Embed Size (px)
Citation preview

ACTA
UNIVERSITATIS
UPSALIENSIS
UPPSALA
2008
Digital Comprehensive Summaries of Uppsala Dissertationsfrom the Faculty of Medicine 371
Cellular design of heparan sulfate
The NDST enzymes and their regulation
PERNILLA CARLSSON
ISSN 1651-6206ISBN 978-91-554-7267-2urn:nbn:se:uu:diva-9231

���������� �������� �� ������ �������� � �� �������� ������� � ���� ������������� �� ������� ��������� ��������� � � �!!" �� !#$%& '� �(� ������ ' ���� ')(����(� *+������ ' �������,- .(� �������� /��� �� ������� � 0����(-
��������
������� )- �!!"- �������� ����� ' (����� ���'���- .(� 1��. �2���� �� �(�����������- 3��� ����������� ���������- ������� ��� � ���� ����� � � ����������� ������� �� �� ������� � � ����� � %- � ��- ������- 4��1 # "5#%5&&�5 �6 5�-
������ ���'��� ���������� ��� ������ /��( ��� �����(�� (����� ���'��� *��,�������(����� �(��� �����(�� � �(��- .(�� ��� '�� ���� ���'���� �� � ��������������� /(��� �(�� ����� �(��� ���� �� ��������� /��( � /��� ���� ' �2���� ��������� �������� �� ��� �(����� ������ � � ���� ' ������ �������� ��( ������������ ��������� �� � ����� �(������-3 ����� ���� ' �(� �������� '��������� ' ���������� �� �� �������� ������� � �(�
�������(����� ����- �� �(��� ������� ���� �������� ���'��� ������� /(��� (��(�� ���'��������� ��� ��������� '� � ����� ���� ' �(� �������� ��������- .(� �����(���� ' �� �� ������� ������ � /(��( � ����� ' �2���� ��� ������- ������ �����(��� ' (/�(�� ������ �� ��������� ���� ������ ����� � (/ '����� ' �� ���'��� ������� �������� �(����� (/ �� '��������� �� �������-.(�� �(���� �� '����� �������� ' � ' �(� �2���� ��������� '� �� ���'����
���������� 15�����������715���'����'����� *1��.,� � � ������� � �������� �(������(����� ������- ��''���� ������� ' 1��. �������� /��� ������� � �(��� ��8����$4, 9������7(����� ���'��� �����(����$ )�������� '����� ' 15���'���� �����:�
/(��� �(� ���'��� �� )3)� �� �(/ � �'����� �(� ���� � /(��( 1��. ���'��� �(����������� �''����� �(� ���� ��������� ' �(� �������(�����-44, 9������ ���'��� �����(����$ �(��������2��� ' � 1��.% ������ ������:� /(��� �
������ ������ ' 1��.% /(��( ������� � �'����� 1��.% ����� ������ �� �''��� ����������� �� ���������-444, 9������ ���'��� �����(���� � 2����'��($ +��� 1��. ���� /��( ������� ��������
������� ����� ������� ���������:� � /(��( '��� 2����'��( 1��.� /��� ���� ���(/ � �� ��������� � � ��������� �� ��������� ��������� ����-
� ������ (����� ���'���� (������ ���������� 1��.� )3)�� ;���
� ������ �������� � ���� �� � � ����� ��� ����� ��� ����!����� " #$%� ����������� ������ �&'(#)%* �������� �� � �
< )������ ������ �!!"
4��1 %6&%56�!64��1 # "5#%5&&�5 �6 5���$�$��$��$����5#��% *(���$77��-=�-��7������>��?��$�$��$��$����5#��%,

En fråga jag ej lyckas besvara hur jag på den än vrider och vänder: Bör ej pluralis av doktorand ändock vara doktoränder?!


List of papers
This thesis is based on the following papers: I Pernilla Carlsson, Jenny Presto, Dorothe Spillmann, Ulf Lin-
dahl and Lena Kjellén (2008), “Heparin/heparan sulfate bio-synthesis: Processive formation of N-sulfated domains”, J. Biol. Chem. 2008 Jul 18;283(29):20008-14
II Pernilla Carlsson, Jenny Presto and Lena Kjellén (2008),
“Heparan sulfate biosynthesis: Characterization of an NDST1 splice variant”, Manuscript
III Pernilla Carlsson, Johan Ledin and Lena Kjellén (2008),
“Heparan sulfate biosynthesis in zebrafish: Five NDST genes with distinct expression patterns during embryonic develop-ment”, Manuscript


Contents
Introduction.....................................................................................................9
Background...................................................................................................10 Glycosaminoglycans ................................................................................10 Heparan sulfate and heparin .....................................................................10 Heparan sulfate proteoglycans .................................................................10 Heparan sulfate biosynthesis ....................................................................12 PAPS ........................................................................................................13 NDST .......................................................................................................15 Heparan sulfate structure..........................................................................16
HS structure .........................................................................................16 Why care about HS structure? .............................................................17 Regulation of HS structure ..................................................................18
Why care about HS at all? - Biological functions of HS..........................20 Why do we need HSPGs and how do they exert their function?.........20 HS, PAPS and human disease .............................................................21 HS-transgenic animals .........................................................................22
Zebrafish as a model system ....................................................................25
Present investigation .....................................................................................27 Aims .........................................................................................................27 Results and discussion..............................................................................27
Paper I - Heparin/heparan sulfate biosynthesis: Processive formation of N-sulfated domains ..........................................................................27 Paper II - Heparan sulfate biosynthesis: Characterization of an NDST1 splice variant .......................................................................................29 Paper III - Heparan sulfate biosynthesis in zebrafish: Five NDST genes with distinct expression patterns during embryonic development .......31
Concluding remarks ......................................................................................33
Populärvetenskaplig sammanfattning ...........................................................34
Acknowledgment ..........................................................................................36
References.....................................................................................................38

Abbreviations
APS Adenosine 5´-phosphosulfate ADP Adenosine diphosphate ATP Adenosine triphosphate Bmp Bone morphogenetic protein EXT Exostosin FGF Fibroblast growth factor FGFR Fibroblast growth factor receptor GAG Glycosaminoglycan Gal Galactose GalN Galactosamine GlcA Glucuronic acid GlcN Glucosamine derivative GlcNAc N-Acetyl-glucosamine GlcNH2 Glucosamine HME Hereditary multiple exostoses HS Heparan sulfate HSPG Heparan sulfate proteoglycan IdoA Iduronic acid NDST N-Deacetylase/N-Sulfotransferase PAP 3´-Phosphoadenosine 5´-phosphate PAPS 3´-Phosphoadenosine 5´-phosphosulfate PAPSS PAPS synthase SGBS Simpson-Golabi-Behmel-syndrome TGF-� Transforming growth factor-� UDP Uridine 5´-diphosphate Xyl Xylose

9
Introduction
Heparan sulfate proteoglycans (HSPGs) are proteins with long, unbranched heparan sulfate (HS) polysaccharide chains attached to them. They are found on cell surfaces and in basement membranes where they exert their action by interacting with a wide range of enzymes and signaling molecules and are thereby involved in a range of various processes both during embryonic de-velopment and in adult physiology (Esko and Selleck, 2002; Hacker et al., 2005; Bishop et al., 2007). With just a few exceptions, all multicellular or-ganisms, from ancient cnidarians to modern mammals, express HSPGs, demonstrating their importance throughout evolution.
A great part of the biological functionality of proteoglycans can be di-rectly related to the polysaccharide part. HS chains display very variable sulfation1 patterns where highly sulfated regions are responsible for a large part of the biological activity. The biosynthesis of HS is a complex process in which a number of enzymes are involved. Better comprehension of how this process is regulated could reveal clues to how formation of HS sulfation patterns occurs, and thereby how HS functionality is controlled.
This thesis is focusing on regulation of one of the enzymes responsible for HS sulfation, glucosaminyl N-deacetylase/N-sulfotransferase (NDST), in an attempt to understand these mechanisms better. Different aspects of NDST regulation were studied in three projects, ranging from substrate level in vitro (paper I), protein level in cells (paper II) and RNA-level in vivo (pa-per III). In Paper I (“Heparin/heparan sulfate biosynthesis: Processive for-mation of N-sulfated domains”) it is shown that the sulfate donor PAPS can influence the manner in which NDST modifies the polysaccharide substrate. Paper II (“Heparan sulfate biosynthesis: Characterization of an NDST1 splice variant”) describes a splice variant of NDST1 which influences NDST1 protein levels and affects HS structure. In the third project (“Heparan sulfate biosynthesis in zebrafish: Five NDST genes with distinct expression patterns during embryonic development”) five zebrafish NDSTs were cloned and shown to be expressed in a temporally and spatially regu-lated manner.
1 The correct term is sulfonation since it refers to sulfonate groups, SO3
-, rather than sulfate, SO4
2-, groups. However, the terms sulfate, sulfated and sulfation are common when referring to sulfotransferases and their actions and will also be used throughout this text.

10
Background
Glycosaminoglycans Glycosaminoglycans, GAGs, are unbranched polysaccharide chains con-
sisting of repeating disaccharide units. One of the sugars in the repeating disaccharide unit is always a derivative of an amino sugar (D-glucosamine, GlcN, or D-galactosamine, GalN) whereas the second sugar residue is either a hexuronic acid (glucuronic acid, GlcA, or iduronic acid, IdoA) or a galac-tose unit (Gal). Four main groups of GAGs can be distinguished: heparan sulfate/heparin (GlcN-GlcA/IdoA), chondroitin sulfate/dermatan sulfate (GalN-GlcA/IdoA), keratan sulfate (GlcN-Gal) and hyaluronan (GlcNAc-GlcA). Except for hyaluronan, all GAGs are sulfated and covalently bound to core proteins in the form of proteoglycans.
Heparan sulfate and heparin Heparan sulfate (HS) and a highly sulfated form of HS called heparin are
GAGs consisting of N-acetyl-D-glucosamine (GlcNAc) and GlcA units that are partly modified by epimerization of GlcA to IdoA and by sulfation at different positions of both GlcNAc and IdoA. The resulting highly sulfated regions of HS are responsible for a large part of its biological activity. By binding various molecules such as extracellular matrix proteins, proteases, protease inhibitors, lipases/lipoproteins, and growth factors, HS influences many important processes, for example organogenesis during embryonic development, angiogenesis, lipid metabolism and cell adhesion (Esko and Selleck, 2002; Bishop et al., 2008). In addition, several different pathogens use HS to attach to and gain entry into the host cell (Rostand and Esko, 1997).
Heparan sulfate proteoglycans The ability of proteoglycans to interact with matrix- and cell-associated
components and growth factors derives from the structural diversity of both the polysaccharide and the core protein. Although this thesis mainly focuses

