Embed Size (px)
Citation preview

ORIGINAL PAPER
Changes in morphology of Rhizopus chinensis in submergedfermentation and their effect on production of mycelium-boundlipase
Yun Teng Æ Yan Xu Æ Dong Wang
Received: 23 May 2008 / Accepted: 19 August 2008 / Published online: 9 September 2008
� Springer-Verlag 2008
Abstract In order to control suitable mycelium mor-
phology to obtain high lipase productivity by Rhizopus
chinensis in submerged fermentation, the effects of fungal
morphology on the lipase production by this strain both in
shake flask and fermentor were investigated. Different
inoculum level and shear stress were used to develop dis-
tinctive morphologies. Analyses and investigations both on
micromorphology and macromorphology were performed.
Study of micromorphology reveals that micromorphologies
for dispersed mycelia and aggregated mycelia are different
in cell shape, biosynthetic activity. Macromorphology and
broth rheology study in fermentor indicate that pellet for-
mation results in low broth viscosity. Under this condition,
the oil can disperse sufficiently in broth which is very
important for lipase production. These results indicate that
morphology changes affected the lipase production sig-
nificantly for R. chinensis and the aggregated mycelia were
suggested to achieve high lipase production.
Keywords Lipase � Morphology � Rheology �Rhizopus chinensis � Mycelia aggregation
Introduction
Fungal morphology is an important parameter for fila-
mentous fungus fermentations, is often considered as one
of the key parameters of physiological and engineering
studies of filamentous fermentations used in the design and
operation of such systems [1]. In submerged culture,
morphology of filamentous microorganisms usually varies
between ‘‘pellet’’, ‘‘clump’’ and ‘‘filamentous’’ forms,
depending on the respective culture conditions and geno-
type of the strain [2]. Often, morphology is correlated with
production of enzymes or secondary metabolites [3].
Filamentous microorganisms are good source for many
primary metabolites and secondary metabolites such as
organic acids and antibiotics. They are also good producer
for many enzymes or microbial proteins production. The
change of mycelial morphology was found to have
remarkably effect on these primary or secondary metabo-
lites [4–7] production, and also on enzymes production.
Therefore, many researchers have focused on this research
field, and some valuable results were obtained. Papagianni
[8] and her coworkers reported that growth of Aspergillus
niger in the form of large pellets was associated with lower
specific protease activities and increased specific gluco-
amylase activities compared with filamentous
morphologies. Jin [9] and his coworkers found that the
formation of small compact pellets under designed culti-
vation conditions favored higher fungal protein yields. El-
Enshasy [10] reported that culture conditions resulting in
the formation of smaller fungal pellets with an increased
mycelial density result in higher yields of extracellular
Y. Teng � Y. Xu � D. Wang
Laboratory of Brewing Microbiology and Applied Enzymology,
School of Biotechnology, Jiangnan University, 1800 Lihu Rd,
214122 Wuxi, Jiangsu, People’s Republic of China
Y. Teng � Y. Xu � D. Wang
Key Laboratory of Industrial Biotechnology,
Ministry of Education, Jiangnan University, 1800 Lihu Rd,
214122 Wuxi, Jiangsu, People’s Republic of China
e-mail: [email protected]
Y. Xu (&)
State Key Laboratory of Food Science and Technology,
Jiangnan University, 1800 Lihu Rd, 214122 Wuxi,
Jiangsu, People’s Republic of China
e-mail: [email protected]
123
Bioprocess Biosyst Eng (2009) 32:397–405
DOI 10.1007/s00449-008-0259-8

glucose oxidase for recombinant A. niger. Other enzymes
production were also affected by the change of mycelial
morphology, they were amylase [11], lipase [12, 13], pol-
ygalacturonase [14], neo-fructosyltransferase [15], and
phytase [16]. The relation between fungal morphology and
process productivities has attracted interest from both
academic and industry and attempts have been made to
manipulate morphology to achieve maximal performance.
Most of researchers focused on two aspects of mor-
phology changes: one is the change of microscopic
morphology which can influence the enzyme biosynthesis
and secretion capacity directly; another one is the change
of macroscopic morphology which can influence broth the
rheology and the mass transfer properties [17]. The
micromorphology deals with the morphology of the
individual hyphal elements, e.g., the diameter, the length
of the hyphal elements and the number of tips on an
individual hyphal. Peberdy suggested that protein secre-
tion in filamentous fungi is intimately associated with the
process of growth at the hyphal tip [18]. Spohr and his
coworkers [11] reported that the enzyme secretion
capacity for A. oryzae have a correlation with the number
of tips. The change of micromorphology for recombinant
strain A. oryzae was also proved to have remarkable
influence on the lipase production [12]. Compare with
micromorphology, the macromorphology could be easily
distinguished by eyes or simple observation methods,
these morphological growth forms can have a significant
effect on the rheology of the fermentation broth and thus
the bioreactor performance. For example, pellet mor-
phologies result in Newtonian broths and better mass
transfer rates compared to viscous and often pseudoplastic
filamentous broths [19]. The different of mass transfer
rate will lead to a different metabolic pathway and dif-
ferent enzyme production.
