Embed Size (px)
Citation preview

OCEANOLOGICA ACTA- VOL. 19- W 3-4 ~ -----~-
Change from planktonic to benthic development:
Polychaeta Terebellidae
Life cycles Dispersal
Structures of retention Evolution
is life cycle evolution an adaptive answer to the constraints of dispersal?
Polychètes Térébellidae Cycle de vie
Dissémination Structures de rétention
Evolution
ABSTRACT
RÉSUMÉ
Michel BHAUD and Jean-Claude DUCHÊNE
Observatoire océanologique, Université Paris VI - Pierre et Marie Curie, Laboratoire Arago, BP 44, 66651 Banyuls-sur-mer Cedex, France.
Received 30/03/95, in revised form 23/10/95, accepted 14/11/95.
Current research on recruitment of benthic marine invertebrates is, to a large extent, concemed with the consequences of larval planktonic development in water masses entailing dispersal, a milieu which appears unfavourable for a successful recruitment. In this paper, the origin of benthic development is tentatively considered as a response to the constraints of dispersal. Examples among annelid polychaetes are taken from persona! work and from the literature. i) Life cycles in the subantarctic area provide good indications of a correlation between direct development and environmental pressure enhancing dispersal (due to turbulence and advection which would carry the planktonic larvae away from a suitable habitat). ii) Experimental analysis of the li fe cycle of Eupolymnia nebulosa in the Mediterranean shows that gelatinous egg masses reduce the ability of larvae to spread and consequently increase retention in areas more suitable for the adults. iii) Examination of a large number of geographie sites where a very reduced habitat area is reported indicates that developmental patterns may involve weak dispersal to secure the return of Iarvae. From an evolutionary point of view, the direction of the transformation of life cycle patterns in the polychaete class may be inferred from the large number of solutions adopted by benthic development. The observations suggest that this pattern is derived. The case of E. nebulosa, which exhibits two developmental strategies, suggests also that the direct development found in populations of the Mediterranean Sea is a derived condition from the free development found elsewhere e.g. the English Channel. Evolution from benthopelagic development to benthic development is frequent, yet the reversion to planktonic development is also observed. We suggest that changes in the physical conditions of the environment disrupt and modify the life cycle; these unbalanced conditions may occur several times over the course of the evolution of a family.
Du développement planctonique au développement benthique : l'évolution des cycles de vie est-elle une réponse aux contraintes de dissémination ?
Les recherches actuelles sur le recrutement s'appliquent pour une large part aux conséquences du développement larvaire planctonique dans un milieu dispersif qui apparaît négatif pour un recrutement satisfaisant. A l'échelle de temps évolutive, ce couplage soulève la question de l'origine du développement benthique. On tente de montrer que ce type de développement chez les annélides polychètes est considéré comme une réponse aux contraintes de dissémination. Pour cette
335

M. BHAUD, J.-C. DUCHÊNE
démonstration on examine plusieurs situations tirées de nos propres travaux et de la littérature. a) Les cycles de vie de la province subantarctique fournissent une bonne corrélation entre le développement direct prédominant et les forces de dissémination extrêmement prononcées. b) L'analyse du cycle de vie de Eupolymnia nebulosa permet de démontrer que les masses gélatineuses contenant les larves réduisent la capacité de dissémination de ces larves et empêchent la colonisation sur des zones défavorables au maintien des adultes. c) L'examen d'un ensemble d'espèces, caractérisées par une surface d'habitat très réduite, indique la nécessité d'une faible dissémination. Du point de vue évolutif, la direction de la transformation du type de développement peut être identifiée. A l'échelle de la classe des Polychètes, le grand nombre de solutions adoptées par le développement direct suggère une origine secondaire de ce mode de développement. Le cas particulier fourni par E. nebulosa avec deux stratégies de développement, suggère que le contrôle des larves en Méditerranée est dérivé par rapport au développement libre connu sur tout le reste de l'aire spécifique. Cependant cette direction d'évolution n'est probablement pas unique. Le retour vers le développement benthoplanctonique est aussi observé et l'on suggère que les variations des conditions physiques de l'environnement peuvent briser le cycle de vie et le modifier. Ce déséquilibre peut se produire plusieurs fois et dans des directions différentes au cours de la vie de la lignée familiale.
OceanologicaActa, 1996, 19,3-4,335-346.
INTRODUCTION
Among marine invertebrates, three spatially defined modes of development are generally recognized: benthoplanktonic, holoplanktonic and holobenthic. The benthoplanktonic pattern takes place in two ecosystems: the plankton and the benthos; the transfer from one to the other occurs with an ontogenetic transformation, namely metamorphosis. Such "complex life cycles" (Sinclair, 1988) are widespread in marine systems where approximately 80 % of species exhibit a life cycle that includes a thorough metamorphosis, often accompanied by a spatial transfer. Severa! conditions are required for a complex life cycle to occur and to ensure the perpetuity of the species. Constraints fall into two large categories: those developed in time relative to the different steps of ontogenesis (Bhaud, 1994); and those linked with space. In this paper we will deal with this second kind of constraint. Sorne difficulties concerning the achievement of bentho-planktonic life cycles can be expected, as development must alternate between two different ecosystems. Though advection and turbulence of water masses can be efficient in spreading passive larvae, these conditions can also be dangerous to the larvae as they may be carried away from a suitable habitat; to ensure adult survival, the larvae must reach a suitable benthic environment. In contrast, holoplanktonic or holobenthic development occurs in only one ecosystem. Species adopting these patterns do not need to change from one ecosytem to another to complete their life cycle; thereby they avoid the quest for a suitable environment.
Many papers deal with biological-physical interactions (as summarized in Mann and Lazier, 1991; Rothschild, 1988) and especially with the importance of physical structures on the maintenance of populations (Banse, 1986; Alldredge and Hamner, 1980; Kingsford, 1990; Lefèvre, 1990; Pineda, 1991; Rothlisberg et al., 1983). Ali these papers come to similar conclusions concerning the need to take watermass circulation into account in order to unders-
336
tand geographical population patterns and levels of recroîtment. There are divergent views on the advantages/disadvantages of dispersal during the larval phase of benthic species. Palmer and Strathmann ( 1981) state that feeding larvae living simultaneously in the plankton receive little if any advantage from an increased scale of dispersal. Grahame and Branch ( 1985) after examining a variety of opinions on the utility of the planktonic period state that with ali these contrasting views, the significance of the dispersal phase linked with pelagie development remains problematic. Sinclair ( 1988) offers good arguments that retention by physical oceanographie processes is a matter of selection rather than of dispersal. Roughgarden ( 1989) argues that if dispersal were the paramount factor in a life cycle, one would expect the dispersal to be carried out by the adult when development is benthic.
If physical oceanographie processes create conditions for successful reproduction, it is of interest to investigate, on the evolutionary time scale, the possible selective pressures of advection and mixing. Most discussions on evolution of life cycles concern strategies developed only by species that are benthic when adult. Holoplanktonic cycles are generally neglected (Chia, 1976; Cazaux, 1981; Jablonski and Lutz, 1983; Bhaud, 1994; Mileikovsky, 1971, 1976; Scheltema, 1989). More often, the problem is limited to the energy requirements in terms of egg size and storage content for species with contrasting life cycles. Environmental conditions relative to spatial constraints are neglected. Another complication is that observed strategies are assumed to have emerged from an ancestral life history pattern. The prevailing opinion is that the benthoplanktonic developmental pattern is probably the ancestral condition (Jagersten, 1972; Schroeder and Hermans, 1975; Strathmann, 1985; Radashevsky, 1994 ). Reproductive strategies take root in both the phyletic history of the group (ultimate causation) and in the changing environmental conditions (proximate causation). Among the latter, constraints of dispersal and energy available for development have been

recognized. Three life-history patterns are available to reduce dispersal (Fig. 1): i) evolution towards benthic development through a reduction in, or the suppression of, the planktonic larval phase; ii) conservation of the benthoplanktonie life cycle but with behavioural adaptation of planktonic larvae (such as vertical migrations) or of adults during the reproductive period (such as timing and placing of spawning relatively to the tidal cycle); these reasons explain why benthoplanktonie development is still so frequent; and iii) evolution towards holoplanktonic Iife cycles. This latter solution of the dispersal problem seems paradoxical but it suppresses the transfer from one ecosystem to another during metamorphosis. Ali Iife history stages of a population are subject to similar transport conditions and may thus develop processes for schooling or aggregation (O'Brien, 1988; Alldredge and Hamner, 1980).
