Embed Size (px)
Citation preview

1
Chapter 3 The Central Auditory System
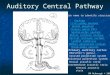
The central auditory pathway is composed of a number of nuclei and complex pathways that ascend within the brainstem (Fig. 1). The pathways are even for experts, terribly complex and the details of the various connections are, for our purposes, not important. The pathway into the brain begins with auditory nerve fibers that project from the cochlea into the brain where they first make synaptic connections with other neurons in the cochlear nucleus (Fig. 1). The cochlear nucleus is the first synaptic station of the auditory system in the brain. The neurons from the cochlear nuclei on both sides of the brain send their axons deep into the brain stem and make synaptic connections in a region of the medulla called the superior olivary complex. Since the superior olivary neurons receive information from both cochlear nuclei, they can be excited (or inhibited) by sounds delivered to either ear. Neurons receiving information from both ears are called binaural neurons. The comparison of information from the two ears is important for determining the location of a sound in space, as we will discuss in the following chapters. Thus, the first place that information from the two ears converges upon common neurons in the auditory pathway is in the superior olivary complex in the medulla. This is considerably from the visual system, where information from the two eyes is first combined in the visual cortex. The axons from the superior olive are then sent to the inferior colliculus in the auditory midbrain. The outputs of the inferior colliculus are sent to the medial geniculate body, the auditory thalamus and counterpart of the lateral geniculate body in the visual system. The projections of the medial geniculate are then sent to the auditory cortex in the temporal lobe.
Fig. 1. Outline of the connections of the central auditory system.

2
Figure 2. More detailed view of central auditory pathway, illustrating its tonotopic organization. Notice that the auditory nerve fibers from the cochlea diverge as they enter the brain, where each fiber innervates two major divisions of the cochlear nucleus, the dorsal and ventral portions of the cochlear nucleus. Each cochlear nucleus division then sends fibers into the brainstem to innerve the superior olivary complex (innervated by the ventral cochlear nucleus) and the inferior colliculus (innervated by direct projections from the dorsal cochlear nucleus. All ascending projections, whether from the dorsal cochlear nucleus, the superior olivary nuclei or the nuclei of the lateral lemniscus, converge upon a common site in the inferior colliculus before being relayed to the medial geniculate body (the auditory thalamus) and from the medial geniculate to the auditory cortex. Notice that each of the lower auditory nuclei are tonotopically organized, and thus remap the frequency organization originally established in the cochlea. The Frequency to Place Transform of the Cochlea is Remapped upon each auditory Nucleus One of the characteristic features of the somatosensory system is somatotopy, and in the visual system it is retinotopy, whereby the sensory surface is remapped upon each successive nucleus and on the somatosensory and visual corticies. This topographic remapping of the sensory surface upon each succeeding higher neural center is also a dominant feature of the auditory system and is called tonotopy. Figure 2 shows that the cochlea has an orderly representation of frequency, tonotopy, and illustrates how tonotopy is created in the cochlear nucleus, the first nucleus in the central auditory system, and how it is preserved in higher regions of the auditory system. Each of the auditory nerve fibers retains its position relative to other auditory nerve fibers as it synapses in the cochlear nucleus. As consequence of these

3
neighborhood relationships, neurons that innervate the basal cochlea, and have high best frequencies, come to lie dorsally in the cochlear nucleus while neurons that innervate the apical cochlea, and have low best frequencies, lie ventrally in the cochlear nucleus. If a microelectrode penetrated the cochlear nucleus from one side to the next, the first neurons sampled would have high best frequencies, and the best frequencies would progressively change to lower best frequencies as the electrode was advanced throughout the nucleus. This basic tonotopy is present in every auditory region, from cochlear nucleus to auditory cortex, and represents the neural basis for our ability to discriminate different pitches of sound.
Figure 3. Simplified drawing to illustrate the anatomical basis of tonotopy.
Numerous Acoustic Attributes are Represented in Each Isofrequency Region The illustration of tonotopy shown in Fig. 3 is in principal correct but oversimplified. The oversimplification is that each auditory nerve fiber is shown as innervating only one cochlear nucleus neuron. What actually happens is that as each auditory nerve fiber enters the cochlear nucleus the fiber bifurcates, or breaks up into two main branches (Fig. 4). Each branch then sends collaterals that make synaptic contact with many different types of neurons, which form a sheet of neurons in the cochlear nucleus. The auditory nerve fiber itself innervates only one segment of the basilar membrane, and has a best frequency determined by the place on the basilar membrane that it innervates. Thus, all of the cochlear nucleus neurons with which the auditory nerve fiber makes synaptic contacts, all of the neurons in the sheet, have the same best