Rhizopus chinensis CCTCC M201021 is a typical fila-
mentous fungus, and the mycelium-bound lipase produced
by this strain was proved to be an efficient biocatalyst in
biosynthesis of short-chain fatty esters in n-heptane [20]. In
our previous studies, we found that the change of mor-
phology could effect the mycelium-bound lipase
production for the mutant strain R. chinensis Y92-M [21]
and the lipase with synthetic activity was membrane-bound
[22]. In order to find out the relationship between the
change of morphology and lipase production by R. chin-
ensis, both effects of micromorphology and
macromorphology on mycelium-bound lipase production
were investigated. The study of micromorphology was
performed using mycelia obtained from shake-flask. And
the rheology study under different macromorphology was
performed in stirred tank reactor. Different morphologies
were manipulated by different inoculum level and stirring
speed.
Material and methods
Microorganism and media
Rhizopus chinensis CCTCC (China Center for Type Cul-
ture Collection) M201021 was isolated from Da Qu (a kind
of traditional leaven for production of Chinese liquor) by
our laboratory. The composition of medium contained
olive oil (23.7 g/L), MgSO4.7H2O (0.5 g/L), K2HPO4 (3 g/
L), maltose (1 g/L), peptone (40.6 g/L), pH was adjusted to
5.5. The medium was inoculated with spores with different
spore concentrations after sterilization. The spores were
washed from a fresh potato dextrose agar slant (72 h) by
deionized water, and the spore concentration was deter-
mined hemacytometer count.
Culture conditions
Shake flask cultures
Erlenmeyer flasks of 250 mL, containing different volume
of medium, were sterilized at 121 �C for 20 min. After
inoculated by different inoculum levels, the flasks were
incubated on shaker incubator for different rotation speed
at 30 �C for 72 h. The cultivated mycelium was separated
from the culture by filtration, washed twice with tap water
and once with 25 mM phosphate buffer (pH 7.0). The
mycelia were then lyophilized for 24 h by a freeze drying
system (Labconco, USA). Biomass was determined from
100 mL fermentation broth.
Stirred tank reactor (STR) cultures
Fermentation was carried out in a stirred-tank fermentor
(7L, Bio Flo110, New Brunswick scientific, USA) at 30 �C
under different stiring speed and different inoculum levels,
the working volume was 4.5 L, and the aeration rate was
1 vvm.
Analytical methods
Synthetic activity determination
Synthetic activity was measured by the ester-synthesis
method in heptane according to the procedure described
previously [22]. Octanoic acid (1.2 M) and ethanol (1.2 M)
in heptane each of 0.5 mL were mixed. The reaction was
started by adding 20 mg dry cell and incubated for 30 min at
40 �C with a shaking speed of 200 rpm. The reaction mix-
ture was then filtered using 0.15 lm membrane to remove
the cells. Samples (400lL) were then drawn and mixed with
2-hexanol of 100lL, as internal-standard, then analyzed by
subsequent gas chromatograph (Agilent 6820, Agilent
398 Bioprocess Biosyst Eng (2009) 32:397–405
123

Technologies Company Ltd, Shanghai, China) which
equipped with a polyethylene glycol capillary column
(AC20, 30 m 9 0.22 mm 9 0.25 lm, SGE International
Pty. Ltd, Australia). Nitrogen was used as a carrier gas, and
the injector and detector temperatures were set at 250 �C.
Oven temperature by programmed temperature was started
at 90 �C for 1 min before being elevated to 200 �C for 5 min
at 10�C/min. One unit of lipase synthetic activity was
defined as the amount of enzyme which catalyzed to produce
1 lmol of ester per minute.
Glucose and maltose determined
Glucose and maltose concentration was determined by
HPLC (Waters 650E, Waters Corp., USA), which equipped
with Sugarpak1 (6.5 mm 9 300 mm) columen (Waters
Corp., USA) and refractive index detector (Waters 2410,
Waters Corp., USA). Pure water was used as mobile phase,
flow rate was 0.5 mL/min and columen temperature was
85 �C. Before injection, the sample was prepared by cen-
trification and filtration by 0.25 lm microporous
membrane, and diluted to a certain concentration.