The idea of considering physical structures in the ocean to be important for success in the life cycle is not new from an evolutionary point of view. Sinclair (1988, p. 184) suggests that evolution to direct development is externally forced, in the oceans, by changes in the physical geography that disrupt the existent tight linkage between the morphologieal and behavioural cycles of the animal and the associated partieular geography. Evolution may lead to holobenthic or holoplanktonic development. According to the same author, speciation in the oceans can be thought of as a repetitive contraction and expansion of complex life cycles in response to a nonequilibrium in the physical environment. The constraint of sexual reproduction is the requirement to find a mate in a dispersive environment and the process of life-cycle selection, in adjusting to such nonequilibrium conditions, results in speciation. Following this idea, it appears that it is possible to go back and forth from one type of life cycle to another. Two questions will be investigated: the role of spatial dispersal as a source of selection in the developmental pattern with a shift from complex development to holobenthie patterns; and the exclusive or non exclusive nature of this direction.
MATERIAL AND METHODS
The interaction between larval behaviour and the physical structure of water masses is increasingly weil understood (Rothschild, 1988; Sinclair, 1988; Lefèvre, 1990; Kingsford, 1990; Pineda, 1991) for many invertebrates but studies are still scarce for polychaetes. For instance, Sinclair (1988, table 6.1, p. 104) does not mention any studies describing retention of the pelagie stages in the life of benthic species among Polychaeta. This class is useful for comparison if we consider that, among the main groups of marine invertebrates, polychaetes exhibit the largest variety of strategies (Giangrande et al., 1994). The aim of this paper is to report on numerous situations in this group. To do so, we will use our own work carried out in both subantarctic and temperate areas. The detailed analysis of Eupo/ymnia nebulosa Iife cycle will be of prime interest for the present objective, which is to identify relations between physieal structures of the environment and specifie features of the !ife cycle. First we will examine the reproductive pattern in
337
LI FE CYCLE EVOLUTION IN POL YCHAETES
Three strategies to esape from dispersal constralnts
/ progressive
reduction of tbe planktonic life
leading to pelagie Iecidtotrophic
development (endotrophy)
direct or holobendtic development
Figure 1
bendto-planktonic development (e<otrophy)
long larval planktonic life
conservation of tbe larval planktonic stage but
development of adaptations in adults (spawning period linked with tide) or larvae
(vertical ntigrations) to hydrodynantic structures.
bendto-planktonic development
integration of dissentination and turbulence into !he adaptive response:
suppression of benthic stage: !he cycle
is holoplanktonic.
holoplanktonic development
Three ways of escaping from dispersal constraints of complex !ife cycles among Polychaeta. First column: the route to direct development with, as an example, Streblospio shrubsolii; the first benthic stage leaving the mother's tube is figured close to her body (from Cazaux, 1985). Middle column: conservation of the benthoplanktonic /ife cycle with development of adaptations in the behaviour of larvae; 1: Sabellaria alveolata, larvae must retum to the reejv built by adults; 2: aulophore of Lanice conchilega (Terebellidae with a long planktonic life supported by a post-larval stage); the consequence of a relative autonomy of these larvae is a decoupling between larval length and geographie displacement. Third column: evolution towards a holoplanktonic /ife cycle (Tomopteridae).
a subantarctic area where turbulence and currents are extreme; then we will describe a specifie case (E. nebulosa) with an analysis of the relation between habitat size and pattern of development; and finally we will attempt to widen the scope by including a series of species for which habitat components and developmental patterns are known.
RESULTS
The Kerguelen Archipelago example
The hypothesis of the emergence of direct development patterns through constraints of dispersal is perfectly illustrated by the study of polychaetes from the Kerguelen province (Duchêne, 1983, 1989, 1992; Bhaud and Duchêne, 1988). Among species with known life cycles, development is most frequently direct with large-sized eggs. lt is generally accepted that there is a relation between developmental pattern and egg size. The size limit is close to lOO-

M. BHAUD, J.-C. DUCHÊNE
150 J.IID, smaller-sized eggs produce larvae with planktonic development. In Kerguelen the majority of species undergo direct development (Tab. 1) and egg size is 200, 450 and 600 J.lffi in Amphicteis gunneri, Platynereis magalhaensis and Neoleprea streptochaeta, respectively. On the other hand, eggs of Boccardia polybranchia reach a diameter of only 80 mm and we therefore expect to find indirect development. In fact development in B. polybranchia is accomplished in capsules enclosing 40 to 90 eggs (Duchêne, 1984 ). Sorne eggs develop into fast growing larvae that feed on abortive eggs, while a few planktotrophic-like larvae are kept inside the egg mass until the opening of the capsule wall (Duchêne, 1992). In this species, characteristics of indirect development are perfectly reflected with marked exotrophy, but spatial dispersal cannot take place. This situation is not a demonstration of evolution going from indirect to direct development, but provides a strong correlation between direct development and the presence of enhanced dispersive conditions. In the same area but for other groups, such as crustaceans, complex development is maintained. In this case an adaption for reduced dispersal is formed by holdfasts of giant kelp like Macrocystis and Durvillea (Bhaud and Duchêne, 1988). These examples of attachment of small eggs on a fixed structure not linked to the animais are a particularly good illustration of adaptation for reduced dispersal. That the oceanographie constraints in the Kerguelen Archipelago are characterized by strong currents and dispersal is demonstrated in Duchêne ( 1989) and Gamberoni et al. ( 1982).
The Eupolymnia nebulosa example
The experimental study of several parts of the life cycle of this species reveals a means of counteracting dispersal. Populations are differentiated between the Atlantic and the Mediterranean Sea in terms of the existence, in the Mediterranean Sea, and absence from the Atlantic and the Channel, of a gelatinous egg mass in which larvae undergo direct development. This divergence is probably rather recent and connected with the history of the Mediterranean Sea (Lenaers and Bhaud, 1992). In the Bay of Banyuls, E. ne bu/osa is one of the most abundant species for which a large body of biological information is available. lts life
Table 1
cycle has been described (Bhaud and Grémare, 1988 and 1991) and experimental studies investigating seulement requirements have also been undertaken (Bhaud, 1988 a, b; 1990, 1991; Cha and Bhaud, 1991; Cha, 1994). Adults are collected in a narrow area, at the periphery of the Banyuls Bay. They inhabit a tube of coarse sediment formed at the interface between stones and the underlying sediment. Spawning occurs between March and the end of May, sometimes between the end of February and early June. Spawning activity is strongly rhythmic, four or five maxima are recorded each year. The highest spawning peak occurs around the last quarter to the new moon in April. In the laboratory the larvae have the ability to use the finest fraction of heterogeneous sediments present in the centre of the Bay of Banyuls: muddy sand from the Nephtys hombergii community (Bhaud and Cha, 1994 ). However adult E. nebulosa are never collected in this community. How then can we reconcile the absence of the species in the axis of the Bay with successful seulement on the same substratum in the laboratory ? We showed, by studying tube-dwelling juveniles in a hydrodynamic canal, that displacement of tubes built on soft substrates of the Nephtys hombergii community may be different according to the thickness of the sediment. When tubes are arranged on a sedimentary bed as in the centre of the Bay, they are easily washed away by water currents. On the other hand, when tubes are anchored on a thin layer of the same sediment deposited on a hard substratum like at the periphery of the bay, they off er a higher resistance to the erosion by currents. In the first case a speed of 16 cm.ç1 detaches the tubes, whereas in the second case a flow rate of 24 cm.s-1, which is the upper limit of the flow reached in the canal, does not detach the tubes, (Cha, 1994; Bhaud et al., 1995). These results suggest that the role of gelatinous egg masses reduces the ability of larvae to spread and prevents colonization in the centre of the Bay. Even if larvae are able to colonize this bay, it does not appear desirable that they reach this area, which forms an unsuitable habitat for adults.
Development in specialized environments
In this part, we apply the situation of E. nebulosa Mediterranean populations (i.e. a reduced adult area) to other spe-
Elements of the !ife history in sorne species among polychaetes from Kerguelen. In al! cases but one, egg size is large, indicating direct development. For the exception B. polybranchia, the small size of oocytes suggests indirect (or planktonic) development but adaptation to a direct development is obtained by encapsulation of eggs; exotrophy is based on feeding on abortive eggs or on eggs that develop late.
