Embed Size (px)
Citation preview

Cellular Reproduction
By:Jeffrey Mahr


Cellular Reproduction
By:Jeffrey Mahr
Online:< http://cnx.org/content/col11635/1.1/ >
C O N N E X I O N S
Rice University, Houston, Texas

This selection and arrangement of content as a collection is copyrighted by Je�rey Mahr. It is licensed under the
Creative Commons Attribution License 3.0 (http://creativecommons.org/licenses/by/3.0/).
Collection structure revised: February 26, 2014
PDF generated: February 26, 2014
For copyright and attribution information for the modules contained in this collection, see p. 40.

Table of Contents
1 Cellular Reproduction - Introduction . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 12 Cellular Reproduction - Genomic DNA . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 33 Cellular Reproduction: The Cell Cycle . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 94 Cellular Reproduction: Control of the Cell Cycle . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 195 Cellular Reproduction: Cancer and the Cell Cycle . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 276 Cellular Reproduction - Prokaryotic Cell Division . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 31Glossary . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 35Index . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 38Attributions . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 40

iv
Available for free at Connexions <http://cnx.org/content/col11635/1.1>

Chapter 1
Cellular Reproduction - Introduction1
Figure 1.1: A sea urchin begins life as a single cell that (a) divides to form two cells, visible by scanningelectron microscopy. After four rounds of cell division, (b) there are 16 cells, as seen in this SEM image.After many rounds of cell division, the individual develops into a complex, multicellular organism, asseen in this (c) mature sea urchin. (credit a: modi�cation of work by Evelyn Spiegel, Louisa Howard;credit b: modi�cation of work by Evelyn Spiegel, Louisa Howard; credit c: modi�cation of work by MarcoBusdraghi; scale-bar data from Matt Russell)
A human, as well as every sexually reproducing organism, begins life as a fertilized egg (embryo) or zygote.Trillions of cell divisions subsequently occur in a controlled manner to produce a complex, multicellularhuman. In other words, that original single cell is the ancestor of every other cell in the body. Once a beingis fully grown, cell reproduction is still necessary to repair or regenerate tissues. For example, new bloodand skin cells are constantly being produced. All multicellular organisms use cell division for growth and themaintenance and repair of cells and tissues. Cell division is tightly regulated, and the occasional failure ofregulation can have life-threatening consequences. Single-celled organisms use cell division as their methodof reproduction.
1This content is available online at <http://cnx.org/content/m48934/1.1/>.
Available for free at Connexions <http://cnx.org/content/col11635/1.1>
1

2 CHAPTER 1. CELLULAR REPRODUCTION - INTRODUCTION
Available for free at Connexions <http://cnx.org/content/col11635/1.1>

Chapter 2
Cellular Reproduction - Genomic DNA1
The continuity of life from one cell to another has its foundation in the reproduction of cells by way of thecell cycle. The cell cycle is an orderly sequence of events that describes the stages of a cell's life from thedivision of a single parent cell to the production of two new daughter cells. The mechanisms involved in thecell cycle are highly regulated.
2.1 Genomic DNA
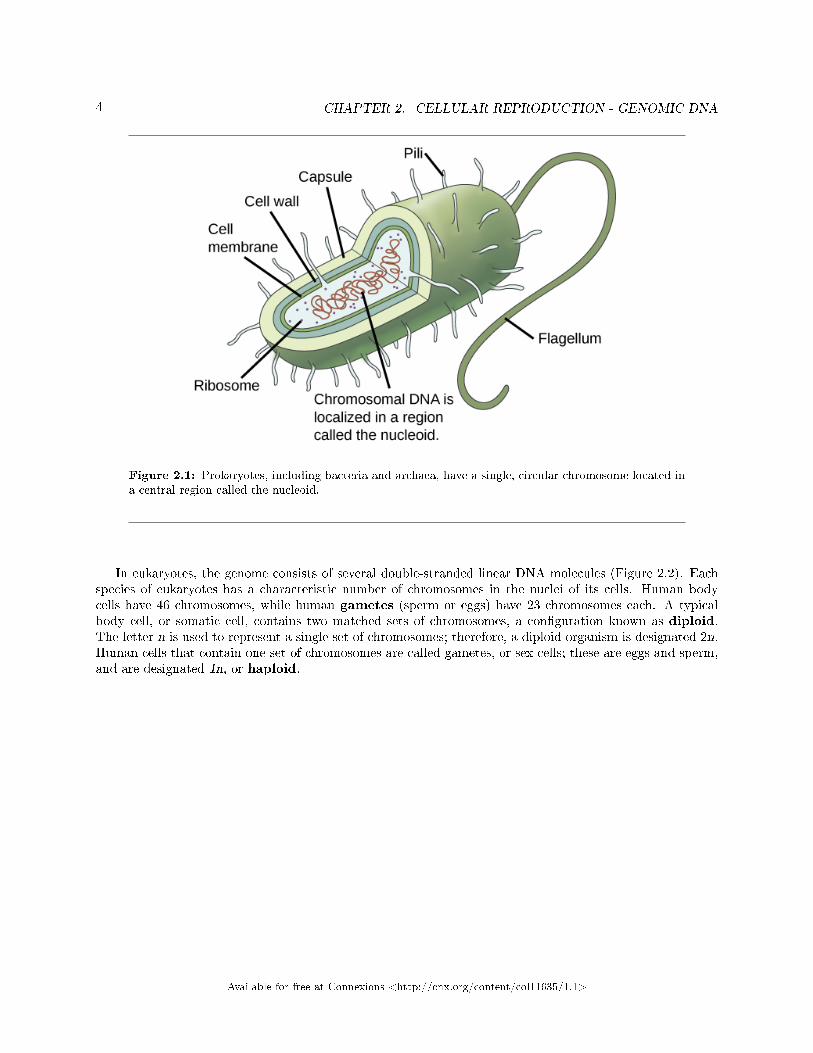
Before discussing the steps a cell must undertake to replicate, a deeper understanding of the structure andfunction of a cell's genetic information is necessary. A cell's DNA, packaged as a double-stranded DNAmolecule, is called its genome. In prokaryotes, the genome is composed of a single, double-stranded DNAmolecule in the form of a loop or circle (Figure 2.1). The region in the cell containing this genetic materialis called a nucleoid. Some prokaryotes also have smaller loops of DNA called plasmids that are not essentialfor normal growth. Bacteria can exchange these plasmids with other bacteria, sometimes receiving bene�cialnew genes that the recipient can add to their chromosomal DNA. Antibiotic resistance is one trait that oftenspreads through a bacterial colony through plasmid exchange.
1This content is available online at <http://cnx.org/content/m48936/1.1/>.
Available for free at Connexions <http://cnx.org/content/col11635/1.1>
3
4 CHAPTER 2. CELLULAR REPRODUCTION - GENOMIC DNA
Figure 2.1: Prokaryotes, including bacteria and archaea, have a single, circular chromosome located ina central region called the nucleoid.
In eukaryotes, the genome consists of several double-stranded linear DNA molecules (Figure 2.2). Eachspecies of eukaryotes has a characteristic number of chromosomes in the nuclei of its cells. Human bodycells have 46 chromosomes, while human gametes (sperm or eggs) have 23 chromosomes each. A typicalbody cell, or somatic cell, contains two matched sets of chromosomes, a con�guration known as diploid.The letter n is used to represent a single set of chromosomes; therefore, a diploid organism is designated 2n.Human cells that contain one set of chromosomes are called gametes, or sex cells; these are eggs and sperm,and are designated 1n, or haploid.
Available for free at Connexions <http://cnx.org/content/col11635/1.1>

5
Figure 2.2: There are 23 pairs of homologous chromosomes in a female human somatic cell. Thecondensed chromosomes are viewed within the nucleus (top), removed from a cell in mitosis and spreadout on a slide (right), and arti�cially arranged according to length (left); an arrangement like this iscalled a karyotype. In this image, the chromosomes were exposed to �uorescent stains for di�erentiationof the di�erent chromosomes. A method of staining called �chromosome painting� employs �uorescentdyes that highlight chromosomes in di�erent colors. (credit: National Human Genome Project/NIH)
Matched pairs of chromosomes in a diploid organism are called homologous (�same knowledge�) chro-mosomes. Homologous chromosomes are the same length and have speci�c nucleotide segments calledgenes in exactly the same location, or locus. Genes, the functional units of chromosomes, determine spe-ci�c characteristics by coding for speci�c proteins. Traits are the variations of those characteristics. Forexample, hair color is a characteristic with traits that are blonde, brown, or black.
Each copy of a homologous pair of chromosomes originates from a di�erent parent; therefore, the genesthemselves are not identical. The variation of individuals within a species is due to the speci�c combinationof the genes inherited from both parents. Even a slightly altered sequence of nucleotides within a gene canresult in an alternative trait. For example, there are three possible gene sequences on the human chromosomethat code for blood type: sequence A, sequence B, and sequence O. Because all diploid human cells havetwo copies of the chromosome that determines blood type, the blood type (the trait) is determined by whichtwo versions of the marker gene are inherited. It is possible to have two copies of the same gene sequence onboth homologous chromosomes, with one on each (for example, AA, BB, or OO), or two di�erent sequences,such as AB.
Minor variations of traits, such as blood type, eye color, and handedness, contribute to the naturalvariation found within a species. However, if the entire DNA sequence from any pair of human homologouschromosomes is compared, the di�erence is less than one percent. The sex chromosomes, X and Y, are thesingle exception to the rule of homologous chromosome uniformity: Other than a small amount of homologythat is necessary to accurately produce gametes, the genes found on the X and Y chromosomes are di�erent.
Available for free at Connexions <http://cnx.org/content/col11635/1.1>