4
frequency as the auditory nerve fiber from which they receive their innervation. Such a sheet of neurons is referred to as being isofrequency, i.e. being of one frequency. Fig. 3.
Figure 4. More realistic illustration of the innervation of the cochlear nucleus by auditory nerve fibers. Each auditory nerve fiber that emanates from a point on the basilar membrane, and thus is tuned to one frequency, branches as the fiber enters the medulla to innervate the cochlear nucleus. The various branches of each fiber then innervate one cell type in the cochlear nucleus. The cell types are shown schematically in the left panel. Each cell type has a unique morphology and a unique set of both ligand gated and voltage gated channels (those features are not shown). In addition, the morphology of the synapse made by each branch of an auditory nerve fiber as it innervates a particular cell type in the cochlear nucleus is also different. For example, the synapse auditory nerve fibers make with bushy cells in the cochlear nucleus (the lowest cells shown) are exceptionally large and are called endbulbs of Held, whereas the synapses made on multipolar cells (next to lowest cells) are much smaller bouton types of synaptic terminals. All of the cochlear nucleus cells that receive innervation from the same auditory nerve fiber are themselves tuned to the same frequency as the fiber that innervated them, and as a group are referred to as being isofrequency. See text for further explanation. In this way, the representation of a particular place along the basilar membrane in the cochlear nucleus (and other, higher regions) is composed not of a single neuron but rather is composed of a sheet of neurons, all having the same best frequency. This sheet of isofrequency cells is the unit module in the auditory system, because it represents the total processing that occurs for that frequency in each nucleus. The idea is that this unit module of isofrequency cells is then reproduced for each frequency (place along the basilar membrane) in virtually identical form across the cochlear nucleus. The only difference among modules (isofrequency sheets) is that each sheet represents a different portion of the sensory surface (Fig. 4). Thus, if we look at the entire cochlear nucleus, we see this basic pattern of unit modules repeated over and over again, each repeating segment deriving its innervation from a different portion of the basilar

5
membrane. The total population of sheets will re-represent the frequency-place map of the basilar membrane. Why is each place along the basilar membrane represented in the cochlear nucleus by a sheet of neurons, where the neurons within a sheet have a variety of morphological characteristics? The answer to this question is that each type of neuron differentially processes the common information brought in by the auditory nerve fiber. All the information contained in a sound is encoded by the vigor and timing of the spike trains in the auditory nerve fibers. Since almost all sounds are composed of a broad range of frequencies, the information of any particular frequency in the sound is encoded by the auditory nerve fibers that innervate the point on the basilar membrane tuned to that frequency. It follows then that each type of neuron in an isofrequency sheet extracts some aspect of the information contained in the spike train of those auditory nerve fibers, and the information extracted by that particular neuronal type is different from the information extracted by the other neuronal types. That each neuronal type processes information differently also means that, although each neuronal type receives the same input (the same spike train from the same auditory nerve fiber), the output from each neuron type, that is the spike train that each type sends to a higher nucleus, is different, and the output of each neuronal type represents a different feature of the sound whose total information was encoded in the spike trains of auditory nerve fibers. The differential processing of information is possible because each neuronal type in an isofrequency sheet is both morphologically distinct and has a particular synaptic arrangement on its cell body or dendritic tree that is different from the synaptic configuration of the other neuronal types. The particular morphology and synaptic arrangement on each neuronal type allows each type to process auditory information in a manner different from the other neuronal types. These features are illustrated in Fig. 5. Here a single auditory nerve fiber projecting upon an isofrequency sheet of neurons in the cochlear nucleus is shown together with the discharge patterns evoked by a tone burst at the auditory nerve fiber’s best frequency. If you look at the spike trains evoked in the auditory nerve fiber, you can see that most tone bursts elicit an initial high burst of discharges whose rate declines as the tone pip continues. This firing pattern is characteristic of all auditory nerve fibers, and all auditory nerve fibers respond to best frequency tone bursts in the same way. Now compare the discharge pattern of the auditory nerve fiber with the discharge patterns evoked in the cochlear nucleus neurons. Notice that the discharge patterns of the cochlear nucleus neurons are different from the discharge pattern of the auditory nerve fiber and are different from each other.

