Embed Size (px)
Citation preview

HUMAN LYSYL HYDROXYLASE ISOFORMSMultifunctionality of human LH3 and the amino acids important for its collagen glycosyltransferase activities
CHUNGUANGWANG
Biocenter Oulu andDepartment of Biochemistry,
University of Oulu
OULU 2002


CHUNGUANG WANG
HUMAN LYSYL HYDROXYLASE ISOFORMSMultifunctionality of human LH3 and the amino acids important for its collagen glycosyltransferase activities
Academic Dissertation to be presented with the assent ofthe Faculty of Science, University of Oulu, for publicdiscussion in Raahensali (Auditorium L10), Linnanmaa, onSeptember 17th, 2002, at 12 noon.
OULUN YLIOPISTO, OULU 2002

Copyright © 2002University of Oulu, 2002
Supervised byProfessor Raili Myllylä
Reviewed byDr. Deborah KaskaProfessor Heather N. Yeowell
ISBN 951-42-6799-0 (URL: http://herkules.oulu.fi/isbn9514267990/)
ALSO AVAILABLE IN PRINTED FORMATActa Univ. Oul. A 388, 2002ISBN 951-42-6798-2ISSN 0355-3191 (URL: http://herkules.oulu.fi/issn03553191/)
OULU UNIVERSITY PRESSOULU 2002

Wang, Chunguang, Human lysyl hydroxylase isoforms. Multifunctionality of humanLH3 and the amino acids important for its collagen glycosyltransferase activitiesBiocenter Oulu, University of Oulu, P.O.Box 5000, FIN-90014 University of Oulu, Finland,Department of Biochemistry, University of Oulu, P.O.Box 3000, FIN-90014 University of Oulu,Finland Oulu, Finland2002
Abstract
Lysyl hydroxylase (EC1.14.11.4, LH) catalyzes post-translationally the hydroxylation of lysylresidues in collagens and other proteins with collagenous domains. Hydroxylysyl residues may alsobe glycosylated by hydroxylysyl galactosyltransferase (EC 2.4.1.50, GT) or galactosylhydroxylysylglucosyltransferase (EC 2.4.1.66, GGT) to form galactosylhydroxylysyl orglucosylgalactosylhydroxylysyl residues, structures unique to collagen.
Three LH isoenzymes (LH1, LH2a/2b, LH3) have been characterized so far. We analyzed mRNAlevels of these isoforms, as well as the mRNAs of the main collagen types (I, III, IV, V) and the αsubunit of PH-4 in different human cell lines. Large variations were found in mRNA expression ofLH1 and LH2 but not LH3. The mRNA levels of LH1, LH2, and the α subunit of PH-4 showedsignificant correlation with each other whereas LH3 correlated with none. No correlation wasobserved between the LH isoforms and individual collagen types.
Three human LH isoforms were expressed in different expression systems. The purifiedrecombinant protein produced by LH3 cDNA was found to be the only one possessing LH, GT andGGT activities. The molecular weight of the partially purified LH3 expressed in Sf9 or Cos-7 cellscorresponded to about 85 kDa whereas that in E.coli cells was about 81 kDa probably due to adeficiency of glycosylation in bacterial cells. The recombinant protein of C. elegans LH cDNA wasexpressed in a cell-free translation system and in E.coli cells. The data indicated that theglycosyltransferase activities, GT and GGT, were also associated with this gene product.
The sequence alignment of LH isoforms from different species revealed that there are 29 aminoacids conserved between human LH3, mouse LH3 and C. elegans LH sequences and scattered evenlyin the molecule, but differing from those of LH1 and LH2. In vitro mutagenesis data showed that theamino acids important for the glycosyltransferase activities were located at the amino-terminal partof the molecule, being separate from the LH active site. Mutation of a conserved LH3 specific, non-disulfide linked cysteine to isoleucine caused a dramatic reduction in GT and GGT activity but hadno effect on LH activity. Mutations of the amino-terminal DxD motif (D187-191) characteristic ofmany glycosyltransferases eliminated both GT and GGT activities, showing the importance of thismotif for collagen glycosyltransferases and suggesting that it might serve as the Mn2+ binding site inthe molecule.
Keywords: collagen biosynthesis, collagen glycosyltransferase, lysyl hydroxylase, post-translational modification


Acknowledgements
This study was carried out at the Biocenter Oulu and Department of Biochemistry, University of Oulu. I wish to first express my heartfelt thanks to my supervisor, Professor Raili Myllylä, for giving the privilege to work in her research group and for her support in many ways during these years. Her everlasting optimism and enthusiasm encouraged me to overcome many frustrating moments and to fulfill this work. I also wish to thank Professor Kalervo Hiltunen and all the other group leaders for creating an inspiring scientific atmosphere and providing excellent research facilities in the department. I am very grateful to Prof. Heather Yeowell and Dr. Deborah Kaska for their critical and valuable comments on the manuscripts of the thesis. Dr. Deborah Kaska is also acknowledged for the careful and rapid rivision of the language. I am indebted to all the former and present members of RM team. My sincere thanks go to Dr. Jari Heikkinen, Dr. Minna Valtavaara, Maija Risteli, M.Sc., and Hanne Luosujärvi, M.Sc., for their collaboration and contribution to the work. Dr. Sakari Kellokumpu is acknowledged for fruitful suggestions. I owe my thanks to Heli Ruotsalainen, M. Sc., Laura Sipilä, M. Sc., Marko Suokas, M. Sc., Dr. Lahja Uitto, Hinni Papponen, M. Sc., Dr. Birgitta Pousi, Antti Salo, M.Sc., and Ms. Anna-Maija Koisti for their friendship, valuable discussions, and assistance with many aspects. I wish to thank Tuula Koret, Anneli Kaattari, Virpi Hannus, Eeva-Liisa Stefanius, Jaakko Keskitalo, Kyösti Keränen, and many others for their expertise in office matters and skillful technical assistance. I would like to thank my friends in the International Women’s Club, especially Ata Bos and Serena Donnini, for sharing many pleasant and relaxing moments in leisure. I am also grateful to my Chinese friends, Tu Hongmin, M. Sc., Drs. Shan Jingdong, Qu Qiang, Qiao Qing, Zheng Aiping, and Qin Zhengdi, the former and present Chinese friends in Dept. of Biochemistry especially Dr. Qin Yongmei, and all the others in the Ping-Pang Club for their valuable discussions concerning all possible matters from the latest news to the Chinese cookings that light up my routine life a lot. My warmest thanks go to all the members of my family and my family-in-law for their encouraging and constant support in many ways whenever needed. My deepest and

dearest thanks belong to my husband Weidong and our son Guanyu for their love, understanding, and for always being the most wonderful part of my life. This work was financially supported by the grants from CIMO, the Sigrid Juselius Foundation, the Academy of Finland, and the University of Oulu. Oulu, April 2002 Chunguang Wang
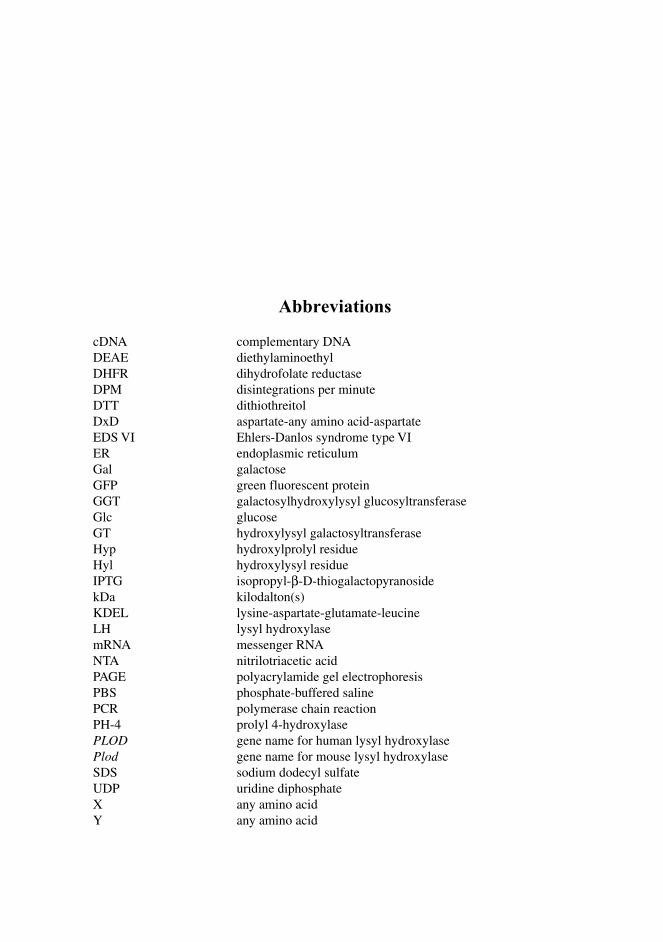
Abbreviations
cDNA complementary DNA DEAE diethylaminoethyl DHFR dihydrofolate reductase DPM disintegrations per minute DTT dithiothreitol DxD aspartate-any amino acid-aspartate EDS VI Ehlers-Danlos syndrome type VI ER endoplasmic reticulum Gal galactose GFP green fluorescent protein GGT galactosylhydroxylysyl glucosyltransferase Glc glucose GT hydroxylysyl galactosyltransferase Hyp hydroxylprolyl residue Hyl hydroxylysyl residue IPTG isopropyl-β-D-thiogalactopyranoside kDa kilodalton(s) KDEL lysine-aspartate-glutamate-leucine LH lysyl hydroxylase mRNA messenger RNA NTA nitrilotriacetic acid PAGE polyacrylamide gel electrophoresis PBS phosphate-buffered saline PCR polymerase chain reaction PH-4 prolyl 4-hydroxylase PLOD gene name for human lysyl hydroxylase Plod gene name for mouse lysyl hydroxylase SDS sodium dodecyl sulfate UDP uridine diphosphate X any amino acid Y any amino acid


List of original articles
This thesis is based on the following articles, which are referred to in the text by their Roman numerals:
I Wang C, Valtavaara M & Myllylä R (2000) Lack of collagen type specificity for lysyl hydroxylase isoforms. DNA and Cell Biology 19: 71-77.
II Heikkinen J, Risteli M∗, Wang C∗, Latvala J, Rossi M, Valtavaara M & Myllylä R (2000) Lysyl hydroxylase is a multifunctional protein possessing collagen glucosyltransferase activity. J Biol Chem 275: 36158-36163. (∗equal contribution)
III Wang C, Risteli M, Heikkinen J, Hussa A.K, Uitto L & Myllylä R (2002) Identification of amino acids important for the catalytic activity of the collagen glucosyltransferase associated with the multifunctional lysyl hydroxylase 3 (LH3). J Biol Chem 277: 18568-18573.
IV Wang C∗, Luosujärvi H∗, Heikkinen J ∗, Risteli M, Uitto L & Myllylä R (2002) The third activity for lysyl hydroxylase 3 (LH3): galactosylation of hydroxylysyl residues in collagens. Matrix Biol, in press. (∗equal contribution)


Contents
Acknowledgements Abbreviations List of original articles Contents 1 Introduction ....................................................................................................................13 2 Review of literature ........................................................................................................14 2.1 Collagens.................................................................................................................. 14
2.1.1 Structure of collagen triple helix ..................................................................... 14 2.1.2 Collagen types................................................................................................. 15 2.1.3 Collagen biosynthesis...................................................................................... 17
2.2 Hydroxylysine.......................................................................................................... 19 2.2.1 Hydroxylysine in collagens and other proteins ............................................... 20 2.2.2 Glycosylation of hydroxylysine in collagens and other proteins..................... 21 2.2.3 Crosslinks in collagen ..................................................................................... 21
2.3 Lysyl hydroxylases................................................................................................... 22 2.3.1 Molecular properties of lysyl hydroxylase ...................................................... 22 2.3.2 Reaction catalysed by lysyl hydroxylase......................................................... 23 2.3.3 Lysyl hydroxylase isoforms ............................................................................ 26 2.3.4 Subcellular localization and distribution of LH isoforms in tissues................ 27 2.3.5 Ehlers-Danlos syndrome type VI .................................................................... 28
2.4 Collagen glycosyltransferases .................................................................................. 29 2.4.1 Purification and molecular properties of collagen glycosyltransferases.......... 30 2.4.2 Catalytic properties of collagen glycosyltransferases ..................................... 31 2.4.3 Intracellular sites of collagen glycosylations .................................................. 33 2.4.4 Collagen glycosyltransferases activities in physiological, pathological, and experimental conditions .................................................................................. 35
3 Aims of the present work ...............................................................................................37 4 Materials and methods ...................................................................................................39 4.1 Cell culture and transfection (I, II)........................................................................... 39 4.2 RNA isolation and Northern blot analysis (I)........................................................... 39 4.3 Production of antibodies (I, II)................................................................................. 40

4.4 Microscopical study (II) ........................................................................................... 40 4.5 Immunoprecipitation (II).......................................................................................... 41 4.6 Immunoblotting (I, II, III, IV) .................................................................................. 41 4.7 In vitro translation (II, III, IV).................................................................................. 41 4.8 Site-directed in vitro mutagenesis (II, III, IV).......................................................... 41 4.9 Expression and purification of the recombinant LH3 proteins (II, III, IV) .............. 42 4.10 GT and GGT activity measurements (II, III, IV).................................................... 44 4.11 Softwares (I, III)..................................................................................................... 44 5 Results ............................................................................................................................45 5.1 Messenger RNA expression levels of LH isoforms and major collagen
types I, III, IV, V in different human cell lines (I) .................................................... 45 5.2 Characterization of the multifunctionality of human LH3 and C. elegans
LH (II, III, IV).......................................................................................................... 46 5.2.1 Human LH3 is a multifunctional protein possessing GGT activity (II) .......... 46 5.2.2 Human LH3 is the only isoenzyme possessing GT activity (IV) .................... 47 5.2.3 GT and GGT activities are associated with C. elegans LH (III, IV) ............... 48
5.3 The intracellular distribution of human LH3 (II) ..................................................... 48 5.4 Identification of the amino acids important for catalytic activities of
GT and GGT associated with human LH3 and C. elegans LH (III, IV)................... 49 5.4.1 Active sites of GGT in human LH3 and C. elegans LH (III)........................... 49 5.4.2 Active sites of GT in human LH3 (IV)............................................................ 50
6 Discussion ...................................................................................................................... 51 6.1 Lack of collagen type specificity for lysyl hydroxylase isoforms (I) ....................... 51 6.2 Characterization of GT and GGT activities associated with human
LH3 and C. elegans LH (II, III, IV) ......................................................................... 52 6.2.1 Human LH3 is a multifunctional protein possessing LH, GT and GGT activities (II, IV)........................................................................ 52 6.2.2 C. elegans LH is multifunctional possessing also glycosyltransferase activities (III, IV)............................................................................................. 53
6.3 Identification of the amino acids important for the glycosyltransferase activities associated with human LH3 and C. elegans LH (III, IV) ......................... 54
6.3.1 Active sites of GGT in human LH3 and C. elegans LH (III)........................... 54 6.3.2 Active sites of GT in human LH3 (IV)............................................................ 55
7 Conclusions.................................................................................................................... 56 8 References...................................................................................................................... 58

1 Introduction
Collagens are a superfamily of glycoproteins, which are found in the extracellular matrix of almost all tissues of the body. They play an important role in maintaining the structures of various tissues and organs, and in regulating cellular behavior. The biosynthesis of collagen is a multi-step process that involves a large number of post-translational modifications, including the hydroxylation of lysyl residues and glycosylation of certain hydroxylysyl residues to galactosylhydroxylysyl and glucosylgalactosylhydroxylysyl residues. The enzymes catalyzing these reactions are lysyl hydroxylase (EC 1.14.11.4, LH), hydroxylysyl galactosyltransferase (EC 2.4.1.50, GT), and galactosylhydroxylysyl glucosyltransferase (EC 2.4.1.66, GGT). The reactions take place within the endoplasmic reticulum. The number of free or glycosylated hydroxylysyl residues varies greatly among different collagen types and within the same collagen type in different tissues and even within the same tissue in different physiological and pathological states. Three isoforms of lysyl hydroxylase (LH1, LH2, LH3) have been identified so far from human and mouse tissues. The expression patterns of the isoforms in different tissues differ from each other. GT and GGT have been purified from chicken embryos and their catalytic and molecular properties have been characterized as well. However, the genes or cDNAs have not yet been cloned. In this work, the mRNA expression levels of the LH isoforms together with the major collagen types I, III, IV, V and the α subunit of prolyl 4-hydroxylase were studied in different human cells and the correlation coefficients between them were analyzed. Both the human LH isoform, LH3, and the C. elegans LH were found to be multifunctional, possessing also GT and GGT activities. These enzymes were characterized and the amino acids critical for GT and GGT activities associated with the human LH3 were identified.

2 Review of literature
2.1 Collagens
Collagens constitute a highly specialized family of extracellular matrix proteins. In addition to the maintenance of the architecture of tissues, they also have other important functions, for instance, in early development and organogenesis, and in the regulation of cell behavior (van der Rest & Garrone 1991, Kielty et al. 1993, Kivirikko 1993, Pihlajaniemi & Rehn 1995, Prockop & Kivirikko 1995). More than twenty collagen types containing altogether at least 38 distinct polypeptide chains have now been identified, and their genes are dispersed among at least fifteen chromosomes. In addition, there are more than fifteen other proteins that have collagen-like domains (Ayad et al. 1998, Myllyharju & Kivirikko 2001).
2.1.1 Structure of collagen triple helix
All collagen molecules are built up from three polypeptide (α) chains, each with a left-handed helical conformation, that are coiled around each other to form a characteristic right-handed collagen triple helix. In addition, they all contain noncollagenous sequences at their termini, and some collagens also have these sequences as interruptions separating adjacent triple-helical regions to make the molecules more flexible. All collagen α chains have repeating -Gly-X-Y- sequences. The occurrence of glycine in every third position is an absolute requirement as glycine is the only residue with a side-chain small enough to fit the restricted space in the center of the triple helix. Proline is frequently in the X-position and Hyp in the Y-position. These residues are required for the correct conformation of the helix because they limit rotation of the polypeptide chains, and their hydrophobic and charged side-chains are located on the surface of the molecule making collagens polymerize into precisely ordered structures. Hyp is also essential for the

15
thermal stability of the helix (van der Rest & Garrone 1991, Kielty et al. 1993, Pihlajaniemi & Rehn 1995, Prockop & Kivirikko 1995).
2.1.2 Collagen types
The collagen superfamily can be divided into two major classes, fibrillar collagens and nonfibrillar collagens, based on the assemblies and other features. There is also a group of proteins called noncollagen proteins that have collagen-like domains but are not defined as collagens.
2.1.2.1 Fibrillar collagens
Collagen types I, II and III are the classical fibril-forming collagens and account for 80-90% of all collagens in the human body. Type V and XI collagens are also classified as fibrillar collagens on the basis of their homology with type I-III collagens (Ayad et al. 1998). The fibrillar collagens have the same overall molecular structures. They all comprise a large triple-helical domain of about 1,000 amino acids, a highly conserved noncollagenous C-terminus and a variable noncollagenous N-terminus. By forming highly ordered quarter-staggered structures, these collagens provide the major mechanical strength for the body in the skeleton, skin, blood vessels, nerves, intestines, and in the fibrous capsules of organs (Vuorio & Crombrugghe 1990, Pihlajaniemi & Rehn 1995). Research suggests that these major collagens can exist as heterotypic fibrils with I and III forming copolymers, and with type V and XI collagens copolymerized largely on the inside of type I and II collagen fibrils, respectively (Ayad et al. 1998).
2.1.2.2 Nonfibrillar collagens
Collagen types IV, VI-X, and XII-XIX do not form fibrils, therefore are defined as nonfibrillar collagens. They exhibit great heterogeneity in structure, tissue distribution, macromolecular organization, and function. One common feature is that they all have one or more imperfections in the collagenous sequences in the triple helical domains which vary in length between about 330 and 1,530 amino acid residues, the shortest being in type VI and the longest being in type VII. Both noncollagenous C- and N- terminal ends are highly variable in sequence and length (Pihlajaniemi & Rehn 1995, Prockop & Kivirikko 1995). Network-forming collagens include collagen types IV, VIII and X. Type IV collagen is only expressed in basement membranes where it is the major component. It aggregates to form three dimensional networks, participates in tissue integrity and filtration. Type VIII and X are very different in structure from type IV but similar to each other. They both

