlike in males, for whom the G6PD phenotype was early-on observed to be binary with individuals being either deficient or nondeficient depending upon which allele was inherited (Beutler et al., 1955), the gene’s X-linked inheritance means that deficiency in females is more complex. Females inherit two copies of the X chromo-some and therefore have two populations of RBCs, each expressing one of the two G6PD alleles they carry. If females inherit two identical alleles (both either normal or deficient), their phenotype and clinical symptoms will be identical to those of hemizygous males. Heterozygous females, however, inherit one wild-type and one deficient allele but display a mosaic effect of expression as only one X chromosome is expressed in each cell. One population of cells will express the normal allele and the other population the deficiency (Beutler et al., 1962). The ratio of normal to deficient cells is variable, due to the phenomenon of Lyonization (Lyon, 1961). Lyoniza-tion is a random process and the resulting proportions of normal and defi-cient cells may deviate significantly from the expected 50:50 ratio (Beutler, 1994), leading some heterozygotes to have virtually normal expression, and others with expression levels comparable with female homozygotes (i.e. entirely deficient). Heterozygotes may therefore express a spectrum of phenotypes; making appropriate diagnoses with standard binary methods much harder than for deficient males, as many heterozygotes will be pheno-typically normal. At the population level, G6PD deficiency is more com-monly expressed in males, though in populations with high frequencies of deficiency, homozygotic inheritance can be common, and the preva-lence of affected heterozygotes may also be of public health concern. More details about the population genetics of the G6PD gene are discussed by Hedrick (2011).

R.�E.�Howes et al.146
3.2. The G6PD EnzymeThe G6PD enzyme consists of either dimer or tetramer forms of a protein subunit consisting of 514 amino acids. Each subunit binds to an NADP+ molecule for its structural stability, which are positioned close to the interface where the two subunits of each dimer bind (Au et al., 2000; Fig. 4.2). The majority of mutations disrupt the enzyme structural stability and thus reduce its overall activity. The effect of each mutation on enzyme structure and func-tion depends on the location of the substituted amino acid. For example, many of the most severe mutations map to exon 10 (Mehta et al., 2000) which encodes the binding interface of the subunits and therefore disrupt its qua-ternary structure and stability. These mutations cause the most severe clinical symptoms and as such do not reach polymorphic frequencies; instead they usually result from independent spontaneous mutations (Fiorelli et al., 2000). Mutations which do not cause such severe reductions in enzyme activity are widely distributed across the gene’s coding region and throughout the enzyme structure (Fig. 4.2), and have been found to reduce the efficacy of protein folding, for example (Gomez-Gallego et al. (1996)). The residual enzyme activity of G6PD variants ranges from <1% to 100%.
As with all enzymes, G6PD activity decreases with cell age: it is esti-mated that in normal blood, reticulocytes have about five times higher activity levels than the oldest 10% of RBCs (Luzzatto, 2006). The oldest cells are therefore most vulnerable to oxidative stress. In individuals with intrinsically reduced G6PD enzyme activity due to genetic mutations, the ageing process is effectively sped up, with larger proportions of cells having lower enzyme levels and being at increased risk of oxidative damage. This has implications for the clinical severity of the mutations, as discussed below. The properties of these enzyme variants correspond to a broad spectrum of enzyme biochemical phenotypes, i.e. electrophoretic properties, heat stabil-ity and enzyme kinetics. WHO guidelines (WHO Working Group, 1989) for standardised biochemical characterisation of the enzyme led to 387 variants of G6PD being described by 1990 (Beutler, 1990), though many of these would later prove to be genetic duplicates.
Few variants have been fully characterised, but a handful of those which are common enough to be of major public health significance have been well researched. The residual enzyme activity of these variants affects cell primaquine susceptibility. Though it is widely accepted, these must be inversely correlated; little evidence informs that presumption (Baird and Surjadjaja, 2011). Three such variants have formed the basis to current drug recommendations, which are further discussed in Section 8.2 (p. 179).

G6PD�Deficiency:�Global�Distribution,�Genetic�Variants�and�Primaquine�Therapy 147
All the earliest evidence about the haemolytic risk of G6PD defi-ciency pertained to the African A- variant (G202A/A376G), due to the racial background of the “primaquine sensitive” patients studied in the 1950s Stateville primaquine experiments (Section 2, p. 136). Although rare as a genetic variant for having a double-point mutation, this type of deficiency is very common among individuals of sub-Saharan African origin. The A- variant characteristically expresses residual enzyme activity about 10% of normal levels (Beutler, 1991). It was studies with this variant which led to the discovery of G6PD deficiency (Carson et al., 1956). The Mahidol variant (G487A) is the predominant allele among many G6PD deficient populations of Myanmar and is also common among Thais (Section 5.3, p. 163 and Fig. 4.6A). Enzyme activity is reduced to 5–32% of normal levels (Louicharoen et al., 2009). Finally, the Mediterranean variant (C563T) was originally known for its association with the clinical pathology of favism, and causes some of the most severely deficient phenotypes (Beutler and Duparc, 2007). This variant usually expresses <1% enzyme activity, with undetectable enzyme levels in older erythrocytes (Piomelli et al., 1968). Despite expressing such low levels of enzyme activity, carriers of this muta-tion are nevertheless asymptomatic until exposed to haemolytic triggers (Beutler, 1991) (Section 3.4, p. 148).
3.3. The Pentose Phosphate Pathway as an Anti-Oxidative Defence
G6PD enzyme activity is necessary for RBC survival as it catalyses the only metabolic pathway capable of generating reducing power to these cells lacking mitochondria (Pandolfi et al., 1995). Reducing power, supplied in the form of NADPH, is necessary as an electron donor, i.e. chemical reduc-tion, for detoxifying oxidative challenges to cells. The metabolic reactions concerned are part of the pentose phosphate pathway (PPP, also called the hexose monophosphate shunt), the first and rate-limiting step of which is catalysed by the G6PD enzyme: the oxidation of glucose-6-phosphate into 6-phosphoglucono-δ-lactone, which simultaneously reduces NADP to NADPH. The electron of NADPH passes to abundant glutathione dimers (GSSG) via another enzyme, glutathione reductase. Reduced glutathione monomers (GSH) represent the primary defence against hydrogen perox-ides, organic peroxidises, and free radicals. When G6PD functions normally, the drain of electrons from the NADPH pool caused by oxidative challenge within the cell prompts the PPP to accelerate according to need, i.e. main-taining an NADP–NADPH equilibrium that strongly favours NADPH.

R.�E.�Howes et al.148
This in turn maintains the oxidised–reduced glutathione (GSSG–2GSH) equilibrium strongly in the direction of the reduced state, i.e. 1:500 at steady state (Greene, 1993).
However, in cells that have a mutant and defective G6PD gene, the PPP may, depending upon the extent of the enzyme activity defect, function at near-maximum rate even at steady-state redox equilibrium. When oxida-tive challenge occurs and the equilibrium of NADP to NADPH shifts to the oxidised direction, the PPP is intrinsically unable to accelerate rapidly enough to force the equilibrium in favour of NADPH. This effectively stymies the flow of electrons to GSH, and that equilibrium shifts in favour of GSSG. The oxidants consuming these reducing equivalents, in turn, over-whelm the ability of the cell to provide them and damage may then occur. Visible evidence of such occurs in the form of Heinz bodies in the RBC membrane that attend acute primaquine-induced haemolytic anaemia (Greene, 1993). Heinz bodies cause the membrane to become rigid, and thus decrease the cells’ lifespans. The mechanism of primaquine-induced haemolysis remains uncertain, but will be discussed in more detail later in this chapter (Section 8.1, p. 175).
3.4. Clinical Manifestations of G6PD DeficiencyIn discussing the clinical manifestations, it is important to note that the majority of G6PD deficient individuals are asymptomatic most of the time. The public health importance of this condition comes from the sheer num-bers affected and at potential risk of developing clinical symptoms: 400 mil-lion globally (Cappellini and Fiorelli, 2008), or 350 million within malaria endemic countries (Howes et al., 2012).
Symptoms are induced when cells are exposed to exogenous oxidative stresses against which they cannot defend themselves. The severity of the clinical symptoms and the subsequent treatment required depends upon the degree of enzyme deficiency (which is variant-dependent), the nature and total dose of the oxidative agent, the time course of exposure, the presence of additional oxidative stresses and pre-existing factors such as age, hae-moglobin concentration and concurrent infection (Cappellini and Fiorelli, 2008). The relative contribution of each to determining the severity of the response is not fully known but is further discussed in Section 8.2, p. 179).
The most clinically serious symptom of G6PD deficiency is neonatal jaundice (NNJ), which peaks 2 to 3 days after birth (Luzzatto, 2010). This is highly variable in severity but can lead to kernicterus (Beutler, 2008) and permanent neurological damage or death if left untreated (Doxiadis and

G6PD�Deficiency:�Global�Distribution,�Genetic�Variants�and�Primaquine�Therapy 149
Valaes, 1964; Luzzatto, 2006). Not all neonates with NNJ are G6PD defi-cient, but this congenital condition greatly increases the risks, and in some countries is the most common cause of NNJ (Luzzatto, 2010).
AHA is the most common manifestation of the deficiency, and may be triggered by a range of exogenous agents causing intravascular haemoly-sis and jaundice, and may include haemoglobinuria (dark urine) (Luzzatto, 2009). The most severe outcome of AHA is acute renal failure (Cappel-lini and Fiorelli, 2008). The longest-known of these triggers are fava beans: “favism” can be very severe or even life-threatening if left untreated with-out transfusion (Beutler, 2008; Luisada, 1941); favism is most common in children. Infection is another important trigger of AHA (Burka et al., 1966), with severe pathology having been previously attributed to hepatitis viruses A and B, cytomegalovirus, pneumonia, and typhoid fever (Cappellini and Fiorelli, 2008). Finally, a number of haemolysis-inducing drugs have also been identified as triggers of AHA (Youngster et al., 2010); in the present context of P. vivax malaria, the most pertinent is primaquine.
The exceptions to G6PD deficiency being asymptomatic until trig-gered by certain exogenous triggers are those sporadically emerging, highly unstable variants expressing very low residual enzyme activity. These variants never reach polymorphic frequencies due to their severe pathology, which is characterised as chronic nonspherocytic haemolytic anaemia (CNSHA). While individuals with these mutations make up only a very small minor-ity of the population affected by G6PD deficiency (almost always males), they are the most clinically severe and may be transfusion-dependent (Luzzatto, 2010). In addition to susceptibility to all the aforementioned trig-gers of AHA, the very low residual enzyme levels mean that cells cannot even protect themselves against oxygen radicals continuously generated by the on-going process of haemoglobin de-oxygenation. CNSHA is therefore a lifelong condition, with haemolysis ongoing even in steady state.
Based on these pathologies, G6PD alleles can be categorised into three types: (1) those sporadic severe variants associated with chronic symptoms, (2) polymorphic types which are typically asymptomatic but susceptible to trigger-induced acute haemolytic episodes, and (3) those with normal activ-ity (Table 4.1). Previous classifications have included additional subdivisions of the polymorphic variants into “mild” and “severe” types (WHO Work-ing Group, 1989; Yoshida et al., 1971). However, as suggested by Luzzatto, the distinctions between these further classes are blurred and are no longer useful (Luzzatto, 2009). As such, we distinguish only three variant types (Table 4.1).

R.�E.�Howes et al.150
In the present context of P. vivax therapy, the generally asymptomatic polymorphic variants vulnerable to AHA (type 2 variants) are the primary threat to safe therapy. These variants are the subject of this review. Prima-quine-induced haemolysis is further discussed later in this chapter (Sec-tion 8.1, p. 175), but can require transfusion even after relatively low doses (Shekalaghe et al., 2010). Identifying this risk is therefore essential.
4. DIAGNOSING G6PD DEFICIENCY
Given the absence of a universally safe drug and the potential severity of primaquine-induced AHA, widespread safe radical treatment of P. vivax is contingent upon reliable diagnosis of G6PD deficiency. There are two types of tests for diagnosing G6PD deficiency: biochemical enzyme activity tests and molecular DNA-based methods. These are suited to different situations, depending upon the type of diagnosis required and the laboratory capaci-ties available. We discuss here those currently available and consider their limitations in respect to the heterogeneity of this condition, then discuss on-going developments towards improving these methods.
4.1. Phenotypic Diagnostic TestsMost tests for G6PD deficiency consider the biochemical phenotype – qualitative or quantitative measures of residual enzyme activity. These tend to use dyes or fluorescence markers serving as direct or proxy indicators of enzyme activity representing the rate of NADP reduction to NADPH (Beutler, 1994); qualitative assessments generally allow classification as nor-mal, intermediate, or deficient, while quantitative tests employ spectropho-tometry to determine exact measures of enzyme activity. One of the earliest screening methods was developed by Motulsky and Campbell–Kraut, using the rate of brilliant cresyl blue decolourisation: if decolourisation had not occurred within a predetermined timeframe (commonly 180 min), the
Table 4.1 G6PD Variant Types and Their Key Characteristics
TypeResidual Enzyme Activity Prevalence Clinical Significance
1 <10% Sporadic, never polymorphic
Severe and chronic: CNSHA
2 1–50% Polymorphic Asymptomatic until triggered: risk of NNJ, AHA, favism
3 Normal (>50%) Polymorphic (wild-type) None

G6PD�Deficiency:�Global�Distribution,�Genetic�Variants�and�Primaquine�Therapy 151
sample was deemed to be G6PD deficient. Developments to this method have included Brewer’s methaemoglobin (metHb) reduction test (Brewer et al., 1962), Bernstein’s DPIP method (2,6-dicholorophenol indophenol dye test (Bernstein, 1962)), Beutler’s fluorescent spot test (FST) (Beutler and Mitchell, 1968; which is the WHO recommended method (Beutler et al., 1979)), and most recently the WST-8/1-methoxy phenazine methosulfate (PMS) method (Tantular and Kawamoto, 2003) created to facilitate field use of the diagnostic.
Applying these diagnostic tests in large-scale population screening is typically feasible in the context of research endeavours. However, such test-ing is very often impracticable for routine care in those settings where most malaria patients live. Most methods still depend on a cold chain and spe-cialised laboratory equipment. Even the FST, perhaps the most widely used test, requires cold storage of reagents, micropipetters, a water bath and a UV light source. Further practical limitations include the time-delay on obtain-ing the results, which may take several hours, and the difficulties of reading the results – naked eye judgements have been found to be subjective with some of the colour changes, which are also influenced by room tempera-ture and humidity. Furthermore, although standardised protocols for using the most common methods were published in 1979, tests are invariably modified by users (for example, in terms of the cut-off times imposed) and adapted to local conditions and survey constraints. This diagnostic varia-tion, therefore, hinders comparative analyses between surveys by adding a level of uncertainty into the results. Furthermore, anaemia is an important potentially confounding factor increasing the probability of false-positive diagnoses. This condition reduces the overall number of RBCs, and there-fore the level of G6PD enzyme per volume of blood. Conversely, increased proportions of reticulocytes following malaria infection can lead to false negatives as reticulocytes have the highest G6PD activity levels. The influ-ence of malaria parasites on qualitative tests has not been evaluated.
The most important limitation to these qualitative tests, however, is their relatively poor ability to diagnose female G6PD deficient heterozygotes. As previously explained (Section 3.2, p. 146), heterozygotes express a mosaic of two RBC populations: cells expressing the normal G6PD gene and cells with the deficiency. The deficient cells carry exactly the same haemolytic risk as those of homo- or hemizygotic individuals. Heterozygote diagnostic outcome is dependent upon: (i) the diagnostic test’s threshold for deter-mining deficiency (the longer the delay, the larger the proportion of sam-ples that will appear “normal”); (ii) the mutation and the level of residual

R.�E.�Howes et al.152
enzyme activity expressed; and (iii) the ratio of normal to deficient cells – determined by Lyonization, as previously described. These issues present serious problems to heterozygote diagnosis, as the normal enzyme expres-sion in one population of cells can mask serious deficiency in others. A heterozygote may have, for example, 70% of her RBCs expressing very low enzyme activity, and therefore exquisitely sensitive to primaquine and vulnerable to harm, but she may test as “normal”.
Although some biochemical tests have been found to be better suited to detecting heterozygosity, including G6PD/6-phosphogluconate- dehydrogenase (6PDG) and G6PD/pyruvate kinase (PK) ratio analysis, and the cytochemical G6PD straining assay (Minucci et al., 2009; Peters and Van Noorden, 2009), these are highly technically challenging and therefore impractical for large-scale, field-based population surveys, and rarely used. Molecular methods, on the other hand, provide an unambiguous diagnosis of genetic heterozygotes. A recently described flow cytometric assay (Shah et al., 2012) appears much more practical, albeit still limited to the setting of relatively sophisticated laboratories.
Haunting all of these methods is the important question of what repre-sents an acceptable level of enzyme activity with respect to risk of prima-quine-induced haemolysis. In other words, at what level of residual activity does each test classify patients as normal, and does this genuinely exclude all at risk of harm? This is the issue of sensitivity. Specificity poses the converse problem by excluding patients who could safely receive effective treatment of their infection. A clinically appropriate sensitivity/specificity balance must be incorporated into the diagnostics. Unfortunately, very little evi-dence regarding primaquine sensitivity phenotypes informs this decision across the broad spectrum of mutant enzymes.
4.2. Molecular Diagnostic TestsA seemingly more clear-cut diagnostic approach examines the gene itself. Molecular methods use variant-specific primers to identify the presence or absence of specific mutations. These direct methods overcome uncertainties associated with variable enzyme activity cut-offs, anaemia, reagent break-down and subjective classifications. Molecular diagnoses allow insight into the severity of the condition for those mutations for which residual enzyme level phenotypes and primaquine sensitivity phenotypes are known. Female heterozygotes will not be dangerously misclassified as G6PD normal.
Irrespective of these benefits, the high-end laboratory requirements and time lags for results leave these methods impracticable for population

