Embed Size (px)
Citation preview

Patricia Ruiz, Christoph Schwarzler and Ursula Günthert
Summary
During mouse early development cell adhesion molecules are indispensable for the embryo organisation. A family of molecules probably involved in development is the transmembrane glycoprotein CD44 family, which exists in multiple isoforms. These are generated by alternative splicing of the pre- mRNA, resulting in the enlargement of the extracellular part of the molecule. The standard form of CD44 is widely expressed in adult tissues and in embryos from day 9.5 post coitum onwards, while the numerous variant isoforms exhibit highly specialised patterns of expression that are already in the egg cylinder at day 6.5 of development. In lymphohemopoiesis, specific variant isoforms also emerge at decisive differentiation stages. Although specific ligands for the variant region still await isolation, the highly organised expression of CD44 variant isoforms suggests they have a pivotal role in cellular interactions during early development, hemopoiesis.
introduction Cell-cell and cell-extracellular matrix (ECM) interactions play a fundamental role in many biological processes including embryonic development, wound healing, the maintenance of tissue integrity and tumour progression. Cells adhere to the ECM and to each other via specific cell surface receptors, e.g. selectins, integrins, cadherins, members of the immunoglobulin superfamily and of the CD44 family (reviewed in refs 1-3). CD44 is a single gene which produces a large number of highly glycosylated transmembrane isoforms involved in lymphocyte recircu- lation, hemopoiesis, leukocyte activation and cancer metastasis. Several reports have been published recently describing the many isoforms of the CD44 receptor and its multiple interactions (reviewed in refs 4-7).
Previously, CD44 has been known as lymphocyte hom- ing receptor (Pgp-1 or Hermes antigen), hyaluronate receptor and extracellular matrix receptor III, but attention on the molecule increased when splice variants were detected and correlated with the invasive and metastatic capacity of tumour cells in animal as well as in human tumour progression (reviewed in refs 6 and 7). Specific CD44 splice variants were found to render metastatic behaviour to non-metastasising rat cells after cDNA trans-
pattern formation and Accepted
16 Septernberr 1994
fe&@. Antibodies specific for these isoforms were abie to inhibit outgrowth of secondary tumours in the lung when they were coinjected with the invasive tumour cells in the spontaneous metastasis assay(9).
It has been postulated for many years that tumour cell and leukocyte migrations involve similar mechanisms to those found in differentiation, pattern formation and devel- opment (reviewed in ref. 10). Adhesion molecules belong- ing to the cadherins (reviewed in refs 11 and 12) and to the i n t e g r i n ~ ( ' ~ ~ ' ~ ) have been identified that serve as cell adhesion receptors during early development as well as being crucial for tumour cell-host interactions. Since tumour cells invade normal healthy tissue, it is thought that they utilise similar adhesion mechanisms to those described for normal lymphocyte recirculation, organo- genesis and embryogenesis. Disseminating tumour cells and activated lymphocytes share many properties, includ- ing the detachment from a primary site, their migration through the blood stream or the lymphatics, their extrava- sation and capacity of organ invasion and colonisation. The development of the blastocyst into early embryonic structures following implantation involves analogous mechanisms, like detachment from primary cites, invasion of surrounding tissue, directed cell migrations and coloni-
sation of secondary sites. The purpose of this review arti- cle is to discuss the potential involvement of CD44 iso- forms in early mammalian development and differen- tiation.
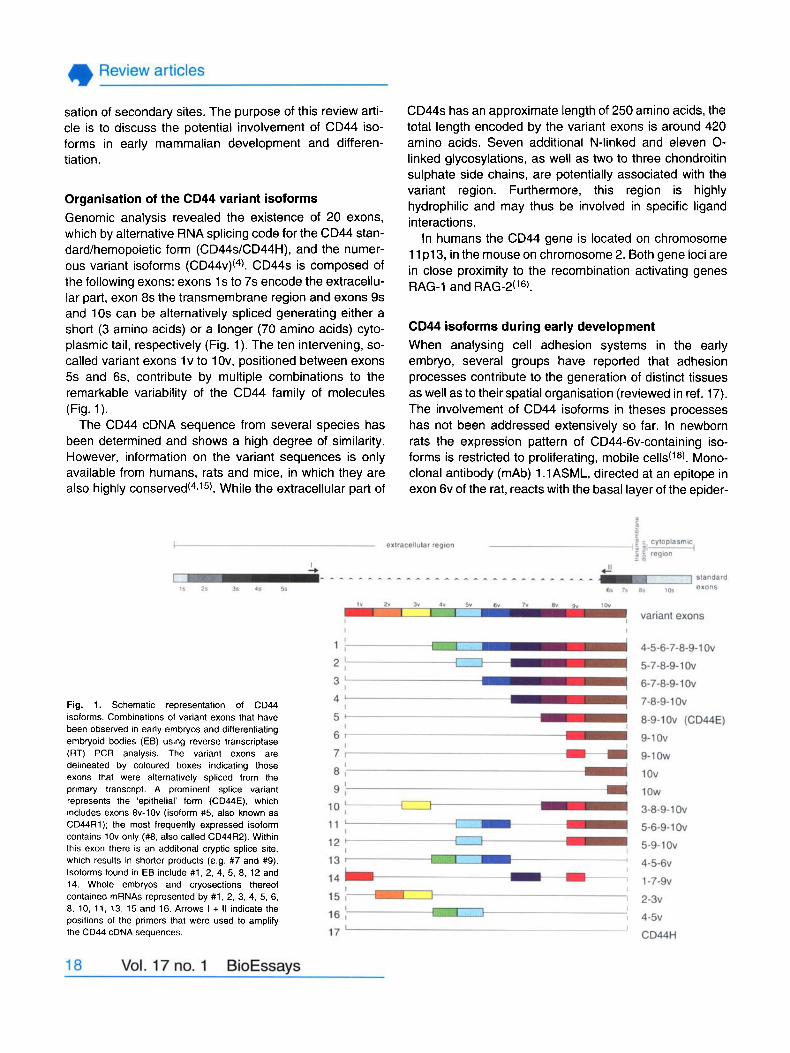
Organisation of the CD44 variant isoforms Genornic analysis revealed the existence of 20 exons, which by alternative RNA splicing code for the CD44 stan- dard/hemopoietic form (CD44s/CD44H), and the numer- ous variant isoforrns ( C D ~ ~ V ) ( ~ ) . CD44s is composed of the following exons: exons 1 s to 7s encode the extracellu- lar part, exon 8s the transmembrane region and exons 9s and 10s can be aiternatively spliced generating either a short (3 amino acids) or a longer (70 amino acids) cyto- plasmic tail, respectively (Fig. 1). The ten intervening, so- called variant exons l v to l Ov, positioned between exons 5s and 6s, contribute by muitiple combinations to the remarkabie variability of the CD44 family of molecules (Fig. 1).
