Embed Size (px)
Citation preview

Immunology and Cell Biology (2002) 80, 1–13
Review Article
CD8+CD28– T cells: Certainties and uncertainties of a prevalent human T-cell subset
F E R N A N D O A A R O S A
Laboratory of Molecular Immunology, Institute for Molecular and Cell Biology (IBMC) and Molecular Immunology and Pathology, Abel Salazar Institute for the Biomedical Sciences (ICBAS), Porto, Portugal
Summary Human peripheral blood CD8+ T cells comprise cells that are in different states of differentiation andunder the control of complex homeostatic processes. In a number of situations ranging from chronic inflammatoryconditions and infectious diseases to ageing, immunodeficiency, iron overload and heavy alcohol intake, majorphenotypic changes, usually associated with an increase in CD8+ T cells lacking CD28 expression, take place.CD8+CD28– T cells are characterized by a low proliferative capacity to conventional stimulation in vitro and bymorphological and functional features of activated/memory T cells. Although the nature of the signals that giveorigin to this T-cell subset is uncertain, growing evidence argues for the existence of an interplay between epithelialcells, molecules with the MHC-class I fold and CD8+ T cells. The possibility that the generation of CD8+CD28–
T cells is the combination of TCR/CD3ζ- and regulatory factor-mediated signals as a result of the sensing ofmodifications of the internal environment is discussed.
Key words: CD8+ T cells, CD28, downmodulation, epithelial cells, homeostasis, major histocompatibility complex-class I.
Introduction
One of the fundamental assets of the T-lymphocyte system, inaddition to its capacity to circulate, is its capacity to undergohomeostatic changes in response to alterations of the internalenvironment. Whether the origin of these alterations is fromoutside or from inside, physical contact between cells andmolecules of the immunological system on the one hand, andof the altered internal milieu on the other, are necessary toelicit activation signals that will ultimately result in qualita-tive and quantitative changes within the peripheral T-cellpool. The signalling events following the primary (e.g. TCR/MHC) and secondary (e.g. CD2/LFA-3, CD28/B7) inter-actions play key roles in the process of T-cell activation andfunction and have been well reviewed elsewhere.1–4 Never-theless, despite the fact that professional antigen-presentingcells (APC), such as dendritic cells, tissue macrophages,monocytes and activated B cells, can initiate an immuno-logical response by primarily activating CD4+ T cells,accumulating evidence indicates that the alterations of theT-lymphocyte system in response to signals of the internalmilieu result in phenotypic changes mostly affecting theCD8+ T-cell pool. The most conspicuous phenotypic changesunder these situations are downmodulation of the CD3ζ chainand complete loss of CD28, which are counteracted by denovo expression of non-specific T-cell markers such asCD11b, CD56, CD57 and CD161. The molecular basis for,
and physiological significance of, the increased tendency ofCD8+ T cells to undergo shifts in response to alterations of theinternal milieu is unclear. However, recent data indicate thatnon-haematopoietic cells, such as epithelial cells, are capableof activating CD8+ T cells. In turn, the activated CD8+ T cellsmay be involved in the regulation of basic physiologicalprocesses within the organ and/or tissue they populate. Toelucidate the mechanisms involved in the activation of CD8+
T cells, loss of CD28 expression and the generation ofdifferentiated CD8+CD28– T cells with regulatory functions isof foremost importance. The purpose of this review is to bringup to date the certainties and uncertainties of this prevalenthuman CD8+ T-cell subset.
CD8+CD28– T cells: From phenotype to function
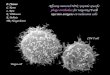
Although CD28 provides a critical costimulus for T-cellactivation,1–4 a large fraction of human peripheral blood Tcells lack CD28, which is due to the different expression ofCD28 by the two major T-cell subsets. While in healthyhumans CD28 is expressed by the majority of CD4+ T cells,the percentage of CD8+ T cells coexpressing CD28 is highlyvariable.5–7 Peripheral blood CD8+CD28– T cells are enrichedfor large, granular lymphocytes (Fig. 1), express high levelsof LFA-1 and variable amounts of CD11b and CD57, aphenotype associated with memory/effector cells.5–7 Freshlyisolated peripheral blood CD8+CD28– T cells from healthyhumans do not express markers of early (e.g. CD69) orchronic (e.g. HLA-DR) activation and proliferate poorly inresponse to conventional mitogenic stimulation in vitro.7 ACD4+ T-cell subset with similar phenotypic and functionalcharacteristics also exists, at very low frequencies, in theperipheral blood of healthy humans.8,9
Correspondence: Dr FA Arosa, Laboratory of Molecular Immu-nology, Institute for Molecular and Cell Biology, Rua do CampoAlegre, 823, 4150-180 Porto, Portugal. Email: [email protected]
Received 19 February 2001; accepted 3 September 2001.

2 FA Arosa
Although the existence of a pool of circulating CD8+CD28–
T cells in humans has been known to exist for more than15 years, specific interest in this CD8+ T-cell subset aroseafter finding increased numbers during infections.10–14
CD8+CD28– T cells were first described as suppressors of B-and T-cell function, but subsequent studies have shown thatCD8+ T cells bearing the CD28– phenotype (CD11b+ and/orCD57+) may also mediate cytotoxicity.15–18 Studies withfreshly isolated peripheral blood lymphocytes demonstratedthat CD8+CD11b+ T lymphocytes contain cells capable ofsuppressing proliferation and IgG synthesis of B cells, andcells that express the IL-2β receptor chain (CD8+CD122+
T cells) and develop into cytotoxic T lympho-cytes (CTL)after culturing with IL-2.18–21 Importantly, IL-2-induced CD8+
T-cell proliferation results in CD11b loss and the generationof CD8+CD28–CD11b– T cells,21 a non-cytotoxic subsetexpanded in the blood of HIV-1-infected patients.22 Recentwork by Suciu-Foca and coworkers has disclosed some of themolecular mechanisms that account for the suppressor activ-ity of CD8+CD28– T cells.23–25 By inhibiting the nuclear factorkappa B (NF-κB) mediated transcription of the CD86 gene,CD8+CD28– T cells hamper the ability of classical APC toelicit CD28-driven CD4+ T activation and proliferation, asuppressor activity that is MHC-class I-restricted.23–25
These data clearly indicate that CD8+CD28– T cells are aheterogeneous subset that contains both memory and effectorcells.26 The most established view, however, considersCD8+CD28– T cells as terminally differentiated or end-stageCTL; a belief based on phenotypic and functional studies.First, CD8+CD28– T cells express CD11b, a member of theβ2-integrin family expressed by monocytes, neutrophils andnatural killer (NK) cells,27 which is considered to be a markerof CD8+ cytotoxic T-cell activation in response to viral
infection in mice.28,29 Yet, together with LFA-1, another β2-integrin, CD11b is believed to play an important role inextravasation of lymphocytes after adhesion to the endothe-lium via intercellular adhesion molecule-1 (ICAM-1).27,30 Inaddition, along with peripheral blood CD8+ T cells, CD11b isexpressed by a significant fraction of CD3+ T cells in the liverand spleen of healthy subjects.31 Recent studies suggest thatthe acquisition of CD11b represents an early event duringhuman CD8+ T-cell differentiation, which may allow extrava-sation to peripheral tissues.32 Second, CD8+CD28– T cellsexpress CD57, a molecule containing the epitope humannatural killer cell carbohydrate antigen-1 (HNK-1), which ispresent on human NK, CD4+ and CD8+ T cells.33,34 CD57, ina manner analogous to CD11b, is a marker associated withCTL effector function due to its increased expression amongCD8+ T cells during immune responses to viral infections.35–38
However, the HNK-1 epitope is also present on glycolipidsand glycoproteins expressed in the central nervous system(e.g. MOG and NCAM) and involved in cell–cell interac-tions.39 In this context it is important to note that CD8+ T cellsexpressing CD57, but not CD11b, are present at low levels innormal bone marrow.40,41 Finally, CD8+CD28– T cells areenriched for large granular lymphocytes that contain per-forin,7,26 a protein involved in target cell apoptosis.42 Thus,these data reveal that CD8+CD28– T cells are a multifacetedpopulation capable of performing many functions, with cyto-toxicity the most extensively studied. Indeed, a recent studyon circulating CD8+CD28– T cells has shown that CTLeffector function correlates with the expression of CD56,43 atypical NK marker also containing the HNK-1 epitope.39
Although the loss of CD28 and gain of CD11b and/orCD57 by peripheral blood CD8+ T cells is the result of adifferentiation process that generates cells with cytotoxic
Figure 1 Human CD8+CD28– Tcells are enriched for large, gran-ular lymphocytes. Freshly col-lected human peripheral bloodlymphocytes from a healthy blooddonor were stained for CD8/CD28and acquired in a FACScaliburaccording to forward and sidescatter characteristics (FSC/SSC,left dot blot). After magnification,two electronic gates were createdaround small/agranular (gate 1)and large/granular (gate 2) lym-phocytes. CD28 expression among(CD3+)CD8bright T cells withineach electronic gate is shown onthe right dot blots. The percent-age of (a) CD8+CD28+ and (b)CD8+CD28– T cells are indicated.

Human CD8+CD28– T cells 3
potential, to consider CD8+CD28– T cells as end-stage CTLeffectors constrains the study of other functions that thisCD8+ T-cell subset may perform. Thus, the increase inCD8+CD28– T cells in peripheral blood could be the result ofa viral infection,10–13,44–49 but may also be a response toalterations of the internal environment,50–52 or even of homeo-static changes within the peripheral T-lymphocyte systemitself. In addition, the CTL effector function detected amongexpanded CD8+CD28– T cells during viral infection43–45 hasbeen difficult to detect by some authors,47–49 unless the bulk ofthe CD8+ T cells were first incubated with IL-2, a cytokinethat upregulates CD3ζ and restores their cytotoxic potential.48
The recently developed peptide-HLA tetramer technology hasallowed researchers to isolate and study in more detailpeptide-specific CD8+ T cells from peripheral blood duringprimary and secondary viral infections.53 Surprisingly, thesestudies have shown that tetramer-specific CD8+ T cells are acomplex population, which in most cases show impairedcytotoxicity activity.54–59 Although the CD8+CD28– T-cellphenotype has been associated with terminal differentiation,unresponsiveness and cytotoxicity it is important to note thatincreased numbers of circulating CD8+CD28– T cells in anumber of disease conditions and situations have beenreported. As shown in Table 1, despite being the most stud-ied, viral infections are only one among the many possiblesituations that could lead to the generation of CD8+CD28–
T cells in humans.
