Embed Size (px)
Citation preview

7
CHAPTER 2
REVIEW OF LITERATURE
Rice (Oryza sativa L.), a member of family Poaceae and subfamily Oryzoidea,
is one of the major staple food consumed by 70% of the world’s population. It occupies
one fifth of the total land covered under cereal crops (Chakravarthi and Naravaneni,
2006). Rice, which is mainly, consumed as a whole grain supplies 20% of daily calories
for the world population (http://apps.fao.org). The adaptation of rice cultivars to wide
climatic conditions has led to the development of thousands of rice varieties having
diverse quality features in terms of physical, cooking, eating and product development
characteristics (Bhattacharya, 2005). Moreover, rice as a model crop with fully
sequenced genome, relatively small genome size, significant level of polymorphism
(McCouch et al., 1988; Wang et al., 1992) and large amount of well conserved
genetically diverse material, gives unique opportunities to researcher for studying its
varietal difference and history of crop improvement using different approaches.
2.1 Classification of Rice Varieties
The genus Oryza has twenty-one wild and two cultivated species, i.e. Oryza
sativa (Asian rice) and Oryza glaberrima (African rice). Rice varieties belong to O.
sativa spp. grown across the world, while the cultivation of the O. glaberrima has been
confined to Africa. O. glaberrima varieties are fastly being replaced by the O. sativa
due to weed tolerance, pest resistance and fast maturing growth attributes of later
varieties. Based on number of morphological, physiological, biochemical and molecular
traits, Asian rice cultivars are broadly classified into two major subspecies, i.e. Oryza

8
sativa japonica and Oryza sativa indica (Second et al., 1982; Glaszmann et al., 1987) as
shown in Figure 1. In addition to these major varieties, the minor groups include Aus
cultivars of India and Bangladesh, Ashinas varieties of Bangladesh and aromatic
basmati varieties of India (Garris et al., 2005).
Figure 2.1 Classification of cultivated rice species
These groups may be further subdivided into more types like aromatic and non-
aromatic, waxy and non-waxy etc. based on their distinct features. Each type of rice
cultivar has different market within same or different region. Moreover, each type has
special quality demands for specific use depending upon the consumer choice and type
of food product.
Cultivated Rice Species
Asian African
Oryza sativa Oryza glaberrima
(Glaszmann, 1987) (Garris et al., 2005) (Chang, 1976)
Indica, Aus, Ashinas,
Rayadas, Aromatic,
Japonica
Indica, Aus, Aromatic,
Temperate Japonica,
Tropical Japonica
Indica, Japonica,
Javanica

9
2.2 Indigenous Rice Cultivars
The archaeological evidence showed that the cultivation of rice in India has been
started in between 1500 and 1000 B.C. With its long history of cultivation and selection
under diverse environments, rice acquired wide adaptability enabling it to grow in wide
range of environmental conditions. About 500,000 accessions of rice genetic resources
are conserved all over the world. The largest six gene banks are in Asia, and together
conserve around 70% of total world rice germplasm. They are International Rice
Research Institution (IRRI) in Philippines, National Bureau of Plant Genetic Resources
(NBPGR) in India, Institute of Crop Germplasm Resources (CAAS) in China, China
National Rice Research Institute (CNRRI), National Institute of Agrobiological
Sciences (NIAS) in Japan, and Rural Development Administration (RDA) gene bank in
the Republic of Korea. India alone has 86,330 accessions, of which 42,004 are in the
national gene bank (Rai, 1999), which is enriched by further explorations, collections
and conservation.
Among tropical Asian countries, India has released the greatest number of
varieties over the last 50 years (www.fao.org). The indigenous rice cultivars have been
enriched with extensive genetic diversity, thus serve as a source for genetic
improvements of cultivated rice varieties (Choudhury et al., 2013). Basmati, the unique
aromatic quality rice is a nature’s gift to Indian sub-continent. The word ‘Basmati’ has
been derived from two words, i.e. ‘Bas’ in Hindi language means “Aroma” and ‘Mati’
means “full of”’, hence basmati, i.e. full of aroma. This rice variety is different from
other varieties because of aroma and post cooking elongation attributes. No other rice
has these combined characteristics. The other distinct characteristics of basmati rice
includes extra long slender grain, lengthwise excessive elongation on cooking, soft and
fluffy texture of cooked rice grain. It has also been reported that the unique quality traits

10
of basmati rice found their gene expression only when they are grown in the north-
western foot hills of the Himalayas in the Indian sub-continent. Due to the geographic
specific manifestation of quality features, basmati is now a Geographical Indication
(GI) belonging to a specific geographical area in the Indian subcontinent grown by
farmers of Punjab, Haryana, Uttar Pradesh and Punjab state of Pakistan for more than
250 years (Nene, 1998; Siddiq et al., 2012).
The market of basmati rice is less than 10 percent of the global rice market. But,
basmati rice captures higher returns as it is priced three times higher (US$ 800-1200 per
metric ton) over non-basmati rice (US$ 200-400 per metric ton) in the international as
well as in the domestic markets. Non- basmati rice has also become a thrust area for
export promotion though there have been fluctuating trends (Rice Knowledge
Management Portal).
In spite of high quality traditional basmati varieties in India, the research is still
ongoing for the development of many new varieties of rice with better quality and
agronomic traits to meet the consumer and farmers demand. The recently developed
basmati varieties are Pusa Basmati 1460 or Improved Pusa Basmati 1 (the first biotech
rice variety), Pusa Basmati 1401 (also known as Pusa Basmati 6) and RH 10 (India’s
first hybrid basmati variety), which are having quality and agronomic features at par or
even better than traditional basmati varieties (http://rkmp.iari.res.in/IARIrole.aspx).
The characterization of these cultivars from cooking and eating quality aspects is
mandatory, as the development of such varieties will lead to a major revolution in the
basmati rice sector for export and quality purposes.
Studies on characterization of quality attributes of traditional; developed and
under development rice varieties are mandatory for the improvement in rice quality
worldwide and to meet the increasingly alternative demands of consumers everywhere.

11
2.3 Multivariate Methods for Characterization of Rice Cultivars
Characterization and quantification of genetic diversity has been a major goal in
evolutionary biology. Characterization is a critical step to be carried out to identify
accessions to find genetic relationships among genotypes and to stop the malpractices of
adulteration. The rice cultivation depends on cultural practices and management of
existing germplasm, which has been collected and conserved by genetic resources
centers (Ng et al., 1988). However, the utilization of rice genetic resources had been
limited to only selected and adaptable genotypes (Caldo et al., 1996). As a result,
the diversity of these genetic resources is being lost to the need for higher yields
and early maturity. However a successful breeding programme will depend on the
genetic diversity of a crop for achieving the goals of improving the crop and
producing high yielding and better resistant varieties (Padulosi, 1993). Therefore,
there is the need to diversify the genetic base of improved rice varieties, and the
first step towards this is to evaluate and characterize available rice germplasm or
genotypes at both the morphological and molecular levels (Ogunbayo et al., 2005).
Systematic study and characterization of high quality germplasm is not only important
for utilizing the appropriate attribute based donors, but also essential in the present era
for protecting the unique rice. Moreover, it is also an essential step to prevent
adulteration of low priced, non-aromatic rice with high priced aromatic rice and
consequently dissemination of right information to the consumers
There are different approaches to achieve characterization of various species of
rice cultivars on basis of its different forms (paddy, milled rice, rice flour etc.). The
different methods of characterization include morphology based, physico-chemical
based, biochemical based and molecular based characterization (Chakravarthi and
Naravaneni, 2006) as shown in Figure 2.2.

