Embed Size (px)
Citation preview

AMINO ACID ACTIVATING ENZYMESIN RED BLOODCELLSOF NORMAL,ANEMIC AND POLYCYTHEMICSUBJECTS*
By G. IZAK, T. WILNERAND J. MAGERWITH THE TECHNICAL ASSISTA.NCE OFA. KARSHAI
(From the Department of Medicine B, and the Hematology Research Laboratory of HadassahUniversity Hospital and from the Department of Biochemistry, Hebreze'
University-Hadassah Medical School, Jerusalem, Israel)
(Submitted for publication May 18, 1960; accepted June 9, 1960)
Following the discovery by Hoagland (1) thatthe soluble cytoplasmic fraction of rat liver ho-mogenates catalyzes an amino acid-dependent py-rophosphate (PP) exchange with adenosine tri-phosphate (ATP), similar systems have been de-scribed in a variety of other tissues and organisms(2-4). These so-called "amino acid activatingenzymes" (AAAE) serve, according to the cur-rently prevailing opinion (5), to convert freeamino acids into the corresponding aminoacyladenylates, thus initiating the chain of reactionsmediating their incorporation into the proteinmolecule.
The present communication provides evidencefor the occurrence of AAAE in human red bloodcells (RBC). In addition, the paper presentsquantitative data indicating the existence of somesignificant deviations from the normal level ofactivity of these enzymes in certain well definedblood disorders.
MATERIALS AND METHODS
Blood samples. Blood was obtained by venipuncturefrom healthy donors and anemic or polycythemic pa-tients. The salient hematological data are recorded inTable I.
Preparation of hemolysates. The cells from freshlydrawn heparinized blood samples were sedimented bycentrifugation, washed 3 times with 2 vol of physiologicalsaline buffered with veronal, pH 7.3, and counted beforethe last washing. All the subsequent operations werecarried out in the cold.
Two ml of the packed blood cells were hemolyzed bydilution with 1.6 vol of cold distilled water. Following10 minutes of stirring, the isotonicity of the hemolysatewas restored by adding concentrated sucrose solutionto a final concentration of 0.25 M, and the volume wasmade up to 10 ml. Ghosts and particles were removedby spinning the hemolysate for 30 minutes at 15,000 G inthe Servall centrifuge. The resulting clear supernatant
* This work was supported by a research grant fromthe Hadassah Medical Organization.
solution was carefully decanted and served as the sourceof amino acid activating enzymes. The hemolysate couldbe stored at - 150 C for at least 2 weeks without ap-preciable loss of activity with respect to amino acid ac-tivation. This preparation was routinely used and noattempt was made to exclude platelets and leukocytes,since the contribution of these cells to the over-all ac-tivity of the lysate ,was found to be negligible (see Re-sults).
In some experiments the washed blood cells suspendedin 0.25 M sucrose were disrupted by sonication in an icewater-cooled 10 KC-Raytheon oscillator.
Chemicals. All chemicals were of analytic or reagentgrade. The radioactive pyrophosphate was prepared bypyrolysis of PU-labeled orthophosphate at 2800 C for 20to 30 hours. The product usually contained less than0.1 per cent orthophosphate.
Standard reaction mixture and assail procedure. Un-less otherwise stated, the following reaction mixture wasused for assaying amino acid activation: Tris buffer, pH7.4, 50 /,moles; MgCl2, 5 Amoles, ATP, 2 &moles; PP',2 ,umoles; amino acid mixture as indicated in the legendto Table II; NaF, 20 /Amoles; hemolysate, 0.3 ml. Thevolume was made up to 1 ml with distilled water and themixture was incubated for 20 minutes at 370 C. The re-action was stopped by the addition of 5 ml of 5 per centtrichloroacetic acid. The precipitated protein was re-moved by centrifugation, and 2 ml samples of the clearsupernates were each treated with 1 ml of a 5 per centsuspension of acid-washed charcoal ("Norit A") in or-der to adsorb the ATP (8). The charcoal was washed5 times with pH 4.5 acetate buffer containing 10 ,AmolesPP per ml, once with distilled water, and was then sus-pended in an alcohol: ether mixture (3: 1 vol/vol) andtransferred quantitatively to planchets. The radioactivitywas measured with an end-window Geiger-Muellercounter and the activity was calculated by dividing thetotal counts per minute incorporated into the ATP bythe specific activity of the pyrophosphate added and ex-pressed in terms of micromoles of PP exchanged per mil-ligram protein of the homolysate or per 10' RBC. Un-der the specified conditions the reaction closely followeda zero order course.