16
form hexagonal networks and belong to short-chain collagens approximately half the size of the fibrillar collagens (van der Rest & Garrone 1991, Kielty et al. 1993, Pihlajaniemi & Rehn 1995, Prockop & Kivirikko 1995). FACITs are fibril associated collagens with interrupted triple helices, which include type IX, XII, XIV, XVI, and XIX collagens. They contain one or two collagenous domains that attach to the surface of preexisting fibrils of fibrillar collagens. Type IX collagen is expressed in cartilage and usually found to be covalently linked to the major fibrillar collagen type II in the same tissue in antiparallel orientation. Types XII and XIV are found not only in tissues rich in type I collagen, but are also present in tissues containing type II collagen. Types XVI and XIX show similarities in structure to the FACITs, and therefore are classified in this subgroup. Type XVI collagen has a broad tissue distribution, being localized predominantly in heart, kidney, smooth muscles, intestine, ovary, testis, eye, and arterial walls. Type XIX is expressed in human rhabdomyosarcoma and fibroblast cell lines (Pihlajaniemi & Rehn 1995, Prockop & Kivirikko 1995, Ayad et al. 1998). Collagen type VI, present in most connective tissues, is the only one known as a beaded filament-forming collagen. The protein contains a rather short unique triple helical domain that accounts for less than half of the total mass of the protein. Another feature is the assembly of collagen type VI monomers into well-defined oligomers which are the building blocks of microfibrils found in tissues and cell cultures and are the best-characterized microfibrillar structures existing in the extracellular matrix (Mayne & Burgeson 1987, van der Rest & Garrone 1991, Prockop & Kivirikko 1995). Collagen type VII, one of the largest known collagens, forms anchoring fibrils that link the basal surface of epithelial cells with the underlying dermis. This network may provide a special trans-basement-membrane route for the transmission of information from the dermis to the epithelial cells or vice versa (Mayne & Burgeson 1987). MACITs are the membrane-associated collagens with interrupted triple helices, including collagen types XIII and XVII (Pihlajaniemi & Rehn 1995). Type XIII collagen is found in many tissues and is a transmembrane component of focal adhesion sites (Hägg et al. 2001). Type XVII collagen is located in hemidesmosomes connecting epithelial cells to the matrix in skin, cornea and lung. It is identified as the autoantigen associated with the blistering skin disease bullous pemphigoid as well as bullous diseases of other epithelia including cornea and mucous membranes (Giudice et al. 1991, Tajima & Tokimitsu 1995, Gordon et al. 1997, Aho et al. 1999, Aho & Uitto 1999, Michelson et al. 2000) Both collagen type XV and XVIII have a highly interrupted triple helix together with large globular domains at both N- and C-termini. They contain several potential attachment sites for serine-linked glycosaminoglycans and asparagines-linked oligosaccharides. They are widely expressed in the basement membrane zones of most tissues, but type XVIII collagen is found at a much higher level in the liver (Pihlajaniemi & Rehn 1995, Prockop & Kivirikko 1995). Type XV collagen is a structural component of the extracellular matrix needed for stabilizing skeletal muscle cells and microvessels (Eklund et al. 2001) whereas type XVIII collagen is needed for the normal development of the eye (Fukai et al. 2002). The C-terminal proteolytic fragment of the type XVIII collagen, endostatin, has been reported being an endogenous inhibitor of angiogenesis and tumor growth (O’Reilly et al. 1997).

17
2.1.2.3 Recent discoveries
Currently complete cDNA sequences for four additional collagen polypeptide chains are available, expanding the collagen superfamily to over 20 members. They encode a fibril-forming collagen-like chain, two FACIT collagen-like chains, and a type XIII collagen-like chain (Koch et al. 2001, Myllyharju & Kivirikko 2001). Furthermore, there are at least fifteen other proteins containing triple helical collagenous domains but not defined as collagens. These include the subcomponent C1q of complement, the tail structure of acetylcholinesterase, the pulmonary surfactant proteins SP-A and SP-D, mannan-binding protein, collectin-43, conglutinin, the ficolins, type I and II macrophage scavenger receptor, MARCO protein, an adipose-specific collagen-like factor apM1, a src-homologous-and-collagen (SHC) protein, aggretin and ectodysplasin (Beck & Brodsky 1998, Kivirikko & Pihlajaniemi 1998, Chung et al. 1999, Ezer et al. 1999, Kraal et al. 2000, Myllyharju & Kivirikko 2001).
2.1.3 Collagen biosynthesis
Collagen biosynthesis is a multistep process that starts with the transcription and translation of the individual collagen gene (Kivirikko & Myllylä 1984, Kielty et al. 1993, Kivirikko 1993). It is characterized by the presence of a large number of co- and post-translational modifications, many of them being unique to collagens or collagen-like proteins. (Kivirikko & Myllylä 1982, Kivirikko & Myllylä 1984, Kielty et al. 1993, Kivirikko 1993, Prockop & Kivirikko 1995). The fibril-forming collagens are synthesized as procollagens on the ribosomes of the rough ER (see Figure 1). The intracellular modifications occur when the procollagens are translocated across the ER membrane into the lumen. These modifications include the removal of signal peptides; hydroxylation of prolyl and lysyl residues to 4-Hyp, 3-Hyp, and Hyl residues; glycosylation of certain hydroxylysyl residues to galactosylhydroxylysyl and glucosylgalactosylhydroxylysyl residues; glycosylation of a mannose-rich oligosaccharide on one or both of the propeptides; chain association; disulfide bonding; and formation of a triple helix (Table 1, for reviews see Kivirikko & Myllylä 1982, Kivirikko & Myllylä 1984, Kielty et al. 1993, Ayad et al. 1998). The mechanism of procollagen secretion is poorly understood, but it is known that procollagen follows the classical secretion route for extracellular proteins, passing through the Golgi complex to the extracellular space (Kielty et al. 1993). Extracellular modifications consist of removal of large peptides from both N- and C-termini of the procollagen, ordered aggregation, and crosslink formation. These events convert the procollagens to collagens and incorporate the collagen molecules into stable cross-linked fibrils or other supramolecular aggregates (Kivirikko & Myllylä 1984, Kielty et al. 1993, Kivirikko 1993, Prockop & Kivirikko 1995, Myllyharju & Kivirikko 2001). The processing and assembly of other collagens basically follow the same steps as for fibrillar collagens but with some exceptions. For example, the N- and/or C-terminal propeptides of many collagens are not cleaved; some collagens undergo N-glycosylation

18
or have additional processing steps such as the addition of glycosaminoglycan side chains (Prockop & Kivirikko 1995, Myllyharju & Kivirikko 2001).
Fig. 1. Biosynthesis of a fibrillar collagen. Procollagen polypeptide chains are synthesized on the ribosomes of the rough ER and secreted into the lumen, where the chains are modified by hydroxylation of certain prolyl and lysyl residues and glycosylation before chain association and triple helix formation. The procollagen molecules are secreted into the extracellular space where the N and C propeptides are cleaved by specific proteases. The collagen molecules then assemble into fibrils that are stabilized by the formation of crosslinks (Modified from Myllyharju and Kivirikko 2001 with permission of Taylor & Francis AB).

19
Table 1. General steps in collagen biosynthesis and the post-translational processing enzymes
Biosynthetic step Enzyme Biological significance
A. Transcription and translation
1. Biosynthesis of pre-mRNA Formation of translatable mRNA
2. Translation Formation of primary structure
B. Intracellular modifications
1. Cleavage of signal peptide of pre-proα chain Signal peptidase Membrane translocation
2. 4-Hydroxylation of proline Prolyl 4-hydroxylase Essential for triple helix at 37°C
3. 3-Hydroxylation of proline Prolyl 3-hydroxylase Unknown
4. Hydroxylation of lysine Lysyl hydroxylase Essential for glycosylation of
hydroxylysine and crosslinks
5. O-glycosylation of hydroxylysine GT Unknown
6. O-glycosylation of galactosylhydroxylysine GGT Unknown
7. Chain association and disulfide bonding Protein disulfide Essential for triple helix formation
isomerase
8. Triple helix formation Essential for secretion and later
molecular function
9. Translocation and secretion of procollagen Transport
C. Extracellular modifications
1. Conversion of procollagen to collagen Procollagen N-proteinase Essential for normal fibril
Procollagen C-proteinase formation
2. Ordered aggregation Formation of fibrils or other
native structures
3. Crosslink formation Lysyl oxidase Essential for stability of native
structure
Modified from the tables presented by Kivirikko and Myllylä 1984 and Kielty and coauthors 1993 with permissions from Elsevier Science and Wiley-Liss, Inc., respectively.
2.2 Hydroxylysine
Hydroxylysine is found only in animal proteins and mostly in collagens. The hydroxyl groups of hydroxylysine residues have two important functions: they serve as attachment sites for carbohydrates and they play a crucial role in stabilizing intra- and intermolecular crosslinks. Hydroxylysine also exists in collagen-like domains of several other proteins that are not defined as collagens. (Kivirikko et al. 1992, Kivirikko & Pihlajaniemi 1998).

20
2.2.1 Hydroxylysine in collagens and other proteins
The content of hydroxylysine in the triple-helical domains of collagen peptide chains varies from 6 [α1(III)] to 67 [α2(VI)] residues per 1,000 amino acids. Only 17% of the lysyl residues are hydroxylated in collagen type III, whereas almost 90% are hydroxylated in collagen type IV and about 80% in collagen type VI. Additional variations are found in hydroxylysine content within the same collagen type and even in the same tissue under different physiological and pathological conditions (Kivirikko & Myllylä 1980, Kivirikko et al. 1992, Kivirikko & Pihlajaniemi 1998). Hydroxylysine is typically found in triple-helical regions almost exclusively in the Y positions of the repeating -X-Y-Gly- sequences in various collagens. In some collagens it is found from the short nontriple-helical regions at the ends of the α chain as -X-Hyl-Ala- or -X-Hyl-Ser- (Kivirikko & Myllylä 1980, Kivirikko et al. 1992). The hydroxylation of many lysyl residues in the Y position is not complete and many of them are not hydroxylated at all in some collagens. This explains the differences observed in hydroxylysine content within the same collagen type from various sources, and also partly explains the differences found among various collagen types. The latter may also be due to the differences in total amounts of lysyl residues incorporated per 1,000 amino acids and in the distribution of incorporated lysyl residues between the X and Y positions (Kivirikko & Myllylä 1980, Kivirikko et al. 1992, Kivirikko & Pihlajaniemi 1998). It seems evident that lysyl residues are more readily hydroxylated in certain triplets than in others. It has been reported that embryonic tissues contain much more hydroxylysine than adult tissues (Kielty et al. 1993, Bateman et al. 1996). There is also evidence that hydroxylation of lysyl residues in collagens is associated with the mineralization of fibrils, the high hydroxylation level preventing a deposit of minerals between fibrils (Wassen et al. 2000). Lysine hydroxylation seems to be increased as well in some diseases, for example, lipodermatosclerosis (Brinckmann et al. 1999), osteoporosis (Köwits et al. 1997, Lo Cascio et al. 1999), and osteogenesis imperfecta (Brenner et al. 1989, Tenni et al. 1993, Lehmann et al. 1995, Bank et al. 2000). Hydroxylysine has also been identified in some ‘noncollagen’ collagens, such as the C1q subcomponent of complement, acetylcholinesterase, mannose binding proteins, type I and II macrophage scavenger receptors and bovine conglutinin (Kivirikko et al. 1992). Angerfish somatostatin-28 contains a single hydroxylysyl residue in the sequence -Trp-Hyl-Gly- (Andrews et al. 1984, Spiess & Noe 1985). It has been reported that the full-length adiponectin produced from mammalian cells acts functionally as an insulin sensitizer. The four lysyl residues conserved in all species within the collagenous domain are all hydroxylated and sequentially glycosylated. It is suggested that the hydroxylation and glycosylation might contribute to the insulin-sensitizing activity of adiponectin with respect to the inhibition of hepatic glucose production (Wang et al. 2002).

21
2.2.2 Glycosylation of hydroxylysine in collagens and other proteins
The glycosylation of hydroxylysyl residues occurs as a post-translational event in collagen biosynthesis. The carbohydrates linked to hydroxylysyl residues are either a monosaccharide galactose or a disaccharide glucosylgalactose. The formation of hydroxylysine linked carbohydrate units is catalyzed by two specific enzymes called hydroxylysyl galactosyltransferase (EC 2.4.1.50, GT) and galactosylhydroxylysyl glucosyltransferase (EC 2.4.1.66, GGT) (details see 2.4). The extent of glycosylation of the hydroxylysyl residues and the ratio of galactosylhydroxylysine to glucosylgalactosylhydroxylysine vary considerably among different collagen types and within the same collagen type from various sources (Kivirikko & Myllylä 1979, Kivirikko & Myllylä 1984, Kivirikko et al. 1992). Hydroxylysyl residues in the Y position of -Gly-X-Y- triplets in collagen type VI are fully glycosylated (Ayad et al. 1998). The chains of collagen type II and IV are also highly glycosylated from 6% in α1(II) to almost 15% in α1(IV) whereas the chains of collagen type I and III contain very low levels of hydroxylysyl-linked carbohydrate, being only 0.5-1% (Kivirikko & Myllylä 1979, Kivirikko & Myllylä 1984). The function of the hydroxylysyl-linked carbohydrate units remains poorly understood. It has been suggested that they may play a role in the control of the organization of the fibrils. Studies on fibrillar collagens have indicated that an increase content of glycosylated hydroxylysyl residues is associated with a decrease of fibril diameter both in vivo and in vitro (Brinckmann et al. 1999, Notbohm et al. 1999). Adiponectin, the fat-derived hormone, has recently been shown to be able to decrease hyperglycemia and to reverse insulin resistance (Berg et al. 2001, Fruebis et al. 2001, Yamauchi et al. 2001). The endogenous adiponectin secreted by adipocytes is post-translationally modified to eight different isoforms and, six of them are glycosylated. The glycosides linked to the four conserved hydroxylysyl residues in the collagenous domain are probably the glucosylgalactosyl group. In vitro mutagenesis data has demonstrated that hydroxylation and glycosylation of the conserved hydroxylysyl residues are required for its insulin-sensitizing action (Wang et al. 2002).
2.2.3 Crosslinks in collagen
Crosslinking takes place in at least two stages. The initial reaction catalyzed by peptidyl lysyl oxidase is oxidative deamination of ε-amino groups of specific lysyl and hydroxylysyl residues in telopeptide regions of collagens to form reactive aldehydes (allysine and hydroxyallysine). These aldehydes subsequently participate in the formation of the various crosslinks, either by aldol condensation of two of the aldehydes or by condensation between one aldehyde and one ε-amino group of a second lysyl, hydroxylysyl, or glycosylated hydroxylysyl residue. The crosslinks formed from hydroxylysine-derived aldehyde are more stable than those formed from lysine-derived aldehyde (Kivirikko & Myllylä 1984). The lysine aldehyde pathway occurs primarily in adult skin, cornea, and sclera whereas the hydroxylysine aldehyde pathway predominates

22
in bone, cartilage, ligament, most tendons, embryonic skin, and most major internal connective tissues of the body (Eyre 1987, Kielty et al. 1993).
2.3 Lysyl hydroxylases
Lysyl hydroxylase (EC 1.14.11.4, LH) catalyzes the hydroxylation of lysyl residues in collagens and some other proteins with collagenous domains (Kivirikko & Myllylä 1980, Kivirikko & Myllylä 1984, Kivirikko et al. 1992, Kielty et al. 1993). At an early stage it was suggested that the same enzyme, protocollagen hydroxylase, catalyzed the hydroxylations of proline and lysine in collagens since they require the same substrates and cofactors (Hausmann 1967, Kivirikko & Prockop 1967, Kivirikko et al. 1968). It was found out later that both enzymatic activities were located at least in different sites in the enzyme (Weinstein et al. 1969). Lysyl hydroxylase was first partially purified from chick embryos and was demonstrated to be a separate enzyme from prolyl hydroxylase by DEAE cellulose chromatography (Kivirikko & Prockop 1972, Popenoe & Aronson 1972). The enzyme was further purified up to several hundred- or thousand-fold from chick embryos and chick embryo cartilage (Ryhänen 1976). By using a procedure involving (NH4)2SO4 fractionation, affinity chromatography on concanavalin A-agarose, affinity chromatography on a collagen-agarose column and gel filtration (Turpeenniemi et al. 1977), lysyl hydroxylase was finally purified as a homogeneous protein from chick embryos (Turpeenniemi-Hujanen et al. 1980) and from human fetal and placenta tissues (Turpeenniemi-Hujanen et al. 1981).
2.3.1 Molecular properties of lysyl hydroxylase
The active lysyl hydroxylase is a dimer with a molecular weight of about 190 kDa by gel filtration, which consists of only one type of monomer with a molecular weight of about 85 kDa seen in SDS-PAGE (Turpeenniemi-Hujanen et al. 1980, Turpeenniemi-Hujanen et al. 1981, Myllylä et al. 1988). It is a glycoprotein containing asparagine-linked carbohydrate units that are essential for maximal enzyme activity (Myllylä et al. 1988, Kivirikko et al. 1992, Kivirikko & Pihlajaniemi 1998). Lysyl hydroxylase was first cloned from chick embryos (Myllylä et al. 1991). The mRNA is 4.0 kb in size and the cDNA encodes a polypeptide of 710 amino acid residues with a signal peptide of 20 amino acids at the N-terminus. Surprisingly there is no significant homology in the primary structures between lysyl hydroxylase and the two types of subunit of prolyl 4-hydroxylase, despite the fact that both enzymes share much similarity in their catalytic properties (Myllylä et al. 1991). Molecular cloning and nucleotide sequencing have been reported later for the human (Hautala et al. 1992, Yeowell et al. 1992) and rat lysyl hydroxylases (Armstrong & Last 1995). The mRNA of human lysyl hydroxylase is 3.2 kb. Its subunit consists of 709 amino acid residues and a signal peptide of 18 amino acids. The overall identity between the sequences of human

23
and chick lysyl hydroxylase is 76% at the amino acid level. The carboxytermini are very well conserved between the two enzymes. The human sequence contains 10 cysteine residues, 9 of which are conserved in the chick sequence. The gene for human lysyl hydroxylase (PLOD) is mapped to chromosome 1p36.2-36.3 (Hautala et al. 1992). The mRNA of rat lysyl hydroxylase is 3.2 kb in size. It encodes a protein consisting of 728 amino acid residues, which might contain a signal peptide of 18 amino acids. The complete cDNA of rat lysyl hydroxylase is 91% and 77% identical to those of human and chick at the amino acid level (Armstrong & Last 1995). The chick, human and rat lysyl hydroxylase all contain four potential attachment sites for asparagine-linked oligosaccharides (Myllylä et al. 1991, Hautala et al. 1992, Armstrong & Last 1995).
2.3.2 Reaction catalyzed by lysyl hydroxylase
Lysyl hydroxylase belongs to the group of 2-oxoglutarate dioxygenases. It requires Fe2+, 2-oxoglutarate, O2 and ascorbate in the reaction, and decarboxylates 2-oxoglutarate, one atom of the O2 molecule being incorporated into the succinate while the other is incorporated into the hydroxyl group of the substrate (Kivirikko & Myllylä 1984, Kivirikko et al. 1992).
COOH | COOH CH2 | | Ascorbate CH2 Substrate-H + CH2 + O2 → Substrate-OH + | + CO2 | Fe 2+ CH2 CO | | COOH COOH
Fig. 2. Hydroxylation reaction catalyzed by lysyl hydroxylase. One atom of oxygen is incorporated into the hydroxyl group of the lysyl residue in the peptide substrate and another one into 2-oxoglutarate, which is decarboxylated to form succinate and liberates CO2.
The hydroxylation reaction involves an ordered binding of Fe2+, 2-oxoglutarate, O2 and the peptide substrate to the enzyme, and an ordered release of the hydroxylated peptide, CO2, succinate, and Fe2+, in which Fe2+ may not necessarily leave the enzyme during each catalytic cycle (Puistola et al. 1980a, 1980b). Lysyl hydroxylase is also able to catalyze the uncoupled decarboxylation of 2-oxoglutarate in the presence of the same cofactors but in absence of the peptide substrate. The reaction rate, however, is only 4% of the one

24
observed in the presence of a saturating concentration of the peptide substrate (Puistola et al. 1980a, 1980b, Myllylä et al. 1984).
2.3.2.1 Peptide substrates
The minimum sequence requirement for lysyl hydroxylase is fulfilled by the -X-Lys-Gly- triplet, but the enzyme purified from chick embryos can also hydroxylate arginine-rich histone, which does not contain any -X-Lys-Gly- triplet but the sequence such as -X-Lys-Ser-, -X-Lys-Ala-, and -X-Lys-Thr- (Ryhänen 1975). This agrees with the presence of the sequences -X-Hyl-Ser- and -X-Hyl-Ala- in the short noncollagenous domains at the end of the α-chains in some collagens (Kivirikko & Myllylä 1980, Kivirikko et al. 1992). It is not yet known whether one single enzyme hydroxylates both -X-Lys-Gly- and other triplets or whether these reactions are catalyzed by different LH isoenzymes in vivo (Ryhänen 1976, Royce & Barnes 1985, Bank et al. 1999). The interaction with lysyl hydroxylase is also influenced by the amino acid sequence around the lysyl residue, the peptide chain length and the peptide conformation. Lysyl hydroxylase does not act on a -Lys-Gly-Pro- tripeptide whereas -Ile-Lys-Gly- can be hydroxylated. The peptide chain length appears to influence only the Km, which decreases with the increasing chain length, whereas the V of the reaction seems to be unaffected (Kivirikko & Myllylä 1980, Kivirikko et al. 1992). The triple-helical conformation of the substrates completely prevents lysine hydroxylation (Kivirikko & Myllylä 1980, Kivirikko et al. 1992). An extended polyproline-II conformation in the peptide substrate may interact at the binding site on lysyl hydroxylase, while a ‘bent’ structure such as the γ- or β-turn in the -Lys-Gly- segment may be necessary for hydroxylation at the catalytic site (Jiang & Ananthanarayanan 1991, Ananthanarayanan et al. 1992).
2.3.2.2 Cofactors
The hydroxylation reaction catalyzed by lysyl hydroxylase requires Fe2+, 2-oxoglutarate, O2 and ascorbate as essential cofactors in an ordered mechanism that leads to the release of a hydroxylated lysyl residue in the procollagen polypeptide, CO2, and succinate (Kivirikko & Myllylä 1980, Puistola et al. 1980b, Kivirikko et al. 1992, Kivirikko & Pihlajaniemi 1998). The Km values of the cofactors for lysyl hydroxylase are: 2 µM for Fe2+, 100 µM for 2-oxoglutarate, 45 µM for O2 and 350 µM for ascorbate (Turpeenniemi-Hujanen et al. 1981, Pirskanen et al.1996, Kivirikko & Pihlajaniemi 1998). The Km values for Fe2+, O2 and ascorbate in the case of lysyl hydroxylase is essentially the same as that of prolyl 4-hydroxylase whereas that for 2-oxoglutarate is five times higher than prolyl 4-hydroxylase (Kivirikko & Pihlajaniemi 1998). The Fe2+ is loosely bound to the enzyme by three side chains (Kivirikko et al. 1992). It doesn’t have to leave the enzyme after every catalytic cycle (Puistola et al. 1980b).