The CD44 cDNA sequence from several species has been deterrnined and shows a high degree of similarity. However, information on the variant sequences is only available from humans, rats and mice, in which they are also highly ~onserved(~1~~). While the extracellular part of
Fig. 1. Schernatic reprecentation of CD44 isoforrns. Cornbinations of variant exons that have been observed in early ernbryos and differentiating embryoid bodies (EB) using reverse transcriptase (RT) PCR analysis. The variant exons are delineated by coloured boxes indicating those exons that were alternatively spliced from the prirnary transcript. A prominent splice variant represents the 'epithelial' form (CD44E). which includes exons 8v-1Ov (isoform #5, also known as CD44R1); the rnost frequently exprecsed isoform contains 1Ov only (#8, also called CD44R2). Within this exon there is an additional cryptic splice site, which results in shorter products (e.g. #7 and #9). Isoforrns found in EB include #1, 2, 4, 5, 8, 12 and 14. Whole embryos and cryosections thereof contained mRNAs represented by #1, 2, 3, 4, 5, 6, 8, 10, 11, 13, 15 and 16. Arrows I + II indicate the positions of the primers that were used to amplify the CD44 cDNA cequences.
CD44s has an approximate length of 250 amino acids, the total length encoded by the variant exons is around 420 amino acids. Seven additional N-linked and eleven 0- linked glycosylations, as well as two to three chondroitin suiphate side chains, are potentially associated with the variant region. Furthermore, this region is highly hydrophilic and may thus be involved in specific ligand interactions.
In humans the CD44 gene is located on chromosome 11 p13, in the mouse on chrornosome 2. Both gene ioci are in close proximity to the recombination activating genes RAG-1 and RAG-2(16).
CD44 isoforms during early development When analysing cell adhesion systems in the early embryo, several groups have reported that adhesion processes contribute to the generation of distinct tissues as weli as to their spatial organisation (reviewed in ref. 17). The involvement of CD44 isoforms in theses processes has not been addressed extensively so far. In newborn rats the expression pattern of CD44-6v-containing iso- forms is restricted to proliferating, mobile cells(' a). Mono- clonal antibody (mAb) 1 .lASML, directed at an epitope in exon 6v of the rat, reacts with the basa1 layer of the epider-

mis and the hair follicles as well as with the lower part of the crypts in the colon mucosa. While this expression pat- tern is kept constant throughout adult life, expression of CD44-6v in ductal epithelia of pancreatic glands is restricted to newborn animals(18). The use of CD44 vari- ant-specific mAbs to analyse expression in human fetal tissue (1 0th week) revealed predominant expression of CD44-9v-containing isoforms in epidermis, trachea, lung, thyroid gland and the rnesonephric and paramesonephric ducts; 6v-containing isoforms could only be detected in epidermis and trachea(Ig). Expression of CD44 in adult murine tissues has been studied by Kennel and col- leagued20). Immunohistochemistry revealed that major sites of expression are spleen, adrenal gland, thymus, liver, intestine, uterus, stomach and choroid of the eyes. Lung, salivary gland, oesophagus and trachea are also reactive with the CD44s-specific mAb, but these tissues express a substantial amount of CD44v isoforms, as shown by Western blot analyses, indicating 180- to 230- kD proteins in addition to the 90-kD standard size(20). The size of the antigen is not strictly indicative of the variant isoforms, because the chondroitin-sulphated form of CD44 has a similar size. Nevertheless, comparison with data obtained for CD44v in human tissue(lg) supports the described findings.
Monoclonal antibodies specific for the murine CD44 variants are not available yet, therefore analysis of CD44v expression in mouse development is restricted to RNA analysis by in situ hybridisation and reverse transcriptase polymerase chain reaction (RT PCR). CD44 expression during early mouse development has been examined by whole-mount immunohistochemistry with a mAb recognis- ing an epitope in the standard regiod2I). Since the stan- dard region is kept in the variant isoforms, mAbs directed at CD44s will recognise not only CD44s, but also al1 vari- ant isoformc (see Fig. 1). To be able to distinguish between the standard and the variant isoforms, the authors performed immunoblots showing larger molecular weight forms, probably indicative of CD44v isoforms(*I). Between days 9.5 and 12.5 of embryonic mouse develop- ment the predominant form is CD44s, but larger molecular weight forms are also detectable. The immunohistochemi- cal analysis revealed that at critica1 stages of organomor- phogenesis, CD44 is expressed at high levels in the heart, the somites and the condensing limb-bud mesenchyme. In these regions hyaluronic acid (HA), the major ligand for CD44s (see below), has been shown to regulate morpho- genetic events. However, it is of particular irnportance that CD44 expression does not always correlate with HA pres- ente, for example in the apical ectodermal ridge of the developing limb bud and the odontogenic placodes of the presurnptive upper and lower jaws, both being instructive
epithelia involved in epithelial-mesenchymal inter- actions(21). It is tempting to speculate that CD44 expressed in instructive epithelia represents variant isoformc. In situ hybridisations using exon 6v-specific sequences revealed indeed expression in the odontogenic placodes(22).