6 CHAPTER 2. CELLULAR REPRODUCTION - GENOMIC DNA
2.2 Eukaryotic Chromosomal Structure and Compaction
If the DNA from all 46 chromosomes in a human cell nucleus was laid out end to end, it would measureapproximately two meters; however, its diameter would be only 2 nm. Considering that the size of a typicalhuman cell is about 10 µm (100,000 cells lined up to equal one meter), DNA must be tightly packaged to�t in the cell's nucleus. At the same time, it must also be readily accessible for the genes to be expressed.During some stages of the cell cycle, the long strands of DNA are condensed into compact chromosomes.There are a number of ways that chromosomes are compacted.
In the �rst level of compaction, short stretches of the DNA double helix wrap around a core of eighthistone proteins at regular intervals along the entire length of the chromosome (Figure 2.3). The DNA-histone complex is called chromatin. The beadlike, histone DNA complex is called a nucleosome, and DNAconnecting the nucleosomes is called linker DNA. A DNA molecule in this form is about seven times shorterthan the double helix without the histones, and the beads are about 10 nm in diameter, in contrast with the2-nm diameter of a DNA double helix. The next level of compaction occurs as the nucleosomes and the linkerDNA between them are coiled into a 30-nm chromatin �ber. This coiling further shortens the chromosomeso that it is now about 50 times shorter than the extended form. In the third level of packing, a variety of�brous proteins is used to pack the chromatin. These �brous proteins also ensure that each chromosome ina non-dividing cell occupies a particular area of the nucleus that does not overlap with that of any otherchromosome (see the top image in Figure 2.2).
Available for free at Connexions <http://cnx.org/content/col11635/1.1>

7
Figure 2.3: Double-stranded DNA wraps around histone proteins to form nucleosomes that have theappearance of �beads on a string.� The nucleosomes are coiled into a 30-nm chromatin �ber. When acell undergoes mitosis, the chromosomes condense even further.
Available for free at Connexions <http://cnx.org/content/col11635/1.1>

8 CHAPTER 2. CELLULAR REPRODUCTION - GENOMIC DNA
DNA replicates in the S phase of interphase. After replication, the chromosomes are composed of twolinked sister chromatids. When fully compact, the pairs of identically packed chromosomes are bound toeach other by cohesin proteins. The connection between the sister chromatids is closest in a region calledthe centromere. The conjoined sister chromatids, with a diameter of about 1 µm, are visible under a lightmicroscope. The centromeric region is highly condensed and thus will appear as a constricted area.
: This animation2 illustrates the di�erent levels of chromosomepacking.
2.3 Section Summary
Prokaryotes have a single circular chromosome composed of double-stranded DNA, whereas eukaryotes havemultiple, linear chromosomes composed of chromatin surrounded by a nuclear membrane. The 46 chro-mosomes of human somatic cells are composed of 22 pairs of autosomes (matched pairs) and a pair of sexchromosomes, which may or may not be matched. This is the 2n or diploid state. Human gametes have 23chromosomes or one complete set of chromosomes; a set of chromosomes is complete with either one of thesex chromosomes. This is the n or haploid state. Genes are segments of DNA that code for a speci�c pro-tein. An organism's traits are determined by the genes inherited from each parent. Duplicated chromosomesare composed of two sister chromatids. Chromosomes are compacted using a variety of mechanisms duringcertain stages of the cell cycle. Several classes of protein are involved in the organization and packing of thechromosomal DNA into a highly condensed structure. The condensing complex compacts chromosomes, andthe resulting condensed structure is necessary for chromosomal segregation during mitosis.
2http://openstaxcollege.org/l/Packaged_DNA
Available for free at Connexions <http://cnx.org/content/col11635/1.1>

Chapter 3
Cellular Reproduction: The Cell Cycle1
The cell cycle is an ordered series of events involving cell growth and cell division that produces two newdaughter cells. Cells on the path to cell division proceed through a series of precisely timed and carefullyregulated stages of growth, DNA replication, and division that produces two identical (clone) cells. The cellcycle has two major phases: interphase and the mitotic phase (Figure 3.1). During interphase, the cellgrows and DNA is replicated. During the mitotic phase, the replicated DNA and cytoplasmic contents areseparated, and the cell divides.
1This content is available online at <http://cnx.org/content/m48937/1.1/>.
Available for free at Connexions <http://cnx.org/content/col11635/1.1>
9

10 CHAPTER 3. CELLULAR REPRODUCTION: THE CELL CYCLE
Figure 3.1: The cell cycle consists of interphase and the mitotic phase. During interphase, the cellgrows and the nuclear DNA is duplicated. Interphase is followed by the mitotic phase. During themitotic phase, the duplicated chromosomes are segregated and distributed into daughter nuclei. Thecytoplasm is usually divided as well, resulting in two daughter cells.
3.1 Interphase
During interphase, the cell undergoes normal growth processes while also preparing for cell division. In orderfor a cell to move from interphase into the mitotic phase, many internal and external conditions must bemet. The three stages of interphase are called G1, S, and G2.
3.1.1 G1 Phase (First Gap)
The �rst stage of interphase is called the G1 phase (�rst gap) because, from a microscopic aspect, littlechange is visible. However, during the G1 stage, the cell is quite active at the biochemical level. The cell isaccumulating the building blocks of chromosomal DNA and the associated proteins as well as accumulatingsu�cient energy reserves to complete the task of replicating each chromosome in the nucleus.
Available for free at Connexions <http://cnx.org/content/col11635/1.1>

11
3.1.2 S Phase (Synthesis of DNA)
Throughout interphase, nuclear DNA remains in a semi-condensed chromatin con�guration. In the S phase,DNA replication can proceed through the mechanisms that result in the formation of identical pairs ofDNA molecules�sister chromatids�that are �rmly attached to the centromeric region. The centrosome isduplicated during the S phase. The two centrosomes will give rise to the mitotic spindle, the apparatusthat orchestrates the movement of chromosomes during mitosis. At the center of each animal cell, thecentrosomes of animal cells are associated with a pair of rod-like objects, the centrioles, which are at rightangles to each other. Centrioles help organize cell division. Centrioles are not present in the centrosomes ofother eukaryotic species, such as plants and most fungi.
3.1.3 G2 Phase (Second Gap)
In the G2 phase, the cell replenishes its energy stores and synthesizes proteins necessary for chromosomemanipulation. Some cell organelles are duplicated, and the cytoskeleton is dismantled to provide resourcesfor the mitotic phase. There may be additional cell growth during G2. The �nal preparations for the mitoticphase must be completed before the cell is able to enter the �rst stage of mitosis.
3.2 The Mitotic Phase
The mitotic phase is a multistep process during which the duplicated chromosomes are aligned, separated,and move into two new, identical daughter cells. The �rst portion of the mitotic phase is called karyokinesis,or nuclear division. The second portion of the mitotic phase, called cytokinesis, is the physical separation ofthe cytoplasmic components into the two daughter cells.
: Revisit the stages of mitosis at this site2 .
3.2.1 Karyokinesis (Mitosis)
Karyokinesis, also known asmitosis, is divided into a series of phases�prophase, prometaphase, metaphase,anaphase, and telophase�that result in the division of the cell nucleus (Figure 3.2). Karyokinesis is alsocalled mitosis.
:
2http://openstaxcollege.org/l/Cell_cycle_mito
Available for free at Connexions <http://cnx.org/content/col11635/1.1>

12 CHAPTER 3. CELLULAR REPRODUCTION: THE CELL CYCLE
Figure 3.2: Karyokinesis (or mitosis) is divided into �ve stages�prophase, prometaphase, metaphase,anaphase, and telophase. The pictures at the bottom were taken by �uorescence microscopy (hence, theblack background) of cells arti�cially stained by �uorescent dyes: blue �uorescence indicates DNA (chro-mosomes) and green �uorescence indicates microtubules (spindle apparatus). (credit �mitosis drawings�:modi�cation of work by Mariana Ruiz Villareal; credit �micrographs�: modi�cation of work by Roy vanHeesbeen; credit �cytokinesis micrograph�: Wadsworth Center/New York State Department of Health;scale-bar data from Matt Russell)
Which of the following is the correct order of events in mitosis?
a.Sister chromatids line up at the metaphase plate. The kinetochore becomes attached to themitotic spindle. The nucleus reforms and the cell divides. Cohesin proteins break down andthe sister chromatids separate.
b.The kinetochore becomes attached to the mitotic spindle. Cohesin proteins break down andthe sister chromatids separate. Sister chromatids line up at the metaphase plate. The nucleusreforms and the cell divides.
c.The kinetochore becomes attached to the cohesin proteins. Sister chromatids line up at themetaphase plate. The kinetochore breaks down and the sister chromatids separate. Thenucleus reforms and the cell divides.
d.The kinetochore becomes attached to the mitotic spindle. Sister chromatids line up at themetaphase plate. Cohesin proteins break down and the sister chromatids separate. Thenucleus reforms and the cell divides.
During prophase, the ��rst phase,� the nuclear envelope starts to dissociate into small vesicles, and themembranous organelles (such as the Golgi complex or Golgi apparatus, and endoplasmic reticulum), fragmentand disperse toward the periphery of the cell. The nucleolus disappears (disperses). The centrosomes beginto move to opposite poles of the cell. Microtubules that will form the mitotic spindle extend between thecentrosomes, pushing them farther apart as the microtubule �bers lengthen. The sister chromatids begin tocoil more tightly with the aid of condensin proteins and become visible under a light microscope.
During prometaphase, the ��rst change phase,� many processes that were begun in prophase continueto advance. The remnants of the nuclear envelope fragment. The mitotic spindle continues to develop asmore microtubules assemble and stretch across the length of the former nuclear area. Chromosomes become
Available for free at Connexions <http://cnx.org/content/col11635/1.1>