25
Sequence alignment analysis of several 2-oxoglutarate dioxygenases and a related enzyme, isopenicillin N synthase, demonstrates a weak homology within two histidine-containing motifs, His-1 and His-2, located about 50-70 amino acids apart being the residue 656 and 708 in human lysyl hydroxylase. Aspartate residue 658 is also conserved in all enzymes compared (Myllylä et al. 1992). Site-directed mutagenesis studies show that mutations of the conserved His656 to serine, Asp658 to alanine in His-1, and His708 to serine in His-2 completely inactivates human lysyl hydroxylase (Pirskanen et al. 1996), suggesting that the three Fe2+ binding ligands in human lysyl hydroxylase are His656, Asp658, and His708 (Kivirikko & Pihlajaniemi 1998). 2-oxoglutarate is a very specific requirement for the hydroxylation reaction (Kivirikko & Myllylä 1980, Kivirikko et al. 1992, Kivirikko & Pihlajaniemi 1998). However, it can be replaced by 2-oxoadipinate although the Km value is significantly higher for the latter (4.8 mM) (Majamaa et al. 1985). The Km value of 2-oxoglutarate for lysyl hydroxylase is much higher than that of other collagen hydroxylases, implying differences in the structures of the 2-oxoglutarate binding sites, which consist of three distinct subsites. Subsite I is supposed to be a positively charged side chain on the enzyme which binds the C5 carboxyl group of the 2-oxoglutarate. Subsite II is assumed to comprise 2 cis-positioned coordination sites of the enzyme-bound Fe2+, which is chelated by the C1-C2 moiety. Subsite III might involve a hydrophobic binding site in the C3-C4 region of the cofactor (Kivirikko & Pihlajaniemi 1998). Site-directed mutagenesis studies suggest that the residue forming subsite I in human lysyl hydroxylase is Arg718 , which is also located in the His-2 motif of the enzyme and binds the C5 carboxyl group of 2-oxoglutarate (Passoja et al. 1998a). The molecular oxygen needed for the hydroxylation reaction comes from the atmosphere. The oxygen atoms present in an enzyme-bound intermediate can be exchanged with water (Kikuchi et al. 1983). The first activated intermediate is a dioxygen unit bound to the Fe2+ of the enzyme whereas the final active intermediate carrying out the hydroxylation reaction is probably a ferryl ion (Kivirikko et al. 1992). Lysyl hydroxylase can complete many catalytic cycles at a maximal rate in the complete absence of ascorbate. Hydroxylation then stops very quickly, however, and ascorbate is required to reactivate the enzyme (Puistola et al. 1980a, Kivirikko et al. 1992, Kivirikko & Pihlajaniemi 1998). Occasionally lysyl hydroxylase catalyzes uncoupled reactions even in the presence of the peptide substrate. In these reactions the reactive iron-oxo complex is probably converted to Fe3+.O-, making the enzyme unavailable for new catalytic cycles until reduced by ascorbate. It is very possible that the main biological function of ascorbate in vivo is to be an alternative oxygen acceptor after such uncoupled cycles (Myllylä et al. 1984). Ascorbate can be replaced by cysteine or dithiothreitol to a minor extent (Puistola et al. 1980a). The studies with prolyl 4-hydroxylase reveal that the ascorbate binding site of the hydroxylation enzyme is partially identical to the binding site of 2-oxoglutarate, and modifications of the ring atoms of ascorbate that abolish the capacity to bind iron destroy the cofactor activity, as in L-galactono γ-lactone and 3-methoxy-L-ascorbate (Majamaa et al. 1986, Kivirikko et al. 1992, Kivirikko & Pihlajaniemi 1998). Dithiothreitol, bovine serum albumin, and catalase are also needed for lysyl hydroxylase in order to obtain maximal activity in vitro (Kivirikko & Myllylä 1980). The stimulatory effect by dithiothreitol suggests that the catalytic site contains free thiol

26
groups essential for the enzyme activity. The action of bovine serum albumin is partly due to the nonspecific ‘protein effect’, but more likely it is because of the presence of many free thiol groups on this protein. Catalase is probably to act partly by destroying peroxide generated non-enzymatically from Fe2+, O2 and ascorbate, and partly by a nonspecific protein effect as well (Kivirikko & Myllylä 1980, Kivirikko et al. 1992).
2.3.3 Lysyl hydroxylase isoforms
Many findings suggest that lysyl hydroxylase may have tissue-specific or collagen type-specific isoenzymes (Ryhänen 1975, Risteli et al. 1980, Turpeenniemi-Hujanen 1981, Puistola 1982, Puistola & Anttinen 1982, Tajima et al. 1983, Ihme et al. 1984, Royce & Barnes 1985). During the past few years, two novel isoenzymes, termed lysyl hydroxylase 2 (LH2) and lysyl hydroxylase 3 (LH3), have been characterized from human (Valtavaara et al. 1997, Passoja et al. 1998b, Valtavaara et al. 1998) and mouse (Ruotsalainen et al. 1999) tissues. The previous known main isoform is now correspondingly termed lysyl hydroxylase 1 (LH1). In addition, LH2 occurs in two alternatively spliced forms, LH2a and LH2b, in which LH2b contains an additional exon of 63 bp to the LH2a sequence (Valtavaara 1999, Yeowell & Walker 1999a). The mRNA of human LH2 is 4.2 kb in size. LH2a cDNA encodes a protein of 737 amino acids that includes a signal peptide of 25 residues at the N-terminus whereas LH2b cDNA encodes a protein of 758 amino acids, of which 21 amino acids are encoded by the additional exon 13A (Valtavaara et al. 1997, Valtavaara 1999, Yeowell & Walker 1999a, Yeowell 2002). The overall similarity of amino acid sequence between LH1 and LH2 is over 75%, being even higher at the C-terminus of the molecules. LH2 has ten cysteine residues that are all conserved in LH1, and seven potential N-glycosylation sites (Valtavaara et al. 1997, Yeowell & Walker 1999a). The corresponding gene (PLOD2) has been localized to chromosome 3q23-24 (Szpirer et al. 1997). The mRNA of human LH3 is 2.8 kb, smaller than that of LH1 and LH2. The cDNA encodes a polypeptide of 738 amino acids including a signal peptide of 24 residues at the N-terminus (Passoja et al. 1998b, Valtavaara et al. 1998, Yeowell 2002). The amino acid sequence shows high identity to LH1 and LH2, both being 59%. The C-terminus is highly conserved in all isoforms. Nine out of ten cysteine residues conserved between LH1 and LH2 are also conserved in LH3. There are only two potential N-glycosylation sites in the molecule. The gene (PLOD3) is localized to chromosome 7q22 (Valtavaara et al. 1998, Valtavaara et al. 2000). The mRNAs of mouse LH1, LH2, LH3 are 3.0 kb, 3.9 kb, and 3.1 kb, respectively. The cDNA for mouse LH1 encodes a 728 amino acid polypeptide including a putative 18 amino acid N-terminal signal peptide. The mouse LH2 cDNA encodes a protein of 737 amino acids including a putative signal peptide of 27 amino acids. The cDNA of mouse LH3 encodes a polypeptide of 741 amino acids with the first 27 amino acids forming a signal peptide at the N-terminus. The amino acid sequences are approximately 60% identical between the mouse LH isoforms and 91% identical to the corresponding human

27
enzymes (Ruotsalainen et al. 1999). The genes encoding mouse LH isoforms (Plod1, Plod2, Plod3) map to chromosome 4, 5, and 9, respectively (Sipilä et al. 2000). The complete gene structures for human LH1 (Heikkinen et al. 1994) and LH3 (Rautavuoma et al. 2000), and for mouse LH2 and LH3 (Ruotsalainen et al. 2001) have been characterized so far. Although they basically all have 19 exons and 18 introns, the LH3 genes (Plod3 and PLOD3) are much smaller than PLOD1 and Plod2 due to the existence of many shorter introns in the sequences. Plod2 is the largest gene with an extra exon 13A being alternatively spliced in the processing of mRNA. The sizes of exon 1 and exon 19, which cover most of the 5’ and 3’ ends of the coding regions and the untranslated regions of mRNAs, are very different among these LH isoforms whereas the sizes of the other exons are very similar to each other. The introns of Plod3/PLOD3 constitute only 69-76% of the gene sequence whereas PLOD1 and Plod2 are about 91-92% (Ruotsalainen et al. 2001).
2.3.4 Subcellular localization and distribution of LH isoforms in tissues
LH1 has been demonstrated by a variety of techniques to reside within the lumen of the ER. It appears to be a luminally oriented peripheral membrane protein that associates with the membrane by weak electrostatic interactions (Kivirikko et al. 1992, Kellokumpu et al. 1994, Kivirikko & Pihlajaniemi 1998). Interestingly LH1, as a resident of the ER, does not contain either of the two previously characterized ER-specific retention signals (KDEL or the double lysine motif) in its primary structure (Myllylä et al. 1991, Hautala et al. 1992). It is shown later that cathepsin D, a soluble lysosomal protease, is able to be converted into a membrane-associated protein by tagging a 40-amino acid C-terminal peptide segment of LH1 into the molecule. The first 25 amino acids seem to play a crucial role in terms of membrane association and ER localization. The data thus reveals a novel retrieval mechanism by which the ER lumen can retain its specific protein components from the bulk flow (Suokas et al. 2000). No data is available thus far concerning the subcellular localization of LH2 and LH3. The human LH1 gene is expressed constitutively in many tissues such as placenta, skin fibroblasts, aorta, lung, vein, artery, cartilage, spleen, gall bladder, brain, liver and skeletal muscle (Heikkinen et al. 1994, Yeowell et al. 1994). Human LH2a is mainly expressed in placenta, pancreas, heart, liver, brain, kidney, skeletal muscle and spleen (Valtavaara et al. 1997, Yeowell & Walker 1999a) whereas LH2b is widely expressed at variable levels in different tissues (Valtavaara 1999, Yeowell & Walker 1999a). LH3 is strongly expressed in heart, placenta and pancreas (Valtavaara et al. 1998). The expression of mouse LH1 is extremely high in liver and heart, as well as in skeletal muscle, kidney and lung (Ruotsalainen et al. 1999). The mouse LH2a and LH2b genes are highly expressed in heart and kidney (Ruotsalainen et al. 1999, Valtavaara 1999) whereas LH3 expression is higher in heart, liver, lung and testis (Ruotsalainen et al. 1999). The phylogenetic analysis of nine LH sequences from five species suggests that the LH isoforms are derived from one ancestral gene by two duplication events. LH1 and LH2 result from more recent duplications and are more closely related to each other whereas

28
LH3 appears to be the ancestral gene and resembles the C. elegans LH more than the other isoforms (Ruotsalainen et al. 1999).
2.3.5 Ehlers-Danlos syndrome type VI
The Ehlers-Danlos syndrome (EDS) is a heterogeneous group of disorders characterized clinically by joint hypermobility, skin fragility and hyperextensibility and other signs of connective tissue involvement. At least ten different subtypes of EDS have been classified based on genetic, biochemical, and clinical characteristics. The type VI variant of EDS is a recessively inherited connective-tissue disorder with specific features such as muscular hypotonia, kyphoscoliosis, and ocular manifestations (Krane 1984, Steinmann et al. 1993, Yeowell & Pinnell 1993, Byers 1994, Kivirikko & Pihlajaniemi 1998). Patients with EDS VI are biochemically divided into two subclasses: those with a low LH1 activity classified as EDS VIA (Krane et al. 1972, Pinnell et al. 1972, Sussman et al. 1974, Steinmann et al. 1975, Chamson et al. 1987, Wenstrup et al. 1989), and those with normal LH1 activity classified as EDS VIB (Judisch et al. 1976, Ihme et al. 1983, Royce et al. 1989, Steinmann et al. 1993). Molecular cloning of the human LH1 has made it possible to characterize mutations responsible for this disorder in detail (Yeowell & Walker 2000). The first mutation leading to a deficiency in LH1 activity in EDS VI patients was reported by Hyland and his coworkers (Hyland et al. 1992). Two patients in a family had a homozygous single base substitution in the LH1 gene that converted the CGA codon for Arg319 to a translation termination codon TGA (R319X). The parents and three healthy siblings of the patients were found to be heterozygous carriers of the same mutation (Hyland et al. 1992). The most common mutation seems to be a large seven exon duplication resulting from recombination of Alu-sequences in the LH1 gene. This has been found in approximately 19% of 35 EDS VI families studied (Hautala et al. 1993, Pousi et al. 1994, Heikkinen et al. 1997). Three other mutations have been shown to occur in more than one unrelated EDS VI patient. These include the Y511X mutation in exon 14 (Yeowell & Walker 1997, Walker et al. 1999, Yeowell & Walker 1999b, Pousi et al. 2000), the 15-bp deletion in exon 11 (Yeowell et al. 2000a), and the Q327X mutation in exon 10 (Yeowell et al. 2000b). Other mutations identified so far include Y142X, R670X, the insertion of a single C nucleotide in exon 16 (Yeowell et al. 2000b), a homozygous splice-site mutation that induces the skipping of exon 9 (Pajunen et al. 1998), and some compound heterozygous mutations, for example, a point mutation (exon 19) and a triplet deletion (exon 15) in the two alleles (Ha et al. 1994), a deletion of exon 17 in one allele and a splicing defect resulting in the skipping of exon 16 in the other allele (Pousi et al. 1998), a point mutation in exon 17 of one allele and an unidentified mutation in the other allele (Brinckmann et al. 1998), a nucleotide deletion in the acceptor splice site of intron 4 in one allele, and an insertion of a C nucleotide in exon 2 of the other allele (Heikkinen et al. 1999).

29
2.4 Collagen glycosyltransferases
Grassman and his coworkers proved in 1957 that neutral sugars are covalently linked to collagen. Butler, Cunningham, and many others in the late 60’s and early 70’s demonstrated the covalent linkage of the sugars to hydroxylysine, and the carbohydrate units turned out to be either a monosaccharide galactose or a disaccharide glucosylgalactose. These are the only sugars found in the triple helical region of collagens (Myllylä 1976, Kivirikko & Myllylä 1979, Kielty et al. 1993, Prockop & Kivirikko 1995, Bateman et al. 1996). These hydroxylysyl glycosides have also been identified in the C1q subcomponent of complement that contains collagen-like amino acid sequences (Kivirikko & Myllylä 1979). The structure of 2-O-α-D-glycopyranosyl-O-β-D-galactopyranosylhydroxylysine involving an unusual α1→2-O-glycosidic linkage between glucose and galactose is shown in Figure 3.
Fig. 3. The structure of the collagen-specific carbohydrate Glc-Gal-Hyl in peptide linkage. The carbohydrate unit may also consist only of the Gal portion (Modified from Kivirikko & Myllylä 1979 with permission of Academic Press).
The carbohydrate units are transferred to collagen as posttranslational modifications catalyzed by two specific enzymes; hydroxylysyl galactosyltransferase (EC 2.4.1.50, GT), transfers galactose to hydroxylysyl residues, and then galactosylhydroxylysyl glucosyltransferase (EC 2.4.1.66, GGT), transfers glucose to galactosylhydroxylysyl residues. Both enzymes have been demonstrated to exist in a number of tissues (Kivirikko & Myllylä 1979). The extent of glycosylation varies between collagen types and within

30
the same collagen type in different tissues and at different ages (Risteli 1977, Anttinen et al. 1977b, Kivirikko & Myllylä 1979, Kivirikko & Myllylä 1982, Kielty et al. 1993).
2.4.1 Purification and molecular properties of collagen
glycosyltransferases
2.4.1.1 Hydroxylysyl galactosyltransferase (GT)
Partial purification of this enzyme has been achieved from guinea-pig skin, rat kidney cortex, and human platelets (Kivirikko & Myllylä 1979, Kivirikko & Myllylä 1982). From chick embryo extract, a method, consisting of six conventional protein purification steps yielded about a 50- to 150-fold increase in specific activity (Risteli et al. 1976a). GT has not yet been isolated as a homogeneous protein, the highest degree of purification, up to about 1000-fold, having been obtained from chick embryo extract by using a procedure consisting of ammonium sulfate fractionation, affinity chromatography on collagen-agarose, and gel filtration (Risteli et al. 1976b). The major problem in the enzyme purification is its marked tendency to be inactivated during purification and in many cases the specific activity decreases during steps in which the purity of the enzyme protein clearly increases (Risteli et al. 1976a, Risteli 1978). The molecular weight of GT is not known as no homogeneous protein has been isolated and the gene or cDNA for the enzyme has not yet been cloned. The activity of the partially purified enzyme is found by gel filtration to fall into two major peaks with molecular weights of about 450 and 200 kDa, and one minor species with a molecular weight of about 50 kDa (Risteli et al. 1976a). It is not known whether they represent an aggregate of the enzyme alone or the enzyme with some other proteins. GT is a glycoprotein as its activity can be inhibited by concanavalin A, and the inhibition can be reduced by methyl α-D-mannoside, methyl α-D-glucoside, mannose, fructose, and glucose (Risteli 1978, Kivirikko & Myllylä 1979). The enzyme binds to concanavalin A coupled to agarose and can be eluted with a buffer containing methyl glucoside and ethylene glycol (Risteli 1978).
2.4.1.2 Galactosylhydroxylysyl glucosyltransferase (GGT)
Partial purification of GGT has been achieved from guinea-pig skin, rat kidney cortex, chick embryo cartilage, bovine arterial tissue, human fetal tissues, plasma and platelets (Kivirikko & Myllylä 1979, Kivirikko & Myllylä 1982). Purifications of over 2000-fold from whole chick embryos and about 160-fold from chick cartilage have also been reported using six conventional protein purification steps (Myllylä et al. 1976). GGT has been isolated as a homogeneous protein from chick embryos by two affinity column