6
Figure 5. Each auditory nerve fiber transmits the same information (spike train) to each of the neurons that it innervates in an isofrequency sheet of the cochlear nucleus. However, each cell type in the isofrequency sheet transforms the incoming information differently, such that the output (the temporal discharges from each cell) is different. Each cell type then sends its axonal projection to a particular set of auditory nuclei in the ascending auditory system. In this way, each cell type in the cochlear nucleus extracts a different aspect of the common information contained in the auditory nerve discharge train, the information extracted by each cell type is then processed further by different higher order nuclei. The only exception is the discharge pattern of the cochlear nucleus fiber on the bottom of the figure (the output of the bushy cell), which is the same as the auditory nerve fiber (as we shall see in a later section, it is not exactly the same). In other words, considerable information processing has occurred, which serves to modify the incoming information and transform it into something different from the original representation. The acoustic information coming into the central auditory system is changed markedly by the different neurons in the cochlear nucleus. Parallel Processing is the Hallmark of the Central Auditory System The general mode of processing that occurs in the cochlear nucleus is called parallel processing, since each cell type in an isofrequency processes information differently and at the same time. Parallel processing is central to the auditory system, and continues through the axonal projections from each of the cell types in the cochlear nucleus. The projections from each cell group form a series of tonotopically organized parallel pathways, where each of the projections innervate different higher order nuclei. Each of those higher order nuclei will also processes information and extract something different from the incoming spike trains such that the spike trains that are output by the cells in higher order nucleus 1 are different from the output

7
of cells in nucleus 2, which are different from the outputs of nucleus 3 and so on. Each successive nucleus is extracting something different from the incoming spike trains, and thus encoding some feature of the sound that evoked the spike train that is different from whatever feature is extracted by the neurons in nucleus 2, which in turn is different yet from the information extracted by nucleus 3 and so on.
Figure 6. Parallel and serial pathways as they emerge from the cochlear nucleus. Notice that all fusiform cells (topmost cells) project to nucleus 1. Thus nucleus 1 will receive the full complement of tuned cells from the cochlear nucleus to the inferior colliculus. The axonal projections from the bushy cells (lowest cells) project to nucleus 5 (the superior olivary complex) and that nucleus will also be tontopically organized. Nucleus 5 (the superior olivary complex) then sends its axonal projections to the inferior colliculus in a tonotopic manner. Thus there is parallel processing (several projections that travel in parallel from the cochlear nucleus to the inferior colliculus) and there is also serial processing (projections from cochlear nucleus to superior olive and from superior olive to inferior colliculus). An example illustrating the outputs from the fusiform cells in the cochlear nucleus (cell number 5 in Fig. 5, and topmost cells in Fig. 6) and the outputs from the bushy cells (cell number 1 in Fig. 5 and lowest cells in Fig. 6) should make the idea of parallel processing clearer. The projections from the fusiform, shown as projecting to nucleus 1, terminate in the inferior colliculus. The axonal projections from fusiform neurons comprise a direct pathway from the cochlear nucleus to the inferior colliculus. The axonal projections from the bushy cells, shown as projecting to nucleus 5, do not go directly to the inferior colliculus, but rather project to the superior olivary complex, the first synaptic region in the auditory system that receives innervation from the two ears. Indeed, the axonal projections from bushy cells go not only to the superior olive on the same side, but also to the superior olive on the other side, thus creating binaural neurons in the superior olive. The projections from the superior olive are then sent to the inferior colliculus. The outputs of these two cell types in the cochlear nucleus travel in parallel, where one set of axons is sent directly to the inferior colliculus while the other is sent first to the superior olive to merge information from the two ears and the superior olive then sends its binaural information to the inferior colliculus.