To analyse expression of CD44v in earlier stages of mouse development we perforrned in situ hybridisations with a probe covering exons 6-1Ov (isoform #3 in Fig. 1). Corresponding RNA was detected in the headfolds of a 7.5 days post coitum (P.C.) embryo, with strongest exprescion in the cell layer most proximal to the amniotic cavity (Fig. 2; Karen Downs and U.G., unpublished data). To unravel the composition of the variant isoforms expressed in the earliest stages of mouse development in more detail, RT PCR analyses of embryos 6.5 and 7.5 days p.c. were performed (Fig. 3). Total RNA was pre- pared from mouse embryos carefully dissected out of the maternal tissues and subjected to randomly primed cDNA synthesis. The cDNAs were equilibrated using the house- keeping gene HPRT (hypoxanthine phosphoribosyltrans- ferase). Using CD44 specific primers I and II (see Fig. l ) , PCR products were blotted and hybridised with CD44 exon-specific radiolabelled probes. This technique allows a more efficient and faster analysis of the exact composi- tion of distinct CD44 mRNA populations, as compared to other published techniques for CD44 isoform ana l~s i s (~~ ) .
Fig. 2. Expreccion of CD44v by in situ hybridication. (A) Longitudinal cection of an early headfold-ctage mouse embryo (7.5 dayc P.C.) and (6) trancverce section through the plane indicated by the dached line. The shaded red area indicatec the neural ectoderm (headfolds), where exprecsion of variant icoformc wac detected using a probe covering CD44 exons 6-1Ov (isoform #3 in Fig. 1). ac. amniotic cavity; al, allantois; am. amnion; ee. embryonic endoderm; hf. headfolds; iem, intra-embryonic (craniai) mesoderm; ng, neural groove; xc, exocoelomic cavity.
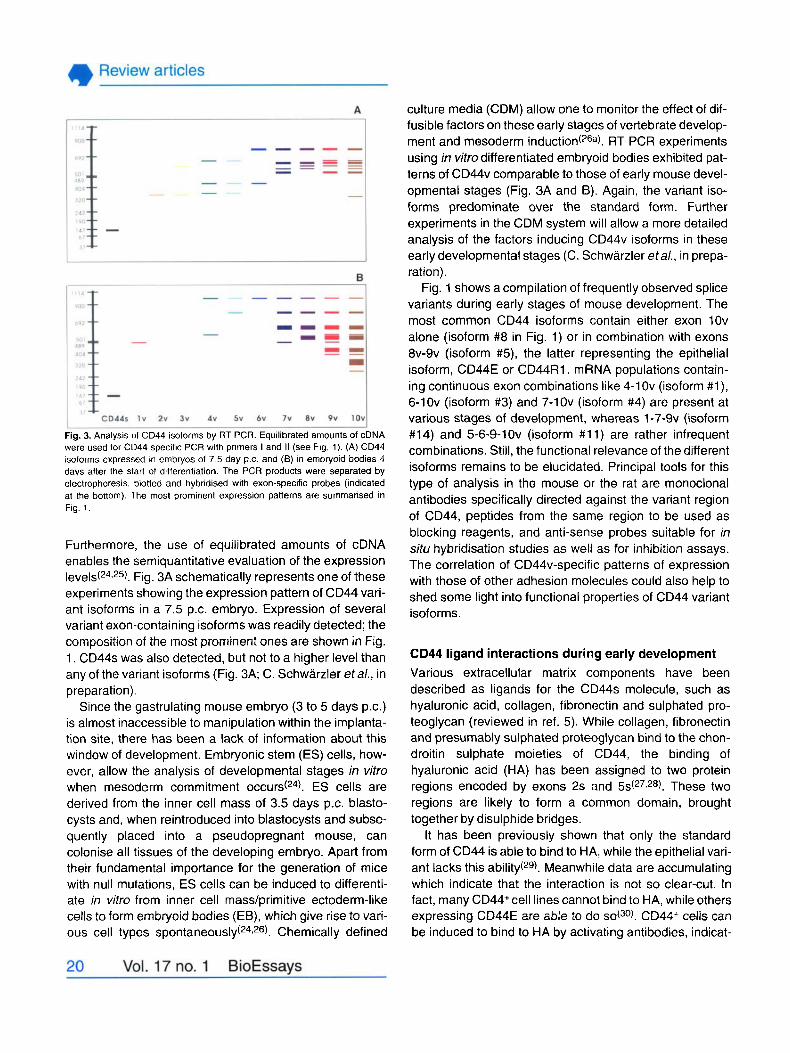
Fig. 3. Analysis of CD44 isoforms by RT PCR. Equilibrated amounts of cDNA were used for CD44 cpecific PCR with prirners I and II (see Fig. 1). (A) CD44 isoforms exprecsed in ernbiyos of 7.5 day p.c. and (6) in embryoid bodies 4 days after the start of differentiation. The PCR products were separated by electrophorecis. blotted and hybridised with exon-specific probes (indicated at the bottom). The most prorninent expression patterns are sumrnarised in Fig. 1.
Furthermore, the use of equilibrated amounts of cDNA enables the semiquantitative evaluation of the expression l e v e l ~ ( ~ ~ ~ ~ ~ ) . Fig. 3A schematically represents one of these experirnents showing the expression pattern of CD44 vari- ant isoforms in a 7.5 p.c. embryo. Expression of several variant exon-containing isoforms was readily detected; the composition of the most prominent ones are shown in Fig. 1. CD44s was also detected, but not to a higher leve1 than any of the variant isoforms (Fig. 3A; C. Schwarzler etal., in preparation).