31
methods. The first procedure includes ammonium sulfate fractionation, collagen-agarose and UDPglucose-derivative-agarose affinity chromatographies, and two gel filtrations (Myllylä et al. 1977). The second one consists of ammonium sulfate fractionation, concanavalin A-agarose, collagen-agarose, and UDPglucose-derivative-agarose affinity chromatographies, and one gel filtration (Anttinen et al. 1978). The molecular weight of GGT from chick embryos is about 72 – 78 kDa seen in SDS-PAGE, and it may consist of only one polypeptide chain (Myllylä et al. 1977). Gel filtration gives a lower and variable molecular weight, probably due to partially adsorption of the molecule to the columns (Anttinen et al. 1977a, Myllylä et al. 1977). Amino acid analysis indicates that the chick GGT is rich in Glu, Gln, Asp, Asn, Gly, and Ala (Anttinen et al. 1978). It is a glycoprotein having a high affinity for concanavalin A-agarose, and this affinity can be reduced by methyl α-D-mannoside (Anttinen et al. 1977a). The chick GGT is capable of generating strong hydrophobic interaction, probably being part of the explanations for adsorption to gel filtration columns and a marked tendency to form aggregates when concentrated (Anttinen et al. 1977a). Antibody against the pure chick embryo GGT inhibits the transferase activity from different tissues, and gives a single precipitation line of identity with the enzyme from many chick embryo tissues in immunodiffusion (Myllylä 1981). However, the gene or cDNA of GGT has not been cloned so far.
2.4.2 Catalytic properties of collagen glycosyltransferases
GT catalyzes the synthesis of galactosylhydroxylysine by transferring galactose from UDP-galactose to hydroxylysyl residues in peptide linkages. GGT catalyzes the formation of glucosylgalactosylhydroxylysine by transferring the glucose from UDP-glucose to galactosylhydroxylysyl residues (see Figure 4). Both enzymes require a bivalent cation, preferably Mn2+ (Kivirikko & Myllylä 1979). The free ε-amino group of the hydroxylysyl residue and a nonhelical polypeptide conformation are the absolute requirements for GT and GGT, as the N-acetylation or deamination of the free ε-amino group completely inhibits both reactions, and the triple helical conformation of the substrate prevents the interactions with both transferases (Kivirikko & Myllylä 1984, Kielty et al. 1993). The glycosylation reactions are also affected by the amino acid sequence of the peptide, and the peptide chain length (Kivirikko & Myllylä 1979, Kivirikko & Myllylä 1984). The number of -X-Hyl-Gly- sequence in a polypeptide chain play a key role in the overall glycosylation (Anttinen & Hulkko 1980) whereas the amino acid sequences adjacent to the hydroxylysyl residue may be a minor factor. Longer peptides are more effective substrates for the transferases (Kivirikko & Myllylä 1979, Kivirikko & Myllylä 1984). The specificity of GT and GGT for their glycosyl acceptors is very high. GT only attaches galactose to hydroxylysyl residues in peptide linkage, and does not attach a second galactose to galactosylhydroxylysyl residues or a galactose to glucosylgalactosylhydroxylysyl residues. Free hydroxylysine does not act as the sugar acceptor. However free hydroxylysine in high concentration and free

32
galactosylhydroxylysine can inhibit the glycosylation reactions (Kivirikko & Myllylä 1979, Kivirikko & Myllylä 1982). GGT does not attach glucose to a number of carbohydrates, glycopeptides or glycoproteins, in which the galactose is linked to other sugars or to amino acids other than hydroxylysine. This enzyme also fails to attach glucose to hydroxylysyl residues or glucosylgalactosylhydroxylysyl residues. Galactosylsphingosine is one exception in that it can act in vitro as a good glucosyl acceptor for GGT, probably due to its structural similarity to galactosylhydroxylysine (Kivirikko & Myllylä 1979, Kivirikko & Myllylä 1982).
Fig. 4. Glycosylation reactions catalyzed by peptidyl GT and GGT (Modified from Kivirikko & Myllylä 1984 with permission of Elsevier Science).
The preferential glycosyl donor for GT or GGT is the corresponding UDP glycoside. The Km of UDP-galactose for GT is about 20-30 µM whereas the Km of UDP-glucose for GGT is about 5-30 µM depending on the source from which the enzyme was isolated. The reaction product, UDP, and several other nucleotides (UTP, UMP, ATP, ADP, AMP, CDP etc.) act as inhibitors of both enzymes (Kivirikko & Myllylä 1979, Kivirikko & Myllylä 1984).

33
The pH optimum for both glycosyltransferases is about 7-7.4. The optimal Mn2+ concentration is 0.2-2 mM for partially purified GGT, and this enzyme binds at least two Mn2+ having the Kd values of 3-5 µM (site I) and 50-70 µM (site II) (Myllylä et al. 1979). The Kd value for Mn2+ at site II is probably much higher than the physiological concentration. It is not known whether this site has any significance in vivo (Kivirikko & Myllylä 1984). Fe2+(Kd 5-7 µM) and Co2+ (Kd 30 µM) are the only other metals that can activate GGT at low concentration. They can, however, inhibit the enzyme when the concentration is high (Myllylä et al. 1979). Several bivalent cations are inhibitors of the Mn2+-activated hydroxylysyl glycosyltransferase (Kivirikko & Myllylä 1982). There are no corresponding data available for GT. The mechanism (shown in figure 5) of the GGT reaction has been investigated in detail with the enzyme purified from chick embryos by analyzing the initial velocity and inhibition kinetics (Myllylä 1976). The data agree with an ordered binding of Mn2+, UDP-glucose, and collagen to GGT, and an ordered release of the glycosylated collagen, UDP, and Mn2+, in which Mn2+ does not need to leave the enzyme between the catalytic cycles (Myllylä 1976). Some kinetic studies have also been carried out on GGT from human platelets (Smith et al. 1977). The data are consistent with the scheme described for chick embryo GGT. No detailed kinetic data is available for GT (Kivirikko & Myllylä 1979, Kivirikko & Myllylä 1982, Kivirikko & Myllylä 1984).
Fig. 5. Schematic representation of the mechanism proposed for the GGT reaction. At high Mn2+ concentrations, GGT probably binds two Mn2+ ions successively before binding to UDP-glucose. Due to the high Kd value at site II, this situation may not occur in vivo and therefore is not shown. Mn2+ need not to leave the enzyme between the catalytic cycles. GGT: galactosylhydroxylysyl glucosyltransferase; UDPglc: UDP-glucose; Glc-pept: glucosylated peptide (Modified from Kivirikko & Myllylä 1984 with permission of Elsevier Science).
2.4.3 Intracellular sites of collagen glycosylations
It was proposed earlier based on work using isolated Hela cell membrane fractions that the glycosyltransferases were located only in the plasma membrane, and that the glycosylations occurred immediately before secretion of procollagen from the cells (Hagopian et al. 1968, Bosmann 1969). The assays used, however, were nonspecific, and more evidence accumulated later that suggested the glycosylations take place in a different intracellular location (Kivirikko & Myllylä 1979). The activities of GT and GGT in most cases were found to be associated with the rough ER (Kivirikko & Myllylä

34
1979), but it was also found to be present in the smooth ER (Harwood et al. 1975b). It has been shown by Blumenkrantz and his coworkers that the distribution of both glycosyltransferases within purified chick embryo bone microsomes is similar to that of LH. About 70-80% of their activities are intramembranous with the remainder intracisternal. The common location of the major portion of LH, GT and GGT activities suggested that they might form a multienzyme complex to preferentially modify certain lysyl residues in nascent procollagen chains as they pass across the membrane of the ER (Blumenkrantz et al. 1984). It has been also demonstrated by Bortolato et al. in the early 90’s that the GGT specific to collagen is located in the rough ER, smooth ER, and Golgi apparatus in the chick embryo liver (Bortolato et al. 1990, Bortolato et al. 1991, Bortolato et al. 1992). Brownell & Veis (1975) and Harwood et al. (1975b) found galactosylhydroxylysyl and glucosylgalactosylhydroxylysyl residues in nascent polypeptide chains, suggesting that glycosylations are initiated while the polypeptide chains are still under assembly on the ribosomes. Oikarinen et al. (1976a) studied the time course of the glycosylation of hydroxylysyl residues in chick embryo cartilage cells, and demonstrated that there was no lag between the hydroxylation of lysyl residues and the glycosylation of hydroxylysyl residues. They also found that after a 5-minute pulse-label with 14C-lysine, the reactions continue for about 10 minutes during the chase period in the chick embryo tendon cells (Oikarinen et al. 1976b) and about 20 minutes in the cartilage cells (Oikarinen et al. 1976a). The triple helix is formed in both cell types at about the same time points (Harwood et al. 1975a), thus suggesting that the glycosylations continue until triple helix formation of the pro-α chains. This conclusion was also reached in the studies, which showed that if the triple helix formation is inhibited, the glycosylations are prolonged, and if it is accelerated, the extent of modification is less (Oikarinen et al. 1976b, Oikarinen et al. 1977). These results agree with the data on hydroxylysyl galactosylation (Risteli et al. 1976a) and galactosylhydroxylysyl glucosylation (Myllylä et al. 1975) in vitro, as the two enzymes do not catalyze reactions with triple helical substrates. The triple helices of procollagen form before they move from the cisternae of the rough ER to the Golgi apparatus, indicating that the hydroxylysyl glycosylations are completed within the cisternae of rough ER (Kivirikko & Myllylä 1979, see Figure 6).

35
Fig. 6. Schematic representation of the events in collagen biosynthesis that occur within the cisternae of the rough ER. The propeptide is removed during the translocation across the membrane and hydroxylation of lysyl residues and glycosylation of hydroxylysyl residues are initiated while the polypeptide chains are still being assembled on the ribosomes. The reactions continue after the synthesis of complete pro α-chains until the triple helix is formed. LH: lysyl hydroxylase; GT: hydroxylysyl galactosyltransferase; GGT: galactosylhydroxylysyl glucosyltransferase; Gal: galactose; Glu: glucose (Modified from Kivirikko & Myllylä 1984 with permission of Elsevier Science).
2.4.4 Collagen glycosyltransferase activities in physiological,
pathological, and experimental conditions
Considerable differences are found in hydroxylysyl glycosyltransferase activities between different tissues, and in the same tissue under different conditions. Both were found to vary with age, being higher in embryonic than in adult tissues (Anttinen et al. 1977b, Kovanen & Suominen 1989). The developmental patterns of both glycosyltransferase activities were also studied in chick embryos (Risteli 1977). GT and GGT increased up to the 16th day and decreased thereafter, the highest activities being found in the leg tendons of the 16-day-old chick embryos, and the activities in cartilage were higher than in skin or skull (Risteli 1977). During cartilage and bone formation induced by demineralized bone matrix in rats, both activities rose dramatically from day one to reach the highest values on day nine and dropped down slowly afterwards (Myllylä et al. 1981). PH-4, GGT and other enzymes in human serum are often regarded as biomarkers of collagen biosynthesis. It has been reported that prolonged or concentric exercise causes an increase in the activity of GGT in human serum. The most likely explanation is that the prolonged heavy exercise results in protein leakage from muscles and probably from the collagen-synthesizing cells of the connective tissue (Takala et al. 1986, Takala et al. 1989, Virtanen et al. 1993).

36
It has been shown under experimental conditions that GT and GGT activities increase in lung of hamsters with bleomycin-induced pulmonary fibrosis (Bolarin et al. 1984a), in liver of CF1 female mice with hepatic murine Schistosomiasis mansoni (Bolarin et al. 1985), and in rat liver with carbon tetrachloride-induced fibrosis (Bolarin et al. 1987) whereas both activities decrease in transformed human and chick-embryo cells (Myllylä et al. 1981), and in isolated chick-embryo tendon cells after the administration of cortisol acetate to the chick embryo (Oikarinen 1977). The GGT activity increases in many pathological conditions, for example dermatological disorders (Kuutti-Savolainen 1979, Kuutti-Savolainen & Kero 1979, Oikarinen et al. 1982a, Oikarinen et al. 1982b, Ala-Kokko et al. 1987), human breast cancer (Bolarin 1983a), human primary hepatocellular carcinoma (Bolarin 1983b), acute viral hepatitis, cirrhotic liver (Bolarin et al. 1984b) and experimentally-induced primary liver carcinoma (Bolarin 1991). The level of GGT is higher than normal in acute myocardial infarction and during subsequent collagen scar formation (Anttinen et al. 1981). GGT activity significantly increases in injured porcine intervertebral discs (Kääpä et al. 1994) or degenerated discs (Kääpä et al. 1995). Increased serum or tissue GGT activity has been also found in fibrosing lung diseases (Anttinen et al. 1985, Anttinen et al. 1986, Poole et al. 1985, Poole et al. 1989), various rheumatic diseases (Myllylä et al. 1989), neuromuscular disorders (Myllylä et al. 1982), and even the adult respiratory distress syndrome (Farjanel et al. 1993). The increase of GGT in these diseases might result from the accelerated collagen biosynthesis or collagen metabolism. Savolainen et al. (1981) have shown that members of a family with dominant epidermolysis bullosa simplex have a deficiency of GGT. The enzyme activity was low in serum, skin tissue, and cultured skin fibroblasts whereas other intracellular enzymes of collagen biosynthesis showed no abnormality. No molecular biological data is available from the patients.

3 Aims of the present work
Many data suggest that LH may have collagen type-specific or tissue-specific isoforms. Considerable differences are found in the extent of lysyl hydroxylation between different collagen types, and even within the same collagen type from different tissues. Lysines in collagen types II, IV, and V are hydroxylated normally in Ehlers-Danlos syndrome type VI patients (Ihme et al. 1984) and the residual lysyl hydroxylase activity of EDS VI cells seems to preferentially hydroxylate lysines in type IV collagen (Risteli et al. 1980). Some data, however, still argue against the existence of collagen type-specific isoenzymes. No differences are observed between the molecular and catalytic features of lysyl hydroxylase purified from various sources (Turpeenniemi-Hujanen et al. 1980, Turpeenniemi-Hujanen et al. 1981, Myllylä et al. 1988) and the catalytic properties of lysyl hydroxylase in crude extracts from cells synthesizing different collagen types (Puistola 1982). It has also been shown that there is a relative excess of lysyl hydroxylase activity in chick-embryo tendon and cartilage compared with skin and bone, implying that the differences in the extent of lysyl hydroxylation could result from differential regulation of a single lysyl hydroxylase (Puistola & Anttinen 1982). Immunological identification of chick lysyl hydroxylase does not support the existence of collagen type-specific or tissue-specific isoforms (Turpeenniemi-Hujanen 1981). During the past few years, two new isoenzymes (LH2 and LH3) for lysyl hydroxylase have been identified from human and mouse tissues, and the previous known isoform is now correspondingly termed LH1. LH2 and LH3 are highly expressed in certain tissues. These tissue-specific isoenzymes may show differences with respect to hydroxylation of various collagen types. However the biological significance of the presence of lysyl hydroxylase isoforms remains unknown. The first aim of the study was 1. To study the mRNA expression patterns and the collagen type specificities of
different LH isoforms in various human cells. It was demonstrated that all LH isoforms lack collagen type specificity and the mRNA levels of LH1, LH2, and the α-subunit of prolyl 4-hydroxylase correlate significantly with each other whereas no correlation was observed between mRNA levels of LH3 and those of the others. As lysyl hydroxylase and glycosyltransferases are the enzymes

38
required for catalyzing three consecutive reactions within the ER in building the unique carbohydrate structures in collagens, we started to test the glycosyltransferase activities with all LH isoforms. Surprisingly, LH3 was found to be the only isoform possessing glycosyltransferase activities. The following more specific goals were then set: 2. To characterize the multifunctionality of human LH3 and C. elegans LH, and to
study the intracellular distribution of human LH3. 3. To identify the amino acids important for collagen glycosyltransferase activities
associated with human LH3.

4 Materials and Methods
More detailed descriptions are present in the original articles I-IV.
4.1 Cell culture and transfection (I, II)
The following human cell lines were used in the study: human skin fibroblasts (K-3), HT-1080 fibrosarcoma cells (ATCC CCL-121), MG-63 osteosarcoma cells (ATCC CRL-1427), CCD-19LU cells from lung (ECACC 90112708), HepG2 heptoblastoma cells (ATCC HB-8065), Hela epitheloid carcinoma cells (ATCC CCL2), JAR-727 cells (ATCC HTB144), pancreas adenocarcinoma cells (ATCC CRL-1687), and ACHN renal adenocarcinoma cells (ECACC 88100508). Cells above were cultured in MEM, DMEM, RPMI (GIBCO BRL) respectively, supplemented with 10% fetal calf serum, 50 µg/ml ascorbate, 100 µg/ml streptomycin and 100 U/ml penicillin. Cells were incubated on tissue culture petri plates at 37°C with 5% CO2 in air atmosphere until confluent, washed with 1× PBS and stored at -70°C until used. Cos-7 cells were grown at 37°C / 5% CO2 in DMEM containing 10% newborn calf serum. 8×105 cells per 100 mm dish or 1×105 cells per 35 mm dish were plated one day before transfection. Cells were transfected by plasmid DNA using FUGENE6 (Boehringer Mannheim). After incubation for 0-48 hours, the cells were harvested and washed with PBS.
4.2 RNA isolation and Northern blot analysis (I)
Total RNA was isolated from cells by Trizol (GIBCO BRL) reagent or using a guanidine isothiocyanate method (Sambrook et al. 1989, Ausubel et al. 1989). The probes used in Northern analysis were listed in Materials and Methods in publication I. The RNA was

40
fractionated in a 0.8% agarose gel containing 0.22 M formaldehyde at 30V overnight, transferred to a nylon membrane and fixed by UV cross-linking. The filter was hybridized at 68 °C for 2 hours with a radioactively labeled probe in ExpressHyb solution (Clontech), washed first with 2 × SSC, 0.05% SDS for 40 min at room temperature and then with 0.1 × SSC, 0.1% SDS for 40 min at 50°C. The probe was removed from the blot by incubating with 0.5% SDS in H2O at 90°C for 10 min. The filter was then re-equilibrated in the ExpressHyb solution and reprobed with a new sequence. The hybridization signals were quantitated by scanning the autoradiograms with a laser densitometer, PhosphoImager (Molecular Dynamics), and normalized to the quantity of β-actin mRNA and the specific activity of different probes.
4.3 Production of antibodies (I, II)
Polyclonal antibodies against human LH2 and LH3 were produced in chickens. The sequence of antigen corresponds to amino acids 284-464 for LH2 (Valtavaara et al. 1997), and amino acids 283-297 for LH3 (Valtavaara et al. 1998). The LH2 antigen was produced as a GST fusion protein in E. coli, purified by Glutathione sepharose 4B matrix, and injected into chickens together with Freund’s complete adjuvant (50% V/V). The injection was repeated 3 times with 2 weeks intervals with incomplete adjuvant. The LH3 antigen was a synthetic peptide coupled via a cysteine residue to keyhole limpet hemocyanin.
4.4 Microscopical study (II)
Cos-7 cells, transfected with the pEGFP-N1-human LH3 plasmid DNA, were grown on coverslips. Forty-eight hours after transfection, the cells were washed with PBS and briefly rinsed in distilled water. The coverslips were mounted on slides using Immuno-mount (Shandon). The expression of the LH3-GFP fusion protein was visualized directly by fluorescence microscopy (Nikon). In order to see the subcellular localization of LH3-GFP protein and compare with protein disulfide isomerase (PDI), the transfected cells were fixed with 4% paraformaldehyde in PBS, pH 7.4 for 20 min, blocked for 1 hour in 0.05% saponin, 0.1% bovine serum albumin in PBS pH 7.4 (IF buffer), followed by a 1-hour incubation at room temperature with a monoclonal antibody (1:100) against PDI (DAKO). Coverslips were washed 3 times with IF buffer and incubated for a further 1-hour at room temperature with a 1:100 dilution of anti-mouse IgG tetramethylrhodamine B isothiocyanate (TRITC) conjugate. After 3 washes with PBS and one brief rinse in distilled water, the coverslips were mounted on slides with Immuno-mount, and the double staining was observed by fluorescence microscopy.