Signals and pathways for CD28 downmodulation
T-lymphocyte activation in vitro and in vivo results inincreased internalization and degradation of components ofthe TCR/CD3ζ complex, namely the CD3ζ chain, and this
results in the downmodulation of cell surface CD3.60 T-cellactivation by MHC-peptide complexes, B7-expressing cellsor mitogens, such as phytohaemagglutinin, also results inCD28 downmodulation by peripheral blood T cells.61–63 Incontrast to CD3ζ, modulation of cell surface CD28 initiatedafter TCR-engagement results in the complete loss of CD28after several cycles of cell division in the presence of γ-chainreceptor signalling cytokines such as IL-2, IL-7 and IL-15.64–67
Hence, CD28 downmodulation initiated after TCR triggeringappears to be an irreversible process that is modulated byvarious factors and cytokines and leads to the generation ofCD8+CD28– T cells in humans. In an elegant study usingCD8+ T cells, Fiorentini et al. showed that after prolongedin vitro stimulation in the presence of IL-2, CD8+CD28+
T cells showed a gradual decrease in CD28 cell surface andmRNA levels and matured into a stable CD8+CD28– pheno-type.66 Similar results were obtained by Labalette et al.which, in addition, showed that IL-4 inhibited this process.68
Thus, loss of CD28 is a hallmark of T cells that haveundergone many cycles of cell division. Indeed, loss oftelomeric DNA resulting from cell division69,70 is observed inCD8+CD28– T cells from healthy and diseased subjects,71,72 anindication of their higher replicative past when comparedwith their CD8+CD28+ T-cell precursors. Telomeric shorten-ing is only evident among CD8+ T cells that have completelylost CD28 expression,73–75 thus, suggesting a link betweenCD28 downmodulation and telomerase activity. Two recentstudies have revealed that telomere shortening in peripheralblood lymphocytes also takes place during the first years oflife,76,77 indicating that T cells undergo a high turnover duringearly childhood, which is a time when internal changes (e.g.organ and tissue remodelling) are still taking place.
To date, CD28 is the only T-cell surface molecule, whichhas been described, that undergoes complete downmodula-tion, a process that is more apparent on CD8+ T cells.Considering the phenotypic changes associated with CD28downmodulation,38 identification and characterization of theactivation signals and factors that regulate CD28 is of greatimportance. In this context, recent studies by Goronzy andcoworkers have shown that the differences in CD28 expres-sion between CD4+ and CD8+ T cells correlate with differ-ences in nuclear protein-binding activities to two motifs, αand β, located within the CD28 promoter.78 In vitro activationvia the TCR/CD3ζ complex correlates with loss of α/β-binding activities both in CD4+ and CD8+ T cells and partialCD28 downmodulation. On the contrary, repeated activationand long-term culture in the presence of IL-2 results in loss ofβ- but not α-binding activities and correlates with a markedmodulation of CD28 expression solely on CD8+ T cells, apattern of expression that mirrors what happens in vivo.78 It isimportant to note that freshly isolated CD4+CD28– andCD8+CD28– T cells lack α/β-binding activities. Further char-acterization of these lymphoid factors may turn out to be ofparamount importance in elucidating the nature of theenvironmental signals and factors that lead to the loss ofCD28 by CD8+ T cells.
Despite the recent advances in our understanding of howCD28 expression is regulated, it is difficult to determinewhich cells provide the original stimulus and the context inwhich CD8+CD28– T cells develop. Still, the well-knownMHC-restriction of CD8+ T cells argues in favour of a
Table 1 Human conditions and disorders associated with anincrease in the percentage and/or number of peripheral bloodCD8+CD28– T cells (defined either as CD28–, CD11b+or CD57+)
Condition/disorder References
CD4+ lymphopenia 132Chagas disease 14Chemotherapy 147Chronic lymphocytic leukaemia 124CMV infection 36, 37, 44, 46, 48, 49, 65Immunodeficiency 122, 123Crohn’s disease 114EBV infection 13, 65Healthy heavy alcohol drinkers 89, 106Healthy humans (aged, including
centenarians)31, 64, 79, 82, 85, 86, 91–93
Healthy humans (with a highpercentage of CD8+ T cells)
87, 89
Hematological tumours 124, 125Hemochromatosis 87Hemodialysis patients 121Hemophilia A 120HIV infection 10, 12, 57, 65, 72Melanoma patients 88Myeloma patients 122, 126Rheumatoid arthritis 115, 127Systemic lupus erythromatosus 73, 117Transplants 50–52Wegener’s granulomatosis 118, 119

4 FA Arosa
predominance of MHC-class I+ versus MHC-class II+ cells indisplaying alterations of the internal environment. Further-more, in the case of continuous encountering with the MHC-class I+ cells, this prevalence would have an immediateoutput: the accumulation within the peripheral CD8+ T-cellpool of CD28– T cells carrying similar TCRαβ specificities.The reported oligoclonality of CD8+(CD57+)CD28– cellsboth in health and in disease is in accordance with thisscenario.79–83 The fact that CD8+CD28– T cells are oligoclonaland have short telomeres,71,83 together with the existence ofidentical clones in CD8+CD28+ and CD8+CD28– Τ cells84
stresses the MHC-class I driven mechanism. CD8+CD28–
T-cell oligoclonality may also suggest that a restricted pool ofMHC-class I/peptide structures account for most of theCD8+CD28– T-cell expansions. Alternatively, conservedstructures in physical association or combination with MHC-class I molecules may also be involved. Finally, the possiblerole played by non-classical MHC-class I molecules indriving the CD28+ to CD28– transition cannot be ruled out. Inany case, and contrary to current beliefs, the preferentialmodulation of CD28 observed in CD8+ T cells may beadvantageous rather than deleterious.
CD8+CD28– T-cell origin and function: Clues from health and disease
The presence of CD8+CD28– Τ cells in peripheral blood ofhealthy individuals is a common event. In some subjectsCD28– T cells may represent more than half of the peripheralCD8+ T-cell pool.6,7 Among the factors implicated in thegeneration of peripheral blood CD8+CD28– T cells in healthysubjects, age is the most important and most studied(Table 1). Posnett et al. first described expanded pools ofoligoclonal CD28– T cells among freshly isolated peripheralblood CD8+ T cells of aged humans.79 In another study, Effroset al. reported that centenarians, and long-term T-cell cul-tures, showed a decline in CD28+ T cells.64 Herein after,Fagnoni et al. studying a group of healthy people thatincluded centenarians showed a direct correlation betweenage and the numbers of peripheral blood CD8+CD28–
T cells.85 Although some groups have reported an increase inCD8+CD28– T cells with age;31,85,86 others have not been ableto find such a correlation.87–89 The basis for these discrepan-cies may result from the study of cohorts with differentgenetic backgrounds or from the unforeseen influence ofenvironmental factors89 on the percentage of CD8+ T cells.The only reliable correlation found with age is a decline in(CD8+)CD28+ T cells.31,64,85–90
The reduction in peripheral blood CD8+CD28+ T cells thattakes place with age, along with the accumulation of a pool ofunresponsive CD8+CD28– T cells with shortened telomeres,has led to the concept of replicative senescence.91–94 However,the weak responsiveness of CD8+CD28– T cells to mitogenicstimulation rather than being a feature of their senescencemay be an indication that they are prepared to receive adifferent stimulus. The recent evidence that telomerase activ-ity is intact in T cells from aged people reinforces thisassumption.95 Thus, study of costimulatory moleculesexpressed on activated/memory T cells, such as ICOS, PD-1,LFA-1, 4-1BB,96–98 may give important clues to the functionalstatus of CD8+CD28– T cells. In this context, special attention
should be given to de novo receptors expressed byCD8+CD28– T cells, such as the family of natural killerreceptors.88,99
Studies of diseases in which increased numbers ofCD8+CD28– T cells have been reported may provide insightsinto the mechanisms that drive the emergence of these CD8+
T cells in peripheral blood and into their physiologicalfunction (Table 1). In haemochromatosis (HFE), a geneticdisorder characterized by an inappropriately high iron absorp-tion in the small intestine and its subsequent accumulation inparenchymal cells of the liver, pancreas and other organs,increased numbers of CD8+CD28– T cells are a hallmark ofthe disease.87 Interestingly, follow-up studies showed thatCD8+CD28– T-cell expansions persist over time and affordprotection from development of hepatocellular carcinoma.87,100
Although HFE, the gene product responsible for the disease,is a non-classical MHC-class I molecule,101,102 there is noformal proof for its direct involvement in the activation ofCD8+ T cells. However, the presence of activated and oligo-clonal CD8+ T cells in these patients87,103 is an indication oftheir continuous priming under an iron-overloaded environ-ment. Furthermore, increased numbers of CD8+CD28– T cells,containing expansions of oligoclonal T cells, have also beenreported in the peripheral blood of heavy alcohol drinkerswithout liver disease.89 Interestingly, high CD8+CD28– T-cellnumbers were associated with low levels of liver injury.These data are in accordance with previous studies in heavyalcohol drinkers without liver disease, which showed thatCD8+ T cells have a typical profile of activated/memory cells,such as expression of CD57, CD45RO and HLA-DR.104–106
Together with the data from haemochromatosis patients theseresults suggest that CD8+CD28– T cells may play a protectiveregulatory role under tissue injury caused either by ethanol oriron, and point to an epithelial-driven CD8+CD28– T-cellgeneration. Through the production of reactive oxygenspecies iron and ethanol mediate protein damage and lipidperoxidation in parenchymal hepatocytes and result in epithe-lial cell injury.107–111 From an immunological point of viewthe stressed and/or injured epithelial cells may modulate theexpression of molecules involved, directly or indirectly, insignalling to either resident or passenger CD8+ T lympho-cytes112 (see next section). Repeated CD8+ T-cell activation inthe presence of γchain signalling cytokines may lead to thegeneration of a progeny of CD8+CD28– T cells with protec-tive regulatory functions as discussed elsewhere.113
Increased numbers of peripheral blood CD8+CD28– T cellshave also been reported in chronic diseases (e.g. Crohn’sdisease, rheumatoid arthritis, Felty’s syndrome, systemiclupus erythromatosus and Wegener’s granulomatosis),114–119
in haemophilic and haemodialysis patients,120,121 and in patientswith haematological tumours and immuno-deficiency122–126
(see Table 1). In some of these disease conditions increasednumbers of CD8+CD28– T cells are concomitant with chronicstimulation of CD4+ T cells. For example, in rheumatoidarthritis the increased number of circulating CD8+CD28–
T cells is paralleled by an increase in oligoclonal CD4+CD28–
T cells.127,128 The CD4+CD28– T-cell subset is a rare humanT-cell subset characterized by autoreactivity and resistance toapoptosis,8,9,128–130 which may preferentially originate in thecontext of certain HLA-DR alleles.131 As mentioned previ-ously, the possibility that increased numbers of circulating

Human CD8+CD28– T cells 5
CD8+CD28– T cells in these diseases may result in part fromhomeostatic changes within the peripheral T-cell pool cannotbe ruled out.