Since the gastrulating mouse embryo (3 to 5 days P.C.) is almost inaccessible to manipulation within the implanta- tion site, there has been a lack of information about this window of development. Embryonic stem (ES) cells, how- ever, allow the analysis of developmental stages in vitro when mesoderm commitment o c c u r ~ ( ~ ~ ) . ES cells are derived from the inner cell mass of 3.5 days p.c. blasto- cysts and, when reintroduced into blastocysts and subse- quently placed into a pseudopregnant mouse, can colonise al1 tissues of the developing embryo. Apart from their fundamental importance for the generation of mice with null mutations, ES cells can be induced to differenti- ate in vitro from inner cell mass/primitive ectoderm-like cells to form ernbryoid bodies (EB), which give rise to vari- ous cell types s p o n t a n e o u ~ l y ( ~ ~ ~ ~ ~ ) . Chernically defined
culture media (CDM) allow one to monitor the effect of dif- fusible factors on these early stages of vertebrate develop- ment and mesoderm induction(26a). RT PCR experiments using in vitro differentiated embryoid bodies exhibited pat- terns of CD44v comparable to those of early mouse devel- opmental stages (Fig. 3A and B). Again, the variant iso- forms predominate over the standard form. Further experirnents in the CDM system will allow a more detailed analysis of the factors inducing CD44v isoforms in these early developmental stages (C. Schwarzler etal., in prepa- ration).
Fig. 1 shows a compilation of frequently observed splice variants during early stages of mouse development. The most common CD44 isoforms contain either exon 1Ov alone (isoform #8 in Fig. 1) or in combination with exons 8v-9v (isoform #5), the latter representing the epithelial isoform, CD44E or CD44R1. mRNA populations contain- ing continuous exon combinations like 4-1Ov (isoforrn #l ) , 6-1Ov (isoform #3) and 7-1Ov (isoform #4) are present at various stages of developrnent, whereas 1 -7-9v (isoform #14) and 5-6-9-1Ov (isoform #11) are rather infrequent combinations. Still, the functional relevance of the different isoforms remains to be elucidated. Principal tools for this type of analysis in the mouse or the rat are monoclonal antibodies specifically directed against the variant region of CD44, peptides from the same region to be used as blocking reagents, and anti-sense probes suitable for in situ hybridisation studies as well as for inhibition assays. The correlation of CD44v-specific patterns of expression with those of other adhesion molecules could also help to shed some light into functional properties of CD44 variant isof orrns .
CD44 ligand interactions during early development Various extracellular matrix components have been described as ligands for the CD44s molecule, such as hyaluronic acid, collagen, fibronectin and sulphated pro- teoglycan (reviewed in ref. 5). While collagen, fibronectin and presumably sulphated proteoglycan bind to the chon- droitin sulphate moieties of CD44, the binding of hyaluronic acid (HA) has been assigned to two protein regions encoded by exons 2s and 5s(27,28). These two regions are likely to form a common domain, brought together by disulphide bridges.
It has been previously shown that only the standard form of CD44 is able to bind to HA, while the epithelial vari- ant lacks this ability(29). Meanwhile data are accumulating which indicate that the interaction is not so clear-cut. In fact, many CD44+cell lines cannot bind to HA, while others exprecsing CD44E are able to do CD44+ cells can be induced to bind to HA by activating antibodies, indicat-

ing that conformational changes or redistribution of CD44 in the plasma membrane can trigger ligand bit1ding(3~ ).
Also stimulation of T cells by antigen or PMA (phorbol myristate acetate) transiently upregulates binding to HA(32333), and concomitantly the expression of variant iso- f orms(34-36).
Signal transduction from extracellular ligands via the cytoplasrnic domain to the cytoskeleton and vice versa not only regulate ligand interactions, but may also influence the cytoskeletal organisation and thus alter cellular motil- ity. Functional implications of the different lengths of the CD44 cytoplasmic tail generated by alternative splicing (either exon 9s or exon 10s) have recently been addressed. Although the longer tail is phosphorylated at two serine positions, these phosphorylations do not influ- ence membrane localisation and cytoskeletal interactions or binding to HA(31,37). Deletion of the cytoplasmic tail, however, leads to a cellular redistribution of the strictly lat- erally expressed rnolecules mainly to the apical surface and also to retention in the cytoplasrn; in addition the half life of the ‘tailless’ rnutant is drastically r e d ~ c e d ( ~ ~ ) .
The linking molecule between the cytoplasmic tail of CD44 and the actin-based cytoskeleton has been postu- lated to be ankyrin, which is specifically accumulated dur- ing HA-stimulated T cell ac t i~a t ion(~~) . Recently Tsukita and col laborat~rs(~~) have demonstrated that a 140 kD CD44 molecule preferentially interacts, via mernbers of the ERM (ezrin-radixin-moesin) family, with the cytoskel- eton. This larger isoforrn, but not CD44s, forms rnultimeric clusters at cleavage furrows and in filopodia, and is proba- bly colocalised with other surface mo lec~ les (~~) . Whether these clusters are the prerequisites for ligand binding and signal transduction, has not yet been dernonstrated. Inter- estingly, it has been shown before, that CD44 from fibrob- lasts (mostly CD44s) and from epithelial cells (rnostly CD44E) exhibit strictly different detergent extraction pro- files; whereas CD44E is essentially detergent soluble, a significant proportion of CD44s rernains in the detergent- insoluble fraction, thus indicating distinct cytoplasrnic part- ners(38).
The major CD44 ligand, the glycosarninoglycan hyaluronic acid, has been implicated in cell rnigrations associated with tumour metastasis and ernbryogenesis (reviewed in refs 41 and 42). HA is present in the ECM of rnost rnamrnalian tissues. lncreases in HA content have been observed in areas of cell proliferation and migration, ¡.e. in ernbryogenesis, tissue rernodelling and tumour pro- gression. The prornoting effect of HA on cell rnigrations is due to its ability to form an expanded, highly hydrated bed which facilitates cell-ECM contacts. Additionally, cytokines and chemokines rnay be bound by the HA matrix
and thus presented in locally different concentrations (reviewed by ref. 43).