G6PD�Deficiency:�Global�Distribution,�Genetic�Variants�and�Primaquine�Therapy 153
screening or routine care in their current form. Even if these limitations could be overcome, deeper limitations remain. Although heterozygosity can be diagnosed, the state of Lyonization, and thus the phenotype remains uncertain, potentially putting individuals with these intermediate genotypes at risk. For instance, a study in Tanzania reported a case of severe haemolysis (defined as <5 g/dl) in a heterozygote A- variant female child following a single dose of primaquine (45 mg dose) (Shekalaghe et al., 2010). Further, although the enzyme activity of three major variants has been characterised (Mediterranean, A-, Mahidol (Baird and Surjadjaja, 2011)), the clinical char-acter of the vast majority remain unknown; in these cases, molecular diag-noses cannot inform clinical severity. Even in the well-studied A- genotype, enzyme activity assays in Uganda identified surprising variation between individuals of the same genotypes (Johnson et al., 2009). Finally, molecular methods look for specific mutations, which is usually only a subset of all described mutations, and hence cannot reliably diagnose all cases of poten-tially clinically significant deficiency. For example, the Tanzanian study pre-viously referred to used primers to identify the common A (A376G) and A- (G202A) mutations, but then reported instances of haemolysis in apparent wild-type B individuals (Shekalaghe et al., 2010). It seems plausible, given greater heterogeneity in the pool of G6PD deficient variants reported from individuals in The Gambia (Clark et al., 2009) and Senegal (De Araujo et al., 2006), that diversity in other populations of sub-Saharan Africa has also been underestimated and that some of the Tanzanian wild-type individuals who haemolysed were in fact deficient, but that the primers employed were not comprehensive enough to identify all cases of phenotypic deficiency.
4.3. The Case for a New Diagnostic for Safe P. vivax Radical Cure
Given the limitations to existing diagnostic methods, it is evident that there is currently no well-adapted point-of-care G6PD test for use prior to primaquine treatment in the field. A new, practical and standardised kit is required to ensure safe use of primaquine – this will require the test not only to identify an enzyme deficiency, but to give a binary assessment of whether a given dosage of primaquine can or cannot be safely adminis-tered. An important prerequisite step will be determining the position of this binary cut-off against the spectrum of haemolytic outcomes, which are known to range from negligible to highly severe.
Although the mechanisms whereby primaquine triggers haemolysis remain uncertain (Section 8.1, p. 175), and the factors determining the

R.�E.�Howes et al.154
severity of haemolysis not conclusively established, is it likely that this test will assess residual enzyme activity levels. Given this, environmental condi-tions are of major concern as these will strongly influence the diagnostic outcome as enzyme activity is heavily dependent on ambient temperature which fluctuates significantly over diurnal and seasonal cycles. Total RBC count – as influenced by anaemia and the blood volume used – will also impact the diagnosis. In terms of practicality, the test needs to be inex-pensive, provide a result rapidly, and be easily used and interpreted with minimal training (Asia Pacific Malaria Elimination Network (APMEN), 2012). Although a binary qualitative test would be easiest to implement, the heterogeneity of this disorder may require quantitative diagnoses; this con-sideration is particularly relevant to diagnosing heterozygotes.
Two visual, qualitative rapid diagnostic test (RDT) kits have been described. The BinaxNOW® G6PD assay (Binax, Inc., Maine, USA) was found to have sensitivity and specificity of 0.98 and 0.98, respectively with a cut-off of 4.0 U/g Hb (Tinley et al., 2010)2; this accuracy may not be suf-ficient for mass screening; furthermore, at around $25 per test, the cost of the test would be prohibitively high for mass screening or routine clinical use where malaria is endemic. However, this test’s major shortcoming is that it must be used within a temperature range of 18–25 °C, and is thus further unsuited to most field-based settings where P. vivax is prevalent. The Care-Start™ G6PD screening test (AccessBio, New Jersey, USA), another quali-tative phenotypic test, is similar to a malaria RDT in appearance and use (Kim et al., 2011), and has been demonstrated to be robust after prolonged storage at high temperatures (up to 45 °C for 90 days). Using a cut-off of 3.5 U/g Hb with ethylenediaminetetraacetic acid (EDTA) blood samples, the test had sensitivity of 0.68 and specificity of 1 (n = 903). The test was able to identify deficient cases up to 2.7 U/g Hb, corresponding to 22% residual enzyme activity (Kim et al., 2011). However, 13 of 903 individuals with very low G6PD activity (<2 U/g Hb) were misdiagnosed as G6PD normal; these false negative cases would therefore be at high risk of harm if administered primaquine. This evaluation study followed the manufacturer recommendations to classify any test showing even slight colour change as “normal”. In practice a far more conservative approach would be taken which would increase this test’s sensitivity and decrease its specificity. The discrepancy between the thresholds used by these two binary tests (4.0 U/g
2 These sensitivity and specificity values were from heparinized blood samples; EDTA blood samples had 0.98 sensitivity and 0.97 specificity. Samples included 50 individuals with G6PD activity ≤4.0 U/g Hb (Trinity Assay) and 196 individuals with G6PD activity >4.0 U/g Hb.

G6PD�Deficiency:�Global�Distribution,�Genetic�Variants�and�Primaquine�Therapy 155
Hb for BinaxNOW vs. 2.7 U/g Hb for AccessBio) demonstrates the uncer-tainty around what exactly determines an intolerance to primaquine and an acceptable level of haemolysis. These basic questions need to be answered before an apparent arbitrary threshold is set. So, while these tests show tan-talizing promise, further development is required for their widespread use to enable more aggressive application of primaquine with the confidence of patients’ safety.
5. MAPPING THE SPATIAL DISTRIBUTION OF G6PD DEFICIENCY
Maps provide an important evidence-base for assessing disease bur-den, for public health decision-making and for efficient resource-allocation through optimal targeting of interventions (Cromley, 2003; Hay and Snow, 2006; McLafferty, 2003), not least in relation to disorders as spatially and genetically heterogeneous as G6PD deficiency. The prevalence of G6PD defi-ciency has been mapped among populations in malaria endemic countries (Howes et al., 2012) to allow identification of where this disorder may be a problem for malaria treatment options. From this modelled map, sex-specific population estimates of deficient individuals were derived and aggregated to national and regional scales. Reported occurrences of the underlying G6PD gene variants have also been assembled (Howes et al., in preparation). The characteristics of the underlying variants are what will determine the sever-ity of potential primaquine-induced haemolysis; while the prevalence map indicates how common the deficient phenotype is. Assembly of the evidence-bases of both types of population data, and the methodological steps involved in generating the maps are summarised here. The following Section 6 (p. 165) then explores in more detail what these maps indicate about the spatial characteristics of G6PD deficiency in relation to the endemicity of P. vivax.
5.1. G6PD Deficiency Prevalence MappingVarious attempts to represent the spatial distribution of G6PD deficiency prevalence have been made. The most recent WHO map dates from 1989, and presented only national-level summaries of available data (Luzzatto and Notaro, 2001; WHO Working Group, 1989). A similar national-level map published by Nhkoma et al. in 2009 updated this effort, but still presented only national summaries, masking any subnational spatial variation in preva-lence. Subnational variation was possible to discern from the impressive com-pilation of human gene maps in Cavalli-Sforza’s History and Geography of

R.�E.�Howes et al.156
Human Genes (Cavalli-Sforza et al., 1994), however the statistical mapping methods used were rudimentary, and modern geostatistics have advanced significantly since their publication. A main limitation to all of these maps is that they give no measure of uncertainty in their predictions, either at the national-level or in a spatially specific manner. Modern geostatistics allow more comprehensive approaches to mapping, particularly in respect to sum-marising relative confidence in the predictions (Patil et al., 2011).
We focus here on the recently developed G6PD deficiency map by Howes et al. 2012, an effort of the Malaria Atlas Project (MAP), which attempted to address some of the limiting factors associated with exist-ing maps. This map was intended to represent the prevalence of clinically significant deficiency, as diagnosed by phenotypic tests, in malaria endemic countries. The evidence-base of surveys and all maps are available for down-load from the MAP website (www.map.ox.ac.uk).
5.1.1. Generating a Map: the Evidence-BaseA total of 1734 spatially unique surveys were included in the map evidence-base which met four criteria: (i) Community representativeness: all poten-tially biased samples were excluded: patients (including malaria cases), all related individuals, and all samples which selected individuals according to ethnicity; (ii) Spatial specificity: surveys had to be geographically specific and possible to geoposition accurately; (iii) Gender specificity: to allow the model to represent the X-linked inheritance mechanism of the G6PD gene, data had to be reported according to sex; (iv) Phenotypic diagnosis: only surveys which had used phenotypic diagnostic methods were included.
5.1.2. Generating a Map: the G6PD Mapping ModelModelling a continuous map of the prevalence of G6PD deficiency had to account for several difficulties: heterogeneity in the data set (both in terms of variable G6PD deficiency prevalence found at nearby locations, and in terms of the uneven distribution of the surveys across the map), the relative reliability of the data set (from highly ranging sample sizes), and the difficulties of predicting deficiency in heterozygotes due to the gene’s X-linked inheritance (Section 4.1, p. 150). A final requirement of the model was that predictions were supported by uncertainty metrics (Patil et al., 2011). A Bayesian geostatistical model was developed to cope with separate input data according to sex and the different outputs required. The core assumption of geostatistics is that of spatial-autocorrelation: namely that populations closer in space would be more similar than populations further apart. While there were some exceptions, the raw data supported

G6PD�Deficiency:�Global�Distribution,�Genetic�Variants�and�Primaquine�Therapy 157
this assumption. Populations were also assumed to be in Hardy–Weinberg equilibrium (Hardy, 1908; Weinberg, 1908).
5.1.3. G6PD Deficiency Prevalence Map: an OverviewG6PD deficiency is widespread across malaria endemic regions (Fig. 4.3). The modelled prevalence map of G6PD deficiency represents the allele fre-quency of phenotypic deficiency, equivalent to the prevalence of deficiency in males. At the continental scale, frequency of the deficiency is highest among the populations of sub-Saharan Africa, where prevalence peaks
Figure 4.3 The prevalence of G6PD deficiency in malaria endemic countries. The preva-lence is the allele frequency, which corresponds to the frequency of deficiency in males. Panels A–D correspond to Asia, Asia-Pacific, the Americas, and Africa+ regions, respec-tively. (The figure is adapted from Howes et al. (2012)). (For a colour version of this figure, the reader is referred to the online version of this book).

R.�E.�Howes et al.158
Figure 4.3—cont’d

G6PD�Deficiency:�Global�Distribution,�Genetic�Variants�and�Primaquine�Therapy 159
around 30% in several areas, but is also absent from parts of southern Africa and communities in the Horn of Africa. Prevalence of G6PD deficiency is less common across the Americas, being concentrated among populations in coastal regions. While prevalence is generally lower among Asian popula-tions than sub-Saharan Africans, the condition is widespread across Asia and particularly patchy and heterogenous in some areas.
The associated uncertainty map of the predictions is shown in Fig. 4.4; and the allele frequency map must be considered alongside these metrics
Figure 4.4 Uncertainty in the prevalence map. Uncertainty is quantified by the inter-quartile range of the model prediction and is closely associated with the proximity of population surveys, which are shown by the black dots. Panels A–D correspond to Asia, Asia-Pacific, the Americas, and Africa+ regions, respectively. (The figure is adapted from Howes et al. (2012)). (For a colour version of this figure, the reader is referred to the online version of this book).

R.�E.�Howes et al.160
Figure 4.4—cont’d

G6PD�Deficiency:�Global�Distribution,�Genetic�Variants�and�Primaquine�Therapy 161
of model confidence, as areas rich in data and with more homogenous frequencies of G6PD deficiency will be easier to model than areas where surveys are scarcely distributed or where available data indicated the under-lying prevalence of deficiency to be heterogeneous. The G6PD deficiency prevalence map’s prediction confidence is quantified in the uncertainty map by the interquartile range (IQR, the 50% confidence interval) around the model prediction. Thus, a smaller IQR is indicative of a more reli-able prediction. Areas of greatest uncertainty are those which would most benefit from new population surveys. Some potential sources of variability, however, are not accounted for by this measure, such as the underlying heterogeneity in the local population (which will not be represented if no surveys are available from that region) and the variation introduced by the diagnostic methods used. These limitations are discussed in greater detail in the original publication of this map (Howes et al., 2012).
Regional prevalence of the deficiency is discussed in detail in relation to P. vivax endemicity in Section 6 (p. 164). Geographic information systems (GIS) grids and high-resolution images of these maps, as well as the input evidence-base of surveys are freely available via the MAP website (www.map.ox.ac.uk).
5.2. G6PD Deficient Population EstimatesEstimates of the population of G6PD deficient individuals needed to account for high-resolution patterns of population density as well as short-scale varia-tion in G6PD deficiency prevalence to ensure that population density was reflected in the overall prevalence estimate. Howes and colleagues (2012) derived these population estimates in a Bayesian framework so as to generate uncertainty metrics around the population estimates (Patil et al., 2011). The aggregated national-level numbers of G6PD deficient individuals are mapped in Fig. 4.5. An overall allele frequency of deficiency of 8.0% (IQR: 7.4–8.8) was predicted across all malaria endemic countries. This corresponded to 220 million affected males (IQR: 203–241) and an estimated 133 million females (IQR: 122–148). The model results indicate that a median estimate of 26% of expected genetic heterozygotes were diagnosed as being phenotypically defi-cient based on the raw survey data. Within the subset of 35 countries target-ing malaria elimination, where primaquine treatment would be particularly beneficial, overall prevalence was lower with a predicted allele frequency of 5.3% (IQR: 4.4–6.7), meaning that in 2010 an estimated 61 million males (IQR: 51–77) and 35 million females (IQR: 29–46) were predicted to be phenotypically G6PD deficient in countries eliminating malaria.

R.�E.�How
es et al.162
Figure 4.5 National allele frequency of G6PD deficiency. (Figure from Howes et al. (2012)).

G6PD�Deficiency:�Global�Distribution,�Genetic�Variants�and�Primaquine�Therapy 163
Although many of the highest frequencies of deficiency were predicted from sub-Saharan Africa, the very high population densities across Asia meant that the overall population burden was largely focussed there. China and India, for instance, were estimated to be home to 41.3% of all G6PD-deficient males across the whole malaria endemic region globally. The high allele frequencies seen in Africa meant that 28.0% of the overall deficient population was in sub-Saharan Africa, while only 4.5% of the global popu-lation burden was in the Americas, and 67.5% across the whole of Asia. National-level estimates for all malaria endemic countries were provided in the original publication (Howes et al., 2012).
5.3. G6PD Deficiency Mutation MappingTo understand the clinical characteristics of G6PD deficiency in different regions, it is also necessary to map the underlying G6PD mutations. A sim-ple map of key genetic variants was published in 2001 (Luzzatto and Notaro, 2001), and a new database of G6PD variants has since been assembled to update this effort (Howes et al., 2012; Howes et al., in preparation). Surveys were collated which provided measures of the proportions of each variant among G6PD deficient individuals in different areas. Exclusion of popula-tion samples from hospitalised or exclusively symptomatic patients avoided a bias towards higher proportions of the more severe variants, which might otherwise have been preferentially identified.
Striking patterns emerged across malaria endemic regions (Fig. 4.6). Genetic heterogeneity was found to be relatively low across populations of the Americas and West Asia, where the A- and Mediterranean vari-ants predominated, respectively. Further east, genetic diversity increased among Indian populations where the Orissa variant, barely reported outside India, predominated among certain communities in east, cen-tral India. Populations in countries east of India carried a completely different set of mutations, and genetic diversity of the G6PD gene was far greater. Although a handful of variants were found to be more com-monly reported from specific populations (such as the Mahidol variant across Myanmar and Thailand; Viangchan variant across Mekong region; Kaiping variant among Chinese populations and the Vanua Lava variant in central and eastern Indonesia), genetic diversity was high among these populations. The proportion of “Unidentified” variants was also great-est among Asian populations, emphasising the inadequacy of molecu-lar methods for diagnosing phenotypic deficiency: a limited number of primers cannot reliably identify all possible cases of deficiency.