Longitudinal studies of peripheral T-cell populations inhealth and in disease have indeed unveiled the existence of aclose relationship between CD8+CD28– T-cell developmentand CD4+ and CD8+ T-cell numbers in peripheral blood. Inother words, it is very likely that a variable fraction of thecirculating pool of CD8+CD28– T cells may, under certaincircumstances, arise without the need of TCR triggering.First, human CD4+ T-cell lymphopenia due to Lck deficiencyresults in the replenishment of the peripheral T-cell pool withCD8+CD28– T cells.132 Second, low CD4/CD8 T-cell ratiosanticipate high percentages of CD8+CD28– T cells in periph-eral blood regardless of age.87,89 Third, the progressive loss ofCD4+ T cells, and naive CD8+ T cells observed in HIV-infected subjects is counteracted by an increase in effector/memory CD8+(CD28–) T cells,133–135 an increase also observedin centenarians in which reductions in naive CD8+ T cells takeplace.136 In all, these data suggest that CD8+CD28– T-celldevelopment is constrained by the size of the peripheral CD8+
T-cell compartment, in relation to the CD4+, and by thenumber of naive CD8+ T cells within this compartment. Thisconstraint is in accord with the genetic control of the periph-eral CD4/CD8 T-cell ratio,137,138 its constancy over time inhumans139,140 and the close relationship observed betweenboth pools in reconstituted mice.141 In this scenario, CD8+
CD28– T cells will expand when a decrease in total CD4+
T cells, or naive CD8+ T cells, takes place within the periph-eral T-cell compartment, regardless of the origin and nature ofthe cause. This increase could be modulated in a positive or anegative manner by the presence or absence, respectively, ofsignalling cytokines both in humans142 and in mice,143–145 orby additional stimuli such as viral infection, iron overloador ethanol consumption. This setting fits well with currentdata indicating that CD8+CD28– T cells divide faster and livelonger than CD8+CD28+ and CD4+ T cells, features likely tobe related to a shorter cell division cycle, a higher resistanceto apoptosis and a different response to regulatorycytokines.65–68,146,147
Are epithelial cells involved in the generation and/or maintenance of CD8+CD28– T cells?
Cells expressing high levels of MHC-class II molecules, suchas dendritic cells, activated monocytes and B cells and tissuemacrophages, are considered the classical APC. However,evidence accumulated during the past decade indicates thatintestinal epithelial cells (iEC) are also capable of activatingT cells.148 Early in vitro studies demonstrated that ex vivo iECfrom rats and humans were capable of activating peripheralblood T cells. Paradoxically, the T cells driven into prolifera-tion by iEC were CD8+ T cells.149,150 It is now known that bothex vivo iEC and iEC tumour cell lines are capable of generat-ing CD8+CD28– T cells in vitro, although using differentmechanisms.150,151 While CD8+CD28– T-cell generation byex vivo iEC involves activation and proliferation of CD8+
T cells, generation by iEC lines entails death of CD4+CD28+
and CD8+CD28+ T cells, thus strengthening the resistance ofCD8+CD28– T cells to apoptosis.151
Evidence gathered during the last few years suggests thatCD1d and gp180 are involved in the activation of humanCD8+ T cells.152–155 CD1d is a non-classical MHC-class Imolecule that is expressed in iEC, parenchymal hepatocytesand stromal cells.156,157 Dendritic cells, monocytes, B cells andactivated T cells express low levels of CD1d.158–160 In con-trast, gp180 is a heavy glycosylated protein that is expressedby the intestinal, thymic and airway epithelium, and introphoblasts.161 While CD1d is thought to interact with theTCR of the CD8+ T cell, gp180 interacts with the CD8molecule and activates its associated tyrosine kinaseLck.152,153 The CD8/gp180 interaction plays an important rolenot only in Lck activation, but also in the adhesion of CD8+
T cells to iEC.151 In this scenario, MHC-class II moleculesappear to modulate the epithelial-driven CD8+ T-cell activa-tion.152 The capability of iEC to activate human CD8+ T cellsin vitro has also been observed with hepatic parenchymalcells,162,163 airway epithelial cells,164 endothelial cells165 andfibroblasts.166 Whether the CD1d/gp180 pair is involved andwhether the proliferating CD8+ T cells become CD28– isunknown, but cocultures of epithelial cell lines from differentorigins with peripheral blood T cells results in accumulationof CD8+CD28– T cells.151
The generation of CD8+CD28– T cells by epithelial cellscould be envisaged as the result of the existence of communi-cation between epithelial tissues and CD8+ T cells. This cross-talk constitutes a unique feature that may help to understandthe phenotypic changes that take place within the peripheralT-lymphocyte system under a variety of situations (Table 1).Thus, altered expression of CD1d by epithelial cells has beenimplicated in the pathogenesis of certain autoimmune dis-orders.170–172 Other studies, however, have suggested thatfaulty expression of gp180, rather than CD1d overexpression,may be responsible for the pathogenesis of certain inflamma-tory diseases.173 On the contrary, when expressed by dendriticcells CD1d seems to preferentially activate CD4+ T cells andCD4–CD8– T cells bearing a conserved Vα chain.174–176 Thepotential of human CD1d to interact and activate oppositeT-cell subsets may reflect presentation of different tissue-specific ligands, as shown in mice,177 or a different physicalassociation with conserved structures. In this context, CD1dhas three distinctive features: (i) a hydrophobic bindinggroove that binds lipid compounds instead of classical pep-tides, in mouse CD1d1;178,179 (ii) the presence of the YQGVmotif in its cytoplasmic tail, a tyrosine-based internalizationsignal that targets the molecule to endosomal compart-ments;180 and (iii) the capability to be expressed in the cellsurface in a β2m-independent manner.181,182 Nevertheless,even though CD1d is considered to be an important signallingmolecule involved in the activation of distinct subsets of Tcells,148,161,167–169,174–176 it must be noted that cell surfaceexpression of CD1d by untransfected human cells is veryweak or absent, and is mainly cytoplasmic. This pattern ofexpression does not meet the classical requirements for anantigen-presenting molecule. Therefore, human CD1d mayhold a specialized biological function that may be intracellu-lar rather than extracellular (for a recent review of CD1d seeJoyce183).
Molecules having the MHC-fold, aside from CD1d, aloneor in combination with conserved monomorphic structuresand/or oxidatively modified proteins are candidate molecules

6 FA Arosa
whose expression on epithelial cells may allow a preferentialCD8+ T-cell activation. Human MICA and MICB molecules(for MHC class I-chain related molecules) are expressed onthe cell surface of stressed iEC and fibroblasts, and have thecapability to activate γδCD8+ T cells.184–186 MICA is alsoexpressed by human keratinocytes, endothelial cells andmonocytes and seems to be expressed in the cell surface in aβ2m-independent manner.187 Although detailed studies ofMICA and MICB expression in the conditions described inTable 1 have not been reported, it is anticipated that theirexpression, along with the expression of other signallingmolecules, could be altered. In this context, increased expres-sion of MHC-class I and ICAM-1 on hepatocytes of patientswith haemochromatosis has been reported.112 By analogy withstudies in humans and mice,188–193 altered expression of con-served molecules and molecules with the MHC-class I fold byepithelial cells under stressed conditions, such as during ironoverload, may activate passenger and/or resident CD8+ T cellsand generate a pool of CD8+CD28– T cells that may act on theinjured epithelial cell immediately or after recirculation. Thecytokines and factors secreted may directly or indirectlyregulate stress/injury and restore normal hepatic function.Although speculative at present, this scenario is not unlikely.Clinical and experimental data accumulated during the past10 years have established the impact that low numbers ofCD8+ T cells, both in peripheral blood and in the liver, havein the clinical setting of haemochromatosis.100,140,194–196
Studies of iron-rich body fluids, such as breast milk and thesynovial fluid of rheumatoid arthritis patients197,198 and of amodel of hepatic injury199 appear to support the assumptionthat CD8+CD28– T cells play a protective role under stressedconditions and thus regulate epithelial physiology.
The majority of T cells populating epithelial tissues inhumans (e.g. small intestine and liver parenchyma) are, likeCD8+CD28– T lymphocytes, phenotypically and functionallymemory CD8+ T cells.31,200–205 Recent work by O’Farrelly andcollaborators203,204 has shown that a large fraction of humanhepatic CD8+ T cells express activating and inhibitory NKreceptors.206 These CD8+ T cells, called NKT cells, are alsopresent in peripheral blood and are characterized by a lack ofCD28 expression.43,88,207,208 Although evidence for the expres-sion of NK receptors by human intestinal intraepitheliallymphocytes (iIEL) is absent, a recent study showed thatBY55, a glycosylphosphatidylinositol-linked protein, isexpressed by both circulating CD8+CD28– T cells and iIEL.209
First described in circulating NK cells, BY55 is expressed bycord blood and bone marrow CD8+ T cells, and in a largefraction of circulating CD8+ T cells in HIV patients.210–212
Therefore, this molecule links the circulating CD8+CD28–
T-cell pool and the mucosal CD8+ T-cell pool, which is alsoCD28–.202,213 In addition, both iIEL and hepatic NK CD8+
T cells lack cytotoxic activity, but gain cytotoxicity afterculture in IL-2,204,214,215 a feature also shared by CD8+CD28–
T cells.19 These phenotypic and functional similarities suggestthat circulating CD8+CD28– T cells may originate fromCD8+CD28+ T-cell precursors that have acquired typical NKreceptors during the differentiation process in epithelial tis-sues, thus, becoming NKT cells, as observed in mice.216,217
It is important to note that NK receptors have also beendescribed in circulating CD4+CD28– T cells from rheumatoidarthritis patients, thus, indicating that induction/acquisition of
NK receptors is linked to the differentiation process of the Tcell.218–220 The likelihood that thymus- and/or bone marrow-derived CD8+ T cells, in the context of signals provided byenvironments such as the liver parenchyma or the intestine,differentiate, lose expression of CD28, transcribe and expresscell surface NK receptors, such as CD161,221 is an intriguing,but as yet, unexplored question.
Conclusions and future prospects
Studies in mice have shown that the maintenance of naive andmemory CD8+ T cells in the periphery requires TCR–MHCinteractions as well as cytokines and growth factors.222
Although similar studies in humans are unfeasible, the nega-tive impact of the absence of MHC-class I molecules orcytokine receptors, such as the γchain, on peripheral T-cellhomeostasis is well known.223,224 Nevertheless, the nature ofthe MHC-class I+ cells that interact with CD8+ T cells in theperipheral tissues and organs and assure their survival isuncertain. Likely candidates are epithelial cells of the intes-tine and liver, two peripheral tissues usually populated byCD8+ T cells. Other organs and tissues, such as bone marrow,can also play an important role. By communicating withepithelial cells, circulating CD8+ T cells could secure not onlytheir own survival but participate in the regulation of basicbiological processes, from modulation of epithelial physio-logy199,225–230 and removal of apoptotic cells231 to the regula-tion of local cell homeostasis.232,233
The primary signals delivered by epithelial tissues to CD8+
T cells can only be envisaged within the context of thecytokines produced in situ;234–237 the nutrients for which CD8+
T cells compete,222 soluble factors and receptors influencingT-cell function and receptivity,238,239 and novel modulators ofT-cell survival.240 Together with the signals delivered whenthe T lymphocyte has changed to the CD28– phenotype,including those provided by engagement of NK receptorssuch as NKG2D via MICA,241–243 this myriad of environmen-tal signals is likely to shape the peripheral T-lymphocytesystem. Classical and non-classical MHC-class I molecules,together with the variety of the NK receptors that recognizethose molecules, are thus central to T-lymphocyte homeo-stasis, namely of the CD8+ T-cell pool.244 These signals areexpected to be delivered in normal conditions (e.g. duringepithelial cell renewal) and in a number of situations duringdegeneration (e.g. senescence), injury (e.g. excess of iron andalcohol, viral infection) or modification (e.g. organ transplan-tation) of the internal environment. In these latter situations,however, a hastened generation of CD8+CD28– T cells wouldtake place. Given their functional heterogeneity and proper-ties, CD8+CD28– T cells are better positioned than other T-cell subsets to counteract these environmental changes. Thisprevalent human CD8+ T-cell subset, contrary to the currentbelief, may encompass highly specialized T cells with thecapability to control and regulate tissue remodelling andrepair within the microenvironment they populate on a day-to-day basis.