Analytical procedure. Total protein was determinedby the method of Lowry, Rosebrough, Farr and Randall(9) and hemoglobin was estimated by the cyanmethemo-globin procedure (10). Inorganic phosphate was deter-
1763
G. IZAK, T. WILNER AND J. MAGER
TABLE I
Summary of hematological data in the various groups of patients (range)
No. ofClinical diagnosis patients Hemoglobin RBC* Hematocrit Reticulocytes Serum iront Vitamin Bi4
gm % 106/cu mm % % pg % ;spg/ml serumNormal 30 13.4-15.2 4.6-5.2 44-49 0.1-0.6 79-108 180-420Macrocytic§
hyperchromic anemia 20 6.5-9.2 1.5-3.2 21-36 1.2-3.1 67-110 60-190Microcytic
hypochromic anemia 16 4.5-7.2 2.7-3.9 19-34 0.8-2.8 0-38 200-450Polycythemia vera 10 15.9-18.0 5.7-7.5 53-70 0.1-1.1 70-115 270-390
* RBC= red blood cells.t Determined according to the method of Davies, Levin and Oberholzer (6).
Determined according to the method of Grossowicz, Aronovitch and Rachmilewitz (7).§ No patients with pernicious anemia (lack of intrinsic factor) or aplastic anemia have been included in this group.
mined by the method of Fiske and Subbarow (11). Allthe experiments were run in duplicate.
RESULTS
Conditions governing PP32-A TP exchange in
hemolysates. As shown in Table II, hemolysatesprepared from normal RBCcatalyzed the incorpo-ration of P32-labeled pyrophosphate into ATP.
Omission of amino acids from the otherwisecomplete reaction mixture resulted in a decrease
TABLE II
PP32-A TP exchange in a normal blood hemolysate;requirements of the reaction *
pmoles pmolesATP3 ATP32
100 mgprotein 101" RBCt
"Complete amino acidmixture"t 11.6 33.0
Amino acid mixture "A"t 11.2 31.0Tryptophan 8.7 25.3No amino acids 5.1 13.7No ATP 2.1 4.5No Mg++ 3.8 6.3p3 Pi§ added instead of pp32 0 0Pyrophosphatase added 0 0
* Conditions as described under Methods; see first sec-tion of text for abbreviations.
f Based on the red cell count performed immediatelyprior to lysis.
"Complete amino acid mixture": arginine, aspartic,glycine, histidine, serine, isoleucine, leucine, lysine, phenyl-alanine, threonine, tryptophan, valine, glutamic, methi-onine, cysteine, tyrosine, proline. All amino acids wereof the L-series and were added at 5 X 10-4 M final con-centration. Amino acid mixture "A": arginine, glycine,histidine, serine, threonine, tryptophan, valine. Inorganicpyrophosphatase (40 ,ug) was added and the reactionmixture was pre-incubated for 5 to 10 minutes prior to theaddition of ATP. (Kindly supplied by Dr. M. Kunitz, theRockefeller Institute for Medical Research.)
§ Pi: inorganic phosphate.
in the rate of exchange to one-fourth to one-halfof that observed with the complete system. Itappeared likely that the considerable "background"activity obtained without the addition of aminoacids was due to the appreciable amounts of aminoacids released from the RBCin the course of lysis( 12), especially since low substrate saturation levelsare characteristic of the amino acid activating en-
zymes (2, 3). The validity of this assumption isstrengthened by the finding that the ratio between
TABLE III
Precipitation of the AAAE at pH 5.0 in the presence ofsodium ribonucleate *
Completeamino No
acid aminoTreatment of mixture acid Ratio Nohemolysatet A B A/B ATP
%activity 5activity
None 100 33 3.0 10pH 5.Ot 95 28 3.4 7pH 5.0 iSupernate 63 23 2.7 12
RNA§ fSediment 28 2.5 11.2 3
* The results are expressed in percentage of the activityobtained with untreated hemolysate, the latter set at 100per cent. See footnotes * and I to Table II for definitions.