Species
Platynereis magalhaensis
Amphicteis gunneri
Thelepus extensus (forrnerly setosus)
Neoleprea streptochaeta
Flabelligera pennigera
Boccardia polybranchia
Cirratulus cirratus
Egg diameter (pm)
450 200
210
600
500
80
500
Observations on larval development
eggs laid in a jelly mass inside the tube built on Macrocystis fronds
no direct observations but likely existence of short planktonic life deduced from development of earl y juveniles
spherical egg masses forrned by the female and attached at the opening of the tube; severa) egg masses forrned by one female
no jelly mass; incubation of embryos and Jarvae in the tube, hatching 20 days at 5° C after spawning; no free-swimming Jife
incubation of embryos and Jarvae in the mucus surrounding the female body
adelphophagy and poecilogony; benthic individuals escaping from the capsules at stage of 10 to 18 setigerous segments
Jarvae held between tentacular cirri; ciliated belt absent
338

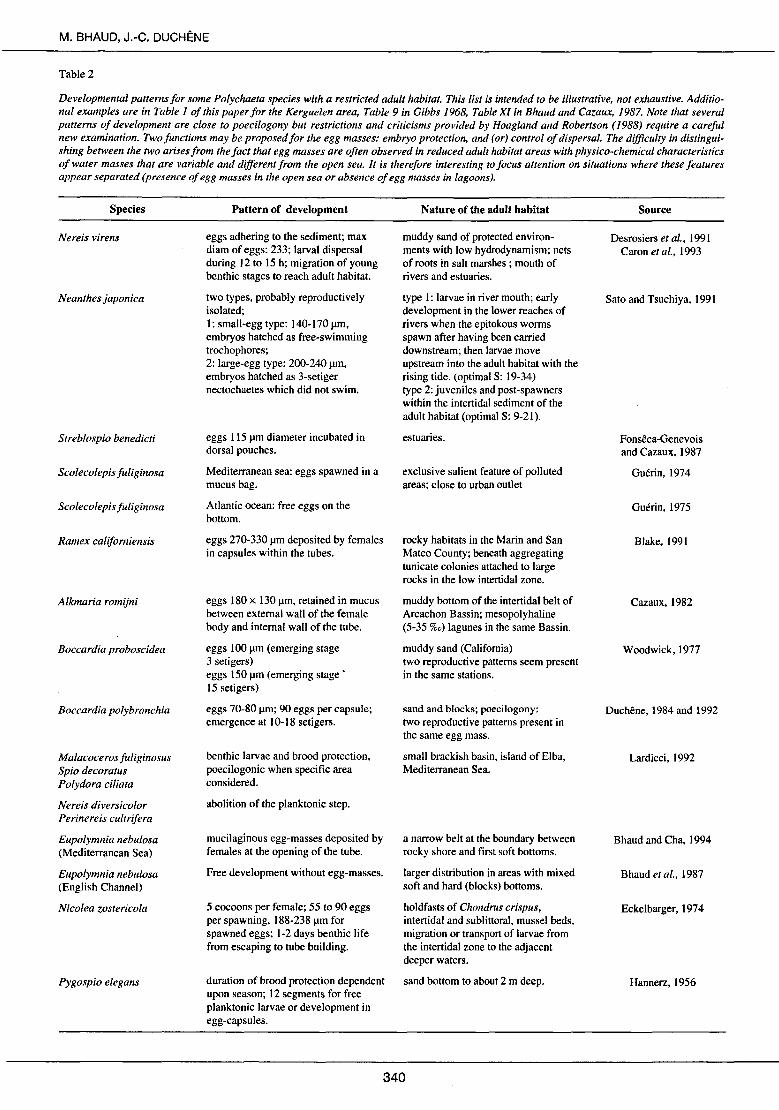
cies. They inhabit a specialized topographical environment (bays, internai neritic zones, lagoons, hydrothermal vents, islands, fissured hard bottom) often coupled with a specialized ecological environment (determined by temperature, hydrodynamics, salinity). The high frequency of spawning mucilaginous structures in species inhabiting particular ecological zones supports the hypothesis of the need to control of dispersal in certain environments. For these species which inhabit an environment characterized by very peculiar ecological features, the reduction of dispersal is a prerequisite for the maintenance of populations (Tab. 2 ). This is the idea developed by Gibbs (1968) with regard to the study of development of Scoloplos · armiger by Chapman (1965). The most important fonction of the cocoon in Scoloplos is thought to prevent the wide dispersal of the larvae (see also Plate and Husemann, 1991 ). The habitat of Scoloplos is restricted to fairly sheltered shores and thus it is probably advantageous to retain the larvae within the favourable conditions of the adult habitat. In the same way, Opheliidae (O. bicornis on the south-west coast of England or O. limacina in the Mediterranean Sea) are polychaetes living in a very restricted and specialized habitat (Harris, 1994): in dean and loose sand when the pelitic fraction is totally absent, at high level in very shallow habitats in tideor current-swept bays or estuaries. To ensure that larvae stay in this restricted habitat, planktonic larval life is short and does not last longer than 4-5 da ys.
The different situations shown in Table 2 may be shared in two kinds of topographie conditions: i) closed areas in which ecological situations are extreme, and where planktonic development does not exist or is present only in a small number of species, whereas benthic development is very varied, and among which egg masses are common; and ii) open sea areas with species developing with a short pelagie phase or with egg masses. With regard to the problem stated in the introduction - benthic development as a response to constraints of dispersal -, the question is to determine whether or not the benthic pattern of development is related to particular conditions of the specialized environment or to the limited dimension of the area in which these conditions occur. In other words, it seems necessary to define the role played by physical or biological structures sheltering larvae. Severa! advantages have been attributed to direct development: to make it easier to survive in a harsh environment (Vance, 1973a, b; Christiansen and Fenchel, 1979; Wilson, 1986); to reduce energy expenditure which was thought to be high in the benthoplanktonic life cycle (Chia, 1976); to reduce the predation thought to be important during planktonic life (Pechenik, 1979); to allow gonad maturation in small-sized adults which are not able to form the minimum number of planktonic larvae required to balance loss by dispersal (Strathmann, 1982); and to prevent removing of juveniles from suitable habitats (Chapman, 1965; Gibbs, 1968). As far as we know, these various hypotheses are not supported by clear evidence even if the latter one is most likely. From situations given in Table 2, a precise and immediate answer is not possible because information on the habitat extent is not always given. However, three observations may be added. Firstly, for a part of the reported cases, particularly in the open sea or when lagoons develop large
339
LI FE CYCLE EVOLUTION IN POL YCHAETES
exchanges with the sea, restricted location is not associated with a modification of the environment bearing, for instance, on the concentration of organic matter or special salinity. Short planktonic life and egg masses have the same finality. This is an indication that control of dispersal is at the outset of the benthic developmental type. Secondly, when, for weil known situations as for E. nebulosa or Scoloplos armiger, the length of the free developmental phase is placed in relation to the adult habitat, the physical dispersal appears negative for success of the life cycle and thus egg masses or short planktonic life are advantageous for the persistence of populations. Lastly, experiments and indirect deductions on the presence of spawning egg masses of E. nebulosa (Cha, 1994; Medernach, 1995) indicate that salinity inside the mucilaginous mass follows external conditions very quickly and that egg masses probably do not form an efficient barrier against sharp variations of salinity. These facts suggest that egg masses are more a protection against dispersal than against predation or variation of osmolarity.
Interestingly, the conclusion on the importance of spatial control of dispersal is in accordance with the findings of Swedmark (1959) who noted similar reproductive modifications to prevent horizontal dislocation in microfaunal species confined to a restricted habitat. Retention of larvae in interstitial water due to lack of reaction to light is considered by this author an absolute necessity for the survival of the populations on account of the low number of gametes.
Species living close to deep-sea hydrothermal vents are led to resolve two constraints which may appear conflicting: it is necessary to secure the survival of the population in this insular environment; but also to be able to colonize new vents as soon as they appear. For the first problem, and in the case of Paralvinella grasslei, the large size of eggs (maximum of 275 Jlm) is connected with lecithotrophic development which restricts dispersal. However, this adaptation seems insufficient and is completed by fertilization secured by the presence of spermatheca in females and seminal vesicles in males. These anatomical adaptations with the mating process diminish the dispersal of gametes. The presence of spermatophores in interstitial species probably bas the same significance (Swedmark, 1959). McHugh and Tunnicliffe (1994) study the hydrothermal vent polychaete Amphisamytha galapagensis (Ampharetidae); they point out that this species occurs in a wide variety of vent habitats and that reproduction is probably continuous, fertilization externat, and larvae demersal and lecithotrophic. For the second problem, two possibilities have been suggested: influence of near-bottom currents and phoresy (Zal et al., 1995).
On the other band, species developing a large habitat at great depth may have a long planktonic larval life. Thus a species of Poecilochaetidae reproduces by planktonic larvae (Bhaud and Cazaux, 1987). These larvae are always caught in the open sea and the surface water layer, never on the shelf, both in the Atlantic and in the Mediterranean. In the latter environment their distribution shows no sign of any concentration in a particular zone. This is probably a rare example among Polychaeta of a deep-sea species with a planktonic larval stage in the reproductive cycle.