Image analysis
Sample preparation was carried using method described by
Haack [12]. Two-milliliter samples were taken during the
cultivations for image analysis, and one or two drops lac-
topenol-blue were added to stop growth and increase the
contrast of the images. The samples were stored at 4 �C for
later analysis. Before analysis, the samples were diluted to
approximately 0.25 g dry weight per litre to get a reason-
ably amount of hyphal element on each slide. Image
capture was carried out via a CCD camera (DXM 1200C,
Nikon Corp, Japan) mounted on a microscope (Nikon
eclipse 50i, Nikon Corp., Japan). Morphological measure-
ments were carried out using the Image-Pro PLUS software
(Media Cybernetics Inc., MD, USA). Measurements of
pellet mean diameter were done on images obtained using
49 objective, and measurements of other hyphal parame-
ters were measured using 109 or 409 objective. The
equivalent diameter of approximately 50 pellets was
measured per sample. For filamentous growth (freely dis-
persed mycelium), the average length of the filaments was
determined by estimating branches in individual mycelial
trees [8]. During the batch phase, the hyphal diameter was
measured approximately 10 lm from the end of the hyphal
tip. The number of tips was measured in individual
mycelial trees. Microscopic morphology was also per-
formed by electron microscope (Quanta-200, FEI,
Netherlands).
Pellets were distinguished from clumps and individual
mycelial trees by greyness levels differences, an approach
which has been used to provide a definition of a pellet [8].
Fully entangled filaments could be easily distinguish by it
irregular shape and tissue like form from pellet and freely
dispersed mycelium.
Fluid dynamics measurements
The rheological measurements were performed in the
250 mL beaker contained 200 mL fermentation broth using
a Brookfield viscometer (Brookfield DV-E, Brookfield,
USA), fitted with a disc-spindle impeller (1#). The shear
stress (s) of the fermentation broth was characterized by
the simple power law model of Ostwald–de Waele power
law approach for non-Newtonian fluids as shown below,
where c is the shear rate.
s ¼ Kcn ð1Þ
The constants, K and n represent the consistency index and
the flow behavior index, respectively. The apparent
viscosity, ga, is given by:
ga ¼sc¼ Kcn�1 ð2Þ
Taking natural logarithms on both sides of Eq. 3:
InðgaÞ ¼ InK þ ðn� 1ÞInc ð3Þ
Because of the DV-E Brookfield viscometer could only
give out the relationship between apparent viscosity ga and
rotational speed Ni. So, a equation was used to describe the
shear rate c by Ni [23].
c ¼ KNcðnÞNi ð4Þ
where KNcðnÞ was a value which could be calculated from
the result reported by Mitschka [23]. After replaced c in
Eq. 3 by KNcðnÞ and Ni, the equation was changed to:
InðgaÞ ¼ InðK � KNcðnÞn�1Þ þ ðn� 1ÞInNi
The values of InðK � KNcðnÞn�1Þ and n were able evaluated
from a natural logarithmic plot of ga versus Ni. And K was
able get from the values of InðK � KNcðnÞn�1Þ:
Result and discussion
Effect of culture condition on the mycelial morphology
and lipase production in shake flask
In our previous studies [21], we found that culture con-
ditions, like inoculum level, rotation speed and
fermentation volume could influence the macromorphol-
ogy of R. chinensis during lipase production in shake flask
cultivation. The culture conditions and results of macro-
morphology and lipase production were listed in Table 1.
Bioprocess Biosyst Eng (2009) 32:397–405 399
123

The synthetic activity of mycelium-bound lipase changed
from 101.2 U/g to 691 U/g with the change of mycelial
morphology from dispersed mycelia to fully entangled
filaments. Inoculum is very important for the mycelial
morphology in this study, when 3.3 9 109 spores L-1
inoculum was used, dispersed mycelia (Fig. 1c) were
formed regardless the rotation speed or volume of broth
for 250 mL flask used. Fully entangled filaments (Fig. 1a)
were obtained in the application of less spore number
3.3 9 108 spores L-1 and 3.3 9 107 spores L-1. Among
the factors that determine the morphology and the general
course of fungal fermentations, the amount, type and age
of the inoculum are of prime importance [1]. Therefore,
many researchers found the inoculum have remark influ-
ence on mycelial morphology. When the inoculum level
was varied from 104 spores mL-1 to 109 spores mL-1, the
mycelial morphology change from pellet, clump to free
mycelial during citric acid production by A. niger. The
production of citric acid was affected by the change of
mycelial morphology [4]. The influence of inoculum on
pellet formation was also studied in detail by Nielsen [24]
and Tucker [25].
Rotation speed and fermentation volume also remark-
ably affected the morphology and lipase production in
shake flask. When the rotation speed was lower than
150 rpm,entangled filaments (Fig. 1b) and dispersed
mycelia (Fig. 1d) were obtained regardless the inoculum
concentration was used. Fully entangled filaments (Fig. 1a)
could be obtained while rotation speed was higher than
200 rpm. Dispersed mycelia also formed when 40 ml broth
was contained in 250 mL flask. Rotation speed and fer-
mentation volume affected the shear force in flask actually,
and the effect of shear force on mycelial morphology has
been studied extensively by many researchers [6, 26].