12
2.3.1 Characterization of Rough Rice/Paddy
A large amount of paddy have been vanished off due to the adoption of old and
outdated methods of drying and milling and furthermore by the use of improper and
unscientific methods of storage, transport and handling (http://www.rkmp.co.in).
Studies on morphological, physical and engineering properties of different paddy
cultivars as well as other cereal crops are important to handle many problems
associated with the design of machines, the analysis of the behavior of the product
during agricultural process operations such as handling, planting, harvesting, threshing,
cleaning, sorting and drying (Varnamkhasti et al., 2007; Abano and Amoah, 2011).
Figure 2.2 Different indicators for characterization of paddy, rice grains and flour
Milling yield is one of the most important criteria of rice quality especially from
a marketing standpoint. A variety should possess a high turnout of whole grain (head)
rice and total milled rice (Webb, 1985). Milling yield of rough rice is the estimate of the
Indicators of Characterization
Paddy Milled Grains Rice Flour
Morphology based
Physical attributes
Engineering attributes
Biochemical based
Molecular based
Morphology based
Physical attributes
Cooking Properties
Sensory evaluation
Molecular based
Morphology based
Physico-chemical based
Thermal and Rheological
Properties
Biochemical based
Product making attributes

13
quantity of head rice and total milled rice that can be produced from a unit of rough rice
(Khush et al., 1979). Thus, the milling quality of rice may be defined as the ability of
rice grain to stand milling and polishing without undue breakage to yield the greatest
amount of total recovery and the highest proportion of head rice to broken rice. The
three main dimensions (length, width and thickness) of paddy grains are useful in
selecting sieve separators to calculate surface area and volume of kernels which are
important during modeling of grain drying, aeration, heating and cooling. It is also
further required in the designing of particular equipment for handling and storage of
grains (Zareiforoush et al., 2009).
The head rice yield, an important parameter of paddy from market point of view
can be calculated with 1000 grain weight of paddy grains. Other parameters like density
and porosity can be useful in studies related to the rate of heat and mass transfer of
moisture during aeration and drying processes through the bulk grain (Correa et al.,
2007). Densities of different types of cereal grain kernels are of paramount interest in
breakage susceptibility and hardness studies (Seifi and Alimardani, 2010). Moreover,
bulk density, true density and porosity (the ratio of inter granular space to the total
space occupied by the grain) can be useful in sizing grain hoppers and storage facilities;
they can affect the rate of heat and mass transfer of moisture during aeration and drying
processes.
The parameters related with frictional properties (angle of repose and coefficient
of friction) of cereal grains on different surfaces are very helpful in design of hoppers
based on the flow ability of agricultural grains and also in safe designing of equipments
related with processing, moving and storage of grains (Mohsenin, 1986; Varnamkhasti
et al., 2008). The coefficient of friction for different surfaces (plywood, glass, steel) is
necessary in designing of conveying structures (Altuntas and Karadag, 2006).

14
A number of experiments have been conducted by researchers to compare the
physical and engineering properties of different rice cultivars (Arora, 1991; Kachru et
al., 1994; Reddy and Chakraverty, 2004; Correa et al., 2007; Varnamkhasti et al., 2007;
Zareiforoush et al., 2009; Kanchana et al., 2012).
2.3.2 Characterization of Milled Rice Grains
The marketing values of rice as an agricultural product depend on its physical
qualities after post harvesting and milling. The appearance of rice with bran (brown)
and without bran (white) is important to the consumers. Rice is marketed according to
quality attributes of raw and cooked rice grains. The quality of rice grain assumes
greater importance because most of the rice produced is cooked and consumed as a
whole kernel., whereas the percentage of rice converted into flour or flakes being very
small (Huang et al.,1998). The genetic diversity among rice cultivars from quality point
of view can be assessed in three broad categories, i.e. physical qualities, processing
qualities and nutritional qualities (Thongbam et al., 2011).
2.3.2.1 Physical Characterization
The rice grain length, width and weight mainly determine the physical quality of
rice grain. The classification of rice quality is based on the length of grain, i.e. short,
medium and long grain. Grain shape is an important agronomic trait in cereal crops
because it is directly or indirectly related to quality and quantity of grain products
(Ayoub et al., 2002; Breseghello and Sorrels, 2006; Zheng et al., 2007). Moreover,
grain size and shape are the first quality characteristics considered in developing new
varieties. The length to breadth ratio ranging from 2.5 to 3.0 is widely acceptable and
the grain length > 6 mm is preferred (Kaul, 1970). The size and shape of the rice grain

15
may vary from one group to another group of different regions. Some groups prefer
short grains; some prefer medium grains and mostly the peoples like long slender grains
in Indian sub-continent regions and in international market. Medium short grains are
preferred by the peoples of South Asian regions (Singh et al., 2000). Thus, components
of appearance traits are one of the first criteria for rice quality breeders in release and
commercial production of rice varieties (Dela and Khush, 2000).
The ratio of length to width is also an important aspect to determine the shape of
the variety (Normita and Cruz, 2002) whereas grain weight gives the information about
the size and density of the grain. The density of different rice grains effect the cooking
quality.
2.3.2.2 Chemical Characterization
The most important quality criterion based on chemical characteristics of rice
grain and flour of different cultivars is moisture, carbohydrate, protein, fat, crude fiber
and ash contents. The moisture content during storage showed changes with the length
of storage and the temperature of storage (Gooding and Davies, 1997). The moisture
content is also important aspect because all grains have to be stored for a certain period
before their end use. The grains having high moisture content are difficult to store safely
because these are more susceptible to attack a pests and diseases (Gooding and Davies,
1997). Proteins and starch are the two major components of rice, with approximately 8
and 80% composition, respectively (Marshall and Wordsworth, 1994). Rice protein is
more nutritious because of its higher lysine content than any other cereal proteins (Bean
and Nishita, 1985). Micro-kjeldahl analysis and different methods are used to determine
the protein content in rice. Fat is the main source of food energy. Rather than giving
energy it also helps in the absorption of fat soluble vitamins. Rice contains very low

16
amount of fat but cooked brown rice contains higher fat contents than white rice. The
ash content of a food sample gives an idea of the mineral elements present in the food
sample.
2.3.2.3 Cooking, Textural, Pasting and Thermal Properties
The rice produced in different parts of India varies significantly in composition
and cooking quality. Genetic and environmental factors are mainly responsible for
variation in composition and cooking quality of rice (Singh et al., 2005). Consumer
preference regarding cooking attributes of rice grains may vary from region to region.
Japanese like sticky rice (Deshpande and Bhattacharya, 1982), while Italians consume
short grain variety, which releases starch during cooking making a creamy and smooth
risotto, Americans prefer a semi milled long grain rice or even brown rice, whereas
Asian dominates spicy and scented Basmati/Jasmine rice and the people of Indian sub-
continent prefer a well milled white rice (Lyon et al., 1999).
Cooking and eating characteristics are largely determined by the properties of
the starch that makes up 90% of milled rice. Gelatinization temperature, amylose
content and gel consistency are the important starch properties, which influence cooking
and eating characteristics of rice grains (Figure 2.3).
Amylose content: Amylose content (AC) is considered as the most important
character for predicting rice cooking and processing behavior (Juliano 1979; Webb,
1985). Many of the cooking and eating characteristics of milled rice are influenced by
the ratio of two kinds of starches, i.e. amylose and amylopectin (Juliano et al., 1964).
Amylose is the linear fraction of starch in the non-glutinous varieties, where as
amylopectin, the branched fraction, makes up the reminder of the starch. Amylose is