R.�E.�Howes et al.164
Figure 4.6 Proportions of G6PD variants among phenotypically deficient community samples. Panels A to D correspond to Asia, Asia-Pacific, the Americas and the Africa+ regions. Pie charts show the local proportions of each variant, sized according to the sur-vey sample sizes and plotted on a logarithmic scale. The individuals from whom these data originate are known to be phenotypically deficient. All samples are therefore deficient, and the “Other/Unidentified” category represents G6PD variants which were too rarely reported to be individually represented in the map, or which could not be identified. (For a colour version of this figure, the reader is referred to the online version of this book).
6. SPATIAL CO-OCCURRENCE OF G6PD DEFICIENCY WITH P. VIVAX ENDEMICITY
We now discuss the spatial epidemiology of G6PD deficiency, its prevalence and genetic variants, in relation to its public health significance in the context of P. vivax transmission. The geographical limits of P. vivax

G6PD�Deficiency:�Global�Distribution,�Genetic�Variants�and�Primaquine�Therapy 165
Figure 4.6—cont’d

R.�E.�Howes et al.166
transmission and its endemicity within those limits have been described in detail in the opening chapter of volume 80 and are shown in Fig. 4.7. The map shows P. vivax endemicity, as quantified by community parasite rate in the 1–99 year age range (PvPR1–99), with grey areas representing unstable transmission where <1 case per 10,000 population occurs per year (Chapter 1 of Volume 80; and Gething et al., 2012).
6.1. G6PD Deficiency in AsiaThe majority of the global population at risk of P. vivax transmission is on the Asian continent (Gething et al., 2012), defined as stretching from Turkey, south to Vietnam and east to the People’s Republic of Korea. The prevalence and genetic variants of G6PD deficiency among these populations were found to be heterogeneous, as was the transmission ende-micity of P. vivax. While P. vivax endemicity is generally very low in Asian countries west of India, being either at no risk or having small patches of low endemicity in areas of unstable transmission, several of these countries had relatively high prevalence of G6PD deficiency compared with other parts of Asia. For instance, the national allele frequency of deficiency across Iran was predicted to be 11.8% (IQR: 9.9–14.1), and Azerbaijan had a national estimate of 10.2% (IQR: 8.9–11.7); both countries had only limited areas of very low/unstable P. vivax transmission. However, G6PD prevalence model uncertainty was high in this region, peaking (up to 37% IQR) in an area devoid of data across southern Pakistan. This high uncer-tainty emphasises the need for additional community surveys to support mapping of G6PD deficiency in this apparently high prevalence region. The underlying G6PD mutation among these populations was reportedly
Figure 4.7 Global endemicity of Plasmodium vivax endemicity. Assembly of this map is described in Chapter 1 of Volume 80. (Figure from Gething et al. (2012)). (For a colour version of this figure, the reader is referred to the online version of this book).

G6PD�Deficiency:�Global�Distribution,�Genetic�Variants�and�Primaquine�Therapy 167
the Mediterranean variant, which was identified in >70% of deficient individuals in all surveys between Turkey and Pakistan (Fig. 4.6A).
Further east, G6PD deficiency was present but at relatively low prevalence (2–5%) across much of the Indian subcontinent, increasing in the eastern states of Chhattisgarh, Orissa and Jharkhand where prevalence reached 5–25%; the common variants in this area were the Mediterranean, Kerala-Kalyan and Orissa variants, the latter being highly restricted in its spatial extent to east India. These areas coincided with high P. vivax endemicity, which reached >7% PvPR1–99 in the neighbouring province of Andhra Pradesh. Another high prevalence area of G6PD deficiency in Asia (approximately 20%) was around the northern Lao–Thai border, an area largely P. vivax free, with only small patches of unstable transmission. Lower G6PD deficiency frequencies were predicted along coastal Myanmar (2–3%), an area of high (>7%) P. vivax endemicity. High frequencies of the two diseases, however, occurred simul-taneously in southern Thailand where endemicity was >7% and G6PD defi-ciency prevalence 5–9%. Prevalence of both disorders was therefore variable with often highly focal P. vivax transmission across this region.
In terms of the G6PD genetic mutations, there was a stark change in variants and greatly increased diversity across countries east of India. G6PD Mahidol was either universal or very common among communities in Myan-mar, remaining prevalent among Thai populations, in whom the Viangchan was also frequently reported. No single variant predominated among Chinese populations, instead the Kaiping, Canton and Gaohe variants were all com-mon. A high proportion of samples were also rare or could not be identified using standard molecular primers, an indicator of high genetic heterogeneity.
6.2. G6PD Deficiency in Asia-PacificSome of the largest continuous regions of high P. vivax endemicity glob-ally were predicted across the Asia-Pacific region, where endemicity reached >7% across much of Papua New Guinea, the Solomon Islands and Vanuatu. Coinciding with this, G6PD deficiency was also prevalent on these islands, peaking at 23% prevalence on the southern reaches of Santa Isabel and Gua-dalcanal islands (where the Union G6PD variant was the main cause of deficiency, Fig. 4.6B). Both diseases were highly heterogeneous across Indo-nesia, by far the most populous nation in this region. Both G6PD deficiency prevalence and P. vivax endemicity were particularly high across the central Nusa Tenggara islands, such as on Flores where endemicity was over 7% and G6PD deficiency approximately 10%. A dearth of G6PD population surveys on Sulawesi introduced relatively high uncertainty to the map; in contrast,

R.�E.�Howes et al.168
numerous parasite rate surveys indicated that P. vivax transmission was mostly unstable in this area. Additional G6PD surveys would be particularly valuable among the south-eastern populations of Sulawesi, where endemicity reached 5%, and G6PD deficiency was predicted at 8% due to the high frequencies observed on nearby islands. G6PD deficiency was heterogeneous across the region. Relatively high genetic diversity was reported with multiple variants commonly co-existing in high proportions alongside important frequencies of unidentified variants. For instance across Papua New Guinea, frequencies of 1% were found along the southern coast which rose to 15% along the East Sepik northern coast. The Vanua Lava G6PD variant was commonly reported from populations in both Indonesia and Papua New Guinea, but was not reported from anywhere outside this region.
A neonatal screening programme for G6PD deficiency exists across the Philippines which contributed high density of prevalence data (n = 636 data points) indicating a spatially variable national prevalence of 2 to 3% (population-adjusted national allele frequency estimate is 2.5% [IQR: 2.4 to 2.5] (Howes et al., 2012)). Across the Philippines, stable P. vivax transmis-sion was only found on islands at the northern and southern ends of the country, peaking in northern Luzon at around 5% endemicity, where G6PD deficiency ranged in prevalence between 1 and 4%.
6.3. G6PD Deficiency in the AmericasThe lowest predictions of G6PD deficiency prevalence globally were across the Americas, where 40.8% of the land area had median prevalence predic-tions of ≤1%. Prevalence ranged from 0% across parts of Mexico, Peru, Bolivia and Argentina, to a national allele frequency prediction of 8.6% in Venezuela. Surveys were relatively scarce across large parts of the continent, particularly in Venezuela which led to high uncertainty around the national allele frequency estimate (IQR: 4.0–18.0). Plasmodium vivax endemicity in this area of high G6PD deficiency prevalence was patchy, with areas of 3 to 4% PvPR1–99 inter-spersed with large expanses of unstable transmission. Deficiency prevalence was also predicted to rise in the central and southern coastal provinces of Bra-zil, such as around the cities of São Paulo and Porto Alegre where P. vivax was absent. The A- variant of G6PD was predominant, causing >80% of deficiency cases across coastal communities of Brazil and Central America, and explaining 62% of deficient cases in a survey in central Mexico (Fig. 4.6C).
Plasmodium vivax endemicity peaked in two areas of the Americas: Honduras and Nicaragua in Central America, and Amazonas province in northwest Brazil (Fig. 4.7); areas where G6PD deficiency prevalence was relatively low compared with other parts of the continent. Parasite rates of 6

G6PD�Deficiency:�Global�Distribution,�Genetic�Variants�and�Primaquine�Therapy 169
to 7% coincided with G6PD deficiency allele frequency national estimates of 2.9% (IQR: 1.5–5.8) in Honduras and 1.5% (IQR: 0.6–3.6) in Nica-ragua. Very few G6PD surveys were found from Amazonian communities, but parts of this region where P. vivax endemicity exceeded 7% were pre-dicted to have G6PD deficiency prevalence of up to 3%. Additional G6PD deficiency surveys in both these areas of high P. vivax endemicity would be valuable, particularly focussed in areas of high population density.
6.4. G6PD Deficiency in Africa, Yemen and Saudi Arabia (Africa+)
Plasmodium vivax epidemiology in the Africa+ region differs starkly from other malaria endemic areas due to the high prevalence of Duffy negativity among these populations (Howes et al., 2011) which depresses transmission to unsta-ble levels across most of the continent (Chapter 2 of this volume). Therefore, in spite of its high prevalence, G6PD deficiency in Africa+ has relatively little bearing on the global picture of P. vivax therapy. The only two areas of stable P. vivax transmission in Africa+ are Ethiopia and close surrounds, and Mada-gascar. G6PD deficiency prevalence in the Horn of Africa is low, with national allele frequency in Ethiopia estimated at 1% (IQR: 0.7–1.5); though stable, the coincident parasite endemicity is equivalently low, with most areas being at 1% PvPR1–99, interspersed with patches of 2% endemicity. The highest single pre-diction of G6PD deficiency prevalence globally is on the Persian Gulf coast of Saudi Arabia where P. vivax is absent. Plasmodium vivax transmission is negligible across this whole peninsula, with only a narrow strip of unstable transmission along the west coast of Saudi Arabia and Yemen. Endemicity on Madagascar is more significant, reaching 2 to 3% PvPR1–99 in the inland and central coastal regions. Only a single-community G6PD survey was available so although the national allele frequency estimate is high at 19.4%, additional surveys would be needed to reduce uncertainty in this estimate (IQR: 11.5–30.3).
Although G6PD deficiency does not present a hurdle to P. vivax radical cure in most parts of Africa+, the main potential application of primaquine across this region is for blocking transmission of P. falciparum (Eziefula et al., 2012; Bousema and Drakeley, 2011). Prevalence estimates of G6PD defi-ciency in Africa+ are the highest globally, with 14 countries predicted to have national allele frequencies >15%, all across sub-Saharan Africa from Ghana (19.6% [IQR: 14.2–27.0]) in the west across to Mozambique in the east (21.1% [IQR: 14.7–29.8]). Several areas were predicted to have particularly high prevalence (approximately 30%), including the coastal areas of West Africa (Ghana to Nigeria), the mouth of the Congo river (western Congo, Democratic Republic of Congo and Angola) and west Sudan. Subnational

R.�E.�Howes et al.170
heterogeneity was important in some areas, for example ranging from 30% around Ibadan to 2% in northwest Nigeria. Prevalence of the deficiency decreased at the continental extremities: in the western Sahel, southern Africa and the Horn of Africa. Prediction uncertainty across the continent was heterogeneous, being very high in areas lacking data such as central Africa between the Democratic Republic of Congo and Madagascar, and the Sudan–Chad border. Additional G6PD community surveys are imperative to reduce the map’s high uncertainty, and caution with primaquine administra-tion must reflect the high prevalence of the condition.
There is a notable absence of G6PD variant surveys from Africa (Fig. 4.6D), which may be in part associated with the presumption of low G6PD genetic heterogeneity among those populations. As a consequence, many of the community G6PD surveys conducted do not use phenotypic diagnostics to identify deficient individuals and instead use only molecular methods to detect a narrow range of variants. These data cannot therefore inform the relative prevalence of variants among deficient individuals. Available surveys indicated that the A- variant was predominant across deficient individuals in sub-Saharan Africa (Burkina Faso and the Comores), with the exception of a Sudanese study which identified a greater diversity, including the Mediterra-nean variant (Saha and Samuel, 1991). The Mediterranean variant was com-mon (>50%) in two investigations of deficient individuals in Saudi Arabia.
7. EVOLUTIONARY DRIVERS OF THE DISTRIBUTION OF G6PD DEFICIENCY
The widespread distribution and frequent high prevalence of G6PD deficiency—a genetic disorder associated with important clinical costs— presents an evolutionary paradox. The spatial overlap between G6PD defi-ciency and the precontrol distribution of malaria (Lysenko’s map of precontrol malaria is reprinted in Chapter 1 of Volume 80) was first remarked upon by Motulsky (1960) and Allison (1960) shortly after the description of G6PD deficiency and led Allison and Clyde (1961) to propose Haldane’s Malaria Hypothesis (Haldane, 1949) as an explanation for the natural selection of this deleterious condition. This idea, which had recently been substantiated by empirical evidence for the sickle-cell mutation (Allison, 1954), implied that the deleterious G6PD deficiency condition carried, at least in some geno-types (homo- or heterozygotes), a selective survival advantage over normal enzyme levels against malaria morbidity or mortality: a possible case of bal-ancing selection (Haldane, 1949). This has attracted much attention over the past half century with evidence from epidemiological observations, in vitro

G6PD�Deficiency:�Global�Distribution,�Genetic�Variants�and�Primaquine�Therapy 171
laboratory findings studies and in vivo clinical studies strongly supporting the hypothesis that this common genetic trait has been selected for by malaria through conferring some degree of resistance against the severity of the infectious disease. A number of excellent reviews of this body of work have been published (Greene, 1993; Hedrick, 2011; Kwiatkowski, 2005; Luzzatto, 1979, 2004; Ruwende and Hill, 1998; Tripathy and Reddy, 2007).
7.1. Evidence of a Selective Advantage7.1.1. Epidemiological EvidenceThe most convincing epidemiological evidence of selection by malaria is the sheer diversity of variants of the G6PD gene, many of which have reached polymorphic frequencies in genetically isolated populations suggesting the independent selection of each variant: an apparent case of convergent evo-lution by a common agent of selection, perhaps. Given that all polymorphic variants are found in historically malaria endemic regions, it would appear that these two diseases, infectious and congenital, are somehow associated. Micro-mapping studies across altitudinal or climatic gradients of malaria endemicity also suggest that the prevalence of this disorder is the result of selection by malaria, rather than random drift or selection (Luzzatto, 1979; Ruwende and Hill, 1998). A recent study from Sumba island in Indonesia provides an impressive example of G6PD deficiency frequencies correlat-ing with a strong malaria endemicity gradient over short spatial distances (Satyagraha, unpublished data). G6PD deficiency prevalence ranged from 2.2 to 3.8% in Central Sumba where malaria endemicity was low, to as high as 11% in highly endemic malaria hotspots to the west and southwest of the island where P. falciparum was found year-round together with seasonable P. vivax endemicity. While these observations provide no evidence of causality, the spatial association between these two diseases is evident.
7.1.2. In vitro EvidenceIn vitro studies have demonstrated unambiguously that parasitaemia is less successful in G6PD deficient cells than in wild-type cells (Cappellini and Fiorelli, 2008; Roth et al., 1983). The clearest demonstration of this was by Luzzatto et al. (1969), who compared parasitaemia in both cell types in heterozygotes (studying heterozygotes overcame the potentially con-founding effect of different levels of acquired immunity which would exist between different individuals). Cells with normal enzyme levels were 2–80 times more likely to be infected than deficient cells. Cappadoro et al. (1998) examined P. falciparum intracellular development and identified selective early-stage phagocytosis of infected G6PD deficient cells as a possible

R.�E.�Howes et al.172
protective mechanism. Although they found no significant difference in the growth and development of P. falciparum parasites between normal and defi-cient cells (Mediterranean variant), infected deficient cells were 2.3 times more intensely phagocytosed when parasites reached the early stages of the schizogonic developmental cycle, early in the erythrocytic stages of infection.
7.1.3. Case-Control In vivo EvidenceLarge-scale case-control studies look for a protective effect against malaria at the population level. Specifically, this in vivo evidence is drawn on to identify which G6PD deficient genotypes the selective advantage is conferred upon: hemi- and homozygotes, or heterozygotes? While studies all concur in iden-tifying a selective advantage associated with G6PD deficiency, the particular genotypes benefitting vary between studies, with data seemingly supporting all scenarios. For instance, a study in southwest Nigeria reported signifi-cantly lower parasitaemia in heterozygous females but not hemizygous males (Bienzle et al., 1972). Subsequent large case-control studies used clinical symptoms rather than parasitaemia as the indicator of protection, and found robust evidence for a protective role of the G6PD A- variant (G202A) in hemizygous males and homozygous females in sub-Saharan Africa (Guindo et al., 2007; Ruwende et al., 1995); the protection extended to heterozygous femals remained contentious. Ruwende et al. (1995) surveyed children in The Gambia and Kenya and found a 46% reduction in risk from severe malaria in A- heterozygotes, similar to the 58% protection estimated against severe malaria in hemizygotes. This study compared mild or severe malaria against community controls who were asymptomatic or parasite-free. A comparable case-control study in most respects was subsequently conducted in Malian children (Guindo et al., 2007), except that the control samples used were of uncomplicated malaria cases, rather than asymptomatic/malaria-free cases, a reflection of the local hyperendemic malaria transmission. This study found no protection conferred by this same A- mutation against heterozygous females (n = 221), in spite of a very similar positive result for hemizygotes.
As well as the different indicators of parasitic protection (parasitaemia vs. clinical symptoms) and the different control groups (asymptomatic/malaria free vs. mild parasitaemia), further difficulty in comparing case-control studies may be introduced by the method used to diagnose the enzyme deficiency. Johnson et al. (2009) demonstrated in Uganda how differences between phenotypic and genotypic diagnostics can affect the apparent sus-ceptibility to malaria of different G6PD statuses, with significant protection

G6PD�Deficiency:�Global�Distribution,�Genetic�Variants�and�Primaquine�Therapy 173
from uncomplicated malaria only identified in phenotypically deficient females.
7.2. Neglect of the Selective Role of P. vivax as a Driver of G6PD Deficiency
Most early studies have focussed on the protective role of G6PD deficiency on malaria in Africa, and thus considered only a narrow representation of the G6PD gene’s overall genetic variation and clearly neglected a potential role for P. vivax, despite this parasite having a wider transmission range than P. falciparum (Guerra et al., 2010) and causing significant morbidity and mortal-ity (Chapter 3 of Volume 80 and Price et al., 2007). An important life cycle difference between these parasites is P. vivax’s preference for infecting reticu-locytes (Anstey et al., 2009; Kitchen, 1938), which could confer a much greater fitness cost on the host by hindering regeneration of the erythrocyte pool. From the host perspective, G6PD enzyme activity levels in reticulo-cytes are at their highest. If a deficiency in enzyme activity can convey a protective advantage against severe clinical symptoms, then the deficiency would need to be particularly severe to be expressed in the reticulocyte stages. In theory, therefore, P. vivax could be exerting much stronger selection pressures on the host than P. falciparum, selecting more severe variants of the G6PD gene; the generally asymptomatic nature of these mutations would mean that the fitness cost of severe deficiency would not always be felt.
Indeed, G6PD Mediterranean, one of the most severe polymorphic variants (<1% residual activity), has been found to offer significant protection against symptomatic P. vivax among male and female Afghan refugees in Pakistan (Leslie et al., 2010). Males and homozygous females were more significantly protected than heterozygotes in whom only weak protection was found. This study offered no comparison with protection against P. falciparum, however, as it accounted for only 5% of infections in the study area. Another study, in an area of co-endemic P. falciparum and P. vivax malaria in Thailand (Louicharoen et al., 2009) found evidence of P. vivax having been the selective agent of the G6PD Mahidol variant, which is common across parts of southeast Asia (Section 6.1, p. 166). In this very comprehensive study, an evolutionary approach using exten-sive single-nucleotide analysis (SNP) around the G6PD gene locus showed high homogeneity between haplotypes of the Mahidol mutation (G487A), indicating that the mutation had undergone recent and strong positive selec-tion, thus suggestive of conferring a strong advantage to human survival. Clini-cal studies indicated that this survival advantage was conferred as protection against P. vivax parasitaemia, having no effect on P. falciparum parasitaemia.

