Embed Size (px)
Citation preview

Catalase Studies on Normal and Cancerous Rats"
DAVIDAPPLEMAN,EDWIN R. SKAVINSKI,ANDABRAHAMM. STEIN
(From the College of Agriculture, L'nirersity of California, Los Angele«)
Several investigators have studied the effect oftumor growths on the activity of catalase in theliver (7, 8, 13, 16), kidney (7, 8, 9), and erythro-cytes (7) of various animals. Brahn (1) originallyreported a decreased catalase activity in the liversof humans with malignant growths. Some of themore recent work has been concerned with the effects of inanition (10, 11) and the dietary proteinlevel (16) upon the liver catalase of normal andcancerous rats. Greenstein and associates (9) havereported that a 3-day starvation period leads to nodecrease of rat liver catalase, whereas Miller (10,11) has shown that a 7-day period produces a definite drop in catalase activity which is greater thanor approximately equal to the liver protein loss.Weil-Malherbe and Schade (16) have reportedthat the dietary protein level has little effect on theability of the tumor to produce drastic reductionsof rat liver catalase. Diets containing 8 per centand 45 per cent casein showed no change in theliver catalase of normal rats, whereas the intra-peritoneal injection of sheep serum produced adefinite increase in the liver catalase of normalrats. The high level of dietary protein has, however, resulted in increased tumor growth and lesssevere losses in original body weight of the rat.
The extensive studies of various liver enzymesin cancerous animals by Greenstein and his associates (3-6) and those mentioned above (1, 13, 16),particularly with respect to catalase, have established the following facts with respect to the effectof tumors on the relative activity of catalase invarious organs:
1. There is a decrease in liver catalase activitywith the progressive growth of the tumor. Within2-3 weeks after tumor implantation, the activitymay be as low as 1/10 to 1/20 of the originalvalue.
2. The effect of the tumor growth is reversible,the liver catalase returning to normal 24-48 hoursafter removal of the tumor.
3. There may be large individual variations of* This investigation was aided by a grant from the Uni
versity of California cancer research funds.Received for publication April 24, 1950.
liver catalase activity in cancerous animals seemingly in the same state of tumor development.
4. The kidney catalase is affected by tumorgrowth toa much lesser degree than liver catalase.The erythrocyte catalase is not affected.
5. Attempts to find activators, toxic substances,or inhibitors in the blood and other organs haveproved fruitless, and it is believed that there hasbeen an actual decrease in concentration of catalase in the liver. This is believed to be due to interference with the synthetic process necessary formaintenance of the catalase concentration in theliver. Recently, Nakahara and Fukuoka (12) havereported a toxic fraction extracted from humantumors which lowers the liver catalase activity inmice when injected intraperitoneally in 100-mg.quantities.
6. The drastic reduction of catalase compared toother liver enzymes and the fact that implantationof growing embryonic tissue and pregnancy (6)do not affect the liver catalase are evidence in favor of the specific ability of the tumor to producea decrease in liver catalase activity. However, theobservations on the effect of the dietary proteinlevel on liver catalase do not preclude the possibility of the nonspecificity of the tumor in producing a decrease liver catalase activity.
The similar structural chemical relationship between catalase and hemoglobin and data in theliterature (14, 15), which indicate a possible correlation between the simultaneous occurrence of adecreased liver catalase activity and a severe anemia, suggested that interference with erythropoie-sis and catalase synthesis were closely related.This hypothesis might indeed be tested readily bydetermining the liver catalase activity of animalswith various types of anemias. The above facts inconjunction with the effect of inanition (10, 11)and the exaggerated protein requirements of arapidly growing tumor, as possible causes for decreased liver catalase activity, led to some quantitative studies to determine the changes in thetotal units of catalase activity in the liver, eryth-rocytes, and kidneys of normal, cancerous, andprotein-depleted rats. An attempt is made to show
498
Research. on August 13, 2019. © 1950 American Association for Cancercancerres.aacrjournals.org Downloaded from

APPLEMANet al.—Catalase in Normal and Cancerous Rata
variations in the major organs containing catalasewhen corrections are made for variations in sizedue to the presence of the tumor or to other experimental conditions.