Therefore, Mycelial aggregation is possible caused by high
shear force in this study. This conclusion could be further
confirmed by using of baffled flask which can provide
higher shear force. Small pellet (Fig. 1d) was formed in
this flask, and lipase activity reached 368 U/g (Table 1).
Same result was reported by Du et al. [27], they found that
in the baffled flask the pellet was formed, and high anti-
biotic production was obtained in R. chinesis 12 culture.
From the above result, it was considered that mycelial
aggregation (pellet and entangled filaments) seems to be a
principal factor in the enhancement of whole-cell lipase
production by R. chinensis. The reason for the different
lipase formation which caused by the morphology changes
need to be studied more in detail.
Growth process and lipase production under two typical
mycelial morphology in shake flask
Two culture conditions were performed in shake flask to
maintain typical mycelial morphology: fully entangled fil-
aments and dispersed mycelia. For fully entangled filaments,
condition A was used: inoculum level 3 9 108 spores/L,
fermentation volume 21 mL/250 mL, rotation speed
200 rpm; For dispersed mycelia, condition B was used:
inoculum level 3 9 109 spores/L, fermentation volume
40 mL/250 mL, rotation speed 150 rpm.
Table 1 Effect of culture
condition on the mycelial
morphology and lipase
production
Conditions Macromorphology Synthetic
activity(U/g)
Inoculum level 106/30 mL Fully entangled filaments 650
107/30 mL Fully entangled filaments 691
108/30 mL Dispersed mycelia (pulp like) 109
Erlenmeyer flasks 150 rpm Entangled filaments 209
200 rpm Fully entangled filaments 691
Baffled flask 150 rpm Entangled filaments 159
200 rpm Fluffy pellets (small) 368
Broth content for 250 mL flask 20 mL Entangled filaments 293
40 mL Dispersed mycelia (pulp like) 101.2
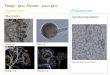
Fig. 1 Macromorphology
under different culture
conditions for Rhizopuschinensis. a Fully entangled
filaments, b Entangled
filaments, c Dispersed mycelia
(pulp like), d Pellets
400 Bioprocess Biosyst Eng (2009) 32:397–405
123

The time courses for biomass, pH, synthetic activities
for mycelium-bound lipase in shake flask are shown in
Fig. 2 and the profiles of glucose and maltose concentra-
tion are shown in Fig. 3. By the change of morphology,
parameters during fermentation process and lipase pro-
duction characteristics are remarkable different. In both
cultures, the pH increased during the fermentation process,
but the final pH within 96 h was 6.4 for fully entangled
filaments and 6.8 for dispersed mycelia. Increased biomass
levels were obtained in the use of the higher inoculum, so
the biomass for condition A was little lower than condition
B. The most remarkable difference for the two processes
were the lipase production with synthetic activity and
maltose consumption. As Fig. 2a shows, synthetic activity
for mycelium-bound lipase production reached 600 U/g at
60 h, and then maintained at 650 U/g when fully entangled
filaments formed. However, for the dispersed mycelia, the
maximal synthetic activity reached 300 U/g at 36 h, and
then the activity decreased sharply to 60 U/g at 96 h
(Fig. 2b). The same as lipase production, maltose
consumption for fully entangled filaments was remarkably
different from dispersed mycelia. Maltose concentration
decreased from 9.5 g/L to 0.3 g/L within 36 h for fully
entangled filaments. The highest concentration of corre-
sponding hydrolysate glucose was 4 g/L at 60 h and then
decreased to 0 g/L at the end of cultivation time. On other
hand, maltose concentration decreased slowly from 9.5 g/L
to 5.5 g/L within 60 h for dispersed mycelia, and glucose
concentration was maintained at 1 g/L after 48 h (Fig. 3).
From this result, it would be considered that, maltose
consumption was relative to the lipase production. Reasons
for these differences will be discussed in the following
content.
Morphological characterization in shake flask
Not only the macroscopic morphology is important for the
level of enzyme production by filamentous fungi but also
the microscopic morphology influences the production.
Therefore, the same as two typical macromorphology, and
the micro-morphology for two conditions were also dif-
ferent for condition A and B. Fig. 4 presents the
micromorphology for the dispersed mycelia and the fully
entangled filaments obtained by optical microscope and
electron microscope. Hypha from the dispersed mycelia
was fragmentated compare with the normal mycelia of R.
chinensis, the shape of mycelia was not asymmetric, dis-
sepiments appeared and some part of hyphal elements
began to swell (Fig. 4a, c). However, hypha from the fully
entangled filaments was asymmetric, no dissepiments
formed and tips of mycelia were clear (Fig. 4b, d).