t Hemolysate: particle-free supernate prepared fromwhole blood as described under Methods.
t The pH of the ice-chilled hemolysate was lowered to5.0 by dropwise addition of 1 N acetic acid with continu-ous stirring. No precipitate was obtained. The pH ofthe hemolysate was immediately readjusted with 1 N Trisbuffer to 7.2.
§ pH 5.0 + RNA: 30 mg/ml sodium ribonucleate[commercial yeast, RNA, purified according to the pro-cedure of Woodward (13)] was added to the hemolysatewith stirring until dissolved. Thereafter the solution wastreated as above. The precipitate formed at pH 5.0 wassedimented by centrifugation and dissolved in a measuredquantity of Tris buffer, 0.025 M, pH 7.2. The pH of thesupernate was readjusted as above.
1764

AMINO ACID ACTIVATING ENZYMESIN MATUREERYTHROCYTES
the endogenous and the amino acid-stimulatedPP32-ATP exchange activity increased from 1: 3in the crude hemolysate to 1: 11 in a fraction ofthis hemolysate isolated by pH precipitation(Table III).
By examining various amino acids singly andin combination, it was found that amino acid mix-ture "A" (see legend to Table II) enhanced theexchange rate almost to the same extent as did a
mixture of 18 natural amino acids, whereas 80 per
cent of the maximal effect was elicited by trypto-phan alone.
The radioactivity yielded without added ATPwas about 15 per cent of that obtained with thecomplete reaction mixture. These counts couldbe almost entirely eliminated by using an isolatedfraction of the hemolysate, indicating that this re-
sidual activity was due to the endogenous ATPcontent of the hemolysate (Table III).
Omission of Mg++ resulted in a roughly 70 per
cent drop of the reaction rate (Table II).P32-orthophosphate failed to induce any detecta-
ble exchange with ATP when substituted for pp32in the standard reaction mixture. The specificityof the system with respect to pyrophosphate was
even more convincingly demonstrated by the find-ing that crystalline inorganic pyrophosphatasecompletely abolished the exchange reaction(Table II).
The pH of optimum PP32-ATP exchange ac-
tivity was found to range from 6.1 to 7.3. Byvarying the concentration of the amino acids inthe reaction mixture the maximal rate of exchangewas attained at levels close to 2 X 10-4 M.
The considerable adenosine triphosphatase andpyrophosphatase activity of the lysates, as meas-
ured by the release of orthophosphate from the re-
spective substrates in the course of incubation, was
entirely eliminated by the fluoride included in thereaction mixture. The stability of the assay sys-
tem was also borne out by the fact that the rateof exchange remained practically constant duringan experimental period which lasted for at least40 minutes (Table IV).
Identification of the reaction product. At theend of the reaction the labeled ATP was adsorbedonto charcoal and the residual PP was removedby extensive washing. Thereafter the ATP was
eluted with 50 per cent ethanol containing 0.5 Mammonia and isolated by paper electrophoresis,
TABLE IV
Rate of PP32-A TP exchange determined after increasinglengths of incubation in normal and polycythemic blood
hemolysates *
Rate of exchangeIncubation
time Normal Polycythemic
min mumoles/hr/mg protein10 35.4 24.920 35.4 21.630 33.045 36.0 23.1
* Rate of exchange (R) was calculated according to thefollowing equation (14):
R=(ATP) X (PP32) 2.3 lg100
(ATP) + (Pp32) t X log 100 - %exchangeConditions as in Table II. Amino acid mixture "A" wasused.
using 0.005 M citrate of pH 3.5 as the mobilephase. Under these conditions radioactivity wasdetectable only in the ATP spot located by itsquenching of ultraviolet light. When the labeledATP was hydrolyzed with 1 N HCl in a boilingwater bath for 10 minutes, the radioactivity was nolonger adsorbable with charcoal. The latter re-sult indicated, therefore, that the P32-label wasentirely confined to the acid-labile terminal pyro-phosphate moiety of ATP.