R.�E.�Howes et al.174
Moving forward, a number of fascinating questions remain unanswered. For instance, the large number of G6PD mutations which have reached polymorphic frequencies is of interest, as is the apparent lower diver-sity among African populations than others. Estimates of the ages of some of these mutations propose relatively recent origins, which may explain part of this diversity. Estimates range from 1000 to 6357 years for the A- variant (Sabeti et al., 2002; Slatkin, 2008; Tishkoff et al., 2001), 3330 years for the Mediterranean variant (Tishkoff et al., 2001), and 1575 years for the Mahi-dol variant (Louicharoen et al., 2009). Further study of the ages of a range of variants would allow a comprehensive picture of the evolutionary history of this condition, and its association with the spread of human Plasmodium infec-tions. Studies should consider both the role of P. vivax as well as P. falciparum to allow the relative selection pressure of the two parasites to be determined with respect to specific variants. The role of P. vivax as a selective agent of human polymorphisms is further discussed in Chapter 2 of this volume, particularly in reference to the protective role of the Duffy negativity blood group.
8. PRIMAQUINE, P. VIVAX AND G6PD DEFICIENCY
Primaquine has a vital and unique role in the malaria elimination toolkit, fulfilling three critical functions: first, it is the only licenced radical cure of P. vivax; second, primaquine is the only drug active against mature, infectious P. falciparum gametocytes making it vital for blocking transmis-sion; and third, in areas of emerging drug resistance, primaquine is being used in containment programmes to prevent the spread of artemisinin resis-tant P. falciparum strains (WHO, 2011b). These invaluable properties make understanding the triangle of interplaying aspects determining primaquine-induced haemolytic risk crucial: the human enzyme, the drug and the par-asite. The relationship between P. vivax and primaquine is considered in detail in Chapter 4 of Volume 80, and not revisited in detail again here. We consider here means of safely administering the 200 mg total dose of primaquine required for P. vivax radical cure (Alving et al., 1953; Baird and Hoffman, 2004; Baird and Rieckmann, 2003; Coatney et al., 1953; Edgcomb et al., 1950).
First, we review the molecular mechanisms by which primaquine trig-gers haemolysis, second how haemolytic risk varies according to prima-quine dosing regimens, and third, how different G6PD variants modulate haemolytic risk and severity. Understanding primaquine’s pharmacological properties and biochemical effects on the cell is necessary for the develop-ment of safer regimens, and alternative safer drugs.

G6PD�Deficiency:�Global�Distribution,�Genetic�Variants�and�Primaquine�Therapy 175
8.1. Mechanism of Primaquine-Induced HaemolysisIt is well established that primaquine-induced haemolysis does not occur in individuals with normal levels of G6PD activity (Baird et al., 2001; Bunnag et al., 1994; Edgcomb et al., 1950). Furthermore, haemolytic risk is greatest in the oldest RBCs, corresponding to an increasing risk as enzyme activ-ity decays over time (Beutler, 1994; Beutler et al., 1954). These indications suggest that primaquine-induced haemolysis is directly associated with the consequences of reduced G6PD enzyme activity. However, the mechanism by which this occurs remains uncertain, and the instability and diversity of primaquine metabolites make studying this system exceedingly difficult. Despite having been in circulation since the 1950s when clinical use of primaquine began, relatively little work has been done to elucidate the pre-cise molecular events leading to primaquine-induced AHA. Understanding these may be critical in rationally disassociating the haemolysing toxicity of the drug from its broad-acting therapeutic properties (Pybus et al., 2012) in developing superior therapies.
Although the focus here of primaquine side-effects is the haemolytic risk to G6PD deficient individuals, it should also be noted that primaquine can cause a number of other side-effects, previously reviewed (Baird and Hoffman, 2004; Hill et al., 2006). For instance, abdominal pain is a common dose-dependent side-effect (Edgcomb et al., 1950; Hill et al., 2006), which can be prevented through simply taking the pills with food (Clayman et al., 1952). Primaquine also routinely causes a relatively mild, although occa-sionally symptomatic methaemoglobinaemia (typically about 6% for as long as dosing lasts), further discussed below.
8.1.1. Primaquine and its MetabolitesPrimaquine is rapidly excreted, with an elimination half-life of about 4 h (Carson et al., 1981; Greaves et al., 1980), and metabolises into a com-plex array of a dozen or so distinct moieties. One metabolite is carboxy-primaquine, which has been detected in plasma within 30 min of dosing, reaching plasma concentrations 10-fold higher than primaquine (Mihaly et al., 1985). Carboxy-primaquine, however, appears physiologically inert, both with respect to toxicity and therapeutic activity. On the other hand, some moieties are highly unstable and oxidatively volatile. For instance, 5-hydroxy-6-methoxy-8-aminoquinoline (5H6MQ) was 2500 times more potent than primaquine in stimulating the PPP in normal RBCs (Baird et al., 1986b). It is likely to be one or several metabolic products of prima-quine which is the active agent against the parasites, rather than the parent compound itself (Beutler, 1969; Carson et al., 1981; Fletcher et al., 1988).

R.�E.�Howes et al.176
The molecular events triggering haemolysis remain unproven, though several mechanisms of action have been proposed, including the build-up of methaemoglobin (metHb) causing damage to the cell membrane, and oxida-tive damage provoked by primaquine metabolites. The relative significance of these pathways remains unclear. In vitro haemotoxicity assays have demonstrated that it is cytochrome P450-linked pathways which metabolise the breakdown of primaquine by redox reactions into its haemotoxic metabolites, including the formation of metHb and the generation of reactive oxygen intermediates (Ganesan et al., 2009), in a dose-dependent response (Ganesan et al., 2012); the assay found no haemotoxic effect of primaquine when exposed to cells in the absence of these cytochromes. Further study with isoenzyme activity screen-ing and steady state kinetic data has identified two cytochrome P450 enzymes (MAO-A and 2D6) to be strongly involved in primaquine metabolism, and reported that the metabolites generated by these enzymes made it likely that the drug’s toxicity and efficacy were enabled by the same single-cytochrome pathway, making it unlikely that these two properties of the drug could be separated (Pybus et al., 2012). Further study is required to support these find-ings and determine the relative influence of inhibitors and inducers of these specific cytochrome enzymes to test their individual effect on the drug’s effi-cacy and toxicity. Similarly, while a number of primaquine metabolites have been identified, none have been definitively associated with activity against the Plasmodium parasite (Baird and Hoffman, 2004; Myint et al., 2011).
8.1.2. A Role for Oxidative StressOxidative stress caused by primaquine metabolites has been long-held as the favoured hypothesis for explaining the drug’s toxicity. Reduced levels of G6PD enzyme activity leave the cell with diminished anti-oxidant reserves, namely NADPH and reduced glutathione (Flanagan et al., 1958), due to the constrained rate of the PPP, as already discussed. Possible oxidative agents which have been proposed include free radicals, such as activated oxygen, and hydroxylated metabolites of primaquine which auto-oxidise into qui-noneimine products, superoxides, hydroxyl radicals and hydrogen peroxide (Brueckner et al., 2001; Fletcher et al., 1988); with oxidative metabolites generated by the cytochrome P450 enzymes previously described (Ganesan et al., 2009, 2012; Pybus et al., 2012). Oxidised glutathione, which accu-mulates during oxidative stress, has been found to be a strong intracellular mediator activating membrane cation channels (Koliwad et al., 1996). The opening of these Ca2+-permeable channels can trigger cell death (apopto-sis) during oxidative stress (Lang et al., 2003, 2006), although a recent study

G6PD�Deficiency:�Global�Distribution,�Genetic�Variants�and�Primaquine�Therapy 177
by Ganesan et al. (2012) did not find this mechanism to be triggered by the primaquine-induced haemolytic pathway. Another suggested mecha-nism whereby depletion of reduced glutathione can lead to cell death was described by Bowman and colleagues, who demonstrated that primaquine-associated oxidative stress (triggered in their experiments by exposure to 5-hydroxyprimaquine) induced oxidative injury to the erythrocyte cyto-skeleton (Bowman et al., 2005b), accelerating the process of cell phagocy-tosis (Bowman et al., 2005a).
8.1.3. A Role for MethaemoglobinThe role of metHb accumulation has been proposed to be highly significant, with oxidised primaquine derivatives, such as quinones or iminoquinones (including 5-hydroxyprimaquine) found strongly associated with the for-mation of metHb (Link et al., 1985). Oxidative stress from primaquine can lead to the oxidation of haemoglobin iron (Fe2+ → Fe3+) and the build-up of metHb. This resulting condition, methaemoglobinaemia, is usually asymptomatic and self-limiting if only low levels accumulate (Brueckner et al., 2001; Fernando et al., 2011). At higher proportions, however, meth-aemoglobinaemia can result in tissue hypoxia, hypoxemia and cyanosis due to metHb’s low affinity for oxygen (Hill et al., 2006; Percy et al., 2005). However, exacerbated methaemoglobinaemia is not a feature of clinical acute haemolytic anaemia in G6PD deficient patients.
8.1.4. A Role for Altered Redox EquilibriumOther experimental evidence points away from damage mediated through oxidative degradation of the RBC cytosol or membrane. Baird and col-leagues observed that the increased PPP activity stimulated by primaquine metabolites occurred independently of glutathione redox activity. They observed that when sodium nitrite was applied to the system, glutathione redox stimulated the PPP. When the dose of sodium nitrite exceeded the ability of the PPP to maintain steady state redox equilibrium, a proteolytic system that degraded irreversibly oxidised proteins became active (Baird et al., 1986b). In contrast, doses of primaquine metabolites that also satu-rated PPP activity came with no such proteolytic activity, and the drugs were not mediating oxidative damage like sodium nitrite. It appeared that the drugs themselves drove NADPH depletion and the PPP response to that disturbed equilibrium (Baird et al., 1986a). In other words, redox equilibrium between a reduced and oxidised species of primaquine would, in a G6PD deficient cell, strongly favour the oxidised species, without

R.�E.�Howes et al.178
necessarily prompting a broad oxidative degradation of cytosol proteins (Brueckner et al., 2001). The oxidised species could be the agent of hae-molysis. Brueckner et al. (2001) proposed a mechanism similar to that demonstrated for the well-characterised molecular events in phenylhydra-zine-induced haemolysis in rats. They postulated that the covalent linkage of a metabolite to the haem moiety of haemoglobin could force the mol-ecule from its globin fold by simple hydrophobic force into the lipid bilayer of the RBC membrane.
Any mechanism of haemolysis must ultimately be reconciled with what is highly likely to be a very brief and quantitatively insubstantial oxidative chal-lenge to the RBC by primaquine. The daily 15 or 30 mg dose may be appre-ciated as an exceedingly small quantity of molecules relative to its distribution in tissue, rapid metabolism to a dominant and inert species, and very rapid elimination. The oxidative challenge must be very insubstantial and fleeting compared with, for example, nitrite poisoning. Although a mild methaemo-globinaemia typically occurs with normal primaquine dosing (Carmona-Fonseca et al., 2009), it does not appear linked to depletion of GSH via a generalised oxidative attack of RBC proteins. The damage being done, what-ever it may be, seems irreversible during the intervals between dosing. The damage appears cumulative across doses, with haemolysis not commencing in earnest until after the third or fourth dose. These features appear incompat-ible with a general oxidative stress and would point to an irreversible capture of the harmful primaquine species, or the damage done by it, in the RBC. An accumulation of displaced haem molecules in the RBC membrane, manifest as Heinz bodies, is compatible with all of these features.
8.1.5. Significance of Primaquine-Induced HaemolysisWhatever the mechanism, be it the build-up of metHb, direct oxidative damage, the balance of redox equilibrium, or any other pathway, the physi-ological damage caused to the cells leads to intravascular haemolysis, mak-ing acute haemolytic anaemia the main clinical symptom. Freely circulating haemoglobin from the haemolysed cells causes the most severe and poten-tially lethal conditions, including haemoglobinuria and acute renal failure (Burgoine et al., 2010).
Understanding the mechanisms by which primaquine induces haemolysis is critical to developing safer administration of prima-quine. In the same way that knowledge of the dose-dependency effect of primaquine (described in the next section) has allowed treatment regimens to be extended over a number of weeks to reduce their tox-icity, further manipulations to improve safe dosing schedules would

G6PD�Deficiency:�Global�Distribution,�Genetic�Variants�and�Primaquine�Therapy 179
likely be possible given a better understanding of its mode of action. For example, primaquine interaction with co-administered schizon-ticides has been suggested to affect its toxicity (Myint et al., 2011). If confirmed, this could prove a relatively straightforward solution towards safer therapy. Improvements to diagnostic methods by ensuring that their target corresponds to the predisposing factor to haemolysis might also be possible with an improved understanding of the predictors of haemolytic susceptibility. Another main motivation for studying the biochemical pro-cesses of primaquine action is pharmaceutical: the potential for disassociating the drug’s therapeutic properties from its toxicity. If feasible, this could lead to safe and effective killing of hypnozoites in P. vivax therapy and control.
8.2. Factors Affecting Haemolytic RiskWhile the biochemical mechanisms of primaquine-induced haemoly-sis remain uncertain, it is clear that the risk and severity of haemolysis is highly variable, affected by both exogenous and endogenous factors (Beutler, 1994). Generalisations about haemolytic risk and its clini-cal severity are frequently cited based on the binary categorisations of “mild” and “severe” enzyme deficiency (Hill et al., 2006; WHO, 2010, 2011a). In reality, a spectrum of clinical severity exists, ranging from asymptomatic to lethal, which is determined by several factors includ-ing the primaquine dose and the genetic and biochemical determinants of enzyme activity levels. Many studies investigating haemolytic risk aggregated all phenotypically “deficient” individuals, sometimes speci-fying “mild” or “severe” deficiency (Hill et al., 2006; Recht et al., 2012); here we consider reports relating to specific genetic variants. The pri-maquine sensitivity phenotypes of three have been investigated in detail, and we discuss these here.
8.2.1. Dose DependencyThe dose dependency of the haemolytic effect of primaquine was noted from the first clinical investigations into primaquine, with observations that toxicity appeared to be cumulative: “in some instances symptoms began late in the course of drug administration and continued for several days after its discontinuance” (Edgcomb et al., 1950). The pharmacokinetics of pri-maquine, however, are unaffected by dose, and its efficacy is contin-gent upon the total dose administered, rather than the total amount per dose or the timescale over which it is administered (Mihaly et al., 1985). A meta-analysis by Schmidt and colleagues of the early data (1946–1975) on dosing regimens in Rhesus monkeys found that the success of radical

R.�E.�Howes et al.180
cure is determined by the overall total primaquine dose, independent from the timescale over which it is administered: 1-, 3-, 7- and 14-day regimens were not found to differ in their therapeutic success (Schmidt et al., 1977). This pharmacokinetic property has the advantageous benefit of permitting extended dosing schedules which reduce the risks of haemolysis without impacting on its effectiveness.
8.2.2. Variant DependencyThe diversity of G6PD genetic variants leads to a spectrum of residual activ-ity levels and associated clinical severity (Section 3, p. 143). We discuss here how drug regimens which can be tolerated by some G6PD deficient indi-viduals were developed in relation to studies on certain key G6PD variants, and as a corollary, how haemolytic risk is variant-dependent.
(a) A- variant. The early Stateville studies into “primaquine sensitivity” were all based on the A- variant, due to the origins of the individual studied. The vulnerability presented by this variant to primaquine-induced hae-molysis was recently re-affirmed by a Brazilian study which resulted in severe haemolysis in three G6PD deficient P. vivax patients treated with 30 mg daily doses of primaquine over 5 or 7 days (Silva et al., 2004). The Stateville studies found that daily dosing of the 30 mg regimen led to hae-molytic symptoms in affected individuals appearing on the second or third day, peaking 4–7 days after starting treatment, continuing for just over a week overall (Clyde, 1981; Hill et al., 2006; Hockwald et al., 1952). Hae-molysis ended within a few days of ceasing treatment, after which haemo-globin levels recovered; recovery even occurred with continued treatment as haemolysis was compensated by erythropoiesis and the regeneration of reticulocytes which have inherently higher G6PD levels.
The drug’s total dose effect has allowed regimens with an acceptable level of toxicity for G6PD deficient individuals to be developed. Studies by Alving et al. found that “intermittent” weekly 45 mg doses of prima-quine over 8 weeks was effective (treating 90% of P. vivax Chesson strain infections; n = 40) and did not produce any clinically significant haemolysis (the number of G6PD deficient individuals was not specified) (Alving et al., 1960). Subsequently, Brewer and Zarafonetis (1967), studying individuals of African origin, confirmed that twice-weekly administration of 45 mg pri-maquine triggered more haemolysis than once-weekly dosing in deficient individuals; no haemolysis was reported from individuals with no deficiency. A review by Clyde (1981) determined that daily 15 mg primaquine doses for 14 days would be safely tolerated, with any side-effects remaining largely