Mycelial fragmentation is common in fungal fermentations
[24, 28, 29]. In certain conditions, dissepiments would
formed in some fungal which were actually typical coe-
nocytic mycelium and vacuolation also appeared. Hypha
under this state will lose their biosynthetic activity and the
Fig. 2 Time courses of biomass, pH, synthetic activities for myce-
lium-bound lipase and hydrolytic activity for mycelium-bound lipase
in shake flask cultures inoculated under different conditions. a Fully
entangled filaments process, b Dispersed mycelia process. Darklyfilled triangle biomass, darkly filled circle synthetic activity, opensquare pH
Fig. 3 The profiles of glucose (filled marker) and maltose (openmarker) concentration in two processes: diamond fully entangled
filaments; triangle dispersed mycelia
Bioprocess Biosyst Eng (2009) 32:397–405 401
123

total number of mitochondria. Even though, the vacuola-
tion and mitochondria were not determined in this study,
but abundant inclusion body was observed in fully entan-
gled filaments. Thus, mycelia from fully entangled
filaments could exhibit high biosynthetic activity which
caused rapid maltose consumption high enzyme produc-
tion, and the situation is reverse for mycelia from dispersed
mycelia.
Parameters of micromorphology for two morphologies
were described in Fig. 5. Mean diameter of mycelia from
dispersed mycelia was little higher than the mycelia from
fully entangled filaments, and the mean length of mycelia
from dispersed mycelia was much higher than the mycelia
from fully entangled filaments. The mumber of tips for
fully entangled filaments was higher than the dispersed
mycelia. Because of the tip growth is important for protein
secretion in filamentous fungi [18, 30], some researchers
has focused on this phenomenon and reported several
compellent results [11, 12]. From these results, it would be
concluded that mycelium-bound lipase production by R.
chinensis was associated with tip growth to some extent.
High number of tips could lead to high branches, and the
high branches could cause high level of cell-to-cell inter-
actions which leads to more mycelial aggregation. In
mycelial aggregates, the high level of cell-to-cell interac-
tion and signaling resulting from short diffusional
distances, leads to a state of differentiation quantitatively
different from that of free dispersed mycelia [8, 31].
Lipase production under two typical mycelial
morphology in STR
In our preliminary experiments, we found that morphology
in stirred tank reactor was different from morphology in
shake flask. Fully entangled filaments were difficult to form
in STR, only dispersed mycelia and pellet were achieved.
In order to investigated the change of morphology on broth
rheology and lipase production in STR, two culture con-
ditions were performed as well as in shake flask. For pellet,
condition A was used: agitation speed 400 rpm, inoculum
level 3 9 108 spores/L; For dispersed mycelia, condition B
Fig. 4 Contrast of microscopic morphology for the dispersed mycelia
and the fully entangled filaments: a microscopic morphology of
dispersed mycelia, b microscopic morphology of fully entangled
filaments, c electron microscope picture of dispersed mycelia, delectron microscope picture of fully entangled filaments
Fig. 5 Time courses of mean diameter, equivalent mean length and
average number of tips for filaments during two typical processes.
Open square fully entangled filaments, darkly filled circle dispersed
mycelia
402 Bioprocess Biosyst Eng (2009) 32:397–405
123

was used: Agitation speed 200 rpm, inoculum level
3 9 109 spores/L.
The effects of inoculum and agitation on morphology in
submerged has been discussed above, and were well-
known effects which have been demonstrated in a number
of studies [4, 6, 24–26, 32, 33]. The same as these reports,
pellet formation by R. chinensis is under condition of low
inoculum and high agitation speed. Morphological deter-
mination showed that smooth pellet and hairiness pellet
were obtained simultaneously, and the diameter of these
pellets was 1,000–1,600 lm under condition A.
The time courses for biomass, pH, synthetic activities
for mycelium-bound lipase and dissolved oxygen (DO) are
shown in Fig. 6. The same as shake flask, parameters
during fermentation process and lipase production charac-
teristics were remarkable different for two morphology
process. Biomass levels for condition A was little lower
than condition B because of the inoculum was different.
Specific growth rate for condition A was 0.15/h and 0.13/h
for condition B, this result might due to the high mass
transfer in condition A. Maximum lipase activity was
270 U/g in the case of condition B at 36 h, and then
decreased to 25 U/g at the end of cultivation time.
However, the maximum lipase activity was 315 U/g in the
case of condition A at 36 h, and decreased slowly to
200 U/g at end of fermentations. These result illustrated the
relationship between the morphological change and lipase
biosynthesis in the fermentation medium, with the pellet
form of growth being associated with high lipase
production.
Morphology and broth rheology study in STR
Compare with unicellular microorganism, the broth rheol-
ogy for filamentous fungi is more complex because of the
complex morphological changes for filamentous fungi [17].