Acknowledgements
I thank EMP Cardoso for invaluable suggestions and helpduring the writing of this manuscript, G Porto and M Santos

Human CD8+CD28– T cells 7
for helpful discussions, AM Carmo for his unwavering beliefin the immune system and AM Fonseca for providing the datafor Fig. 1. Last, but not least, I want to thank M de Sousa forher continuous support and for teaching me to look at theimmunological system in a different way. Some of the workdescribed in this review was supported by the PortugueseScience Funding Agency (FCT) and by the American Portu-guese Biomedical Research Fund (APBRF). Regretfully, anddue to space constraints, some original work was notreferenced.
References
1 Rudd CE. Upstream downstream: CD28 co-signaling pathwaysand T cell function. Immunity 1996; 4: 527–34.
2 Lenschow DJ, Walunas TL, Bluestone JA. CD28/B7 system ofT cell costimulation. Annu. Rev. Immunol. 1996; 14: 233–58.
3 Salomon B, Bluestone JA. Complexities of CD28/B7: CTLA-4costimulatory pathways in autoimmunity and transplantation.Annu. Rev. Immunol. 2001; 19: 225–52.
4 Salazar-Fontana LI, Bierer BE. T-lymphocyte coactivator mole-cules. Curr. Opin. Hematol. 2001; 8: 5–11.
5 Hansen JA, Martin PJ, Nowinski RC. Monoclonal antibody iden-tifying a novel T-cell antigen and Ia antigens of human lym-phocytes. Immunogenetics. 1980; 10: 247–52.
6 Yamada H, Martin PJ, Braun MP, Beatty PG, Sadamoto K,Hansen JA. Monoclonal antibody 9.3 and anti-CD11 antibodiesdefine reciprocal subsets of lymphocytes. Eur. J. Immunol. 1985;15: 1164–8.
7 Azuma M, Phillips JH, Lanier LL. CD28– lymphocytes: anti-genic and functional properties. J. Immunol. 1993; 150:1147–59.
8 Morishita Y, Martin PJ, Bean MA, Yamada H, Hansen JA.Antigen-specific functions of a CD4+ subset of human T lym-phocytes with granular morphology. J. Immunol. 1986; 136:2095–102.
9 Morishita Y, Sao H, Hansen JA, Martin PJ. A distinct subset ofhuman CD4+ cells with a limited alloreactive T cell receptorrepertoire. J. Immunol. 1989; 143: 2783–9.
10 Saukkonen JJ, Kornfeld H, Bergman JS. Expansion of aCD8+CD28– cell population in the blood and lung of HIV-positive patients. J. Acquir. Immune Defic. Syndr. 1993; 6:1194–204.
11 Borthwick NJ, Bofill M, Gombert WM et al. Lymphocyteactivation in HIV-1 infection. II. Functional defects of CD28–
T cells. AIDS. 1994; 8: 431–41.12 Choremi-Papadopoulou H, Viglis V, Gargalianos P, Kordossis TK,
Iniotaki-Theodoraki A, Kosmidis J. Downregulation of CD28surface antigen on CD4+ and CD8+ T lymphocytes during HIV-1infection. J. Acquir. Immune Defic. Syndr. 1994; 7: 245–53.
13 Callan MF, Tan L, Annels N et al. Direct visualization ofantigen-specific CD8+ T cells during the primary immuneresponse to Epstein–Barr virus in vivo. J. Exp. Med. 1998; 187:1395–402.
14 Dutra WO, Martins-Filho OA, Cançado JR et al. Chagasicpatients lack CD28 expression on many of their circulating Tlymphocytes. Scand. J. Immunol. 1996; 43: 88–93.
15 Lum LG, Orcutt-Thordarson N, Seigneuret MC, Hansen JA. Invitro regulation of immunoglobulin synthesis by T cell subpopu-lations defined by a new T cell antigen (9.3). Cell. Immunol.1982; 72: 122–31.
16 Damle NK, Mohagheghpour N, Hansen JA, Engleman EG.Alloantigen-specific cytotoxic and suppressor T lymphocytes are
derived from phenotypically distinct precursors. J. Immunol.1983; 131: 2296–300.
17 Koide J, Engleman EG. Differences in surface phenotype andmechanism of action between alloantigen-specific CD8+ cytotoxicand suppressor T cell clones. J. Immunol. 1990; 144: 32–40.
18 Takeuchi T, DiMaggio M, Levine H, Schlossman SF,Morimoto C. CD11 molecule defines two types of suppressor cellswithin the T8+ population. Cell. Immunol. 1988; 111: 398–409.
19 Dianzani U, Zarcone D, Pistoia V et al. CD8+CD11b+ peripheralblood T lymphocytes contain lymphokine-activated killer cellprecursors. Eur. J. Immunol. 1989; 19: 1037–44.
20 Zupo S, Azzoni L, Massara R, D’Amato A, Perussia B,Ferrarini M. Coexpression of Fcγ receptor IIIA and interleukin 2receptor β chain by a subset of human CD3+/CD8+/CD11b+ lym-phocytes. J. Clin. Immunol. 1993; 13: 228–36.
21 Freedman MS, Ruijs TCG, Blain M, Antel JP. Phenotypic andfunctional characteristics of activated CD8+ cells: a CD11b–
CD28– subset mediates noncytolytic functional suppression.Clin. Immunol. Immunopathol. 1991; 60: 254–67.
22 Caruso A, Fiorentini S, Licenziati S et al. Expansion of rareCD8+CD28–CD11b– T cells with impaired effector functions inHIV-1-infected patients. J. Acquir. Immune Defic. Syndr. 2000;24: 465–74.
23 Liu Z, Tugulea S, Cortesini R, Suciu-Foca N. Specific suppres-sion of T helper alloreactivity by allo-MHC class I-restrictedCD8+CD28– T cells. Int. Immunol. 1998; 10: 775–83.
24 Liu Z, Tugulea S, Cortesini R, Lederman S, Suciu-Foca N.Inhibition of CD40 signaling pathway in antigen presenting cellsby T suppressor cells. Hum. Immunol. 1999; 60: 568–74.
25 Li J, Liu Z, Jiang S, Cortesini R, Lederman S, Suciu-Foca N.T suppressor lymphocytes inhibit NF-kappa B-mediated transcrip-tion of CD86 gene in APC. J. Immunol. 1999; 163: 6386–92.
26 Hamann D, Baars PA, Rep MHG et al. Phenotypic and func-tional separation of memory and effector human CD8+ T cells.J. Exp. Med. 1997; 186: 1407–18.
27 Springer TA. Adhesion receptors of the immune system. Nature1990; 346: 425–34.
28 McFarland HI, Nahill SR, Maciaszek JW, Welsh RM. CD11b(Mac-1): a marker for CD8+ cytotoxic T cell activation andmemory in virus infection. J. Immunol. 1992; 149: 1326–33.
29 Christensen JE, Andreasen SO, Christensen JP, Thomsen AR.CD11b expression as a marker to distinguish between recentlyactivated effector CD8+ T cells and memory cells. Int. Immunol.2001; 13: 593–600.
30 Hynes RO. Integrins: versatility, modulation, and signaling incell adhesion. Cell. 1992; 69: 11–25.
31 Hoshino T, Yamada A, Honda J et al. Tissue-specific distribu-tion and age-dependent increase of human CD11b+ T cells.J. Immunol. 1993; 151: 2237–46.
32 Fiorentini S, Licenziati S, Alessandri G et al. CD11b expressionidentifies CD8+CD28+ T lymphocytes with phenotype and func-tion of both naïve/memory and effector cells. J. Immunol. 2001;166: 900–7.
33 Abo T, Balch CM. A differentiation antigen of human NK and Kcells identified by a monoclonal antibody (HNK-1). J. Immunol.1981; 127: 1024–9.
34 Lanier LL, Loken MR. Human lymphocyte subpopulationsidentified using three colour immunofluorescence and flowcytometry analysis. correlation of Leu-2, Leu-7, Leu-8, andLeu-11. J. Immunol. 1984; 132: 151–6.
35 Dupuy d’Angeac A, Monier S, Pilling D, Travaglio-Encinoza A,Rème T, Salmon M. CD57+ T lymphocytes are derived fromCD57– precursors by differentiation occurring in late immuneresponses. Eur. J. Immunol. 1994; 24: 1503–11.

8 FA Arosa
36 Wang ECY, Taylor-Wiedeman J, Perera P, Fisher J,Borysiewicz LK. Subsets of CD8+, CD57+ cells in normal,healthy individuals: correlations with HCMV carrier status, phe-notypic and functional analyses. Clin. Exp. Immunol. 1993; 94:297–305.
37 Labalette M, Salez F, Pruvot FR, Noel C, Dessaint JP. CD8 lym-phocytosis in primary cytomegalovirus (CMV) infection of allo-graft recipients: expansion of uncommon CD8+CD57– subsetand its progressive replacement by CD8+CD57– T cells. Clin.Exp. Immunol. 1994; 95: 465–71.
38 Hamann D, Roos MTL, van Lier RAW. Faces and phases ofhuman CD8+ T cell development. Immunol. Today 1999; 20:177–80.
39 Jungalwala FB. Expression and biological functions of sulfoglu-curonyl glycolipids (SGGLs) in the nervous system – A review.Neurochem. Res. 1995; 19: 945–57.
40 Abo T, Miller CA, Gartland GL, Balch CM. Differentiationstages of human natural killer cells in lymphoid tissues from fetalto adult life. J. Exp. Med. 1983; 157: 273.
41 Clark P, Normansell DE. Phenotypic analysis of lymphocytesubsets in normal human bone marrow. Am. J. Clin. Pathol.1990; 94: 632–6.
42 Liu CC, Walsh CM, Young JD. Perforin: structure and function.Immunol. Today 1995; 16: 194–201.
43 Pittet MJ, Speiser DE, Valmori D, Cerottini JC, Romero P.Cutting edge: cytolytic effector function in human circulatingCD8+ T cells closely correlates with CD56 surface expression.J. Immunol. 2001; 164: 1148–52.
44 Wang ECY, Moss PAH, Frodsham P, Lehner PJ, Bell JI,Borysiewicz LK. CD8highCD57+ T lymphocytes in normal,healthy individuals are oligoclonal and respond to humancytomegalovirus. J. Immunol. 1995; 155: 5046–56.
45 Fiorentino S, Dalod M, Olive D, Guillet JG, Gomard E. Predom-inant involvement of CD8+CD28– lymphocytes in humanimmunodeficiency virus-specific cytotoxic activity. J. Virol.1996; 70: 2022–6.