G6PD�Deficiency:�Global�Distribution,�Genetic�Variants�and�Primaquine�Therapy 181
asymptomatic; but recommended the extended 8-weekly 45 mg dosing schedule. As such, current WHO recommendations for adults with “mild” G6PD deficiency are for 8-weekly 45 mg doses of primaquine (Hill et al., 2006; WHO, 2010, 2011a). As well as averting the severity of haemolysis, another benefit of extending the dosing schedule is the opportunity for affected G6PD deficient patients to discontinue treatment before the sever-ity of haemolysis becomes too serious. In these “mild” cases, haemolysis is self-limiting and ends a few days after stopping treatment.
However, a review by the American Centers for Disease Control and Prevention (CDC) recommends great caution when administering prima-quine to individuals with any degree of G6PD deficiency, and emphasises that very careful risk assessments and strict medical supervision are essential (Hill et al., 2006). The authors note that this weekly dosing schedule for G6PD deficient individuals has not been approved by the FDA. Indeed, case reports of severe adverse effects to these “safe” regimens exist, including even severe reactions in individuals once deemed to be at especially low risk. For instance, a heterozygote with the A- variant (G202A mutation) was reported to suffer a clinically severe haemoglobin drop to <5 g/dl following a single 45 mg dose of primaquine (Shekalaghe et al., 2010). A possible reason for this unexpected outcome may stem from a misdiagnosis, as only one locus in the gene was tested in the mutation analysis and other more severe variants would not have been identified.
Recent studies of primaquine in individuals with the A- variant are few (Eziefula et al., 2012) and focus on the drug’s application to P. falciparum gametocyte clearance – single dose of 0.75 mg/kg primaquine in combina-tion with artemisinin combination therapy (ACT) (Shekalaghe et al., 2007, 2010). However, a clinical trial in several African countries of individuals with the A- variant with another drug-trigger of AHA in G6PD deficient individuals, dapsone, resulted in high rates of haemolysis, with 10.9% of hemi- and homozygotes requiring transfusion following 150 mg/day dos-ing over 3 days.3 Authors analysing these outcomes conclude that, contrary to current perception, the A- variant cannot be considered “mild” (Pamba et al., 2012).
(b) Mahidol variant. This is an important variant among populations from Myanmar and Thailand, and is reported to cause residual activity to drop to
3 Direct comparison on the haemolysing effect of dapsone in relation to primaquine was investigated during the Stateville trials, when a G6PD deficient African male was exposed on different occasions to daily doses of 100 mg dapsone for 21 days, and 30 mg of daily primaquine doses for 18 days (Degowin et al., 1966). The haemolytic effect was less marked with dapsone.

R.�E.�Howes et al.182
5–32% of normal levels (Louicharoen et al., 2009). A study investigating the effects of primaquine has been conducted in a relatively small number of P. vivax-positive G6PD deficient individuals (n = 22) in Thailand who were given 15 mg primaquine for 14 days with standard chloroquine treatment (Buchachart et al., 2001). No serious adverse effects were subsequently reported during the 28-day follow-up period, though a 24.5% (±13.9) drop in haematocrit was observed in the G6PD deficient patients with an equivalent only 1.2% (±14.4) drop in G6PD normal individuals. No patient required transfusion. This study concludes that standard primaquine therapy would be safe in Thailand, even for those G6PD deficient (Buchachart et al., 2001). Although these individuals were only diagnosed as deficient pheno-typically, the authors reported the Mahidol variant predominant in the study area. Polymerase chain reaction (PCR) diagnosed individuals from a simi-lar study in Thailand were all diagnosed as carrying Mahidol variant; no serious adverse effects were reported from this study of 14-day primaquine therapy (Takeuchi et al., 2010). A similar attitude is presented from an economic perspective by Wilairatana et al. (2010) who determined that in the absence of widely available G6PD testing at the malaria clinic level, only patients who develop black urine or anaemia should be tested for G6PD deficiency, and that “mild-moderate” deficient cases should be pre-scribed the 8-weekly dose. A study from Myanmar where 22 G6PD defi-cient individuals were given the 8-weekly 45 mg primaquine dosage also reported no severe adverse effects (Myat Phone et al., 1994), and similarly concludes that weekly dosing is safe for G6PD deficient individuals in Myanmar. The authors of these studies therefore encourage blind prima-quine therapy in this region, where G6PD deficiency allele frequency is high: estimated to be 13.6% (IQR: 11.9–15.5) in Thailand and 6.1% (IQR: 4.1–9.3) across Myanmar (Howes et al., 2012). The WHO specifies that primaquine should be used for P. vivax radical cure in this region, but that prior G6PD screening is necessary (WHO, 2010). Given the difficulties of field-based G6PD diagnosis, it is uncertain whether clinicians actually risk administering primaquine ( John et al., 2012). The evidence-base for these recommendations, however, is relatively meagre (Recht et al., 2012). The heterogeneity of local variants reported from this region (Fig. 4.6A) is also an important reason to exercise great caution, even if a handful of studies have reported no serious adverse reactions to primaquine, these studies may not have included patients with severe G6PD variants. For instance, a more severe variant, Viangchan, is also common in Thailand, and along the Cambodian and Laos borders.

G6PD�Deficiency:�Global�Distribution,�Genetic�Variants�and�Primaquine�Therapy 183
In spite of these risks, the use of primaquine in mass drug administration has been investigated in Cambodia for P. falciparum transmission control. Very low dosing (9 mg/10 days for 6 months; n = 6040) administered alongside an ACT without G6PD testing (despite local prevalence being 18.6% in males) was not found to cause any severe adverse effects (Song et al., 2010). Importantly, however, no active monitoring was in place to detect these. This very low dos-ing, therefore, was deemed safe for P. falciparum transmission control; however, this dosing bears little relation to P. vivax radical cure, and would not be logis-tically feasible as a standard therapy due to its very resource-intensive dosing.
(c) Mediterranean variant. Originally known for its association with the clinical pathology of favism, the Mediterranean variant causes one of the most severely deficient phenotypes, reducing enzyme activity to <1% of normal levels (Beutler and Duparc, 2007). Serious haemolysis is caused by 15 mg daily or 45 mg weekly courses of primaquine (Clyde, 1981) and, unlike with the A- variant, haemolysis in G6PD Mediterranean individuals is not self-limiting. A review by Clyde concluded that individuals affected by the Mediterranean variant should be administered supervised weekly doses of 30 mg for 15 weeks (Clyde, 1981). WHO guidelines today, how-ever, state that no primaquine should be given to individuals with so severe a deficiency (WHO, 2010, 2011a).
8.2.3. Red Blood Cell Age DependencySenescent RBCs are most likely to succumb to haemolytic challenges (Beutler et al., 1954). Wild-type erythrocytes appear to have a large surplus of potential G6PD activity, allowing the PPP to be significantly upregu-lated when exposed to oxidative stress (Salvador and Savageau, 2003). This enzyme activity decays naturally with RBC age (as erythrocytes lack nuclei and therefore have no mechanism for regenerating enzyme levels), correlating with susceptibility to haemolytic risk. This decay was found to be exponential, with a half-life of 62 days for wild-type enzyme and 13 days for A- enzyme (Piomelli et al., 1968). The more severe variant, Mediterranean, had such a rapid decline that no enzyme activity could be detected in mature erythrocytes. Cells with the A- genetic variant were demonstrated to be insensitive to primaquine when 8–21 days old, but were rapidly destroyed 55 days later (corresponding to slightly older than half their normal lifespans) when re-exposed to primaquine (Beutler et al., 1954). In wild-type individuals natural enzyme decay does not reach levels which put the individual at clinical risk; in G6PD deficient cells, however, the ageing process being so much more marked than in normal

R.�E.�Howes et al.184
cells, means that even moderately deficient cells will have enzyme activ-ity levels that drop to clinically at-risk levels. If erythropoiesis can replace haemolysed cells, then the clinical effect will be negligible and the hae-molysis self-limiting once all susceptible cells have been destroyed (Alving et al., 1960; Dern et al., 1954a).
8.2.4. Sex DependencyHaemolytic risk is also sex-dependent. Although primaquine’s funda-mental pharmacokinetics are not affected by gender (Cuong et al., 2006; Elmes et al., 2006), the majority of affected females are heterogeneous and therefore present less commonly with severe symptoms than males. In areas of high prevalence, however, homozygote inheritance of deficiency can be common (Howes et al., 2012), and heterozygous females can also suffer severe haemolysis (Pamba et al., 2012; Shekalaghe et al., 2010). The pop-ulation of RBCs carrying the deficiency are at equal haemolytic risk as homozygous or hemizygous cells. The relative proportion of wild-type and deficient cells will have a major influence in determining the overall clinical severity of haemolytic stress at the individual level. Interactions with other genetic blood disorders may also exacerbate the effects of the deficiency in females (Chopra, 1968).
8.3. Predicting Haemolytic RiskThe core motivation for understanding G6PD deficiency in the context of P. vivax control is in being able to ascertain haemolytic risk and pre-vent adverse drug events from primaquine therapy. However, while much research has been done and the complexity of interacting factors has been clearly demonstrated, key knowledge gaps still hinder reliable predictions of haemolytic risk. The main assumption made when assessing haemolytic risk is that enzyme activity levels can be a reliable indicator. Although this would appear to be generally true, exceptions exist, such as an Iranian boy with 19.5% residual activity requiring a transfusion after a single 45 mg dose of primaquine (Ziai et al., 1967), and a heterozygote Tan-zanian experiencing severe adverse reaction also following a single dose (Shekalaghe et al., 2010); a detailed record of adverse drug events has been compiled (Recht et al., 2012). While these cases are exceptions, from a clinical perspective, interpretation of diagnostic outcomes must allow for the uncertainties in these relationships. The practical implica-tions of these considerations and difficulties in assessing haemolytic risk are discussed in the next section.

G6PD�Deficiency:�Global�Distribution,�Genetic�Variants�and�Primaquine�Therapy 185
9. TOWARDS A RISK FRAMEWORK FOR P. VIVAX RELAPSE TREATMENT
In this final section, we consider how the current state of under-standing about G6PD deficiency and primaquine may be practically applied to promoting safe radical cure of P. vivax. G6PD deficiency is widespread, predicted in all malaria endemic countries, with an overall estimated allele frequency of 8.0% (IQR: 7.4–8.8), as discussed in Section 5.2 (p. 161). Given its potential clinical severity, primaquine cannot be administered without careful prior assessment of risk. This haemolytic risk may be con-sidered at two scales: (i) large regional scales for public health perspectives, and (ii) directly by clinicians in relation to individual-level treatment deci-sions. Once risk has been satisfactorily judged, primaquine can be adminis-tered, or withheld, accordingly. Important limitations hinder risk assessment at both scales, however, and we discuss necessary developments towards overcoming these and improving safe access to P. vivax radical cure.
9.1. Assessing National-Level Haemolytic Risk of Primaquine Therapy
Knowledge of the spatial characteristics of G6PD deficiency can be cou-pled with information about clinical phenotypes to allow comparisons of haemolytic risk between regions at scales of public health significance. It is important to note that assessment of risk at such large spatial scales cannot inform risk at the level of the individual, and can never replace the need for careful oversight of primaquine therapy by clinicians (Hill et al., 2006), even in areas considered to be at low risk from a public health perspective.
A simple national-level framework assessing large-scale risk has been proposed (Howes et al., 2012), but important limitations to the data inform-ing this analysis make it only a coarse-scaled and crude framework: one which must be refined as our understanding of clinical risk improves.
9.1.1. Proposed Framework for Ranking National-Level Risk from G6PD Deficiency
Current WHO treatment guidelines consider haemolytic risk to dif-fer between “mild” and “severe” classes of deficiency. “Mild to moder-ate” cases of deficiency may be treated with 8 weekly 45 mg doses, while “severely” d eficient individuals should not be administered any primaquine

R.�E.�Howes et al.186
( WHO, 2010, 2011a). Consequently, relative haemolytic risk from prima-quine due to G6PD deficiency at the population level may be considered to be contingent upon two factors: (i) the overall prevalence of deficiency within that population, and (ii) the relative composition of mild and severe genetic G6PD variants reported from that population. The simple framework suggested by Howes et al. (2012) scores the national prevalence of deficiency based on the national allele frequency estimates described in Section 5.2 (p. 161), and assigns variant severity scores using a database of documented reports of G6PD variants to assess the relative proportion of Class II and Class III variants nationally (WHO-endorsed subdivisions of the Type 2 category of variants; Table 4.1 and WHO Working Group, 1989). An overall risk score was obtained for each country by multiplying the prevalence score by the vari-ant severity score to give six categories of risk across a spectrum from “rare and mild” G6PD deficiency to “common and severe” G6PD deficiency (Fig. 4.8).
Overall, this simple risk analysis ranked the highest level of G6PD deficiency risk as being in the Asia and Asia–Pacific regions where severe variants were reported and population prevalence of deficiency was com-mon (>1% allele frequency). High prevalence of deficiency caused by pre-dominantly Class III variants across sub-Saharan African countries led to moderate levels of risk in this region. Risk scores in the Americas ranged from low to moderate. National-level scores are mapped in Fig. 4.8; further details about the methodology employed are given in the original publica-tion (Howes et al., 2012).
9.1.2. Important Limitations to Predicting National-Level Haemolytic Risk
Both the underlying assumptions and the value of the described risk stratification are questionable. It is nonetheless useful to attempt to do so with regard to identifying weaknesses and the knowledge gaps that cause them. The analysis’ main assumption is that the severity of haemolysis can be predicted from the genetic variant and that haemolytic severity is adequately represented by the subjective categorisations of Classes II and III. Further, this assumes that primaquine sensitivity phenotypes inversely correlate with residual enzyme activity (categorised here as Classes II and III). Given that the mechanism of primaquine-induced haemolysis remains uncertain (Section 8.1, p. 175) only isolated case reports exist to support this assumption. Although the data from the three variants described in Section 8.2 (p. 179) would appear to support this correlation, the evi-dence underpinning this relationship across all variants is not robust, and

G6PD�Deficiency:�Global�Distribution,�Genetic�Variants�and�Primaquine�Therapy 187
extrapolating risk to all variants based on their WHO classifications is therefore very uncertain. Furthermore, the evidence used to determine the classifications of variants into their corresponding WHO Class (tabu-lated by Beutler, 1993; Kwok et al., 2002; Luzzatto et al., 2001; Minucci et al., 2012; UCL Bioinformatics Group website; Vulliamy et al., 1997; Yoshida et al., 1971) is weak in many cases, being determined from small numbers of samples with variable laboratory techniques (Recht et al., 2012). Given this uncertainty in the risk analysis, only a very coarse clas-sification with three scores of variant severity was used. Finally, the data set informing the national classifications of variant severity is often poor, with data reported from only 54 of 90 malaria endemic countries, meaning that severity scores had to be inferred for many of them.
If a robust understanding of the relationship between genetic variants and their primaquine sensitivity phenotypes could be incorporated in this analysis with a more complete background picture of the spatial hetero-geneity of the G6PD variants and their prevalence, the conclusions drawn would be much more valuable. Given these gaps in our current understand-ing, the main robust predictor of haemolysis is the phenotypic deficiency in G6PD enzyme activity. The modelled prevalence map may therefore be the most detailed, reliable, and appropriate risk assessment of overall G6PD deficiency-associated harm – be it moderate or severe – which is relevant to informing public health policy for P. vivax radical cure.
9.2. Assessing Haemolytic Risk at the Level of the IndividualGiven the numerous testing kits which have been developed to diagnose G6PD deficiency (Section 4, p. 150), assessing haemolytic risk at the level of the individual ought to be straightforward. However, logistical con-straints hinder the widespread use of these diagnostic methods in the rural communities where P. vivax is most common and point-of-care testing is most needed. Further, it would appear that many of the available diagnos-tic methods have not been calibrated to any predetermined and clinically relevant measures of risk, and are seemingly arbitrary in their classifications of deficiency. For instance, the threshold for distinguishing deficient from nondeficient cases ranges from 10% to 60% residual enzyme activity. These assessments of haemolytic risk also assume that enzyme activity is a suitable indicator of primaquine-induced haemolysis. Their poor suitability to diag-nosing deficiency in females (Section 4.1, p. 150) is a further hindrance to their suitability for discerning haemolytic risk. No molecular methods are currently suited to field-based settings.