MATERIALS AND METHODSAnimals.—Adult rats of both sexes of a Long-
Evans black-hooded strain were used for all theexperimental work. Individual groups were usuallyselected so that they were all the same age, withinvery close limits. They were maintained on a routine diet of Rockland pellets with bi-weekly supplements of lettuce or cabbage greens. (The routine diet contained approximately 21 per cent protein.)
The essentially protein-free diet consisted of 80per cent cornstarch, 15 per cent Wesson oil, 4 percent U.S.P. salt mixture No. 2,1 per cent cod liveroil, and appropriate quantities of thiamine, ribo-fiavin, pyridoxine, nicotinamide, niacin, choline,and calcium pantothenate.
Tumors.—All experimental animals were implanted subcutaneously with active fibrosarcomatransplants (Series No. 8), originally induced bysubcutaneous injections of methylcholanthrene.Data are available on the amino acid compositionof this fibrosarcoma and homologous normal tissue (2). Methods of tumor transplantation weresimilar to those reported in the preceding reference. One series of experimental animals was implanted with the Jensen rat sarcoma.
Preparation of tissues for analysis.—The animalwas anesthetized with ether, and 1 ml. of a 1.5 percent solution of vital red dye in 0.85 per cent salinesolution was injected into the jugular vein using a27-gauge needle and a tuberculin syringe. The abdominal cavity was exposed, and after approximately 6 minutes the animal was exsanguinated byremoving the blood from the abdominal aorta bymeans of a syringe rinsed with a saline solution ofheparin, of suitable concentration to prevent clotting. Aliquots of the blood samples were used asfollows: (a) A 1-ml. sample for catalase estimationwas placed into volumetric flasks and stored in thefrozen state. The cells were hemolyzed and appropriately diluted with buffer for the estimation ofcatalase. (b) A 1-ml. sample was diluted to 100 ml.with 0.85 per cent saline, aliquots centrifuged, andthe concentration of dye in the supernatant solution compared with a standard using the Klett-Summerson colorimeter with a No. 54 green filter.With reasonable care no significant error is introduced due to hemolysis of erythrocytes. This procedure supplied the data necessary for calculationof the blood volume, (c) A \-m\. sample dilutedwith 0.85 JXTcent saline was centrifuged for 30
minutes at 2,500-3,000 r.p.m. to déterminethecell volume. Hemoglobin, nitrogen, and henmto-crit determinations performed on saline-washederythrocyte samples of cancerous and normal ratsshowed that the hemoglobin and nitrogen contentcould be closely correlated with the volume ofpacked blood cells. Thus, the hematocrit value ortotal red blood cell volume would be directly proportional to the corresponding hemoglobin value.
The liver, kidneys, and spleen were removed,rapidly weighed, and immediately placed in a deepfreeze unit. The liver and erythrocyte catalase activity was determined on the same day that theanimal was sacrificed. Kidney catalase was determined after 1-4 days of storage in the deep freeze,most cases not exceeding 2 days of storage. Preliminary tests indicated that whole liver tissuestored for 1 week showed no loss of catalase activity. As preliminary tests on the catalase activityof spleen and bone marrow of normal and cancerous rats showed no distinctly significant differences, the spleen weight was determined only tofind the degree of enlargement associated with tumor growth.
The whole liver was ground 10 minutes at 0°C.
with 5 times its weight of M/15 phosphate bufferof pH 7.0 in a small size monel metal Waringblendor. This homogenate was appropriately diluted with the buffer for the catalase determination, and 5-ml. aliquots were taken for the determination of nitrogen by the customary semi-microKjeldahl method. The kidneys were homogenizedsimilarly with 25 ml. of phosphate buffer regardless of weight, and similar methods of catalase andnitrogen determinations employed.