M. BHAUD, J.-C. DUCHÊNE
Table 2
Developmental patterns for sorne Polychaeta species with a restricted adult habitat. This list is intended to be illustrative, not exhaustive. Additional examples are in Table 1 of this paper for the Kerguelen area, Table 9 in Gibbs 1968, Table Xl in Bhaud and Cazaux, 1987. Note that severa[ patterns of development are close to poecilogony but restrictions and criticisms provided by Hoagland and Robertson ( 1988) require a careful new examination. Two functions may be proposed for the egg masses: embryo protection, and (or) control of dispersal. The difficulty in distinguishing between the two arises from the fact that egg masses are often observed in reduced adult habitat areas with physico-chemical characteristics of water masses that are variable and different from the open sea. It is therefore interesting to focus attention on situations where these features appear separated (presence of egg masses in the open sea or absence of egg masses in lagoons).
Species
Ne reis virens
Neanthes japonica
Streblospio benedicti
Scolecolepis fuliginosa
Scolecolepis fuliginosa
Ramex californiensis
Alkmaria romijni
Boccardia proboscidea
Boccardia polybranchia
Malacocerosfuliginosus Spio decoratus Polydora ciliata
Ne reis diversicolor Perinereis cultrifera
Eupolymnia nebulosa (Mediterranean Sea)
Eupolymnia nebulosa (English Channel)
Nicolea zostericola
Pygospio elegans
Pattern of development
eggs adhering to the sediment; max diam of eggs: 233; larval dispersal during 12 to 15 h; migration of young benthic stages to reach adult habitat.
two types, probably reproductively isolated; 1: small-egg type: 140-170 ~m. embryos hatched as free-swimming trochophores; 2: large-egg type: 200-240 ~m. embryos hatched as 3-setiger nectochaetes which did not swim.
eggs 115 ~rn diameter incubated in dorsal pouches.
Mediterranean sea: eggs spawned in a mucus bag.
Atlantic ocean: free eggs on the bottom.
eggs 270-330 ~rn deposited by females in capsules within the tubes.
eggs 180 x 130 ~m. retained in mucus between externat wall of the female body and internai wall of the tube.
eggs 100 ~rn (emerging stage 3 setigers) eggs 150 ~rn (emerging stage' 15 setigers)
eggs 70-80 f.tm; 90 eggs per capsule; emergence at 1 0-18 setigers.
benthic larvae and brood protection, poecilogonic when specifie area considered.
abolition of the planktonic step.
mucilaginous egg-masses deposited by females at the opening of the tube.
Free development without egg-masses.
5 cocoons per female; 55 to 90 eggs per spawning, 188-238 ~rn for spawned eggs; 1-2 days benthic life from escaping to tube building.
duration of brood protection dependent upon season; 12 segments for free planktonic larvae or development in egg-capsules.
Nature of the adult habitat
muddy sand of protected environmenis with low hydrodynamism; nets of roots in salt marshes ; mouth of rivers and estuaries.
type 1: larvae in river mouth; earl y development in the lower reaches of ri vers when the epitokous worms spawn after having been carried downstream; then larvae move upstream into the adult habitat with the rising tide. (optimal S: 19-34) type 2: juveniles and post-spawners within the intertidal sediment of the adult habitat (optimal S: 9-21).
estuaries.
exclusive salient feature of polluted areas; close to urban outlet
rocky habitats in the Marin and San Mateo County; beneath aggregating tunicate colonies attached to large rocks in the low intertidal zone.
muddy bottom of the intertidal belt of Arcachon Bassin; mesopolyhaline (5-35 %o) lagunes in the same Bassin.
muddy sand (California) two reproductive patterns seem present in the same stations.
sand and blocks; poecilogony: two reproductive patterns present in the same egg mass.
small brackish basin, island of El ba, Mediterranean Sea.
a narrow belt at the boundary between rocky shore and first soft bottoms.
larger distribution in areas with mixed soft and hard (blocks) bottoms.
holdfasts of Chondrus crispus, intertidal and sublittoral, musse) beds, migration or transport of Iarvae from the intertidal zone to the adjacent deeper waters.
sand bottom to about 2 rn deep.
340
Source
Desrosiers et al., 1991 Caron et al., 1993
Sato and Tsuchiya, 1991
Fonsêca-Genevois and Cazaux, 1987
Guérin, 1974
Guérin, 1975
Blake, 1991
Cazaux, 1982
Woodwick, 1977
Duchêne, 1984 and 1992
Lardicci, 1992
Bhaud and Cha, 1994
Bhaud et al., 1987
Eckelbarger, 1974
Hannerz, 1956
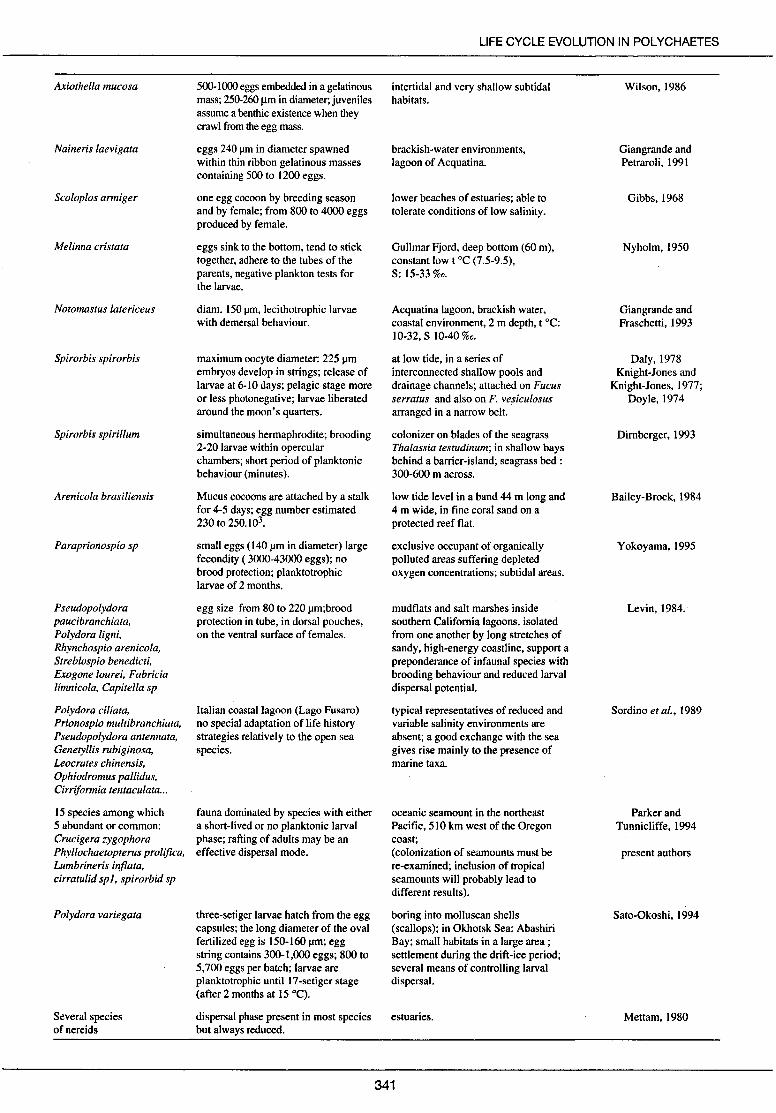
Axiothella mucosa
Naineris laevigata
Scoloplos armiger
Melinna cristata
Notomastus latericeus
Spirorbis spirorbis
Spirorbis spirillum
Arenicola brasiliensis
Paraprionospio sp
Pseudopolydora paucibranchiata, Polydora ligni, Rhynchospio arenicola, Streblospio benedicti, Exogone lourei, Fabricia limnicola, Capite lia sp
Polydora ciliata, Prionospio multibranchiata, Pseudopolydora antennata, Genetyllis rubiginosa, Leocrates chinensis, Ophiodromus pallidus, Cirriformia tentaculata ...
15 species among which 5 abundant or common: Crucigera zygophora Phyllochaetopterus prolifica, Lumbrineris injlata, cirratulid spi, spirorbid sp
Polydora variegata
Severa! species ofnereids
500-1000 eggs embedded in a gelatinous mass; 250-260 Jlm in diameter;juveniles assume a benthic existence when they crawl from the egg mass.
eggs 240 J.lffi in diameter spawned within thin ribbon gelatinous masses containing 500 to 1200 eggs.
one egg cocoon by breeding season and by female; from 800 to 4000 eggs produced by female.
eggs sink to the bottom, tend to stick together, adhere to the tubes of the parents, negative plankton tests for the larvae.
diam. 150 J.IID, lecithotrophic larvae with demersal behaviour.
maximum oocyte diameter: 225 J.lffi embryos develop in strings; release of larvae at 6-10 da ys; pelagie stage more or Jess photonegative; larvae liberated around the moon's quarters.
simultaneous hermaphrodite; brooding 2-20 larvae within opercular chambers; short period of planktonic behaviour (minutes).