11
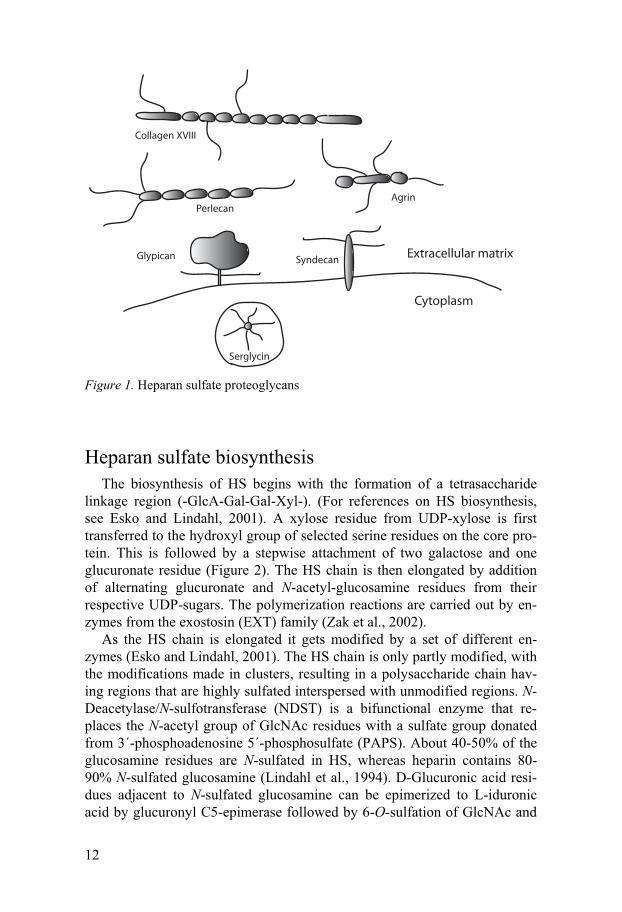
on the GAG part of the heparan sulfate proteoglycans the different classes of HSPGs will be described shortly below. (Figure 1)
Syndecans are transmembrane proteoglycans with rather large ectodo-mains (Tumova et al., 2000; Alexopoulou et al., 2007). The cytoplasmic part is small and comprises two conserved domains with a variable region in between allowing for both general and isoform specific interactions with cytoplasmic molecules. Four different syndecans, which show low sequence similarity between the ectodomains, have been identified in vertebrates, syn-decan-1–syndecan-4. All syndecans carry HS chains and syndecan-1 and syndecan-3 can also be substituted with chondroitin sulfate.
Glypicans are attached to the cell membrane via a glycosylphosphatidy-linositol anchor and thus are not integral proteoglycans (Filmus and Selleck, 2001). The glypicans have a large extracellular domain rich in cysteine. The cysteine residues probably bind to each other through disulfide bonds, mak-ing the glycoprotein compact and globular. Glypicans are substituted exclu-sively with HS chains which in contrast to syndecan HS are located in close proximity to the cell surface. Six glypicans have been identified, glypican 1-glypican 6.
Perlecan is present in virtually all basement membranes of the body and also in other extracellular matrices (Iozzo, 2005). The protein core consists of five domains and has three GAG attachment sites at the N-terminal do-main and one in the C-terminus. Perlecan is important for matrix organiza-tion and cell-matrix interactions.
Collagen XVIII is the only known member of the collagen family carry-ing HS chains (Iozzo, 2005). It is expressed in cartilage and in most base-ment membranes where it may function as a structural organizer. It shows a wide variety of molecular interactions with different growth factors and ad-hesion molecules.
Agrin was originally described as an organizer of the neuromuscular junc-tion (Bezakova and Ruegg, 2003). It is also found in various basement membranes, for instance in the glomerulus basement membrane where it may play a role in renal filtration.
Serglycin is the only known heparin proteoglycan (Kolset et al., 2004). When substituted with heparin chains, serglycin is found exclusively in con-nective tissue type mast cells where it is suggested to concentrate positively charged proteases, histamine and other molecules within the storage granules (Humphries et al., 1999; Henningsson et al., 2002). In other cell types, ser-glycin can carry chondroitin sulfate chains instead of heparin.
12
Cytoplasm
Extracellular matrix
Serglycin
Glypican Syndecan
PerlecanAgrin
Collagen XVIII
Figure 1. Heparan sulfate proteoglycans
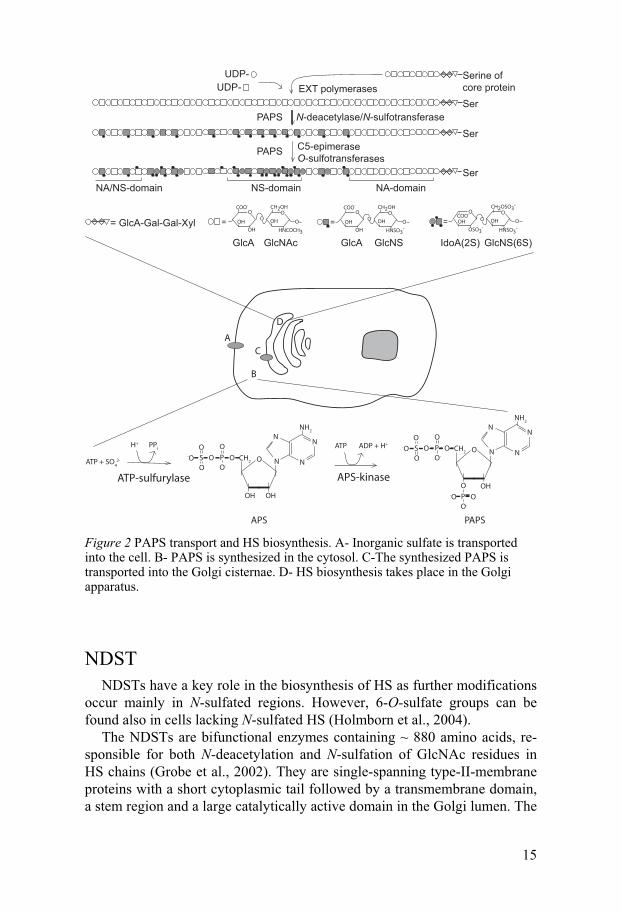
Heparan sulfate biosynthesis The biosynthesis of HS begins with the formation of a tetrasaccharide
linkage region (-GlcA-Gal-Gal-Xyl-). (For references on HS biosynthesis, see Esko and Lindahl, 2001). A xylose residue from UDP-xylose is first transferred to the hydroxyl group of selected serine residues on the core pro-tein. This is followed by a stepwise attachment of two galactose and one glucuronate residue (Figure 2). The HS chain is then elongated by addition of alternating glucuronate and N-acetyl-glucosamine residues from their respective UDP-sugars. The polymerization reactions are carried out by en-zymes from the exostosin (EXT) family (Zak et al., 2002).
As the HS chain is elongated it gets modified by a set of different en-zymes (Esko and Lindahl, 2001). The HS chain is only partly modified, with the modifications made in clusters, resulting in a polysaccharide chain hav-ing regions that are highly sulfated interspersed with unmodified regions. N-Deacetylase/N-sulfotransferase (NDST) is a bifunctional enzyme that re-places the N-acetyl group of GlcNAc residues with a sulfate group donated from 3´-phosphoadenosine 5´-phosphosulfate (PAPS). About 40-50% of the glucosamine residues are N-sulfated in HS, whereas heparin contains 80-90% N-sulfated glucosamine (Lindahl et al., 1994). D-Glucuronic acid resi-dues adjacent to N-sulfated glucosamine can be epimerized to L-iduronic acid by glucuronyl C5-epimerase followed by 6-O-sulfation of GlcNAc and

13
2-O-sulfation of IdoA and, more rarely, of GlcA, by 6-O-sulfotransferase and 2-O-sulfotransferase, respectively. 3-O-Sulfotransferase adds sulfate groups to GlcNAc, although this modification is not very common. Also GlcNH2 residues can be found in HS, although not very frequently (Westling and Lindahl, 2002). How these units are formed is not yet known.
All mammalian HS biosynthesis enzymes, except for 2-O-sulfotransferase and C5-epimerase exist in different isoforms with three 6-O-sulfotransferases, seven 3-O-sulfotransferases and four NDSTs reported. The fruitfly Drosphila melanogaster and the round-worm Caenorhabditis ele-gans have orthologues to the mammalian enzymes, but they hold only one isoform of each. In zebrafish (Danio rerio) one 2-O-sulfotransferase, two C5-epimerases, four 6-O-sulfotransferases and eight 3-O-sulfotransferases have been reported (Bink et al., 2003; Chen et al., 2005; Ghiselli and Farber, 2005; Cadwallader and Yost, 2006b; Cadwallader and Yost, 2006a; Cadwal-lader and Yost, 2007). In paper III of this thesis five zebrafish NDSTs are described.
Post-synthetic modification of HS also occurs, performed by two endosul-fatases, Sulf1 and Sulf2, located at the cell surface (Lamanna et al., 2007). These sulfatases act on the HS chain after extracellular translocation, by cleaving off 6-O-sulfate groups in the internal part of the polysaccharide chain.
PAPS PAPS is the universal sulfate donor for all biochemical sulfotransferase
reactions, including sulfation of HS. For sulfation reactions taking place in the Golgi compartment, as in HS biosynthesis, inorganic sulfate must be transported into the cell, transformed into its activated form, PAPS, and be translocated into the Golgi where it is used as a substrate (Figure 2).
Cellular uptake of sulfate is performed by a number of transmembrane an-tiporter and symporter molecules (ul Haque et al., 1998). A spectrum of recessively inherited disorders affecting bone and cartilage development have been related to mutations of genes encoding such sulfate transporters (Superti-Furga et al., 1996).
In the cytosol, PAPS is synthesized from adenosine triphosphate (ATP) and inorganic sulfate in a two-step reaction by PAPS synthase (PAPSS). In bacteria, fungi, yeast and plants, the two catalytic activities are located to separate polypeptide chains whereas in animals PAPSS is found as a bifunc-tional enzyme containing both an ATP sulfurylase domain and an APS kinase domain needed for the reaction. The first reaction step, in which inor-ganic sulfate reacts with ATP to form adenosine 5´-phosphosulfate (APS), is catalyzed by ATP sulfurylase, followed by formation of PAPS and adeno-sine diphosphate (ADP) from ATP and the preformed APS, catalyzed by

14
APS kinase (Strott, 2002; Venkatachalam, 2003) (Figure 2). Whereas only one PAPS synthase has been found in D. melanogaster (Jullien et al., 1997) and C. elegans (Dejima et al., 2006), two isoforms exist in vertebrates, PAPSS1 and PAPSS2 (Fuda et al., 2002; Strott, 2002; Venkatachalam, 2003). PAPSS2 has also been found as two differentially spliced products, differing only by the presence or absence of a five-amino-acid-stretch in the ATP sulfurylase domain. The expression of the isoforms differs, with PAPSS1 being found ubiquitously while PAPSS2 has a more restricted ex-pression pattern (Fuda et al., 2002).
Brachymorphic mice were found to have a mutation in PAPSS2 causing dwarfism and undersulfation of GAGs in cartilage extracellular matrix (Lane and Dickie, 1968; Orkin et al., 1976; Kurima et al., 1998; ul Haque et al., 1998). In humans, a mutation in the PAPSS2 gene, resulting in a truncated protein lacking the ATP sulfurylase domain, has been reported to cause an autosomal recessive skeletal defect associated with short stature at birth and degenerative joint disease (Ahmad et al., 1998; ul Haque et al., 1998). Total deficiency in PAPS-synthase in Caenorhabditis elegans results in larval lethality whereas moderate inhibition by RNA interference causes epidermal elongation defects (Dejima et al., 2006).
The PAPS synthases are localized to the cytosol and the nucleus and thus the PAPS utilized in HS sulfation must be transported into the Golgi com-partment after synthesis. Two such PAPS transporters have been identified in both human and Drosophila melanogaster (Kamiyama et al., 2003; Goda et al., 2006; Kamiyama et al., 2006) and very recently the zebrafish gene pinscher was also revealed to encode for a PAPS translocase (Clement et al., 2008). The Drosophila PAPS-transporters have been shown to be important for several signaling pathways, including Wingless, Hedgehog and Decapen-taplegic pathways, which are dependent on HS sulfation (Luders et al., 2003; Goda et al., 2006), as discussed below. Pinscher was first isolated in a large-scale screen for genes involved in pectoral fin development and was later found to be important also for jaw development and axon sorting processes (Karlstrom et al., 1996; Schilling et al., 1996; Trowe et al., 1996; Clement et al., 2008). PAPS transporters probably act by an antiport mechanism shuf-fling PAPS into the Golgi while transporting PAP back to the cytosol (Ozeran et al., 1996).
15
EXT polymerases
N-deacetylase/N-sulfotransferase
C5-epimerase O-sulfotransferases
Ser
NS-domain NA-domainNA/NS-domain
= GlcA-Gal-Gal-Xyl
Ser
Ser
GlcA GlcNAc
=
GlcNS(6S)IdoA(2S)
=
GlcA GlcNS
=
Serine of core proteinUDP-
UDP-
O
HNSO3-
OOCH2OSO3
-
OH OHCOO-
OSO3-
OO
HNSO3-
OOCH2OH
OH OH
COO-
OH
OO
HNCOCH3
OOCH2OH
OH OH
COO-
OH
O
PAPS
PAPS
C
B
A
D
O
OHOH
CH2
O
P OOO-
O
S-OO
N
NN
N
NH2
O
OH
CH2
O
P OOO-
O
S-OO
N
NN
N
NH2
O
P O-OO-
H+ PPi
ATP + SO4
2-
ATP-sulfurylase
ADP + H+ATP
APS-kinase
APS PAPS Figure 2 PAPS transport and HS biosynthesis. A- Inorganic sulfate is transported into the cell. B- PAPS is synthesized in the cytosol. C-The synthesized PAPS is transported into the Golgi cisternae. D- HS biosynthesis takes place in the Golgi apparatus.
NDST NDSTs have a key role in the biosynthesis of HS as further modifications
occur mainly in N-sulfated regions. However, 6-O-sulfate groups can be found also in cells lacking N-sulfated HS (Holmborn et al., 2004).
The NDSTs are bifunctional enzymes containing ~ 880 amino acids, re-sponsible for both N-deacetylation and N-sulfation of GlcNAc residues in HS chains (Grobe et al., 2002). They are single-spanning type-II-membrane proteins with a short cytoplasmic tail followed by a transmembrane domain, a stem region and a large catalytically active domain in the Golgi lumen. The

16
N-sulfotransferase domain has been located to the carboxyl part of the pro-tein and has been crystallized (Berninsone and Hirschberg, 1998; Kakuta et al., 1999). The deacetylase domain has not yet been possible to crystallize, but from a study expressing residues A66-P604 of human NDST2 as a pro-tein with retained N-deacetylase activity, the domain could be located to this part of the protein (Duncan et al., 2006). A cysteine at position 486 of rat NDST1 has been shown to be located close to the active site, as mutation of this residue to an amino acid with a large side chain abolishes N-deacetylase activity, while replacing it with alanine or valine results in enhanced enzyme activity, probably due to steric influence (Wei and Swiedler, 1999).
As mentioned above, many of the HS biosynthesis enzymes exist in dif-ferent isoforms. In the case of NDST, four vertebrate isoforms have been identified and cloned, NDST1-NDST4 (Hashimoto et al., 1992; Eriksson et al., 1994; Orellana et al., 1994; Kusche-Gullberg et al., 1998; Aikawa and Esko, 1999; Aikawa et al., 2001). In addition, NDST1 also exists in a shorter form, arising from a differently spliced version of the transcript, described in Paper II.
NDST1 and NDST2 transcripts are widely spread in various tissues both during the embryonic stage and in adult mice whereas NDST3 and NDST4 show a more modest mRNA expression (Kusche-Gullberg et al., 1998; Ai-kawa et al., 2001; Pallerla et al., 2008). NDST3 and NDST4 are mostly ex-pressed during embryonic development with NDST3 also found in adult brain. The splice variant of NDST1 was found to be expressed in lung, liver, kidney and in small amounts also in testis (Paper II). However, transcription levels may not necessarily correlate with translation levels. In fact, it seems that expression of the NDST isoforms may be translationally regulated (Grobe and Esko, 2002).
From computer modeling, using the crystal structure of the NDST1 sul-fotransferase domain as a template, it was shown that the four isoforms dif-fered in surface charge of amino acids constituting the substrate binding cleft, indicating that the isoforms have differing substrate preferences (Aikawa et al., 2001). In the same study, the four NDSTs were also shown to have different kinetic properties, as measured as N-deacetylase/N-sulfotransferase ratios.
Heparan sulfate structure HS structure
HS molecules are very complex. Except for being attached to a number of different core proteins they also show an enormous variation regarding chain length, domain structure, GlcA/IdoA content and sulfation pattern (Esko and Lindahl, 2001). Moreover, IdoA residues can shift between two different