Employing a soluble CD44 imrnunoglobulin fusion pro- tein for screening of the predorninant ligand HA in the developing mouse embryo, Fenderson and c ~ l l e a g u e s ( ~ ~ ) observed drastic changes in the distribution of HA. HA was first observed in embryos of 5.5 to 6.5 p.c. (pre-primi- tive streak stage) in the basernent rnernbranes of the primitive ecto- and endoderm and in the yolk sac cavity. At the neural plate and pre-somite stage (day 7.5 P.C.), HA was present in the basernent membrane of embryonic ectoderm and visceral endoderm, as well as in the amnion and the extraembryonic visceral yolk sac; but was absent in the region of the presumptive neural plate and parietal e n d ~ d e r m ( ~ ~ ) . At day 9.5 of development onwards, HA was detected throughout the cranial mes- enchyrn, but not in the branchial bars, sornites or lirnb buds and in the neural plate. At later stages of develop- rnent the pattern of HA expression changed significantly: while at earlier stages HA was detected primarily in cavi- ties, from day 9.5 p.c. on, HA was rnostly found in the intercellular spaces between mesenchymal cells of neural crest and mesodermal cell ~ r i g i n ( ~ ~ ) . At rnid-gesta- tion HA is expressed in the axial skeleton, craniofacial mesenchyme, endocardial cushions of the heart, srnooth muscle of the gastrointestinal tract and connective tissue throughout the b ~ d y ( ~ ~ ) . Similar data have been obtained using a biotin-labelled bovine nasal cartilage proteogly- can, which binds with high affinity to HA(45). Dissecting primitive streak embryos (7.5 days P.C.) and cultivating the cells for a short time, HA was secreted frorn meso- derrn, embryonic ectoderrn and endoderm, but was absent in fragments from the ectoplacental cone, parietal yolk sac and extraernbryonic ectoderm and e n d ~ d e r m ( ~ ~ ) . Since the amount of HA synthesis was rnarkedly increased in cultures containing serum, it is likely that additional growth factors present in the serurn trigger HA synthesis in vivo, t00 (~~) .
The distribution of CD44 and HA in later developrnental stages (from day 9.5 p.c. onwards) matches, largely. How- ever, in situ hybridisations using variant-specific sequences revealed expression of CD44v in the headfolds of a 7.5 day p.c. embryo, a region devoid of HA at this time of development (Fig. 2 and ref. 44). RT PCR analyses of the CD44 exon cornposition revealed that several variant isoforrns are present at that stage and that CD44s is more abundant than CD44v isoforms (Fig. 3A). It is ternpting to speculate that at the earliest developmental stages before organogenesis commences, CD44 does not interact with HA, but HA receptor interactions (if existing) are rnediated via other receptors like RHAMM (receptor for HA-medi-

ated mobility) and LEC (liver endothelial ceii) receptor (reviewed in ref. 42).
Cell-cell and cell-ECM interplays are not static events but are constantly changing. Alterations are mediated by growth factors and/or chemokines which play pivotal roles in signalling and instruction (reviewed in refs 46 and 47). Some factors have been described to induce epithelial morphogenesis, e.g. scatter factor/hepatocyte growth fac- tor, activin and basic fibroblast growth factor (reviewed in refs 47 and 48). Cell adhesion molecules like a& integrin and E-cadherin are involved in mesoderm differentiation and migration at the mammalian primitive streak, and dur- ing conversion of mesenchyme to epi thel i~m(~~). How- ever, rather than being involved in direct induction processes, the expression and function (e.g. activation state) of these molecules seem to be modulated in a fine- tuned series of events leading to embryonic induction and tissue differentiation. Expression of CD44 variant isoforms can be modulated by cytokines: tumour necrosis factor a as well interferon y upregulate CD44-9v- and CD44-6v- containing isoforms on myelomonocytic cells and CD44- 6v on come epithelial cell lines(36~~~). Whether these or other factors are involved in CD44v induction processes in the developing embryo has to await further analysis.
CD44 isoformc on differentiating hemopoietic cells As in organogenecis and tumour progression, lympho- hemopoiesis is also strictly dependent on cell adhesion molecules, including CD44. Developing blood cells have to encounter stromal cells in the bone marrow where they receive stimulatory signals, for example cytokines and growth factors. The earliest hernatopoietic precursors in the embryo are detected in the yolk sac and the liver (reviewed in ref. 51). In newborn animals, the bone mar- row takes over as the principal site of blood cell develop- ment. Concomitantly, the irnmature thymus is seeded with T-cell precursors in several distinct waves (reviewed in ref. 51).
Aimed at identifying earliest developmental stages for the analysis of function, pluripotent and multipotent hemo- poietic stem cells from bone marrow and fetal liver have been described (reviewed in ref. 51). In the thymus, a pop- ulation of progenitor cells which express CD44 has been described. The earliest thymocyte population identified so far is defined by the phenotype CD4+ CD8- CD3- Thyl+ HSA+ CD44s+ IL2Ra-, cornprising 0.05% of al1 thymo- cyted5*). The differentiated progeny of these cells are the double negative (CD4-CD8-) thymocytes, which can be subdived by their surface expression of CD44 and IL-2 receptor (IL-2R) rnolecules. Before acquiring the double positive phenotype (CD4+ CD8+), double negative thymo-
cytes undergo the transition from CD44+ IL-2R- to CD44- IL-2R+ to CD44- IL-2R-(53). So far, no evidence has been presented concerning the presence of CD44 variants on T-cell precursors. In human ten- to eleven-weeks-old fetal thymus, CD44s was expressed by cortical as well as by medullary cells and CD44-9v containing isoforrns were only detected on medullary cells. In general, no further CD44v isoforms were expressed by lymphoid cells within the embryonic thymus, except for cellc in the Hassall’s cor- pusc~es(’g).
A role in thymus colonisation by pro T-cells has been ascribed to CD44. It is expressed on bone rnarrow cells able to repopulate the thymus of irradiated mice and in long-term bone marrow reconstitution. Anti-CD44 anti- bodies inhibit thymus repopulation by bone marrow prog- enitors and homing of fluorescently labelled bone marrow cells to the t h y m u ~ ( ~ ~ 7 ~ ~ ) . Further evidence for the role as a homing receptor of CD44 on intrathymic precursor cells was given by Wu and collaborat~rs(~~). The dynamic inter- action between the adhesion receptors expressed by leukocytes and endothelium is viewed as a cascade of receptor/ligand interactions, in which each event is a pre- requisite for the next. This carefully orchestrated series of events operates most stringently when subsets of memory T-cells are guided to their ‘home’ in the tissues of the body, and less stringently when neutrophils, monocytes and lymphocytes have to combat an inflammatory reaction (reviewed in ref. 3).