17
almost absent from the waxy (glutinous) rice cultivars which do not expand in volume,
are glossy and sticky and remain firm when cooked (Juliano, 1979). High amylose rice
grains show high volume expansion and a high degree of flakiness. They cook dry, are
less tender and become hard upon cooling. Rice grains with intermediate amylose
content cook moist and tender and do not become hard upon cooling. Rice varieties are
grouped on the basis of their amylose content into waxy (0-2%); very low (3-9%), low
(10-19%), intermediate (20-25%) and high with >25% amylose content (Kumar and
Khush, 1986).
Gelatinization Temperature: The different diffusive processes like heating, water
uptake and swelling of the rice particle take place during the cooking of rice grains
(Oko et al., 2012). Starch granules are heated in water beyond a critical temperature; the
granules absorb a large amount of water and swell to many times their original size.
Over a critical temperature range, the starch granules undergo an irreversible process
known as gelatinization, which is marked by crystalline melting, loss of birefringence
and starch solubilization (Rani et al., 2006). As soon as the temperature rises above the
gelatinization temperature (GT), the starch granules begin to swell and viscosity
increases. The gelatinization temperature provides an indication of the minimum
temperature required to cook for a given rice sample. The gelatinization temperature of
rice varieties may be classified as low (55 to 69 oC), intermediate (70 to 74
oC) and high
with >74 oC (Rani et al., 2006). The gelatinization temperature can be estimated by
alkali digestibility test (Little et al., 1958) in which the degree of spreading value of
individual milled rice kernels in a weak alkali solution (1.7% KOH) is very closely
correlated with gelatinization temperature. Rice with low gelatinization temperature
disintegrates completely, whereas rice with intermediate gelatinization temperature

18
shows only partial disintegration. Rice with high gelatinization temperature remains
largely unaffected in the alkali solution.
Linear elongation of rice on cooking is one of the major characteristics of good
rice. Some varieties expand more in size than others upon cooking. Length-wise
expansion without increase in girth is considered a highly desirable trait of high quality
rice (Sood and Sadiq, 1979). Soaked milled rice of high gelatinization temperature
elongates less during cooking than low and intermediate gelatinizing rice. Thus,
gelatinization temperature correlates positively with grain elongation (Perez et al.,
1987). However, gelatinization temperature does not correlate with the texture of
cooked rice (Rani et al., 2006).
Figure 2.3 Effect of amylose content, gel consistency and gelatinization
temperature on cooked rice grains
Gel Consistency: The texture of cooked rice grains is mainly dependent upon the
amylose content, however the cohesiveness, tenderness, colour and gloss of cooked
Amylose Content
• Effect the texture of cooked rice. Rice with high AC cook dry, less tender and hard, whereas with low AC are soft and sticky
Gelatinization Temperature
• Determines the time required for cooking the rice
Gel Consistency
• Measures the tendency of cooked rice to harden on cooling

19
grains is greatly based on gel consistency (GC), when the amylose content is high.
Varietal differences in gel consistency exist among varieties of similar amylose content
(Cagampang et al., 1973).
The gel consistency test is based on the consistency of the rice paste and
differentiates among varieties with high amylose content. It measures the tendency of
cooked rice to harden when it cools down. The test separates high amylose rice grains
into three categories (Singh et al., 2000):
(a) Very flaky rice grains with hard gel consistency (length of gel, 40 mm or less);
(b) Flaky rice grains with medium GC (length of gel, 41 to 60);
(c) Soft rice grains with soft gel consistency (length of gel more than 61 mm).
Cooked Kernel Elongation: The elongation ratio and elongation index are the
important cooking quality traits of rice and are much preferred by the consumers
(Parikh et al., 2012). The quality of cooked rice grain is mainly associated with its
length wise expansion as compared to widthwise (Khush et al., 1979). It has been also
observed that if rice elongates more lengthwise, it gives a finer appearance whereas
gives coarse look if expands girthwise (Srivastava and Jaiswal, 2013).
Aroma: Aromatic long grain basmati rices are known for their pleasant aroma, which
plays a role in its consumer acceptability and it draws a premium price in certain
specialty markets (Sood et al., 1983; Ahn et al., 1993). Most of the high quality and
preferred varieties in rice growing countries are aromatic in nature (Singh et al., 2000).
Jasmine rice from Thailand, basmati rice from Pakistan and India whereas Della from
U.S.A are the main sources of aromatic imports (Bryant and McClung, 2011). The scent
of aromatic rice is a highly heritable trait and reportedly is under the control of one to

20
four genes, depending on the population studied (Dhulappanavar, 1976; Sood and
Siddiq, 1979; Tripathi and Rao, 1979; Reddy and Sathyanarayanaiah, 1980; Pinson,
1994). More than 100 compounds that contribute to the aroma of rice have been
identified (Tsugita, 1985; Widjaja et al., 1996). Some of these volatile compounds
contribute to consumer acceptance of certain type of rice whereas other compounds
contribute to consumer rejection. The popcorn-like smell of aromatic rice grains due to
2-acetyl-1-pyrroline content is considered desirable by many consumers through out the
world (Buttery et al., 1986).
Water Uptake Ratio: Optimum water uptake ratio on cooking is an important
parameter for a variety, as yield of cooked rice is directly proportional to water uptake
ratio, which is an important consideration for the catering industry. Rice with high water
binding capacity normally yields soft textured cooked product. As degree of milling
influences cooking qualities like water uptake ratio, length expansion ratio, moreover,
related to the profit of the farmers and rice milling industry, it is imperative to choose a
suitable milling degree for the cultivars in order to minimize losses and improve
cooking/eating qualities.
Pasting Attributes: In terms of pasting behavior and functionality, starch is
generally regarded as the most important constituent of rice (Juliano, 1985). The starch
pasting characteristics determines the cooking quality which can be determined by using
Brabender Visco-Amylograph (BVA) or the Rapid Visco Analyzer which is a latest
technique. The validity of pasting behavior of starch by visco-amylograph has been
reported by many researchers (Halik and Kelly, 1959; Doublier, 1987; Deffenbaugh and
Walker, 1989; Aerts and Verspaille, 2001).

21
Different researchers have conducted experiments to study the genetic
divergence among rice cultivars based on physico-chemical, cooking and pasting
attributes and have been shown in Table 2.1.
Table 2.1 Characterization of rice cultivars based on grain quality attributes
S. No. Parameters Studied No. of Rice Cultivars and
their geographical place
Reference(s)
1 Physico-chemical 12 indica and 4 japonica Juliano et al. (1964)
2 Physico-chemical Dwarf indica, tall indica and
japonica
Swamy et al. (1978)
3 Physico-chemical and
Thermal
Indica, Japonica and
Japonica waxy
Singh et al. (2000)
4 Physico-chemical
Properties
20, Vietnamese and Japanese Tran et al. (2001)
5 Physico-chemical and
Cooking
6, Bangladesh Dipti et al. (2002)
6 Physico-chemical and
Cooking
8, Argentina Gonzalez et al. (2004)
7 Physico-chemical, textural
and Thermal
15, Jasmine rice cultivars Suwansri and
Meullenet (2004)
8 Physico-chemical 5 Indica, 6 Japonica and 3
waxy
Chen et al. (2004)
9 Physico-chemical,
Cooking and Textural
23, Indigenous Singh et al. (2005)
10 Physico-chemical 8, U.S long grain rice
cultivars
Cameron and Wang
(2005)
11 Physicochemical and
Cooking
5, Morogoro Shayo et al. (2006)
12 Physico-chemical and
Starch Structure
16 red rice accessions,
Southern United States
Patindol et al. (2006)
13 Physico-chemical and
cooking
56 rice cultivars from
different countries
Vanaja and Babu
(2006)
14 Physico-chemical,
Cooking and Pasting
2, Japonica waxy and
Japonica non-waxy
Ibanez et al. (2007)
15 Physico-chemical and
Cooking
4 non-basmati and 2 basmati,
Indigenous
Yadav et al. (2007)
16 Mineral and Pasting 4, Pakistani Shabbir et al. (2008)
17 Physico-chemical 23, Indigenous Bhat Upadya et al.
(2008)
18 Physico-chemical and
Pasting
4, non-glutinuous, Korean Kang et al. (2009)