13
more condensed and discrete. Each sister chromatid develops a protein structure called a kinetochorein the centromeric region (Figure 3.3). The proteins of the kinetochore attract and bind mitotic spindlemicrotubules. As the spindle microtubules extend from the centrosomes, some of these microtubules comeinto contact with and �rmly bind to the kinetochores. Once a mitotic �ber attaches to a chromosome, thechromosome will be oriented until the kinetochores of sister chromatids face the opposite poles. Eventually,all the sister chromatids will be attached via their kinetochores to microtubules from opposing poles. Spindlemicrotubules that do not engage the chromosomes are called polar microtubules. These microtubules overlapeach other midway between the two poles and contribute to cell elongation. Astral microtubules are locatednear the poles, aid in spindle orientation, and are required for the regulation of mitosis.
Figure 3.3: During prometaphase, mitotic spindle microtubules from opposite poles attach to eachsister chromatid at the kinetochore. In anaphase, the connection between the sister chromatids breaksdown, and the microtubules pull the chromosomes toward opposite poles.
Duringmetaphase, the �change phase,� all the chromosomes are aligned in a plane called themetaphaseplate, or the equatorial plane, midway between the two poles of the cell. The sister chromatids are stilltightly attached to each other by cohesin proteins. At this time, the chromosomes are maximally condensed.
During anaphase, the �upward phase,� the cohesin proteins degrade, and the sister chromatids separateat the centromere. Each chromatid, now called a chromosome, is pulled rapidly toward the centrosome towhich its microtubule is attached. The cell becomes visibly elongated (oval shaped) as the polar microtubulesslide against each other at the metaphase plate where they overlap.
During telophase, the �distance phase,� the chromosomes reach the opposite poles and begin to de-condense (unravel), relaxing into a chromatin con�guration. The mitotic spindles are depolymerized intotubulin monomers that will be used to assemble cytoskeletal components for each daughter cell. Nuclearenvelopes form around the chromosomes, and nucleosomes appear within the nuclear area.
Available for free at Connexions <http://cnx.org/content/col11635/1.1>

14 CHAPTER 3. CELLULAR REPRODUCTION: THE CELL CYCLE
3.2.2 Cytokinesis
Cytokinesis, or �cell motion,� is the second main stage of the mitotic phase during which cell division iscompleted via the physical separation of the cytoplasmic components into two daughter cells. Division is notcomplete until the cell components have been apportioned and completely separated into the two daughtercells. Although the stages of mitosis are similar for most eukaryotes, the process of cytokinesis is quitedi�erent for eukaryotes that have cell walls, such as plant cells.
In cells such as animal cells that lack cell walls, cytokinesis follows the onset of anaphase. A contractilering composed of actin �laments forms just inside the plasma membrane at the former metaphase plate. Theactin �laments pull the equator of the cell inward, forming a �ssure. This �ssure, or �crack,� is called thecleavage furrow. The furrow deepens as the actin ring contracts, and eventually the membrane is cleavedin two (Figure 3.4).
In plant cells, a new cell wall must form between the daughter cells. During interphase, the Golgiapparatus accumulates enzymes, structural proteins, and glucose molecules prior to breaking into vesiclesand dispersing throughout the dividing cell. During telophase, these Golgi vesicles are transported onmicrotubules to form a phragmoplast (a vesicular structure) at the metaphase plate. There, the vesicles fuseand coalesce from the center toward the cell walls; this structure is called a cell plate. As more vesiclesfuse, the cell plate enlarges until it merges with the cell walls at the periphery of the cell. Enzymes use theglucose that has accumulated between the membrane layers to build a new cell wall. The Golgi membranesbecome parts of the plasma membrane on either side of the new cell wall (Figure 3.4).
Available for free at Connexions <http://cnx.org/content/col11635/1.1>

15
Figure 3.4: During cytokinesis in animal cells, a ring of actin �laments forms at the metaphase plate.The ring contracts, forming a cleavage furrow, which divides the cell in two. In plant cells, Golgi vesiclescoalesce at the former metaphase plate, forming a phragmoplast. A cell plate formed by the fusion ofthe vesicles of the phragmoplast grows from the center toward the cell walls, and the membranes of thevesicles fuse to form a plasma membrane that divides the cell in two.
3.3 G0 Phase
Not all cells adhere to the classic cell cycle pattern in which a newly formed daughter cell immediately entersthe preparatory phases of interphase, closely followed by the mitotic phase. Cells in G0 phase are notactively preparing to divide. The cell is in a quiescent (inactive) stage that occurs when cells exit the cellcycle. Some cells enter G0 temporarily until an external signal triggers the onset of G1. Other cells thatnever or rarely divide, such as mature cardiac muscle and nerve cells, remain in G0 permanently.
: Determine the Time Spent in Cell Cycle Stages
Problem: How long does a cell spend in interphase compared to each stage of mitosis?
Background: A prepared microscope slide of blastula cross-sections will show cells arrested invarious stages of the cell cycle. It is not visually possible to separate the stages of interphase fromeach other, but the mitotic stages are readily identi�able. If 100 cells are examined, the number
Available for free at Connexions <http://cnx.org/content/col11635/1.1>
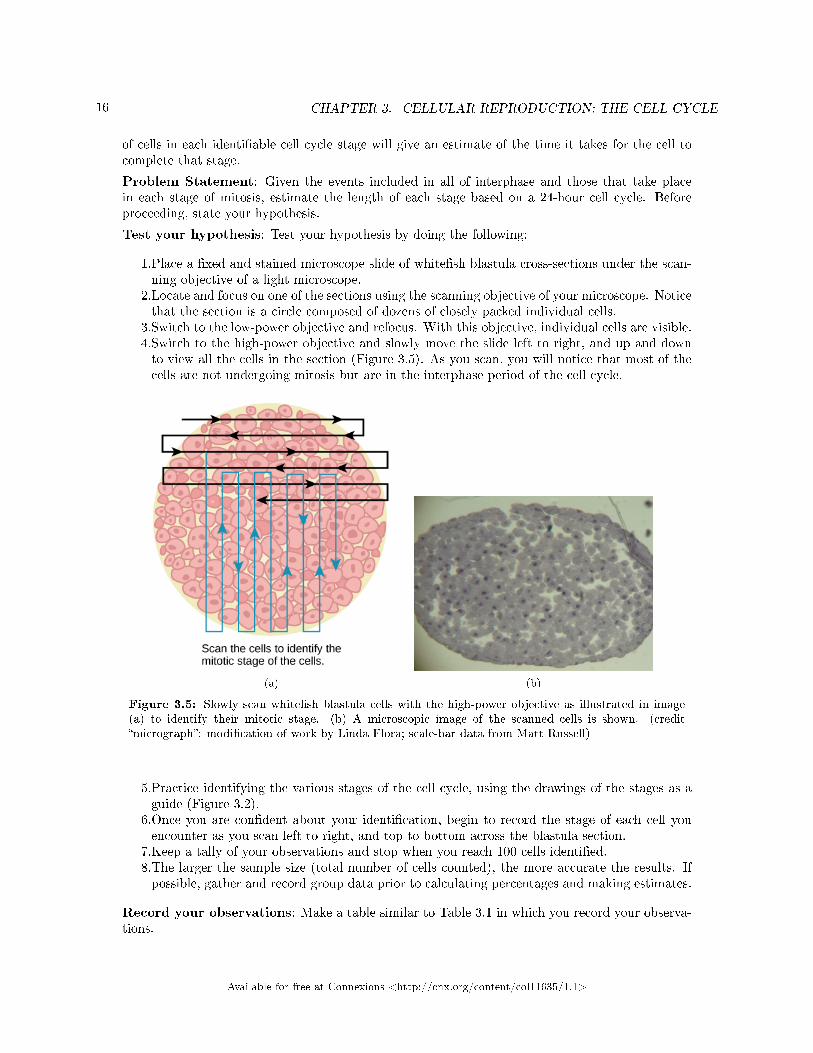
16 CHAPTER 3. CELLULAR REPRODUCTION: THE CELL CYCLE
of cells in each identi�able cell cycle stage will give an estimate of the time it takes for the cell tocomplete that stage.
Problem Statement: Given the events included in all of interphase and those that take placein each stage of mitosis, estimate the length of each stage based on a 24-hour cell cycle. Beforeproceeding, state your hypothesis.
Test your hypothesis: Test your hypothesis by doing the following:
1.Place a �xed and stained microscope slide of white�sh blastula cross-sections under the scan-ning objective of a light microscope.
2.Locate and focus on one of the sections using the scanning objective of your microscope. Noticethat the section is a circle composed of dozens of closely packed individual cells.
3.Switch to the low-power objective and refocus. With this objective, individual cells are visible.4.Switch to the high-power objective and slowly move the slide left to right, and up and downto view all the cells in the section (Figure 3.5). As you scan, you will notice that most of thecells are not undergoing mitosis but are in the interphase period of the cell cycle.
(a) (b)
Figure 3.5: Slowly scan white�sh blastula cells with the high-power objective as illustrated in image(a) to identify their mitotic stage. (b) A microscopic image of the scanned cells is shown. (credit�micrograph�: modi�cation of work by Linda Flora; scale-bar data from Matt Russell)
5.Practice identifying the various stages of the cell cycle, using the drawings of the stages as aguide (Figure 3.2).
6.Once you are con�dent about your identi�cation, begin to record the stage of each cell youencounter as you scan left to right, and top to bottom across the blastula section.
7.Keep a tally of your observations and stop when you reach 100 cells identi�ed.8.The larger the sample size (total number of cells counted), the more accurate the results. Ifpossible, gather and record group data prior to calculating percentages and making estimates.
Record your observations: Make a table similar to Table 3.1 in which you record your observa-tions.
Available for free at Connexions <http://cnx.org/content/col11635/1.1>