41
4.5 Immunoprecipitation (II)
Cos-7 cells transfected with the pEGFP-N1-human LH3 construct or the vector alone were lysed with 5 mM EDTA, 0.5% Triton X-100 in 1 × PBS for 30 min at +4°C. The crude cell lysates were pretreated with protein A-Sepharose CL-4B (Amersham Pharmacia Biotech) to remove non-specific binding and then immunoprecipitated with a monoclonal anti-GFP antibody (Clontech). The precipitated proteins were separated by SDS-PAGE and detected by immunoblotting.
4.6 Immunoblotting (I, II, III, IV)
The proteins separated by SDS-PAGE were electroblotted onto Immobilon-P transfer membrane (Millipore). Detection of the proteins was carried out first by incubating with the primary antibody, and then with the secondary antibody coupled to horseradish peroxidase. The recognized epitopes were detected by an enhanced chemiluminescence detection system (Amersham Pharmacia Biotech).
4.7 In vitro translation (II, III, IV)
The in vitro translation was performed with the single tube protein system 3 kit, STP 3 (Novagen). For transcription, 200 ng of plasmid were mixed with 3.2 µl of transcription mix in a total volume of 4 µl and incubated at 30°C for 15 min. For translation, 1.0 µl of [35S] methionine (10 mCi/ml, Amersham Pharmacia Biotech) or 0.8 µl of cold methionine in the kit and 12 µl of translation mix were added to the transcription reaction and incubated at 30°C for 1 hour. The products were analyzed by SDS-PAGE and autoradiography. GGT activity was directly measured from the reaction mixture containing unlabeled methionine.
4.8 Site-directed in vitro mutagenesis (II, III, IV)
The QuickChange site-directed mutagenesis kit (Stratagene) was used to introduce site-specific point mutations into cDNA. PCR reactions were performed using a supercoiled, double stranded DNA as a template with two synthetic oligonucleotide primers containing the desired mutation encoding a mutated amino acid residue in the protein molecule. PfuTurbo DNA polymerase was used to replicate both DNA strands with high fidelity.

42
4.9 Expression and purification of the recombinant LH3 proteins (II,
III, IV)
4.9.1 Constructs
Commercially available baculovirus transfer vector pFastBacI in the BAC-TO-BACTM expression system (Life Technologies, Inc.) was modified to contain the human LH1 signal peptide (Hautala et al. 1992), a His6 tag, and a BamHI restriction site for insertion of the desired cDNA. The inserts cover the nucleotides from 255 to 2405 for LH1 (Hautala et al. 1992), 76-2267 for both LH2a and LH2b, in which the LH2b construct also contains the alternatively spliced exon sequence between nucleotides 1500 and 1501 (Valtavaara et al. 1997, Valtavaara 1999, Yeowell & Walker 1999a), and the nucleotides from 289 to 2455 for LH3 (Valtavaara et al. 1998). LH1, LH2a, LH2b, LH3 coding sequences without signal peptides were also cleaved with BamHI/HindIII from pFastBacI constructs (Valtavaara 1999) and subcloned into the same sites in the E. coli expression vector pQE30 (Qiagen) containing a His6 tag at the amino terminus. The insert for C. elegans LH covering the amino acids 14-730 (Wixon et al. 2000) was generated by PCR and ligated into the SacI/PstI site of the pQE30 vector. Constructs were transformed into the E. coli XL1-Blue strain. Human LH3 cDNA coding for the amino acid 25-738 (Valtavaara et al. 1998) and C. elegans LH cDNA coding for the amino acid 17-730 (Wixon et al. 2000) were subcloned at NdeI/BamHI sites and NdeI/XhoI sites, respectively, into a pET-15b expression vector (Novagen). Both constructs contain a His6 tag at the amino terminus and were transformed into E. coli BL21(DE3)pLysS cells.
4.9.2 Protein expression
4.9.2.1 Human LH isoforms in insect cells
Sf9 (Spodoptera frugiperda) insect cells were grown in Sf-900 II serum-free medium (Life Technologies, Inc.). Expression of recombinant protein was carried out by the baculovirus transfer vector pFastBacI in the BAC-TO-BACTM expression system (Life Technologies, Inc.). The cells were harvested 0-72 hours after infection and homogenized according to protocol described for human lysyl hydroxylase (Pirskanen et al. 1996).

43
4.9.2.2 Human LH isoforms and C. elegans LH with pQE30 vector in E.coli XL1-Blue cells
E. coli XL1-Blue cells transformed with the human LH1, LH2a/2b, LH3-pQE30 or C. elegans LH-pQE30 constructs, respectively, were grown in LB medium in the presence of ampicillin (100 µg/ml) to an OD600 of 0.6-0.7 at 37°C with vigorous shaking. Protein expression was induced by addition of 1 mM IPTG to the cultures and incubation at 30°C for 5 hours. The cells were washed, harvested and stored at +4°C. The cells were suspended at room temperature for 30 minutes in lysis buffer containing 0.4 M NaCl, 0.5% Nonidet P-40 and 20 mM Tris-HCl, pH 7.8 at 20°C in the presence of 50 µg/ml lysozyme, 10 µg/ml DNase I and RNase A. Lysis was completed by sonication and the soluble proteins were collected by centrifugation.
4.9.2.3 Human LH3 and C. elegans LH with pET-15b vector in E. coli BL21(DE3) pLysS cells
E. coli BL21(DE3)pLysS cells transformed with the human LH3-pET-15b or C. elegans LH-pET-15b construct were grown in M9ZB-medium in the presence of ampicillin (100 µg/ml) and chloramphenicol (34 µg/ml) to an OD600 of 0.7-0.9 at 25 °C. Protein expression was induced by addition of 0.3 mM IPTG and incubation at 18°C overnight, after which the cells were washed, harvested and stored at +4°C. The cells were lysed in lysis buffer containing 50 mM Tris-HCl, pH 8.4 at +4°C, 5% glycerol, 0.5% triton X-100, 1 mM DTT, in the presence of one proteinase inhibitor cocktail tablet, 200 µg/ml lysozyme, 20 µg/ml DNase I and RNase A. Lysis was completed by freezing the cells at -70°C over an hour and thawing at 30°C for 40 minutes. The soluble proteins were collected by centrifugation.
4.9.3 Purification of His-tag protein by Nickel-NTA-Agarose
Recombinant proteins containing a His-tag at the N-terminus were purified on a Nickel-NTA agarose column (Qiagen). For the LH-pFastBacI constructs, the homogenate was mixed with Ni-NTA agarose equilibrated with 20 mM Tris-HCl, pH 7.8, 0.3 M NaCl, 5% glycerol, and 10 mM imidazole at +4°C for 45 min. Washing was carried out in a buffer containing 20 mM imidazole, and elution in a buffer containing 100 mM, 200 mM, and 300 mM imidazole, respectively. For the LH-pQE30 constructs, the agarose was mixed with the soluble fraction of the E. coli homogenate and incubated for 45 min at +4°C with a solution containing 20 mM Tris-HCl, pH 7.8, 0.4 M NaCl, 1% glycerol, and 5 mM imidazole. Elution was carried out in an elution buffer containing 100 mM imidazole. For the LH-pET-15b constructs, the Nickel-NTA beads were mixed with the cleared cell lysates for 2 hours at +4°C in a buffer containing 50 mM Tris-HCl, pH 8.4, 5% glycerol,

44
0.5% Triton X-100, 1 mM DTT and 5 mM imidazole. Elution was performed with the elution buffer containing 250 mM imidazole.
4.9.4 UDP-glucuronic acid affinity chromatography
UDP-glucuronic acid affinity matrix (Uridine 5’-diphosphoglucuronic acid immobilized on cross-linked 4% beaded agarose, Sigma) was equilibrated in the lysis buffer containing 50 mM Tris-HCl, pH 8.4 at +4°C, 5% glycerol, 0.5% Triton X-100 and 1 mM DTT. The fractions eluted from the nickel column were mixed with the matrix for 2 hours at +4°C. The affinity matrix was washed 6 times with 2ml of buffer containing 0.3 M NaCl, 10 mM MnCl2 in the lysis buffer. The elution was carried out by 1 ml of buffer containing 50% ethylene glycol, 0.5 M NaCl, 5 mM UDP-glucuronic acid sodium salt, 10 mM MnCl2, 0.1 M citrate buffer, pH 6.4, by mixing the matrix with the buffer overnight at +4°C. A PD-10 column (Pharmacia) was used to remove UDP-glucuronic acid, and the purified human LH3 protein was concentrated by Centricon® Centrifugal Filter Devices YM-10 (Millipore) column.
4.10 GT and GGT activity measurements (II, III, IV)
The activities were measured by a method based on the transfer of radioactively-labeled (tritium) sugar from a UDP-sugar to hydroxylysyl or galactosylhydroxylysyl residues in a calf skin gelatin substrate, respectively. The amount of UDP-sugar in the reaction was 66 µM and the specific activity 139 Ci/mol. The specific detection of the reaction products (dpm counting of ion exchange chromatography elution) was performed after alkaline hydrolysis.
4.11 Software (I, III)
The correlation coefficients analysis was carried out by using SPSS 8.0 for Windows. The ClustalX program was used to align 11 amino acid sequences of the LHs from 7 species (see Figure 1 in publication III).

5 Results
5.1 Messenger RNA expression levels of LH isoforms and major
collagen types I, III, IV, V in different human cell lines (I)
Total RNAs isolated from confluent cultures of nine human cell lines were first hybridized with cDNA probes for different LH isoforms. The LH1 probe hybridized with a 3.2 kb mRNA, LH2 with a 4.2 kb mRNA, and LH3 with a 2.8 kb mRNA. Three lysyl hydroxylase isoenzymes are present in all cells tested. LH1 is mainly expressed in normal skin fibroblasts, HepG2, and MG-63 cells. LH2 is expressed at very high levels in HepG2, MG-63, and HT-1080 cells. The amount of LH3 is lower than that of the other isoforms in all cell lines, the highest levels being in HT-1080 cells and normal skin fibroblasts. The immunoblotting results with antibodies against LH2 and LH3 roughly reflect the data obtained from the hybridization analysis. The mRNA levels of the main collagen types I, III, IV, V and the α subunit of prolyl 4-hydroxylase were measured in the same cells. The probe for type I collagen, α1(I), hybridized with mRNAs of 4.7 kb and 6.0 kb in MG-63, normal skin fibroblasts, and CCD-19LU cells. Hybridization with an α1(III) probe revealed mRNAs of 5.6 kb and 4.6 kb in MG-63 cells, and weak signals in normal skin fibroblasts and CCD-19LU cells. Hybridization with an α2(IV) probe revealed one mRNA band of 6.5 kb in HT-1080, ACHN, Hela, JAR-727, CCD-19LU, and MG-63 cells. A probe for α1(V) cDNA hybridized with three mRNAs of 8.3 kb, 7.2 kb, and 6.5 kb, which were mainly observed in Hela, and MG-63 cells. The probe for the α subunit of prolyl 4-hydroxylase hybridized with one mRNA of about 3 kb in size in all cells, and higher mRNA levels were detected in HepG2, MG-63, and HT-1080 cells. After quantitation of the hybridization signals, expression levels of the LH isoforms were compared with those of prolyl 4-hydroxylase, another post-translational enzyme of collagen biosynthesis, and with those of collagens. Correlation coefficient analysis showed that the enzyme mRNAs did not correlate with collagen mRNAs, and that the mRNA levels of LH1, LH2 and the α subunit of prolyl 4-hydroxylase significantly

46
correlated with each other whereas no correlation was observed between mRNA levels of LH3 and those of LH1, LH2, and the α subunit of prolyl 4-hydroxylase.
5.2 Characterization of the multifunctionality of human LH3 and C.
elegans LH (II, III, IV)
5.2.1 Human LH3 is a multifunctional protein possessing GGT activity
(II)
Sf9 insect cells and Cos-7 cells were transfected with human LH3 cDNA constructs. The GGT activity measurements were carried out 72 hours after transfection in Sf9 cells and 48 hours in Cos-7 cells. The GGT activity was about 2.0 × 106 dpm/plate in Sf9 cell lysates and 1.2 × 106 dpm/plate in medium over the background. One plate corresponded to about 10 × 106 cells. The activity in mammalian expression system was about 2 × 105 dpm/plate in Cos-7 cell lysates and 1.1 × 106 dpm/plate in medium over the background. One plate corresponded to about 1.4 × 106 cells. LH3 was expressed in Sf9 cells as a protein with a His-tag at the amino terminus. Sixty percent of the GGT activity was bound to the nickel column and all activity could be eluted from the column by imidazole. The eluate was analyzed by SDS-PAGE and immunostained by anti-His-tag antibodies on a Western blot. One major band was present corresponding to the molecular weight of about 85 kDa. A cell-free translation system was also used to study whether the GGT activity was linked to human LH3. Only one protein was found when human LH3 cDNA was added to the incubation mixture. The molecular weight corresponded again to that of LH3. The GGT activity was about 34,000 dpm/translation mixture supplemented with LH3 cDNA. Human LH3 peptide antibodies and antibodies against highly purified chicken GGT were used to study whether the antibody binding inhibits the catalytic activity of the GGT produced by the His-tagged LH3 cDNA construct in Sf9 cells. Both antibodies partially inhibited the GGT activity whereas no inhibition was observed when tested with His-tag antibody and antibodies against human Dpb11 protein used as a control. The other lysyl hydroxylase isoforms, human LH1, LH2a, and LH2b, were expressed in Sf9 cells under identical conditions to those used for LH3. Although LH activity was present in all cell lysates, none of these recombinant proteins had detectable GGT activity.

47
5.2.2 Human LH3 is the only isoenzyme possessing GT activity (IV)
A baculovirus system was first used to express all human LH isoforms and test for GT activity. There was a reasonably high endogeneous GT activity in Sf9 cells, about 80,000 dpm/mg. LH3 cDNA expression generated additional GT activity, varying from 50,000 to 170,000 dpm/mg, over the background in different experiments. No additional GT activity over background was found when human LH1, LH2a, and LH2b were expressed in insect cells. The LH3 protein was also produced in an in vitro translation system. A small but reproducible amount of GT activity over the background was detected (550 dpm/translation mixture), but only after adding dog pancreas microsomes to the incubation mixture. Two LH3 cDNA constructs were prepared in E. coli cells. The pQE30 vector and XL1-Blue strain were first used to produce a His-tagged LH3 protein. After Nickel-NTA purification, both GT and GGT activities were bound to the nickel column. Elution was carried out by 100 mM imidazole. Analysis of the nickel-column beads and the eluates by immunoblotting using His-tag antibody revealed one major protein band with a molecular weight of 81 kDa, corresponding in size to full length LH3. Activity measurements showed a good recovery of GGT and a lower recovery of GT in the elution fractions. In order to confirm that the full length LH3 is responsible for both activities, the pET-15b expression vector and BL21(DE3) pLysS cells were then used to produce recombinant protein on a large scale. Pure LH3 protein was achieved with a yield of about 1 mg soluble LH3 in a 200 ml E. coli cell culture after optimizing the expression conditions. The His-tagged recombinant LH3 protein was first purified by a nickel column and eluted by 250 mM imidazole. Further purification by UDP-glucuronic acid column chromatography revealed a pure LH3 on SDS-PAGE. The activity measurements demonstrated that the purified LH3 possessed not only LH, GGT, but also GT activities, with the specific activity of 1.5 × 105 dpm/mg, 1.2 × 109 dpm/mg and 2.72 × 107 dpm/mg respectively. The Km value of the purified LH3 was 35 µM for UDP-galactose and and 17 µM for UDP-glucose. One milligram of the pure recombinant LH3 was able to transfer 1.6 µmoles of galactose and 19.7 µmoles of glucose, respectively, from corresponding UDP-sugar in one hour at 37°C in the glycosylation reactions when the activity values were corrected to a saturating concentration of citrate soluble collagen. These values are similar to those reported earlier (Myllylä et al. 1976, Kivirikko & Myllylä 1979). Similarly, one milligram of the pure LH3 with LH activity was able to decarboxylate 0.8 µmoles of 2-oxoglutarate, the decarboxylation linked to hydroxylation of lysyl residues, in the reaction when the activity value was corrected to saturating substrate and cosubstrate concentrations. The human LH1, LH2a, and LH2b isoforms were expressed in E. coli cells as well. Immunostaining by His-tag antibodies indicated the presence of the LH isoforms in the supernatant fraction. However, no GT or GGT activity was obtained from any of these recombinant proteins, showing again that LH3 is the only isoform with glycosyltransferase activities among the human LH isoforms. Very low LH activity was detected in the crude cell extracts of all these LH isoforms in bacteria system.

48
5.2.3 GT and GGT activities are associated with C. elegans LH (III, IV)
There is only one gene for lysyl hydroxylase in the C. elegans genome. Two cDNA constructs, C. elegans LH-pQE30 in E. coli XL1-Blue cells and C. elegans LH-pET-15b in E. coli BL21(DE3)pLysS cells, were prepared in order to express the protein in bacterial cells and to determine whether the LH gene product possesses the GT and GGT activities. Control E. coli XL1-Blue cells, containing only the pQE30 vector, as well as cells expressing DHFR have only very low residual GT or GGT activity, less than 300 dpm/mg soluble protein. When C. elegans LH cDNA was expressed, both GT and GGT activities were present in the supernatant. Due to different expression levels, the activities varied in different experiments. The C. elegans LH-pET-15b construct transformed in E. coli BL21(DE3)pLysS cells was also used to express the recombinant protein. The His-tagged protein was first partially purified on a Ni-NTA column and then both GT and GGT activities were analyzed. The enzyme activity assays showed GT activity in the recombinant C. elegans LH, 2.84 × 104 dpm/mg, the corresponding value for GGT was 4.76 × 106 dpm/mg. A cell-free translation system was used as well to test if GT and GGT activities are associated with the C. elegans LH gene product. Only one protein corresponding to full length LH was synthesized and GGT activity was present in the translation mixture. GT activity was not detectable. However, the addition of dog pancreas microsomes to the incubation mixture revealed a small but reproducible amount of GT activity over background in the reaction mixture (2,300 dpm/translation mixture, the corresponding value for GGT was 23,800 dpm/translation).
5.3 The intracellular distribution of human LH3 (II)
A cDNA construct for the human LH3-GFP fusion protein was prepared, in which GFP was inserted into the carboxy-terminal end of human LH3. Cos-7 cells were transfected with the construct and produced a fluorescent protein that can be visualized directly by fluorescence microscopy. Forty eight hours after transfection most of the fusion protein was located in the ER whereas a small fraction of the fluorescent protein entered the Golgi complex, in agreement with the findings that a part of the expressed protein was secreted due to overexpression. GGT activity measurement was carried out with the cells collected at forty eight hours after transfection, and showed a 2-3-fold increase in the activity, suggesting the correct three-demensional conformation of the human LH3-GFP fusion protein for catalysis in the enzyme reaction. This result that LH3 is retained in the ER in most cells is similar to the data reported earlier for LH1 (Kellokumpu et al. 1994), and is in good agreement with the hydroxylation and glycosylation sites in collagen biosynthesis (Kivirikko & Myllylä 1979, Kivirikko & Pihlajaniemi 1998)

49
5.4 Identification of the amino acids important for catalytic activities
of GT and GGT associated with human LH3 and C. elegans LH (III,
IV)
5.4.1 Active sites of GGT in human LH3 and C. elegans LH (III)
Sequence alignment of LH isoforms from bovine, rat, chicken, human, mouse, C. elegans indicated that there are 29 amino acids conserved between the LH3 and C. elegans sequences (LH3-C. elegans specific amino acids), but differing from those of LH1 and LH2 (see figure 1 in publication III). These amino acids are scattered evenly throughout the molecule. In vitro mutagenesis combined with a cell-free translation system was first used to study the importance of these amino acids for GGT activity. The LH3-C. elegans specific amino acids were mutated in the C. elegans LH sequence. The most inhibitory changes and one as a control from carboxy-terminal area were tested further with human LH3. The data was then confirmed with human LH3 in an E. coli system. The LH3-C. elegans specific amino acids were mutated to the ones present in LH1 and/or LH2. Mutations at the amino-terminal end of the molecule caused an inhibition of GGT activity, whereas mutations at the carboxy-terminal end of the molecule had no effect. Two mutations, the Cys144Ile and Leu208Ile in the human LH3 sequence, and Cys132Ile and Leu196Ile in C. elegans LH sequence, inhibited GGT activity markedly in both species. Some differences exist between human and C. elegans. For example, the Leu124Val and Leu125Val changes in the C. elegans LH inhibited GGT activity whereas the corresponding changes in human LH3 increased the activity. Furthermore, a mutation of Ala453Ile in the C. elegans LH caused a remarkable inhibition of GGT activity whereas the corresponding changes in human LH3 had no effect on the activity. The most effective mutations of human LH3, i.e. Cys144Ile and Leu208Ile, were studied further in a baculovirus expression system to determine whether these changes have an effect on LH activity. Similar inhibitory results were obtained for GGT activity in the mutated molecules whereas no inhibitory effects on LH activity were observed. The human Cys144 was investigated further to determine if this cysteine is disulfide-linked by using an alkylating agent, iodoacetamide, to block free sulfhydryl groups in the molecule. The His-tagged human LH3 and three Cys mutants, Cys144Ile, Cys494Ser, and Cys577Ser, were expressed in E. coli XL1-Blue cells. The alkylated proteins purified on a nickel column were then analyzed under non-reducing and reducing condition by SDS-PAGE combined with Western blotting. Based on the mobility shift on the gel, the results suggest that Cys144 is not disulfide-linked whereas Cys494 and Cys577 seem to be disulfide bonded amino acids in human LH3. Alkylation of Cys144 does not reduce GGT activity. The truncated molecules of human LH3 were generated in order to determine if the carboxy-terminal part of human LH3 is required for GGT activity. The amino acids from the C-terminal portion were removed by creating a translation stop codon in different part of human LH3. The data indicate that a 355 amino acid long portion of the N-terminal moiety is still able to generate a low GGT activity whereas none of the truncated molecules were able to hydroxylate lysyl residues.