17
conformations which further contributes to conformal flexibility of the HS chain (Torri et al., 1985).
A somewhat overlooked aspect of HS structure is the three-dimensional conformation of the polysaccharide chain. The tendency to write HS struc-tures as static, linear molecules with all N-sulfate groups on one side of the chain results in loss of view in spatial orientation which obviously is an im-portant factor when it comes to ligand binding. NMR studies and molecular modelling of heparin have shown that the polysaccharide has a helical con-formation with sulfate groups clustered at specific intervals alternating on opposite sides of the helix (Mulloy et al., 1993), as opposed to the common way of drawing 6-O-sulfates and 2-O-sulfates of adjacent monosaccharides as if they were located on opposite planes with respect to each other.
As mentioned above, modifications of the HS chain are made in clusters resulting in regions that are highly sulfated (NS-domains) interspersed with non-sulfated regions (NA-domains) and with intermediately sulfated regions (NA/NS-domains) usually surrounding the NS-domains (Gallagher, 2001). The sulfated regions are however not completely modified at all possible sites, resulting in specific patterns depending on cell type and at which time point the HS is produced (David et al., 1992; Lindahl et al., 1995; Maccarana et al., 1996; Brickman et al., 1998; van Kuppevelt et al., 1998; Jenniskens et al., 2000; Jenniskens et al., 2002; Allen and Rapraeger, 2003; ten Dam et al., 2003; Ledin et al., 2004; Warda et al., 2006). Notably, very little structural variation is seen when HS from the same tissue but from different individu-als is compared (Lindahl et al., 1995; Ledin et al., 2004), suggesting that HS synthesis is a highly regulated process.
Why care about HS structure? The question of HS specificity for ligand binding and biological activity
has long been debated. Since the finding of the pentasaccharide sequence with high affinity for antithrombin, resulting in a conformational change of the protein and enhanced interaction with thrombin leading to inhibition of blood coagulation (Petitou et al., 2003), the search has been ongoing for other such highly specific HS interactions. However, the result has been discouraging. Except for the herpes simplex gD protein requiring unusual 3-O-sulfated GlcNH2 residues for binding to HS (Shukla et al., 1999), most proteins seem to bind HS sequences with less specificity. 3-O-Sulfation is also a crucial component of the anti-thrombin-binding pentasaccharide. The involvement of 3-O-sulfate groups in these interactions, where a very spe-cific HS sequence is essential for binding, indicates that 3-O-sulfation probably is dedicated to interactions involving very specific requirements of HS structure, considering the general infrequency of this type of sulfation in HS. Notably, there are seven isoforms of the enzyme responsible for 3-O-sulfation, also suggesting a crucial role for this type of HS modification.

18
The interaction of fibroblast growth factors (FGFs) with their receptors (FGFRs) has been shown to be greatly enhanced when HS is acting as a co-receptor (Yayon et al., 1991). Both the FGF molecule and the FGFRs can bind to HS and the formation of a ternary complex including all three com-ponents is necessary for adequate signalling. A vast amount of studies re-garding the interactions between HS, FGFs and FGFRs have been per-formed, reporting differences in preferential HS sequences for the different FGFs and their receptors. However, no unique binding sequence has yet emerged, and the specificity of HS fine structure in the preferred sequences is being questioned since recent reports suggest that different FGFs can bind to a set of common HS motifs and that total charge density may be of greater importance than the actual positioning of sulfate groups (Kreuger et al., 2005; Jastrebova et al., 2006).
As shown in some cases, as for endostatin, interleukin-8, interferon-� and platelet factor 4 (Lortat-Jacob et al., 1995; Stringer and Gallagher, 1997; Spillmann et al., 1998; Kreuger et al., 2002), also the organization of NS- and NA-domains can be an important factor in HS-protein interactions.
In this context it should probably also be mentioned that neither high specificity, nor high affinity, necessarily has to be accompanied by high biological activity. Whether a protein binds to HS gives clues to its function but the results should always be interpreted with caution in respect to bio-logical activity.
Although the interplay between HS and most of its ligands may not be dependent on such highly fine-tuned sulfation patterns as previously be-lieved, but be more dependent on overall charge load and domain structure, it is beyond doubt that HS biosynthesis is a regulated process of great impor-tance in various biological events. To achieve better comprehension of HS binding and function and how HS structure is dictated, it is of importance to study regulation of the enzymes responsible for generating the different HS structures.
Regulation of HS structure How is the formation of distinct HS structures regulated? Why do not all
positions available for sulfation become modified? As opposed to DNA in protein synthesis, there is no template to determine the design of the final product. So far, we have fairly little knowledge of the regulatory mecha-nisms but it is generally believed that the relative expression of the different enzymes involved in HS biosynthesis is of importance. By regulating the abundance of the enzymes, at transcriptional or translational levels, or by changing the degradation level of specific enzymes, the HS structure could be influenced in a regulated manner. Along this line, a GAGosome model has been proposed, suggesting close assembly of the enzymes in a physical complex leading to different outcomes of HS structure depending on the

19
composition of the complex. Supporting this hypothesis, the HS polymerases EXT1 and EXT2 are known to exert their action as a complex (Kobayashi et al., 2000; McCormick et al., 2000; Senay et al., 2000). Physical association has also been observed between GlcA C5-epimerase and IdoA 2-O-sulfotransferase (Pinhal et al., 2001) and, recently, we reported an interaction between EXT2 and NDST1 (Presto et al., 2008). In this study, it was also shown that the expression levels of EXT polymerases can affect the amount of NDST1 protein in cells, in turn influencing HS structure. NDST1 protein levels appear to be affected also by expression of an NDST1 splice variant, as described in Paper II.
As mentioned above, the NDST isoforms have different N-deacetylase/N-sulfotransferase ratios and probably also differing substrate preferences, as indicated by dissimilarities in the substrate binding cleft (Aikawa et al., 2001). In fact, HS from HEK-293 cells over-expressing either NDST1 or NDST2 showed isoform-dependent structural differences (Pikas et al., 2000). Tentatively, this could contribute to HS regulation at a more refined level.
The knowledge of transcriptional control of HS biosynthesis enzymes is scarce, but the NDST2 gene has been shown to be under regulation of the GA-binding protein, which is a transcription factor (Morii et al., 2001). Regulation of the NDST proteins probably also occurs at the translational level as suggested by differential expression of constructs in which the dif-ferent NDST 5’-untranslated regions were ligated to a reporter gene (Grobe and Esko, 2002). Presumptively, also posttranslational modifications such as glycosylation could play a role as was recently shown for one of the chon-droitin sulfate sulfotransferases (Yusa et al., 2005).
The relative abundance of enzymes differing in substrate preference could possibly explain the origin of HS fine-structure, i.e. variations seen in sul-fation at the different positions of monosaccharides, but it could hardly ex-plain the formation of alternating NA- and NS-domains along the polysac-charide chain. Why is sulfation not spread evenly along the HS chain instead of being scattered in clusters? This question is yet far from being answered but the results in Paper I suggest that PAPS may act as one regulatory factor in domain pattern formation.
Another factor that will affect the outcome of HS biosynthesis is the ac-cess to the different substrates involved, i.e. UDP-sugars (UDP-GlcNAc and UDP-GlcA) and the sulfate donor PAPS, which in turn is influenced by ac-cess to the enzymes and substrates involved in their respective synthesis, and the transport of these molecules into the ER and Golgi where HS synthesis takes place. This is, however, a relatively unexplored field of the HS arena.
In addition to regulation during HS synthesis, the endosulfatases acting post-synthetically might be regarded as regulators of HS structure, since their action can affect different signaling pathways, such as Wnt, Bone-morphogenetic-protein (Bmp) and FGF2, differently (Lamanna et al., 2007).

20
Why care about HS at all? - Biological functions of HS Why do we need HSPGs and how do they exert their function?
As mentioned earlier, HSPGs are involved in a range of various proc-esses, both during embryonic development and in adult physiology, such as cell adhesion, lipid metabolism, inflammatory responses and angiogenesis (Hacker et al., 2005; Bishop et al., 2007), just to mention a few. Complete abolishment of HS is non-compatible with viability and alterations of HS modifications lead to a variation of different defects, as described below.
There are many different ways in which HSPGs can exert their function (Figure 3). Upon binding of HS, molecules may change conformation, which can influence downstream reactions, as in the case of antithrombin binding to heparin (Petitou et al., 2003). Several molecules can bind to the same HS chain, thereby being brought together which will enhance the likelihood for interaction between them (Lander, 1998; Nybakken and Perrimon, 2002). By facilitating the interaction between FGFs and their receptors, HSPGs act as co-receptors. HSPGs are also suggested to function in storage of growth factors in the extracellular matrix. By binding to HS, growth factors could be protected from proteolytic degradation and be released by cleavage of the HS chain when needed.
Another important process in which HSPGs is an important player is the formation of morphogen gradients (Lin, 2004; Hacker et al., 2005). Morpho-gens are signaling molecules acting directly on cells in a concentration de-pendent manner. Upon secretion from the cell where it is expressed, the morphogen spreads by restricted diffusion, creating a gradient with the high-est concentration close to the morphogen-producing cell. The shape of this gradient is depending on the rate of production, degradation and movement of the morphogen. While diffusing, the secreted molecule interacts with other molecules that can bind to it and thereby immobilize it, target it to degradation or prevent its degradation (Lander, 1998; Nybakken and Perri-mon, 2002). HSPGs contribute to this process, mainly by sequestering the morphogen and stabilizing the gradient. HSPGs may also play a more active role by transporting the morphogen across cells as demonstrated by the in-ability of Decapentaplegic and Hedgehog molecules to traverse cell clones mutant for the glypican homologue dally in Drosophila (Hacker et al., 2005). Some of the most studied morphogens include members of the Hedgehog, Wnt and transforming growth factor-� (TGF-�) families, all shown to be important players in pattern formation.
21
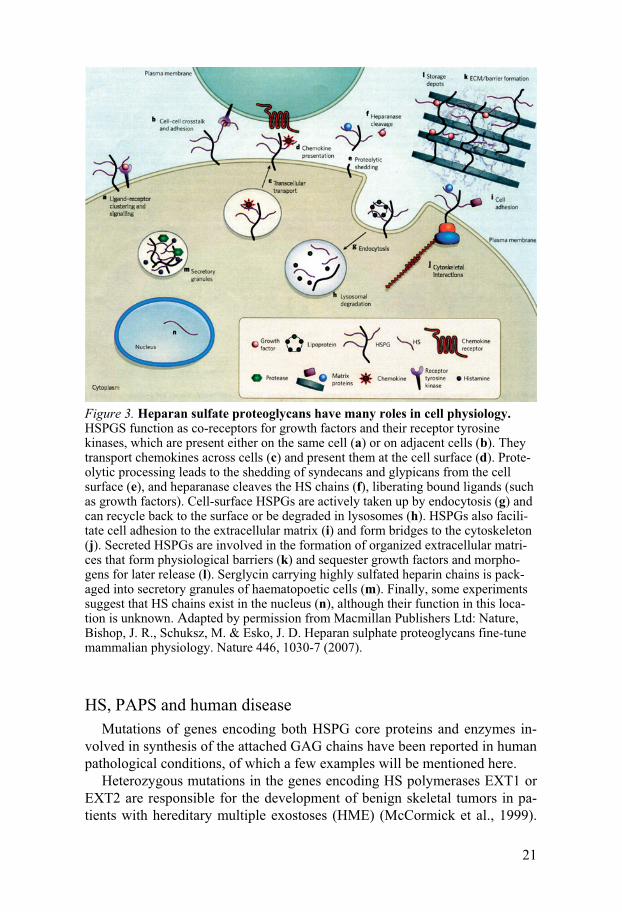
Figure 3. Heparan sulfate proteoglycans have many roles in cell physiology. HSPGS function as co-receptors for growth factors and their receptor tyrosine kinases, which are present either on the same cell (a) or on adjacent cells (b). They transport chemokines across cells (c) and present them at the cell surface (d). Prote-olytic processing leads to the shedding of syndecans and glypicans from the cell surface (e), and heparanase cleaves the HS chains (f), liberating bound ligands (such as growth factors). Cell-surface HSPGs are actively taken up by endocytosis (g) and can recycle back to the surface or be degraded in lysosomes (h). HSPGs also facili-tate cell adhesion to the extracellular matrix (i) and form bridges to the cytoskeleton (j). Secreted HSPGs are involved in the formation of organized extracellular matri-ces that form physiological barriers (k) and sequester growth factors and morpho-gens for later release (l). Serglycin carrying highly sulfated heparin chains is pack-aged into secretory granules of haematopoetic cells (m). Finally, some experiments suggest that HS chains exist in the nucleus (n), although their function in this loca-tion is unknown. Adapted by permission from Macmillan Publishers Ltd: Nature, Bishop, J. R., Schuksz, M. & Esko, J. D. Heparan sulphate proteoglycans fine-tune mammalian physiology. Nature 446, 1030-7 (2007).
HS, PAPS and human disease Mutations of genes encoding both HSPG core proteins and enzymes in-
volved in synthesis of the attached GAG chains have been reported in human pathological conditions, of which a few examples will be mentioned here.
Heterozygous mutations in the genes encoding HS polymerases EXT1 or EXT2 are responsible for the development of benign skeletal tumors in pa-tients with hereditary multiple exostoses (HME) (McCormick et al., 1999).