R.�E.�Howes et al.188
Although WHO treatment recommendations distinguish “mild” from “severe” deficiency, no details are provided to indicate what these categories correspond to in terms of enzyme activity or associated degree of hae-molysis. Given the difficulties with diagnosing these severity types, and the largely unknown clinical risk which “mild” deficient individuals would be exposed to, it has been argued that any G6PD deficient phenotype, regard-less of severity, should suffice as a contraindication for primaquine therapy (Baird and Surjadjaja, 2011). Distinguishing the degree of severity is not possible with most diagnostic tests, including the binary point-of-care tests currently in development (Section 4.3, p. 153). Attempting to account for the severity of deficiency in a framework determining safe primaquine therapy may therefore be an unnecessary complication at this stage, and instead it may be more appropriate to re-assign the WHO treatment guide-lines with binary options, removing the additional subjective mild/severe distinction. Anecdotally, the importance of simplicity and great caution in determining drug guidelines is well illustrated by a recent case report of a Burmese P. vivax patient, undiagnosed as G6PD deficient, misunderstand-ing his primaquine regimen guidelines due to linguistic barriers, and taking almost his full course of primaquine (165 mg) in one go, resulting in severe haemoglobinuria (Burgoine et al., 2010). Risk assessment with a potentially harmful drug must be especially cautious.
Until the evidence underpinning the mechanism of haemolytic risk (Sec-tion 8, p. 174) allows predictability of the haemolytic outcome of different variants with specific primaquine dosages, simple and safe guidelines based on practicable diagnoses appear the most appropriate end-points of haemolytic risk assessments. A better understanding of the mechanism of drug-induced
Figure 4.8 National risk index from G6PD deficiency. Two aspects of G6PD deficiency epidemiology were used to define the national risk of G6PD deficiency: (1) the national prevalence of deficiency was stratified into three classes (≤1%; >1–10%; >10%) based on the national allele frequency estimates described in Section 5.2 (Fig. 4.5); (2) the severity of local variants was classified at the national level based on a literature search of reports of occurrences of G6PD variants. Variants were classified into mild and severe types, in accordance with the WHO-endorsed classification (WHO Working Group, 1989). The relative proportion of reported Class II and Class III variants was used to categorise the severity of variants nationally (Class III variants only; minority of Class II variants, ≤1/3; Class II variants common, >1/3), as shown in Panel A. The two scores were multiplied to given an overall risk score (Panel B and C). Uncertainty in the two scores was scored and ranked between countries (Panel D and E). (Figure from Howes et al. (2012)). (For a colour version of this figure, the reader is referred to the online version of this book).

G6PD�Deficiency:�Global�Distribution,�Genetic�Variants�and�Primaquine�Therapy 189

R.�E.�Howes et al.190
haemolysis might allow diagnostic methods to be engineered as predictors of haemolysis, rather than a direct indicator of enzyme activity (which may not prove to be the only determinant of haemolytic risk). The heterogeneity of genetic variants (Fig. 4.6A,B), particularly across areas where P. vivax radical cure is most relevant (Fig. 4.7), supports the potential benefits of improving the diagnostic resolution and tailoring treatments to different variants. How-ever, until the underlying haemolytic mechanisms can be understood, the short-term primary goal must be refinement of binary diagnostics.
For individuals diagnosed as G6PD normal, a different set of consid-erations apply to optimising primaquine treatment options. For instance, there is pressure to reduce treatment duration from 14 days to a 7-day regi-men with equivalent efficacy so as to improve compliance (Fernando et al., 2011; Krudsood et al., 2008; Schmidt et al., 1977; Takeuchi et al., 2010). Prerequisite to promoting this higher dosage schedule is a high sensitivity diagnostic of haemolytic risk for G6PD deficiency.
10. CONCLUSIONS
The aim of studying G6PD deficiency in the context of P. vivax therapy and malaria elimination is to support safe use of 8-aminoquinoline drugs for radical cure that will enable access to effective, life-saving therapy. At this pivotal time in malaria control, when important progress is being made and vital funding cuts imposed (Garrett, 2012), it is more imperative than ever to maximise efficient use of available tools. As discussed in Chapter 6 of Volume 80, the relapsing P. vivax hypnozoite reservoir makes this para-site life-form the major challenge to patient health and malaria elimination programmes. The only licenced drug active against these parasites is prima-quine. However, widespread G6PD deficiency (estimated allele frequency of 5.3% [IQR: 4.4–6.7] in declared malaria eliminating countries, Howes et al. (2012)) and poor associated diagnostics result in primaquine being under-used. While great caution must be taken in dealing with a potentially haemolysing drug, informed caution may increase access to this drug.
Although a good deal is known about the molecular characteristics of the G6PD enzyme and its clinical manifestations as favism and NNJ, for instance, the disorder’s interactions with P. vivax and primaquine remain much less well understood. This is likely to be a symptom of the higher priority which has been placed in recent decades on therapy against the asexual blood stages of P. falciparum (Baird, 2010), to the neglect of most other therapeutic targets, such as P. vivax radical cure. As such, a recurring

G6PD�Deficiency:�Global�Distribution,�Genetic�Variants�and�Primaquine�Therapy 191
theme emerging from many sections of this review is a lack of data, tools and understanding. Prioritising these gaps to increase access to safe pri-maquine can be considered in three steps, though fundamental to all of these is an understanding of how residual enzyme activity levels and genetic variants interact with the underlying mechanisms triggering haemolysis, thereby providing evidence that can guide rationally developed and practi-cal solutions to the problem. 1. Improving point-of-care assessment of haemolytic risk through devel-
opment of a highly sensitive, practical diagnostic which can be con-sidered a conservative indicator of tolerance of the standard 14-day regimen. It may be necessary to develop separate methods to adequately diagnose deficient males and females (Peters and Van Noorden, 2009; Shah et al., 2012). Adequate diagnostics would greatly increase access to primaquine and could be available in the near future.
2. Extending access to primaquine by identifying dosing regimens of reduced toxicity for G6PD deficient individuals by leveraging unex-plored synergies with other drugs.
3. Developing alternative therapies (likely non-8-aminoquinolines) which present no risk to G6PD deficient individuals.
The operational inadequacy and potentially mortal threat posed by pri-maquine due to G6PD deficiency, renders it unfit for purpose in endemic zones in its current form. The dawning realization that acute P. vivax malaria is associated with significant burdens of severe illness and death in endemic zones and no longer misclassified as benign (Price et al., 2007) may finally crystalise the determination of the scientific community to address this 60-year-old problem. There may be no higher priority for malaria research than the triangular G6PD-primaquine-P. vivax problem.
If elimination is to be the focus, perspectives on future malaria therapy need to shift away from treatment of symptomatic parasitaemia towards comprehen-sive treatment for all parasites and multiple life stages (Baird, 2012). Given their unique therapeutic action, the importance of overcoming the dangers of the 8-aminoquinolines cannot be over-emphasised towards meeting this target.
ACKNOWLEDGEMENTSThe authors are particularly grateful to Lucio Luzzatto and Pete Zimmerman for valu-able comments on the manuscript, and to Jennie Charlton and David Pigott for proof-reading. This work was supported by a Wellcome Trust Biomedical Resources Grant (#085406), which funded R.E.H.; S.I.H. is funded by a Senior Research Fellowship from the Wellcome Trust (#095066) that supports K.E.B. also. A.W.S. is supported by grant #107-13 from the Asia Pacific Malaria Elimination Network (APMEN). J.K.B. is supported

R.�E.�Howes et al.192
by grant #B9RJIXO of the Wellcome Trust. This work forms part of the output of the Malaria Atlas Project (MAP, http://www.map.ox.ac.uk/), principally funded by the Wellcome Trust, UK.
REFERENCESAllison, A.C., 1954. Protection afforded by sickle-cell trait against subtertian malarial infec-
tion. Br. Med. J. 1, 290–294.Allison, A.C., 1960. Glucose-6-phosphate dehydrogenase deficiency in red blood cells of
East Africans. Nature 186, 531–532.Allison, A.C., Clyde, D.F., 1961. Malaria in African children with deficient erythrocyte glu-
cose-6-phosphate dehydrogenase. Br. Med. J. 1, 1346–1349.Alving, A.S., Arnold, J., Robinson, D.H., 1952. Mass therapy of subclinical vivax malaria
with primaquine. JAMA 149, 1558–1562.Alving, A.S., Craige, B., Pullman, T.N., Whorton, C.M., Jones, R., Eichelberger, L., 1948.
Procedures used at Stateville Penitentiary for the testing of potential antimalarial agents. J. Clin. Investig. 27, 2–5.
Alving, A.S., Hankey, D.D., Coatney, G.R., Jones Jr., R., Coker, W.G., Garrison, P.L., Dono-van, W.N., 1953. Korean vivax malaria. II. Curative treatment with pamaquine and pri-maquine. Am. J. Trop. Med. Hyg. 2, 970–976.
Alving, A.S., Johnson, C.F., Tarlov, A.R., Brewer, G.J., Kellermeyer, R.W., Carson, P.E., 1960. Mitigation of the haemolytic effect of primaquine and enhancement of its action against exoerythrocytic forms of the Chesson strain of Plasmodium vivax by intermittent regi-mens of drug administration: a preliminary report. Bull. W H O 22, 621–631.
Anstey, N.M., Russell, B., Yeo, T.W., Price, R.N., 2009. The pathophysiology of vivax malaria. Trends Parasitol. 25, 220–227.
Asia Pacific Malaria Elimination Network (APMEN), 2012. Vivax Working Group Meeting – Incheon, Republic of Korea. URL http://apmen.org/vxwg-2012/.
Au, S.W., Gover, S., Lam, V.M., Adams, M.J., 2000. Human glucose-6-phosphate dehydroge-nase: the crystal structure reveals a structural NADP(+) molecule and provides insights into enzyme deficiency. Structure 8, 293–303.
Baird, J.K., 2010. Eliminating malaria–all of them. Lancet 376, 1883–1885.Baird, J.K., 2011. Resistance to chloroquine unhinges vivax malaria therapeutics. Antimi-
crob. Agents Chemother. 55, 1827–1830.Baird, J.K., 2012. Elimination therapy for the endemic malarias. Curr. Infect. Dis. Rep.Baird, J.K., Davidson Jr., D.E., Decker-Jackson, J.E., 1986a. Oxidative activity of hydroxyl-
ated primaquine analogs. Non-toxicity to glucose-6-phosphate dehydrogenase-deficient human red blood cells in vitro. Biochem. Pharmacol. 35, 1091–1098.
Baird, J.K., Hoffman, S.L., 2004. Primaquine therapy for malaria. Clin. Infect. Dis. 39, 1336–1345.
Baird, J.K., Lacy, M.D., Basri, H., Barcus, M.J., Maguire, J.D., Bangs, M.J., Gramzinski, R., Sismadi, P., Krisin, Ling, J., Wiady, I., Kusumaningsih, M., Jones, T.R., Fryauff, D.J., Hoff-man, S.L., 2001. Randomized, parallel placebo-controlled trial of primaquine for malaria prophylaxis in Papua, Indonesia. Clin. Infect. Dis. 33, 1990–1997.
Baird, J.K., McCormick, G.J., Canfield, C.J., 1986b. Effects of nine synthetic putative metab-olites of primaquine on activity of the hexose monophosphate shunt in intact human red blood cells in vitro. Biochem. Pharmacol. 35, 1099–1106.
Baird, J.K., Rieckmann, K.H., 2003. Can primaquine therapy for vivax malaria be improved? Trends Parasitol. 19, 115–120.
Baird, J.K., Surjadjaja, C., 2011. Consideration of ethics in primaquine therapy against malaria transmission. Trends Parasitol. 27, 11–16.
Bernstein, R.E., 1962. A rapid screening dye test for the detection of glucose-6-phosphate dehydrogenase deficiency in red cells. Nature 194, 192–193.

G6PD�Deficiency:�Global�Distribution,�Genetic�Variants�and�Primaquine�Therapy 193
Beutler, B., 2009. Obituary: Ernest Beutler (1928–2008). Haematologica 94, 154–156.Beutler, E., 1959. The hemolytic effect of primaquine and related compounds: a review.
Blood 14, 103–139.Beutler, E., 1969. Drug-induced hemolytic anemia. Pharmacol. Rev. 21, 73–103.Beutler, E., 1990. The genetics of glucose-6-phosphate dehydrogenase deficiency. Semin.
Hematol. 27, 137–164.Beutler, E., 1991. Glucose-6-phosphate dehydrogenase deficiency. N. Engl. J. Med. 324,
169–174.Beutler, E., 1993. Study of glucose-6-phosphate dehydrogenase: history and molecular biol-
ogy. Am. J. Hematol. 42, 53–58.Beutler, E., 1994. G6PD deficiency. Blood 84, 3613–3636.Beutler, E., 1996. G6PD: population genetics and clinical manifestations. Blood Rev. 10,
45–52.Beutler, E., 2008. Glucose-6-phosphate dehydrogenase deficiency: a historical perspective.
Blood 111, 16–24.Beutler, E., Blume, K.G., Kaplan, J.C., Lohr, G.W., Ramot, B., Valentine, W.N., 1979. Inter-
national Committee for Standardization in Haematology: recommended screening test for glucose-6-phosphate dehydrogenase (G-6-PD) deficiency. Br. J. Haematol. 43, 465–467.
Beutler, E., Dern, R.J., Alving, A.S., 1954. The hemolytic effect of primaquine. IV. The relationship of cell age to hemolysis. J. Lab. Clin. Med. 44, 439–442.
Beutler, E., Dern, R.J., Alving, A.S., 1955. The hemolytic effect of primaquine. VI. An in vitro test for sensitivity of erythrocytes to primaquine. J. Lab. Clin. Med. 45, 40–50.
Beutler, E., Duparc, S., 2007. Glucose-6-phosphate dehydrogenase deficiency and antima-larial drug development. Am. J. Trop. Med. Hyg. 77, 779–789.
Beutler, E., Mitchell, M., 1968. Special modifications of the fluorescent screening method for glucose-6-phosphate dehydrogenase deficiency. Blood 32, 816–818.
Beutler, E., Vulliamy, T.J., 2002. Hematologically important mutations: glucose-6-phosphate dehydrogenase. Blood Cells Mol. Dis. 28, 93–103.
Beutler, E., Yeh, M., Fairbanks, V.F., 1962. The normal human female as a mosaic of X-chromosome activity: studies using the gene for G-6-PD-deficiency as a marker. Proc. Natl. Acad. Sci. U S A 48, 9–16.
Bienzle, U., Ayeni, O., Lucas, A.O., Luzzatto, L., 1972. Glucose-6-phosphate dehydrogenase and malaria. Greater resistance of females heterozygous for enzyme deficiency and of males with non-deficient variant. Lancet 1, 107–110.
Bousema, T., Drakeley, C., 2011. Epidemiology and infectivity of Plasmodium falciparum and Plasmodium vivax gametocytes in relation to malaria control and elimination. Clin. Microbiol. Rev. 24, 377–410.
Bowman, Z.S., Jollow, D.J., McMillan, D.C., 2005a. Primaquine-induced hemolytic anemia: role of splenic macrophages in the fate of 5-hydroxyprimaquine-treated rat erythrocytes. J. Pharmacol. Exp. Ther. 315, 980–986.
Bowman, Z.S., Morrow, J.D., Jollow, D.J., McMillan, D.C., 2005b. Primaquine-induced hemolytic anemia: role of membrane lipid peroxidation and cytoskeletal protein altera-tions in the hemotoxicity of 5-hydroxyprimaquine. J. Pharmacol. Exp. Ther. 314, 838–845.
Brewer, G.J., Tarlov, A.R., Alving, A.S., 1962. The methemoglobin reduction test for prima-quine-type sensitivity of erythrocytes. A simplified procedure for detecting a specific hypersusceptibility to drug hemolysis. JAMA 180, 386–388.
Brewer, G.J., Zarafonetis, C.J., 1967. The haemolytic effect of various regimens of prima-quine with chloroquine in American Negroes with G6PD deficiency and the lack of an effect of various antimalarial suppressive agents on erythrocyte metabolism. Bull. W H O 36, 303–308.