22
19 Physico-chemical and
Cooking
32 aromatic, 5 exotic
aromatic and 3 non-aromatic
from different countries
Shahidullah et al.
(2009)
20 Physico-chemical,
Structure of starches
10, non-waxy Chinese Wang et al. (2010)
21 Physico-chemical and
Cooking
22, Indigenous Bhonsle and Krishnan
(2010)
22 Physico-chemical, Textural
and Nutritional
Mexican Murillo et al. (2011)
23 Physico-chemical and
Cooking
Jasmine brown rice, Thailand Daomukda et al.
(2011)
24 Physico-chemical,
Cooking and Pasting
9, Brazalian Garcia et al. (2011)
25 Physico-chemical and
Cooking
15 Indigenous and 5
Nigerian
Oko et al. (2012)
26 Physico-chemical and
Pasting
12, Korean Lee et al. (2012)
27 Physical, Cooking and
Nutritional
18, Indigenous Thongbam et al., 2012
28 Physico-chemical and
Cooking
Traditional Indigenous Asian
rice variety
Ravi et al. (2012)
29 Physico-chemical and
Cooking
24, Indigenous aromatic
cultivars
Srivastava et al.
(2013)
30 Physico-chemical and
Cooking
18, Japanese Nagano et al. (2013)
2.3.3 Characterization on Basis of Proteins
The polymorphism in proteins has wide range of applications in plant
classification, screening of mutants for seed storage proteins (Kumamaru et al., 1988),
germplasm resource analysis (Sun et al., 2000), variety identification (Huebner et al.,
1990), genetic diversity analysis (Thanh et al., 2000). Different types of methods like
alkali method, sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-
PAGE), capillary electrophoresis etc. have been used for extraction and separation of
proteins from rice cultivars.
A large number of rice cultivars with different ecotypes in japonica rice were
analyzed for endosperm storage proteins by SDS-PAGE to estimate their genetic

23
diversity and variety identification (Wei-dong, 2006). Similarly, analysis of rice seed
proteins was performed by SDS-PAGE in different rice varieties followed by cluster
analysis using SPSS to find out the diversity among them (Ansari et al., 2011).
The studies on protein polymorphism in rice varieties of Chhattisgarh (India) indicated
that the genetic variation among them related to grain storage protein polymorphism
(Meena and Shukla, 2013).
Capillary electrophoresis technique has been used to differentiate rice cultivars
on basis of protein polymorphism in less than 12 minutes. This method was observed to
be faster than other methods (Lookhart and Bean, 1995). The 2D-DIGE (2-Dimensional
Fluorescence Difference Gel Electrophoresis) method was applied to phenotype rice
varieties on basis of proteins. This method has been useful for analyzing natural
varieties of different cultivars and also for comparing the expression of allergen proteins
(Teshima et al., 2010).
2.3.4 Molecular Characterization of Rice
Morphology-based characterization has some limitations in the accurate
identification of the accessions, such as limited number of traits to characterize (Rao,
2004), highly heritable traits show no variation over much of the material studied (Karp
et al., 1997) and trait expression is subjected to strong environmental influence, mainly
in quantitative traits (Karp et al. 1997; Rao, 2004). Biochemical markers such as
isozymes, allozymes and storage proteins are effective in avoiding environmental
influence, but they are unable to detect low levels of variation (Rao et al., 2004)
because they screen a small section of the genome. DNA-based techniques overcome
both disadvantages, they identify polymorphism at DNA sequence level, therefore they

24
are independent of environmental influence, and furthermore they sample the whole
genome (Ovesna et al., 2002).
2.3.4.1 Extraction of DNA for Molecular Characterization
A unique advantage of molecular techniques is the rapid DNA analysis of many
plant samples using small quantities of DNA. Thus, a simple and rapid DNA extraction
method is needed for estimation of genetic divergence among genotypes by minimizing
the DNA extraction steps. Most of the reported methods for extraction of DNA, require
a large amount of plant tissue and it’s grinding in liquid nitrogen. But, with the
advancement in technologies, DNA extraction method using the dry half seeds of rice
and wheat has been developed (Chunwongse et al., 1993; Kang et al., 1998). A
simplified and modified Cetyl trimethyl ammonium bromide (CTAB) method has been
used for extraction of DNA from 6 days etiolated seedling (Wen-yue et al., 2006). A
protocol for extracting genomic DNA from young or old, fresh or dry rice leaves
that is applicable to a variety of plants, regardless of the complexity of their
genomes has been developed (Ahmadikhah, 2009). A modified CTAB based procedure
has been standardized for isolation of high quality and quantity of DNA from single rice
grain, seed and leaf tissue for deployment in hybrid seed and basmati grain purity
assessments (Rajenderakumar et al., 2011). A rapid and inexpensive DNA extraction
protocol has been described for different tissues of color rice and other plant species
which contain pigment and polyphenolic compound. This protocol is simple and fast
compared to other methods and no liquid nitrogen is required. The extracted DNA
applicable to marker assisted selection, DNA fingerprinting, quantitative traits loci
analysis, screening of transformants and enzymatic digestion (Ferdous et al., 2012).

25
2.3.4.2 Role of Molecular Markers in Estimation of Genetic
Relationships
Molecular markers have demonstrated a potential to detect genetic diversity and
to aid the management of plant genetic resources (Virk et al., 2000; Song et al., 2003).
In contrast to morphological traits, molecular markers can reveal differences among
genotypes at the DNA level, providing a more direct, reliable and efficient tool for
germplasm characterization, conservation and management (Botstein et al., 1980;
Tanksley et al., 1989). Molecular markers are randomly distributed throughout the
genome and the identified marker-trait associations are prone to recombination, possibly
leading to breakage of linkage between marker(s) and genes of interest. Plant genome
analysis in a number of plant species including rice have shown that genetic relationship
between different varieties using different markers can vary significantly (Parson et al.,
1997 and Virk et al., 2000).
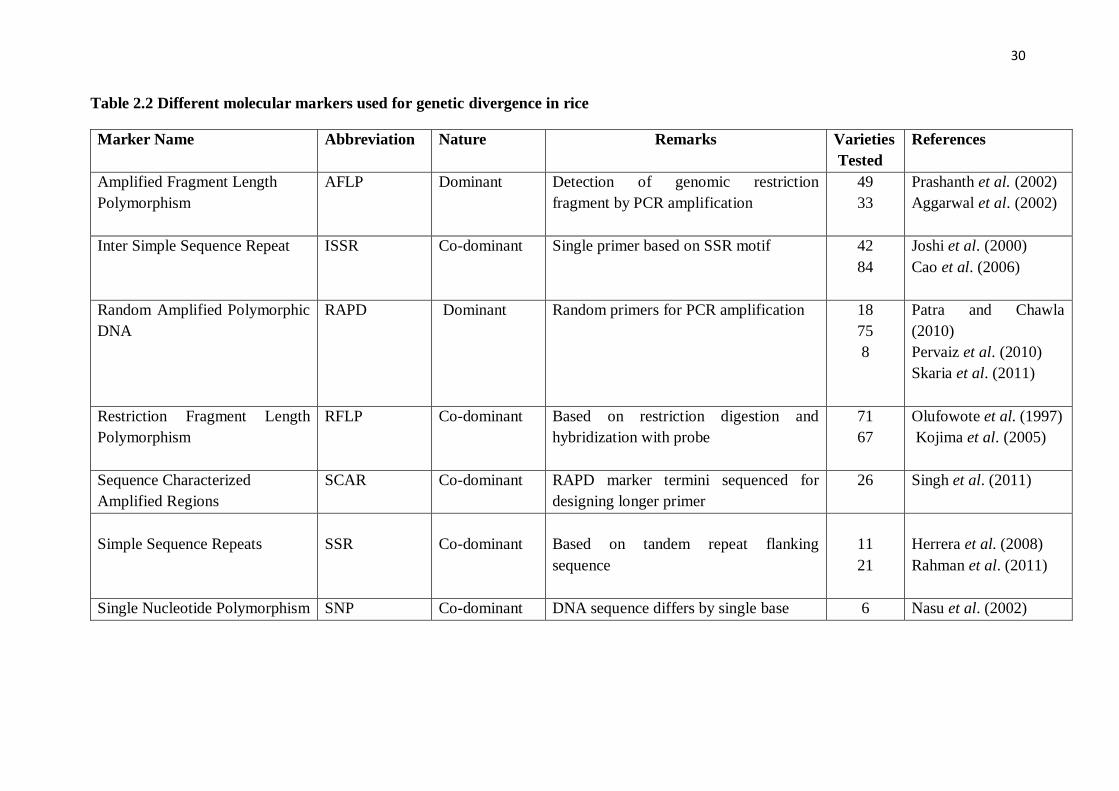
The most widely adopted markers used for genetic divergence studies among
various genotypes are Amplified fragment length polymorphism (AFLP), Random
amplified polymorphic DNA (RAPD), Simple sequence repeat (SSR) and Restriction
fragment length polymorphism (RFLP), having their own advantages and disadvantages
and needs careful evaluation before effectively developed for diversity analysis (Saini et
al., 2004). This applies to rice, where the infraspecific O. sativa groups can now be
identified using RAPD (Virk et al., 1994), as well as other molecular marker strategies
including RFLP (Wang and Tanksley, 1989), ISSR (Parsons et al., 1997), AFLP (Vos et
al.,1995; Zhu,1998) and SSR markers (Wu and Tanksley, 1993) as shown in Table 2.2.
Among different markers, SSR markers have been effectively used for characterization
of rice cultivars from various aspects and are the marker of choice for researchers. The
details and broad range of applications of SSR markers in rice species have been
discussed in subsequent paragraphs.