22
Although most of the tested HME patients bear heterozygotic mutations of either of the EXTs, the cause of the remaining cases is unknown. In a recent study, it was suggested that PAPS transporters might be candidate genes for HME in these patients, since the phenotype of zebrafish with mutations in a zebrafish PAPS-transporter-orthologue, pinscher, have defects resembling those of HME patients (Clement et al., 2008).
A mutation in the PAPS synthetase-2 gene has, as mentioned above, been reported to cause an autosomal recessive skeletal defect associated with short stature at birth and degenerative joint disease (Ahmad et al., 1998; ul Haque et al., 1998). The Simpson-Golabi-Behmel-syndrome (SGBS) instead causes overgrowth and heart and skeletal defects due to loss-of-function-mutations in the glypican-3 gene (Pilia et al., 1996; Neri et al., 1998).
In agreement with the effects described in humans, mice deficient in glypican-3 exhibit several of the clinical features demonstrated in SGBS patients (Cano-Gauci et al., 1999) and a PAPSS2 mutation was found to cause dwarfism also in mice (Kurima et al., 1998). Interestingly, no exosto-ses have been observed in mice heterozygous for EXT1 (Lin et al., 2000) and in EXT2 heterozygotic mice exostoses are located merely to the ribs (Stickens et al., 2005).
HS-transgenic animals Drosophila melanogaster and Caenorhabditis elegans
Most clues to HS function come from studies of transgenic animals where different HSPGs or HS biosynthesis enzymes have been modified. A lot of studies have been made in the fruitfly Drosophila melanogaster, especially focusing on the EXT polymerases tout-velu (orthologue of EXT1), sister-of-tout-velu (orthologue of EXT2), brother-of-tout-velu (orthologue of EXTL3) and on sulfateless, which is an NDST orthologue (Nybakken and Perrimon, 2002; Lin, 2004). Two Drosophila glypicans, dally and dally-like, have also been extensively studied. These studies have revealed the involvement of HSPGs in many important signalling pathways, including those of Wnt, Hedgehog and TGF-� family members. In most cases, HSPGs seem to func-tion as weak-affinity-binders of the secreted signalling molecules, required for the establishment and maintenance of morphogen gradients.
Mutations in the gene encoding HS polymerase (rib-2) in the nematode Caenorhabditis elegans affect egg-laying in F2 progeny and cause embry-onic lethality in F3 progeny (Morio et al., 2003), while 6-O-sulfotransferase and C5-epimerase are dispensable for viability but mutants show cell migra-tion and axon outgrowth defects (Bulow et al., 2002; Bulow and Hobert, 2004). Mutation of C. elegans 2-O-sulfotransferase results in cell migration defects (Bulow and Hobert, 2004; Kinnunen et al., 2005). So far, no muta-tions in NDST or 3-O-sulfotransferase have been described in C. elegans.

23
Zebrafish The zebrafish genes dackel and boxer, coding for exostosin proteins
EXT2 and EXTL3 respectively, were originally isolated in a large-scale genetic screen for pectoral fin development phenotypes (van Eeden et al., 1996). Later, these genes were also found to be important for proper axon guidance (Lee et al., 2004). In a recent study, zebrafish with mutations in dackel as well as zebrafish mutated in a PAPS transporter, pinscher, were shown to have cartilage defects resembling those in HME patients (Clement et al., 2008), suggesting pinscher to be a novel candidate gene for the devel-opment of HME.
Zebrafish commonly have duplicate orthologues to mammalian genes, due to a genome duplication event after the divergence of teleost and mam-malian ancestors, more than 300 million years ago (Jaillon et al., 2004). For instance, the zebrafish genome contains two C5-epimerases. Overexpression of the C5-epimerases gives a ventralized phenotype while reduced expres-sion after injection with morpholinos against the epimerases results in dor-salization (Ghiselli and Farber, 2005). 6-O-Sulfotranferase-2 has been sug-gested to be important for muscle development, since silencing of this gene resulted in disturbed somite formation (Bink et al., 2003). Another study targeting 6-O-sulfotranferase-2, showed its involvement in vessel formation, the morpholino especially affecting branching morphogenesis of the caudal vein (Chen et al., 2005). On the contrary, embryos injected with a mor-pholino targeting 6-O-sulfotranferase-1 did not show any obvious phenotype (Chen et al., 2005).
Mutations and morpholino-injections not only affecting the GAG chain, but the whole proteoglycan, have also been reported. For example, the ze-brafish glypican knypek has been found to be involved in the modulation of convergent extension movements during gastrulation (Topczewski et al., 2001). Syndecan-2 was found to play a role in angiogenesis (Chen et al., 2004) and perlecan is suggested to function both in skeletal muscle and car-diovascular development (Zoeller et al., 2008).
Mice deficient in HS biosynthesis enzymes Mouse mutant embryos deficient in EXT1 lack HS and fail to gastrulate
(Lin et al., 2000). When EXT1 is selectively disrupted in the central nervous system the mice exhibit several brain defects and die within the first day after birth (Inatani et al., 2003). Mice lacking EXT2 also die before gastrula-tion. Heterozygotes on the other hand, are fertile and have a normal lifespan (Stickens et al., 2005). However, exostoses develop in one third of EXT2+/- mice but are restricted to the ribs.
Mice lacking NDST1 show significant reduction of N-sulfated HS in most tissues, accompanied by a reduction in 2-O-sulfation, whereas 6-O-sulfation levels are less affected (Ledin et al., 2004). They die at or shortly after birth

24
due to lung failure and a minor proportion die already at the embryonic stage (Fan et al., 2000; Ringvall et al., 2000). Atelectasis2 is observed in lungs from NDST1-/- pups and their condition resembles the respiratory distress syndrome of premature infants. Moreover, skeletal and eye defects have been observed (Ringvall et al., 2000; Grobe et al., 2005; Pallerla et al., 2007). When NDST1 is inactivated selectively in endothelial cells and leu-kocytes, mice show impaired neutrophil infiltration during inflammatory responses (Wang et al., 2005). NDST1 has also been selectively disrupted in the central nervous system (Grobe et al., 2005). These mice show severe developmental defects of the forebrain, including lack of olfactory bulbs and axon guidance errors.
Since both NDST1 and NDST2 are highly expressed in most organs one could expect structural alterations of HS also in mice lacking NDST2. How-ever, experiments have shown that the HS structure in a number of tissues from such animals appear to be unaffected (Forsberg et al., 1999; Ledin et al., 2004). The only obvious defects are abnormalities in the morphology and composition of connective-tissue-type mast cells which are fewer and have reduced amounts of histamine and proteases (Forsberg et al., 1999; Humphries et al., 1999). They contain few secretory granules and instead have large, apparently empty, vacuoles. These effects are probably caused by a total lack of sulfated heparin, normally found in mast cells. Tentatively, other NDST isoforms may be more important in HS biosynthesis in cells other than mast cells and thus being able to compensate for a lack of NDST2. In connective-tissue-type mast cells, however, NDST2 is expressed at significant levels while very little or no NDST1-transcript is found (Kusche-Gullberg et al., 1998), explaining the dramatic effect of NDST2 deficiency on these cells.
NDST1 is obviously an important enzyme in HS biosynthesis of most tis-sues while NDST2 appears to be more involved in the production of heparin, in connective-tissue-type mast cells. However, mouse embryos lacking both NDST1 and NDST2 die during early embryogenesis, clearly pointing to a functional role also for NDST2 during embryonic development (Forsberg and Kjellen, 2001).
A recent study on NDST3 knockout mice reported no major phenotypes, but lack of both NDST1 and NDST3 results in a somewhat more severe phe-notype than that of NDST1 deficient mice (Pallerla et al., 2008). No NDST4 transgenic animals have so far been reported.
When C5-epimerase is eliminated in mice, the HS is completely devoid of IdoA and has a lower amount of 2-O-sulfate groups than HS from wildtype mice (Li et al., 2003). The overall sulfation level is however increased be-cause of elevated N- and 6-O-sulfation. The animals have phenotypes similar to NDST1-/- mice, i.e. neonatal death due to respiratory failure, lung defects 2 collapse of a portion of the lung or of an entire lung

25
and skeletal malformations. In addition, they lack kidneys, a phenotype also observed in mice deficient in 2-O-sulfotransferase but not in NDST1-/- ani-mals. As opposed to NDST1-/- mice the brain seems to be normally devel-oped. Heterozygous mice are indistinguishable from wildtype littermates, both regarding HS structure and phenotypic features.
2-O-Sulfotransferase null mice share most phenotypes of C5-epimerase and NDST1 knock-outs, but have apparently normal lungs and brain (Bullock et al., 1998). HS from embryonic fibroblasts derived from these mice lacks 2-O-sulfate groups but shows an increase in total sulfation due to elevated N- and 6-O-sulfation levels (Merry et al., 2001).
Transgenic mice lacking one of the 6-O-sulfotransferases, 6-O-sulfotransferase-1, die between embryonic day 15.5 and the perinatal stage, possibly due to abnormal angiogenesis of the placenta (Habuchi et al., 2007). They are smaller than their wildtype littermates and also exhibit abnormal navigation of retinal ganglion cell axons at the optic chiasm (Pratt et al., 2006).
3-O-sulfotransferase-1, although suggested to be the most critical isoform for producing the antithrombin-binding HS sequence, does not result in a procoagulant phenotype when deleted in mice (Shworak et al., 2002). In-stead, genetic-background-dependent lethality and intrauterine growth retar-dation are observed, but the cause of these abnormalities is not yet known.
Summary In conclusion, through studies of both invertebrate and vertebrate animal
models, HSPGs have proved to be important for development and mainte-nance of various important biological processes. Elimination of enzymes acting early in HS biosynthesis, such as EXT polymerases is incompatible with embryo development and the embryos die at very early stages. Surpris-ingly, animals lacking enzymes responsible for HS modification processes can fulfill embryogenesis, with defects restricted to certain organs and parts of the body. As many enzymes are expressed as different isoforms this may be explained by redundancy between the isoforms.
Zebrafish as a model system (This text is based on the book “Zebrafish”, (Nusslein-Volhard and Dahm, 2002))
Zebrafish was established as a model system in the 1970s and has since then become one of the most important organisms for studying biological processes in vivo. There are many reasons to why zebrafish have become so popular in research. First of all, the fish are quite cheap and easy to breed and keep. More importantly, they are relatively easy to manipulate geneti-cally and well suited for developmental studies. Fertilization and subsequent

26
embryonic development is external and study of the embryos does thus not require dissection. From a single breeding pair, large clutches of embryos can be obtained at the same occasion, facilitating genetic analysis and large-scale experiments. During the first 24 hours, the embryos are transparent and their development, even of internal organs, can be followed by simple light microscopy techniques. Moreover, embryonic development is rapid. All major body features, including brain, eyes, ears and internal organs, can be seen after only two days. The heart starts beating around 24 hours after fer-tilization, and as soon as five days after fertilization, zebrafish are able to swim and search for food. However, compared to other vertebrate model systems, the generation time of 2-4 months is not particularly short.
Ectopic expression of specific gene products can be performed by injec-tion of either DNA of mRNA. Injection of DNA results in highly mosaic expression patterns. The exact cause of this mosaicism is not known but one probable explanation is that the DNA, as opposed to mRNA, needs to enter the nucleus for expression. It is therefore advisable to perform the injection in the forming cell as early as possible. When mRNA is used for overexpres-sion, it can be injected into the yolk rather than the forming cells, since a process called ooplasmic streaming will transport the injected substance into the cell cytoplasm. Expression from injected mRNA is ubiquitous and does not result in the mosaic patterns obtained when DNA is used.
There is not yet any method to permanently delete a selected gene in ze-brafish, but by using the morpholino technique it is fairly easy to silence genes transiently. Morpholinos are synthetic antisense oligonucleotides with a morpholine ring instead of a ribose ring linked to the nucleotide bases. Upon binding to the corresponding sense mRNA, translation is blocked. By injection into the newly fertilized egg, morpholinos can eliminate functions of a specific gene at very early stages and have the advantage of reducing expression also of maternally contributed mRNAs. Although the effect is transient it is most often fully penetrant through the first two days of devel-opment. Though the morpholino technique has been useful for elucidating important functions of targeted genes, it is of vital importance that the ex-periments are performed with adequate controls (Eisen and Smith, 2008). To avoid phenotypes due to off-target effects, i.e. when the morpholino anneals to and blocks a gene other than the intended, rescue experiments by injec-tions with capped mRNA corresponding to the targeted gene should be per-formed when applicable. Moreover, in a recent paper it was described that non-specific neural defects can arise from morpholino injections due to acti-vation of the p53 gene with subsequent apoptosis (Robu et al., 2007). This problem can however be overcome by the co-injection with a morpholino against p53. The mechanism behind the p53 activation, and why only a frac-tion of the morpholinos results in such activation, is so far unknown.