Fractionation. The AAAEof the rat liver cy-toplasmic fraction are referred to by Hoagland,Keller and Zamecnik (15) as pH 5.0 enzymes be-cause of their precipitating property at this pH asa nucleoprotein complex. The same procedureapplied to whole blood hemolysates failed to pro-duce a precipitate without affecting, however, theactivity, provided that the pH was quickly read-justed to neutrality. It appeared likely that thisdifference in behavior is attributable to the virtual
TABLE V
Distribution of amino acid activating enzymes in variousformed elements of the peripheral blood *
jLmoles pAmolesATP32 ATP32
%of100 mg loll totaltprotein cells activity
Red blood cells 13.0 30.0 95.6White blood cells 69.0 78.0 0.4Platelets 30.0 37.0 4.0Whole blood 13.57 31.3 100.0
* Conditions as described under Methods.t The activity of the whole blood hemolysate was set at
100 per cent.
1765
G. IZ.AkK, T. WILN'ER AND J. MAGER
TABLE VI
Mean values of PP32-A TI' exchange in blood lysates from normlal subjects and fromanemic and polycythemic patients
Normal Macrocytic anemia Microcytic anemia Polycythemia
pmoles ATP32 ymoles ATP32 Mmoles ATP32 Jlmoles ATP32 Mmoles ATP32 Mimoles ATP32 pmoles ATP32 Amoles ATP32
100 mg 100 mg 100 mg 100 mgprotein 1011 RBC protein 1011 RBC protein 101" RBC protein 1011 RBC
Mean SD Mean SD Mean SD Mean SD Mean SD Mean SD Mean SD Mean SD
Amino acidmixture "A"* 10.6 3.0 31.0 6.8 17.8 4.7 42.3 6.6 24.4 5.3 50.0 14.5 6.9 1.1 15.3 3.2
Tryptophan 8.1 1.9 24.9 6.2 14.4 4.6 35.0 7.3 20.0 5.2 40.0 10.2 7.0 2.1 15.6 4.7No amino acids 4.5 1.4 12.3 2.7 7.5 2.1 18.3 4.4 8.0 1.5 14.1 4.1 2.9 0.4 6.7 1.3No ATP 2.0 0.8 5.0 2.0 2.1 0.5 6.0 1.5 1.9 0.6 3.5 1.5 1.2 0.2 2.7 0.4
* Amino acid mixture "A"; see footnote: to Table II. Conditions otherwise as in Table II.
absence of nucleic acids in the hemolysate. Thisseems to b)e confirmed by the finding that whenRNAwas added to the hemolysate, a precipitatewas obtained at pH 5.0 which contained about25 per cent of the total AAAEactivity present inthe original hemolysate (Table III). No furtherfractionation was attempted.
The distribution of AAAE in the peripheralblood cells. The distribution of the AAAE in thevarious formed elements of the peripheral bloodwas studied by determining the PP32-ATP ex-change in lysates prepared separately from eryth-rocytes, leukocytes and platelets isolated by amodification of a conventional technique (16).As evident from Table V, the bulk of the total
blood cell activity resides in the RBC. On theother hand, the specific activity of the lysates cal-culated per unit number of cells is highest in theleukocytes and lowest in the RBC.