17
Results of Cell Stage Identi�cation
Phase or Stage Individual Totals Group Totals Percent
Interphase
Prophase
Metaphase
Anaphase
Telophase
Cytokinesis
Totals 100 100 100 percent
Table 3.1
Analyze your data/report your results: To �nd the length of time white�sh blastula cellsspend in each stage, multiply the percent (recorded as a decimal) by 24 hours. Make a table similarto Table 3.2 to illustrate your data.
Estimate of Cell Stage Length
Phase or Stage Percent (as Decimal) Time in Hours
Interphase
Prophase
Metaphase
Anaphase
Telophase
Cytokinesis
Table 3.2
Draw a conclusion: Did your results support your estimated times? Were any of the outcomesunexpected? If so, discuss which events in that stage might contribute to the calculated time.
3.4 Section Summary
The cell cycle is an orderly sequence of events. Cells on the path to cell division proceed through a seriesof precisely timed and carefully regulated stages. In eukaryotes, the cell cycle consists of a long preparatoryperiod, called interphase. Interphase is divided into G1, S, and G2 phases. The mitotic phase begins withkaryokinesis (mitosis), which consists of �ve stages: prophase, prometaphase, metaphase, anaphase, andtelophase. The �nal stage of the mitotic phase is cytokinesis, during which the cytoplasmic components ofthe daughter cells are separated either by an actin ring (animal cells) or by cell plate formation (plant cells).
Available for free at Connexions <http://cnx.org/content/col11635/1.1>

18 CHAPTER 3. CELLULAR REPRODUCTION: THE CELL CYCLE
Available for free at Connexions <http://cnx.org/content/col11635/1.1>

Chapter 4
Cellular Reproduction: Control of the
Cell Cycle1
The length of the cell cycle is highly variable, even within the cells of a single organism. In humans, thefrequency of cell turnover ranges from a few hours in early embryonic development, to an average of two to�ve days for epithelial cells, and to an entire human lifetime spent in G0 by specialized cells, such as corticalneurons or cardiac muscle cells. There is also variation in the time that a cell spends in each phase of the cellcycle. When fast-dividing mammalian cells are grown in culture (outside the body under optimal growingconditions), the length of the cycle is about 24 hours. In rapidly dividing human cells with a 24-hour cellcycle, the G1 phase lasts approximately nine hours, the S phase lasts 10 hours, the G2 phase lasts aboutfour and one-half hours, and the M phase lasts approximately one-half hour. In early embryos of fruit �ies,the cell cycle is completed in about eight minutes. The timing of events in the cell cycle is controlled bymechanisms that are both internal and external to the cell.
4.1 Regulation of the Cell Cycle by External Events
Both the initiation and inhibition of cell division are triggered by events external to the cell when it is aboutto begin the replication process. An event may be as simple as the death of a nearby cell or as sweepingas the release of growth-promoting hormones, such as human growth hormone (HGH). A lack of HGH caninhibit cell division, resulting in dwar�sm, whereas too much HGH can result in gigantism. Crowding ofcells can also inhibit cell division. Another factor that can initiate cell division is the size of the cell; as acell grows, it becomes ine�cient due to its decreasing surface-to-volume ratio. The solution to this problemis to divide.
Whatever the source of the message, the cell receives the signal, and a series of events within the cellallows it to proceed into interphase. Moving forward from this initiation point, every parameter requiredduring each cell cycle phase must be met or the cycle cannot progress.
4.2 Regulation at Internal Checkpoints
It is essential that the daughter cells produced be exact duplicates of the parent cell. Mistakes in theduplication or distribution of the chromosomes lead to mutations that may be passed forward to every newcell produced from an abnormal cell. To prevent a compromised cell from continuing to divide, there areinternal control mechanisms that operate at three main cell cycle checkpoints. A checkpoint is one ofseveral points in the eukaryotic cell cycle at which the progression of a cell to the next stage in the cycle
1This content is available online at <http://cnx.org/content/m48939/1.1/>.
Available for free at Connexions <http://cnx.org/content/col11635/1.1>
19

20CHAPTER 4. CELLULAR REPRODUCTION: CONTROL OF THE CELL
CYCLE
can be halted until conditions are favorable. These checkpoints occur near the end of G1, at the G2/Mtransition, and during metaphase (Figure 4.1).
Figure 4.1: The cell cycle is controlled at three checkpoints. The integrity of the DNA is assessed atthe G1 checkpoint. Proper chromosome duplication is assessed at the G2 checkpoint. Attachment ofeach kinetochore to a spindle �ber is assessed at the M checkpoint.
4.2.1 The G1 Checkpoint
The G1 checkpoint determines whether all conditions are favorable for cell division to proceed. The G1
checkpoint, also called the restriction point (in yeast), is a point at which the cell irreversibly commits tothe cell division process. External in�uences, such as growth factors, play a large role in carrying the cellpast the G1 checkpoint. In addition to adequate reserves and cell size, there is a check for genomic DNAdamage at the G1 checkpoint. A cell that does not meet all the requirements will not be allowed to progressinto the S phase. The cell can halt the cycle and attempt to remedy the problematic condition, or the cellcan advance into G0 and await further signals when conditions improve.
Available for free at Connexions <http://cnx.org/content/col11635/1.1>

21
4.2.2 The G2 Checkpoint
The G2 checkpoint bars entry into the mitotic phase if certain conditions are not met. As at the G1
checkpoint, cell size and protein reserves are assessed. However, the most important role of the G2 checkpointis to ensure that all of the chromosomes have been replicated and that the replicated DNA is not damaged.If the checkpoint mechanisms detect problems with the DNA, the cell cycle is halted, and the cell attemptsto either complete DNA replication or repair the damaged DNA.
4.2.3 The M Checkpoint
The M checkpoint occurs near the end of the metaphase stage of karyokinesis. The M checkpoint is also knownas the spindle checkpoint, because it determines whether all the sister chromatids are correctly attached tothe spindle microtubules. Because the separation of the sister chromatids during anaphase is an irreversiblestep, the cycle will not proceed until the kinetochores of each pair of sister chromatids are �rmly anchoredto at least two spindle �bers arising from opposite poles of the cell.
: Watch what occurs at the G1, G2, and M checkpoints by visitingthis website2 to see an animation of the cell cycle.
4.3 Regulator Molecules of the Cell Cycle
In addition to the internally controlled checkpoints, there are two groups of intracellular molecules thatregulate the cell cycle. These regulatory molecules either promote progress of the cell to the next phase(positive regulation) or halt the cycle (negative regulation). Regulator molecules may act individually, orthey can in�uence the activity or production of other regulatory proteins. Therefore, the failure of a singleregulator may have almost no e�ect on the cell cycle, especially if more than one mechanism controls the sameevent. Conversely, the e�ect of a de�cient or non-functioning regulator can be wide-ranging and possiblyfatal to the cell if multiple processes are a�ected.
4.3.1 Positive Regulation of the Cell Cycle
Two groups of proteins, called cyclins and cyclin-dependent kinases (Cdks), are responsible for theprogress of the cell through the various checkpoints. The levels of the four cyclin proteins �uctuate throughoutthe cell cycle in a predictable pattern (Figure 4.2). Increases in the concentration of cyclin proteins aretriggered by both external and internal signals. After the cell moves to the next stage of the cell cycle, thecyclins that were active in the previous stage are degraded.
2http://openstaxcollege.org/l/cell_checkpnts
Available for free at Connexions <http://cnx.org/content/col11635/1.1>