27
Present investigation
Aims It is well established that HS structure varies depending on the cell type
and the stage at which the HS is produced. Interactions with proteins and growth factors are affected by the HS sulfation pattern but how the forma-tion of specific sulfate structures is controlled is far from understood. As NDST can be regarded a key enzyme in HS modification, with other modifi-cations located mainly to NS-domains, it is of great importance to elucidate its regulation. In this thesis different aspects of NDST regulation have been studied:
Paper I - Influence of PAPS on NDST action Paper II - Influence of a splice variant of NDST1 on HS sulfation Paper III - Expression patterns of different NDST isoforms in zebrafish
Results and discussion Paper I - Heparin/heparan sulfate biosynthesis: Processive formation of N-sulfated domains
PAPS is obviously an important component in HS biosynthesis since it is responsible for donating the sulfate group needed in sulfation reactions. In this project we studied how PAPS can influence NDST action, and thereby affect HS structure.
Recombinant NDST1 and NDST2 were incubated with bacterial K5 poly-saccharide, which consists of unmodified GlcNAc and GlcA residues in alternating sequence. When PAPS was excluded from the reaction mixture, the polysaccharide chain was modified in a random fashion. However, GlcNAc residues in neighboring disaccharides were rarely attacked by the enzyme under these conditions. Addition of PAPS, on the other hand, re-sulted in the generation of long, contiguous, N-sulfated stretches composed of �8 disaccharide units. These sulfated stretches were formed by an appar-ently processive action of the enzyme, as suggested by the almost exclusive production of consecutive, N-sulfated disaccharides, the amount of which increased with time. This suggests a process in which the enzyme acts by

28
working its way along the polysaccharide chain, rather than by initial ran-dom modification with subsequent sulfation of unmodified gaps.
The NS-domains formed were similar in size to those present in heparin and considerably longer than in naturally occurring HS. Heparin N-sulfation is performed by NDST2 (Kusche-Gullberg et al., 1998; Forsberg et al., 1999), while N-sulfation in HS producing cells seems to rely mostly on NDST1 (Ringvall et al., 2000; Ledin et al., 2006). It was therefore interest-ing to note that the extended NS-domains were formed both in the presence of NDST1 and NDST2. In intact cells, however, it probably matters whether NDST1 or NDST2 performs the N-sulfation reaction, as shown in HEK-293 cells where isoform-dependent differences in sulfation levels and structures were obtained (Pikas et al., 2000). Thus, proper formation of HS domain structure seem to demand additional factors associated with intact cells, which are lost when purified enzymes or subcellular fractions are used. This was also observed when rat liver microsomes were incubated with UDP-sugar precursors and PAPS, resulting in HS chains that were more heavily sulfated than HS produced in rat hepatocytes (Riesenfeld et al., 1982).
The drastic change in modification pattern upon PAPS addition, prompted us to study the effect of different PAPS-concentrations on HS structure. The relative abundance of consecutively N-sulfated disaccharides was higher after incubation with 5�M than with 1�M PAPS, indicating that variations in PAPS-concentration may have an influence on HS domain structure. Tenta-tively, HS structure may be regulated through variation in intracellular PAPS concentrations also in vivo. The physiological concentration of PAPS has been measured in animals and humans. In the few studies available, the es-timated PAPS-concentrations were in the range of 5-80 nmol/g tissue (Strott, 2002), which corresponds quite well to the concentrations used in our study. Measuring concentrations from whole tissues is however a crude method, not revealing putatively important intracellular local variations.
Uncoupling of N-deacetylation and N-sulfation reactions was performed by incubating K5 polysaccharide with NDST2 for one hour in the absence of PAPS followed by an additional hour after addition of the sulfate donor. The additional hour in the presence of PAPS only marginally lowered the amount of N-unsubstituted residues. Some sulfation had however occurred. This suggests that preformed N-deacetylated residues are not primary targets for sulfation by NDST2. Instead, N-sulfation seemed to occur in uninterrupted, unmodified regions of extended length.
Taken together, these results suggest that the NDST enzymes act in a processive manner and that the formation of extended NS-domains may be ascribed to mechanims inherent to the NDST enzymes without the involve-ment of other proteins. Proper regulation of domain structure, however, is dependent also on other factors.

29
Future perspectives The finding that NDST works in a processive manner raises further ques-
tions. Does the enzyme cling to the polysaccharide chain after attachment, working its way along the chain, or does the enzyme, after releasing the sub-strate, reattach to the chain with higher preference for GlcNAc residues ad-jacent to already sulfated ones? Since the process probably is directional it would be of interest to determine whether the enzyme works from the reduc-ing or the non-reducing end of the HS chain. This would give an important clue to how the HS biosynthesis machinery is organized, i.e. how the indi-vidual polymerization and modification enzymes are co-coordinated.
Brachymorphic mice, which are defective in PAPS synthetase-2 and thus produce lower amounts of PAPS, show defects in postnatal skeletal devel-opment and proteoglycans in the cartilage extracellular matrix are undersul-fated (Lane and Dickie, 1968; Orkin et al., 1976; Kurima et al., 1998; ul Haque et al., 1998). Since chondroitin sulfate is abundant in cartilage, the skeletal phenotype is probably mostly depending on alterations in chondro-itin sulfate, rather than heparan sulfate, structure. However, by studying the structure of HS isolated from such mice, which has not previously been done, we hope to obtain interesting results regarding the influence of altered PAPS concentrations on HS biosynthesis. The relative abundances of N-, 2-O-, and 6-O-sulfates could indicate whether the different HS sulfotrans-ferases are equally sensitive to lowered PAPS concentrations in an in vivo system. The domain structure should also be analyzed and compared with our in vitro data.
Also other animal models could be of interest for studying PAPS influ-ence on HS structure. For instance, the zebrafish mutant pinscher, which is lacking a PAPS-transporter, could be used for this purpose. It would also be beneficial to study PAPS-effects in C. elegans, since chondroitin sulfate from this organism appears to be devoid of sulfate groups (Yamada et al., 1999; Toyoda et al., 2000), and would thus not interfere with HS sulfation.
Paper II - Heparan sulfate biosynthesis: Characterization of an NDST1 splice variant Results
In this project we characterized a splice variant of NDST1, NDST1S. The mRNA is transcribed from the first five exons of the NDST1 gene and a unique exon, 5b, present in the intron between exon 5 and exon 6 in the NDST1 gene. NDST1S cDNA clones were originally identified in a mouse urinary bladder cDNA library and were later also isolated as mouse embryo (E11.5) EST-clones. By RT-PCR we confirmed the expression of NDST1S

30
in urinary bladder as well as in other tissues of both adult mice and E18.5 embryos.
NDST1S contains the first 479 amino acids of the full-length NDST1 pro-tein followed by a stretch of 16 additional amino acid residues unique to the splice variant. NDST1S is devoid of N-sulfotransferase activity since it lacks the C-terminal half of the full-length protein where this domain is located. The truncated protein could however potentially contain N-deacetylase activ-ity. To test this, we cloned NDST1S and expressed it in HEK-293 cells. Un-expectedly, the cell extract showed decreased rather than increased N-deacetylase activity. Thus, although the spliced version appears to be devoid of enzyme activity, it can apparently influence the action of endogenously expressed full-length NDST1.
Since endogenous levels of NDST1 are too low to detect by immunostain-ing, we co-expressed the truncated and the full-length versions of NDST1 in HEK-293 cells. An inverse relationship of their expression levels was found in these cells, as shown by Western blotting. When NDST1S was present at high levels, expression of NDST1 was low. The decreased N-deacetylase activity levels seen in cells overexpressing NDST1S alone may thus be ex-plained by inhibition of NDST1 expression. The mechanism behind this interference remains to be elucidated.
Stable clones of cells over-expressing NDST1S alone were analyzed for protein levels by Western blotting and HS structure by RPIP-HPLC. The level of total N-sulfation was similar in all clones independent of NDST1S expression levels. Correspondingly, mice which lack one NDST1 allele and are completely deficient of NDST2 produce heparan sulfate structurally identical to HS from wildtype mice although the enzyme activity is reduced by 70% (Ledin et al., 2006). On the other hand, 6-O-sulfation was greatly increased and 2-O-sulfation slightly decreased in cells expressing high levels of NDST1S. Tentatively, the GAGosome model (explained in the Chapter “Regulation of HS structure”) previously proposed for HS biosynthesis can explain this phenomenon. When present at high concentrations, NDST1S may displace other HS biosynthesis enzymes and change the composition of the GAGosome protein complex, resulting in altered HS structure.
Future perspectives The present results indicating that NDST1S is devoid of enzyme activity
are indirect and should be established by measuring enzyme activity of puri-fied protein.
It would be of particular interest to look for the expression of NDST1S during embryonic stages since changes in HS structure must be regulated both temporally and spatially during this time, and expression of NDST1S may be a rapid way to downregulate N-sulfation. Both RT-PCR, in situ hy-bridization and immunohistochemistry methods could be used for this pur-pose.

31
Splicing may be a general way to regulate HS biosynthesis enzymes and it would therefore be worthwhile to investigate if also transcripts of the other modification enzymes are alternatively spliced.
Paper III - Heparan sulfate biosynthesis in zebrafish: Five NDST genes with distinct expression patterns during embryonic development Five NDST genes in zebrafish
Since NDST is regarded a key enzyme in HS biosynthesis, and regulation of its expression possibly is an important factor in determining HS structure, we set out to complete the map of zebrafish HS biosynthesis enzymes by cloning the NDSTs in zebrafish. Through searching the Ensembl database using mouse NDST sequences as templates we found five zebrafish orthologues. When mouse, human and zebrafish NDSTs were aligned, two of the zebrafish sequences were found to be most similar to NDST1 and thus named NDST1a and NDST1b, whereas yet two others clustered with NDST2 and were named NDST2a and NDST2b. The presence of duplicate ortologues to mammalian genes is not uncommon in zebrafish, since a whole-genome duplication event took place after the divergence of teleost and mammalian ancestors (Jaillon et al., 2004). The fifth zebrafish NDST was called NDST3/4 as it is equally similar to mouse NDST3 and NDST4. The fact that mammalian NDST3 and NDST4 are equally similar to a single zebrafish orthologue suggests that these forms have arisen from a gene du-plication event in the mammalian lineage after the divergence from teleosts and since then have probably acquired more specified functions.
All five zebrafish NDSTs have similar structures as previously character-ized NDSTs, a short cytoplasmic part followed by a hydrophobic transmem-brane domain and a large catalytic part containing the N-deacetylase and N-sulfotransferase domains. The PAPS-binding sites in the N-sulfotransferase domain, as well as a lysine residue (corresponding to position 614 in human NDST1) shown to be important for N-sulfotransferase activity, are con-served in all zebrafish orthologues. Also the N-deacetylase domain is highly conserved.
mRNA expression of the zebrafish NDSTs in distinct patterns To study the expression of the different isoforms during zebrafish em-
bryogenesis we performed in situ hybridization on embryos from different developmental stages. Earlier studies using Northern blotting and RT-PCR on mouse tissues have shown that both NDST1 and NDST2 are ubiquitously expressed in adult animals (Kusche-Gullberg et al., 1998; Aikawa et al., 2001; Pallerla et al., 2008). We were therefore expecting the zebrafish

32
orthologues of these isoforms to be expressed more or less in the entire em-bryo. Surprisingly, they were instead expressed in very distinct patterns of minor structures. All isoforms were located to the brain whereas only NDST1b and NDST3/4 were also found in the tail, more specifically in the somites and the spinal chord, respectively.
The different variants of the same isoform, i.e. NDST1a/NDST1b and NDST2a/NDST2b, are expressed differently, suggesting that after the ge-nome duplication they have individually acquired more specified functions in comparison to the mammalian version.
HS structure does not vary much globally during zebrafish embryogenesis
We also analyzed the structure of HS from zebrafish at different devel-opmental stages. Interestingly, overall HS fine structure does not seem to change much during zebrafish embryogenesis. This implies that structural variations due to regulated expression of NDSTs and other HS modifying enzymes are more likely to be a highly spatially restricted phenomenon. Such local changes in HS structure would not be detected when analyzing whole embryos but will need more refined analysis of dissections. The in situ hybridization results suggest that expression of NDSTs, as well as other HS modifying enzymes, in the central nervous system is highly regulated throughout embryogenesis. In the brain, very specific expression patterns were seen for the different enzymes and isoforms also at later stages, whereas only very few enzymes were strongly expressed in the tail at 48 hours postfertilization. Since the tail makes up a large proportion of the ani-mal at later stages, putative local structural variations in the brain might be drowning in the overall signal from the whole fish at the stages analyzed.
Future perspectives By just looking at mRNA expression patterns it is difficult to draw con-
clusions of the functional contributions of the individual NDST isoforms. It will therefore be of great importance to study the effects from elimination of the enzymes, which can be done by using the morpholino knockdown tech-nique. It will be of interest also to express different NDST isoforms ectopi-cally. HS isolated from zebrafish can be analyzed structurally by RPIP-HPLC, and, as proven by pilot experiments, it is also possible to measure both N-deacetylase and N-sulfotransferase activities in zebrafish. Thus, em-bryo morphology, NDST activity and HS structure can be analyzed in the morpholino-injected and overexpressing fish. In this way, the relation be-tween enzymatic activity and HS structure, can be compared in an in vivo system.

33
Concluding remarks
Although in most cases the exact HS sequence for promotion of HS-protein interactions appears to be of less importance than previously be-lieved, HS structure must be strictly regulated to function in a proper way. The HS biosynthesis machinery is complex and there is so far no satisfactory model for describing how the individual polymerization and modification enzymes are coordinated. HS biosynthesis has traditionally been looked upon as a very hierarchical process, with the enzymes involved acting in a sequential manner, but during recent years different studies have indicated that synthesis may occur less hierarchically. N-Sulfation, for example, has long been considered an absolute requirement for further modification reac-tions. HS from mouse embryonic stem cells lacking NDST enzymes does contain 6-O-sulfate groups, however, demonstrating that 6-O-sulfation can occur without prior N-sulfation (Holmborn et al., 2004). Moreover, findings from mutant mice show that manipulation of “downstream” enzymes actu-ally influence “upstream” reactions (Merry et al., 2001; Li et al., 2003). In a model where the enzymes are strictly separated, as to result in a spatially predicted hierarchical biosynthesis machinery, this would hardly be possible. The GAGosome model would, however, explain this more satisfactorily. By changing the balance of the enzymes involved, the GAGosome composition and the resulting HS structure would also change. But what is regulating the constitution of such GAGosomes under normal conditions? Is it merely a question of abundance of the enzymes in the Golgi apparatus at a certain time point? What factors regulate the transcription and translation of these enzymes? Does posttranslational regulation occur? And, what are the roles of the many different isoforms? Speculatively, also inactive splice forms and splice variants with other kinetics or substrate specificities than the full-length protein could interfere with the GAGosome structure, resulting in changed HS design.
The finding that PAPS appears to influence the mode of NDST action fur-ther adds to the complexity. Can the substrate affect also other HS sul-fotransferases? What is the in vivo relevance of this finding? How are PAPS-concentrations proper of HS structure for a certain time point or cell type controlled?
Until we manage to place more pieces of the big HS puzzle, the question of HS structure regulation remains enigmatic, but worthwhile to explore further, perhaps in zebrafish?