8
The pathway from the cochlear nucleus to superior olive to inferior colliculus illustrates two other organizational principles of the auditory system. The first is that the processing in any one of the parallel pathways also incorporates serial processing. In other words, the connections of the pathway from the bushy cells in the cochlear nucleus to superior olive to inferior colliculus is serial, in that there is initial processing in bushy cells of the cochlear nucleus, which is followed by the binaural processing in the superior olive, which then is followed by additional processing in the inferior colliculus. The second principle is that all of the processing that occurred in each of the parallel pathway s that emerged from the cochlear nucleus then converges in the inferior colliculus. The inferior colliculus is a nexus in the ascending auditory system, since it receives the projections from almost all lower auditory nuclei, and thus the nucleus where a massive amount of integration and processing occurs. In later chapters we shall see how acoustic information from lower nuclei is integrated and transformed in the inferior colliculus with specific examples of how binaural information is transformed. Additional Features of the Ascending Auditory Pathway The general flow of information along the ascending auditory system was shown in Figs.1 and 2 . Two additional features should be mentioned, one that is inherent in the two parallel projection described above and another that is not apparent, but are illustrated in Fig. 7.
Figure 7. Wiring diagram of the auditory system showing both monaural and binaural pathways and that some pathways use glutamate (black dashed lines) as a neurotransmitter, and thus are excitatory to their targets, whereas other pathways use either GABA (red dashed lines or glycine (orange dashed lines) as neurotransmitters, and provide inhibition to their targets.

9
The first is that some of the parallel pathways are monaural in that they keep the information from the two ears segregated and convey information that is received only by one ear. An example is the first of the pathways described above, the pathway from the fusiform cells in Fig. 4 that project directly to the inferior colliculus. This pathway is also shown in Fig. 7 as the dashed black line emanating from the uppermost arrow in the cochlear nucleus. Since the projections originate only from the cochlear nucleus cells on one side, the information conveyed to the inferior colliculus is about sound received only by one ear, and thus is monaural information. Other parallel pathways are binaural, in that they convey information from both ears. The projections from the bushy cells in the cochlear nucleus to superior olive to inferior colliculus comprise a pathway that conveys binaural information.
The second feature, also illustrated in Fig. 7, is that while some of the ascending projections from the various nuclei are excitatory, many others are inhibitory, where some are GABAergic and others are glycinergic. For example, all of the projections from the cochlear nucleus utilize glutamate as a neurotransmitter, and the impact of the projections from the cochlear nucleus on their targets are always excitatory. In contrast, the projections from the superior olive are mixed, where some projections are excitatory (black dashed lines emanating from superior olive in Fig. 7) and others are inhibitory (orange dashed lines emanating from superior olive in Fig. 7). Thus the impact of the projections from the superior olive on the inferior colliculus is complex. Principal Organizational Features of the Ascending Auditory System The first is a well-defined tonotopic organization, a remapping of the cochlear surface upon the nuclei of the primary auditory pathway. The second is that the principal ascending pathways originate from the cochlear nucleus. Each pathway ascends in parallel with the others, and has a unique pattern of connectivity with the subdivisions of the superior olivary complex and other higher nuclei. The third feature is that the projections emanating from the cochlear nuclei create some pathways designed to provide convergence of signals from the two ears (binaural pathways) whereas others provide separation of the inputs from the two ears (monaural pathways). The fourth feature is that virtually all of the brainstem auditory pathways, whether binaural or monaural, converge at a common destination in the central nucleus of the inferior colliculus (ICc). In summary, we can identify 9 major organizational features of the ascending auditory system.
1) Each nucleus in the auditory pathway remaps the frequency representation of the cochlea and thus each auditory nucleus, from cochlear nucleus to auditory cortex is tonotopically organized.
2) Each incoming auditory nerve fiber innervates a number of different neuronal types in the cochlear nucleus, and thus is the first site where acoustic information is processed in a parallel fashion.
3) A series of parallel pathways emerges from the cochlear nucleus, where each pathway originates from a particular neuronal type in the cochlear nucleus.