22CHAPTER 4. CELLULAR REPRODUCTION: CONTROL OF THE CELL
CYCLE
Figure 4.2: The concentrations of cyclin proteins change throughout the cell cycle. There is a directcorrelation between cyclin accumulation and the three major cell cycle checkpoints. Also note the sharpdecline of cyclin levels following each checkpoint (the transition between phases of the cell cycle), ascyclin is degraded by cytoplasmic enzymes. (credit: modi�cation of work by "WikiMiMa"/WikimediaCommons)
Cyclins regulate the cell cycle only when they are tightly bound to Cdks. To be fully active, the Cdk/cyclincomplex must also be phosphorylated in speci�c locations. Like all kinases, Cdks are enzymes (kinases) thatphosphorylate other proteins. Phosphorylation activates the protein by changing its shape. The proteinsphosphorylated by Cdks are involved in advancing the cell to the next phase. (Figure 4.3). The levels ofCdk proteins are relatively stable throughout the cell cycle; however, the concentrations of cyclin �uctuateand determine when Cdk/cyclin complexes form. The di�erent cyclins and Cdks bind at speci�c points inthe cell cycle and thus regulate di�erent checkpoints.
Available for free at Connexions <http://cnx.org/content/col11635/1.1>
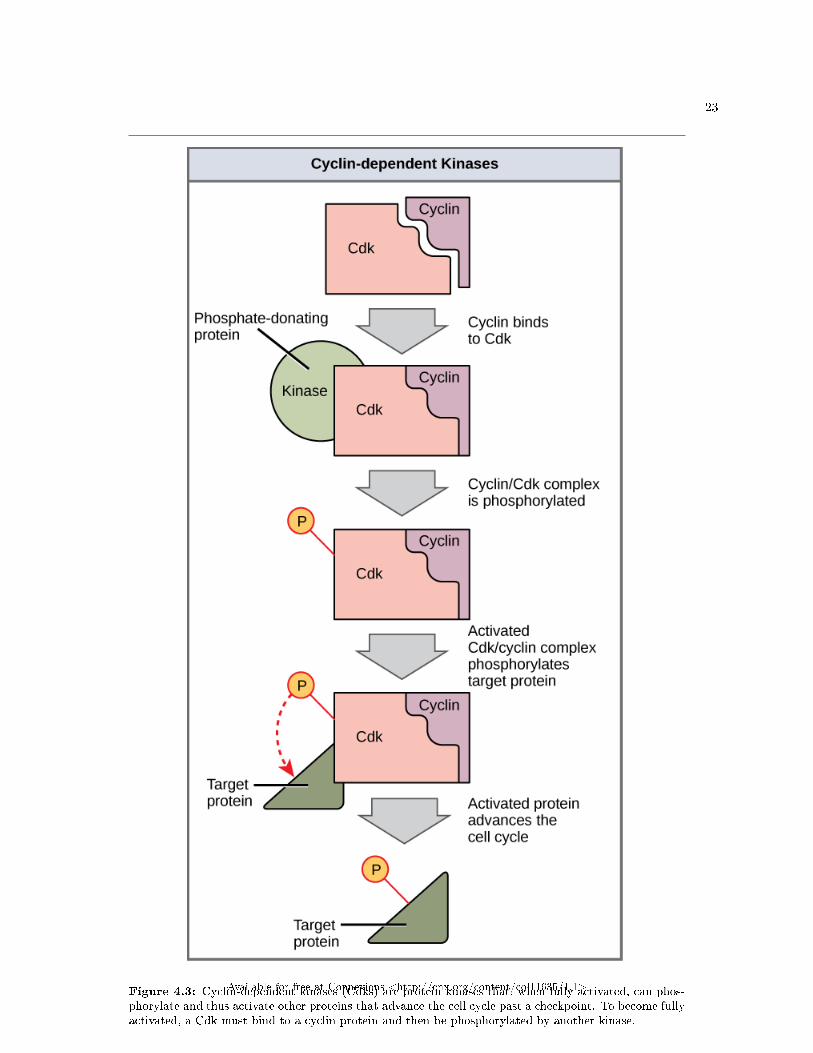
23
Figure 4.3: Cyclin-dependent kinases (Cdks) are protein kinases that, when fully activated, can phos-phorylate and thus activate other proteins that advance the cell cycle past a checkpoint. To become fullyactivated, a Cdk must bind to a cyclin protein and then be phosphorylated by another kinase.
Available for free at Connexions <http://cnx.org/content/col11635/1.1>

24CHAPTER 4. CELLULAR REPRODUCTION: CONTROL OF THE CELL
CYCLE
Since the cyclic �uctuations of cyclin levels are based on the timing of the cell cycle and not on speci�cevents, regulation of the cell cycle usually occurs by either the Cdk molecules alone or the Cdk/cyclincomplexes. Without a speci�c concentration of fully activated cyclin/Cdk complexes, the cell cycle cannotproceed through the checkpoints.
Although the cyclins are the main regulatory molecules that determine the forward momentum of thecell cycle, there are several other mechanisms that �ne-tune the progress of the cycle with negative, ratherthan positive, e�ects. These mechanisms essentially block the progression of the cell cycle until problematicconditions are resolved. Molecules that prevent the full activation of Cdks are called Cdk inhibitors. Manyof these inhibitor molecules directly or indirectly monitor a particular cell cycle event. The block placedon Cdks by inhibitor molecules will not be removed until the speci�c event that the inhibitor monitors iscompleted.
4.3.2 Negative Regulation of the Cell Cycle
The second group of cell cycle regulatory molecules are negative regulators. Negative regulators halt the cellcycle. Remember that in positive regulation, active molecules cause the cycle to progress.
The best understood negative regulatory molecules are retinoblastoma protein (Rb), p53, and p21.Retinoblastoma proteins are a group of tumor-suppressor proteins common in many cells. The 53 and 21designations refer to the functional molecular masses of the proteins (p) in kilodaltons. Much of whatis known about cell cycle regulation comes from research conducted with cells that have lost regulatorycontrol. All three of these regulatory proteins were discovered to be damaged or non-functional in cells thathad begun to replicate uncontrollably (became cancerous). In each case, the main cause of the uncheckedprogress through the cell cycle was a faulty copy of the regulatory protein.
Rb, p53, and p21 act primarily at the G1 checkpoint. p53 is a multi-functional protein that has a majorimpact on the commitment of a cell to division because it acts when there is damaged DNA in cells thatare undergoing the preparatory processes during G1. If damaged DNA is detected, p53 halts the cell cycleand recruits enzymes to repair the DNA. If the DNA cannot be repaired, p53 can trigger apoptosis, or cellsuicide, to prevent the duplication of damaged chromosomes. As p53 levels rise, the production of p21 istriggered. p21 enforces the halt in the cycle dictated by p53 by binding to and inhibiting the activity of theCdk/cyclin complexes. As a cell is exposed to more stress, higher levels of p53 and p21 accumulate, makingit less likely that the cell will move into the S phase.
Rb exerts its regulatory in�uence on other positive regulator proteins. Chie�y, Rb monitors cell size. Inthe active, dephosphorylated state, Rb binds to proteins called transcription factors, most commonly, E2F(Figure 4.4). Transcription factors �turn on� speci�c genes, allowing the production of proteins encoded bythat gene. When Rb is bound to E2F, production of proteins necessary for the G1/S transition is blocked.As the cell increases in size, Rb is slowly phosphorylated until it becomes inactivated. Rb releases E2F,which can now turn on the gene that produces the transition protein, and this particular block is removed.For the cell to move past each of the checkpoints, all positive regulators must be �turned on,� and all negativeregulators must be �turned o�.�
:
Available for free at Connexions <http://cnx.org/content/col11635/1.1>

25
Figure 4.4: Rb halts the cell cycle and releases its hold in response to cell growth.
Rb and other proteins that negatively regulate the cell cycle are sometimes called tumor suppressors.Why do you think the name tumor suppressor might be appropriate for these proteins?
4.4 Section Summary
Each step of the cell cycle is monitored by internal controls called checkpoints. There are three majorcheckpoints in the cell cycle: one near the end of G1, a second at the G2/M transition, and the third duringmetaphase. Positive regulator molecules allow the cell cycle to advance to the next stage. Negative regulatormolecules monitor cellular conditions and can halt the cycle until speci�c requirements are met.
Available for free at Connexions <http://cnx.org/content/col11635/1.1>

26CHAPTER 4. CELLULAR REPRODUCTION: CONTROL OF THE CELL
CYCLE
Available for free at Connexions <http://cnx.org/content/col11635/1.1>

Chapter 5
Cellular Reproduction: Cancer and the
Cell Cycle1
Cancer comprises many di�erent diseases caused by a common mechanism: uncontrolled cell growth. Despitethe redundancy and overlapping levels of cell cycle control, errors do occur. One of the critical processesmonitored by the cell cycle checkpoint surveillance mechanism is the proper replication of DNA during theS phase. Even when all of the cell cycle controls are fully functional, a small percentage of replication errors(mutations) will be passed on to the daughter cells. If changes to the DNA nucleotide sequence occur withina coding portion of a gene and are not corrected, a gene mutation results. All cancers start when a genemutation gives rise to a faulty protein that plays a key role in cell reproduction. The change in the cell thatresults from the malformed protein may be minor: perhaps a slight delay in the binding of Cdk to cyclin oran Rb protein that detaches from its target DNA while still phosphorylated. Even minor mistakes, however,may allow subsequent mistakes to occur more readily. Over and over, small uncorrected errors are passedfrom the parent cell to the daughter cells and ampli�ed as each generation produces more non-functionalproteins from uncorrected DNA damage. Eventually, the pace of the cell cycle speeds up as the e�ectivenessof the control and repair mechanisms decreases. Uncontrolled growth of the mutated cells outpaces thegrowth of normal cells in the area, and a tumor (�-oma�) can result.
5.1 Proto-oncogenes
The genes that code for the positive cell cycle regulators are called proto-oncogenes. Proto-oncogenes arenormal genes that, when mutated in certain ways, become oncogenes, genes that cause a cell to becomecancerous. Consider what might happen to the cell cycle in a cell with a recently acquired oncogene. Inmost instances, the alteration of the DNA sequence will result in a less functional (or non-functional) protein.The result is detrimental to the cell and will likely prevent the cell from completing the cell cycle; however,the organism is not harmed because the mutation will not be carried forward. If a cell cannot reproduce,the mutation is not propagated and the damage is minimal. Occasionally, however, a gene mutation causesa change that increases the activity of a positive regulator. For example, a mutation that allows Cdk tobe activated without being partnered with cyclin could push the cell cycle past a checkpoint before all ofthe required conditions are met. If the resulting daughter cells are too damaged to undergo further celldivisions, the mutation would not be propagated and no harm would come to the organism. However, if theatypical daughter cells are able to undergo further cell divisions, subsequent generations of cells will probablyaccumulate even more mutations, some possibly in additional genes that regulate the cell cycle.
The Cdk gene in the above example is only one of many genes that are considered proto-oncogenes. Inaddition to the cell cycle regulatory proteins, any protein that in�uences the cycle can be altered in such a
1This content is available online at <http://cnx.org/content/m48940/1.1/>.
Available for free at Connexions <http://cnx.org/content/col11635/1.1>
27