Leukocytes and RBCinjured by a single hypo-tonic treatment retained about as much activity asthat released into the supernatant solution. Theresidual activity could be solubilized by subjectingthe ghosts resedimented by centrifugation to re-peated osmotic lysis or to sonication for 2 to 3minutes. On the other hand, ghosts of plateletsobtained after a single hypotonic lysis exhibitedno detectable PP32-ATP exchange. It may bepointed out in this connection that no PP32-ATPexchange could be demonstrated with any of the
ATP AuM100mg Protein
1st 2nd 3d 4th I 2nd 3d
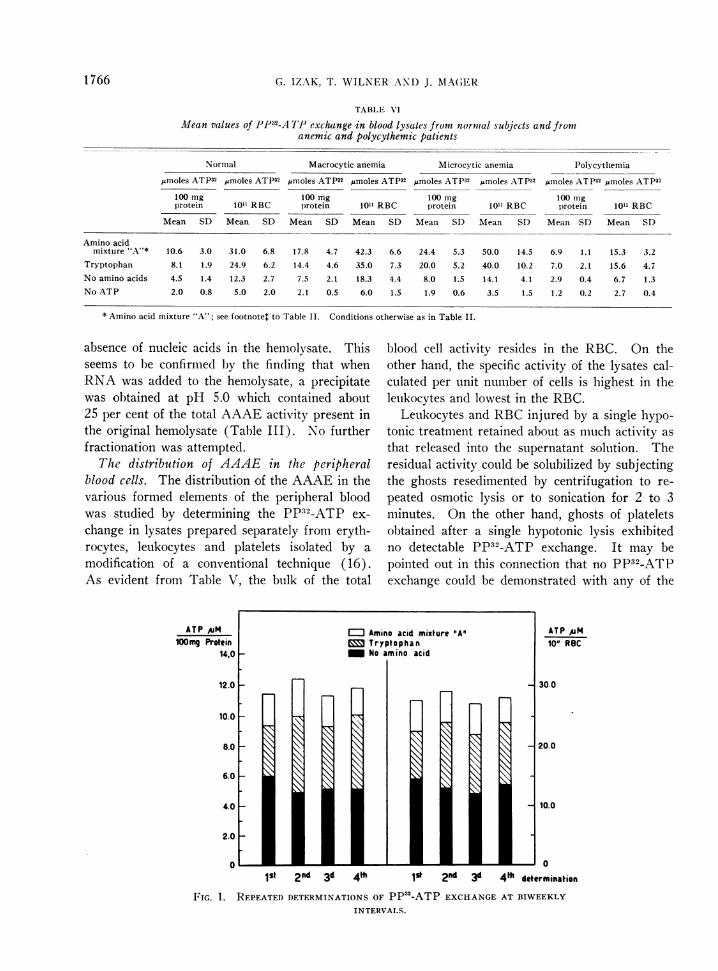
FIG. 1. REPEATEDDETERMINATIONSOF PP52-ATP EXCHANGEINTERVALS.
ATP AM10" RBC
- 30.0
20.0
* 10.0
041 determination
AT BIWEEKLY
1 766
AMINO ACID ACTIVATING ENZYMESIN MATUREERYTHROCYTES
ATP puM 17.0100 mg Prdein
16.0
15.0
14.0
13.0
12.0
11.0
10.00 10 20 30 40
Hemoglobingm%
13.0
12 .
11.0
10.0
9.0
8D
7.0
D a y s
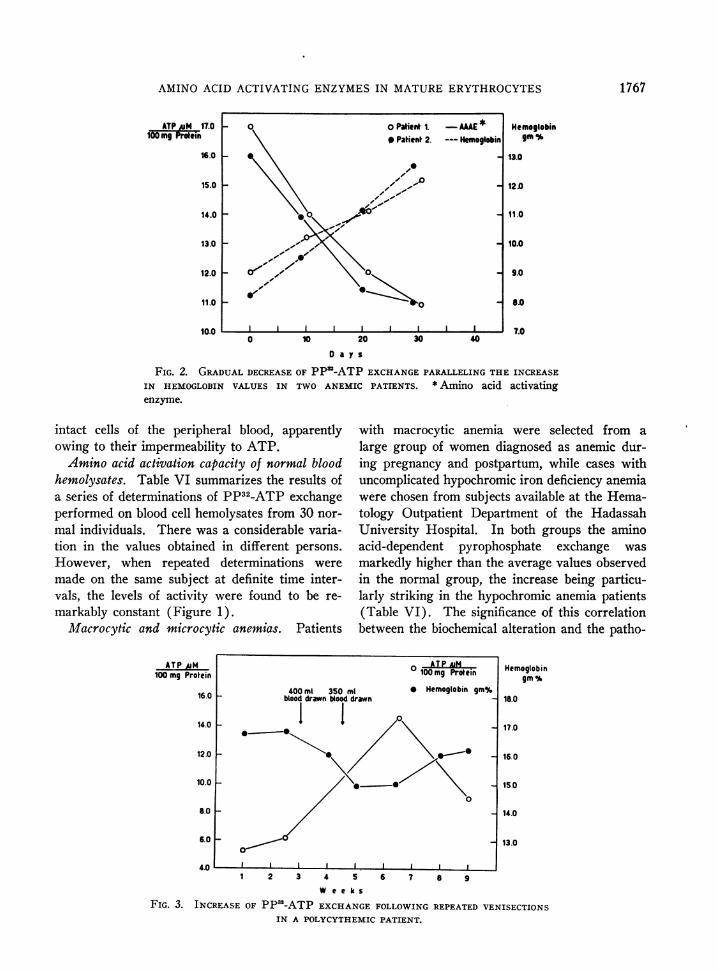
FIG. 2. GRADUALDECREASEOF PPS-ATP EXCHANGEPARALLELING THE INCREASEIN HEMOGLOBIN VALUES IN TWO ANEMIC PATIENTS. * Amino acid activatingenzyme.