34
Populärvetenskaplig sammanfattning
Heparansulfat (HS) är långa sockerkedjor som fästade till proteiner tillverkas av kroppens alla celler. En del av dessa återfinns inuti cellerna, andra sitter fast i cellmembranen medan några transporteras ut ur cellerna (Figur 1). Genom att sulfatgrupper sätts fast på sockerkedjorna vid tillverkningen av HS blir heparansulfatkedjorna negativt laddade (Figur 2). Eftersom sulfat-grupperna sprids ut ojämnt längs HS-kedjan bildas olika mönster av laddade regioner. Mönstrets utseende och laddningens storlek gör att olika molekyler kan binda till HS-kedjan. Genom att studera vad som händer om man förstör sulfatmönstrets utseende eller helt stoppar tillverkningen av heparansulfat i möss och andra djur har man kunnat visa att HS är viktigt redan mycket ti-digt under fosterutvecklingen. Dessutom har en del sjukdomar hos männi-skor visats bero på defekt produktion eller distribution av heparansulfat.
Sulfateringen av heparansulfat, dvs då kedjans negativt laddade mönster bildas, sker genom samverkan av många olika enzymer. Ett av dessa enzy-mer är N-deacetylas/N-sulfotransferas, förkortat NDST, vilket är det första enzymet som sätter fast sulfatgrupper på sockerkedjan. Eftersom de andra enzymerna kan fästa sulfatgrupper bara till redan sulfaterade enheter, är NDST mycket viktigt för den slutliga designen av sockerkedjan. Innan NDST sätter fast en sulfatgrupp avlägsnar enzymet en acetylgrupp från sockermolekylen det ska sulfatera. NDST är alltså ett enzym med två funk-tioner, deacetylering och sulfatering. Det finns fyra stycken olika varianter av NDST, NDST1- NDST4, som skiljer sig något vad gäller struktur och funktion.
Man har visat att heparansulfat-strukturen skiljer sig åt mellan kroppens olika vävnader men är väldigt lik när man jämför samma vävnad från olika individer. Detta tyder på att sulfateringen av heparansulfat är strikt reglerad. I motsats till proteinproduktion, som styrs av DNA i cellerna, finns ingen mall för hur den färdiga HS-kedjan ska se ut och idag vet man väldigt lite om hur designen av heparansulfat är kontrollerad. Man tror att de relativa koncentrationerna av de olika enzymen som sulfaterar kedjan kan påverka dess slutgiltiga mönster.
Genom sin diversitet spelar heparansulfat många olika roller i biologiska processer (Figur 3) och regleringen av heparansulfat-designen är viktig för att dessa processer ska fungera på ett riktigt sätt. För att få mer kunskap om denna reglering har vi i tre projekt studerat NDST-enzymet på olika sätt. I det första projektet har vi tittat på hur den molekyl som bidrar med sulfat-

35
gruppen vid sulfateringsreaktionen, PAPS, påverkar NDST-funktionen. Om man utesluter PAPS vid reaktionen angrips heparansulfatet tillsynes slump-mässigt, lite här och var längs kedjan, av enzymet. I närvaro av PAPS verkar däremot NDST-enzymet vandra utmed sockerkedjan och sulfatera denna i långa, oavbrutna sträckor. Vid PAPS-tillsatser av olika koncentrationer fås också olika sulfateringsmönster. Reglering av PAPS-koncentration föreslås därför vara ett möjligt sätt att reglera heparansulfat-struktur.
I ett andra projekt studerades en kortare version av NDST1-proteinet, NDST1S. Denna version saknar den del av enzymet som ansvarar för sulfa-teringen, men kan eventuellt utföra deacetyleringsreaktionen. När man över-uttrycker NDST1S i celler, dvs gör så att cellerna producerar mer av protei-net, påverkas även produktionen av det långa NDST1-enzymet. En hypotes som tidigare framlagts är att de enzymer som ansvarar för heparansulfatpro-duktion sitter tillsammans i ett stort proteinkomplex och att sammansättning-en av detta komplex beror av hur mycket av de respektive enzymerna som cellen producerar. Genom att påverka mängden NDST1 skulle alltså NDST1S kunna delta i regleringen av HS-sulfatering.
För att studera enzymers funktioner i organismer, och inte enbart deras beteenden i provrör eller i cellkulturer, krävs djurstudier. Eftersom den tidiga embryoutvecklingen är ungefär likadan i många djur kan man genom att studera andra organismer få ledtrådar till funktioner även hos människa. Zebrafisk har börjat användas en hel del på senare tid eftersom denna djur-modell har många fördelar. Till exempel sker befruktningen utanför kroppen och embryon behövs därför inte opereras ut för att studeras. Äggen är dess-utom genomskinliga och man kan således studera embryoutvecklingen från befruktning till färdigutvecklad fisk genom att använda ett vanligt mikro-skop. I det tredje projektet har vi studerat olika NDST-enzymer i just zebra-fisk. Vi visade att zebrafisk har fem stycken olika NDST-enzymer istället för däggdjurens fyra. Mängden av de olika enzymerna skiljer sig åt mellan olika delar av zebrafisken, vilket troligtvis tyder på att de är olika viktiga i olika vävnader.
Genom dessa projekt har vi visat på tidigare icke kända aspekter avseende heparansulfats reglering. Fortfarande återstår dock att finna fler ledtrådar för att fullständigt förstå hur produktionen och regleringen av denna komplexa struktur fungerar.

36
Acknowledgment
I want to thank all the people who, in different ways and at different times, have taken part in the work that lies behind this thesis. Both those who have contributed in the scientific process, by helping out in the lab, sharing ideas and discussing scientific issues, and also the ones that have taken my mind off work from time to time. Lena, en förebild på många sätt. Tack för att du alltid tar dig tid, oavsett hur stressigt du har det. Tack för alla idéer, all peppning och allt stöd. Och inte minst för att du är en så trygg och bra person. D9:4, During my very many years in this place sooo many great people have passed through. You are just too many to mention by name so I will simply settle for a GREAT THANKS to you all!! Some names should however be mentioned: Ulf, tack för stöd, entusiasm och synpunkter kring mina projekt Marion, tack för att du ställt upp som biträdande handledare Dorothe, som har bidragit med många tankar, idéer och förslag till min forskning, tack för all hjälp Kjelléngruppen: Katarina, Anders, Maria R, Maria T, Mehdi, Håkan och Maria W. Särskilt tack till Inger, för att du håller koll på oss alla valpar i labbet. Och allt annat också för den delen, du är en klippa! Tack också för alla intressanta samtal om stort och smått. Jenny och Johan, mina roligaste perioder under denna tid har varit vid mina båda intensiva samarbeten med er, stort tack för det!! Anne, min tryckkokarventil genom doktorandåren. Tack! Other IMBIM-colleagues, researchers, students, and administrative staff, with whom I have teached, chatted in the lunchroom, travelled to meetings, and had a lot of fun! The SLU-gang in the Red Sofa (my second home at BMC), past and pre-sent members, for always being so helpful, friendly and full of fun! Everyone at Evolutionary Organism Biology, EBC, for letting me sneak around in your corridors and drink your coffee Carl Neumann group, EMBL Heidelberg, for friendly welcoming me during summer 2007 at the start of the zebrafish project GLIBS, for having arranged so many interesting and fun research meetings and also made travels to international meetings possible

37
Flicksen, ni e ju så goa! Tack också till alla mini-flicks & -boys och deras fäder i Flicksen Fanclub Ullis, för all positiv energi du ger, genom tankar, prat, dans och musik. Tack också till Mikael. Och Majken & Love såklart…. Lousie, för allt kul på Trastvägen och även efteråt. Hoppas vi får chans att slåss snart igen… Eva-Lotta, min musketörsyster under den senaste tiden. Delad ångest är dubbel ångest, eller hur var det nu?! Carpe Diem ;-) Tack också till Akka som fått mig att upptäcka nya underbara Uppsalaställen Ronak, för att du är så vis och klok och ändå härligt galen Jenny, för att du alltid har ett öra att lyssna med och en axel att luta mig mot Josefin, Johanna, Lena, Gunnar, Claes, Micke, Erik, Katrin och övriga Uppsalavänner, för allt skoj genom åren, hoppas jag kan ha tid att fixa fler kubbkvällar m.m. nu Familjen Nord, för alla middagar jag snyltat till mig. Ni är min lilla vardagstillflykt, glad att ni finns Mina Granada/Stockholmsvänner, vi träffas tyvärr inte så ofta men att ni finns kvar i mitt liv efter alla år betyder mycket Willy, Anita och Anna, tack för oerhört många roliga resor och upplevelser tillsammans IM-gänget, som gör så gott vi kan och mäktar med… Tack för Cappuccino-fikor och roliga evenemang genom åren IM, för att ni gör skillnad! (www.manniskohjalp.se) Farmor och Farfar, som ändå finns med Mormor, som jag haft mycket roligt med, särskilt som litet pyre. Stor kram till Dig! Anders och Barbro, för att ni tror att jag kan klara det mesta Per och Nina, för att jag alltid är välkommen och för alla goda råd Lisa, Du Solstråle och Glädjekälla Lilla BrorSyster, som jag längtar efter att lära känna även Dig! Mamma och Pappa, som fått utstå mycket gnäll och negativt tjat under denna tid, ett särskilt stort tack till er! Tack för att ni älskar mig, inte för vad jag gör, utan för vem jag är Thanks also those of you who are not mentioned, but in one or another way, at one or another time, have taken part in my life. You have all contributed, made me the one I am, and brought me to where I find myself today.

38
References
Ahmad M, Haque MF, Ahmad W, Abbas H, Haque S, Krakow D, Rimoin DL, Lachman RS, Cohn DH. 1998. Distinct, autosomal recessive form of spondyloepimetaphyseal dysplasia segregating in an inbred Pakistani kindred. Am J Med Genet 78:468-473.
Aikawa J, Esko JD. 1999. Molecular cloning and expression of a third mem-ber of the heparan sulfate/heparin GlcNAc N-deacetylase/ N-sulfotransferase family. J Biol Chem 274:2690-2695.
Aikawa J, Grobe K, Tsujimoto M, Esko JD. 2001. Multiple isozymes of heparan sulfate/heparin GlcNAc N-deacetylase/GlcN N-sulfotransferase. Structure and activity of the fourth member, NDST4. J Biol Chem 276:5876-5882.
Alexopoulou AN, Multhaupt HA, Couchman JR. 2007. Syndecans in wound healing, inflammation and vascular biology. Int J Biochem Cell Biol 39:505-528.
Allen BL, Rapraeger AC. 2003. Spatial and temporal expression of heparan sulfate in mouse development regulates FGF and FGF receptor as-sembly. J Cell Biol 163:637-648.
Berninsone P, Hirschberg CB. 1998. Heparan sulfate/heparin N-deacetylase/N-sulfotransferase. The N-sulfotransferase activity do-main is at the carboxyl half of the holoenzyme. J Biol Chem 273:25556-25559.
Bezakova G, Ruegg MA. 2003. New insights into the roles of agrin. Nat Rev Mol Cell Biol 4:295-308.
Bink RJ, Habuchi H, Lele Z, Dolk E, Joore J, Rauch GJ, Geisler R, Wilson SW, den Hertog J, Kimata K, Zivkovic D. 2003. Heparan sulfate 6-o-sulfotransferase is essential for muscle development in zebrafish. J Biol Chem 278:31118-31127.
Bishop JR, Schuksz M, Esko JD. 2007. Heparan sulphate proteoglycans fine-tune mammalian physiology. Nature 446:1030-1037.
Bishop JR, Stanford KI, Esko JD. 2008. Heparan sulfate proteoglycans and triglyceride-rich lipoprotein metabolism. Curr Opin Lipidol 19:307-313.
Brickman YG, Ford MD, Gallagher JT, Nurcombe V, Bartlett PF, Turnbull JE. 1998. Structural modification of fibroblast growth factor-binding heparan sulfate at a determinative stage of neural development. J Biol Chem 273:4350-4359.
Bullock SL, Fletcher JM, Beddington RS, Wilson VA. 1998. Renal agenesis in mice homozygous for a gene trap mutation in the gene encoding heparan sulfate 2-sulfotransferase. Genes Dev 12:1894-1906.