Memory T-cells can be distinguished from naive T cells (CD45RO+ versus CD45RA+) by their CD45 surface expression. CD44 also displays a differential expression on these cells: memory T cells have much higher levels of CD44 than naive T cells and do express CD44-9v contain- ing isoforms (see ref. 36, reviewed in ref. 57). In addition, certain splice variants of CD44, including CD44E and exon 6v-containing isoforms, are upregulated following anti- genic ~t imulat ion(~~) or allogenic response (refs 35,36; R. Stauder etal., submitted).
The genesis, proliferation and differentiation of B-cells is likewise a complex process directed through interac- tions with many other cell t y p e ~ ( ~ ~ ) . CD44s defines two discrete subpopulations among bone marrow B-cells that differ 10- to 15-fold in the levels of CD44 expres~ion(~~). The earliest pre-B cells (CDI9+CD10+) have a CD44-low phenotype with the transition to the CD44-high phenotype occurring relatively late in development, when cells become immature IgM+ B cells. CD44 is expressed at medium densities on normal blood B cells and increases to high density with B-cell activation (reviewed in ref. 52). Although most resting IgD+ B cells are positive for CD44, a significant fraction of B cells in tonsils are CD44c-low to -negative; these cells are predominantly sessile B cells in

germina1 centres, which also do not express any variant isoforms(55~60).
On B cells, CD44-variant isoforms are transiently expressed upon activation and this process is inhibitable by incubation with rnonoclonal antibodies specific for exon 6v(34). Furtherrnore, CD44s-specific mAbs were found to completely block B-cell lymphopoiesis in long- term bone marrow cultures, but did not have such an effect when added later, indicating that CD44 is a crucial effector in a prirnary cell adhesion event and may interact with ligands, for example HA, on the sutface of stromal cells(61). Recently it has been shown that CD34+ myeloid precursor cells spontaneously bind to HA via CD44, sug- gesting that CD44 is involved in the interaction of these precursor cells with the bone marrow stroma(62). When a panel of non-Hodgkin’s lymphomas, possibly represent- ing a complete spectrum of B-cell development, was analysed for the presence of CD44 isoforms, the CD44- 6v isoform in combination with other variant exons was observed predominantly in aggressive lymphomas and was associated with a shorter overall survival of patients (R. Stauder et al., submitted). This study re-emphasises the major role of CD44 isoforms during turnour progres- sion as well as during developmentally regulated processes. In conclusion, it is reasonable to hypothesise that specific CD44 variant isoforms transiently establish specific ligand interactions during development and pat- tern formation. These interactions may guide the expressing cells to their new destinations in a kind of area code system, as has been described for lymphocyte recir- cuiation(3).
Concluding remarks So far, broad information has been collected about the structural details of the CD44 molecule. Currently, the functional relevance of the CD44 receptor molecule, espe- cially al1 its isoforms, is still unknown. The generation of antibodies directed to, and peptides frorn, the mouse and rat CD44 sequences will doubtlessly facilitate the elucida- tion of the complex distribution pattern of the isoforms and will hopefully provide clues for a functional interpretation in development and ernbryogenesis. The latter rnight like- wise be obtained by loss-of-function experiments, either with competing antibodies or peptides, or by knocking-out specific exons by targeted hornologous recombination. Assuming a crucial role for CD44 isoforms during mouse development (e.9. ectoderm/mesoderm transitions, epithelial rnigrations and T/B cell differentiation), we expect a possible arrest at an early developmental stage or a detectably altered phenotype by interfering with the normal interactions of a particular variant exon.
Acknowiedgment We are grateful to Dr Michael V. Wiles for help with ES cell culture and embryoid body differentiation, as well as for critically reviewing the manuscript.
Due to a limit set for the references cited, we have indi- cated several review articles, in which the original publica- tions will be found.
The Basel Institute for lmmunology was founded and is supported by F. Hoffmann-La Roche, Ltd., Basel, Switzer- land.
References 1 Adams, J. C. and Watt, F. M. (1993). Regulation of development and differentiation by the extracellular matrix. Developmentl 17, 11 83-1 198. 2 Lin, C. Q. and Bissell, M. J. (1993). Multi-faceted regulation of cell differentiation by extracellular matrix. FASE6 J. 7, 737-743. 3 Springer, T. A. (1994). Traffic signals for lymphocyte recirculation and leukocyte emigration: the multistep paradigm. Ce//76,301-314. 4 Günthert, U. (1993). CD44: a multitude of isoforms with diverse functions. Curr. Top. Microbiol. immunol. 184,47-63. 5 Lesley, J., Hyman, R. and Kincade, P. W. (1993). CD44 and its interactions with the extracellular matrix. Adv. lmmunol. 54,271 -335. 6 Pals, S. T., Koopman, G., Griffioen, A., Ponta, H., Herrlich, P., van den Berg, F. and Horst, E. (1993). CD44: a variety of isoforms with functions in cell adhesion and tumor metastasis. In Lymphocyte Adhesion Molecules (ed. Y. Shimizu), pp. 135-150. R.G. LandesComp., Austin. 7 Günthert, U., Stauder, R., Mayer, B., Terpe, H.-J., Finke, L. and Friedrichs, K. (1994). Are CD44 variant isoforms involved in human tumour progression? Cancer Surveys (in press). 8 Günthert, U., Hofmann, M., Rudy, W., Reber, S., Zoller, M., Haussmann, I., Matzku, S., Wenzel, A., Ponta, H. and Herrlich, P. (1991). A new variant of glycoprotein CD44 confers metastatic potential to rat carcinoma cells. Cell65. 13-24. 9 Ceiter, S., Arch, R., Reber, S., Komitowski, D., Hofmann, M., Ponta, H., Herrlich, P., Matzku, S. and Zoller, M. (1993). Prevention of tumor metastasis formation by anti-variant CD44. J. Exp. Med. 177,443-455. 10 Mareei, M. M., Van Roy, F. M. and De Baetselier, P. (1990). The invasive phenotypes. Cancer Metast. Rev. 9.45-62. 11 Birchmeier, W. and Behrens, J. (1994). Cadherin expression in carcinomas: role in the formation of cell junctions and the prevention of invasiveness. Biochem. Biophys. Acta 11 98, 1 1-26. 12 Mareel, M., Bracke, M. and Van Roy, F. (1994). Invasion promoter versus invasion suppressor molecules: the paradigm of E-cadherin. Mol. Bol. Rep. 19,