Mucus cocoons are attached by a stalk for 4-5 days; egg number estimated 230 to 250.103•
small eggs (140 J.lffi in diameter) large fecondity ( 3000-43000 eggs); no brood protection; planktotrophic larvae of 2 months.
egg size from 80 to 220 J.lm;brood protection in tube, in dorsal pouches, on the ventral surface of females.
ltalian coastallagoon (Lago Fusaro) no special adaptation of life history strategies relatively to the open sea species.
fauna dominated by species with either a short-lived or no planktonic Jarval phase; rafting of adults may be an effective dispersal mode.
three-setiger larvae hatch from the egg capsules; the long diameter of the oval fertilized egg is 150-160 J.lm; egg string con tains 300-1,000 eggs; 800 to 5,700 eggs per batch; larvae are planktotrophic until 17-setiger stage (after 2 months at 15 °C).
dispersal phase present in most species but al ways reduced.
LI FE CYCLE EVOLUTION IN POL YCHAETES
intertidal and very shallow subtidal habitats.
brackish-water environments, lagoon of Acquatina.
lower beaches of estuaries; able to tolerate conditions of low salinity.
Gullmar Fjord, deep bottom (60 rn), constant low t °C (7.5-9.5), S: 15-33 %o.
Acquatina lagoon, brackish water, coastal environment, 2 rn depth, t oc: 10-32, S 10-40 %o.
at low tide, in a series of interconnected shallow pools and drainage channels; attached on Fucus serratus and also on F. vesiculosus arranged in a narrow belt.
colonizer on blades of the seagrass Thalassia testudinum; in shallow hays behind a barrier-island; seagrass bed : 300-600 rn across.
low tide leve! in a band 44 rn long and 4 rn wide, in fine coral sand on a protected reef flat.
exclusive occupant of organically polluted areas suffering depleted oxygen concentrations; subtidal areas.
mudflats and salt marshes inside southem Califomia lagoons, isolated from one another by long stretches of sandy, high-energy coastline, support a preponderance of infaunal species with brooding behaviour and reduced larval dispersal potential.
typical representatives of reduced and variable salinity environments are absent; a good exchange with the sea gives rise main! y to the presence of marine taxa.
oceanic seamount in the northeast Pacifie, 510 km west of the Oregon coast; (colonization of seamounts must be re-examined; inclusion of tropical seamounts will probably lead to different results ).
boring into molluscan shells (scallops); in Okhotsk Sea: Abashiri Bay; small habitats in a large area ; seulement during the drift-ice period; severa! means of controlling larval dispersal.
estuaries.
341
Wilson, 1986
Giangrande and Petraroli, 1991
Gibbs, 1968
Nyholm, 1950
Giangrande and Fraschetti, 1993
Daly, 1978 Knight-Jones and
Knight-Jones, 1977; Doyle, 1974
Dimberger, 1993
Bailey-Brock, 1984
Y okoyama, 1995
Levin, 1984.
Sordino et al., 1989
Parker and Tunnicliffe, 1994
present authors
Sato-Okoshi, 1994
Mettam, 1980

M. BHAUD, J.-C. DUCHÊNE
The sampling area suggests that this species is present at a depth of more than 1500-2000 rn and lives on more or less homogeneous bottoms. At settling, the morphological organization of the larvae with more than fifty setigerous segments is very elaborate; this permits the use of a wide variety of sediment particles; consequently, the larvae can dispose of a large area for settling.
DISCUSSION
The direction of evolution and its persistence
According to prevailing opinion, the benthoplanktonic developmental pattern is probably the ancestral condition from which a developmental pattern emerged that is characterized by reduced dispersal ability and limited fecundity. lndeed, Agassiz (1874, p. 11) observed that "the higher we rise in the scale of animallife the more restricted is the number of offspring". Jagersten (1972) presented several arguments in favour of this opinion, among which is that the benthoplanktonic iife cycle is spread over the whole phylogenetic tree of the metazoans. It is not the high number of species with planktonic development which forms the centre of the argument; there is no link between the frequency of a situation expressed by the number of species and its primary or secondary position on a genealogie axis. Rather, the representation of the benthoplanktonic pattern in nearly ali phyletic lines forms the basis of the argument. Moreover, in several groups with a holobenthic life cycle, larval development reveals traces of features from the primary larva which are adaptations to pelagie life. We wish to add two arguments with regard to this evolutionary trend (cf. Mc Edward and Janies, 1993, for echinoderms).
a) In Polychaetes, benthic development uses a large number of solutions (Wilson, 1991; Duchêne, 1992; Bhaud, 1994; Premoli and Sella, 1995) such as: gelatinous egg masses outside the tube of the female, associated or not with this tube; brooding in the maternai tube, inside egg masses or capsules, or free; conservation of the initial number of eggs in each capsule, or decrease of this number by adelphophagy; incubation in dorsal brood pouches; larvae not incubated in an individualized structure, but kept in the tube by adult tentacles; viviparity with fracture of the tegument. Accordingly, the large spectrum of solutions adopted by direct development can only be derived and leads us to propose an evolutionary direction from benthoplanktonic to benthic development.
b) From the precise situation fumished by the two strategies found in E. nebulosa it is possible to argue that the protection of larvae by mucilaginous masses in the Mediterranean Sea is secondary relative to free development. It has been proposed (Lenaers and Bhaud, 1992) that during the last lee Age, relict populations survived during a period of generally low sea temperatures in the Mediterranean Sea. During subsequent global warming, E. nebulosa began a graduai colonization of Atlantic waters with development of the free larval pattern. However, three reasons prevent us from accepting this interpretation: i) populations from low latitudes are not affected by ice-age fluctuations (Scavotto, 1986; Ruddiman, 1990); ii) tropical populations
342
of E. nebulosa do not produce brooding egg-masses (persona! observations of J. D. Delgado, La Laguna, Canarias); and iii) the large geographical distribution area in the Atlantic, Pacifie and Indian Oceans (Lang, 1986) in which mucilaginous masses have never been observed. It is more than likely that the Mediterranean pattern with control of larval dispersal is secondary. This pattern evolved during colonization following the salinity crisis of the Messinian (6.5-5.3 MY BP) (Hsü, 1972; Busson, 1990)
To illustrate the persistence or interruption of this direction, the Terebellidae are used once again. This family is an example which displays a secondary planktonic life and shows a remarkable heterogeneity in terms of developmental strategies and larval ecology. From this basis it is possible to separate species belonging to the genera Lanice and Loimia from ail other species (Marcano and Bhaud, 1995). Representatives of these two genera exhibit a long planktonic larvallife related to the presence of a thick-walled slime-tube (Fig. 2). This aulophore stage, probably a recent acquisition in terms of life style (the aulophore morphology being "neutra]" in this respect), corresponds to a relatively late morphological stage which is benthic in other species of the family. This new feeding planktonic stage is not a retum to the first pattern of the trochophoremetatrochophore series, but represents a new developmental type with the addition of a planktonic phase during the postlarval or juvenile stage. However, with regard to the
B
6
7 1
Figure 2
Diagram of the different events in development represented in the Terebellidae. Terms from Fauchald 1983. 1: spawning and fertilization; 2: demersal planktonic phase or 2': development in retention structures; 3: first contact with the sediment, or release from the tube or from egg-masses; 4: secretion of a mucous tube including detritic material; 5: start of the second planktonic phase in a transparent mucous tube; 6: different aulophore stages with a post-larval morpho/ogy; 7: seulement and definitive benthic establishment; 8: juvenile growth; 9: gamete maturation; 4': equivalent of 8 + 9 in holobenthic cycles. Modifiedfrom Marcano and Bhaud 1995. A: common part for ali Terebellidae; B: added part in species with aulophore larvae. Example of a holobenthic cycle: Neoleprea streptochaeta (Duchêne, 1982) or Ramex califomiensis (Blake, 1991 ); example of a mixed cycle: Eupolymnia nebulosa (Bhaud et al., 1995), Thelepus extensus (Duchêne, 1983 ); example of a /ife cycle with planktonic /ife at a post-larval stage: the species Lanice conchilega (Bhaud, 1988 b) and probably also other species of this genus and species of the genus Loimia.

dispersal process, there is a return to an ancestral phase. Accordingly, there is no definitive establishment of benthic development. If the probability of an evolution from benthopelagic development to direct development is high, taking into account the diversity of solutions in direct development, retum to benthopelagic development is also observed, as in the case of Terebellidae. In either case, changes in physical conditions may break the life cycle and this disequilibrium may occur in severa! phases of the life history of a family. Movement back and forth from one type of life cycle to another may be possible, as is observed in Terebellidae. To explain the secondary acquisition of a dispersal stage by severa! species of this family, we do not have a definitive theory. However, we suggest that the extension of the planktonic phase acts to postpone and to decrease seulement constraints which are exerted during benthic seulement. If there is a decrease of seulement constraints with age, as has been demonstrated for E. ne bulosa (Cha, 1994), seulement at a late developmental stage allows an increase of the area compatible with the presence of the species. In such a context, it is not surprising that in the Bay of Banyuls Lanice conchilega which postpone planktonic life during juvenile development are collected in four communities out of a total of seven. This is not the first example of a secondary evolution. Westheide ( 1982) described a new species of Hesionidae which he argued convincingly to be an example of evolution leading from the interstitial fauna, generally assumed to have originated from larger forms living in other marine biotopes (Westheide, 1987), to the macrofaunal mode of life.