intact cells of the peripheral blood, apparentlyowing to their impermeability to ATP.
Amino acid activation capacity of normal bloodhemolysates. Table VI summarizes the results ofa series of determinations of PP32-ATP exchangeperformed on blood cell hemolysates from 30 nor-mal individuals. There was a considerable varia-tion in the values obtained in different persons.However, when repeated determinations weremade on the same subject at definite time inter-vals, the levels of activity were found to be re-markably constant (Figure 1).
Macrocytic and microcytic anemias. Patients
ATPAiM100 mg Protein
16.0 _
14.0
12.0
10.0 I
8.0
6.0
400 ml 350 mlblood drawn blood drawn
I
with macrocytic anemia were selected from alarge group of women diagnosed as anemic dur-ing pregnancy and postpartum, while cases withuncomplicated hypochromic iron deficiency anemiawere chosen from subjects available at the Hema-tology Outpatient Department of the HadassahUniversity Hospital. In both groups the aminoacid-dependent pyrophosphate exchange wasmarkedly higher than the average values observedin the normal group, the increase being particu-larly striking in the hypochromic anemia patients(Table VI). The significance of this correlationbetween the biochemical alteration and the patho-
Hemoglobingm%
18.0
17.0
16.0
15.0
14.0
13.0
W e e k s
FIG. 3. INCREASE OF PP32-ATP EXCHANGEFOLLOWINGREPEATEDVENISECTIONS
IN A POLYCYTHEMICPATIENT.
1767
O AT P AiM100mg Protein
* Hemoglobin gm%
I I 0
0~~~~~~~
I I I ~~~~~~~I I
1 2 3 4 5 6 7 8 94.U * ' ' IAA L

G. IZAK, T. WILNER AND J. MAGER
logical condition is further borne out by the fol-low-up of six treated patients in whom progres-sive decline of activity down to the normal rangeparalleled the improvement in the hematologicalstatus. Two representative cases are illustratedin Figure 2.
Polycythemia vera. In untreated patients withpolycythemia vera the pyrophosphate exchangewas of a considerably lower order than in normalsubjects. Addition of tryptophan alone to the re-action mixture elicited maximal response whichwas not increased by further addition of otheramino acids (Table VI). In these patients theactivity rose substantially after repeated venesec-tions, gradually returning, however, to the originallow level within several weeks following the lastphlebotomy (Figure 3).
Osmnotic fragility and AAAE. The high ac-tivity found in anemic patients suggested the pos-sibility that the elevated values were due to a"shift to the left" in the age of the red cell popu-lation in view of the shortened RBC life spancharacterizing these conditions (17, 18). Thispossibility was explored by examining the enzymicactivity as a function of the osmotic fragility whichis known to vary directly with the age of the redcells (19).