R.�E.�Howes et al.194
Brueckner, R.P., Ohrt, C., Baird, J.K., Milhous, W.K., 2001. 8-Aminoquinolines. In: Rosen-thal, P.J. (Ed.), Antimalarial Chemotherapy: Mechanisms of Action, Resistance, and New Directions in Drug Discovery, Humana Press, Totowa, NJ.
Buchachart, K., Krudsood, S., Singhasivanon, P., Treeprasertsuk, S., Phophak, N., Srivilairit, S., Chalermrut, K., Rattanapong, Y., Supeeranuntha, L., Wilairatana, P., Brittenham, G., Looareesuwan, S., 2001. Effect of primaquine standard dose (15 mg/day for 14 days) in the treatment of vivax malaria patients in Thailand. Southeast Asian J. Trop. Med. Public Health 32, 720–726.
Bunnag, D., Karbwang, J., Thanavibul, A., Chittamas, S., Ratanapongse, Y., Chalermrut, K., Bangchang, K.N., Harinasuta, T., 1994. High dose of primaquine in primaquine resistant vivax malaria. Trans. R Soc. Trop. Med. Hyg. 88, 218–219.
Burgoine, K.L., Bancone, G., Nosten, F., 2010. The reality of using primaquine. Malaria J. 9, 376.Burka, E.R., Weaver, Z., Marks, P.A., 1966. Clinical spectrum of hemolytic anemia asso-
ciated with glucose-6-ghosphate dehydrogenase deficiency. Ann. Intern. Med. 64, 817–825.
Cappadoro, M., Giribaldi, G., O’Brien, E., Turrini, F., Mannu, F., Ulliers, D., Simula, G., Luzzatto, L., Arese, P., 1998. Early phagocytosis of glucose-6-phosphate dehydrogenase (G6PD)-deficient erythrocytes parasitized by Plasmodium falciparum may explain malaria protection in G6PD deficiency. Blood 92, 2527–2534.
Cappellini, M.D., Fiorelli, G., 2008. Glucose-6-phosphate dehydrogenase deficiency. Lancet 371, 64–74.
Carmona-Fonseca, J., Alvarez, G., Maestre, A., 2009. Methemoglobinemia and adverse events in Plasmodium vivax malaria patients associated with high doses of primaquine treatment. Am. J. Trop. Med. Hyg. 80, 188–193.
Carson, P.E., Flanagan, C.L., Ickes, C.E., Alving, A.S., 1956. Enzymatic deficiency in prima-quine-sensitive erythrocytes. Science 124, 484–485.
Carson, P.E., Hohl, R., Nora, M.V., Parkhurst, G.W., Ahmad, T., Scanlan, S., Frischer, H., 1981. Toxicology of the 8-aminoquinolines and genetic factors associated with their toxicity in man. Bull. W H O 59, 427–437.
Cavalli-Sforza, L.L., Menozzi, P., Piazza, A., 1994. The History and Geography of Human Genes. Princeton University Press, Princeton, NJ.
Chevion, M., Navok, T., Glaser, G., Mager, J., 1982. The chemistry of favism-inducing com-pounds. Eur. J. Biochem. 127, 405–409.
Chopra, S.A., 1968. Haemolytic crisis in a Zanzibari Arab girl with G6PD deficiency and sickle cell trait. East Afr. Med. J. 45, 726–727.
Clark, T.G., Fry, A.E., Auburn, S., Campino, S., Diakite, M., Green, A., Richardson, A., Teo, Y.Y., Small, K., Wilson, J., Jallow, M., Sisay-Joof, F., Pinder, M., Sabeti, P., Kwiatkowski, D.P., Rockett, K.A., 2009. Allelic heterogeneity of G6PD deficiency in West Africa and severe malaria susceptibility. Eur. J. Hum. Genet. 17, 1080–1085.
Clayman, C.B., Arnold, J., Hockwald, R.S., Yount Jr., E.H., Edgcomb, J.H., Alving, A.S., 1952. Toxicity of primaquine in Caucasians. JAMA 149, 1563–1568.
Clyde, D.F., 1981. Clinical problems associated with the use of primaquine as a tissue schizon-tocidal and gametocytocidal drug. Bull. W H O 59, 391–395.
Coatney, G.R., Alving, A.S., Jones Jr., R., Hankey, D.D., Robinson, D.H., Garrison, P.L., Coker, W.G., Donovan, W.N., Di Lorenzo, A., Marx, R.L., Simmons, I.H., 1953. Korean vivax malaria. V. Cure of the infection by primaquine administered during long-term latency. Am. J. Trop. Med. Hyg. 2, 985–988.
Coatney, G.R., Cooper, W.C., Ruhe, D.S., 1948. Studies in human malaria; the organization of a program for testing potential antimalarial drugs in prisoner volunteers. Am. J. Hyg. 47, 113–119.
Comfort, N., 2009. The prisoner as model organism: malaria research at Stateville Peniten-tiary. Stud. Hist. Philos. Biol. Biomed. Sci. 40, 190–203.

G6PD�Deficiency:�Global�Distribution,�Genetic�Variants�and�Primaquine�Therapy 195
Condon-Rall, M.E., 1992. U.S. Army medical preparations and the outbreak of war: the Philippines, 1941–6 May 1942. J. Mil. Hist. 56, 35–56.
Condon-Rall, M.E., 1994. The Army’s war against malaria: collaboration in drug research during World War II. Armed Forces Soc. 21, 129–143.
Craige, B., Eichelberger, L., Jones, R., Alving, A.S., Pullman, T.N., Whorton, C.M., 1948. The toxicity of large doses of pentaquine (Sn-13,276), a new antimalarial drug. J. Clin. Investig. 27, 17–24.
Craige Jr., B., Alving, A.S., et al., 1947. The Chesson strain of Plasmodium vivax malaria; relationship between prepatent period, latent period and relapse rate. J. Infect. Dis. 80, 228–236.
Crockett, M., Kain, K.C., 2007. Tafenoquine: a promising new antimalarial agent. Expert Opin. Investig. Drugs 16, 705–715.
Cromley, E.K., 2003. GIS and disease. Annu. Rev. Public Health 24, 7–24.Cuong, B.T., Binh, V.Q., Dai, B., Duy, D.N., Lovell, C.M., Rieckmann, K.H., Edstein, M.D.,
2006. Does gender, food or grapefruit juice alter the pharmacokinetics of primaquine in healthy subjects? Br. J. Clin. Pharmacol. 61, 682–689.
De Araujo, C., Migot-Nabias, F., Guitard, J., Pelleau, S., Vulliamy, T., Ducrocq, R., 2006. The role of the G6PD A-376G/968C allele in glucose-6-phosphate dehydrogenase defi-ciency in the seerer population of Senegal. Haematologica 91, 262–263.
Degowin, R.L., Eppes, R.B., Powell, R.D., Carson, P.E., 1966. The haemolytic effects of diaphenylsulfone (DDS) in normal subjects and in those with glucose-6-phosphate-dehydrogenase deficiency. Bull. W H O 35, 165–179.
Dern, R.J., Beutler, E., Alving, A.S., 1954a. The hemolytic effect of primaquine. II. The natu-ral course of the hemolytic anemia and the mechanism of its self-limited character. J. Lab. Clin. Med. 44, 171–176.
Dern, R.J., Weinstein, I.M., Leroy, G.V., Talmage, D.W., Alving, A.S., 1954b. The hemolytic effect of primaquine. I. The localization of the drug-induced hemolytic defect in prima-quine-sensitive individuals. J. Lab. Clin. Med. 43, 303–309.
Downs, W.G., Harper, P.A., Lisansky, E.T., 1947. Malaria and other insect-borne diseases in the South Pacific campaign, 1942–1945. II. Epidemiology of insect-borne diseases in Army troops. Am. J. Trop. Med. 27, 69–89.
Doxiadis, S.A., Valaes, T., 1964. The clinical picture of glucose 6-phosphate dehydrogenase deficiency in early infancy. Arch. Dis. Child 39, 545–553.
Earle, D.P., Bigelow, F.S., Zubrod, C.G., Kane, C.A., 1948. Studies on the chemotherapy of the human malarias. IX. Effect of pamaquine on the blood cells of man. J. Clin. Investig. 27, 121–129.
Edgcomb, J.H., Arnold, J., Yount Jr., E.H., Alving, A.S., Eichelberger, L., Jeffery, G.M., Eyles, D., Young, M.D., 1950. Primaquine, SN 13272, a new curative agent in vivax malaria; a preliminary report. J. Natl. Malar. Soc. 9, 285–292.
Editorial, 1952. Primaquine for vivax malaria. JAMA 149, 1573.Editorial, 1955. Drug-induced hemolytic anemia. JAMA 158, 310.Elderfield, R.C., Mertel, H.E., Mitch, R.T., Wempen, I.M., Werble, E., 1955. Synthesis of
primaquine and certain of its analogs. J. Am. Chem. Soc. 77, 4816–4819.Elmes, N.J., Bennett, S.M., Abdalla, H., Carthew, T.L., Edstein, M.D., 2006. Lack of
sex effect on the pharmacokinetics of primaquine. Am. J. Trop. Med. Hyg. 74, 951–952.
Elyazar, I.R., Hay, S.I., Baird, J.K., 2011. Malaria distribution, prevalence, drug resistance and control in Indonesia. Adv. Parasitol. 74, 41–175.
Eziefula, A.C., Gosling, R., Hwang, J., Hsiang, M.S., Bousema, T., von Seidlein, L., Drakeley, C., on behalf of the Primaquine in Africa Discussion Group, 2012. Rationale for short course primaquine in Africa to interrupt malaria transmission. Malaria J. 11, 360.
Fermi, C., Martinetti, P., 1905. Studio sul favismo. Annali di Igiene Sperimentale 15, 76–112.

R.�E.�Howes et al.196
Fernando, D., Rodrigo, C., Rajapakse, S., 2011. Primaquine in vivax malaria: an update and review on management issues. Malaria J. 10, 351.
Fiorelli, G., Martinez di Montemuros, F., Cappellini, M.D., 2000. Chronic non-spherocytic haemolytic disorders associated with glucose-6-phosphate dehydrogenase variants. Bail-liere’s Best Pract. Res. Clin. Haematol. 13, 39–55.
Flanagan, C.L., Schrier, S.L., Carson, P.E., Alving, A.S., 1958. The hemolytic effect of prima-quine. VIII. The effect of drug administration on parameters of primaquine sensitivity. J. Lab. Clin. Med. 51, 600–608.
Fletcher, K.A., Barton, P.F., Kelly, J.A., 1988. Studies on the mechanisms of oxidation in the erythrocyte by metabolites of primaquine. Biochem. Pharmacol. 37, 2683–2690.
Ganesan, S., Chaurasiya, N.D., Sahu, R., Walker, L.A., Tekwani, B.L., 2012. Understanding the mechanisms for metabolism-linked hemolytic toxicity of primaquine against glu-cose 6-phosphate dehydrogenase deficient human erythrocytes: evaluation of eryptotic pathway. Toxicology 294, 54–60.
Ganesan, S., Tekwani, B.L., Sahu, R., Tripathi, L.M., Walker, L.A., 2009. Cytochrome P(450)-dependent toxic effects of primaquine on human erythrocytes. Toxicol. Appl. Pharma-col. 241, 14–22.
Garrett, L., 2012. Global health hits crisis point. Nature 482, 7.Gething, P.W., Elyazar, I.R.F., Moyes, C.L., Smith, D.L., Battle, K.E., Guerra, C.A., Patil, A.P.,
Tatem, A.J., Howes, R.E., Myers, M.F., George, D.B., Horby, P., Wertheim, H.F.L., Price, R.N., Mueller, I., Baird, J.K., Hay, S.I., 2012. A long neglected world malaria map: Plas-modium vivax endemicity in 2010. PLoS Negl. Trop. Dis. 6, e1814.
Gomez-Gallego, F., Garrido-Pertierra, A., Mason, P.J., Bautista, J.M., 1996. Unproductive folding of the human G6PD-deficient variant A. Faseb. J. 10, 153–158.
Graves, P.M., Gelband, H., Garner, P., 2012. Primaquine for reducing Plasmodium falciparum transmission. Cochrane Database Syst. Rev. (Issue 9) Art. No.: CD008152.
Greaves, J., Evans, D.A., Gilles, H.M., Fletcher, K.A., Bunnag, D., Harinasuta, T., 1980. Plasma kinetics and urinary excretion of primaquine in man. Br. J. Clin. Pharmacol. 10, 399–404.
Greene, L.S., 1993. G6PD deficiency as protection against falciparum-malaria: an epide-miologic critique of population and experimental studies. Yearb. Phys. Anthropol. 36, 153–178.
Guerra, C.A., Howes, R.E., Patil, A.P., Gething, P.W., Van Boeckel, T.P., Temperley, W.H., Kabaria, C.W., Tatem, A.J., Manh, B.H., Elyazar, I.R., Baird, J.K., Snow, R.W., Hay, S.I., 2010. The international limits and population at risk of Plasmodium vivax transmission in 2009. PLoS Negl. Trop. Dis. 4, e774.
Guindo, A., Fairhurst, R.M., Doumbo, O.K., Wellems, T.E., Diallo, D.A., 2007. X-linked G6PD deficiency protects hemizygous males but not heterozygous females against severe malaria. PLoS Med. 4, e66.
Haldane, J.B.S., 1949. The rate of mutation of human genes. Hereditas 35, 267–273.Harcourt, B.E., 2011. Making willing bodies: the University of Chicago human experiments
at Stateville penitentiary. Soc. Res. 78, 443–478.Hardgrove, M., Applebaum, I.L., 1946. Plasmochin toxicity; analysis of 258 cases. Ann.
Intern. Med. 25, 103–112.Hardy, G.H., 1908. Mendelian proportions in a mixed population. Science 28, 49–50.Hay, S.I., Snow, R.W., 2006. The Malaria Atlas Project: developing global maps of malaria
risk. PLoS Med. 3, e473.Hedrick, P.W., 2011. Population genetics of malaria resistance in humans. Heredity 107,
283–304.Hill, D.R., Baird, J.K., Parise, M.E., Lewis, L.S., Ryan, E.T., Magill, A.J., 2006. Primaquine:
report from CDC expert meeting on malaria chemoprophylaxis I. Am. J. Trop. Med. Hyg. 75, 402–415.

G6PD�Deficiency:�Global�Distribution,�Genetic�Variants�and�Primaquine�Therapy 197
Hockwald, R.S., Arnold, J., Clayman, C.B., Alving, A.S., 1952. Toxicity of primaquine in Negroes. JAMA 149, 1568–1570.
Howes, R.E., Dewi, M., Piel, F.B., Hay, S.I., Baird, J.K. Geographic distribution of clini-cally significant G6PD deficiency variants within P. vivax malaria endemic countries, in preparation.
Howes, R.E., Patil, A.P., Piel, F.B., Nyangiri, O.A., Kabaria, C.W., Gething, P.W., Zimmerman, P.A., Barnadas, C., Beall, C.M., Gebremedhin, A., Menard, D., Williams, T.N., Weatherall, D.J., Hay, S.I., 2011. The global distribution of the Duffy blood group. Nature Commun. 2, 266.
Howes, R.E., Piel, F.B., Patil, A.P., Nyangiri, O.A., Gething, P.W., Hogg, M.M., Battle, K.E., Padilla, C.D., Baird, J.K., Hay, S.I., G6PD deficiency prevalence and estimates of affected populations in malaria endemic countries: a geostatistical model-based map. PLoS Med. 2012. 9, e1001339.
John, G.K., Douglas, N.M., von Seidlein, L., Nosten, F., Baird, J.K., White, N.J., Price, R.N., 2012. Primaquine radical cure of Plasmodium vivax: a critical review of the literature. Malaria J. 11, 280.
Johnson, M.K., Clark, T.D., Njama-Meya, D., Rosenthal, P.J., Parikh, S., 2009. Impact of the method of G6PD deficiency assessment on genetic association studies of malaria suscep-tibility. PLoS One 4, e7246.
Jones, R., Craige, B., Alving, A.S., Whorton, C.M., Pullman, T.N., Eichelberger, L., 1948. A study of the prophylactic effectiveness of several 8-aminoquinolines in sporozoite-induced vivax malaria (Chesson strain). J. Clin. Investig. 27, 6–11.
Joy, R.J., 1999. Malaria in American troops in the South and Southwest Pacific in World War II. Med. Hist. 43, 192–207.
Kellermeyer, R.W., Tarlov, A.R., Schrier, S.L., Carson, P.E., Alving, A.S., 1961. The hemolytic effect of primaquine. XIII. Gradient susceptibility to hemolysis of primaquine-sensitive erythrocytes. J. Lab. Clin. Med. 58, 225–233.
Kim, S., Nguon, C., Guillard, B., Duong, S., Chy, S., Sum, S., Nhem, S., Bouchier, C., Tichit, M., Christophel, E., Taylor, W.R., Baird, J.K., Menard, D., 2011. Performance of the CareStart G6PD deficiency screening test, a point-of-care diagnostic for primaquine therapy screening. PLoS One 6, e28357.
Kitchen, S.F., 1938. The infection of reticulocytes by Plasmodium vivax. Am. J. Trop. Med. Hyg. 18, 347–359.
Koliwad, S.K., Elliott, S.J., Kunze, D.L., 1996. Oxidized glutathione mediates cation channel activation in calf vascular endothelial cells during oxidant stress. J. Physiol. 495 (Pt 1), 37–49.
Krudsood, S., Tangpukdee, N., Wilairatana, P., Phophak, N., Baird, J.K., Brittenham, G.M., Looareesuwan, S., 2008. High-dose primaquine regimens against relapse of Plasmodium vivax malaria. Am. J. Trop. Med. Hyg. 78, 736–740.
Kwiatkowski, D.P., 2005. How malaria has affected the human genome and what human genetics can teach us about malaria. Am. J. Hum. Genet. 77, 171–192.
Kwok, C.J., Martin, A.C., Au, S.W., Lam, V.M., 2002. G6PDdb, an integrated database of glucose-6-phosphate dehydrogenase (G6PD) mutations. Hum. Mutat. 19, 217–224.
Lang, F., Lang, K.S., Lang, P.A., Huber, S.M., Wieder, T., 2006. Mechanisms and significance of eryptosis. Antioxid. Redox Signal. 8, 1183–1192.
Lang, P.A., Kaiser, S., Myssina, S., Wieder, T., Lang, F., Huber, S.M., 2003. Role of Ca2+-activated K+ channels in human erythrocyte apoptosis. Am. J. Physiol. Cell Physiol. 285, C1553–C1560.
Leslie, T., Briceno, M., Mayan, I., Mohammed, N., Klinkenberg, E., Sibley, C.H., Whitty, C.J., Rowland, M., 2010. The impact of phenotypic and genotypic G6PD deficiency on risk of Plasmodium vivax infection: a case-control study amongst Afghan refugees in Pakistan. PLoS Med. 7, e1000283.