Energy available and pattern of development
It has been proposed (Chia, 1976) that the main selective force for shifting from feeding to non-feeding larvae (or from benthoplanktonic to holobenthic) is the limitation of energy for gamete production. The major part of energy expenditure of an individual is threefold: maintenance (respiration); growth; and gamete production. This third activity is the Jast to be provided with energy and it may therefore be limited. The animal bas a choice, evo1utionarily speaking, in gamete production: it may produce numerous small eggs or a small number of large eggs. This hypothesis states that the shifting from planktotrophic to lecithotrophic development is due to the limitation of available energy for gamete production. The condition for this shift is that the energy for gamete production is at a minimum and, in order to maintain a steady population state, the reproductive strategy shifts to a more efficient way of reproduction in terms of per unit energy cost, but incurs the penalty of limited dispersal and mutation. For Chia (1976), this means that in the long run, lecithotrophic development is an inferior method of reproduction when compared with planktotrophy. The question calls for three comments.
a) The choice, in terms of evolution, is not as straightforward as is supposed. The choice of direction must originate from something; that means evolutionary trends towards benthoplanktonic or holobenthic development are not equally probable. Basically, the pattern of development probably does not depend on the energy expenditure for gamete production at the levet of the organism as a who le,
343
LI FE CYCLE EVOLUTION IN POL YCHAETES
but on the constraints of packaging energy in a variable number of eggs.
b) Among Polychaeta, Bridges (1993) showed from measuring reproductive investment in four developmental morphs of Streblospio, that lecithotrophic development involved twice the C and N investment in reproduction as in the planktotrophic development.
c) Efforts to determine which reproductive pattern, planktotrophy or lecithotrophy, uses more energy have yielded equivocal results (Grahame and Branch, 1985; Strathmann, 1985). Grahame and Branch (1985) reported an equa1 number of works in both directions and concluded that there is as yet no consensus as to whether lecithotrophy or planktotrophy require more energy from the parent. Grahame ( 1982) from his work on Lacuna suggests that there may be very little difference, and Grant ( 1990) raises the question of whether there is a Iink between mode of development and reproductive effort. In short, these works suggest that there may be little overall difference in energy allocation between the two different modes of reproduction (see also Reznick, 1992).
CONCLUSION
The constraint of dispersal is linked to the necessity of returning to a suitable substratum for adult life. Accordingly, a connection must exist between habitat area and the duration of the planktonic larval stage. From the example of E. nebulosa, the length of the planktonic larval li fe and, therefore, the ability for dispersal, seem correlated with the locally available area for the adults. To attempt a generalization from this situation requires us to query, not the absolute value of the length of the planktonic life, but the meaning of this length integrated with ali the elements forming the life cycle. Against such a background a paradoxical result emerges. A species with a reduced distributional area in terms of adult habitat and with a limited ability to disseminate does not act differently from another species with larvae which wander for severa! weeks in the plankton without really leaving the adult habitat. lt is a mistake to argue from absolute values: number of days of larval life, the supposed travelled distance, the mean current speed, the use of a residual vector without taking the variability into account.
However the relation between habitat area and the length of life of planktonic larvae is confused by the adaptation of larvae to the hydrodynamic structures of water masses (Boicourt, 1982; Christy and Stancyk, 1982; Rothschild, 1988). Through these adaptations, control of dispersal is carried out without any relation to the size of the adult habitat. The observation of a short larval life leads us to suppose the probability of a control of dispersal and a reduced area of adult habitat. We cannot however, infer the reverse: a long larval !ife may be coupled with a large adult area as well as a reduced adult area. The first situation allows the reception of a sufficient number of larvae probably at the priee of large Joss by passing over the adult area. The second situation occurs only when the physical structure taken by larvae is developed on a large spatial scale (Rothlisberg et al., 1983) and when retention process

M. BHAUD, J.-C. DUCHËNE
bas been selected for, by means of larval behaviour. The fundamental necessity in a life cycle is to secure mating; for that, in gonochoric species, the important point is not to reduce the ability to disseminate but to find the adult habitat or not to leave it. This goal is achieved by different strategies such as reduction of dispersal period and development of elaborate larval behaviour. Exploration of the polychaete life cycle leads to the identification of two sources for selection: one related to larval behaviour with control of spatial dispersal, the other related to reduction or suppression of dispersal. In both cases, larvae resist being swept along; even in the case of a long course, they are able by their behaviour, to control advective strength.
These conclusions are partly speculative and are based upon a relatively reduced amount of data, at least with regard to polychaetes. In this class, development and auto-
REFERENCES
Agassiz L. (1874). Evolution and permanence of type. Atlantic Monthly 33, 3-12.
Arnoux, S., M. Bhaud, C. Cazaux, J.H. Cha, J.C. Duchêne, P. Fraunié, G. Marcano, D. Martin, M.J. Nattero, C. Nozais, C. Rey, S. Pinedo, R. Sarda (1995). Recruitment in the light of biologicalphysical interactions in coastal waters: results of PNDR action at the Arago Laboratory in 1994-1995. Vie Milieu 45, 2, 85-105.
Alldredge A.L., W.M. Hamner (1980). Recurring aggregation of zoop1akton by tidal current. Estuar. coast. mar. Sei. 10,31-37.
Bai1ey-Brock J.H. (1984). Spawning and development of Arenicola brasiliensis (Nonato) in Hawaii (Polychaeta; Arenicolidae). Proceedings of the First International Polychaete Conference, Sydney, P.A. Hutchings, ed. publ. by The Linnean Society of New South Wales. 439-449.
Banse K. (1986). Vertical distribution and horizontal transport of planktonic larvae of echinoderms and benthic polychaetes in an open coastal sea. Bull. marine Sc. 39. 162-175.
Bhaud M. ( 1988 a). Experimental studies of influence of temperature and food available on development rate of Eupolymnia nebulosa (Polychaete, Terebellidae ). J. Exp. mar. Biol. Eco!. 118, 103-113.
Bhaud M. (1988 b). The two planktonic Iarval periods of Lanice conchilega: a peculiar example of the irreversibility of Evolution. Ophelia 29, 141-152.
Bhaud M. ( 1990). Acquisition de la vic benthique par Eupolymnia nebulosa (Polychète Terebellidae): dispositifs expérimentaux et premiers résultats. Vie Milieu. 40, 1, 17-28.
Bhaud M. (1991). Larval release from egg mass and seulement of Eupolymnia nebulosa (Polychaeta, Terebellidae). Bull. mar. Sc. 48, 2, 420-431.
Bhaud M. (1994). Les contraintes dans la réalisation des cycles de développement en mer. Bull. Soc. zoo!. Fr. 119, 3, 243-262.
Bhaud M., C. Cazaux (1987). Description and identification of polychaete larvae: their implications in current biological problems. Oceanis 13, 596-753.
Bhaud M., J.C. Duchêne ( 1988). Biologie larvaire et stratégie de reproduction des Annélides Polychètes en province subantarctique. ln : Actes du Colloque sur la Recherche Française dans les Terres Australes: Strasbourg 1987, C.N.F.R.A. Paris. 145-152.
Bhaud M., A. Grémare (1988). Larval development of the Terebellid Polychaeta Eupolymnia nebulosa. Zoo!. scripta. 17, 4, 347-356.
Bhaud M., A. Grémare (1991). Reproductive cycle of Eupolymnia nebulosa (Polychaete Terebellidae) in the Mediterranean Sea. Ophelia. Suppl. 5, 295-304.
344
noroy of larvae, implication of advection in the dispersal, are neglected fields of investigation. The interpretation of the selective advantages of the wide range of life cycles of polychaetes is, however, consistent with the literature on other marine forms. Excessive dispersal in an oceanic environment is a constraint to the persistence of benthic populations with specifie adult substratum requirements.
Acknowledgments
This work is a contribution to the French National Programme on Determinants of Recruitment (PNDR/FranceGLOBEC) and was financed by grants to the authors from CNRS and IFREMER.
Bhaud M., J.H. Cha (1994). Larvae-substrate relationships of Eupolymnia nebulosa (Montagu, 1818) (Polychaeta, Terehellidae): an experimental analysis. Mém. Mus. nat. Hist. Nat.(A) 162, 371-381.