13 Qian, F., Vaux, D. L. and Weissman, 1. L. (1994). Expression of the integrin a4B1 on melanoma cells can inhibit the invasive stage of metastasis formation. Cel/77,335-347. 14 Ruiz, P., Dunon, D., Sonnenberg, A. and Imhof, B. A. (1993). Suppression of mouse melanoma metastasis by EA-1, a rnonoclonal antibody specific for a6 integrins. Ceil Adh. Comm. 1,6741. 15 Screaton, G. R., Bell, M. V., Bell, J. 1. and Jackson, D. G. (1993). The identification of a new alternative exon with highly restricted tissue expression in transcripts encoding the mouse Pgp-1 (CD44) homing receptor. Cornparison of al1 10 variable exons between mouse, hurnan, and rat. J. Bid. Chern. 268,
16 Sherrington, P. D., Forster, A., Seawright, A., van Heyningen, V. and Rabbitts, T. H. (1 992). Human RAG2, like RAGI, is on chromosome 11 band p l3 and therefore not linked to ataxia telangiectasia complementation groups. Genes, Chrom. Cancer5,404-406. 17 Fleming, T. P. etal. (1994). Molecular maturation of cell adhesion systems during mouse early development. HistochemistfylOl, 1-7. 18 Wirth, K. etal. (1993). Expression of CD44 isoforms carfying metastasis- associated sequences in newborn and adult rats. Eur. J. Cancer 29A, 1 172- 1177.
45-67.
12235-12238.

19 Terpe, H.-J., Stark, H., Prehm, P. and Günthert, U. (1994). CD44 variant isoforms are expressed preferentially in basa1 epithelia of non-malignant human fetal and adult tiscues. Histochemistry101,79-89. 20 Kennel, S. J., Lankford, T. K., Foote, L. J., Shinpock, S. G. S. and Stringer, C. (1993). CD44 expreccion on murine ticsues. J. Cell Sci. 104, 373- 382. 21 Wheatley, S. C., Isacke, C. M. and Crossley, P. H. (1993). Restricted expreccion of the hyaluronan receptor, CD44. during postimplantation mouse ernbryogenecis cuggestc key roles in tissue formation and patterning. Development119.295-306. 22 Herrlich, P. et al. (1993). CD44 and splice variants of CD44 in normal differentiation and tumor progression. In Cell Adhesion Molecules (ed. M. E. Hemler and E. Mihich). pp. 265-288. Plenurn Press, New York. 23 van-Weering, D. H., Baas, P. D. and Bos, J. L. (1993). A PCR-based method for the analycic of human CD44 splice products. PCRMethods Appl. 3,
24 Keller, G., Kennedy, M., Papayannopoulou, T. and Wiles, M. (1993). Hematopoietic cornmitrnent during embryonic stem cell differentiation in culture. Mol. Cell Biol. 13.473-486. 25 Ruiz, P., Haasner, D. and Wiles, M. V. (1994). Use of polymerase chain reactions (PCR) on limited arnountc of material. in Manual of lmmunological Mefbods (ed. l. Levkovits) (in press). Academic Press, San Diego. 26 Wiles, M. V. (1993). Embryonic stem cell differentiation in vitro. In Methods in Enzymology (ed. P. M. Wassarmann and M. L. DePamphilis), pp. 900-918. Academic Press, San Diego. 26a Johansson, B. M. and Wiles, M. V. (1995). Evidence for the involvernent of activin A and BMP-4 in rnammalian rnesoderm and hematopoietic development. Mol. Cell Biol. In press. 27 Peach, R. J., Hollenbaugh, D., Stamenkovic, 1. and Aruffo, A. (1993). ldentification of hyaluronic acid binding sites in the extracellular domain of CD44. J. Cell Biol. 122,257-264. 28 Yang, B., Yang, B. L., Savani, R. C. and Turley, E. A. (1994). ldentification of a common hyaluronan binding motif in the hyaluronan binding proteins RHAMM, CD44 and link protein. EMBO J. 13,286-96. 29 Starnenkovic, 1.. Aruffo, A., Amiot, M. and Seed, B. (1991). The hematopoietic and epithelial forms of CD44 are dictinct polypeptides with different adhesion potentials for hyaluronate-bearing cellc. EMBO J. 10, 343-
30 He, Q., Lesley, J., Hyman, R., Ishihara, K. and Kincade, P. W. (1992). Molecular isoforms of murine CD44 and evidence that the rnembrane proximal domain is not critica1 for hyaluronate recognition. J. CellBiol. 119, 171 1-1719. 31 Lesley, J., He, Q., Miyake, K., Hamann, A,, Hyman, R. and Kincade, P. W. (1992). Requirements for hyaluronic acid binding by CD44: A role for the cytoplasrnic domains and activation by antibody. J. Exp. Med. 175, 257-266. 32 Galandrini, R., Galluuo, E., Albi, N., Grossi, C. E. and Velardi, A. (1 994). Hyaluronate is costimulatory for human T cell effector functions and binds to CD44 on activated T ceils. J. lmmunol. 153,21-31. 33 Lesley, J., Howes, N., Perschl, A. and Hyman, R. (1994). Hyaiuronan binding function of CD44 is transiently activated on T cells during an in vivo irnmune response. J. Exp. Med. 180,383-387. 34 Arch, R., Wirth, K., Hofmann, M., Ponta, H., Matzku, S., Herrlich, P. and Zoller, M. (1992). Participation in normal immune responses of a metactasic- inducing splice variant of CD44. Science257.682-685. 35 Koopman, G. et al. (1993). Activated human lymphocytes and aggressive non-Hodgkin's lymphornac exprecs a homoiogue of the rat metastasis- accociated variant of CD44. J. Exp. Med. 177,897-904. 36 Mackay, C. R. et al. (1994). Expression and modulation of CD44 variant isoforms in humans. J. CellBiol. 124,71-82. 37 Neame, S. J. and Isacke, C. M. (1992). Phocphorylation of CD44 in vivo requires both Ser 323 and Ser 325, but doec not regulate membrane localization or cytoskeletal interaction in epithelial cells. EMBO J. 11, 4733- 4738. 38 Neame, S. J. and Isacke, C. M. (1993). The cytoplasmic tail of CD44 is required for bacolateral localization in epithelial MDCK cells but doec not mediate association with the detergent-insoluble cytockeleton of fibroblasts. J. CeliBiol. 121. 1299-1310. 39 Bourguignon, L. Y. W., Lokeshwar, V. B., Chen, X. and Kerrick, W. G. L. (1 993). Hyaluronic acid-induced lymphocyte signal transduction and HA receptor (GP85/CD44)-cytockeleton interaction. J. lmmunol. 151,6634-6644.