26
2.3.4.3 Simple Sequence Repeat Markers
SSR markers are actually non-coding regions which remained conserved during
the course of evolution and are ideal for DNA fingerprinting and varietal identification.
These are also known as the microsatellites (Litt and Lutty, 1989) and short tandem
repeat (STR) that contain tandem repeats (di, tri, tetra or penta), which vary in the
number of repeat units between genotypes and are referred to as variable number of
tandem repeats (VNTRs) or hypervariable regions (Mittal and Dubey, 2009). Any
variation in DNA sequence of priming sites and repeat number may help in diversity
analysis among genotypes. The SSR markers reveal polymorphism due to variations in
the length of microsatellite at particular loci among different genotypes. Since these are
abundant, hypervariable, multiallelic and evenly distributed throughout the nuclear
genome, they provide a valuable source of polymorphism, making them important class
of genetic markers (Varshney et al., 2005).
In addition, the highly polymorphic nature of many SSR markers is of particular
value when analyzing closely related varieties (McCouch et al., 2002). SSRs were first
studied in humans and have now been found in a wide array of other eukaryotes,
including mammals, birds, fish, insects, yeast and plant species (Mccouch, 1997). SSR
marker have wide range of merits like quickness, simplicity, rich polymorphism
and stability. The multifold nature of these markers make them widely applied in
genetic diversity analysis, gene mapping (Ma et al., 2011), construction of fingerprints
(Xiao et al., 2006; Ma et al., 2011), genetic purity test (Peng et al 2003), analysis of
germplasm diversity (Zhou et al 2003; Jin et al 2010) and identification of closely
related species . Furthermore, it has been stated that the data produced by SSR markers
analysis was helpful in providing a simple, fast and safe mean of genome assay in rice
(Upadhyay et al., 2011).

27
2.3.4.4 Mapping of Simple Sequence Repeats in Rice
The current level of genome coverage provided by SSR markers in rice is
sufficient for genotype identification, gene and quantitative trait locus (QTL) analysis,
screening of large insert libraries, and marker-assisted selection in breeding
(McCouch et al., 1997). SSRs were characterized and mapped in rice genome from
partial sequencing of cDNA and genomic DNA clones and it was observed that the
most frequently repeated SSR motif was d(CCG/CGG)n (Miyao et al., 1996).
Mapping of 121 SSR markers having genome-wide coverage of the 12
chromosomes, with an average distance of 1 SSLP (simple sequence repeat
polymorphism) per 16-20 cM has been done (Chen et al., 1997). These SSLP loci
included 86 poly (GA), 6 poly (TCT), 1 poly (ATT), and 1 poly (AATT) motifs.
Similarly, mapping of 312 microsatellite markers developed on the basis of partial
cDNA sequences available in GenBank and from Tsp509-digested small-insert genomic
library has been completed (Temnykh et al., 2000). A total of 2414 non-redundant SSR
primer pairs, representing 2240 unique marker loci, have been developed and
experimentally validated for Oryza sativa L (McCouch et al., 2002).
The sequence of rice chromosome 4 was screened for the chromosomal
positions, and composition of SSR, to produce a detailed map displaying all possible
SSR motifs and to use them for comparative genome analysis of the indica and japonica
subspecies (Li et al., 2004). Total of 1,844 SSR markers with SSR motifs of 20 bp and
repeated unit length of 1-6 base pairs were mapped and these SSRs were found to be
occurred once in every 18.8 kb, with one SSR per 23.8 kb and 16 kb on the short and
long arms, respectively while no SSR was detected in the core region of the centromere.

28
2.3.4.5 Application of Simple Sequence Repeat Markers
SSR markers have a large number of applications in rice cultivar identification,
genetic relationships, marker assisted selections, detection of genes and quantitative
trait loci analysis. These are ideal markers which would be able to reveal multiple
alleles, as they are evenly distributed in rice genome and are easy to score.
Assessment of Genetic Variability in Rice Cultivars: Thousands of new
accessions of rice are introducing into germplasm every year, thereby necessitating
assessment of their molecular diversity before elimination of the redundant genotypes.
The knowledge regarding the amount of genetic variation in germplasm accessions and
genetic relationships between genotypes are important considerations for designing
effective breeding programs, an essential prerequisite for the preservation of endangered
species. SSR markers are efficient in detecting genetic polymorphisms among rice
genotypes. Sequence variations in SSRs were observed between two cultivars which
provided a great opportunity for SSR-based marker development and comparative
genome analysis of the subspecies (Wu and Tanksley, 1993; Yang et al., 1994; Panaud
et al., 1996; Olufowote et al., 1997).
The use of microsatellite polymorphism for the identification of Australian
breeding lines of rice was investigated and showed that most of the cultivars were
uniquely identified by at least one microsatellite marker (Garland et al., 1999). ISSR-
PCR and SSR markers unambiguously distinguish the traditional basmati rice varieties
from the evolved and non-basmati rice varieties which can be exploited in future
breeding program and authentification (Nagaraju et al., 2002). SSR marker has been
utilized in detecting hybridization between rice and red rice in Arkansas (Rajguru et al.
2002). The rice cultivars of particular interest to U.S. breeding programs and two wild