39
Bulow HE, Berry KL, Topper LH, Peles E, Hobert O. 2002. Heparan sulfate proteoglycan-dependent induction of axon branching and axon mis-routing by the Kallmann syndrome gene kal-1. Proc Natl Acad Sci U S A 99:6346-6351.
Bulow HE, Hobert O. 2004. Differential sulfations and epimerization define heparan sulfate specificity in nervous system development. Neuron 41:723-736.
Cadwallader AB, Yost HJ. 2006a. Combinatorial expression patterns of heparan sulfate sulfotransferases in zebrafish: I. The 3-O-sulfotransferase family. Dev Dyn 235:3423-3431.
Cadwallader AB, Yost HJ. 2006b. Combinatorial expression patterns of heparan sulfate sulfotransferases in zebrafish: II. The 6-O-sulfotransferase family. Dev Dyn 235:3432-3437.
Cadwallader AB, Yost HJ. 2007. Combinatorial expression patterns of heparan sulfate sulfotransferases in zebrafish: III. 2-O-sulfotransferase and C5-epimerases. Dev Dyn 236:581-586.
Cano-Gauci DF, Song HH, Yang H, McKerlie C, Choo B, Shi W, Pullano R, Piscione TD, Grisaru S, Soon S, Sedlackova L, Tanswell AK, Mak TW, Yeger H, Lockwood GA, Rosenblum ND, Filmus J. 1999. Glypican-3-deficient mice exhibit developmental overgrowth and some of the abnormalities typical of Simpson-Golabi-Behmel syn-drome. J Cell Biol 146:255-264.
Chen E, Hermanson S, Ekker SC. 2004. Syndecan-2 is essential for angio-genic sprouting during zebrafish development. Blood 103:1710-1719.
Chen E, Stringer SE, Rusch MA, Selleck SB, Ekker SC. 2005. A unique role for 6-O sulfation modification in zebrafish vascular development. Dev Biol 284:364-376.
Clement A, Wiweger M, von der Hardt S, Rusch MA, Selleck SB, Chien CB, Roehl HH. 2008. Regulation of zebrafish skeletogenesis by ext2/dackel and papst1/pinscher. PLoS Genet 4:e1000136.
David G, Bai XM, Van der Schueren B, Cassiman JJ, Van den Berghe H. 1992. Developmental changes in heparan sulfate expression: in situ detection with mAbs. J Cell Biol 119:961-975.
Dejima K, Seko A, Yamashita K, Gengyo-Ando K, Mitani S, Izumikawa T, Kitagawa H, Sugahara K, Mizuguchi S, Nomura K. 2006. Essential roles of 3'-phosphoadenosine 5'-phosphosulfate synthase in embry-onic and larval development of the nematode Caenorhabditis ele-gans. J Biol Chem 281:11431-11440.
Duncan MB, Liu M, Fox C, Liu J. 2006. Characterization of the N-deacetylase domain from the heparan sulfate N-deacetylase/N-sulfotransferase 2. Biochem Biophys Res Commun 339:1232-1237.
Eisen JS, Smith JC. 2008. Controlling morpholino experiments: don't stop making antisense. Development 135:1735-1743.
Eriksson I, Sandback D, Ek B, Lindahl U, Kjellen L. 1994. cDNA cloning and sequencing of mouse mastocytoma glucosaminyl N-

40
deacetylase/N-sulfotransferase, an enzyme involved in the biosyn-thesis of heparin. J Biol Chem 269:10438-10443.
Esko JD, Lindahl U. 2001. Molecular diversity of heparan sulfate. J Clin Invest 108:169-173.
Esko JD, Selleck SB. 2002. Order out of chaos: assembly of ligand binding sites in heparan sulfate. Annu Rev Biochem 71:435-471.
Fan G, Xiao L, Cheng L, Wang X, Sun B, Hu G. 2000. Targeted disruption of NDST-1 gene leads to pulmonary hypoplasia and neonatal respi-ratory distress in mice. FEBS Lett 467:7-11.
Filmus J, Selleck SB. 2001. Glypicans: proteoglycans with a surprise. J Clin Invest 108:497-501.
Forsberg E, Kjellen L. 2001. Heparan sulfate: lessons from knockout mice. J Clin Invest 108:175-180.
Forsberg E, Pejler G, Ringvall M, Lunderius C, Tomasini-Johansson B, Ku-sche-Gullberg M, Eriksson I, Ledin J, Hellman L, Kjellen L. 1999. Abnormal mast cells in mice deficient in a heparin-synthesizing en-zyme. Nature 400:773-776.
Fuda H, Shimizu C, Lee YC, Akita H, Strott CA. 2002. Characterization and expression of human bifunctional 3'-phosphoadenosine 5'-phosphosulphate synthase isoforms. Biochem J 365:497-504.
Gallagher JT. 2001. Heparan sulfate: growth control with a restricted se-quence menu. J Clin Invest 108:357-361.
Ghiselli G, Farber SA. 2005. D-glucuronyl C5-epimerase acts in dorso-ventral axis formation in zebrafish. BMC Dev Biol 5:19.
Goda E, Kamiyama S, Uno T, Yoshida H, Ueyama M, Kinoshita-Toyoda A, Toyoda H, Ueda R, Nishihara S. 2006. Identification and characteri-zation of a novel Drosophila 3'-phosphoadenosine 5'-phosphosulfate transporter. J Biol Chem 281:28508-28517.
Grobe K, Esko JD. 2002. Regulated translation of heparan sulfate N-acetylglucosamine N-deacetylase/n-sulfotransferase isozymes by structured 5'-untranslated regions and internal ribosome entry sites. J Biol Chem 277:30699-30706.
Grobe K, Inatani M, Pallerla SR, Castagnola J, Yamaguchi Y, Esko JD. 2005. Cerebral hypoplasia and craniofacial defects in mice lacking heparan sulfate Ndst1 gene function. Development 132:3777-3786.
Grobe K, Ledin J, Ringvall M, Holmborn K, Forsberg E, Esko JD, Kjellen L. 2002. Heparan sulfate and development: differential roles of the N-acetylglucosamine N-deacetylase/N-sulfotransferase isozymes. Biochim Biophys Acta 1573:209-215.
Habuchi H, Nagai N, Sugaya N, Atsumi F, Stevens RL, Kimata K. 2007. Mice deficient in heparan sulfate 6-O-sulfotransferase-1 exhibit de-fective heparan sulfate biosynthesis, abnormal placentation, and late embryonic lethality. J Biol Chem 282:15578-15588.
Hacker U, Nybakken K, Perrimon N. 2005. Heparan sulphate proteoglycans: the sweet side of development. Nat Rev Mol Cell Biol 6:530-541.

41
Hashimoto Y, Orellana A, Gil G, Hirschberg CB. 1992. Molecular cloning and expression of rat liver N-heparan sulfate sulfotransferase. J Biol Chem 267:15744-15750.
Henningsson F, Ledin J, Lunderius C, Wilen M, Hellman L, Pejler G. 2002. Altered storage of proteases in mast cells from mice lacking heparin: a possible role for heparin in carboxypeptidase A processing. Biol Chem 383:793-801.
Holmborn K, Ledin J, Smeds E, Eriksson I, Kusche-Gullberg M, Kjellen L. 2004. Heparan sulfate synthesized by mouse embryonic stem cells deficient in NDST1 and NDST2 is 6-O-sulfated but contains no N-sulfate groups. J Biol Chem.
Humphries DE, Wong GW, Friend DS, Gurish MF, Qiu WT, Huang C, Sharpe AH, Stevens RL. 1999. Heparin is essential for the storage of specific granule proteases in mast cells. Nature 400:769-772.
Inatani M, Irie F, Plump AS, Tessier-Lavigne M, Yamaguchi Y. 2003. Mammalian brain morphogenesis and midline axon guidance require heparan sulfate. Science 302:1044-1046.
Iozzo RV. 2005. Basement membrane proteoglycans: from cellar to ceiling. Nat Rev Mol Cell Biol 6:646-656.
Jaillon O, Aury JM, Brunet F, Petit JL, Stange-Thomann N, Mauceli E, Bouneau L, Fischer C, Ozouf-Costaz C, Bernot A, Nicaud S, Jaffe D, Fisher S, Lutfalla G, Dossat C, Segurens B, Dasilva C, Salanou-bat M, Levy M, Boudet N, Castellano S, Anthouard V, Jubin C, Cas-telli V, Katinka M, Vacherie B, Biemont C, Skalli Z, Cattolico L, Poulain J, De Berardinis V, Cruaud C, Duprat S, Brottier P, Cou-tanceau JP, Gouzy J, Parra G, Lardier G, Chapple C, McKernan KJ, McEwan P, Bosak S, Kellis M, Volff JN, Guigo R, Zody MC, Mesi-rov J, Lindblad-Toh K, Birren B, Nusbaum C, Kahn D, Robinson-Rechavi M, Laudet V, Schachter V, Quetier F, Saurin W, Scarpelli C, Wincker P, Lander ES, Weissenbach J, Roest Crollius H. 2004. Genome duplication in the teleost fish Tetraodon nigroviridis reveals the early vertebrate proto-karyotype. Nature 431:946-957.
Jastrebova N, Vanwildemeersch M, Rapraeger AC, Gimenez-Gallego G, Lindahl U, Spillmann D. 2006. Heparan sulfate-related oligosaccha-rides in ternary complex formation with fibroblast growth factors 1 and 2 and their receptors. J Biol Chem 281:26884-26892.
Jenniskens GJ, Hafmans T, Veerkamp JH, van Kuppevelt TH. 2002. Spatio-temporal distribution of heparan sulfate epitopes during myogenesis and synaptogenesis: a study in developing mouse intercostal muscle. Dev Dyn 225:70-79.
Jenniskens GJ, Oosterhof A, Brandwijk R, Veerkamp JH, van Kuppevelt TH. 2000. Heparan sulfate heterogeneity in skeletal muscle basal lamina: demonstration by phage display-derived antibodies. J Neu-rosci 20:4099-4111.
Jullien D, Crozatier M, Kas E. 1997. cDNA sequence and expression pattern of the Drosophila melanogaster PAPS synthetase gene: a new sali-vary gland marker. Mech Dev 68:179-186.

42
Kakuta Y, Sueyoshi T, Negishi M, Pedersen LC. 1999. Crystal structure of the sulfotransferase domain of human heparan sulfate N-deacetylase/ N-sulfotransferase 1. J Biol Chem 274:10673-10676.
Kamiyama S, Sasaki N, Goda E, Ui-Tei K, Saigo K, Narimatsu H, Jigami Y, Kannagi R, Irimura T, Nishihara S. 2006. Molecular cloning and characterization of a novel 3'-phosphoadenosine 5'-phosphosulfate transporter, PAPST2. J Biol Chem 281:10945-10953.
Kamiyama S, Suda T, Ueda R, Suzuki M, Okubo R, Kikuchi N, Chiba Y, Goto S, Toyoda H, Saigo K, Watanabe M, Narimatsu H, Jigami Y, Nishihara S. 2003. Molecular cloning and identification of 3'-phosphoadenosine 5'-phosphosulfate transporter. J Biol Chem 278:25958-25963.
Karlstrom RO, Trowe T, Klostermann S, Baier H, Brand M, Crawford AD, Grunewald B, Haffter P, Hoffmann H, Meyer SU, Muller BK, Rich-ter S, van Eeden FJ, Nusslein-Volhard C, Bonhoeffer F. 1996. Ze-brafish mutations affecting retinotectal axon pathfinding. Develop-ment 123:427-438.
Kinnunen T, Huang Z, Townsend J, Gatdula MM, Brown JR, Esko JD, Turnbull JE. 2005. Heparan 2-O-sulfotransferase, hst-2, is essential for normal cell migration in Caenorhabditis elegans. Proc Natl Acad Sci U S A 102:1507-1512.
Kobayashi S, Morimoto K, Shimizu T, Takahashi M, Kurosawa H, Shirasawa T. 2000. Association of EXT1 and EXT2, hereditary mul-tiple exostoses gene products, in Golgi apparatus. Biochem Biophys Res Commun 268:860-867.
Kolset SO, Prydz K, Pejler G. 2004. Intracellular proteoglycans. Biochem J 379:217-227.
Kreuger J, Jemth P, Sanders-Lindberg E, Eliahu L, Ron D, Basilico C, Salmivirta M, Lindahl U. 2005. Fibroblast growth factors share binding sites in heparan sulphate. Biochem J 389:145-150.
Kreuger J, Matsumoto T, Vanwildemeersch M, Sasaki T, Timpl R, Claes-son-Welsh L, Spillmann D, Lindahl U. 2002. Role of heparan sulfate domain organization in endostatin inhibition of endothelial cell func-tion. Embo J 21:6303-6311.
Kurima K, Warman ML, Krishnan S, Domowicz M, Krueger RC, Jr., Deyrup A, Schwartz NB. 1998. A member of a family of sulfate-activating enzymes causes murine brachymorphism. Proc Natl Acad Sci U S A 95:8681-8685.
Kusche-Gullberg M, Eriksson I, Pikas DS, Kjellen L. 1998. Identification and expression in mouse of two heparan sulfate glucosaminyl N-deacetylase/N-sulfotransferase genes. J Biol Chem 273:11902-11907.
Lamanna WC, Kalus I, Padva M, Baldwin RJ, Merry CL, Dierks T. 2007. The heparanome--the enigma of encoding and decoding heparan sul-fate sulfation. J Biotechnol 129:290-307.
Lander AD. 1998. Proteoglycans: master regulators of molecular encounter? Matrix Biol 17:465-472.

43
Lane PW, Dickie MM. 1968. Three recessive mutations producing dispro-portionate dwarfing in mice: achondroplasia, brachymorphic, and stubby. J Hered 59:300-308.
Ledin J, Ringvall M, Thuveson M, Eriksson I, Wilen M, Kusche-Gullberg M, Forsberg E, Kjellen L. 2006. Enzymatically active N-deacetylase/N-sulfotransferase-2 is present in liver but does not con-tribute to heparan sulfate N-sulfation. J Biol Chem.
Ledin J, Staatz W, Li JP, Gotte M, Selleck S, Kjellen L, Spillmann D. 2004. Heparan sulfate structure in mice with genetically modified heparan sulfate production. J Biol Chem.
Lee JS, von der Hardt S, Rusch MA, Stringer SE, Stickney HL, Talbot WS, Geisler R, Nusslein-Volhard C, Selleck SB, Chien CB, Roehl H. 2004. Axon sorting in the optic tract requires HSPG synthesis by ext2 (dackel) and extl3 (boxer). Neuron 44:947-960.
Li JP, Gong F, Hagner-McWhirter A, Forsberg E, Abrink M, Kisilevsky R, Zhang X, Lindahl U. 2003. Targeted disruption of a murine glucuro-nyl C5-epimerase gene results in heparan sulfate lacking L-iduronic acid and in neonatal lethality. J Biol Chem 278:28363-28366.
Lin X. 2004. Functions of heparan sulfate proteoglycans in cell signaling during development. Development 131:6009-6021.
Lin X, Wei G, Shi Z, Dryer L, Esko JD, Wells DE, Matzuk MM. 2000. Dis-ruption of gastrulation and heparan sulfate biosynthesis in EXT1-deficient mice. Dev Biol 224:299-311.
Lindahl B, Eriksson L, Lindahl U. 1995. Structure of heparan sulphate from human brain, with special regard to Alzheimer's disease. Biochem J 306 ( Pt 1):177-184.
Lindahl U, Lidholt K, Spillmann D, Kjellen L. 1994. More to "heparin" than anticoagulation. Thromb Res 75:1-32.
Lortat-Jacob H, Turnbull JE, Grimaud JA. 1995. Molecular organization of the interferon gamma-binding domain in heparan sulphate. Biochem J 310 ( Pt 2):497-505.
Luders F, Segawa H, Stein D, Selva EM, Perrimon N, Turco SJ, Hacker U. 2003. Slalom encodes an adenosine 3'-phosphate 5'-phosphosulfate transporter essential for development in Drosophila. Embo J 22:3635-3644.
Maccarana M, Sakura Y, Tawada A, Yoshida K, Lindahl U. 1996. Domain structure of heparan sulfates from bovine organs. J Biol Chem 271:17804-17810.
McCormick C, Duncan G, Goutsos KT, Tufaro F. 2000. The putative tumor suppressors EXT1 and EXT2 form a stable complex that accumu-lates in the Golgi apparatus and catalyzes the synthesis of heparan sulfate. Proc Natl Acad Sci U S A 97:668-673.
McCormick C, Duncan G, Tufaro F. 1999. New perspectives on the molecu-lar basis of hereditary bone tumours. Mol Med Today 5:481-486.
Merry CL, Bullock SL, Swan DC, Backen AC, Lyon M, Beddington RS, Wilson VA, Gallagher JT. 2001. The molecular phenotype of