Table VII shows that the activity per unit num-ber of red cells lysed remained roughly constant.The anemic cells showed an essentially similar be-havior. It may be relevant to mention in thiscontext that there was no correlation between thedegree of reticulocytosis and the PP32-ATP ex-change activity in the various groups studied.Thus the AAAE exhibits a uniform pattern ofdistribution in the RBC population, differing in
TABLE VIIPP32-A TP exchange in relation to
osmotic fragility
Microcytic hypo-Normal chromic anemia
%Hemo- % Hemo-
Activ- globin Activ- globinNaCi ity in in ity in in
solutioni supernate sul)ernate supernate sulernate
0.4 30 39 14 170.35 90 92 60 5 10.30 95 93 90 84
100* 100 100 100
* The values obtained after lysis with distilled water arearbitrarily conisidered as 100 per cenit.
this respect from the behavior of some other en-zymes, as described by Marks, Johnson, Hirsch-berg and Banks (20).
DISCUSSION
The occurrence of amino acid activating en-zymes in mature red blood cells which are knownto be incapable of protein production is surprisingin view of the presumed essential role of these en-zymes in the biosynthesis of protein (5). It ispossible, of course, that the AAAE perform inthe mature RBC a different function which re-mains to be elucidated. It is likewise conceivable,however, that the system concerned represents anonfunctional rudiment carried over from the nu-cleated stages of erythropoiesis.
The pronounced differences in the AAAE ac-tivity observed between blood cells from normalsubjects and those from patients with various he-matological disorders seem to reflect early disturb-ances occurring at somiie stage of the RBCmatura-tion rather than a change in the type of the ma-tture blood cells. The uniform distribution of thisactivity within the blood cell population, regard-less of the differences in the osmotic fragility inboth normal subjects and anemic patients, indi-cates a lack of correlation between the level ofAAAE and the age of RBC. This conclusion isalso in line with the observation that the reticu-locyte count did not seem to affect significantly theactivity observed within a given group.
The nature of the biochemical lesion responsiblefor the observed deviations from the normal levelof activity is not clear. It is tempting to speculatethat the enhanced AAAE activity found in theanemic disorders described manifests an attemptof the hematopoietic system to compensate for thedeficient hemoglobin synthesis. This hypothesiswould also account for the gradual decrease of theactivity, down to normal values, occurring sometime after a normal hemoglobin level had been at-tained. On the other hand, the decrease in theAAAE level observed in patients with polycy-themia vera may reflect an opposite tendency,aimed to restrain the excessive activity of the(errytbroloietic tissule. Tt is interestino to men-tiOn in this colllectioln that the AA.\I activity de-termined in the IRBC of a limited number of pa-tients with secondary polycythemia was found to
1 768

AMINO ACID ACTIVATING ENZYMESIN MATUREERYTHROCYTES
be within the normal range. As to the transientincrease of the PP32-ATP exchange observed inpolycythemic patients following repeated venesec-tions, this represents a response of the protein syn-thesizing system to the drastic reduction in thecirculating red cell mass.
Thus the mechanisms discussed above can beviewed as two facets of a homeostatic device operat-ing through a restrictive effect exerted by the redcell proteins on the formation of the enzymic sys-tem mediating their own biosynthesis. No in-formation is available as to the nature of the regu-latory factor (s) involved in this mechanism.
Further work is required to test the validity ofthis interpretation.
SUMMARY
Human whole blood hemolysates were found tocatalyze an amino acid-dependent exchange ofpyrophosphate with adenosine triphosphate. Outof a number of amino acids tested, tryptophanshowed highest activity, amounting to about 80per cent of the total exchange rate elicited by amixture of 18 natural amino acids. Magnesiumions proved to be an essential component of thereaction.
Other factors influencing the reaction rate, suchas amino acid concentration and pH optimum,were determined. The product of the exchangereaction was characterized as adenosine triphos-phate labeled in the two terminal phosphategroups.
The bulk of the total amino acid activating en-zyme content of the hemolysates was present inthe red blood cells, while leukocytes showed thehighest specific activity. The distribution ofthese enzymes in the red blood cell populationshowed no correlation with its osmotic fragilitypattern.
The level of amino acid activating enzyme inerythrocytes derived from patients with macrocyticand microcytic anemias was markedly increasedas compared with normal values. On the otherhand, a significant decrease of this activity wasobserved in red cells obtained from patients withpolycythenlia vera. These elevations above thenormal range tended to regress parallel with he-matological improvement.