10
4) The processing of information that occurs along each of the parallel pathways is serial, in that projections from one nucleus are then sent to a higher nucleus whose projections are then sent to yet a higher nucleus.
5) There are two major types of parallel pathways, monaural pathways that convey information from only one ear and binaural pathways that convey information resulting from the merger of the information from the two ears.
6) Some parallel pathways are excitatory to their targets whereas others are mixed, and have both excitatory and inhibitory influences on their targets.
7) All of the parallel pathways converge on a common target in the midbrain, the inferior colliculus.
8) Projections from the inferior colliculus are sent to the auditory thalamus, the medial geniculate body.
9) The projections from the medial geniculate are sent to the auditory cortex. Binaural Information Is Topographically Mapped in Isofrequency Regions at Higher Levels of the Auditory Pathway One of the most intensively studied aspects of hearing is sound localization. Sounds are localized by comparing the arrival time or intensity of sound received at one ear with the same features received by the other ear. Thus the ability to determine the location of a sound depends on binaural processing, a topic that we take up in all the subsequent chapters. Recall that information from the two ears initially converges in the superior olive, the synaptic station that follows the cochlear nuclei (Figs. 7 and 8). There are actually a large number of distinct auditory nuclei in the superior olive and for this reason it is more appropriate to refer to it as the superior olivary complex.
Figure 8. Location and innervation of the medial superior olive (MSO) and lateral superior olive (LSO). The LSO and MSO are the first nuclei in the central auditory system that are binaurally innervated (innervated by both ears). Notice that the MSO receives direct inputs from the cochlear nuclei (CN) on both sides. The inputs from both sides are excitatory and thus MSO neurons are excited by stimulation of either ear. Such neurons are often referred to as excitatory-excitatory (E-E) neurons. The lateral superior olive (LSO) is also binaurally innervated but through a slightly different circuit than the MSO. The inputs from the

11
cochlear nucleus on the ipsilateral (same) side are excitatory to LSO neurons. Notice that the inputs from the contralateral (opposite) side first make a synaptic connection in the medial nucleus of the trapezoid body (MNTB). The axonal projections from MNTB neurons then provide the innervation from the opposite ear to the LSO cells. MNTB neurons utilize glycine as a neruotransmitter and thus provide inhibitory innervation to the their targets in the LSO. Thus, LSO neurons are excited by stimulation of the ipsilateral ear and inhibited by stimulation of the contralateral ear. Such neurons are referred to as excitatory-inhibitory (E-I) neurons.
Although the superior olive complex contains many auditory nuclei, the two principal nuclei in the complex are the lateral superior olive (LSO) and the medial superior olive (MSO). They are located next to each other in the medulla, where one is located more laterally (the lateral superior olive) while the other is closer to the midline (the medial superior olive). As we a preview, the majority of neurons in the medial superior olive are tuned to low frequencies and are involved in the processing of interaural time disparities, the cues animals use to localize low frequency sounds. In contrast, most neurons in the lateral superior olive are tuned to high frequencies and are well suited to code interaural intensity disparities, the cues animals use to localize high frequency sounds. In later chapters we will discuss why interaural time and intensity disparities convey information about the location of low and high frequency sounds, and how the MSO and LSO process these sounds.
Fig. 9. Schematic diagram illustrating E-E and E-I neurons. In each panel, recordings are made from a neuron in either the left MSO (left panels) or left LSO (right panels). In each case, speakers are positioned in both ears. If sound is delivered only to the left ear (top left and right panels), both E-E neurons in the MSO and E-I neurons in LSO