100-1 06.
348.
40 Tsukita, S., Oishi, K., Sato, N., Sagara, J., Kamai, A. and Tsukita, S. (1994). ERM family members as molecular linkers between the cell curface glycoprotein CD44 and actin-based cytoskeletonc. J. Cell Biol. 126,391 -401. 41 Laurent, T. C. and Fraser, J. R. (1992). Hyaluronan. FASEB J. 6, 2397- 2404. 42 Knudson, C. B. and Knudson, W. (1993). Hyaluronan-binding proteins in development, tissue homeostasis, and disease. FASEB J. 7, 1233-1241. 43 Tanaka, Y., Adams, D. H. and Shaw, S. (1993). Proteoglycans on endothelial cells present adhecion-inducing cytokines to leukocytec. lmmunol. Today l4 , l l 1-1 15. 44 Fenderson, B. A., Stamenkovic, l. and Aruffo, A. (1993). Localization of hyaluronan in mouse embryos during implantation, gastruiation and organogenesis. Differentiation 54,8598. 45 Brown, J. J. G. and Papaioannou, V. E. (1993). Ontogeny of hyaluronan secretion during eariy mouse development. Developmenf 117,483-492. 46 Juliano, R. L. and Haskill, S. (1993). Signal transduction from the extracellular matrix. J. CellBiol. 120, 577-585. 47 Smith, J. C. (1993). Mesoderm-inducing factors in early vertebrate developrnent. EMBO J. 12,4463-4470. 48 Gherardi, E., Sharpe, M., Lane, K., Sirulnik, A. and Stoker, M. (1993). Hepatocyte growth factorkcatter factor (HGFISF), the c-met receptor and the behaviour of epithelial cells. Symp. Soc. Exp. Biol. 47, 163-1 81. 49 Burdsal, C. A., Damsky, C. H. and Pedersen, R. A. (1 993). The role of E- cadherin and integrins in mesoderm differentiation and migration at the mammalian primitive streak. Development 118,829-844. 50 Haegel, H., Tolg, C., Hofmann, M. and Ceredig, R. (1993). Activated mouse astrocytes and T cells express similar CD44 variantc. Role of CD44 in actrocytefi cell binding. J. CellBiol. 122. 1067-1077. 51 Weissmann, 1. L. (1994). Developmental switches in the immune systern. Ce//76,207-218. 52 Kincade, P. W. (1993). Cell adhecion mechanicms utilized for lympho- hematopoiesis. In Lymphocyte Adhesion Molecules (ed. Y. Shimizu). pp. 249- 279. R.G. Landes Comp., Austin,Texas. 53 Petrie, H. T., Hugo, P., Scollay, R. and Shortman, K. (1990). Lineage relationships and developrnental kinetics of irnrnature thymocytes: CD3, CD4, and CD8 acquisition in vivo and in vitro. J. Exp. Med. 172, 1583-1 588. 54 O'Neill, H. C. (1 989). Antibody which defines a subset of bone marrow cellc that can migrate to the thymus. lmmunology68.59-65. 55 Horst, E. etai. (1990). Expression of a hurnan homing receptor (CD44) in lymphoid maiignancies and related stages of lymphoid development. Leukemia
56 Wu, L., Kincade, P. W. and Shortman, K. (1993). The CD44 expresced on the earliest intrathymic precursor population functions as a thymuc homing molecule but does not bind to hyaluronate. lmmunol. iett. 38,69-75. 57 Mackay, C. R. (1 993). lmmunological memory. Adv. lmmunol. 53,217-265. 58 Rolink, T. and Melchers, F. (1993). B lymphopoiesis in the rnouse. Adv. lmmunol. 53. 123-1 56. 59 Kansas, G. S. and Dailey, M. O. (1989). Expreccion of adhesion structures during B-cell development in man. J. lmmunol. 142,3058-3062. 60 Terpe, A., Koopmann, R., Imhof, B. and Günthert, U. (1994). Expression of integrins and CD44 in non-Hodgkin's lymphomas: CD44 variants are preferentially expressed in high-grade malignant lymphomas. J. Pathology (in press). 61 Miyake, K. et al. (1990). Monoclonal antibodiec to Pgp-l/CD44 block lympho-hemopoiesis in long-term bone marrow cultures. J. Exp. Med. 171,
62 Morimoto, K. et al. (1994). CD44 mediates hyaluronan binding by human myeloid KG1A and KG1 cells. Blood83. 657-62.
4, 383-389.
477-488.
Patricia Ruiz', Christoph Schwarzler and Ursula Günthert are at the Base1 Inctitute for Imrnunology, Grenzacherstr. 487, CH-4005 Bacel, Switzerland. *Present addresc: Max-Delbrück Centre for Molecular Medicine, Robert- Rocsle Str. 10, D-13122 Berlin. Germany.





