46 Weekes MP, Carmichael AJ, Wills MR, Mynard K, Sissons JGP.Human CD28–CD8+ T cells contain greatly expanded functionalvirus-specific memory CTL clones. J. Immunol. 1999; 162:7569–77.
47 Dalod M, Dupuis M, Deschemin JC et al. Weak anti-HIV CD8+T-cell effector activity in HIV primary infection. J. Clin. Invest.1999; 104: 1431–9.
48 Trimble LA, Lieberman J. Circulating CD8 T lymphocytes inhuman immunodeficiency virus-infected individuals have impairedfunction and downmodulate CD3 zeta, the signaling chain of theT-cell receptor complex. Blood 1998; 91: 585–94.
49 Trimble LA, Kam LW, Friedman RS, Xu Z, Lieberman J.CD3zeta and CD28 down-modulation on CD8 T cells duringviral infection. Blood 2000; 96: 1021–9.
50 Leroy E, Calvo CF, Divine M et al. Persistence of T8+/HNK-1+suppressor lymphocytes in the blood of long term survivingpatients after allogeneic bone marrow transplantation. J. Immu-nol. 1986; 137: 2180–9.
51 Fregona I, Guttman RD, Jean R. HNK1+ (Leu-7) and other lym-phocyte subsets in long-term survivors with renal transplant.Transplantation 1985; 39: 25–9.
52 Maher P, O’Toole CM, Wreghitt TG, Spiegelhalter DJ,English TA. Cytomegalovirus infection in cardiac transplantrecipients associated with chronic T cell subset ratio inversionwith expansion of a Leu-7+ TS-C+ subset. Clin. Exp. Immunol.1985; 62: 515–23.
53 Ogg GS, McMichael AJ. HLA-peptide tetrameric complexes.Curr. Opin. Immunol. 1998; 10: 393–6.
54 Kelleher AD, Rowland-Jones SL. Functions of tetramers-stainedHIV-specific CD4+ and CD8+ T cells. Curr. Opin. Immunol.2000; 12: 370–4.
55 Altman J, Moss PAH, Goulder P et al. Direct visualization andphenotypic analysis of virus-specific T lymphocytes in HIV-infected individuals. Science 1996; 274: 94–6.
56 Gillespie GMA, Wills MR, Appay V et al. Functional hetero-geneity and high frequencies of cytomegalovirus-specific CD8+T lymphocytes in healthy seropositive donors. J. Virol. 2000; 74:8140–50.
57 Appay V, Nixon DF, Donahoe SM et al. HIV-specific CD8+T cells produce antiviral cytokines but are impaired in cytolyticfunction. J. Exp. Med. 2000; 192: 63–75.
58 Lim DG, Bieganowska K, Freeman GJ, Hafler DA. Examinationof CD8+ T cell function in humans using MHC class I tetramers:similar cytotoxicity but variable proliferation and cytokine pro-duction among different clonal CD8+ T cells specific to singleviral epitope. J. Immunol. 2000; 165: 6214–20.
59 Tussey L, Speller S, Gallimore A, Vessey R. Functionally dis-tinct memory T cell subsets in persistent EBV infection aredifferentiated by migratory receptor expression. Eur. J. Immunol.2000; 30: 1823–9.
60 Alcover A, Alarcon B. Internalization and intracellular fate ofTCR-CD3 complexes. Crit. Rev. Immunol. 2000; 20: 325–46.
61 Lake RA, O’Hehir RE, Verhoef A, Lamb JR. CD28 mRNArapidly decays when activated T cells are functionally anergizedwith specific peptide. Int. Immunol. 1993; 5: 461–6.
62 Linsley PS, Bradshaw J, Urnes M, Grosmaire L, Ledbetter J.CD28 engagement by B7/BB1 induces transient downregulationof CD28 synthesis and prolonged unresponsiveness to CD28 sig-naling. J. Immunol. 1993; 150: 3161–9.
63 Eck SC, Chang D, Wells AD, Turka LA. Differential downregu-lation of CD28 by B7–1 and B7–2 engagement. Transplantation1997; 64: 1497–9.
64 Effros RB, Boucher N, Porter V et al. Decline in CD28+ T cellsin centenarians and in long-term T cell cultures: a possible causefor both in vivo and in vitro immunosenescence. Exp. Gerontol.1994; 9: 601–9.
65 Posnett DN, Edinger JW, Manavalan JS, Irwin C, Marodon G.Differentiation of human CD8 T cells. implications for in vivopersistence of CD8+CD28– cytotoxic effector clones. Int. Immu-nol. 1999; 11: 229–41.
66 Fiorentini S, Malacarne F, Ricotta D et al. Generation of CD28–
cells from long term-stimulated CD8+CD28+ T cells: a possiblemechanism accounting for the increased number of CD8+CD28–
T cells in HIV-1-infected patients. J. Leukoc. Biol. 1999; 65:641–8.
67 Borthwick NJ, Lowdell M, Salmon M, Akbar AN. Loss of CD28expression on CD8+ T cells is induced by IL-2 receptor gammachain signalling cytokines and type I IFN, and increases suscep-tibility to activation-induced apoptosis. Int. Immunol. 2000; 12:1005–13.
68 Labalette M, Leteurtre E, Thumerelle C, Grutzmacher C, Tour-vieille B, Dessaint JP. Peripheral human CD8+CD28+ T lym-phocytes give rise to CD28– progeny, but IL-4 prevents loss ofCD28 expression. Int. Immunol. 1999; 11: 1327–36.
69 Harley CB. Telomere loss: mitotic clock or genetic timebomb?Mutat. Res. 1991; 256: 271–82.
70 Aspinall R. Does the immune system of a mouse age faster thanthe immune system of a human? Bioessays 1999; 21: 519–24.
71 Monteiro J, Batliwalla F, Ostrer H, Gregersen PK. Shortenedtelomeres in clonally expanded CD28-CD8+ T cells imply a rep-licative history that is different from their CD28+CD8+ counter-parts. J. Immunol. 1996; 156: 3587–90.

Human CD8+CD28– T cells 9
72 Effros RB, Allsopp R, Chiui CP et al. Shortened telomeres inthe expanded CD28–CD8+ cell subset in HIV disease implicatereplicative senescence in HIV pathogenesis. AIDS 1996; 10:F17–F22.
73 Honda M, Mengesha E, Albano S et al. Telomere shortening anddecreased replicative potential, contrasted by continued prolifer-ation of telomerase-positive CD8+CD28lo T cells in patients withsystemic lupus erythematosus. Clin. Immunol. 2001; 99:191–201.
74 Speiser DE, Migliaccio M, Pittet MJ et al. Human CD8(+) Tcells expressing HLA-DR and CD28 show telomerase activityand are distinct from cytolytic effector T cells. Eur. J. Immunol.2001; 31: 459–66.
75 Plunkett FJ, Soares MVD, Annels N et al. The flow cytometricanalysis of telomere length in antigen-specific CD8+ T cellsduring acute Epstein–Barr virus infection. Blood 2001; 97:700–7.
76 Rufer N, Brummendorf TH, Kolvraa S et al. Telomere fluores-cence measurements in granulocytes and T lymphocyte subsetspoint to a high turnover of hematopoietic stem cells and memoryT cells in early childhood. J. Exp. Med. 1999; 190: 157–67.
77 Zeichner SL, Palumbo P, Feng YR et al. Rapid telomere short-ening in children. Blood 1999; 93: 2824–30.
78 Vallejo AB, Brandes JC, Weyand CM, Goronzy JJ. Modulationof CD28 expression: distinct regulatory pathways during activa-tion and replicative senescence. J. Immunol. 1999; 162: 6572–9.
79 Posnett DN, Sinha R, Kabak S, Russo C. Clonal populations ofT cells in normal elderly humans: the T cell equivalent to‘benign monoclonal gammapathy’. J. Exp. Med. 1994; 179:609–18.
80 Fitzgerald JE, Ricalton NS, Meyer AC et al. Analysis of clonalCD8+ T cell expansions in normal individuals and patients withrheumatoid arthritis. J. Immunol. 1995; 154: 3538–47.
81 Morley JK, Batliwalla FM, Hingorani R, Gregersen PK. Oligo-clonal CD8+ T cells are preferentially expanded in the CD57+
subset. J. Immunol. 1995; 154: 6182–90.82 Batliwalla F, Monteiro J, Serrano D, Gregersen PK. Oligo-
clonality of CD8+ T cells in health and disease: aging, infection,or immune regulation? Hum. Immunol. 1996; 48: 68–72.
83 Batliwalla FM, Rufer N, Lansdorp PM, Gregersen PK. Oligo-clonal expansions in the CD8+CD28– T cells largely explain theshorter telomeres detected in this subset: analysis by flow FISH.Hum. Immunol. 2000; 61: 951–8.
84 Mugnaini EN, Egeland T, Spurkland A, Brinchmann JE. The Tcell receptor repertoire of CD8+CD28– T lymphocytes is domi-nated by expanded clones that persist over time. Clin. Exp.Immunol. 1999; 117: 298–303.
85 Fagnoni FF, Vescovini R, Mazzola M et al. Expansion of cyto-toxic CD8+CD28– T cells in healthy ageing people, includingcentenarians. Immunology 1996; 88: 501–7.
86 Nociari MM, Telford W, Russo C. Postthymic development ofCD28–CD8+ T cell subset: age-associated expansion and shift frommemory to naïve phenotype. J. Immunol. 1999; 162: 3327–35.
87 Arosa FA, Oliveira L, Porto G et al. Anomalies of the CD8+ Tcell pool in hemochromatosis: HLA-A3-linked expansions ofCD8+CD28– T cells. Clin. Exp. Immunol. 1997; 107: 548–54.
88 Speiser DE, Valmori D, Rimoldi D et al. CD28-negative cyto-lytic effector T cells frequently express NK receptors and arepresent at variable proportions in circulating lymphocytes fromhealthy donors and melanoma patients. Eur. J. Immunol. 1999;29: 1990–9.
89 Arosa FA, Porto G, Cabeda JM et al. Expansions of CD8+CD28–
and CD8+TcRVβ5.2+ T cells in peripheral blood of alcoholheavy drinkers. Alcohol Clin. Exp. Res. 2000; 214: 519–27.
90 Merino J, Martinez-Gonzalez MA, Rubio M, Inoges S, Sanchez-Ibarrola A, Subira ML. Progressive decrease of CD8high+ CD28+CD57- cells with ageing. Clin. Exp. Immunol. 1998; 112: 48–51.
91 Effros RB. Loss of CD28 expression on T lymphocytes: amarker of replicative senescence. Dev. Comp. Immunol. 1997;21: 471–8.
92 Effros RB. Replicative senescence in the immune system:impact of the Hayflick limit on T-cell function in the elderly. Am.J. Hum. Genet. 1998; 62: 1003–7.
93 Globerson A, Effros RB. Ageing of lymphocytes and lym-phocytes in the aged. Immunol. Today 2000; 21: 515–21.
94 Pawelec G. Hypothesis: loss of telomerase inducibility and sub-sequent replicative senescence in cultured human T cells is aresult of altered costimulation. Mech. Ageing Dev. 2000; 121:181–5.