28CHAPTER 5. CELLULAR REPRODUCTION: CANCER AND THE CELL
CYCLE
way as to override cell cycle checkpoints. An oncogene is any gene that, when altered, leads to an increasein the rate of cell cycle progression.
5.2 Tumor Suppressor Genes
Like proto-oncogenes, many of the negative cell cycle regulatory proteins were discovered in cells that hadbecome cancerous. Tumor suppressor genes are segments of DNA that code for negative regulatorproteins, the type of regulators that, when activated, can prevent the cell from undergoing uncontrolleddivision. The collective function of the best-understood tumor suppressor gene proteins, Rb, p53, and p21,is to put up a roadblock to cell cycle progression until certain events are completed. A cell that carries amutated form of a negative regulator might not be able to halt the cell cycle if there is a problem. Tumorsuppressors are similar to brakes in a vehicle: Malfunctioning brakes can contribute to a car crash.
Mutated p53 genes have been identi�ed in more than one-half of all human tumor cells. This discoveryis not surprising in light of the multiple roles that the p53 protein plays at the G1 checkpoint. A cell with afaulty p53 may fail to detect errors present in the genomic DNA (Figure 5.1). Even if a partially functionalp53 does identify the mutations, it may no longer be able to signal the necessary DNA repair enzymes. Eitherway, damaged DNA will remain uncorrected. At this point, a functional p53 will deem the cell unsalvageableand trigger programmed cell death (apoptosis). The damaged version of p53 found in cancer cells, however,cannot trigger apoptosis.
:
Figure 5.1: The role of normal p53 is to monitor DNA and the supply of oxygen (hypoxia is a con-dition of reduced oxygen supply). If damage is detected, p53 triggers repair mechanisms. If repairs areunsuccessful, p53 signals apoptosis. A cell with an abnormal p53 protein cannot repair damaged DNAand thus cannot signal apoptosis. Cells with abnormal p53 can become cancerous. (credit: modi�cationof work by Thierry Soussi)
Human papillomavirus can cause cervical cancer. The virus encodes E6, a protein that binds p53.Based on this fact and what you know about p53, what e�ect do you think E6 binding has on p53activity?
Available for free at Connexions <http://cnx.org/content/col11635/1.1>

29
a.E6 activates p53b.E6 inactivates p53c.E6 mutates p53d.E6 binding marks p53 for degradation
The loss of p53 function has other repercussions for the cell cycle. Mutated p53 might lose its ability to triggerp21 production. Without adequate levels of p21, there is no e�ective block on Cdk activation. Essentially,without a fully functional p53, the G1 checkpoint is severely compromised and the cell proceeds directly fromG1 to S regardless of internal and external conditions. At the completion of this shortened cell cycle, twodaughter cells are produced that have inherited the mutated p53 gene. Given the non-optimal conditionsunder which the parent cell reproduced, it is likely that the daughter cells will have acquired other mutationsin addition to the faulty tumor suppressor gene. Cells such as these daughter cells quickly accumulate bothoncogenes and non-functional tumor suppressor genes. Again, the result is tumor growth.
: Go to this website2 to watch an animation of how cancer resultsfrom errors in the cell cycle.
5.3 Section Summary
Cancer is the result of unchecked cell division caused by a breakdown of the mechanisms that regulate thecell cycle. The loss of control begins with a change in the DNA sequence of a gene that codes for one ofthe regulatory molecules. Faulty instructions lead to a protein that does not function as it should. Anydisruption of the monitoring system can allow other mistakes to be passed on to the daughter cells. Eachsuccessive cell division will give rise to daughter cells with even more accumulated damage. Eventually, allcheckpoints become nonfunctional, and rapidly reproducing cells crowd out normal cells, resulting in a tumoror leukemia (blood cancer).
2http://openstaxcollege.org/l/cancer
Available for free at Connexions <http://cnx.org/content/col11635/1.1>

30CHAPTER 5. CELLULAR REPRODUCTION: CANCER AND THE CELL
CYCLE
Available for free at Connexions <http://cnx.org/content/col11635/1.1>

Chapter 6
Cellular Reproduction - Prokaryotic Cell
Division1
Prokaryotes, such as bacteria, propagate by binary �ssion. For unicellular organisms, cell division is the onlymethod to produce new individuals. In both prokaryotic and eukaryotic cells, the outcome of cell reproductionis a pair of daughter cells that are genetically identical to the parent cell. In unicellular organisms, daughtercells are individuals.
To achieve the outcome of cloned o�spring, certain steps are essential. The genomic DNA must bereplicated and then allocated into the daughter cells; the cytoplasmic contents must also be divided to giveboth new cells the machinery to sustain life. In bacterial cells, the genome consists of a single, circular DNAchromosome; therefore, the process of cell division is simpli�ed. Karyokinesis is unnecessary because thereis no nucleus and thus no need to direct one copy of the multiple chromosomes into each daughter cell. Thistype of cell division is called binary (prokaryotic) �ssion.
6.1 Binary Fission
Due to the relative simplicity of the prokaryotes, the cell division process, called binary �ssion, is a less com-plicated and much more rapid process than cell division in eukaryotes. The single, circular DNA chromosomeof bacteria is not enclosed in a nucleus, but instead occupies a speci�c location, the nucleoid, within the cell( here2). Although the DNA of the nucleoid is associated with proteins that aid in packaging the moleculeinto a compact size, there are no histone proteins and thus no nucleosomes in prokaryotes. The packingproteins of bacteria are, however, related to the cohesin and condensin proteins involved in the chromosomecompaction of eukaryotes.
The bacterial chromosome is attached to the plasma membrane at about the midpoint of the cell. Thestarting point of replication, the origin, is close to the binding site of the chromosome to the plasmamembrane (Figure 6.1). Replication of the DNA is bidirectional, moving away from the origin on bothstrands of the loop simultaneously. As the new double strands are formed, each origin point moves awayfrom the cell wall attachment toward the opposite ends of the cell. As the cell elongates, the growingmembrane aids in the transport of the chromosomes. After the chromosomes have cleared the midpoint ofthe elongated cell, cytoplasmic separation begins. The formation of a ring composed of repeating units ofa protein called FtsZ directs the partition between the nucleoids. Formation of the FtsZ ring triggers theaccumulation of other proteins that work together to recruit new membrane and cell wall materials to thesite. A septum is formed between the nucleoids, extending gradually from the periphery toward the centerof the cell. When the new cell walls are in place, the daughter cells separate.
1This content is available online at <http://cnx.org/content/m48941/1.1/>.2"Cell Division", Figure 1 <http://cnx.org/content/m44459/latest/#�g-ch10_01_01>
Available for free at Connexions <http://cnx.org/content/col11635/1.1>
31