Both of the increasing biomass and morphological changes
[34, 35] would lead to non-Newtonian broths, especially
for dispersed mycelia process. Therefore, pellets were
preferred in many researches [15, 36, 37], because of the
most important advantage of the pellet morphology is the
decrease in the viscosity of the culture fluid, resulting in
improved mixing and mass transfer properties.
To investigate the relationship between morphology
change and broth rheology, offline broth rheology study
was performed for two morphologies. As shown in Fig. 7,
it was observed that broth from both processes exhibit a
typical pseudoplastic non-Newtonian behavior and follow
power law model. The viscosity of the broth increased in
the early hours of fermentation and reached to maximum at
about 60 h for pellet and 72 h for dispersed mycelia.
However, broth rheology properties for pellet and dis-
persed mycelia process were significant different. For
dispersed mycelia process, the consistency index (K) was
changed from 1 to 3.5 which indicate high viscosity, and
Fig. 6 The profiles of cell growth, DO, pH, and lipase production
under condition A and B. Condition A, agitation speed 400 rpm,
inoculum level 3 9 108 spores/L; Condition B, agitation speed
200 rpm, inoculum level 3 9 109 spores/L; darkly filled trianglesynthetic activity, open circle DO, darkly filled circle biomass, darklyfilled square pH
Fig. 7 Variations in flow behavior index (n) and consistency index
(K) of fermentation under condition A (dark square) and B (opensquare). Conditon A, agitation speed 400 rpm, inoculum level
3 9 108 spores/L; Conditon B, agitation speed 200 rpm, inoculum
level 3 9 109 spores/L
Bioprocess Biosyst Eng (2009) 32:397–405 403
123

the flow behavior index (n) was changed from 0.2 to 0.4
which indicate a typical pseudoplastic broth. Under this
condition, the mass transfer and broth rheology were
affected significantly by biomass concentration and mor-
phyology change. When the biomass was 18 g/L at
stationary phase (48 h), it would be found that broth around
the wall of fermentor was nearly immovable and the olive
oil in the fermentor could not disperse sufficiently. The oil
clump which caused by the insufficient oil dispersion
resulted in the insufficient contact between oil and cell.
Because the oil is very important for the mycelium-bound
lipase biosynthesis [21], the effect of broth rheology on oil
dispersion would affect the lipase production directly.
For pellet process, the consistency index (K) was
changed from 0.7 to 2.5 which is little lower than the
dispersed mycelia process and the flow behavior index (n)
was changed from 0.4 to 0.7. This result indicated that
viscosity of the broth is lower than dispersed mycelia
process, and non-Newtonian characteristic is also not
obviously. The mixing and mass transfer properties were
improved under this condition, and oil could disperse suf-
ficiently in the fermentation system. Corresponding high
and stable mycelium-bound lipase production was
achieved.
Moreover, DO concentration in the culture broth was
different for different morphology process because of the
different agitation speeds and broth rheology, and high DO
concentration was obtained for pellets process. These
results suggest that high DO concentration may play a key
role in the enhancement of mycelium-bound lipase pro-
duction. It was well known that, even at a high DO
concentration, oxygen transfer would be limited in the
central part of the pellet. In Hilles study [38], they found
that in the pellet growth by A. niger, DO concentration was
nearly zero at pellet core (r = 0–200 lm). Other
researchers also reported that 100 lm is a limitation dis-
tance for mycelium which with biofilm [39]. Investigations
have shown that pellet formation is necessary for the pro-
duction of some enzymes from filamentous
microorganisms, such as polygalacturonase [14, 37], phy-
tase [16], glucoamylase [8], fructosyltransferase [15], and
other proteins [33]. Based on other researchers and their
results, Papagianni concluded that such phenomena may be
related to diffusional limitations in pellets, which either
reduce the extent of catabolic repression in pellets or limit
the oxygen supply, preventing oxidative inactivation of a
specific set of enzymes [1, 8]. Thus, for fully entangled
filaments in shake flask in our study, the mycelium was
more compact than pellet and mass transfer was limited
strictly and a much higher lipase production was obtained
compare with pellet. It is interesting that an enhancement
in intracellular lipase production has been observed with
immobilized cultures or aggregated format in earlier work
[13, 40]. However, those previous works were focused on
the comparison between free and immobilized or aggre-
gated mycelium and did not include studies on the
morphological changes.
Conclusion
The present study on morphological variation during
mycelium-bound lipase production by R. chinensis both in
shake flask and fermentor suggests a direct correlation of
macromorphology and micromorphology changes with the
lipase production. Morphology change is strongly influ-
enced by inoculum level and shear stress both in shake
flask and fermentor. Low inoculum level and high shear
stress results in aggregated mycelia which can produce
more mycelium-bound lipase. Study of micromorphology
reveals that microscopic morphologies for dispersed
mycelia and aggregated mycelia are different in cell shape
and biosynthetic activity. Hypha from fully entangled fil-
aments is asymmetric, no dissepiments formed and tips of
mycelia were clear, nearly 650 U/g activity is achieved
which is ten times higher than the dispersed mycelia.