95 Son NH, Murray S, Yanovski J, Hodes RJ, Weng NP. Lineage-specific telomere shortening and unaltered capacity for telomereexpression in human T and B lymphocytes with age. J. Immunol.2000; 165: 1191–6.
96 Watts TH, DeBenedette MA. T cell costimulatory moleculesother than CD28. Curr. Opin. Immunol. 1999; 11: 286–93.
97 Vinay DS, Kwon BS. Role of 4-1BB in immune responses.Semin. Immunol. 1998; 10: 481–9.
98 Lens SMA, Tesselaar K, van Oers MHJ, van Lier RAW. Controlof lymphocyte function through CD27–CD70 interaction. Semin.Immunol. 1998; 10: 491–9.
99 Barten R, Torkar M, Haude A, Trowsdale J, Wilson MJ. Diver-gent and convergent evolution of NK-cell receptors. TrendsImmunol. 2001; 22: 52–7.
100 De Sousa M, Porto G, Arosa FA et al. T lymphocyte expressionand function in hemochromatosis. In: Barton JC, Edwards C(eds). Hemochromatosis. London: Cambridge University Press,2000; 396–407.
101 Feder JN, Gnirke A, Thomas W et al. A novel MHC class I-likegene is mutated in patients with hereditary haemochromatosis.Nat. Genet. 1996; 13: 399–408.
102 Braud VM, Allan DSJ, McMichael AJ. Functions of nonclassi-cal MHC and non-MHC-encoded class I molecules. Curr. Opin.Immunol. 1999; 11: 100–8.
103 Cardoso C, Porto G, Lacerda R et al. T cell receptor repertoire inhereditary hemochromatosis: a study of 32 hemochromatosispatients and 274 healthy subjects. Hum. Immunol. 2001; 62:488–99.
104 Cook RT, Garvey MJ, Booth BM, Goeken JA, Stewart B,Noel M. Activated CD8 cells and HLA-DR expression in alco-holics without overt liver disease. J. Clin. Immunol. 1991; 11:246–53.
105 Cook RT, Ballas ZK, Waldschmidt TJ, Vandersteen D,LaBrecque DR, Cook BL. Modulation of T cell adhesion mark-ers, and the CD45R and CD57 antigens in human alcoholics.Alcohol Clin. Exp. Res. 1995; 19: 555–63.
106 Cook RT. Alcohol abuse, alcoholism, and damage to the immunesystem. A review. Alcohol Clin. Exp. Res. 1998; 21: 974–80.
107 Kamimura S, Gaal K, Britton RS, Bacon BR, Triadafipoulos G,Tsukamoto H. Increased 4-hydroxynonenal levels in experimen-tal alcoholic liver disease: association of lipid peroxidation withliver fibrogenesis. Hepatology 1992; 16: 448–53.
108 Niederau C, Stremmel W, Strohmeyer GWW. Clinical spectrumand management of haemochromatosis. Ballière’s Clin. Haema-tol. 1994; 7: 881–901.
109 Niemela O, Parkkila S, Yla-Herttuala S, Villanueva J,Ruebner B, Halsted CH. Sequential acetaldehyde production,lipid peroxidation, and fibrogenesis in micropig model ofalcohol-induced liver disease. Hepatology 1995; 22: 1208–14.

10 FA Arosa
110 Tsukamoto H, Horne W, Kamimura S et al. Experimental livercirrhosis induced by alcohol and iron. J. Clin. Invest. 1995; 96:620–30.
111 Houglum K, Ramm GA, Crawford DHG, Witztum JL, Powell LW,Chojkier M. Excess iron induces hepatocyte oxidative stress andtransforming growth factor β1 in genetic hemochromatosis.Hepatology 1997; 26: 605–12.
112 Stal P, Broome U, Scheynius A, Befrits R, Hultcrantz R.Kupffer cell iron overload induces intercellular adhesionmolecule-1 expression on hepatocytes in genetic hemochromato-sis. Hepatology 1995; 21: 1308–16.
113 Arosa FA. Biochemical and functional characterization ofhuman peripheral blood CD8+ T lymphocytes: Hemochromato-sis as a model. PhD thesis. 1999. University of Porto, Portugal(unpubl.).
114 Neil GA, Summers RW, Cheyne BA, Carpenter C, Huang W,Waldschmidt TJ. Analysis of T-lymphocyte subpopulations ininflammatory bowel diseases by three-color flow cytometry.Digest. Dis. Sci. 1994; 39: 1900–8.
115 Dupuy d’Angeac A, Monier S, Jorgensen C et al. Increased per-centage of CD3+, CD57+ lymphocytes in patients with rheuma-toid arthritis: correlation with duration of disease. Arthritis.Rheum. 1993; 36: 608–12.
116 Bowman SJ, Bhavnani M, Geddes GC et al. Large granular lym-phocyte expansions in patients with Felty’s syndrome: analysisusing anti-T cell receptor Vβ-specific monoclonal antibodies.Clin. Exp. Immunol. 1995; 101: 18–24.
117 Kaneko H, Saito K, Hashimoto H, Yagita H, Okumura K,Azuma M. Preferential elimination of CD28+ T cells in systemiclupus erythematosus (SLE) and the relation with activation-induced apoptosis. Clin. Exp. Immunol. 1996; 106: 218–29.
118 Ikeda M, Watanabe Y, Kitahara S, Inouye T. Distinctiveincreases in HLA-DR+ and CD8+CD57+ lymphocyte subsets inWegener’s granulomatosis. Int. Arch. Allergy Immunol. 1993;102: 205–12.
119 Moosig F, Csernok E, Wang G, Gross WL. Costimulatory mole-cules in Wegener’s granulomatosis (WG). lack of expression ofCD28 and preferential up-regulation of its ligands B7–1 (CD80)and B7–2 (CD86) on T cells. Clin. Exp. Immunol. 1998; 114:113–18.
120 Dianzani U, Pileri A, Bianchi A, Camponi A, Tamponi G,Massaia M. Biochemical and immunologic abnormalities inperipheral blood T lymphocytes of patients with hemophilia A.Eur. J. Haematol. 1988; 41: 334–40.
121 Nishimoto A, Matsumoto Y. Increase of peripheral natural killerT cells in hemodialysis patients. Clin. Nephrol. 2001; 55: 121–6.
122 Walchner M, Wick M. Elevation of CD8+CD11b+Leu-8– T cellsis associated with the humoral immunodeficiency in myelomapatients. Clin. Exp. Immunol. 1997; 109: 310–16.
123 Serrano D, Becker K, Cunningham-Rundles C, Mayer L. Char-acterization of the T cell receptor repertoire in patients withcommon variable immunodeficiency. Oligoclonal expansion ofCD8+ T cells. Clin. Immunol. 2000; 97: 248–528.
124 Van den Hove LE, Vandenberghe P, Van Gool SW et al. Periph-eral blood lymphocyte subset shifts in patients with untreatedhematological tumors: evidence for systemic activation of theT cell compartment. Leukoc. Res. 1998; 22: 175–84.
125 Montagna D, Arico M, Montini E, De Benedetti F, Maccario R.Identification of HLA-unrestricted CD8+/CD28- cytotoxic T-cell clones specific for leukemic blasts in children with acuteleukemia. Cancer Res. 1995; 55: 3835–9.
126 Raitakari M, Brown RD, Sze D et al. T-cell expansions inpatients with multiple myeloma have phenotype of cytotoxic Tcells. Br. J. Haematol. 2000; 110: 203–9.
127 Imberti L, Sottini A, Signorini S, Gorla R, Primi D. OligoclonalCD4+CD57+ T-cell expansions contribute to the imbalanced T-cell receptor repertoire of rheumatoid arthritis patients. Blood1997; 89: 2822–32.
128 Schmidt D, Goronzy JJ, Weyand CM. CD4+CD7–CD28– T cellsare expanded in rheumatoid arthritis and are characterized byautoreactivity. J. Clin. Invest. 1996; 97: 2027–37.
129 Schirmer M, Vallejo AN, Weyand CM, Goronzy JJ. Resistanceto apoptosis and elevated expression of Bcl-2 in clonallyexpanded CD4+CD28– T cells from rheumatoid arthritispatients. J. Immunol. 1998; 161: 1018–25.
130 Vallejo AN, Schirmer M, Weyand CM, Goronzy JJ. Clonalityand longevity of CD4+CD28null T cells are associated withdefects in apoptotic pathways. J. Immunol. 2000; 165: 6301–7.
131 Chapman A, Stewart SJ, Nepom GT et al. CD11b+CD28–CD4+
human T cells. Activation requirements and association withHLA-DR alleles. J. Immunol. 1996; 157: 4771–80.
132 Goldman FD, Ballas ZK, Schutte BC et al. Defective expressionof p56lck in an infant with severe combined immunodeficiency.J. Clin. Invest. 1998; 102: 421–9.
133 Caruso A, Licenziati S, Canaris AD et al. Contribution of CD4+,CD8+CD28+, and CD8+CD28– T cells to CD3+ lymphocytehomeostasis during the natural course of HIV-1 infection. J.Clin. Invest. 1998; 101: 137–44.
134 Rabin RL, Roederer M, Maldonado Y, Petru A, Herzenberg LA,Herzenberg LA. Altered representation of naïve and memoryCD8 T cell subsets in HIV-infected children. J. Clin. Invest.1995; 95: 2054–60.
135 Roederer M, Dubs JG, Anderson MT, Raju PA, Herzenberg LA.CD8 naïve T cell counts decrease progressively in HIV-infectedadults. J. Clin. Invest. 1995; 95: 2061–6.
136 Fagnoni FF, Vescovini R, Passeri G et al. Shortage of circulat-ing naïve CD8+ T cells provides new insights on immuno-deficiency in aging. Blood 2000; 95: 2860–8.
137 Amadori A, Zamarchi R, de Silvestro G et al. Genetic control ofthe CD4/CD8 T-cell ratio in humans. Nat. Med. 1995; 1:1279–83.
138 Kraal G, Weissman IL, Butcher EC. Genetic control of T-cellsubset representation in inbred mice. Immunogenetics 1983; 18:585–92.
139 Reimão R, Porto G, de Sousa M. Stability of CD4/CD8 ratios inman: new correlation between CD4/CD8 profiles and iron over-load in idiopathic haemochromatosis patients. C. R. Acad. Sci.1991; 313: 481–7.
140 Porto G, Vicente C, Teixeira MA et al. Relative impact of HLAphenotype and CD4-CD8 ratios on the clinical expression ofhemochromatosis. Hepatology 1997; 25: 397–402.
141 Rocha B, Dautigny N, Pereira P. Peripheral T lymphocytes:expansion potential and homeostatic regulation of pool sizes andCD4/CD8 ratios in vivo. Eur. J. Immunol. 1989; 19: 905–11.
142 Gollob JA, Schnipper CP, Orsini E et al. Characterization of anovel subset of CD8+ T cells that expands in patients receivinginterleukin 12. J. Clin. Invest. 1998; 102: 561–75.
143 Kieper WC, Prlic M, Schmidt CS, Mescher MF, Jameson SC.IL-12 enhances CD8 T cell homeostatic expansion. J. Immunol.2001; 166: 5515–21.