44
heparan sulfate in the Hs2st-/- mutant mouse. J Biol Chem 276:35429-35434.
Morii E, Ogihara H, Oboki K, Sawa C, Sakuma T, Nomura S, Esko JD, Handa H, Kitamura Y. 2001. Inhibitory effect of the mi transcription factor encoded by the mutant mi allele on GA binding protein-mediated transcript expression in mouse mast cells. Blood 97:3032-3039.
Morio H, Honda Y, Toyoda H, Nakajima M, Kurosawa H, Shirasawa T. 2003. EXT gene family member rib-2 is essential for embryonic de-velopment and heparan sulfate biosynthesis in Caenorhabditis ele-gans. Biochem Biophys Res Commun 301:317-323.
Mulloy B, Forster MJ, Jones C, Davies DB. 1993. N.m.r. and molecular-modelling studies of the solution conformation of heparin. Biochem J 293 ( Pt 3):849-858.
Neri G, Gurrieri F, Zanni G, Lin A. 1998. Clinical and molecular aspects of the Simpson-Golabi-Behmel syndrome. Am J Med Genet 79:279-283.
Nusslein-Volhard C, Dahm R. 2002. Zebrafish. Oxford University Press. Nybakken K, Perrimon N. 2002. Heparan sulfate proteoglycan modulation
of developmental signaling in Drosophila. Biochim Biophys Acta 1573:280-291.
Orellana A, Hirschberg CB, Wei Z, Swiedler SJ, Ishihara M. 1994. Molecu-lar cloning and expression of a glycosaminoglycan N-acetylglucosaminyl N-deacetylase/N-sulfotransferase from a hepa-rin-producing cell line. J Biol Chem 269:2270-2276.
Orkin RW, Pratt RM, Martin GR. 1976. Undersulfated chondroitin sulfate in the cartilage matrix of brachymorphic mice. Dev Biol 50:82-94.
Ozeran JD, Westley J, Schwartz NB. 1996. Kinetics of PAPS translocase: evidence for an antiport mechanism. Biochemistry 35:3685-3694.
Pallerla SR, Lawrence R, Lewejohann L, Pan Y, Fischer T, Schlomann U, Zhang X, Esko JD, Grobe K. 2008. Altered heparan sulfate structure in mice with deleted NDST3 gene function. J Biol Chem.
Pallerla SR, Pan Y, Zhang X, Esko JD, Grobe K. 2007. Heparan sulfate Ndst1 gene function variably regulates multiple signaling pathways during mouse development. Dev Dyn 236:556-563.
Petitou M, Casu B, Lindahl U. 2003. 1976-1983, a critical period in the his-tory of heparin: the discovery of the antithrombin binding site. Bio-chimie 85:83-89.
Pikas DS, Eriksson I, Kjellen L. 2000. Overexpression of different isoforms of glucosaminyl N-deacetylase/N-sulfotransferase results in distinct heparan sulfate N-sulfation patterns. Biochemistry 39:4552-4558.
Pilia G, Hughes-Benzie RM, MacKenzie A, Baybayan P, Chen EY, Huber R, Neri G, Cao A, Forabosco A, Schlessinger D. 1996. Mutations in GPC3, a glypican gene, cause the Simpson-Golabi-Behmel over-growth syndrome. Nat Genet 12:241-247.
Pinhal MA, Smith B, Olson S, Aikawa J, Kimata K, Esko JD. 2001. Enzyme interactions in heparan sulfate biosynthesis: uronosyl 5-epimerase

45
and 2-O-sulfotransferase interact in vivo. Proc Natl Acad Sci U S A 98:12984-12989.
Pratt T, Conway CD, Tian NM, Price DJ, Mason JO. 2006. Heparan sul-phation patterns generated by specific heparan sulfotransferase en-zymes direct distinct aspects of retinal axon guidance at the optic chiasm. J Neurosci 26:6911-6923.
Presto J, Thuveson M, Carlsson P, Busse M, Wilen M, Eriksson I, Kusche-Gullberg M, Kjellen L. 2008. Heparan sulfate biosynthesis enzymes EXT1 and EXT2 affect NDST1 expression and heparan sulfate sul-fation. Proc Natl Acad Sci U S A 105:4751-4756.
Riesenfeld J, Hozok M, Lindahl U. 1982. Biosynthesis of heparan sulfate in rat liver. Characterization of polysaccharides obtained with intact cells and with a cell-free system. J Biol Chem 257:7050-7055.
Ringvall M, Ledin J, Holmborn K, van Kuppevelt T, Ellin F, Eriksson I, Olofsson AM, Kjellen L, Forsberg E. 2000. Defective heparan sul-fate biosynthesis and neonatal lethality in mice lacking N-deacetylase/N-sulfotransferase-1. J Biol Chem 275:25926-25930.
Robu ME, Larson JD, Nasevicius A, Beiraghi S, Brenner C, Farber SA, Ek-ker SC. 2007. p53 activation by knockdown technologies. PLoS Genet 3:e78.
Rostand KS, Esko JD. 1997. Microbial adherence to and invasion through proteoglycans. Infect Immun 65:1-8.
Schilling TF, Piotrowski T, Grandel H, Brand M, Heisenberg CP, Jiang YJ, Beuchle D, Hammerschmidt M, Kane DA, Mullins MC, van Eeden FJ, Kelsh RN, Furutani-Seiki M, Granato M, Haffter P, Odenthal J, Warga RM, Trowe T, Nusslein-Volhard C. 1996. Jaw and branchial arch mutants in zebrafish I: branchial arches. Development 123:329-344.
Senay C, Lind T, Muguruma K, Tone Y, Kitagawa H, Sugahara K, Lidholt K, Lindahl U, Kusche-Gullberg M. 2000. The EXT1/EXT2 tumor suppressors: catalytic activities and role in heparan sulfate biosyn-thesis. EMBO Rep 1:282-286.
Shukla D, Liu J, Blaiklock P, Shworak NW, Bai X, Esko JD, Cohen GH, Eisenberg RJ, Rosenberg RD, Spear PG. 1999. A novel role for 3-O-sulfated heparan sulfate in herpes simplex virus 1 entry. Cell 99:13-22.
Shworak NW, HajMohammadi S, De Agostini AI, Rosenberg RD. 2002. Mice deficient in heparan sulfate 3-O-sulfotransferase-1: Normal hemostasis with unexpected perinatal phenotypes. Glycoconj J 19:355-361.
Spillmann D, Witt D, Lindahl U. 1998. Defining the interleukin-8-binding domain of heparan sulfate. J Biol Chem 273:15487-15493.
Stickens D, Zak BM, Rougier N, Esko JD, Werb Z. 2005. Mice deficient in Ext2 lack heparan sulfate and develop exostoses. Development.
Stringer SE, Gallagher JT. 1997. Specific binding of the chemokine platelet factor 4 to heparan sulfate. J Biol Chem 272:20508-20514.
Strott CA. 2002. Sulfonation and molecular action. Endocr Rev 23:703-732.

46
Superti-Furga A, Hastbacka J, Rossi A, van der Harten JJ, Wilcox WR, Cohn DH, Rimoin DL, Steinmann B, Lander ES, Gitzelmann R. 1996. A family of chondrodysplasias caused by mutations in the di-astrophic dysplasia sulfate transporter gene and associated with im-paired sulfation of proteoglycans. Ann N Y Acad Sci 785:195-201.
ten Dam GB, Hafmans T, Veerkamp JH, van Kuppevelt TH. 2003. Differen-tial expression of heparan sulfate domains in rat spleen. J Histochem Cytochem 51:727-739.
Topczewski J, Sepich DS, Myers DC, Walker C, Amores A, Lele Z, Ham-merschmidt M, Postlethwait J, Solnica-Krezel L. 2001. The zebraf-ish glypican knypek controls cell polarity during gastrulation movements of convergent extension. Dev Cell 1:251-264.
Torri G, Casu B, Gatti G, Petitou M, Choay J, Jacquinet JC, Sinay P. 1985. Mono- and bidimensional 500 MHz 1H-NMR spectra of a synthetic pentasaccharide corresponding to the binding sequence of heparin to antithrombin-III: evidence for conformational peculiarity of the sul-fated iduronate residue. Biochem Biophys Res Commun 128:134-140.
Toyoda H, Kinoshita-Toyoda A, Selleck SB. 2000. Structural analysis of glycosaminoglycans in Drosophila and Caenorhabditis elegans and demonstration that tout-velu, a Drosophila gene related to EXT tu-mor suppressors, affects heparan sulfate in vivo. J Biol Chem 275:2269-2275.
Trowe T, Klostermann S, Baier H, Granato M, Crawford AD, Grunewald B, Hoffmann H, Karlstrom RO, Meyer SU, Muller B, Richter S, Nus-slein-Volhard C, Bonhoeffer F. 1996. Mutations disrupting the or-dering and topographic mapping of axons in the retinotectal projec-tion of the zebrafish, Danio rerio. Development 123:439-450.
Tumova S, Woods A, Couchman JR. 2000. Heparan sulfate proteoglycans on the cell surface: versatile coordinators of cellular functions. Int J Biochem Cell Biol 32:269-288.
ul Haque MF, King LM, Krakow D, Cantor RM, Rusiniak ME, Swank RT, Superti-Furga A, Haque S, Abbas H, Ahmad W, Ahmad M, Cohn DH. 1998. Mutations in orthologous genes in human spondy-loepimetaphyseal dysplasia and the brachymorphic mouse. Nat Genet 20:157-162.
van Eeden FJ, Granato M, Schach U, Brand M, Furutani-Seiki M, Haffter P, Hammerschmidt M, Heisenberg CP, Jiang YJ, Kane DA, Kelsh RN, Mullins MC, Odenthal J, Warga RM, Nusslein-Volhard C. 1996. Genetic analysis of fin formation in the zebrafish, Danio rerio. De-velopment 123:255-262.
van Kuppevelt TH, Dennissen MA, van Venrooij WJ, Hoet RM, Veerkamp JH. 1998. Generation and application of type-specific anti-heparan sulfate antibodies using phage display technology. Further evidence for heparan sulfate heterogeneity in the kidney. J Biol Chem 273:12960-12966.

47
Venkatachalam KV. 2003. Human 3'-phosphoadenosine 5'-phosphosulfate (PAPS) synthase: biochemistry, molecular biology and genetic defi-ciency. IUBMB Life 55:1-11.
Wang L, Fuster M, Sriramarao P, Esko JD. 2005. Endothelial heparan sul-fate deficiency impairs L-selectin- and chemokine-mediated neutro-phil trafficking during inflammatory responses. Nat Immunol 6:902-910.
Warda M, Toida T, Zhang F, Sun P, Munoz E, Xie J, Linhardt RJ. 2006. Isolation and characterization of heparan sulfate from various mur-ine tissues. Glycoconj J 23:555-563.
Wei Z, Swiedler SJ. 1999. Functional analysis of conserved cysteines in heparan sulfate N-deacetylase-N-sulfotransferases. J Biol Chem 274:1966-1970.
Westling C, Lindahl U. 2002. Location of N-unsubstituted glucosamine resi-dues in heparan sulfate. J Biol Chem 277:49247-49255.
Yamada S, Van Die I, Van den Eijnden DH, Yokota A, Kitagawa H, Suga-hara K. 1999. Demonstration of glycosaminoglycans in Caenorhab-ditis elegans. FEBS Lett 459:327-331.
Yayon A, Klagsbrun M, Esko JD, Leder P, Ornitz DM. 1991. Cell surface, heparin-like molecules are required for binding of basic fibroblast growth factor to its high affinity receptor. Cell 64:841-848.
Yusa A, Kitajima K, Habuchi O. 2005. N-linked oligosaccharides are re-quired to produce and stabilize the active form of chondroitin 4-sulphotransferase-1. Biochem J 388:115-121.
Zak BM, Crawford BE, Esko JD. 2002. Hereditary multiple exostoses and heparan sulfate polymerization. Biochim Biophys Acta 1573:346-355.
Zoeller JJ, McQuillan A, Whitelock J, Ho SY, Iozzo RV. 2008. A central function for perlecan in skeletal muscle and cardiovascular devel-opment. J Cell Biol 181:381-394.

Acta Universitatis UpsaliensisDigital Comprehensive Summaries of Uppsala Dissertationsfrom the Faculty of Medicine 371
Editor: The Dean of the Faculty of Medicine
A doctoral dissertation from the Faculty of Medicine, UppsalaUniversity, is usually a summary of a number of papers. A fewcopies of the complete dissertation are kept at major Swedishresearch libraries, while the summary alone is distributedinternationally through the series Digital ComprehensiveSummaries of Uppsala Dissertations from the Faculty ofMedicine. (Prior to January, 2005, the series was publishedunder the title “Comprehensive Summaries of UppsalaDissertations from the Faculty of Medicine”.)
Distribution: publications.uu.seurn:nbn:se:uu:diva-9231
ACTA
UNIVERSITATIS
UPSALIENSIS
UPPSALA
2008