32CHAPTER 6. CELLULAR REPRODUCTION - PROKARYOTIC CELL
DIVISION
Figure 6.1: These images show the steps of binary �ssion in prokaryotes. (credit: modi�cation of workby �Mcstrother�/Wikimedia Commons)
Available for free at Connexions <http://cnx.org/content/col11635/1.1>
33
: Mitotic Spindle Apparatus
The precise timing and formation of the mitotic spindle is critical to the success of eukaryoticcell division. Prokaryotic cells, on the other hand, do not undergo karyokinesis and thereforehave no need for a mitotic spindle. However, the FtsZ protein that plays such a vital role inprokaryotic cytokinesis is structurally and functionally very similar to tubulin, the building blockof the microtubules that make up the mitotic spindle �bers that are necessary for eukaryotes. FtsZproteins can form �laments, rings, and other three-dimensional structures that resemble the waytubulin forms microtubules, centrioles, and various cytoskeletal components. In addition, both FtsZand tubulin employ the same energy source, GTP (guanosine triphosphate), to rapidly assembleand disassemble complex structures.
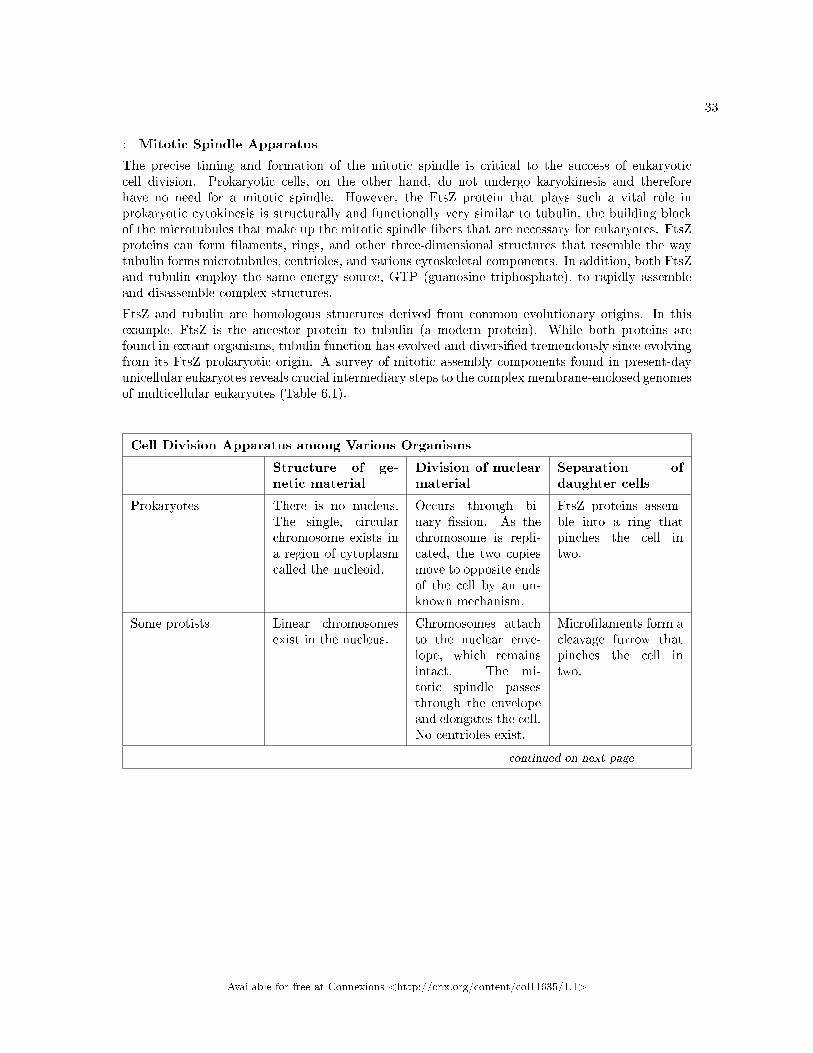
FtsZ and tubulin are homologous structures derived from common evolutionary origins. In thisexample, FtsZ is the ancestor protein to tubulin (a modern protein). While both proteins arefound in extant organisms, tubulin function has evolved and diversi�ed tremendously since evolvingfrom its FtsZ prokaryotic origin. A survey of mitotic assembly components found in present-dayunicellular eukaryotes reveals crucial intermediary steps to the complex membrane-enclosed genomesof multicellular eukaryotes (Table 6.1).
Cell Division Apparatus among Various Organisms
Structure of ge-netic material
Division of nuclearmaterial
Separation ofdaughter cells
Prokaryotes There is no nucleus.The single, circularchromosome exists ina region of cytoplasmcalled the nucleoid.
Occurs through bi-nary �ssion. As thechromosome is repli-cated, the two copiesmove to opposite endsof the cell by an un-known mechanism.
FtsZ proteins assem-ble into a ring thatpinches the cell intwo.
Some protists Linear chromosomesexist in the nucleus.
Chromosomes attachto the nuclear enve-lope, which remainsintact. The mi-totic spindle passesthrough the envelopeand elongates the cell.No centrioles exist.
Micro�laments form acleavage furrow thatpinches the cell intwo.
continued on next page
Available for free at Connexions <http://cnx.org/content/col11635/1.1>

34CHAPTER 6. CELLULAR REPRODUCTION - PROKARYOTIC CELL
DIVISION
Other protists Linear chromosomesexist in the nucleus.
A mitotic spindleforms from the cen-trioles and passesthrough the nuclearmembrane, whichremains intact. Chro-mosomes attach tothe mitotic spindle,which separates thechromosomes andelongates the cell.
Micro�laments form acleavage furrow thatpinches the cell intwo.
Animal cells Linear chromosomesexist in the nucleus.
A mitotic spindleforms from thecentrosomes. Thenuclear envelopedissolves. Chro-mosomes attach tothe mitotic spindle,which separates thechromosomes andelongates the cell.
Micro�laments form acleavage furrow thatpinches the cell intwo.
Table 6.1
6.2 Section Summary
In both prokaryotic and eukaryotic cell division, the genomic DNA is replicated and then each copy isallocated into a daughter cell. In addition, the cytoplasmic contents are divided evenly and distributed tothe new cells. However, there are many di�erences between prokaryotic and eukaryotic cell division. Bacteriahave a single, circular DNA chromosome but no nucleus. Therefore, mitosis is not necessary in bacterial celldivision. Bacterial cytokinesis is directed by a ring composed of a protein called FtsZ. Ingrowth of membraneand cell wall material from the periphery of the cells results in the formation of a septum that eventuallyconstructs the separate cell walls of the daughter cells.
Available for free at Connexions <http://cnx.org/content/col11635/1.1>

GLOSSARY 35
Glossary
A anaphase
stage of mitosis during which sisterchromatids are separated from each other
B binary �ssion
prokaryotic cell division process
C cell cycle checkpoint
mechanism that monitors the preparednessof a eukaryotic cell to advance throughthe various cell cycle stages
cell cycle
ordered sequence of events that a cellpasses through between one cell divisionand the next
cell cycle
ordered series of events involving cellgrowth and cell division that producestwo new daughter cells
cell plate
structure formed during plant cellcytokinesis by Golgi vesicles, forming atemporary structure (phragmoplast) andfusing at the metaphase plate; ultimatelyleads to the formation of cell walls thatseparate the two daughter cells
centriole
rod-like structure constructed ofmicrotubules at the center of each animalcell centrosome
centromere
region at which sister chromatids arebound together; a constricted area incondensed chromosomes
chromatid
single DNA molecule of two strands ofduplicated DNA and associated proteinsheld together at the centromere
cleavage furrow
constriction formed by an actin ringduring cytokinesis in animal cells thatleads to cytoplasmic division
condensin
proteins that help sister chromatids coilduring prophase
cyclin
one of a group of proteins that act inconjunction with cyclin-dependentkinases to help regulate the cell cycle byphosphorylating key proteins; theconcentrations of cyclins �uctuatethroughout the cell cycle
cyclin-dependent kinase
one of a group of protein kinases that helpsto regulate the cell cycle when bound tocyclin; it functions to phosphorylateother proteins that are either activated orinactivated by phosphorylation
cytokinesis
division of the cytoplasm following mitosisthat forms two daughter cells.
D diploid
cell, nucleus, or organism containing twosets of chromosomes (2n)
F FtsZ
tubulin-like protein component of theprokaryotic cytoskeleton that isimportant in prokaryotic cytokinesis(name origin: Filamentingtemperature-sensitive mutant Z)
G G0 phase
distinct from the G1 phase of interphase; acell in G0 is not preparing to divide
G1 phase
(also, �rst gap) �rst phase of interphasecentered on cell growth during mitosis
G2 phase
Available for free at Connexions <http://cnx.org/content/col11635/1.1>

36 GLOSSARY
(also, second gap) third phase ofinterphase during which the cellundergoes �nal preparations for mitosis
gamete
haploid reproductive cell or sex cell(sperm, pollen grain, or egg)
gene
physical and functional unit of heredity, asequence of DNA that codes for a protein.
genome
total genetic information of a cell ororganism
H haploid
cell, nucleus, or organism containing oneset of chromosomes (n)
histone
one of several similar, highly conserved,low molecular weight, basic proteinsfound in the chromatin of all eukaryoticcells; associates with DNA to formnucleosomes
homologous chromosomes
chromosomes of the same morphologywith genes in the same location; diploidorganisms have pairs of homologouschromosomes (homologs), with eachhomolog derived from a di�erent parent
I interphase
period of the cell cycle leading up tomitosis; includes G1, S, and G2 phases(the interim period between twoconsecutive cell divisions
K karyokinesis
mitotic nuclear division
kinetochore
protein structure associated with thecentromere of each sister chromatid thatattracts and binds spindle microtubulesduring prometaphase
L locus
position of a gene on a chromosome
M metaphase plate
equatorial plane midway between the twopoles of a cell where the chromosomesalign during metaphase
metaphase
stage of mitosis during which chromosomesare aligned at the metaphase plate
mitosis
(also, karyokinesis) period of the cell cycleduring which the duplicatedchromosomes are separated into identicalnuclei; includes prophase, prometaphase,metaphase, anaphase, and telophase
mitotic phase
period of the cell cycle during whichduplicated chromosomes are distributedinto two nuclei and cytoplasmic contentsare divided; includes karyokinesis(mitosis) and cytokinesis
mitotic spindle
apparatus composed of microtubules thatorchestrates the movement ofchromosomes during mitosis
N nucleosome
subunit of chromatin composed of a shortlength of DNA wrapped around a core ofhistone proteins
O oncogene
mutated version of a normal gene involvedin the positive regulation of the cell cycle
origin
(also, ORI) region of the prokaryoticchromosome where replication begins(origin of replication)
P p21
cell cycle regulatory protein that inhibitsthe cell cycle; its levels are controlled byp53
p53
cell cycle regulatory protein that regulatescell growth and monitors DNA damage;it halts the progression of the cell cycle incases of DNA damage and may induceapoptosis
Available for free at Connexions <http://cnx.org/content/col11635/1.1>