29
species accessions were evaluated by means of 111 microsatellite markers distributed
over the whole rice genome and a total of 753 alleles were detected (Ni et al., 2002).
Different studies have shown that SSR markers are useful in identification of
rice hybrids and their respective parents, assessment of plant to plant variation within
parental lines and testing the genetic purity of rice hybrids (Yashitola et al., 2002;
Nandakumar et al., 2004; Sundaram et al., 2007). It has been suggested that a relatively
small number of microsatellite markers could be used for the estimation of genetic
diversity and the identification of rice cultivars (Ni et al., 2002). Two land races from
four commercial high yielding varieties were differentiated by using sequence tagged
microsatellite sites (STMS) markers which revealed a high degree of genetic diversity
(Kohli et al., 2004).
DNA fingerprint database of 24 rice genotypes including 3 traditional basmati, 9
cross-bred basmati, 8 indica and 3 japonica rice varieties, was developed on basis of
allelic diversity using 50 microsatellite markers (Siwach et al., 2004). Genetic diversity
and pattern of relationships were studied among the 18 rice genotypes representative of
the traditional basmati, cross-bred basmati and non-basmati (indica and japonica) rice
varieties using AFLP, ISSR and SSR markers and concluded that all the three marker
systems generated higher levels of polymorphism and could distinguish between all the
rice cultivars (Saini et al., 2004). Genetic diversity of seven Egyptian rice genotypes
using eight RAPD, six SSR and eight AFLP primer combinations has been observed
and concluded that some varieties originated from closely related ancestors possessing
high degree of genetic similarity (Saker et al., 2005). Analysis of the genetic variation
and DNA fingerprinting of 15 elite rice genotypes using 30 SSR primers on
chromosome numbers 7-12 has been done (Chakravarthi and Naravaneni, 2006).
30
Table 2.2 Different molecular markers used for genetic divergence in rice
Marker Name Abbreviation Nature Remarks Varieties
Tested
References
Amplified Fragment Length
Polymorphism
AFLP Dominant Detection of genomic restriction
fragment by PCR amplification
49
33
Prashanth et al. (2002)
Aggarwal et al. (2002)
Inter Simple Sequence Repeat ISSR Co-dominant Single primer based on SSR motif 42
84
Joshi et al. (2000)
Cao et al. (2006)
Random Amplified Polymorphic
DNA
RAPD Dominant Random primers for PCR amplification 18
75
8
Patra and Chawla
(2010)
Pervaiz et al. (2010)
Skaria et al. (2011)
Restriction Fragment Length
Polymorphism
RFLP Co-dominant Based on restriction digestion and
hybridization with probe
71
67
Olufowote et al. (1997)
Kojima et al. (2005)
Sequence Characterized
Amplified Regions
SCAR Co-dominant RAPD marker termini sequenced for
designing longer primer
26 Singh et al. (2011)
Simple Sequence Repeats
SSR
Co-dominant
Based on tandem repeat flanking
sequence
11
21
Herrera et al. (2008)
Rahman et al. (2011)
Single Nucleotide Polymorphism SNP Co-dominant DNA sequence differs by single base 6 Nasu et al. (2002)

31
Microsatellites or SSR marker are also useful in identifying indigenous non-
basmati aromatic rice genotypes which can help in genetic conservation management
and support intellectual property protection (Joshi and Behera, 2006). Different
NERICA varieties have been grouped with 102 microsatellite polymorphic markers and
this revealed the presence of a wide range of genetic differences among all the
NERICAs (Semagn et al., 2006). SSR markers are found to be fundamental in
determining the relationship between Brazilian rice land races (Brondani et al., 2006).
DNA fingerprints of the Bangladeshi cultivars of rice using SSR marker has been
determined and observed large differences in the polymorphism between indica and
japonica rice sub-species (Jalaluddin et al., 2007). Molecular analysis of colored Korean
rice line using SSR primer pairs has been done and polymorphic bands (20) with an
average of three alleles per SSR markers has been obtained (Ji et al., 2007).
Genetic variance of Cuban traditional rice cultivars using microsatellite markers
and the microsatellite loci revealed a poor relation of improved varieties with majority
of traditional varieties (Alvarez et al., 2007). SSR markers grouped the Oryza sativa
accessions of Argentine rice cultivars into two major groups, indica and japonica and
the japonica group into the sub-groups, tropical and temperate (Giarrocco et al., 2007).
The assessment of genetic variance of 11 Venezuelan rice cultivars using 48 SSR
markers has been done (Herrera et al., 2008). Similarly 12 SSR markers were used to
characterized 417 landraces collected in 1986, 1987 and 2003, in the state of Goias
(Brazil) to study the impact of rural exodus or replacement of rural varieties by
commercial cultivars (de Oliveira Borba et al., 2009).
Genetic diversity was measured using 35 microsatellite markers in 75 genotypes
of rice grown in Pakistan (Perviaz et al., 2010). A total of 142 alleles were detected at
32 polymorphic SSR loci. A dendrogram based on total microsatellite polymorphism

32
grouped 75 genotypes into four major clusters at 0.40 similarity coefficient,
differentiating aromatic from non-aromatic ones. Similar study was carried out to
evaluate the genetic relationship among 41 traditional and improved cultivars of
Pakistani rice by means of 30 microsatellite markers distributed over the whole rice
genome. It has been observed that all the tested markers were polymorphic and a total of
104 alleles were detected (Rabbani et al., 2010).
The analysis of genetic diversity among 16 aromatic short grain local land races
collected from different parts of India and 30 basmati cultivars collected from the
traditional basmati growing areas has been done using 26 SSR markers (Sivaranjani et
al., 2010). Single markers (RM28102) or combination of two (RM577, RM30) markers
were found to be effective in discrimination between both the aromatic groups. Based
on the similarity co-efficient values, genotypes were classified into two major clusters
with 70% dissimilarity revealing presence of high diversity.
Identification and discrimination of 17 HYVs and 17 local rice cultivars
including two wild rice cultivars has been done and it was observed that all the analyzed
microsatellite markers were polymorphic in nature with an average number of 6.33
alleles per locus (Rahman et al., 2009). A total of three variety specific alleles (RM-
11/147, RM-151/289 and RM-153/178) were identified for BR-11, Badshabhog and
BR-19 cultivars. Genetic variability in Spanish rice cultivars was evaluated by
fluorescently labeled primer pairs (Wankhade et al., 2010). In the same regard, 54
microsatellite markers were used to estimate genetic diversity of 53 Sarawak rice
cultivars in which a total of 43 alleles were detected with an average of 3.58 alleles per
locus (Lee et al., 2011). A total of 24 SSR markers were used across 12 elite aromatic
rice genotypes for their characterization and discrimination (Sajib et al., 2012). 48 rice
genotypes were grouped into different clusters using 7 microsatellite markers tightly

33
linked to major QTLs controlling three major components of rice cooking and eating
quality, i.e. amylase content, gelatinization temperature and gel consistency (Tabkhkar
et al., 2012). Genetic diversity among 15 indica basmati advanced line and 5 basmati
improved varieties has been detected by using 28 SSR markers covering all the 2 rice
chromosomes (Ashfaq et al., 2012). The genetic diversity of 64 rice genotypes using 20
SSR primers on chromosome number 7-12 was investigated (Kumar et al., 2012). Rice
accessions were evaluated by means of 24 microsatellite markers distributed over the
whole rice genome to evaluate the genetic diversity within a diverse collection of rice
(Oryza sativa L.) accessions and to determine differences in the patterns of diversity
within the aromatic and non-aromatic rice varieties (Shah et al., 2013)
Detection of Adulteration of Basmati with Non-Basmati Rice
Cultivars: Basmati rice has both aroma and post elongation properties, mainly
preferred by the consumers and no other variety in the world have both these
characteristics. But since the yield of basmati rice, per acre of land, is less than half of
that of non- basmati rice and because of higher inputs, basmati rice has become
unaffordable for most people. Basmati traders have made basmati rice affordable for
people of various income brackets-with its varietal difference (Lopez, 2008).
Many methods have been developed to detect adulteration in rice, like smelling
of grains after boiling in water or treated with potassium hydroxide and
chromatographic analysis of aromatic compounds (Lorieux et al., 1996). It’s very
difficult on morphological or chemical basis to identify a true basmati variety, since
many evolved cross bred non basmati varieties also resembles the basmati on
morphological basis. So techniques based on molecular basis proved effective in