Bhaud M., A. Grémare, F. Lang, C. Rétière ( 1987). Comparaison des caractères reproductifs du Terebellien Eupolymnia nebulosa (Montagu) en deux points de son aire géographique. C. R. Acad. Sei. Paris, série D. 119-122.
Bhaud M., J.H. Cha, J.C. Duchêne, D. Martin, C. Nozais (1995). Larval biology and benthic recruitment: new prospects on the role of egg masses and proposai on regulation mode! of the !ife cycle. Sei. Mar. 59, 103-117.
Blake J.A. (1991). Larval developmcnt of polychaeta from the northem California coast v. Ramex californiensis Hartman (Polychaeta: Terebellidae). Bull. Mar. Sc. 48, 2. 448-460.
Boicourt W.C. ( 1982). Estuarine larval retention mechanisms on two scales. ln: Estuarine Comparisons, V.S. Kennedy eds. Academie Press, New York, 445-456.
Bridges T.S. (1993). Reproductive investment in four developmental morphs of Streblospio (Polychaeta: Spionidae). Biol. Bull. 184, 144-152.
Busson G. ( 1990). Le Messinien de la Méditerranée vingt ans après. Bull. Soc. Géol. Fr. 3-4, 3-57.
Caron A., M. Olivier, G. Derosiers, E. Hudier, S. Côté, V. Koutitonsky, G. Miron, C. Rétière (1993). Distribution spatiale d'une espèce benthique épitoque en zone intcrtidale: rôle de l'hydrodynamisme? Vie Milieu 43, 85-93.
Cazaux C. (1981). Évolution et adaptation larvaires chez les Polychètes. Océanis 1, 43-77.
Cazaux C. (1982). Développement larvaire de l' Ampharetidae lagunaire Alkmaria romijni Horst 1919. Cah. Biol. Mar. 23, 143-158.
Cazaux C. ( 1985). Reproduction et développement larvaire de l' Annélide Polychète saumâtre Streblospio shrubsolii (Buchanan 1890). Cah. Biol. Mar. 26, 207-211.
Cha J.H. (1994). Analyse expérimentale du recrutement benthique chez Eupolymnia nebulosa (Annélide Polychète Terebellidae) en Méditerranée occidentale. Thèse de Doctorat, Université P. et M. Curie, 268 p.
Cha J.H., M. Bhaud (1991 ). Étude expérimentale du recrutement benthique en canal hydrodynamique et en milieu calme. C. R. Acad. Sei. 313, 3, 113-118.
Chapman G. ( 1965). The egg cocoons of Scoloplos armiger O.F. Müller. Biol. Bull. 128, 189-197.

Chia F.S. ( 1976). Classification and adaptive significance of developmental patterns in marine invertebrates. Thalassia Jugoslavica 10, 121-130.
Christiansen F.B., T.M. Fenchel ( 1979). Evolution of marine invertebrate reproductive patterns. Theor. Popul. Biol. 16, 267-282.
Christy J.H., S.E. Stancyk ( 1982). Timing of larval production and flux of invertebrate larvae in a well-mixed estuary. In : Estuarine Comparisons, V.S. Kennedy eds. Academie Press, New York, 489-503.
Daly J.M. (1978). The annual cycle and the short term periodicity of breeding in a Northumberland population of Spirorbis spirorbis (Polychaeta: Serpulidae). J. mar. biol. Ass. U.K. 58, 161-176.
Doyle R.W. (1974). Choosing between darkness and light: the ecological genetics of photic behavior in planktonic larva of Spirorbis borealis. Mar. Biol. 25,311-317.
Desrosiers G., M. Olivier, B. Vincent (1991). Variation de la densité et de la croissance des recrues de l'annélide polychète Nereis virens (Sars) en zone intertidale. Can.J. Zoo!. 66, 1454-1459.
Dirnberger J.M. ( 1993). Dispersal of larvae with a short planktonic phase in the polychaete Spirorbis spirillum (Linnaeus). Bull. Mar. Sc. 52, 3, 898-910.
Duchêne J.C. ( 1982). Étude comparée de la biologie d'Annélides Polychètes considérées en différentes zones climatiques. Thèse de Doctorat d'État, Océanographie biol., Univ. P. et M. Curie (Paris VI) 315 p.
Duchêne J.C. (1983). Développement larvaire et fixation chez The lepus setosus (Annélide Polychète) à Kerguelen, Province subantarctique. Vie Milieu 33, 2, 65-77.
Duchêne J.C. (1984). Reproductive biology of Boccardia polybranchia (Carazzi) in Kerguelen (Subantarctic province). Polar Biol. 2, 251-257.
Duchêne J.C. (1989). Kerguelen: recherches au bout du monde. Territoire des Terres Australes et Antarctiques Françaises: Mission de Recherche, Paris, l-360.
Duchêne J.C. (1992). Adelphophagie et biologie larvaire chez Boecardia polybranchia (Carazzi) (Annélide Polychète Spionidae) en province subantarctique. Vie Milieu 39, 143-152.
Eckelbarger K.J. (1974). Population biology and larval development of the terebellid polychaete Nicolea zostericola. Mar. Biol. 27, 101-113.
Fauchald K. (1983). Life diagram patterns in benthic polychaetes. Proceedings of the Biological Society of Washington, 96, 160-177.
Fonseca-Genevois V., C. Cazaux (1987). Streblospio benedicti Webster, 1879 (Annélide Polychète) dans l'estuaire de la Loire: biologie et écologie. Cah. Biol. Mar. 28, 231-261.
Gamberoni L., J. Geronimi, P.F. Jeannin, J.F. Murait (1982). Study of the frontal zones in the Crozet-Kerguelen region. Oceanologica Acta 5, 288-299.
Giangrande A., S. Geraci, G. Belmonte (1994). Life-cycle and lifehistory diversity in marine invertebrates and implications in community dynamics. Oceanography and Marine Bio/ogy: an Annual Review 32, 305-333.
Giangrande A., A. Petraroli ( 1991 ). Reproduction, larval development and post larval growth of Naineris laevigata (polychaeta, Orbiniidae) in the Mediterranean Sea. Mar. Biol.1ll, 129-137.
Giangrande A., S. Fraschetti (1993). Life cycle, growth and secondary production in a brackish-water population of the polychaete Notomastus latericeus (Capitellidae) in the Mediterranean Sea. P.S.Z.N.: Marine Ecology 14, 313-327.
Gibbs P.E. (1968). Observations on the population of Scoloplos armiger at Whitstable. J. mar. biol. Ass. U.K. 48, 225-254.
Grahame J. (1982). Energy flow and breeding in two species of Lacuna: comparative costs of egg production and maintenance. /nt. J. lnvert. Reprod. 5, 91-99.
Grahame J,, G.M. Branch ( 1985). Reproductive patterns of marine invertebrates. Oceanogr. Mar. Biol. Ann. Rev. 23, 373-398.
345
LI FE CYCLE EVOLUTION IN POL YCHAETES
Grant A. (1990). Mode of development and reproductive effort in marine invertebrates: should there be any relationship? Functional Eco/ogy 4, l, 128-130.
Guerin J.P. (1974). Rôle du mucus entourant la ponte dans la survie des embryons et la vitesse de croissance des larves de Scolelepis fuliginosa Claparède (Annélide Polychète). C.R. Acad. Sc. Paris, 279, D, 363-366.
Guerin J.P. (1975). Redescription des adultes et comparaison des divers stades ontogénétiques des populations méditerranéennes et atlantiques de Scolelepis fuliginosa Claparède (Annélide Polychète). Cah. Biol. Mar. 16,21-37.
Hannerz L. ( 1956). Larval development of polychaete families Spionidae Sars, Disomidae Mesnil and Poecilochaetidae n. fam. in the Gullmar Fjord, Sweden. Zoo!. Bidr. 31, l-204.
Harris T. (1994). Localisation of Ophelia bicomis Savigny in nonestuarine sandy shores in Devon, UK. Polychaete Research 16,41-43.
Hoagland K.E., R. Robertson ( 1988). An assessment of poecilogony in marine invertebrates: phenomenon or fantasy? Biol. Bull. 174, 109-125.
Hsü K.L. (1972). When the mediterranean dried up. Scient. Am. 227, 27-36.
Jablonski D., R.A. Lutz (1983). Larval ecology of marine benthic invertebrates: paleobiological implications. Biol. Rev. 58,21-89.
Jiigersten G. (1972). Evolution of the metazoan life cycle. A comprehensive theory. Academie Press, New York, l-282.
Kingsford M.J. (1990). Linear oceanographie features: a focus for research on recruitment processes. Austral. Eco!. 15, 391-40 l.
Knight-Jones P., E.W. Knight-Jones (1977). Taxonomy and ecology of British Spirorbidae (Polychaeta). J. mar. biol. Ass. U.K. 51, 453-499.