R.�E.�Howes et al.198
Link, C.M., Theoharides, A.D., Anders, J.C., Chung, H., Canfield, C.J., 1985. Structure–activity relationships of putative primaquine metabolites causing methemoglobin for-mation in canine hemolysates. Toxicol. Appl. Pharmacol. 81, 192–202.
Longo, L., Vanegas, O.C., Patel, M., Rosti, V., Li, H., Waka, J., Merghoub, T., Pandolfi, P.P., Notaro, R., Manova, K., Luzzatto, L., 2002. Maternally transmitted severe glucose 6-phosphate dehydrogenase deficiency is an embryonic lethal. EMBO J. 21, 4229–4239.
Louicharoen, C., Patin, E., Paul, R., Nuchprayoon, I., Witoonpanich, B., Peerapittayamon-gkol, C., Casademont, I., Sura, T., Laird, N.M., Singhasivanon, P., Quintana-Murci, L., Sakuntabhai, A., 2009. Positively selected G6PD-Mahidol mutation reduces Plasmodium vivax density in Southeast Asians. Science 326, 1546–1549.
Luisada, A., 1940. Favism. JAMA 115, 632.Luisada, A., 1941. A singular disease chiefly affecting the red blood cells. Medicine 20,
229–250.Luzzatto, L., 1979. Genetics of red cells and susceptibility to malaria. Blood 54, 961–976.Luzzatto, L., 2004. Malaria and Darwinian selection in human populations. In: Keynes,
M., Edwards, A. W. F., Peel, R. (Eds.), A Century of Mendelism in Human Genetics, CRC Press, Boca Raton, Florida, pp. 73–84.
Luzzatto, L., 2006. Glucose 6-phosphate dehydrogenase deficiency: from genotype to phe-notype. Haematologica 91, 1303–1306.
Luzzatto, L., 2009. Glucose-6-phosphate dehydrogenase deficiency. In: Orkin, S.H., Nathan, D.G., Ginsburg, D. (Eds.), Nathan and Oski’s Hematology of Infancy and Childhood, Saunders, Philadelphia.
Luzzatto, L., 2010. Glucose 6-phosphate dehydrogenase deficiency. In: Warrell, D., Cox, T.M., Firth, J.D. (Eds.), Oxford Textbook of Medicine, OUP, Oxford, pp. 4474–4479.
Luzzatto, L., Mehta, A., Vulliamy, T.J., 2001. Glucose-6-phosphate dehydrogenase defi-ciency. eighth ed. In: Scriver, C.R., Beaudet, A.L., Sly, W.S., Valle, D. (Eds.), The Meta-bolic and Molecular Bases of Inherited Disease, vol. iii. McGraw-Hill Inc, New York, pp. 4517–4553.
Luzzatto, L., Notaro, R., 2001. Malaria. Protecting against bad air. Science 293, 442–443.Luzzatto, L., Usanga, F.A., Reddy, S., 1969. Glucose-6-phosphate dehydrogenase deficient
red cells: resistance to infection by malarial parasites. Science 164, 839–842.Lyon, M.F., 1961. Gene action in the X-chromosome of the mouse (Mus musculus L.). Nature
190, 372–373.Martini, G., Toniolo, D., Vulliamy, T., Luzzatto, L., Dono, R., Viglietto, G., Paonessa, G.,
D’Urso, M., Persico, M.G., 1986. Structural analysis of the X-linked gene encoding human glucose 6-phosphate dehydrogenase. EMBO J. 5, 1849–1855.
Mason, P.J., Bautista, J.M., Gilsanz, F., 2007. G6PD deficiency: the genotype-phenotype asso-ciation. Blood. Rev. 21, 267–283.
Mason, P.J., Vulliamy, T.J., 2005. Glucose-6-phosphate dehydrogenase (G6PD) deficiency: genetics. Encyclopedia of Life Sciences, John Wiley & Sons, Ltd.
McLafferty, S.L., 2003. GIS and health care. Annu. Rev. Public Health 24, 25–42.Mehta, A., Mason, P.J., Vulliamy, T.J., 2000. Glucose-6-phosphate dehydrogenase deficiency.
Bailliere’s Best Pract. Res. Clin. Haematol. 13, 21–38.Meloni, T., Forteleoni, G., Dore, A., Cutillo, S., 1983. Favism and hemolytic anemia in glu-
cose-6-phosphate dehydrogenase-deficient subjects in North Sardinia. Acta Haemato-logica 70, 83–90.
Mihaly, G.W., Ward, S.A., Edwards, G., Nicholl, D.D., Orme, M.L., Breckenridge, A.M., 1985. Pharmacokinetics of primaquine in man. I. Studies of the absolute bioavailability and effects of dose size. Br. J. Clin. Pharmacol. 19, 745–750.
Minucci, A., Giardina, B., Zuppi, C., Capoluongo, E., 2009. Glucose-6-phosphate dehydro-genase laboratory assay: how, when, and why? IUBMB Life 61, 27–34.

G6PD�Deficiency:�Global�Distribution,�Genetic�Variants�and�Primaquine�Therapy 199
Minucci, A., Moradkhani, K., Hwang, M.J., Zuppi, C., Giardina, B., Capoluongo, E., 2012. Glucose-6-phosphate dehydrogenase (G6PD) mutations database: review of the “old” and update of the new mutations. Blood Cells Mol. Dis. 48, 154–165.
Most, H., Kane, C.A., et al., 1946. Combined quinine-plasmochin treatment of vivax malaria; effect of relapse rate. Am. J. Med. Sci. 212, 550–560.
Motulsky, A.G., 1960. Metabolic polymorphisms and the role of infectious diseases in human evolution. Hum. Biol. 32, 28–62.
Mühlens, V.P., 1926. Die Behandlung der natürlichen menschlichen Malaria-Infektion mit Plasmochin. Naturwissenschaften 14, 1162–1166.
Myat Phone, K., Myint, O., Aung, N., Aye Lwin, H., 1994. The use of primaquine in malaria infected patients with red cell glucose-6-phosphate dehydrogenase (G6PD) deficiency in Myanmar. Southeast Asian J. Trop. Med. Public Health 25, 710–713.
Myint, H.Y., Berman, J., Walker, L., Pybus, B., Melendez, V., Baird, J.K., Ohrt, C., 2011. Review: improving the therapeutic index of 8-aminoquinolines by the use of drug combinations: review of the literature and proposal for future investigations. Am. J. Trop. Med. Hyg. 85, 1010–1014.
Nkhoma, E.T., Poole, C., Vannappagari, V., Hall, S.A., Beutler, E., 2009. The global preva-lence of glucose-6-phosphate dehydrogenase deficiency: a systematic review and meta- analysis. Blood Cells Mol. Dis. 42, 267–278.
Notaro, R., Afolayan, A., Luzzatto, L., 2000. Human mutations in glucose 6-phosphate dehy-drogenase reflect evolutionary history. Faseb. J. 14, 485–494.
Office of the Surgeon General, 1943. The drug treatment of malaria, suppressive and clinical. Circular letter no. 153 JAMA 123, 205–208.
Pamba, A., Richardson, N.D., Carter, N., Duparc, S., Premji, Z., Tiono, A.B., Luzzatto, L., 2012. Clinical spectrum and severity of hemolytic anemia in glucose 6-phosphate dehy-drogenase-deficient children receiving dapsone. Blood.
Pandolfi, P.P., Sonati, F., Rivi, R., Mason, P., Grosveld, F., Luzzatto, L., 1995. Targeted disrup-tion of the housekeeping gene encoding glucose 6-phosphate dehydrogenase (G6PD): G6PD is dispensable for pentose synthesis but essential for defense against oxidative stress. EMBO J. 14, 5209–5215.
Patil, A.P., Gething, P.W., Piel, F.B., Hay, S.I., 2011. Bayesian geostatistics in health cartogra-phy: the perspective of malaria. Trends Parasitol. 27, 246–253.
Percy, M.J., McFerran, N.V., Lappin, T.R., 2005. Disorders of oxidised haemoglobin. Blood Rev. 19, 61–68.
Persico, M.G., Viglietto, G., Martini, G., Toniolo, D., Paonessa, G., Moscatelli, C., Dono, R., Vulliamy, T., Luzzatto, L., D’Urso, M., 1986. Isolation of human glucose-6-phosphate dehydrogenase (G6PD) cDNA clones: primary structure of the protein and unusual 5’ non-coding region. Nucleic Acids Res. 14, 2511–2522.
Peters, A.L., Van Noorden, C.J., 2009. Glucose-6-phosphate dehydrogenase deficiency and malaria: cytochemical detection of heterozygous G6PD deficiency in women. J. Histo-chem. Cytochem. 57, 1003–1011.
Piomelli, S., Corash, L.M., Davenport, D.D., Miraglia, J., Amorosi, E.L., 1968. In vivo lability of glucose-6-phosphate dehydrogenase in GdA- and GdMediterranean deficiency. J. Clin. Investig. 47, 940–948.
Price, R.N., Tjitra, E., Guerra, C.A., Yeung, S., White, N.J., Anstey, N.M., 2007. Vivax malaria: neglected and not benign. Am. J. Trop. Med. Hyg. 77, 79–87.
Pybus, B.S., Sousa, J.C., Jin, X., Ferguson, J.A., Christian, R.E., Barnhart, R., Vuong, C., Sciotti, R.J., Reichard, G.A., Kozar, M.P., Walker, L.A., Ohrt, C., Melendez, V., 2012. CYP450 phenotyping and accurate mass identification of metabolites of the 8-amino-quinoline, anti-malarial drug primaquine. Malaria J. 11, 259.
Recht, J., Ashley, E., White, N.J., 2012. 8-aminoquinolines Safety Review for WHO Prima-quine ERG, August 2012 (unpublished).

R.�E.�Howes et al.200
Roth Jr., E.F., Raventos-Suarez, C., Rinaldi, A., Nagel, R.L., 1983. Glucose-6-phosphate dehydrogenase deficiency inhibits in vitro growth of Plasmodium falciparum. Proc. Natl. Acad. Sci. U S A 80, 298–299.
Ruwende, C., Hill, A., 1998. Glucose-6-phosphate dehydrogenase deficiency and malaria. J. Mol. Med. (Berlin) 76, 581–588.
Ruwende, C., Khoo, S.C., Snow, R.W., Yates, S.N., Kwiatkowski, D., Gupta, S., Warn, P., Allsopp, C.E., Gilbert, S.C., Peschu, N., et al., 1995. Natural selection of hemi- and het-erozygotes for G6PD deficiency in Africa by resistance to severe malaria. Nature 376, 246–249.
Sabeti, P.C., Reich, D.E., Higgins, J.M., Levine, H.Z., Richter, D.J., Schaffner, S.F., Gabriel, S.B., Platko, J.V., Patterson, N.J., McDonald, G.J., Ackerman, H.C., Campbell, S.J., Altshuler, D., Cooper, R., Kwiatkowski, D., Ward, R., Lander, E.S., 2002. Detecting recent positive selection in the human genome from haplotype structure. Nature 419, 832–837.
Saha, N., Samuel, A.P., 1991. Characterization of glucose-6-phosphate dehydrogenase vari-ants in the Sudan–including Gd Khartoum, a hyperactive slow variant. Hum. Hered. 41, 17–21.
Salvador, A., Savageau, M.A., 2003. Quantitative evolutionary design of glucose 6-phosphate dehydrogenase expression in human erythrocytes. Proc. Natl. Acad. Sci. U S A 100, 14463–14468.
Schmidt, L.H., Fradkin, R., Vaughan, D., Rasco, J., 1977. Radical cure of infections with Plasmodium cynomolgi: a function of total 8-aminoquinoline dose. Am. J. Trop. Med. Hyg. 26, 1116–1128.
Shah, S.S., Diakite, S.A.S., Traore, K., Diakite, M., Kwiatkowski, D.P., Rockett, K.A., Wellems, T.E., Fairhurst, R.M., 2012. A novel cytofluorometric assay for the detection and quanti-fication of glucose-6-phosphate dehydrogenase deficiency. Scientific Rep. 2.
Shanks, G.D., Oloo, A.J., Aleman, G.M., Ohrt, C., Klotz, F.W., Braitman, D., Horton, J., Brueckner, R., 2001. A new primaquine analogue, tafenoquine (WR 238605), for pro-phylaxis against Plasmodium falciparum malaria. Clin. Infect. Dis. 33, 1968–1974.
Shekalaghe, S., Drakeley, C., Gosling, R., Ndaro, A., van Meegeren, M., Enevold, A., Alifrangis, M., Mosha, F., Sauerwein, R., Bousema, T., 2007. Primaquine clears submicroscopic Plasmodium falciparum gametocytes that persist after treatment with sulphadoxine–pyrimethamine and artesunate. PLoS One 2, e1023.
Shekalaghe, S.A., ter Braak, R., Daou, M., Kavishe, R., van den Bijllaardt, W., van den Bosch, S., Koenderink, J.B., Luty, A.J., Whitty, C.J., Drakeley, C., Sauerwein, R.W., Bousema, T., 2010. In Tanzania, hemolysis after a single dose of primaquine coadministered with an artemisinin is not restricted to glucose-6-phosphate dehydrogenase-deficient (G6PD A-) individuals. Antimicrob. Agents Chemother. 54, 1762–1768.
Silva, M.C., Santos, E.B., Costal, E.G., Filho, M.G., Guerreiro, J.F., Povoa, M.M., 2004. [Clini-cal and laboratorial alterations in Plasmodium vivax malaria patients and glucose-6-phos-phate dehydrogenase deficiency treated with primaquine at 0.50 mg/kg/day]. Rev. Soc. Bras. Med. Trop. 37, 215–217.
Simoons, F.J., 1998. Plants of Life, Plants of Death. The University of Wisconsin Press, 216–249.
Slatkin, M., 2008. A Bayesian method for jointly estimating allele age and selection intensity. Genet. Res. 90, 129–137.
Song, J., Socheat, D., Tan, B., Dara, P., Deng, C., Sokunthea, S., Seila, S., Ou, F., Jian, H., Li, G., 2010. Rapid and effective malaria control in Cambodia through mass administration of artemisinin-piperaquine. Malaria J. 9, 57.
Takeuchi, R., Lawpoolsri, S., Imwong, M., Kobayashi, J., Kaewkungwal, J., Pukrittayakamee, S., Puangsa-art, S., Thanyavanich, N., Maneeboonyang, W., Day, N.P., Singhasivanon, P., 2010. Directly-observed therapy (DOT) for the radical 14-day primaquine treatment of Plasmodium vivax malaria on the Thai-Myanmar border. Malaria J. 9, 308.

G6PD�Deficiency:�Global�Distribution,�Genetic�Variants�and�Primaquine�Therapy 201
Takizawa, T., Huang, I.Y., Ikuta, T., Yoshida, A., 1986. Human glucose-6-phosphate dehy-drogenase: primary structure and cDNA cloning. Proc. Natl. Acad. Sci. U S A 83, 4157–4161.
Tantular, I.S., Kawamoto, F., 2003. An improved, simple screening method for detection of glucose-6-phosphate dehydrogenase deficiency. Trop. Med. Int. Health 8, 569–574.
Tinley, K.E., Loughlin, A.M., Jepson, A., Barnett, E.D., 2010. Evaluation of a rapid qualitative enzyme chromatographic test for glucose-6-phosphate dehydrogenase deficiency. Am. J. Trop. Med. Hyg. 82, 210–214.
Tishkoff, S.A., Varkonyi, R., Cahinhinan, N., Abbes, S., Argyropoulos, G., Destro-Bisol, G., Drousiotou, A., Dangerfield, B., Lefranc, G., Loiselet, J., Piro, A., Stoneking, M., Tagarelli, A., Tagarelli, G., Touma, E.H., Williams, S.M., Clark, A.G., 2001. Haplotype diversity and linkage disequilibrium at human G6PD: recent origin of alleles that confer malarial resistance. Science 293, 455–462.
Tripathy, V., Reddy, B.M., 2007. Present status of understanding on the G6PD deficiency and natural selection. J. Postgrad. Med. 53, 193–202.
UCL Bioinformatics Group website, Andrew C. R. Martin’s Bioinformatics Group at UCL. URL: http://www.bioinf.org.uk/g6pd/db/.
Vulliamy, T., Luzzatto, L., Hirono, A., Beutler, E., 1997. Hematologically important muta-tions: glucose-6-phosphate dehydrogenase. Blood Cells Mol. Dis. 23, 302–313.
Vulliamy, T.J., D’Urso, M., Battistuzzi, G., Estrada, M., Foulkes, N.S., Martini, G., Calabro, V., Poggi, V., Giordano, R., Town, M., et al., 1988. Diverse point mutations in the human glucose-6-phosphate dehydrogenase gene cause enzyme deficiency and mild or severe hemolytic anemia. Proc. Natl. Acad. Sci. U S A 85, 5171–5175.
Weinberg, W., 1908. Über den nachweis der vererbung beim menschen. Jahreshefte des Ver-eins für vaterländische Naturkunde in Württemberg 64, 368–382.
WHO, 2010. Guidelines for the Treatment of Malaria, second ed.WHO, 2011a. Country Antimalarial Drug Policies: By Region. URL: http://www.who.int/
malaria/am_drug_policies_by_region_afro/en/index.html.WHO, 2011b. Global Plan for Artemisinin Resistance Containment (GPARC).WHO Working Group, 1989. Glucose-6-phosphate dehydrogenase deficiency. Bull. W H O
67, 601–611.Wilairatana, P., Tangpukdee, N., Kano, S., Krudsood, S., 2010. Primaquine administration
after falciparum malaria treatment in malaria hypoendemic areas with high incidence of falciparum and vivax mixed infection: pros and cons. Korean J. Parasitol. 48, 175–177.
Yoshida, A., Beutler, E., Motulsky, A.G., 1971. Human glucose-6-phosphate dehydrogenase variants. Bull. W H O 45, 243–253.
Youngster, I., Arcavi, L., Schechmaster, R., Akayzen, Y., Popliski, H., Shimonov, J., Beig, S., Berkovitch, M., 2010. Medications and glucose-6-phosphate dehydrogenase deficiency: an evidence-based review. Drug Saf. 33, 713–726.
Ziai, M., Amirhakimi, G.H., Reinhold, J.G., Tabatabee, M., Gettner, M.E., Bowman, J.E., 1967. Malaria prophylaxis and treatment in G-6-PD deficiency. An observation on the toxicity of primaquine and chloroquine. Clin. Pediatr. 6, 242–243.