34
characterizing basmati and non- basmati varieties (Lopez, 2008). Two classes of DNA
markers, i.e. fluorescence-based ISSR markers and SSR markers were developed for the
purpose of carrying out the genetic analysis of basmati rice varieties (Nagaraju et al.,
2002). They have found such markers efficient, cost-effective and helpful in detecting a
higher degree of polymorphism in rice. Traditional basmati was differentiated from
cheap cross bred and long rice varieties by using 35 SSR markers (Pal et al., 2004). Out
of 123 alleles, 25 alleles were found to be present only in basmati varieties. Basmati,
indica and japonica rice genotypes were analyzed for genetic diversity and a total of
235 alleles were detected by 30 SSR loci having 62 (26.4%) of them were present only
in basmati (Jain et al., 2004).
Capillary electrophoresis based multiplex microsatellite marker assay was
reported fist time for detection as well as quantification of adulteration in basmati rice
samples by Archak et al. (2007). They used single-tube multiplex assay for eight
microsatellite loci to generate variety-specific allele profiles that can detect adulteration
from 1% upwards.
Marker Assisted Breeding: Traditionally, plant breeding is based on phenotypic
selection of superior individuals for crop improvement, yet many limitations
encountered during this process viz. effect of environment, genotype, unreliability,
expensive in terms of time and cost. But in recent times, the development of molecular
marker-assisted selection (MAS), involves selection of plants carrying genomic regions
that are involved in the expression of traits of interest through molecular markers (Babu
et al., 2004). With the development and availability of an array of molecular markers
and genetic maps in crop plants, MAS has become possible for traits both governed by
major genes as well as quantitative trait loci (QTLs). Many reports have been published

35
about the linkage between SSR and agronomically important QTLs and genes which are
discussed as below:
Mapping and detection of quantitative trait loci: Currently, many SSR markers are
available for detection of QTL in rice, linked to important quality and agronomic trait
which aid in breeding superior lines that can grow well in local climates and have good
yield under stress conditions.
Mapping of quantitative tait loci related to quality traits: The gelatinization
temperature (GT), gel consistency (GC) and amylose content (AC) are the three major
rice traits that are directly related to cooking and eating quality (Little et al. 1958). GT
is the temperature at which starch irreversibly loses its crystalline order during cooking
which is mainly responsible for cooking time and the capacity to absorb water during
the cooking process, whereas GC and AC are responsible for softness and for texture
and appearance in rice. Hence, regulation of these three traits in rice has been a major
concern of rice breeders. To facilitate the development of new varieties with high
cooking and eating qualities, it is necessary to understand the genetic and quantitative
inheritance of such traits. QTLs that accounted for 80% and 57% phenotypic variation
for AC and GC, respectively on chromosome three, four, six and seven were detected
(Lanceras et al., 2000). Similarly, 4 QTLs for AC, 3 for GT and 5 for GC were found
using backcross-inbred lines (Li et al., 2004). The QTL on chromosome six covered the
wx gene region and mainly contributed to the variance between japonica and indica
varieties. The effect of QTLs corresponding to wax locus and alk locus has been
observed and found that wx locus showing a major effect on AC and GC whereas alk
locus showed a major effect on GT (Fan et al., 2005). One QTL for texture on

36
chromosome 3 and four QTL for AC on chromosome 3, 7, 9 and 12 were mapped
(Wada et al., 2006). Using 110 polymorphic SSR markers and 209 recombinant inbred
lines QTLs related to AC, GC and GT were mapped on seven different chromosomes
(Amarawathi et al., 2008).
Mapping of quantitative trait loci related to agronomic traits: A total of 11 QTLs
located at chromosome 2, 3, 4, 6, 11, and 12 were detected in eight traits, i.e., heading
date, flag leaf length, plant height, panicle length, panicle weight, seed set, weight of
100 grains, and grain weight by using 93 SSR markers (Susanto et al., 2008). Similarly,
the parental screening of IR64 and Tarome molaei was done with 235 SSR markers; out
of which 114 markers gave clear polymorphic bands which were used to construct a
linkage map to search for QTLs associated with panicle length, number of grain per
panicle, and panicle grain sterility (Ahamadi et al., 2008).
Drought is one of the major limited factors that affect the growth of rice in many
countries. A total of 525 SSR markers were chosen to screen a F2 mapping population
derived from a cross between Taichung 189, a susceptible japonica line, and Milyang
23, a tolerant indica line to find out the chromosome region associated with drought
tolerance (Lin et al., 2007). Further, 121 SSR markers were performed to search
potential QTL regions and a total of four QTL associating to drought sensitivity index
were detected. Applying these SSR DNA markers that are closely linked with drought
tolerance, it will be possible to facilitate early selection of drought tolerant lines and
shorten breeding period. Similarly, 90 polymorphic makers were used for QTL mapping
for salinity tolerance at seedling stage of a F2 breeding population derived from the

37
cross between BRRI dhan40, a moderately tolerant female parent with IR61920-3B-22-
2-1, a highly tolerant male parent (Islam et al., 2011).
Bulked segregant analysis (BSA) serves as an alternative approach for rapid
identification of markers associated with drought resistance traits (Kanagaraj et al.,
2010). BSA was carried out to identify markers linked to drought resistance using 23
recombinant inbred (RI) lines of IR20/Nootripathu, two indica ecotypes with extreme
drought response. The parent plants were screened for polymorphism using 1206 rice
microsatellite primer pairs. Out of 134 SSR polymorphic primers between parents, three
primers showed polymorphism between bulks. It was found that three primers co-
segregated among the individual RI lines constituting the respective bulks. The genomic
regions flanked by SSR markers have been reported to be associated with several
drought resistance component traits, that will aid in marker assisted breeding for
drought resistance in rice.
Mapping of Genes: Genetic improvement of important quality and agronomic traits are
of major concern for rice breeders to attain high yields and better quality characteristics.
The development of molecular marker techniques has enabled researchers to map
important traits in the rice genome and can be used in marker-aided selection of these
traits. Simple sequence repeat (SSR) markers are particularly useful for gene mapping
and marker-based selection, since these markers are amenable to high-throughput
analysis and are informative in many types of genetic crosses.
Cytoplasmic male sterility (CMS) is a common plant reproductive feature that
has been extensively used as an important tool to exploit heterosis and to develop
hybrids in many crops. Rf genes are needed for restoring fertility to CMS lines.
Searching for and molecular tagging of restorer genes are of high importance where

38
phenotyping is very time consuming and requires the determination of spikelet sterility
in testcross progeny. A fertility restorer gene was mapped by using SSR and CAPS
markers in rice line IR36 in a F2 population developed from the cross Neda-A×IR36
(Alavi et al., 2009). The genetic linkage analysis indicated three SSR markers (RM1,
RM3233, RM3873) and one CAPS marker (RG140/EcoRI) on the short arm of
chromosome 1 were linked to Rf3. Seven SSR markers linked with eui locus at the
genetic distance from 1.0 to 7.1 cM were identified (Khera et al., 2009). Furthermore,
these markers were validated in F2 (IR58025A/IR91-1591-3) and backcross
(IR58025B/IR91-1591-2//IR58025B) population segregating for eui gene. The SSR
markers reported in this study might be useful to the rice breeders interested in
transferring eui gene into their promising parental lines of hybrid rice.
2.4 Rice Adulteration
Basmati rice has both aroma and post elongation properties, mainly preferred by
the consumers and no other variety in the world have both these characteristics.
Difficulty in differentiating genuine basmati from other types of rice and the significant
price difference between them has led fraudulent traders to adulterate basmati rice with
non basmati rice varieties. Moreover, the unsuitability of traditional basmati for
intensive cultivation compelled the farmers and researchers to develop long grain
aromatic rice varieties, leading to the presence of both varieties in the market in addition
to a relatively inferior non-aromatic long-grain rice varieties (Nagaraju et al., 2002).
Since it is difficult to differentiate between traditional and evolved or long
grained aromatic grains based on visual test or physicochemical tests, fraudulent traders