50
There are at least three DxD like motifs in the human LH3 sequence. In vitro mutagenesis result and the data from the shortened molecules indicate that the DxD motifs at positions 392-394 and 489-491 are not important for GGT activity. Mutations of the short conserved motif containing five aspartate residues at position 187-191 eliminated GGT activity. Mutation analyses were used to determine whether LH1 and/or LH2 would possess GGT activity if some of LH3-C. elegans specific amino acids were introduced into their structure. Single Cys, single Leu, or Cys plus Leu were introduced respectively into LH1, or LH2a, or LH2b molecules (corresponding to the change Cys144Ile or Cys144Leu between LH3 and LH1/LH2, and Leu208Ile between LH3 and LH1/LH2). The mutated proteins were expressed in E. coli XL1-Blue cells and the activity measurements showed that no GGT activity was regained after these mutations.
5.4.2 Active sites of GT in human LH3 (IV)
The same strategy was utilized to search for the amino acids critical for catalytic activity of GT associated with human LH3. The LH3-C. elegans specific amino acids were mutated to the ones present in LH1 and/or LH2. Potential DxD motifs were also mutated by in vitro mutagenesis. The LH3 and its mutants with pET-15b vector were expressed in E. coli BL21(DE3)pLysS cells and GT activity was measured from partially purified enzyme eluates after nickel purification. GGT activity was measured for comparison. The data indicate that the mutations of Cys144Ile, and aspartates at position 187-191 reduced the GT activity remarkably whereas the mutations at carboxy-terminal end of the molecule and the DxD motif at position 392-394 had no effect on GT activity. The Leu208Ile mutation inhibited GT activity slightly (see figure 2 in publication IV). Similar results were obtained in GGT activity measurements as well.

6 Discussion
6.1 Lack of collagen type specificity for lysyl hydroxylase isoforms (I)
The mRNA expression levels of three LH isoforms (LH1, LH2, and LH3) have been studied in several different human cell lines. They were present in all human cells tested. There is a five-fold variation in the LH1 mRNA levels and a 20 to 30-fold variation in LH2 mRNA levels, whereas the variation for LH3 is only two-fold among different human cells. LH1 is the major isoform in skin fibroblasts, pancreas adenocarcinoma cells, and placenta trophoblastic tumor cells, being about 50% of the total lysyl hydroxylases. This explains the finding that the hydroxylysine content of collagens is most noticeably reduced in the skin when tissue samples are analyzed from patients with EDS VI, a disease caused by a LH1 gene defect (Hyland et al. 1992, Hautala et al. 1993, Ha et al. 1994, Heikkinen et al. 1997, Yeowell & Walker 1997, Brinckmann et al. 1998, Pajunen et al. 1998, Pousi et al. 1998, Heikkinen et al. 1999, Walker et al. 1999, Yeowell & Walker 1999b, Pousi et al. 2000, Yeowell et al. 2000a, Yeowell et al. 2000b). LH2 is the major form in fibrosarcoma, osteosarcoma, and hepatoblastoma cells, constituting about 60% of total LH in those cells. LH3 is a major form in kidney adenocarcinoma cells, about half of the LH within these cells being LH3. Todate no heritable disorders have been reported associated with a gene defect in LH2 and/or LH3. The hydroxylysine content varies in the triple-helical regions of different collagen types. Type III and type I collagens contain only 6-10 hydroxylysyl residues per polypeptide chain, whereas type V and type IV contain a three- to five-fold higher amount of hydroxylysine (Kivirikko et al. 1992, Kielty et al. 1993). The initial suggestion for the existence of collagen type-specific LH isoforms was based on the findings that lysines in collagen type II, IV, V were hydroxylated normally in EDS VI patients (Ihme et al. 1984), and that the residual LH activity of EDS VI cells appears to preferentially hydroxylate lysines in type IV collagen (Risteli et al. 1980). The mRNA levels of LH isoforms and the α subunit of prolyl 4-hydroxylase, another enzyme involved in collagen biosynthesis were compared with the mRNAs of the most abundant collagen types (I, III, IV, V). The data do not indicate any significant correlation between individual collagen types and LH isoforms or the α subunit of prolyl 4-hydroxylase, arguing against collagen type

52
specificity in the enzyme reactions. The amino acid sequences surrounding the individual lysyl residues, rather than collagen type, are probably more important for determining which lysyl hydroxylase isoform catalyzes the hydroxylation reaction. Prolyl 4-hydroxylase activity has been widely used to estimate the rate of total collagen synthesis in many experimental models and in patients with various diseases (Kivirikko et al. 1992, Kielty et al. 1993, Prockop & Kivirikko 1995). The mRNA levels of LH1, LH2, and the α subunit of prolyl 4-hydroxylase are significantly correlated with each other in various human cell lines, suggesting that LH1 and LH2, together with prolyl 4-hydroxylase are coregulated with total collagen synthesis. It is remarkable that the mRNA levels of LH3 do not correlate with those of LH1, LH2, and the α subunit of prolyl 4-hydroxylase, implying a difference in their substrates and in the regulation of their gene expression. According to the phylogenetic analysis, LH3 is the oldest form and the others result from more recent duplication events (Ruotsalainen et al. 1999). It is highly possible that LH3 acts on other types of sequences in vivo in addition to collagenous sequences.
6.2 Characterization of GT and GGT activities associated with
human LH3 and C. elegans LH (II, III, IV)
6.2.1 Human LH3 is a multifunctional protein possessing LH, GT and
GGT activities (II, IV)
Both LH and GGT activities were observed when human LH3 cDNA was expressed in baculovirus and mammalian expression systems. The activities appeared simultaneously without any lag, suggesting the expression of human LH3 resulted in the simultaneous induction of the GGT activity. GT activity was also found when human LH3 was expressed in insect cells although the endogenous GT activity was higher. Purification of the recombinant protein in insect cells and in E. coli cells with nickel column chromatography demonstrated that the GT and GGT activities bound to the column and could be eluted by imidazole, indicating that the GT or GGT activity is present on a protein having a His-tag in its structure. Results from in vitro translation experiments further support the finding that both GT and GGT activities are associated with human LH3, as only one protein, corresponding in size to the full length LH3, is translated in the mixture supplemented with LH3 cDNA, and this protein has glycosyltransferase activities. Immunoblot analysis of the eluates by using anti-His-tag antibody demonstrated the recombinant protein with molecular weight of about 85 kDa in a baculovirus system and about 81 kDa in an E. coli system, corresponding to the full size of human LH3. The lower molecular weight in the E. coli system is probably due to a deficiency of glycosylation in bacterial cells. GGT has been reported earlier to be purified from chick embryos, having a molecular weight of 78 kDa (Myllylä et al. 1976) whereas GT has never been isolated as a homogenous protein due to a marked tendency to lose activity

53
(Kivirikko & Myllylä 1979). It is not clear whether there are more than one GT and GGT in vivo, and whether the molecular weight of the enzymes differs in different species. It should be noticed that the glycosyltransferase activities with LH3 correspond to the activity levels of the enzymes isolated or highly purified from chick embryos (Kivirikko & Myllylä 1979), and the Km values for UDP-sugars are similar as reported earlier to the enzymes of chick embryos (Kivirikko & Myllylä 1979). Human LH3 polyclonal antibodies were raised against a synthetic peptide corresponding to the amino acids 283-297 of human LH3 sequence (Valtavaara et al. 1998), a region that is highly dissimilar between LH isoforms. GGT antibodies were prepared earlier against chick glucosyltransferase isolated as a homogenous protein from chick embryos (Myllylä 1981). Both LH3 and GGT antibodies were tested with the protein produced by human His-tagged LH3 cDNA in insect cells and with the GGT protein in human fibroblasts. Similar inhibitions of GGT activities were observed in both cases. The result provides evidence for similar behavior of fibroblastic and LH3 derived GGT. Furthermore, GGT antibodies behaved similarly in binding to chicken (Myllylä 1981) and human GGT. A trifunctional character for a protein is a remarkable feature. It is found in enzymes involved in lipid metabolism, as well as in purine and pyrimidine metabolism (Kan et al. 1993, Iwahana et al. 1996, Eaton et al. 2000). Protein disulfide isomerase is also an example of a multifunctional protein. By association with different proteins it acts as a chaperone, as a protein thiol oxidoreductase and as a subunit for prolyl 4-hydroxylase and the microsomal triglyceride transfer protein (Noiva 1999). It should be noted that the activity measurements of GT and GGT in vitro do not necessarily correspond to the full activity in vivo. It is known that many post-translational modifications, for instance phosphorylation and glycosylation, are defective in E. coli cells. Therefore the recombinant LH3 protein produced in bacterial system may not represent the enzyme with maximal activities. It has been reported that treatment of LH1 with endoglycosidase H (Myllylä et al. 1988) and mutation of the glycosylated asparagines of LH1 (Pirskanen et al. 1996) reduced the enzyme activity, indicating the N-glycosylation is probably important for LH activity. Other factors may affect the activities measured in vitro. This is seen for example by the addition of microsomes in cell-free translation system. A small, reproducible GT activity was detectable only when dog pancreas microsomes were added to the incubation mixture, suggesting additional factor(s) present in microsomes increases GT activity.
6.2.2 C. elegans LH is multifunctional possessing also
glycosyltransferase activities (III, IV)
The genome of C. elegans, a primitive eukaryotic organism, was the first animal genome to be completely sequenced (Wixon et al. 2000). It contains only about 19,000 genes, and is widely used in studies concerning gene expression and gene silencing. C. elegans has only one gene for LH, which is an ortholog with the vertebrate LH isoforms, LH1, LH2, and LH3 (Norman & Moerman 2000, Wixon et al. 2000). All three activities,

54
hydroxylation-galactosylation-glucosylation, which are associated with human LH3 as indicated above, are also associated with the C. elegans LH gene. The phylogenetic data have shown that LH3 is more closely related to C. elegans LH than the other isoforms (Ruotsalainen et al. 1999), suggesting that LH1 and LH2 have lost the glycosylating functions during evolution although they have still retained the LH activity. It is not known whether they have acquired any other functions during evolution.
6.3 Identification of the amino acids important for the
glycosyltransferase activities associated with human LH3 and C.
elegans LH (III, IV)
6.3.1 Active sites of GGT in human LH3 and C. elegans LH (III)
The data from site-directed mutagenesis combined with in vitro translation and E. coli expression system revealed that the active sites of GGT are located in the amino-terminal parts of the molecule in C. elegans LH and human LH3. There are evolutionary changes between C. elegans LH and human LH3, the active sites are not identical in these species although GT and GGT activities are present in both molecules. C. elegans LH seems to be more sensitive to the LH3-C. elegans specific amino acid mutations, as more inhibitory changes were found at the amino-terminal part of the molecule for C. elegans LH, and the inhibitions were more effective compared with that of human LH3. Two mutations, the Cys144Ile and Leu208Ile in human LH3 sequence, decreased GGT activity dramatically but had no effect on LH activity. The shortening of the carboxy-terminal portion of LH3 eliminates LH activity, but only reduces GGT activity. These findings provide further evidence that LH and GGT active sites are separated on the LH3 molecule. This is consistent with the earlier data obtained from LH1 that the amino acids responsible for LH activity are located in the carboxy-terminal portion of the molecule (Pirskanen et al. 1996, Passoja et al. 1998a), the most conserved region among LH isoforms (Hautala et al. 1992, Valtavaara et al. 1997, Passoja et al. 1998b, Valtavaara et al. 1998, Ruotsalainen et al. 1999). This result is also confirmed by a recent study in which LH3 was produced as a secreted form in baculovirus system (Rautavuomo et al. 2002). Our data indicated that the Cys144 is not disulfide linked whereas the Cys494 and the Cys577 used as controls are disulfide bonded. Furthermore our data suggest that although the Cys144 is required for the functional integrity of GGT, it probably does not participate directly in the enzyme catalysis. Sequence alignments and X-ray crystal structures have indicated a so-called DxD motif in many glycosyltransferases, and it has been found in different glycosyltransferase families (Ünligil et al. 2000). This motif is thought to stabilize the Mn2+ and thus indirectly stabilize the binding of the diphosphate moiety of the UDP-sugar (Ünligil et al. 2000, Gastinel et al. 2001). At least three DxD motifs exist in the LH3 molecule. Only the mutations of the most amino-terminal DxD motif, which consists of five aspartates at

55
position 187-191, eliminated GGT activity, indicating that this region is important for GGT activity and might act as a Mn2+ binding site in the molecule. However, this needs to be verified by crystallization of LH3 protein. The replacement of the two most conserved amino acids among the LH3-C. elegans specific amino acids, the Cys144 and Leu208, in the LH1 and LH2 structures is not sufficient to restore GGT activity to these isoforms, suggesting that LH1 and LH2 have diverged so much during evolution from the ancestral LH3 (Ruotsalainen et al. 1999) that single amino acid changes are not sufficient to restore glycosyltransferase activity.
6.3.2 Active sites of GT in human LH3 (IV)
The search for GT active sites was carried out by in vitro mutagenesis combined with protein expression in E. coli BL21(DE3)pLysS cells with the pET-15b vector. The data indicate that the mutations of Cys144Ile, and aspartates at position 187-191 reduced the GT activity remarkably whereas the mutations at the carboxy-terminal end of the molecule and the DxD motif at position 392-394 had no effect on GT activity. The results thus suggest that the amino acids located at the amino-terminal part of the molecule, important for GGT activity, are also required for GT activity. The data indicate similar results for GT and GGT. More in vitro mutagenesis studies are needed to see whether GT active site is identical to that of GGT.

7 Conclusions
The collagen type specificity of the LH isoforms in different human cell lines was first investigated in this study. The mRNA expression levels of the LH isoforms do not correlate with those of any major individual collagen type (I, III, IV, V), indicating a lack of collagen type specificity among the LH isoforms. The mRNA expression level of LH1, LH2 and the α-subunit of prolyl 4-hydroxylase showed a high correlation with each other whereas LH3 mRNA did not show any correlation to the mRNA levels of LH1, LH2 or the α-subunit of prolyl 4-hydroxylase. This suggests that LH3 may act on a different substrate in vivo. More studies are required to make clear the difference in substrate specificity of the LH isoforms. It was also demonstrated in this study that the full length LH3 and C. elegans LH are multifunctional proteins, possessing LH, GT and GGT activities, which are able to catalyze three consecutive steps in building the unique hydroxylysine linked carbohydrate units in collagens. It reveals for the first time the cDNA sequence of GT and GGT, which enables us to search for the gene mutations in abnormal glycosylation-related diseases, although no heritable disorders related to this gene have been reported so far. Subcellular localization of human LH3 was also studied in this work. It is located mainly in the ER whereas some is found in the Golgi complex probably due to overexpression, being similar to human LH1. This is in good agreement with the known intracellular sites of hydroxylation and glycosylation of lysyl residues in collagen biosynthesis. No corresponding data are available for human lysyl hydroxylase isoform 2 (LH2a and LH2b). The functions of human LH2a and LH2b remain to be elucidated. The amino acids important for the catalytic activity of glycosyltransferases associated with human LH3 have been shown in this study to be localized at the amino-terminal part of the molecule, separate from the active site of LH. This enables us to design a knock out or transgenic study to manipulate LH and glycosyltransferase activities separately in mice to see if these activities associated with LH3 have the functions in vivo. Furthermore, the studies may lead to the elucidation of the functions of the glycosylation of hydroxylysyl residues in vivo. The His-tagged constructs of the LH isoforms were expressed in E. coli or insect cells. Purification was carried out easily and conveniently by Nickel-NTA-Agarose chromatography. UDP-glucuronic acid affinity chromatography resulted in pure

57
recombinant LH3/C. elegans LH protein. This approach will facilitate crystallization of human LH3 and C. elegans LH to study in detail the molecular structure of these multifunctional proteins.

8 References
Aho S, Rothenberger K & Uitto J (1999) Human p120ctn catenin: tissue-specific expression of isoforms and molecular interactions with BP180/type XVII collagen. J Cell Biochem 73: 390-399.
Aho S & Uitto J (1999) 180-kD bullous pemphigoid antigen/type XVII collagen: tissue-specific expression and molecular interactions with keratin 18. J Cell Biochem 72: 356-367.
Ala-Kokko L, Rintala A & Savolainen ER (1987) Collagen gene expression in keloids: analysis of collagen metabolism and type I, III, IV, and V procollagen mRNAs in keloid tissue and keloid fibroblast cultures. J Invest Dermatol 89: 238-244.
Ananthanarayanan VS, Saint-Jean A & Jiang P (1992) Conformation of a synthetic hexapeptide substrate of collagen lysyl hydroxylase. Arch Biochem Biophys 298: 21-28.
Andrews PC, Hawke D, Shively JE & Dixon JE (1984) Anglerfish preprosomatostatin II is processed to somatostatin-28 and contains hydroxylysine at residue 23. J Biol Chem 259: 15021-15024.
Anttinen H & Hulkko A (1980) Regulation of the glycosylations of collagen hydroxylysine in chick embryo tendon and cartilage cells. Biochim Biophys Acta 632: 417-427.
Anttinen H, Järvensivu PM & Savolainen ER (1981) Serum galactosylhydroxylysyl glucosyltransferase in acute myocardial infarction and during subsequent collagen scar formation. Eur J Clin Invest 11: 375-379.
Anttinen H, Myllylä R & Kivirikko KI (1977a) Hydrophobic and carbohydrate-recognition chromatographies of collagen glucosyltransferase. Eur J Biochem 78: 11-17.
Anttinen H, Myllylä R & Kivirikko KI (1978) Further characterization of galactosylhydroxylysyl glucosyltransferase from chick embryos. Amino acid composition and acceptor specificity. Biochem J 175: 737-742.
Anttinen H, Oikarinen A & Kivirikko KI (1977b) Age-related changes in human skin collagen galactosyltransferase and collagen glucosyltransferase activities. Clin Chim Acta 76: 95-101.
Anttinen H, Terho EO, Järvensivu PM & Savolainen ER (1985) Elevated serum galactosylhydroxylysyl glucosyltransferase, a collagen synthesis marker, in fibrosing lung diseases. Clin Chim Acta 148: 3-8.
Anttinen H, Terho EO, Myllylä R & Savolainen ER (1986) Two serum markers of collagen biosynthesis as possible indicators of irreversible pulmonary impairment in farmer's lung. Am Rev Respir Dis 133: 88-93.
Armstrong LC & Last JA (1995) Rat lysyl hydroxylase: molecular cloning, mRNA distribution and expression in a baculovirus system. Biochim Biophys Acta 1264: 93-102.
Ausubel FM, Brent R, Kingston RE, Moore DD, Smith JA, Seidman JG & Struhl K (1989) Current protocols in molecular biology. John Wiley & Sons, New York.
Ayad S, Boot-Handford RP, Humphries MJ, Kadler KE & Shuttleworth CA (1998) The extracellular matrix FactsBook, second edition. Academic Press, London.