144 Tough DF, Zhang X, Sprent J. An IFN-γ-dependent pathwaycontrols stimulation of memory phenotype CD8+ T cell turnoverin vitro by IL-12, IL-18, and IFN-γ. J. Immunol. 2001; 166:6007–11.
145 Wong P, Pamer EG. Cutting edge: antigen-independent CD8T cell proliferation. J. Immunol. 2001; 166: 5864–8.
146 Borthwick NJ, Bofill M, Hassan I et al. Factors influencing acti-vated CD8+ T cell apoptosis in patients with acute herpesvirus

Human CD8+CD28– T cells 11
infection: loss of costimulatory molecules CD28, CD5 and CD6but relative maintenance of Bax and Bcl-X expression. Immunol-ogy 1996; 88: 508–15.
147 Mackall CL, Fleisher TA, Brown MR et al. Distinctionsbetween CD8+ and CD4+ T-cell regenerative pathways result inprolonged T-cell subset imbalance after intensive chemotherapy.Blood 1997; 89: 3700–7.
148 Hershberg RM, Mayer LF. Antigen processing and presentationby intestinal epithelial cells – polarity and complexity. Immunol.Today 2000; 21: 123–8.
149 Bland PW, Warren LG. Antigen presentation by epithelial cellsof the rat small intestine. II. Selective induction of suppressorT cells. Immunology 1986; 58: 9–14.
150 Mayer L, Shlien R. Evidence for function of Ia molecules on gutepithelial cells in man. J. Exp. Med 1987; 166: 1471–83.
151 Arosa FA, Irwin C, Mayer L, de Sousa M, Posnett DN. Inter-actions between peripheral blood CD8 T lymphocytes andintestinal epithelial cells (iEC). Clin. Exp. Immunol. 1998; 112:226–36.
152 Panja A, Blumberg RS, Balk SP, Mayer L. CD1d is involved inT cell–intestinal epithelial cell interactions. J. Exp. Med. 1993;178: 1115–19.
153 Li Y, Yio XY, Mayer L. Human intestinal epithelial cell-induced CD8+ T cell activation is mediated through CD8 and theactivation of CD8-associated p56lck. J. Exp. Med. 1995; 182:1079–88.
154 Yio XY, Mayer L. Characterization of a 180 kDa intestinal epi-thelial cell membrane glycoprotein, gp180. A candidate mole-cule mediating T cell–epithelial interactions. J. Biol. Chem.1997; 272: 12786–92.
155 Campbell NA, Kim HS, Blumberg RS, Mayer L. The nonclassi-cal class I molecule CD1d associates with the novel CD8 ligandgp180 on intestinal epithelial cells. J. Biol. Chem. 1999; 274:26259–665.
156 Blumberg RS, Terhorst C, Bleicher P et al. Expression of a non-polymorphic MHC class I-like molecule, CD1D, by humanintestinal epithelial cells. J. Immunol. 1991; 147: 2518–24.
157 Canchis PW, Bhan AK, Landau SB, Yang L, Balk SP,Blumberg RS. Tissue distribution of the non-polymorphic majorhistocompatibility complex class I-like molecule, CD1d. Immu-nology 1993; 80: 561–5.
158 Yang OO, Racke FK, Nguyen PT et al. CD1d on myeloiddendritic cells stimulates cytokine secretion from and cyto-lytic activity of V alpha 24J alpha Q T cells: a feedbackmechanism for immune regulation. J. Immunol. 2000; 165:3756–62.
159 Exley M, Garcia J, Wilson SB et al. CD1d structure and regula-tion on human thymocytes, peripheral blood T cells, B cells andmonocytes. Immunology 2000; 100: 37–47.
160 Salamone MC, Rabinovich GA, Mendiguren AK, SalamoneGV, Fainboim L. Activation-induced expression of CD1dantigen on mature T cells. J. Leukoc. Biol. 2001; 69: 207–14.
161 Campbell N, Yio XY, So LP, Li Y, Mayer L. The intestinalepithelial cell: processing and presentation of antigen to themucosal immune system. Immunol. Rev. 1999; 172: 315–24.
162 Bumgardner GL, Chen S, Almond PS, Bach FH, Ascher NL,Matas AJ. Cell subsets responding to purified hepatocytes andevidence of indirect recognition of hepatocyte major histo-compatibility complex class I antigen. II. In vitro-generated‘memory’ cells to class I+ class II– hepatocytes. Transplantation1992; 53: 863–8.
163 Volpes R, van den Oord JJ, Desmet VJ. Can hepatocytes serveas ‘activated’ immunomodulating cells in the immune response?J. Hepatol. 1992; 16: 228–40.
164 Kalb TH, Yio XY, Mayer L. Human airway epithelial cells stim-ulate T-lymphocyte Lck and Fyn tyrosine kinase. Am. J. Respir.Cell Mol. Biol. 1997; 17: 561–70.
165 Dengler TJ, Pober JS. Human vascular endothelial cells stimu-late memory but not naive CD8+ T cells to differentiate intoCTL retaining an early activation phenotype. J. Immunol. 2000;164: 5146–55.
166 Roberts AI, Nadler SC, Ebert EC. Mesenchymal cells stimulatehuman intestinal intraepithelial lymphocytes. Gastroenterology1997; 113: 144–50.
167 Balk SP, Ebert EC, Blumenthal RL et al. Oligoclonal expansionand CD1 recognition by human intestinal intraepithelial lym-phocytes. Science 1991; 253: 1411–15.
168 Mayer L. Current concepts in mucosal immunity I. Antigenpresentation in the intestine. new rules and regulations. Am. J.Physiol. 1998; 274: G7–G9.
169 Mayer L. Current concepts in mucosal immunity II. One size fitsall: nonclassical MHC molecules fulfill multiple roles in epithe-lial cell function. Am. J. Physiol. 1998; 274: G227–G231.
170 Tsuneyama K, Yasoshima M, Harada K et al. Increased CD1dexpresion on small bile duct epithelium and epithelioid granu-loma in livers in primary biliary cirrhosis. Hepatology. 1998; 28:620–3.
171 Page MJ, Poritz LS, Tilberg AF et al. CD1d-restricted cellularlysis by peripheral blood lymphocytes: relevance to the inflam-matory bowel disease. J. Surg. Res. 2000; 92: 214–21.
172 Bonish B, Jullien D, Dutronc Y et al. Overexpression of CD1dby keratinocytes in psoriasis and CD1d-dependent IFN-γ produc-tion by NK-T cells. J. Immunol. 2000; 165: 4076–85.
173 Toy LS, Yio XY, Lin AL, Honig S, Mayer L. Defective expres-sion of gp180, a novel CD8 ligand on intestinal epithelial cells,in inflammatory bowel disease. J. Clin. Invest. 1997; 100:2026–71.
174 Exley M, Garcia J, Balk SP, Porcelli S. Requirements for CD1drecognition by human invariant Vα24+ CD4–CD8– T cells. J.Exp. Med. 1997; 186: 109–20.
175 Takahashi T, Nieda M, Koezuka Y et al. Analysis of humanVα24+ CD4+ NKT cells activated by α-glycosylceramide-pulsedmonocyte-derived dendritic cells. J. Immunol. 2000; 164:4458–64.
176 Yang OO, Racke FK, Nguyen PT et al. CD1d on myeloid den-dritic cells stimulates cytokine secretion from and cytolyticactivity of Vα24JαQ T cells: a feedback mechanism for immuneregulation. J. Immunol. 2000; 165: 3756–62.
177 Park SH, Roark JH, Bendelac A. Tissue-specific recognition ofmouse CD1 molecules. J. Immunol. 1998; 160: 3128–34.
178 Zeng ZH, Castano AR, Segelke BW et al. Crystal structure ofmouse CD1d: an MHC-like fold with a large hydrophobicbinding groove. Science 1997; 277: 339–45.
179 Joyce S, Woods AS, Yewdell JW et al. Natural ligand of mouseCD1d1: cellular glycosylphosphatidylinositol. Science 1998;279: 1541–4.
180 Rodionov DG, Nordeng TW, Pedersen K, Balk SP, Bakke O. Acritical tyrosine residue in the cytoplasmic tail is important forCD1d internalization but not for its basolateral sorting in MDCKcells. J. Immunol. 1999; 162: 1488–95.
181 Balk SP, Burke S, Polischuk JE et al. β2-microglobulin-independent MHC class Ib molecule expressed by humanintestinal epithelium. Science 1994; 265: 259–65.
182 Kim HS, Garcia J, Exley M et al. Biochemical characterizationof CD1d expression in the absence of beta2-microglobulin. J.Biol. Chem. 1999; 274: 9289–95.
183 Joyce S. CD1d and natural T cells: how their properties jump-start the immune system. Cell. Mol. Life Sci. 2001; 58: 442–69.

12 FA Arosa
184 Groh V, Bahram S, Bauer S, Herman A, Beauchamp M, Spies T.Cell stress-regulated human major histocompatibility complexclass I gene expressed in gastrointestinal epithelium. Proc. NatlAcad. Sci. USA 1996; 93: 12445–50.
185 Groh V, Steinle A, Bauer S, Spies T. Recognition of stress-induced MHC molecules by intestinal epithelial γδ T cells.Science 1998; 279: 1737–9.
186 Groh V, Rhinehart R, Secrist H et al. Broad tumor-associatedexpression and recognition by tumor-derived γδ T cells of MICAand MICB. Proc. Natl Acad. Sci. USA 1999; 96: 6879–84.
187 Zwirner NW, Fernandez-Vina MA, Stastny P. MICA, a newpolymorphic HLA-related antigen, is expressed mainly by kerat-inocytes, endothelial cells, and monocytes. Immunogenetics1998; 47: 139–48.
188 Paroli M, Carloni G, Franco A, De Petrillo G, Alfani E,Barnaba V. Human hepatoma cells expressing HLA class I mole-cules stimulate primary responses of purified CD8+ T lym-phocytes. Res. Virol. 1993; 144: 327–32.
189 Paroli M, Carloni G, Franco A et al. Human hepatoma cellsexpressing MHC antigens display accessory cell function:dependence on LFA-1/ICAM−1 interaction. Immunology 1994;82: 215–21.
190 Bertolino P, Trescol-Biemont MC, Rabourdin-Combe C. Hepa-tocytes induce functional activation of naive CD8+ T lym-phocytes but fail to promote survival. Eur. J. Immunol. 1998; 28:221–36.
191 Bumgardner GL, Li J, Apte S, Heininger M, Frankel WL. Effectof tumor necrosis factor α and intercellular adhesion molecule-1expression on immunogenicity of murine liver cells in mice.Hepatology 1998; 28: 466–74.
192 Bertolino P, Bowen DG, McCaughan W. Fazekas de St Groth B.Antigen-specific primary activation of CD8+ T cells within theliver. J. Immunol. 2001; 166: 5430–8.
193 Chen T, Goldstein JS, O’Boyle K, Whitman MC, Brunswick M,Kozlowski S. ICAM-1 co-stimulation has differential effects onthe activation of CD4+ and CD8+ T cells. Eur. J. Immunol.1999; 29: 809–14.