The significance of these enzymes in mature red
cells and the possible mechanism underlying thedeviations from the normal level in various he-matologic disorders are discussed.
REFERENCES
1. Hoagland, M. B. An enzymic mechanism for aminoacid activation in animal tissues. Biochim. bio-phys. Acta 1955, 16, 288.
2. DeMoss, J. A., and Novelli, G. D. An amino aciddependent exchange between inorganic pyrophos-phate and ATP in microbial extracts. Biochim.biophys. Acta 1955, 18, 592.
3. Davies, E. W., Konigsberger, V. V., and Lipmann, F.The isolation of a tryptophan-activating enzymefrom pancreas. Arch. Biochem. 1956, 65, 21.
4. Mager, J., and Lipmann, F. Amino acid incorpora-tion and the reversion of its initial phase with cell-free tetrahymena preparations. Proc. nat. Acad.Sci. (Wash.) 1958, 44, 305.
5. Lipmann, F., Hulsmann, W. C., Hartmann, G.,Boman, H. G., and Acs, G. Amino acid activationand protein synthesis. J. cell. comp. Physiol. 1959,54, 75.
6. Davies, G., Levin, B., and Oberholzer, V. G. Themicroestimation of serum iron and unsaturatediron-binding capacity in normals and in disease.J. clin. Path. 1952, 5, 312.
7. Grossowicz, N., Aronovitch, J., and Rachmilewitz,M. Determination of vitamin B,2 in human serumby a mutant of Escherichia coli. Proc. Soc. exp.Biol. (N. Y.) 1954, 87, 513.
8. Crane, R. K., and Lipmann, F. The effect of arsenateon aerobic phosphorylation. J. biol. Chem. 1953,201, 235.
9. Lowry, 0. H., Rosebrough, N. J., Farr, A. L., andRandall, R. J. Protein measurement with Folinphenol reagent. J. biol. Chem. 1951, 193, 265.
10. Duchateau, N. J. A comparative study of oxyhemo-globin and cyanmethemoglobin determinations byphotometric and spectrophotometric methods.Amer. J. med. Technol. 1957, 23, 17.
11. Fiske, C. H., and Subbarow, Y. The colorimetricdetermination of phosphorus. J. biol. Chem. 1925,66, 375.
12. Behrendt, H. J. Chemistry of Erythrocytes: ClinicalAspects. Springfield, Ill., Charles C Thomas, 1957,p. 58.
13. Woodward, G. E. The ribonuclease activity of Pas-teurella pestis (Plague Bacillus). J. biol. Chem.1944, 156, 143.
14. Duffield, R. B., and Calvin, M. The stability ofchelate compounds, III. Exchange reactions ofcopper chelate comrpouinds. J. Amer. chem. Soc.1946, 68, 557.
15. Hoagland, M. B., Keller, E. B., and Zaimecnik, P. C.Enzymatic carboxyl activation of amino acids. J.biol. Chem. 1956, 218, 345.
1769

G. IZAK, T. WILNER AND J. MAGER
16. Nelken, D., Gilboa, N. (Garber) and Gurevitch, J.A method for the simultaneous separation of hu-man thrombocytes and leukocytes. J. clin. Path.In press.
17. Scott, J. M., and Govan, A. B. Anaemia simulatingpre-eclamptic toxaemia. J. Obstet. Gynaec. Brit.Emp. 1949, 56, 27.
18. Fairley, N. H., Bromfield, R. J., Foy, H., and Kondi,A. Nutritional macrocytic anaemia in Macedonia;
preliminary report. Trans. roy. Soc. trop. Med.Hyg. 1938, 32, 132.
19. Chalfin, D. Differences between young and maturerabbit erythrocytes. J. cell. comp. Physiol. 1956,47, 215.
20. Marks, P. A., Johnson, A. B., Hirschberg, E., andBanks, J. Studies on the mechanism of aging ofhuman red blood cells. Ann. N. Y. Acad. Sci.1958, 75, 95.
1770





