59
Bank RA, Robins SP, Wijmenga C, Breslau-Siderius LJ, Bardoel AF, van der Sluijs HA, Pruijs HE & TeKoppele JM (1999) Defective collagen crosslinking in bone, but not in ligament or cartilage, in Bruck syndrome: indications for a bone-specific telopeptide lysyl hydroxylase on chromosome 17. Proc Natl Acad Sci U S A 96: 1054-1058.
Bank RA, Tekoppele JM, Janus GJ, Wassen MH, Pruijs HE, Van der Sluijs HA & Sakkers RJ (2000) Pyridinium cross-links in bone of patients with osteogenesis imperfecta: evidence of a normal intrafibrillar collagen packing. J Bone Miner Res 15: 1330-1336.
Bateman JF, Lamande SR & Ramshaw JAM (1996) In: Comper WD (ed) Extracellular Matrix. Harwood Academic Publishers GmbH, Amsterdam, p22-67.
Beck K & Brodsky B (1998) The collagen triple helix and the alpha-helical coiled coil. J Struct Biol 122: 17-29.
Berg AH, Combs TP, Du X, Brownlee M and Scherer PE (2001) The adipocyte-secreted protein Acrp30 enhances hepatic insulin action. Nat Med 7: 947-953.
Blumenkrantz N, Assad R & Peterkofsky B (1984) Characterization of collagen hydroxylysyl glycosyltransferases as mainly intramembranous microsomal enzymes. J Biol Chem 259: 854-859.
Bolarin DM (1983a) Immunoreactive prolyl hydroxylase protein and galactosyl-hydroxylysyl glucosyltransferase activity in breast tissues and sera of patients with primary breast cancer. Res Commun Chem Pathol Pharmacol 39: 493-502.
Bolarin DM (1983b) Elevation of liver-galactosylhydroxylysyl glucosyltransferase activity in human primary hepatocellular carcinoma. Int J Biochem 15: 1291-1293.
Bolarin DM (1991) Liver collagen glucosyltransferase in experimental primary liver carcinoma. Acta Physiol Hung 77: 113-120.
Bolarin DM, Barker K & Fuller GC (1985) Serum and liver enzymes of collagen synthesis in hepatic murine schistosomiasis mansoni. Trans R Soc Trop Med Hyg 79: 826-830.
Bolarin DM, Barker K & Fuller GC (1987) Enzyme markers of collagen synthesis in carbon tetrachloride-induced fibrosis and during colchicine modification of CCl4-induced liver injury. Exp Mol Pathol 46: 145-152.
Bolarin DM, Palicharla P & Fuller GC (1984a) Enzymes of collagen synthesis in lung tissues of bleomycin-induced pulmonary fibrosis. Toxicol Appl Pharmacol 73: 188-191.
Bolarin DM, Savolainen ER and Kivirikko KI (1984b) Three serum markers of collagen biosynthesis in Nigerians with cirrhosis and various infectious diseases. Eur J Clin Invest 14: 90-95.
Bosmann HB (1969) Collagen-galactosyl transferase: subcellular localization and distribution in fibroblasts transformed by oncogenic viruses. Life Sci 8: 737-746.
Bortolato M, Azzar G, Farjanel J & Got R (1990) Evidence of partial localization in Golgi apparatus of UDP-glucose collagen glucosyltransferase from chick embryo liver. Int J Biochem 22: 1039-1046.
Bortolato M, Azzar G, Farjanel J & Got R (1991) Solubilization of UDP-glucose collagen glucosyltransferase activities from chick embryo liver: Golgi apparatus, smooth and rough endoplasmic reticulum. Int J Biochem 23: 897-900.
Bortolato M, Radisson J, Azzar G & Got R (1992) Studies on the mechanism of different collagen glucosyltransferase reactions (Golgi apparatus, smooth endoplasmic reticulum, rough endoplasmic reticulum) in chick embryo liver. Int J Biochem 24: 243-248.
Brenner RE, Vetter U, Nerlich A, Worsdorfer O, Teller WM & Müller PK (1989) Osteogenesis imperfecta: insufficient collagen synthesis in early childhood as evidenced by analysis of compact bone and fibroblast cultures. Eur J Clin Invest 19: 159-166.
Brinckmann J, Acil Y, Feshchenko S, Katzer E, Brenner R, Kulozik A & Kugler S (1998) Ehlers-Danlos syndrome type VI: lysyl hydroxylase deficiency due to a novel point mutation (W
612C).
Arch Dermatol Res 290: 181-186. Brinckmann J, Notbohm H, Tronnier M, Acil Y, Fietzek PP, Schmeller W, Müller PK & Batge B
(1999) Overhydroxylation of lysyl residues is the initial step for altered collagen cross-links and fibril architecture in fibrotic skin. J Invest Dermatol 113: 617-621.
Brownell AG & Veis A (1975) The intracellular location of the glycosylation of hydroxylysine of collagen. Biochem Biophys Res Commun 63: 371-377.

60
Byers PH (1994) Ehlers-Danlos syndrome: recent advances and current understanding of the clinical and genetic heterogeneity. J Invest Dermatol 103: 47S-52S.
Chamson A, Berbis P, Fabre JF, Privat Y & Frey J (1987) Collagen biosynthesis and Isomorphism in a case of Ehlers-Danlos syndrome type VI. Arch Dermatol Res 279: 303-307.
Chung CH, Au LC & Huang TF (1999) Molecular cloning and sequence analysis of aggretin, a collagen-like platelet aggregation inducer. Biochem Biophys Res Commun 263: 723-727.
Eaton S, Bursby T, Middleton B, Pourfarzam M, Mills K, Johnson AW & Bartlett K (2000) The mitochondrial trifunctional protein: centre of a beta-oxidation metabolon? Biochem Soc Trans 28: 177-182.
Eklund L, Piuhola J, Komulainen J, Sormunen R, Ongvarrasopone C, Fassler R, Muona A, Ilves M, Ruskoaho H, Takala TE & Pihlajaniemi T (2001) Lack of type XV collagen causes a skeletal myopathy and cardiovascular defects in mice. Proc Natl Acad Sci U S A 98: 1194-1199.
Eyre D (1987) Collagen cross-linking amino acids. Methods Enzymol 144: 115-139. Ezer S, Bayés M, Elomaa O, Schlessinger D & Kere J (1999) Ectodysplasin is a collagenous
trimeric type II membrane protein with a tumor necrosis factor-like domain and colocalizes with cytoskeletal structures at lateral and apical surfaces of cells. Hum Mol Genet 8: 2079-2086.
Farjanel J, Hartmann DJ, Guidet B, Luquel L & Offenstadt G (1993) Four markers of collagen metabolism as possible indicators of disease in the adult respiratory distress syndrome. Am Rev Respir Dis 147: 1091-1099.
Fruebis J, Tsao TS, Javorschi S, Ebbets-Reed D, Erickson MR, Yen FT, Bihain BE & Lodish HF (2001) Proteolytic cleavage product of 30-kDa adipocyte complement-related protein increases fatty acid oxidation in muscle and causes weight loss in mice. Proc Natl Acad Sci U S A 98: 2005-2010.
Fukai N, Eklund L, Marneros AG, Oh SP, Keene DR, Tamarkin L, Niemela M, Ilves M, Li E, Pihlajaniemi T & Olsen BR (2002) Lack of collagen XVIII/endostatin results in eye abnormalities. EMBO J 21: 1535-1544.
Gastinel LN, Bignon C, Misra AK, Hindsgaul O, Shaper JH & Joziasse DH (2001) Bovine alpha1, 3-galactosyltransferase catalytic domain structure and its relationship with ABO histo-blood group and glycosphingolipid glycosyltransferases. EMBO J 20: 638-649.
Giudice GJ, Squiquera HL, Elias PM & Diaz LA (1991) Identification of two collagen domains within the bullous pemphigoid autoantigen, BP180. J Clin Invest 87: 734-738.
Gordon MK, Fitch JM, Foley JW, Gerecke DR, Linsenmayer C, Birk DE & Linsenmayer TF (1997) Type XVII collagen (BP 180) in the developing avian cornea. Invest Ophthalmol Vis Sci 38: 153-166.
Ha VT, Marshall MK, Elsas LJ, Pinnell SR & Yeowell HN (1994) A patient with Ehlers-Danlos syndrome type VI is a compound heterozygote for mutations in the lysyl hydroxylase gene. J Clin Invest 93: 1716-1721.
Hagopian A, Bosmann HB & Eylar EH (1968) Glycoprotein biosynthesis: the localization of polypeptidyl: N-acetylgalactosaminyl, collagen: glucosyl, and glycoprotein: galactosyl transferases in HeLa cell membrane fractions. Arch Biochem Biophys 128: 387-396.
Harwood R, Bhalla AK, Grant ME & Jackson DS (1975a) The synthesis and secretion of cartilage procollagen. Biochem J 148: 129-138.
Harwood R, Grant ME & Jackson DS (1975b) Studies on the glycosylation of hydroxylysine residues during collagen biosynthesis and the subcellular localization of collagen galactosyltransferase and collagen glucosyltransferase in tendon and cartilage cells. Biochem J 152: 291-302.
Hausmann E (1967) Cofactor requirements for the enzymatic hydroxylation of lysine in a polypeptide precursor of collagen. Biochim Biophys Acta 133: 591-593.
Hautala T, Byers MG, Eddy RL, Shows TB, Kivirikko KI & Myllylä R (1992) Cloning of human lysyl hydroxylase: Complete cDNA-derived amino acid sequence and assignment of the gene (PLOD) to chromosome 1p36.3-36.2. Genomics 13: 62-69.
Hautala T, Heikkinen J, Kivirikko KI & Myllylä R (1993) A large duplication in the gene for lysyl hydroxylase accounts for the type VI variant of Ehlers-Danlos syndrome in two siblings. Genomics 15: 399-404.

61
Heikkinen J, Hautala T, Kivirikko KI & Myllylä R (1994) Structure and expression of the human lysyl hydroxylase gene (PLOD): introns 9 and 16 contain Alu sequences at the sites of recombination in Ehlers-Danlos syndrome type VI patients. Genomics 24: 464-471.
Heikkinen J, Toppinen T, Yeowell H, Krieg T, Steinmann B, Kivirikko KI & Myllylä R (1997) Duplication of seven exons in the lysyl hydroxylase gene is associated with longer forms of a repetitive sequence within the gene and is a common cause for the type VI variant of Ehlers-Danlos syndrome. Am J Hum Genet 60: 48-56.
Heikkinen J, Pousi B, Pope M & Myllylä R (1999) A null-mutated lysyl hydroxylase gene in a compound heterozygote British patient with Ehlers-Danlos syndrome type VI. Hum Mutat 14: 351.
Hyland J, Ala-Kokko L, Royce P, Steinmann B, Kivirikko KI & Myllylä R (1992) A homozygous stop codon in the lysyl hydroxylase gene in two siblings with Ehlers-Danlos syndrome type VI. Nat Genet 2: 228-231.
Hägg P, Väisänen T, Tuomisto A, Rehn M, Tu H, Huhtala P, Eskelinen S & Pihlajaniemi T (2001) Type XIII collagen: a novel cell adhesion component present in a range of cell-matrix adhesions and in the intercalated discs between cardiac muscle cells. Matrix Biol 19: 727-742.
Ihme A, Krieg T, Nerlich A, Feldmann U, Rauterberg J, Glanville RW, Edel G & Müller PK (1984) Ehlers-Danlos syndrome type VI: collagen type specificity of defective lysyl hydroxylation in various tissues. J Invest Dermatol 83: 161-165.
Ihme A, Risteli L, Krieg T, Risteli J, Feldmann U, Kruse K & Müller PK (1983) Biochemical characterization of variants of the Ehlers-Danlos syndrome type VI. Eur J Clin Invest 13: 357-362.
Iwahana H, Fujimura M, Ii S, Kondo M, Moritani M, Takahashi Y, Yamaoka T, Yoshimoto K & Itakura M (1996) Molecular cloning of a human cDNA encoding a trifunctional enzyme of carbamoyl-phosphate synthetase-aspartate transcarbamoylase-dihydroorotase in de Novo pyrimidine synthesis. Biochem Biophys Res Commun 219: 249-255.
Jiang P & Ananthanarayanan VS (1991) Conformational requirement for lysine hydroxylation in collagen. Structural studies on synthetic peptide substrates of lysyl hydroxylase. J Biol Chem 266: 22960-22967.
Judisch GF, Waziri M & Krachmer JH (1976) Ocular Ehlers-Danlos syndrome with normal lysyl hydroxylase activity. Arch Ophthalmol 94: 1489-1491.
Kan JL, Jannatipour M, Taylor SM & Moran RG (1993) Mouse cDNAs encoding a trifunctional protein of de novo purine synthesis and a related single-domain glycinamide ribonucleotide synthetase. Gene 137: 195-202.
Kellokumpu S, Sormunen R, Heikkinen J & Myllylä R (1994) Lysyl hydroxylase, a collagen processing enzyme, exemplifies a novel class of luminally-oriented peripheral membrane proteins in the endoplasmic reticulum. J Biol Chem 269: 30524-30529.
Kielty CM, Hopkinson I & Grant ME (1993) Collagen: structure, assembly and organization in the extracellular matrix. In: Royce PM and Steinmann B (eds) Connective tissue and its heritable disorders. Molecular, genetics and medical aspects. Wiley-Liss, Inc., New York, p103-147.
Kikuchi Y, Suzuki Y & Tamiya N (1983) The source of oxygen in the reaction catalysed by collagen lysyl hydroxylase. Biochem J 213: 507-512.
Kivirikko KI (1993) Collagens and their abnormalities in a wide spectrum of diseases. Ann Med 25: 113-126.
Kivirikko KI, Bright HJ & Prockop DJ (1968) Kinetic patterns of protocollagen hydroxylase and further studies on the polypeptide substrate. Biochim Biophys Acta 151: 558-567.
Kivirikko KI & Myllylä R (1979) Collagen glycosyltransferases. In: Hall DA & Jackson DS (eds) Int Rev Connect Tissue Res. Academic Press, Inc., 8: 23-72.
Kivirikko KI & Myllylä R (1980) The hydroxylation of prolyl and lysyl residues. In: Freedman RB & Hawkins HC (eds) The enzymology of post-translational modifications of protein. Academic Press, London 1: 53-104.
Kivirikko KI & Myllylä R (1982) Post-translational enzymes in the biosynthesis of collagen: Intracellular enzymes. Methods Enzymol 82: 245-304.
Kivirikko KI & Myllylä R (1984) Biosynthesis of the collagens. In: Piez KA & Reddi AH (eds) Extracellular matrix biochemistry. Elsevier, New York, p 83-118.

62
Kivirikko KI, Myllylä R & Pihlajaniemi T (1992) Hydroxylation of proline and lysine residues in collagens and other animal and plant proteins. In: Harding JJ and Grabbe MJC (eds) Post-translational modifications of proteins. CRC Press, Boca Raton, FL, p 1-51.
Kivirikko KI & Pihlajaniemi T (1998) Collagen hydroxylase and the protein disulfide isomerase subunit of prolyl 4-hydroxylases. Adv Enzymol Related Areas Mol Biol 72: 325-398.
Kivirikko KI & Prockop DJ (1967) Hydroxylation of proline in synthetic polypeptides with purified protocollagen hydroxylase. J Biol Chem 242: 4007-4012.
Kivirikko KI & Prockop DJ (1972) Partial purification and characterization of protocollagen lysine hydroxylase from chick embryos. Biochim Biophys Acta 258: 366-379.
Koch M, Foley JE, Hahn R, Zhou P, Burgeson RE, Gerecke DR & Gordon MK (2001) Alpha 1 (Xx) collagen, a new member of the collagen subfamily, fibril-associated collagens with interrupted triple helices. J Biol Chem 276: 23120-23126.
Kovanen V & Suominen H (1989) Age- and training-related changes in the collagen metabolism of rat skeletal muscle. Eur J Appl Physiol Occup Physiol 58: 765-771.
Kraal G, van der Laan LJ, Elomaa O, & Tryggvason K (2000) The marcophage receptor MARCO. Microbes Infect 2: 313-316.
Krane SM (1984) Genetic and acquired disorders of collagen deposition. In: Piez KA & Reddi AH (eds) Extracellular matrix biochemistry. Elsevier, New York, p 413-463.
Krane SM, Pinnell SR & Erbe RW (1972) Lysyl-protocollagen hydroxylase deficiency in fibroblasts from siblings with hydroxylysine-deficient collagen. Proc Natl Acad Sci U S A 69: 2899-2903.
Kuutti-Savolainen ER (1979) Enzymes of collagen biosynthesis in skin and serum in dermatological diseases. II. Serum enzymes. Clin Chim Acta 96: 53-58.
Kuutti-Savolainen ER & Kero M (1979) Enzymes of collagen biosynthesis in skin and serum in dermatological diseases. I. Enzymes of the skin. Clin Chim Acta 96: 43-51.
Kääpä E, Han X, Holm S, Peltonen J, Takala T & Vanharanta H (1995) Collagen synthesis and types I, III, IV, and VI collagens in an animal model of disc degeneration. Spine 20: 59-66; discussion 66-67.
Kääpä E, Holm S, Han X, Takala T, Kovanen V & Vanharanta H (1994) Collagens in the injured porcine intervertebral disc. J Orthop Res 12: 93-102.
Köwitz J, Knippel M, Schuhr T & Mach J (1997) Alteration in the extent of collagen I hydroxylation, isolated from femoral heads of women with a femoral neck fracture caused by osteoporosis. Calcif Tissue Int 60: 501-505.
Lehmann HW, Rimek D, Bodo M, Brenner RE, Vetter U, Worsdorfer O, Karbowski A & Müller PK (1995) Hydroxylation of collagen type I: evidence that both lysyl and prolyl residues are overhydroxylated in osteogenesis imperfecta. Eur J Clin Invest 25: 306-310.
Lo Cascio V, Bertoldo F, Gambaro G, Gasperi E, Furlan F, Colapietro F, Lo Cascio C & Campagnola M (1999) Urinary galactosylhydroxylysine in postmenopausal osteoporotic women: A potential marker of bone fragility. J Bone Miner Res 14: 1420-1424.
Majamaa K, Günzler V, Hanauske-Abel HM, Myllylä R & Kivirikko KI (1986) Partial identity of the 2-oxoglutarate and ascorbate binding sites of prolyl 4-hydroxylase. J Biol Chem 261: 7819-7823.
Majamaa K, Turpeenniemi-Hujanen TM, Latipää P, Günzler V, Hanauske-Abel HM, Hassinen IE & Kivirikko KI (1985) Differences between collagen hydroxylases and 2-oxoglutarate dehydrogenase in their inhibition by structural analogues of 2-oxoglutarate. Biochem J 229: 127-133.
Mayne R & Burgeson RE (1987) Structure and function of collagen types. Academic Press, Orlando, Florida.
Michelson PH, Tigue M & Jones JC (2000) Human bronchial epithelial cells secrete laminin 5, express hemidesmosomal proteins, and assemble hemidesmosomes. J Histochem Cytochem 48: 535-544.
Myllyharju J & Kivirikko KI (2001) Collagens and collagen-related diseases. Ann Med 33(1): 7-21.
Myllylä R (1976) Collagen glucosyltransferase. Acta Univ Oul A44: 1-47.