194 Cardoso EMP, Hagen K, de Sousa M, Hultcrantz R. Hepaticdamage in C282Y homozygotes relates to low numbers of CD8+cells in the liver lobuli. Eur. J. Clin. Invest. 2001; 31: 45–53.
195 De Sousa M, Porto G. The immunological system in hemochro-matosis. J. Hepatol. 1998; 28: 1–7.
196 Cardoso C, Porto G, Arosa FA et al. Relative impact of HFEmutations and lymphocyte numbers on the expression of ironoverload in non-C282Y homozygous subjects. In: Proceedingsof the World Congress on Iron Metabolism, 1999.
197 Eglinton BA, Roberton DM, Cummins AG. Phenotype of Tcells, their soluble receptor levels, and cytokine profile of humanbreast milk. Immunol. Cell. Biol. 1994; 72: 306–13.
198 Arai K, Yamamura S, Seki S et al. Increase of CD57+ T cells inknee joints and adjacent bone marrow of rheumatoid arthritis(RA) patients: implications for an anti-inflammatory role. Clin.Exp. Immunol. 1998; 111: 345–52.
199 Minagawa M, Oya H, Yamamoto S et al. Intensive expansion ofnatural killer T cells in the early phase of hepatocyte regenera-tion after partial hepatectomy in mice and its association withsympathetic nerve activation. Hepatology 2000; 31: 907–15.
200 Jarry A, Cerf-Bensussan N, Brousse N, Selz F, Guy-Grand D.Subsets of CD3+ (T cell receptor alpha/beta or gamma/delta) andCD3– lymphocytes isolated from normal human gut epitheliumdisplay phenotypical features different from their counterparts inperipheral blood. Eur. J. Immunol. 1990; 20: 1097–103.
201 Cerf-Bensussan N, Guy-Grand D. Intestinal intraepithelial lym-phocytes. Gastroenterol. Clin. North. Am. 1991; 20: 549–76.
202 Lundquist C, Baranov V, Hammarstrom S, Athlin L, Ham-marstrom ML. Evidence for regional specialization and extra-thymic T cell maturation in the human gut epithelium. Int.Immunol. 1995; 7: 1473–8.
203 Norris S, Collins C, Doherty DG et al. Resident human hepaticlymphocytes are phenotypically different from circulating lym-phocytes. J. Hepatol. 1998; 28: 84–90.
204 Doherty DG, Norris S, Madrigal-Estebas L et al. The humanliver contains multiple populations of NK cells, T cells, andCD3+CD56+ natural T cells with distinct cytotoxic activities andTh1, Th2, and Th0 cytokine secretion patterns. J. Immunol.1999; 163: 2314–21.
205 Ishihara S, Nieda M, Kitayama J et al. CD8+NKR-P1A+ T cellspreferentially accumulate in human liver. Eur. J. Immunol. 1999;29: 2406–13.
206 Moretta A, Bottino C, Vitale M et al. Activating receptors andcoreceptors involved in human natural killer cell-mediated cyto-lysis. Ann. Rev. Immunol. 2001; 19: 197–223.
207 Mingari MC, Schiavetti F, Ponte M et al. Human CD8+ Tlymphocyte subsets that express HLA class I-specific inhibi-tory receptors represent oligoclonally or monoclonallyexpanded cell populations. Proc. Natl Acad. Sci. USA 1996;93: 12433–8.
208 Lanier LL, Chang C, Phillips JH. Human NKR-P1A. Adisulfide-linked homodimer of the C-type lectin superfamilyexpressed by a subset of NK and T lymphocytes. J. Immunol.1994; 153: 2417–28.
209 Anumanthan A, Bensussan A, Boumsell L et al. Cloning ofBY55, a novel Ig superfamily member expressed on NK cells,CTL, and intestinal intraepithelial lymphocytes. J. Immunol.1998; 161: 2780–90.
210 Maiza H, Leca G, Mansur IG, Schiavon V, Boumsell L, Bensus-san A. A novel 80-kD cell surface structure identifies human cir-culating lymphocytes with natural killer activity. J. Exp. Med.1993; 178: 1121–6.
211 Bensussan A, Gluckman E, el Marsafy S et al. BY55 mono-clonal antibody delineates within human cord blood and bonemarrow lymphocytes distinct cell subsets mediating cytotoxicactivity. Proc. Natl Acad. Sci. USA 1994; 91: 9136–40.
212 Bensussan A, Rabian C, Schiavon V, Bengoufa D, Leca G,Boumsell L. Significant enlargement of a specific subset ofCD3+CD8+ peripheral blood leukocytes mediating cytotoxic T-lymphocyte activity during human immunodeficiency virusinfection. Proc. Natl Acad. Sci. USA 1993; 90: 9427–30.
213 Russell GJ, Parker CM, Sood A et al. p126 (CDw101), a costim-ulatory molecule preferentially expressed on mucosal T lym-phocytes. J. Immunol. 1996; 157: 3366–3374.
214 Chott A, Gerdes D, Spooner A et al. Intraepithelial lymphocytesin normal human intestine do not express proteins associatedwith cytolytic function. Am. J. Pathol. 1997; 151: 435–42.
215 Ebert EC, Roberts AI. Lymphokine-activated killing by humanintestinal lymphocytes. Cell. Immunol. 1993; 146: 107–16.
216 Slifka MK, Pagarigan RR, Whitton L. NK markers areexpressed on a high percentage of virus-specific CD8+ and CD4+
T cells. J. Immunol. 2000; 164: 2009–15.217 Assarsson E, Kambayashi T, Sandberg JK et al. CD8+ T cells
rapidly acquire NK1.1 and NK cell-associated molecules uponstimulation in vitro and in vivo. J. Immunol. 2000; 165: 3673–9.
218 Namekawa T, Snyder MR, Yen JH et al. Killer cell activatingreceptors function as costimulatory molecules on CD4+CD28null
T cells clonally expanded in rheumatoid arthritis. J. Immunol.2000; 165: 1138–45.
219 Warrington KJ, Takemura S, Goronzy JJ, Weyand CM.CD4+CD28– T cells in rheumatoid arthritis patients combine

Human CD8+CD28– T cells 13
features of the innate and adaptive immune systems. ArthritisRheum. 2001; 44: 13–20.
220 Vely F, Peyrat MA, Couedel C et al. Regulation of inhibitoryand activating killer-cell Ig-like receptor expression occurs inT cells after termination of TCR rearrangements. J. Immunol.2001; 166: 2487–94.
221 Azzoni L, Zatsepina O, Abele B et al. Differential transcrip-tional regulation of CD161 and a novel gene, 197/15a, by IL-2,IL-15, and IL-12 in NK and T cells. J. Immunol. 2000; 161:3493–500.
222 Freitas AA, Rocha B. Population biology of lymphocytes. Theflight for survival. Annu. Rev. Immunol. 2000; 18: 83–111.
223 De la Salle H, Donato L, Zimmer J et al. HLA class I deficien-cies. In: Ochs HD, Smith CEI, Puck JM (eds). PrimaryImmunodeficiency Diseases: a Molecular and GeneticApproach. New York: Oxford University Press, 1998; 181–8.
224 Sugamura K, Asao H, Kondo M et al. The interleukin-2 receptorγ chain: its role in the multiple cytokine receptor complexes andT cell development. Annu. Rev. Immunol. 1996; 14: 179–205.
225 Croituru K, Ernst PB. Intraepithelial lymphocyte lineage andfunction. The interactions between the intestinal epithelium andthe intraepithelial lymphocyte. In: Kiyono H, McGhee JR (eds).Mucosal Immunology: Intraepithelial Lymphocytes. New York:Raven Press, Ltd., 1993; 79–88.
226 Madara JL, Stafford J. Interferon–γ directly affects barrier func-tion of cultured intestinal epithelial monolayers. J. Clin. Invest.1989; 83: 724–7.
227 Dignass AU, Podolsky DK. Cytokine modulation of intestinalepithelial cell restitution: central role of transforming growthfactor β. Gastroenterology 1993; 105: 1323–32.
228 Boismenu R, Havran WL. Modulation of epithelial cell growthby intraepithelial γδ T cells. Science 1994; 266: 1253–5.
229 Purdue MH, McKay DM. Integrative immunophysiology in theintestinal mucosa. Am. J. Physiol. 1994; 267: G151–G165.
230 McKay DM, Croituru K, Perdue MH. T cell–monocyte inter-actions regulate epithelial physiology in a coculture model ofinflammation. Am. J. Physiol. 1996; 270: C418–C428.
231 Takahashi-Iwanaga H, Iwanaga T, Sakamoto Y, Fujita T.Ultrastructural and time-lapse observations of intraepitheliallymphocytes in the small intestine of the guinea pig: their
possible role in the removal of effete enterocytes. Cell TissueRes. 1995; 280: 491–7.
232 Wang J, Whetsell M, Klein JR. Local hormone networks andintestinal T cell homeostasis. Science 1997; 275: 1937–9.
233 Vinci G, Vernant JP, Nakaza WA et al. In vitro inhibition ofnormal human hematopoiesis by marrow CD3+, CD8+, HLA-DR+, HNK-1+ lymphocytes. Blood 1988; 72: 1616–21.
234 Stadnyk AW. Cytokine production by epithelial cells. FASEB J.1994; 8: 1041–7.
235 Colgan SP, Hershberg RM, Furuta GT, Blumberg RS. Ligationof the intestinal epithelial CD1d induces bioactive IL-10: criticalrole of the cytoplasmic tail in autocrine signaling. Proc. NatlAcad. Sci. USA 1999; 96: 13938–43.
236 Rowbottom AW, Lepper MW, Garland RJ et al. Interleukin10-induced CD8 cell proliferation. Immunology 1999; 98: 80–9.
237 Nathan C, Sporn M. Cytokines in context. J. Cell Biol. 1991;113: 981–6.
238 Levite M. Nervous immunity. neurotransmitters, extracellularK+ and T cell function. Trends Immunol. 2001; 22: 2–5.
239 Karaszewski JW, Reder AT, Anlar B, Kim WC, Arnason BG.Increased lymphocyte beta-adrenergic receptor density in pro-gressive multiple sclerosis is specific for the CD8+,CD28– sup-pressor cell. Ann. Neurol. 1991; 30: 42–7.
240 Fonseca AM, Porto G, Uchida K, Arosa FA. Red blood cellsinhibit activation induced cell death and oxidative stress inhuman peripheral blood T lymphocytes. Blood 2001; 97:3152–60.
241 Bauer S, Groh V, Wu J et al. Activation of NK cells and T cellsby NKG2D, a receptor for stress-inducible MICA. Science 1999;285: 727–9.
242 Groh V, Rhinehart R, Randolph-Habecker J et al. Costimulationof CD8αβ T cells by NKG2D via engagement by MIC inducedon virus-infected cells. Nat. Immunol. 2001; 2: 255–60.
243 Ugolini S, Arpin C, Anfossi N et al. Involvement of inhibitoryNKRs in the survival of a subset of memory-phenotype CD8+
T cells. Nat. Immunol. 2001; 2: 430–5.244 Ugolini S, Vivier E. Multifaceted roles of MHC class I and
MHC class I-like molecules in T cell activation. Nat. Immunol.2001; 2: 198–200.