GLOSSARY 37
prometaphase
stage of mitosis during which the nuclearmembrane breaks down and mitoticspindle �bers attach to kinetochores
prophase
stage of mitosis during whichchromosomes condense and the mitoticspindle begins to form
proto-oncogene
normal gene that when mutated becomesan oncogene
Q quiescent
refers to a cell that is performing normalcell functions and has not initiatedpreparations for cell division
R retinoblastoma protein (Rb)
regulatory molecule that exhibits negativee�ects on the cell cycle by interacting
with a transcription factor (E2F)
S S phase
second, or synthesis, stage of interphaseduring which DNA replication occurs
septum
structure formed in a bacterial cell as aprecursor to the separation of the cellinto two daughter cells
T telophase
stage of mitosis during which chromosomesarrive at opposite poles, decondense, andare surrounded by a new nuclear envelope
tumor suppressor gene
segment of DNA that codes for regulatorproteins that prevent the cell fromundergoing uncontrolled division
Available for free at Connexions <http://cnx.org/content/col11635/1.1>

38 INDEX
Index of Keywords and Terms
Keywords are listed by the section with that keyword (page numbers are in parentheses). Keywordsdo not necessarily appear in the text of the page. They are merely associated with that section. Ex.apples, � 1.1 (1) Terms are referenced by the page they appear on. Ex. apples, 1
A anaphase, � 1(1), � 3(9), 13
B binary (prokaryotic) �ssion, 31binary �ssion, � 1(1), � 6(31)
C cancer, � 1(1), � 5(27)Cdk, � 1(1), � 4(19)cell cycle, � 1(1), � 2(3), 3, � 3(9), 9, � 5(27)cell cycle checkpoint, � 1(1), � 4(19)cell cycle checkpoints, 19cell cycle control, � 1(1), � 4(19)cell cycle regulation, � 1(1), � 4(19), � 5(27)cell division, � 1(1), � 2(3)cell motion, � 1(1), � 3(9)cell plate, � 1(1), � 3(9), 14cell reproduction, � 1(1), � 2(3), � 3(9),� 4(19), � 5(27), � 6(31)centriole, � 1(1), � 3(9)centrioles, 11centromere, � 1(1), � 2(3), 8checkpoint, � 1(1), � 4(19)chromatid, � 1(1), � 2(3)chromatids, 8chromosome, � 1(1), � 2(3)chromosome compaction, � 1(1), � 2(3)chromosomes, 5cleavage furrow, � 1(1), � 3(9), 14condensin, � 1(1), � 3(9), 12cyclin, � 1(1), � 4(19)cyclin-dependent kinase, � 1(1), � 4(19)cyclin-dependent kinases, 21cyclins, 21cytokinesis, � 1(1), � 3(9), 14cytoplasm, � 1(1), � 3(9)
D diploid, � 1(1), � 2(3), 4DNA, � 1(1), � 2(3)double helix, � 1(1), � 2(3)
E eukaryotic genome, � 1(1), � 2(3)
F �rst gap, � 1(1), � 3(9), � 4(19)FtsZ, � 1(1), � 6(31), 31
G G0 phase, � 1(1), � 3(9), 15G1, � 1(1), � 4(19)G1 checkpoint, � 1(1), � 4(19)G1 phase, � 1(1), � 3(9), 10G2, � 1(1), � 4(19)G2 checkpoint, � 1(1), � 4(19)G2 phase, � 1(1), � 3(9), 11gamete, � 1(1), � 2(3)gametes, 4gene, � 1(1), � 2(3)genes, 5genome, � 1(1), � 2(3), 3genomic DNA, � 1(1), � 2(3)Golgi apparatus, � 1(1), � 3(9)
H haploid, � 1(1), � 2(3), 4histone, � 1(1), � 2(3)histone protein, � 1(1), � 2(3)histone proteins, 6homologous, 5homologous chromosome, � 1(1), � 2(3)
I inhibition of cell division, � 1(1), � 4(19)initiation of cell division, � 1(1), � 4(19)interphase, � 1(1), � 3(9), 9
K karyokinesis, � 1(1), � 3(9), 11kinetochore, � 1(1), � 3(9), 13
L locus, � 1(1), � 2(3), 5
M M checkpoint, � 1(1), � 4(19)metaphase, � 1(1), � 3(9), 13, � 4(19)metaphase plate, � 1(1), � 3(9), 13mitosis, � 1(1), � 3(9), 11mitotic phase, � 1(1), � 3(9), 9mitotic spindle, � 1(1), � 3(9), 11mutation, � 1(1), � 5(27)
N negative regulation, � 1(1), � 4(19)nucleosome, � 1(1), � 2(3), 6
O oncogene, � 1(1), � 5(27)
Available for free at Connexions <http://cnx.org/content/col11635/1.1>

INDEX 39
oncogenes, 27origin, � 1(1), � 6(31), 31
P p21, � 1(1), � 4(19), 24p53, � 1(1), � 4(19), 24prokaryote cell division, � 6(31)prokaryotic cell division, � 1(1), � 6(31)prokaryotic genome, � 1(1), � 2(3)prometaphase, � 1(1), � 3(9), 12prophase, � 1(1), � 3(9), 12proto-oncogene, � 1(1), � 5(27)proto-oncogenes, 27
Q quiescent, � 1(1), � 3(9), 15
R Rb, � 1(1), � 4(19)
retinoblastoma protein, � 1(1), � 4(19)retinoblastoma protein (Rb), 24
S S phase, � 1(1), � 3(9), 11second gap, � 1(1), � 3(9), � 4(19)septum, � 1(1), � 6(31), 31sister chromatid, � 1(1), � 2(3)somatic cell, � 1(1), � 2(3)spindle checkpoint, � 1(1), � 4(19)
T telophase, � 1(1), � 3(9), 13trait, � 1(1), � 2(3)tumor suppressor, � 1(1), � 5(27)tumor suppressor gene, � 1(1), � 5(27)Tumor suppressor genes, 28
Available for free at Connexions <http://cnx.org/content/col11635/1.1>

40 ATTRIBUTIONS
Attributions
Collection: Cellular ReproductionEdited by: Je�rey MahrURL: http://cnx.org/content/col11635/1.1/License: http://creativecommons.org/licenses/by/3.0/
Module: "Cellular Reproduction - Introduction"By: Je�rey MahrURL: http://cnx.org/content/m48934/1.1/Page: 1Copyright: Je�rey MahrLicense: http://creativecommons.org/licenses/by/3.0/Based on: IntroductionBy: OpenStax CollegeURL: http://cnx.org/content/m44457/1.2/
Module: "Cellular Reproduction - Genomic DNA"By: Je�rey MahrURL: http://cnx.org/content/m48936/1.1/Pages: 3-8Copyright: Je�rey MahrLicense: http://creativecommons.org/licenses/by/3.0/Based on: Cell DivisionBy: OpenStax CollegeURL: http://cnx.org/content/m44459/1.7/
Module: "Cellular Reproduction: The Cell Cycle"By: Je�rey MahrURL: http://cnx.org/content/m48937/1.1/Pages: 9-17Copyright: Je�rey MahrLicense: http://creativecommons.org/licenses/by/3.0/Based on: The Cell CycleBy: OpenStax CollegeURL: http://cnx.org/content/m44460/1.7/
Module: "Cellular Reproduction: Control of the Cell Cycle"By: Je�rey MahrURL: http://cnx.org/content/m48939/1.1/Pages: 19-25Copyright: Je�rey MahrLicense: http://creativecommons.org/licenses/by/3.0/Based on: Control of the Cell CycleBy: OpenStax CollegeURL: http://cnx.org/content/m44466/1.6/
Available for free at Connexions <http://cnx.org/content/col11635/1.1>

ATTRIBUTIONS 41
Module: "Cellular Reproduction: Cancer and the Cell Cycle"By: Je�rey MahrURL: http://cnx.org/content/m48940/1.1/Pages: 27-29Copyright: Je�rey MahrLicense: http://creativecommons.org/licenses/by/3.0/Based on: Cancer and the Cell CycleBy: OpenStax CollegeURL: http://cnx.org/content/m44463/1.6/
Module: "Cellular Reproduction - Prokaryotic Cell Division"By: Je�rey MahrURL: http://cnx.org/content/m48941/1.1/Pages: 31-34Copyright: Je�rey MahrLicense: http://creativecommons.org/licenses/by/3.0/Based on: Prokaryotic Cell DivisionBy: OpenStax CollegeURL: http://cnx.org/content/m44467/1.6/
Available for free at Connexions <http://cnx.org/content/col11635/1.1>

About ConnexionsSince 1999, Connexions has been pioneering a global system where anyone can create course materials andmake them fully accessible and easily reusable free of charge. We are a Web-based authoring, teaching andlearning environment open to anyone interested in education, including students, teachers, professors andlifelong learners. We connect ideas and facilitate educational communities.
Connexions's modular, interactive courses are in use worldwide by universities, community colleges, K-12schools, distance learners, and lifelong learners. Connexions materials are in many languages, includingEnglish, Spanish, Chinese, Japanese, Italian, Vietnamese, French, Portuguese, and Thai. Connexions is partof an exciting new information distribution system that allows for Print on Demand Books. Connexionshas partnered with innovative on-demand publisher QOOP to accelerate the delivery of printed coursematerials and textbooks into classrooms worldwide at lower prices than traditional academic publishers.