12
discharge. However, if sound is delivered only to the right ear (middle panels), only cells in the MSO fire. LSO cells do not fire because they are receiving inhibitory innervation from the MNTB through the right ear. However, if sound is presented simultaneously from both speakers, MSO cells fire vigorously because of the summation of the excitatory inputs from both ears. LSO cells, in contrast, either fire much less vigorously or not at all to binaural stimulation because the excitatory innervation from the ipsilateral ear is canceled by the inhibition from the contralateral ear.
Neurons in the medial superior olive receive excitatory inputs from the two ears, and thus sound delivered from either ear evokes discharges. These are called excitatory-excitatory or E-E neurons. When best frequency tone pips are delivered simultaneously to both ears, the responses summate and a very large response is evoked. In contrast to neurons in the medial superior olive, neurons in the lateral superior olive receive excitatory inputs from one ear and inhibitory inputs from the other, and are called excitatory-inhibitory or E-I neurons. When a best frequency tone pip is delivered to the excitatory ear, brisk discharges are evoked from these neurons. However, when best frequency tone pips are delivered simultaneously to both ears, the discharges evoked by the excitatory ear are suppressed by the inhibition produced by the inhibitory ear, and the neuron can be totally silenced with binaural stimulation. The degree to which the neuron is inhibited depends upon the relative sound intensities at the two ears. Some E-I neurons are completely inhibited when the sounds at the two ears are equally intense, while others require a louder sound at the inhibitory ear than the excitatory ear to silence activity. E-E and E-I Neurons are also Common in Higher Regions of the Auditory System Both the lateral and medial superior olives send projections to the inferior colliculus, and the inferior colliculus in turn projects to the medial geniculate body, which then projects to the auditory cortex. Recent investigations have shown that E-E and E-I types are topographically ordered within each isofrequency region of the inferior colliculus, medial geniculate body and auditory cortex. One of the clearest illustrations of this binaural representation is seen in the primary auditory cortex of the cat (that cortical region is called AI). Notice first that the primary auditory cortex (AI) is tonotopically organized (Fig. 2, 10). In the auditory cortex this is somewhat analogous to the repeating ocular dominance columns in the visual cortex. All of the neurons in a dorsoventral column in the cortex have the same best frequency. This is equivalent to the similar receptive field locations that neurons in a column of the visual cortex have. Furthermore, there is a region of cortex where all columns have the same best frequency, an isofrequency region, which is the equivalent of a hypercolumn in the visual cortex where all neurons have similar receptive field positions. Each isofrequency region consists of a strip of cortex running from the top of the cortex towards the bottom. Shown in Figure 7 is a portion of the cortex showing the tonotopic organization and binaural representation. If you look at a particular isofrequency region or contour, it is evident that within the contour are repeating areas of E-E and E-I neurons. Furthermore, the E-E and E-I areas occur in each isofrequency region and thus run perpendicular to the isofrequency contours.

13
Figure 10. Orderly rrangement of E-E and E-I cells within isofrequency contours of the auditory cortex. Unlike the visual cortex, the functional significance of these binaural strips is not well understood. However, one function consequence, but certainly not the only one, may be related to our ability to associate a sound with the region in space from which it emanated. One of the important cues for sound localization is the relative sound intensity at the two ears. If you put on earphones and present two tone pips of equal intensity, they would appear to be centered in your head. If the pip to the left ear is made louder, the sound appears to move towards the right. Due to the shadowing effect of the head, sounds above 2000 cycles coming from the right side will be louder in the right ear than the left, and so on for different positions in space. Sounds located directly ahead will be equally loud in the two ears. Recall that the discharge vigor of E-I neurons depends upon the intensity of sound in the excitatory ear relative to the intensity in the ipsilateral ear. Some E-I neurons are only inhibited

14
when sounds are equally loud in the two ears. Others require louder sounds in the ipsilateral for inhibition while others required louder sounds in the contralateral ear for inhibition: there is a graded continuum of relative binaural intensity requirements among the population of E-I cells. It is for this reason that EI cells are thought to be important for the coding of sound location. A more detailed examination of exactly how EI cells process intensity differences at the two ears and how that acoustic cue is represented in the brain is the topic of the next chapter.









![Index [link.springer.com]978-1-4757-4219-0/1.pdfeffects on central auditory system, 103-104 ... 314 Index Auditory system development ... Central nervous system,](https://img.pdfslide.us/doc/110x75/5b3d10337f8b9a26728dc19c/index-link-978-1-4757-4219-01pdfeffects-on-central-auditory-system-103-104.jpg)