63
Myllylä R (1981) Preparation of antibodies to chick-embryo galactosylhydroxylysyl glucosyltransferase and their use for an immunological characterization of the enzyme of collagen synthesis. Biochim Biophys Acta 658: 299-307.
Myllylä R, Alitalo K, Vaheri A & Kivirikko KI (1981) Regulation of collagen post-translational modification in transformed human and chick-embryo cells. Biochem J 196: 683-692.
Myllylä R, Anttinen H & Kivirikko KI (1979) Metal activation of galactosylhydroxylysyl glucosyltransferase, an intracellular enzyme of collagen biosynthesis. Eur J Biochem 101: 261-269.
Myllylä R, Anttinen H, Risteli L & Kivirikko KI (1977) Isolation of collagen glucosyltransferase as a homogeneous protein from chick embryos. Biochim Biophys Acta 480: 113-121.
Myllylä R, Becvar R, Adam M & Kivirikko KI (1989) Markers of collagen metabolism in sera of patients with various rheumatic diseases. Clin Chim Acta 183: 243-252.
Myllylä R, Günzler V, Kivirikko KI & Kaska DD (1992) Modification of vertebrate and algal prolyl 4-hydroxylases and vertebrate lysyl hydroxylase by diethyl pyrocarbonate. Evidence for histidine residues in the catalytic site of 2-oxoglutarate-coupled dioxygenases. Biochem J 286: 923-927.
Myllylä R, Majamaa K, Günzler V, Hanauske-Abel HM & Kivirikko KI (1984) Ascorbate is consumed stoichiometrically in the uncoupled reactions catalyzed by prolyl 4-hydroxylase and lysyl hydroxylase. J Biol Chem 259: 5403-5405.
Myllylä R, Myllylä VV, Tolonen U & Kivirikko KI (1982) Changes in collagen metabolism in diseased muscle. I. Biochemical studies. Arch Neurol 39: 752-755.
Myllylä R, Pajunen L & Kivirikko KI (1988) Polyclonal and monoclonal antibodies to human lysyl hydroxylase and studies on the molecular heterogeneity of the enzyme. Biochem J 253: 489-496.
Myllylä R, Pihlajaniemi T, Pajunen L, Turpeenniemi-Hujanen T & Kivirikko KI (1991) Molecular cloning of chick lysyl hydroxylase. Little homology in primary structure to the two types of subunit of prolyl 4-hydroxylase. J Biol Chem 266: 2805-2810.
Myllylä R, Risteli L & Kivirikko KI (1975) Glucosylation of galactosylhydroxylysyl residues in collagen in vitro by collagen glucosyltransferase. Inhibition by triple-helical conformation of the substrate. Eur J Biochem 58: 517-521.
Myllylä R, Risteli L & Kivirikko KI (1976) Collagen glucosyltransferase. Partial purification and characterization of the enzyme from whole chick embryos and chick-embryo cartilage. Eur J Biochem 61: 59-67.
Noiva R (1999) Protein disulfide isomerase: the multifunctional redox chaperone of the endoplasmic reticulum. Semin Cell Dev Biol 10: 481-493.
Norman KR & Moerman DG (2000) The let-268 locus of Caenorhabditis elegans encodes a procollagen lysyl hydroxylase that is essential for type IV collagen secretion. Dev Biol 227: 690-705.
Notbohm H, Nokelainen M, Myllyharju J, Fietzek PP, Müller PK & Kivirikko KI (1999) Recombinant human type II collagens with low and high levels of hydroxylysine and its glycosylated forms show marked differences in fibrillogenesis in vitro. J Biol Chem 274: 8988-8992.
Oikarinen A (1977) Effect of cortisol acetate on collagen biosynthesis and on the activities of prolyl hydroxylase, lysyl hydroxylase, collagen galactosyltransferase and collagen glucosyltransferase in chick-embryo tendon cells. Biochem J 164: 533-539.
Oikarinen A, Anttinen H & Kivirikko KI (1976a) Hydroxylation of lysine and glycosylation of hydroxylysine during collagen biosynthesis in isolated chick-embryo cartilage cells. Biochem J 156: 545-551.
Oikarinen A, Anttinen H & Kivirikko KI (1976b) Effect of L-azetidine-2-carboxylic acid on glycosylations ofcollagen in chick-embryo tendon cells. Biochem J 160: 639-645.
Oikarinen A, Anttinen H & Kivirikko KI (1977) Further studies on the effect of the collagen triple-helix formation on the hydroxylation of lysine and the glycosylations of hydroxylysine in chick-embryo tendon and cartilage cells. Biochem J 166: 357-362.

64
Oikarinen A, Palatsi R, Linna SL & Peltonen L (1982a) Types I and III collagens and the activities of prolyl hydroxylase and galactosylhydroxylysyl glucosyltransferase in skin lesions of tuberous sclerosis. Br J Dermatol 107: 659-664.
Oikarinen A, Savolainen ER, Tryggvason K, Foidart JM & Kiistala U (1982b) Basement membrane components and galactosylhydroxylysyl glucosyltransferase in suction blisters of human skin. Br J Dermatol 106: 257-266.
O'Reilly MS, Boehm T, Shing Y, Fukai N, Vasios G, Lane WS, Flynn E, Birkhead JR, Olsen BR & Folkman J (1997) Endostatin: an endogenous inhibitor of angiogenesis and tumor growth. Cell 88: 277-285.
Pajunen L, Suokas M, Hautala T, Kellokumpu S, Tebbe B, Kivirikko KI & Myllylä R (1998) A splice-site mutation that induces exon skipping and reduction in lysyl hydroxylase mRNA levels but does not create a nonsense codon in Ehlers-Danlos syndrome type VI. DNA Cell Biol 17: 117-123.
Passoja K, Myllyharju J, Pirskanen A & Kivirikko KI (1998a) Identification of arginine-700 as the residue that binds the C-5 carboxyl group of 2-oxoglutarate in human lysyl hydroxylase 1. FEBS Lett 434: 145-148.
Passoja K, Rautavuoma K, Ala-kokko L, Kosonen T & Kivirikko KI (1998b) Cloning and characterization of a third human lysyl hydroxylase isoform. Proc Natl Acad Sci USA 95: 10482-10486.
Pihlajaniemi T & Rehn M (1995) Two new collagen subgroups: membrane-associated collages and type XV and XVIII. Prog Nucleic Acid Res Mol Biol 50: 225-262.
Pinnell SR, Krane SM, Kenzora JE & Glimcher MJ (1972) A heritable disorder of connective tissue. Hydroxylysine-deficient collagen disease. N Engl J Med 286: 1013-1020.
Pirskanen A, Kaimio AM, Myllylä R & Kivirikko KI (1996) Site-directed mutagenesis of human lysyl hydroxylase expressed in insect cells. Identification of histidine residues and an aspartic acid residue critical for catalytic activity. J Biol Chem 271: 9398-9402.
Poole A, Myllylä R & Davies BH (1989) Activities of enzyme of collagen biosynthesis and levels of type III procollagen peptide in the serum of patients with sarcoidosis. Life Sci 45: 319-326.
Poole A, Myllylä R, Wagner JC & Brown RC (1985) Collagen biosynthesis enzymes in lung tissue and serum of rats with experimental silicosis. Br J Exp Pathol 66: 567-575.
Popenoe EA & Aronson RB (1972) Partial purification and properties of collagen lysine hydroxylase from chick embryos. Biochim Biophys Acta 258: 380-386.
Pousi B, Hautala T, Heikkinen J, Pajunen L, Kivirikko KI & Myllylä R (1994) Alu-Alu recombination results in a duplication of seven exons in the lysyl hydroxylase gene in a patient with the type VI variant of Ehlers-Danlos syndrome. Am J Hum Genet 55: 899-906.
Pousi B, Hautala T, Hyland JC, Schroter J, Eckes B, Kivirikko KI & Myllylä R (1998) A compound heterozygote patient with Ehlers-Danlos syndrome type VI has a deletion in one allele and a splicing defect in the other allele of the lysyl hydroxylase gene. Hum Mutat 11: 55-61.
Pousi B, Heikkinen J, Schroter J, Pope M & Myllylä R (2000) A nonsense codon of exon 14 reduces lysyl hydroxylase mRNA and leads to aberrant RNA splicing in a patient with Ehlers-Danlos syndrome type VI. Mutat Res 432: 33-37.
Prockop DJ & Kivirikko KI (1995) Collagens: Molecular biology, diseases, and potentials for therapy. Annu Rev Biochem 64: 403-434.
Puistola U (1982) Catalytic properties of lysyl hydroxylase from cells synthesizing genetically different collagen types. Biochem J 201: 215-219.
Puistola U & Anttinen H (1982) Evidence for a relative excess of lysyl hydroxylase in chick embryo tendon and cartilage compared with bone and skin. Biochim Biophys Acta 717: 118-123.
Puistola U, Turpeenniemi-Hujanen TM, Myllylä R & Kivirikko KI (1980a) Studies on the lysyl hydroxylase reaction. I. Initial velocity kinetics and related aspects. Biochim Biophys Acta 611: 40-50.
Puistola U, Turpeenniemi-Hujanen TM, Myllylä R & Kivirikko KI (1980b) Studies on the lysyl hydroxylase reaction. II. Inhibition kinetics and the reaction mechanism. Biochim Biophys Acta 611: 51-60.

65
Rautavuoma K, Passoja K, Helaakoski T & Kivirikko KI (2000) Complete exon-intron organization of the gene for human lysyl hydroxylase 3 (LH3). Matrix Biol 19: 73-79.
Rautavuoma K, Takaluoma K, Passoja K, Pirskanen A, Kvist AP, Kivirikko KI & Myllyharju J (2002) Characterization of three fragments that constitute the monomers of the human lysyl hydroxylase isoenzymes 1-3. The 30-kDa N-terminal fragment is not required for lysyl hydroxylase activity. J Biol Chem 277: 23084-23091.
van der Rest M & Garrone R (1991) Collagen family of proteins. FASEB J 5: 2814-2823. Risteli L (1977) Developmental changes in collagen glycosyltransferase activities in chick
embryos. Biochim Biophys Acta 497: 673-681. Risteli L (1978) Further characterization of collagen galactosyltransferase from chick embryos.
Biochem J 169: 189-196. Risteli L, Myllylä R & Kivirikko KI (1976a) Partial purification and characterization of collagen
galactosyltransferase from chick embryos. Biochem J 155: 145-153. Risteli L, Myllylä R & Kivirikko KI (1976b) Affinity chromatography of collagen
glycosyltransferases on collagen linked to agarose. Eur J Biochem 67: 197-202. Risteli L, Risteli J, Ihme A, Krieg T & Müller PK (1980) Preferential hydroxylation of type IV
collagen by lysyl hydroxylase from Ehlers-Danlos syndrome type VI fibroblasts. Biochem Biophys Res Commun 96: 1778-1784.
Royce PM & Barnes MJ (1985) Failure of highly purified lysyl hydroxylase to hydroxylate lysyl residues in the non-helical regions of collagen. Biochem J 230: 475-480.
Royce PM, Moser U & Steinmann B (1989) Ehlers-Danlos syndrome type VI with normal lysyl hydroxylase activity cannot be explained by a defect in cellular uptake of ascorbic acid. Matrix 9: 147-149.
Ruotsalainen H, Sipilä L, Kerkelä E, Pospiech H & Myllylä R (1999) Characterization of cDNAs for mouse lysyl hydroxylase 1, 2, and 3, their phylogenetic analysis and tissue-specific expression in the mouse. Matrix Biol 18: 325-329.
Ruotsalainen H, Vanhatupa S, Tampio M, Sipilä L, Valtavaara M & Myllylä R (2001) Complete genomic structure of mouse lysyl hydroxylase 2 and lysyl hydroxylase 3/collagen glucosyltransferase. Matrix Biol 20: 137-146.
Ryhänen L (1975) Hydroxylation of lysyl residues in lysine-rich and arginine-rich histones by lysyl hydroxylase in vitro. Biochim Biophys Acta 397: 50-57.
Ryhänen L (1976) Lysyl hydroxylase. Further purification and characterization of the enzyme from chick embryos and chick embryo cartilage. Biochim Biophys Acta 438: 71-89.
Sambrook J, Fritsch EF & Maniatis T (1989) Molecular Cloning. A Laboratory manual, second edition. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, New York, USA
Savolainen ER, Kero M, Pihlajaniemi T & Kivirikko KI (1981) Deficiency of galactosylhydroxylysyl glucosyltransferase, an enzyme of collagen synthesis, in a family with dominant epidermolysis bullosa simplex. N Engl J Med 304: 197-204.
Sipilä L, Szatanik M, Vainionpää H, Ruotsalainen H, Myllylä R & Guenet JL (2000) The genes encoding mouse lysyl hydroxylase isoforms map to chromosomes 4,5, and 9. Mamm Genome 11: 1132-1134.
Smith DF, Kosow DP, Wu C & Jamieson GA (1977) Characterization of human platelet UDPglucose-collagen glucosyltransferase using a new rapid assay. Biochim Biophys Acta 483: 263-278.
Spiess J & Noe BD (1985) Processing of an anglerfish somatostatin precursor to a hydroxylysine-containing somatostatin 28. Proc Natl Acad Sci USA 82: 277-281.
Steinmann B, Gitzelmann R, Vogel A, Grant ME, Harwood R & Sear CH (1975) Ehlers-Danlos syndrome in two siblings with deficient lysyl hydroxylase activity in cultured skin fibroblasts but only mild hydroxylysine deficit in skin. Helv Paediatr Acta 30: 255-274.
Steinmann B, Royce PM & Superti-Furga A (1993) The Ehlers-Danlos syndrome. In: Royce PM & Steinmann B (eds) Connective tissue and its heritable disorders. Molecular, genetic, and medical aspects. Wiley-Liss, Inc., New York, p 351-407.
Suokas M, Myllylä R & Kellokumpu S (2000) A single C-terminal peptide segment mediates both membrane association and localization of lysyl hydroxylase in the endoplasmic reticulum. J Biol Chem 275: 17863-17868.

66
Sussman M, Lichtenstein JR, Nigra TP, Martin GR & McKusick VA (1974) Hydroxylysine-deficient skin collagen in a patient with a form of the Ehlers-Danlos syndrome. J Bone Joint Surg Am 56: 1228-1234.
Szpirer C, Szpirer J, Riviere M, Vanvooren P, Valtavaara M & Myllylä R (1997) Localization of the gene encoding a novel isoform of lysyl hydroxylase. Mamm Genome 8: 707-708.
Tajima S, Murad S & Pinnell SR (1983) A comparison of lysyl hydroxylation in various types of collagen from type VI Ehlers-Danlos syndrome fibroblasts. Coll Relat Res 3: 511-515.
Tajima S & Tokimitsu I (1995) Regulation of BPAG2 (180 kDa) gene expression in spontaneously transformed murine keratinocytes: tissue distribution, modulation during cell adhesion, and up-regulation by ascorbic acid. J Biochem (Tokyo) 118: 734-737.
Takala TE, Vuori J, Anttinen H, Väänänen K & Myllylä R (1986) Prolonged exercise causes an increase in the activity of galactosylhydroxylysyl glucosyltransferase and in the concentration of type III procollagen aminopropeptide in human serum. Pflugers Arch 407: 500-503.
Takala TE, Vuori JJ, Rahkila PJ, Hakala EO, Karpakka JA, Alen MJ, Orava YS & Väänänen HK (1989) Carbonic anhydrase III and collagen markers in serum following cross-country skiing. Med Sci Sports Exerc 21: 593-597.
Tenni R, Valli M, Rossi A & Cetta G (1993) Possible role of overglycosylation in the type I collagen triple helical domain in the molecular pathogenesis of osteogenesis imperfecta. Am J Med Genet 45: 252-256.
Turpeenniemi TM, Puistola U, Anttinen H & Kivirikko KI (1977) Affinity chromatography of lysyl hydroxylase on concanavalin A-agarose. Biochim Biophys Acta 483: 215-219.
Turpeenniemi-Hujanen TM (1981) Immunological characterization of lysyl hydroxylase, an enzyme of collagen synthesis. Biochem J 195: 669-676.
Turpeenniemi-Hujanen TM, Puistola U & Kivirikko KI (1980) Isolation of lysyl hydroxylase, an enzyme of collagen synthesis, from chick embryos as a homogeneous protein. Biochem J 189: 247-253.
Turpeenniemi-Hujanen TM, Puistola U & Kivirikko KI (1981) Human lysyl hydroxylase: purification to homogeneity, partial characterization and comparison of catalytic properties with those of a mutant enzyme from Ehlers-Danlos syndrome type VI fibroblasts. Coll Relat Res 1: 355-366.
Ünligil UM, Zhou S, Yuwaraj S, Sarkar M, Schachter H & Rini JM (2000) X-ray crystal structure of rabbit N-acetylglucosaminyltransferase I: catalytic mechanism and a new protein superfamily. EMBO J 19: 5269-5280.
Valtavaara M (1999) Novel lysyl hydroxylase isoforms. Acta Univ Oul A334: 1-63. Valtavaara M, Papponen H, Pirttilä AM, Hiltunen K, Helander H & Myllylä R (1997) Cloning and
characterization of a novel human lysyl hydroxylase isoform highly expressed in pancreas and muscle. J Biol Chem 272: 6831-6834.
Valtavaara M, Szpirer C, Szpirer J & Myllylä R (1998) Primary structure, tissue distribution and chromosome localization of a novel isoform of lysyl hydroxylase (lysyl hydroxylase 3). J Biol Chem 273: 12881-12886.
Valtavaara M, Szpirer C, Szpirer J & Myllylä R (2000) Primary structure, tissue distribution and chromosome localization of a novel isoform of lysyl hydroxylase (lysyl hydroxylase 3). J Biol Chem 275: 20956.
Virtanen P, Viitasalo JT, Vuori J, Väänänen K & Takala TE (1993) Effect of concentric exercise on serum muscle and collagen markers. J Appl Physiol 75: 1272-1277.
Vuorio E & De Crombrugghe B (1990) The family of collagen genes. Annu Rev Biochem 59: 837-872.
Walker LC, Marini JC, Grange DK, Filie J & Yeowell HN (1999) A patient with Ehlers-Danlos syndrome type VI is homozygous for a premature termination codon in exon 14 of the lysyl hydroxylase 1 gene. Mol Genet Metab 67: 74-82.
Wang Y, Xu A, Knight C, Xu LY & Cooper GJ (2002) Hydroxylation and glycosylation of the four conserved lysine residues in the collagenous domain of adiponectin. Potential role in the modulation of its insulin-sensitizing activity. J Biol Chem 277: 19521-19529.

67
Wassen MH, Lammens J, Tekoppele JM, Sakkers RJ, Liu Z, Verbout AJ & Bank RA (2000) Collagen structure regulates fibril mineralization in osteogenesis as revealed by cross-link patterns in calcifying callus. J Bone Miner Res 15: 1776-1785.
Weinstein E, Blumenkrantz N & Prockop DJ (1969) Hydroxylation of proline and lysine in protocollagen involves two separate enzymatic sites. Biochim Biophys Acta 191: 747-750.
Wenstrup RJ, Murad S & Pinnell SR (1989) Ehlers-Danlos syndrome type VI: clinical manifestations of collagen lysyl hydroxylase deficiency. J Pediatr 115: 405-409.
Wixon J, Blaxter M, Hope I, Barstead R & Kim S (2000) Caenorhabditis elegans. Yeast 17: 37-42. Yamauchi T, Kamon J, Waki H, Terauchi Y, Kubota N, Hara K, Mori Y, Ide T, Murakami K,
Tsuboyama-Kasaoka N, Ezaki O, Akanuma Y, Gavrilova O, Vinson C, Reitman ML, Kagechika H, Shudo K, Yoda M, Nakano Y, Tobe K, Nagai R, Kimura S, Tomita M, Froguel P & Kadowaki T (2001) The fat-derived hormone adiponectin reverses insulin resistance associated with both lipoatrophy and obesity. Nat Med 7: 941-946.
Yeowell HN (2002) Lysyl hydroxylases. In: Creighton TE (ed) Wiley encyclopedia of molecular medicine. John Wiley and Sons Inc., New York, p 1980-1984.
Yeowell HN, Allen JD, Walker LC, Overstreet MA, Murad S & Thai SF (2000a) Deletion of cysteine 369 in lysyl hydroxylase 1 eliminates enzyme activity and causes Ehlers-Danlos syndrome type VI. Matrix Biol 19: 37-46.
Yeowell HN, Ha V, Walker LC, Murad S & Pinnell SR (1992) Characterization of a partial cDNA for lysyl hydroxylase from human skin fibroblasts; lysyl hydroxylase mRNAs are regulated differently by minoxidil derivatives and hydralazine. J Invest Dermatol 99: 864-869.
Yeowell HN, Ha V, Clark WL, Marshall MK & Pinnell SR (1994) Sequence analysis of a cDNA for lysyl hydroxylase isolated from human skin fibroblasts from a normal donor: differences from human placental lysyl hydroxylase cDNA. J Invest Dermatol 102: 382-384.
Yeowell HN & Pinnell SR (1993) The Ehlers-Danlos syndromes. Semin Dermatol 12: 229-240. Yeowell HN & Walker LC (1997) Ehlers-Danlos syndrome type VI results from a nonsense
mutation and a splice site-mediated exon-skipping mutation in the lysyl hydroxylase gene. Proc Assoc Am Phys 109: 383-396.
Yeowell HN & Walker LC (1999a) Tissue specificity of a new splice form of the human lysyl hydroxylase 2 gene. Matrix Biol 18: 179-187.
Yeowell HN & Walker LC (1999b) Prenatal exclusion of Ehlers-Danlos syndrome type VI by mutational analysis. Proc Assoc Am Phys 111: 57-62.
Yeowell HN & Walker LC (2000) Mutations in the lysyl hydroxylase 1 gene that result in enzyme deficiency and the clinical phenotype of Ehlers-Danlos syndrome type VI. Mol Genet Metab 71: 212-224.
Yeowell HN, Walker LC, Farmer B, Heikkinen J & Myllylä R (2000b) Mutational analysis of the lysyl hydroxylase 1 gene (PLOD) in six unrelated patients with Ehlers-Danlos syndrome type VI: prenatal exclusion of this disorder in one family. Hum Mutat 16: 90.