Lang F. (1986). Peuplement des fonds durs du bassin maritime de la Rance; rôle fonctionnel de Eupolymnia nebulosa (Annélide Polychète). Thèse de doctorat, Université de Rennes, 240 p.
Lardicci C. (1992). General characteristics of recruitment and recolonization in Polychaeta assemblage in a small brackish basin on island ofElba (Western Mediterranean). Oebalia 18,69-81.
Lefevre M. (1990). Interférence sur le recrutement benthique entre hydrodynamisme des masses d'eau et comportement larvaire. Oceanis 16, 135-148.
Lenaers G., M. Bhaud (1992). Molecular phylogeny of sorne polychaete annelids: an initial approach to the Atlantic-Mediterranean speciation problem. J. Mol. Evol. 35, 429-435.
Levin L.A. (1984 ). Li fe his tory and dispersal patterns in a dense infaunal polychaete assemblage: community structure and response to disturbance. Eco/ogy 65, 1185-1200.
McHugh D., V. TunniclitTe (1994). Ecology and reproductive biology of the hydrothermal vent polychaete Amphisamytha galapagensis (Ampharetidae).Mar. Eco!. Progr. Ser.106, 1-2,111-120.
Mann K.H., J.R.N. Lazier (1991). Dynamics of marine ecosystems: Biological-Physical Interactions in the Oceans. Blackwell Scientific Publications, Boston, 466 p.
Marcano G., M. Bhaud (1995). New observations on the terebellid (Polychaeta) aulophore larvae on the French coasts. Ophelia 43, 3, 128-135.
McEdward L.R., D.A. Janies ( 1993). Life cycle evolution in Asteroids: what is a larva? Biol. Bull. 184, 255-268.
Medernach L. (1995). Rôle du cocon muqueux dans la protection des larves de Eupolymnia nebulosa (Annélide Polychète). Rapport de Maîtrise (Ecologie Marine) Laboratoire Arago, 34 p.
Menge B.A. ( 1975). Brood or broadcast? The adaptive significance of different reproductive strategies in two intertidal sea stars Leptasterias hexactis and Pisaster ochraceus. Mar. Biol. 31, 87-100.
Mettam C. (1980). Survival strategies in estuarine nereids. In Feeding and survival strategies of estuarine organisms. N.V. Jones and J. Wolff, eds. New York: Plenum Press, 65-77.

M. BHAUD, J.-C. DUCHËNE
Mileikovsky S.A. (1971). Types of larval development in marine bottom invertebrates, their distribution and ecological significance: a re-evaluation. Mar. Biol. 10, 193-213.
Mi1eikovsky S.A. (1976). Types of larval development in marine bottom invertebrates: an integrated ecological scheme. Thalassia Jugoslavica 10, 171-179.
Nyho1m K.G. ( 1950). Contributions to the life-history of the Ampharetid, Melinna cristata. Zoo/. Bidrag. 29, 79-91.
O'Brien D.P. (1988). Direct observations of clustering (schooling and swarming) behavior in mysids (Crustacea: Mysidacea). Mar. Ecot. Prog. Ser. 42, 235-246.
Palmer A.R., R.R. Strathmann ( 1981 ). Sc ale of dispersal in varying environments and its implication for life histories of marine invertebrates. Oecolologia 48, 308-318.
Parker T., V. Tunnicliffe (1994). Dispersal strategies of biota on an oceanic seamount: implications for ecology and biogeography. Biol. Bull. 187, 336-345.
Parry G.D. (1981). The meanings of r-and K-selection. Oecologia 48, 260-264.
Pechenik J.A. ( 1979). Rote of encapsulation in invertebrate life histories. Am. Nat. 114, 859-870.
Pineda J. (1991). Predictable upwelling and the shoreward transport of planktonic 1arvae by internai tidal bores. Science 253, 548-551.
Plate S., E. Husemann (1991). An alternative mode of larval development in Scoloplos armiger (O.F. Müller, 1776) (Polychaeta, Orbiniidae). Helgol. Meeresunter. 45,487-492.
Premoli M.C., G. Sella ( 1995). Sex economy in benthic polychaetes. Ethol. Ecot. Evol. 1, 27-48.
Radashevsky V.I. (1994). Life history of a new Polydora species from the Kurile Islands and evolution of lecithotrophy in polydorid genera (Polychaeta: Spionidae). Ophelia 39, 2, 121-136.
Reznick D. (1992). Measuring the costs of reproduction. TREE 1, 2. 42-45.
Rothlisberg P.C., J.A. Church, A.M.G. Forbes (1983). Modelling the advection of vertically migrating shrimp larvae. J. Mar. Res. 41, 511-538.
Rothschild B.J. ( 1988). Dynamics of the sea: the Ecology of High Dimensionality Systems. ln: Toward a Theory on Biologicai-Physical Interactions in the World Ocean. B.J. Rothschild ed. Kluwer Academie Publishers, Dordrecht, Boston, 527-548.
Roughgarden J. (1989). The evolution of marine life cycles. P 270-300 in Mathematical Evolutionary Theory, M. Feldman, ed. Princeton University Press, Princeton, New Jersey.
Ruddiman W.F. (1990). Changes in climate and biota on geologie time scales. Trends Eco/. Evol. 5, 285-288.
Sato-Okoshi W. (1994). Life history of the Polychaete Polydora variegata that bores into the shells of Scallops in northern Japan. Mem. Mus. nat. Hist. nat. 162, 541-547.
Sato M., M. Tsuchiya (1991). Two patterns of early development in nereidid polychaetes keying out to Neanthes japon ica (lzuka). Ophelia suppl. 5, 371-382.
346
Scavotto E.D. (1986). The little ice age. Oceanus 29,38-39.
Sche1tema R.S. (1989). Planktonic and non-planktonic development among prosobranch gastropods and its relationship to the geographie range of species. ln: Reproduction, genetics and distribution of marine organisms, J.S. Ryland and P.A. Tyler eds. Olsen and Olsen, Fredensborg, 183-188.
Schroeder P.C., C.O. Hermans (1975). Annelida: Polychaeta. ln A. Giese and J. Pearse (eds): Reproduction of marine invertebrates. Vol. III. Annelids and Echiurans, 1-213. Academie Press, New York.
Sinclair M. ( 1988). Marine populations: an essay on population regulation and speciation. Washington Sea Grant Program, University of Washington Press, Seattle and London, 1-252.
Sordino P., M.C. Gambi, G.C. Carrada (1989). Spatio-temporal distribution of Polychaetes in an italian coastal lagoon (Lago Fusaro, Naples). Cah. Biol. Mar. 30, 375-391.
Strathmann R.R. (1982). Selection for retention or export of larvae in estuarines. ln: Estuarine Comparisons, V.S. Kennedy eds. Academie Press, New York, 521-536.
Strathmann R.R. (1985). Feeding and nonfeeding larval development and life-history evolution in marine invertebrates. Ann. Rev. Ecot. Syst. 16, 339-361.
Swedmark B. ( 1959). On the biol ogy of sexual reproduction of the interstitial fauna of marine sand. Proc. XV th Congr. intern. Zoo!., London, 327-330.
Vance R.R. (1973 a). On reproductive strategies in marine benthic invertebrates. Am. Nat. 107, 339-352.
Vance R.R. (1973 b). More on reproductive strategies in marine benthic invertebrates. Am. Nat. 107, 353-361.
Westheide W. (1982). Microphthalmus hamosus sp.n. (Polychaeta, Hesionidae). An example of evolution leading from the interstitial fauna to a macrofaunal interspecific relationship. Zoo/. Scripta 11, 189-193.
Westheide W. (1987). Progenesis as a principle in meiofauna evolution. J. nat. Hist. London 21, 843-854.
Wilson W.H. (1986). Detachment of egg masses of a polychaete: environmental risks of benthic protective development. Eco/ogy. 67, 3, 810-815.
Wilson W.H. (1991). Sexual reproductive modes in Polychaetes: classification and diversity. Bull. Mar. Sei. 48, 500-516.
Woodwick K.H. (1977). Lecithotrophic Jarval development in Boecardia proboscidea Hartman. ln: Essay on polychaetous annelids in memory of Dr. O. Hartman. A. Hancock Foundation, 347-371.
Yokoyama H. (1995). Occurrence of Paraprionospio sp. (forrn A) larvae (polychaeta: Spionidae) in hypoxic water of an enclosed bay. Est. Coast. Shelf Sc. 40, 9-19.
Zal F ., D. Jollivet, P. Chevaldonne, D. Desbruyères ( 1995). Reproductive strategy and population structure of the deep-sea hydrothermal vent worrn Paralvinella grasslei (Polychaeta: Alvinellidae) at 13° Non the East Pacifie Rise. Mar. Biol. 122, 4, 637-648.





