39
make a sizeable profit by adulterating basmati rice grains and exploit consumers. In
Britain, the Food Standards Agency found in 2005 that about half of all basmati rice
sold was adulterated with other strains of long-grain rice, prompting rice importers to
sign up to a code of practice. Considering that some inadvertent admixture of grains
during harvest or post-harvest processing is not unusual, rice importing countries allow
certain limit (5-15%) of mixture. If any lot of basmati rice exceeds the permitted limit of
admixture, it is considered intentionally adulterated. Many basmati importing countries
particularly European nations have made it mandatory today for all consignments to
have an authentication certificate based on DNA test for importing basmati. Agricultural
Processed Food Products Export Developmental Authority (APEDA) and the Centre for
DNA Fingerprinting and Diagnostics (CDFD), Hyderabad, India, have established
jointly ‘‘APEDA-CDFD Centre for basmati DNA Analysis’’ to carryout DNA testing
and certification of the export consignments.
2.4.1 Methods for Detection of Adulteration
Basmati varieties have been sold at 2-3 time higher price, as compared to other
rice varieties. The high price of basmati led to its adulterations with other long grain
non-basmati varieties (Bligh, 2000). Different types of detection methods, based on
morphological, cooking and organoleptic attributes of basmati rice grains, have been
developed to authenticate and to check for adulterants.
Basmati grains are generally distinguished by their cooking quality traits.
Among various cooking quality parameters, differential l/b ratio has been reported to be
a reliable indicator of adulteration of basmati rice with Improved Sabarmati, Pusa-169,
PR-106, Kali-much, Lakra and Parimal, which are frequently used for adulteration. A

40
minimum value of l/b ratio of 3.92 ± 0.09 to 4.09 ± 0.09 was found to be an indicative
of pure Basmati-370 (Vaingankar and Kulkarni, 1989). The other cooking quality
parameters like apparent water uptake and loss of solids in the cooking gruel did not
seem to hold any promise as indicators of adulteration as no consistent behaviour has
been observed for these parameters. Certain other methods have been developed to
detect adulteration in rice like smelling of grains after boiling in water (Sood and Sidiq,
1978) or treated with potassium hydroxide and chromatographic analysis of aromatic
compounds (Lorieux et al., 1996). The study has been tried to investigate more rapid
and accurate method to detect Jasmine rice’s adulteration. Jasmine rice, KDML 105
variety, was adulterated with CNT 1 and PTT 1 at level 5, 8, 10, 15, 20, 25 and 30 %.
Classification of these adulterations of Jasmine rice using its textural and pasting
properties was achieved by applying Principal Component Analysis (PCA). PCA could
classify adulteration of Jasmine rice efficiently. The first two principal components
explained 81.6% and 90.5% of the total variance explained for CN1 and PTT1
adulteration, respectively (Pitiphunpong et al., 2011).
It is very difficult on morphological or chemical basis to identify a true basmati
variety, since many evolved cross bred long grained non-basmati varieties also
resembles the basmati on morphological basis. Work in this area using sensitive
analytical techniques is seriously warranted. Polymerase chain reaction (PCR) is very
useful for analytical purposes, and could detect adulteration in basmati rice. SSR
markers based polymorphisms have been studied for mapping the rice genome (Chen et
al., 1997) and for identification of rice accessions (Yang et al., 1994), which could be
helpful in detecting such fraudulent practices. The fluorescent labeled simple sequence
length polymorphism (SSLP) markers have been used to find the adulteration among
basmati and non-basmati rice cultivars. The centre for DNA fingerprinting and

41
Diagnostics (CDFD), Hyderabad (India) developed a capillary electrophoresis (CE)
based micro-satellite DNA profiling protocol, which facilitates quick and accurate
detection and quantification of adulteration in basmati rice consignments. The single-
tube assay multiplexes to generate variety specific alleles, which can detect adulteration
in basmati rice (Archak et al., 2007). A setup consists of a DNA based method coupled
with High Resolution Melting (HRM) analysis for rice products traceability using five
different microsatellite markers to genotyping basmati and non-basmati varieties has
been developed (Ioannis et al., 2012). In this set up, 8 bp deletion in badh2 gene in
combination with HRM to both DNA-typing of the basmati and non-basmati varieties to
quantify adulteration of Basmati rice products with non-basmati rice.
A fingerprint database of 27 rice varieties including commercially important
traditional basmati (TB), cross-bred (evolved) basmati and some non-basmati indica
rice varieties have been prepared using 50 microsatellite markers. A total of 271
electromorphs (alleles) were detected at 50 SSR (simple sequence repeat) loci, 92 of
which were observed only in 2–10 of the 16 basmati rice varieties. A large number of
unique (22) and null (16) electromorphs were noticed in basmati rice varieties. SSR
database reported in this paper broaden the list of SSR markers already recommended
for varietal identification and detection of adulteration in basmati rice supplies (Kaushik
et al., 2011).
2.5 Importance of Database Designing
The organized collection and management of any kind of data is called database.
Designing of database involves the process of producing a detailed data model, which
contains all the needed logical and physical design choices and physical storage
parameters needed to generate a design in a Data Definition Language, which can then

42
be used to create a database. The compilation and management of research data into
database is required for data integrity and consistency, to secure and maintain primary
research data, allow easy access and thereby increasing the overall utility of research
work. Different computer based programs are available, which help to organize and
analyze data. Visual Basic (VB) 6 is a high level programming language, which evolved
from the earlier DOS version called BASIC (Kiong, 2006). This program is a visual
and events driven programming language. VB Program is made up of many
subprograms, each has its own program code, and each can be executed independently
and at the same time each can be linked together in one way or another.
An online database system for Indonesian rice cultivars has been developed by
using VB.Net (Pertiwi et al., 2008). The database provides information of improved rice
varieties and their characteristics, application program for identifying the recommended
rice variety for certain location and preference, and application program for estimating
harvest time and yield based on transplanting time input. Similarly, a set of simple
Visual Basic 6.0 scripts was developed to construct the rice DNA polymorphism
database (Shen et al., 2004).
TNAURICE, a database released from Tamil Nadu Agricultural University
(TNAU), Coimbatore (India) provides information on both quantitative and qualitative
descriptors of the rice varieties inclusive of their parental details. The database is
available to the plant breeders involved in the varietal improvement programs to decide
on the choice of parental lines. TNAURICE can be accessed at
http://www.btistnau.org/germdefault.aspx for public domain (Ramalingam et al., 2010).
Similarly, a database for Chinese rice varieties and their genealogy has been developed
by Chinese National Rice Data Center (http://www.ricedata.cn/). Retrieval for rice

43
variety and pedigree search were developed as two core components of the database
(Zhi-guo and Lei, 2011).
The documentation of the physico-chemical and molecular attributes of basmati
and non-basmati rice cultivars facilitated by an easy and efficient search and retrieval
system is an absolute necessity considering the wide range of consumer taste and rice
based food products. Moreover, the management of research data into database is also
helpful to estimate the genetic divergence and to control the adulteration practices
among rice cultivars.





