Macromorphology and broth rheology study in fermentor
indicates that pellet formation results in low broth viscos-
ity. Under this condition, the oil can disperse sufficiently in
broth which is very important for lipase inducement.
However, lipase activity in fermentor is much lower
compare to shake flask. In order to further improve the
lipase activity in fermentor, a deeper understanding of the
aggregated mycelia formation mechanism and detailed
study on effect of different culture conditions on mor-
phology change are necessary.
Acknowledgment Financial supports of National Natural Science
Foundation of China (3047006), the Program for Changjiang Scholars
and Innovative Research Team in University (IRT0532), the Ministry
of Education, PR China under Program for New Century Excellent
Talents in University (NCET-04-0498), The National High Tech-
nology Research and Development Program of China (863)
(2006AA020202) and Program for Hi-Tech Research of Jiangsu
Province (BG2006011) are gratefully acknowledged.
References
1. Papagianni M (2004) Fungal morphology and metabolite pro-
duction in submerged mycelial processes. Biotechnol Adv
22:189–259
2. Thomas CR (1992) Image analysis: putting filamentous micro-
organisms in the picture. Trends Biotechnol 10:343–348
3. Grimm LH, Kelly S, Krull R, Hempel DC (2005) Morphology
and productivity of filamentous fungi. Appl Microbiol Biotechnol
69:375–384
4. Papagianni M, Mattey M (2006) Morphological development of
Aspergillus niger in submerged citric acid fermentation as a
function of the spore inoculum level. Application of neural
404 Bioprocess Biosyst Eng (2009) 32:397–405
123

network and cluster analysis for characterization of mycelial
morphology. Microb Cell Fact 5:12
5. Patnaik PR (2000) Penicillin fermentation: mechanisms and
models for industrial-scale bioreactors. Crit Rev Biotechnol
20:1–15
6. Heydarian SM, Mirjalili N, Ison AP (1999) Effect of shear on
morphology and erythromycin production in Saccharopolysporaerythraea fermentations. Bioprocess Eng 21:31–39
7. Tamura S, Park Y, Toriyama M, Okabe M (1997) Change of
mycelial morphology in tylosin production by batch culture of
Streptomyces fradiae under various shear conditions. J Ferment
Bioeng 83:523–528
8. Papagianni M, Moo-Young M (2002) Protease secretion in glu-
coamylase producer Aspergillus niger cultures: fungal
morphology and inoculum effects. Process Biochem 37:1271–
1278
9. Jin B, van Leeuwen JH, Patel B (1999) Mycelial morphology and
fungal protein production from starch processing wastewater in
submerged cultures of Aspergillus oryzae. Process Biochem
34:335–340
10. El-Enshasy H, Hellmuth K, Rinas U (1999) Fungal morphology
in submerged cultures and its relation to glucose oxidase excre-
tion by recombinant Aspergillus niger. Appl Biochem Biotechnol
81:1–11
11. Spohr A, Carlsen M, Nielsen J, Villadsen J (1997) Morphological
characterization of recombinant strains of Aspergillus oryzaeproducing alpha-amylase during batch cultivations. Biotechnol
Lett 19:257–261
12. Haack MB, Olsson L, Hansen K, Lantz AE (2006) Change in
hyphal morphology of Aspergillus oryzae during fed-batch cul-
tivation. Appl Microbiol Biotechnol 70:482–487
13. Nakashima T, Kyotani S, Izumoto E, Fukuda H (1990) Cell
aggregation as a trigger for enhancement of intracellular lipase
production by a Rhizopus species. J Ferment Bioeng 70:85–89
14. Oncul S, Tari C, Unluturk S (2007) Effect of various process
parameters on morphology, rheology, and polygalacturonase
production by Aspergillus sojae in a batch bioreactor. Biotechnol
Prog 23:836–845
15. Lim JS, Lee JH, Kim JM, Park SW, Kim SW (2006) Effects of
morphology and rheology on neo-fructosyltransferase production
by Penicillium citrinum. Biotechnol Bioprocess Eng 11:100–104
16. Papagianni M, Nokes SE, Filer K (1999) Production of phytase
by Aspergillus niger in submerged and solid-state fermentation.
Process Biochem 35:397–402
17. Olsvisk ES, Kristiansen B (1994) Rheology of filamentous fer-
mentations. Biotechnol Adv 12:1–39
18. Peberdy JF (1994) Protein secretion in filamentous fungi-Trying
to understand a highly productive black-box. Trends Biotechnol
12:50–57
19. Metz B, Kossen NWF (1977) Growth of molds in form of pellets-
Literature-review. Biotechnol Bioeng 19:781–799
20. Xu Y, Wang D, Mu XQ, Zhao GA, Zhang KC (2002) Biosyn-
thesis of ethyl esters of short-chain fatty acids using whole-cell
lipase from Rhizopus chinensis CCTCC M201021 in non-aqueous
phase. J Mol Catal B-Enzym 18:29–37
21. Teng Y, Xu Y (2008) Culture condition improvement for whole-
cell lipase production in submerged fermentation by Rhizopuschinensis using statistical method. Bioresour Technol 99:3900–
3907
22. Dong W, Yan X, Yun T (2007) Synthetic activity enhancement of
membrane-bound lipase from Rhizopus chinensis by pretreatment
with isooctane. Bioprocess Biosys Eng 30:147–155
23. Mitschka P (1982) Simple conversion of brookfield Rvt readings
into viscosity functions. Rheol Acta 21:207–209
24. Nielsen J, Johansen CL, Jacobsen M, Krabben P, Villadsen J
(1995) Pellet formation and fragmentation in submerged cultures
of Penicillium chrysogenum and its relation to penicillin pro-
duction. Biotechnol Prog 11:93–98
25. Tucker KG, Thomas CR (1992) Mycelial morphology: the effect
of spore inoculum level. Biotechnol Lett 14:1071–1074
26. Kelly S, Grimm LH, Hengstler J, Schultheis E, Krull R, Hempel
DC (2004) Agitation effects on submerged growth and product
formation of Aspergillus niger. Bioprocess Biosyst Eng 26:315–
323
27. Du LX, Jia SJ, Lu FP (2003) Morphological changes of Rhizopuschinesis 12 in submerged culture and its relationship with anti-
biotic production. Process Biochem 38:1643–1646
28. McNeil B, Berry DR, Harvey LR, Grant A, White S (1998)Measurement of autolysis in submerged batch cultures of Peni-cillium chrysogenum. Biotechnol Bioeng 57:297–305
29. Harvey LM, McNeil B, Berry DR, White S (1998) Autolysis in
batch cultures of Penicillium chrysogenum at varying agitation
rates. Enzyme Microb Technol 22:446–458
30. Wosten HAB, Moukha SM, Sietsma JH, Wessels JGH (1991)
Localization of growth and secretion of proteins in Aspergillusniger. J Gen Microbiol 137:2017–2023
31. Braun S, Vechtlifshitz SE (1991) Mycellal morphology and
metabolite production. Trends Biotechnol. Trends Biotechnol
9:63–68
32. Amanullah A, Christensen LH, Hansen K, Nienow AW, Thomas
CR (2002) Dependence of morphology on agitation intensity in
fed-batch cultures of Aspergillus oryzae and its implications for
recombinant protein production. Biotechnol Bioeng 77:815–826
33. El-Enshasy H, Kleine J, Rinas U (2006) Agitation effects on
morphology and protein productive fractions of filamentous and
pelleted growth forms of recombinant Aspergillus niger. Process
Biochem 41:2103–2112
34. Olsvik E, Tucker KG, Thomas CR, Kristiansen B (1993) Corre-
lation of Aspergillus niger broth rheological properties with
biomass concentration and the shape of mycelial aggregates.
Biotechnol Bioeng 42:1046–1052
35. Riley GL, Tucker KG, Paul GC, Thomas CR (2000) Effect of
biomass concentration and mycelial morphology on fermentation
broth rheology. Biotechnol Bioeng 68:160–172
36. Lopez JLC, Perez JAS, Sevilla JMF, Porcel EMR, Chisti Y
(2005) Pellet morphology, culture rheology and lovastatin pro-
duction in cultures of Aspergillus terreus. J Biotechnol 116:61–77
37. Gogus N, Tari C, Oncu S, Unluturk S, Tokatli F (2006) Rela-
tionship between morphology, rheology and polygalacturonase
production by Aspergillus sojae ATCC 20235 in submerged
cultures. Biochem Eng J 32:171–178
38. Hille A, Neu TR, Hempel DC, Horn H (2005) Oxygen profiles
and biomass distribution in biopellets of Aspergillus niger. Bio-
technol Bioeng 92:614–623
39. Viniegra-Gonzalez G, Favela-Torres E (2006) Why solid-state
fermentation seems to be resistant to catabolite repression. Food
Technol Biotechnol 44:397–406
40. Nakashima T, Fukuda H, Nojima Y, Nagai S (1989) Intracellular
lipase production by Rhizopus chinensis using biomass support
particles in a circulating bed fermenter. J Ferment Bioeng 68:19–
24
Bioprocess Biosyst Eng (2009) 32:397–405 405
123










![SHELL MYCELIUM [degradation movement manifesto]](https://img.pdfslide.us/doc/110x75/6191c09297e7f8291a6419a6/shell-mycelium-degradation-movement-manifesto.jpg)

