Estimation of catalase activity.—The enzymaticactivity of the various tissues was determined bymeasuring the rate of oxygen evolution from H2O2manometrically. The conditions for the determinations were as follows: temperature, 0°C., pH, 7.0;
5 ml. of 1.2 N H2O2; 1 ml. enzyme.All dilutions of the substrate and enzyme prepa
rations were made with M/15 phosphate buffer.The enzyme concentration was adjusted by properdilution to approximately the same range of activity in order that the ratio of substrate to enzyme concentration was fairly constant. Preliminary investigations were performed with erythrocyte and liver catalase solutions in order to selecta substrate to enzyme concentration which showeda constant proportionality with enzyme concentration even with appreciable variations of substrate. The proportionality between enzyme activity and concentration, at any reaction rate overa limited range, makes it possible to express enzyme activity in terms of arbitrary units. The ar-
Research. on August 13, 2019. © 1950 American Association for Cancercancerres.aacrjournals.org Downloaded from

500 Cancer Research
bitrary unit is defined as that amount of catalasewhich will liberate 1 nil. of oxygen per second froma 1.0 N H2O2 solution at 0°C.
Enzyme and nitrogen determinations were performed in duplicate, the average values of the results obtained being used to calculate the unitenzyme content. The usual rate of oxygen evolution corresponded to approximately 2 ml. perminute, and calculation of the enzyme activitywas obtained by determining the maximum initialreaction rate. The error in the catalase determination is less than 5 per cent.
RESULTS
AH the pertinent data are given in the tables.Catalase activity is calculated on the bases of nitrogen, fresh tissue weight, total weight of the animal, and a corrected body weight which does notinclude the weight of the tumor.
Liver catalase activity.—Our experimental resultsconfirm in a general way the observations of otherinvestigators—namely, that rats with growing
tumors suffer a loss of liver catalase activity, butthe decrease in catalase is neither as large nor asconsistent as those reported by Greenstein and co-workers. In no case did liver catalase drop to lessthan 20 per cent of normal, even in animals wherethe tumor composed 30 per cent of body weight.And only rarely did we obtain so large a decrease. The failure to obtain as low values as otherinvestigators may be an apparent one, occasionedby a difference in the method used for the determination of catalase activity.
A comparison of Tables 1 and 9 for the UCLAfibrosarcoma and the Jensen rat sarcoma showsvery clearly that the catalase loss from the liver ismuch greater and more consistent in the lattertumor. In fact, when the liver catalase activity iscalculated on the basis of units per 100 gm. bodyweight, as we have done in these tables, a largenumber of rats bearing the UCLA fibrosarcomaapproach the normal value for liver catalase.
It will be pointed out later that usually whenthe liver catalase is only slightly reduced, otherphenomena characteristic of tumor-bearing ratsare not apparent.
Eri/tlirocyte catalase.—In the UCLA fibrosarcoma the erythrocyte catalase activity per unit cellvolume remains practically the same as in normalanimals. The same is essentially true of the ratswith the Jensen sarcoma. It should be noted, however, that in the UCLA fibrosarcoma the volumeof erythrocytes per 100 gm. of total weight iswithin the normal range; this is not the case inrats with the Jensen sarcoma. These show a drop
in the unit volume of erythrocytes to between 60per cent and 80 per cent of normal; this, in spiteof the relatively large increase in blood volume.
Kidney catalase.—The activity of catalase in the
kidneys of cancerous rats suffers a decrease withboth tumors. However, when the catalase is calculated in units per 100 gm. of body weight, mostof the values fall within the normal range.
Other effects of cancer on the host.—In general,the liver of a tumor-bearing rat is increased in sizeabove that of normal—the increase may be asgreat as 60 per cent. This is more striking in theJensen sarcoma than in the UCLA fibrosarcoma.A high degree of spleen enlargement was also observed.
In most cases where the tumor is advancing tothe stage where there is a definite decrease in livercatalase, the blood volume of the animal per 100gm. of total weight is increased 25-50 per cent
above the normal values. The erythrocyte volumeper 100 gm. total weight either remains virtuallynormal, as in the case of the rats with the UCLAfibrosarcoma (Table 1), or suffers an appreciabledecrease as in the case of animals with the Jensensarcoma. In either case, the higher blood volumeexaggerates the drop in the hematocrit, and it mayindicate an apparent anemia when no true anemiaexists.
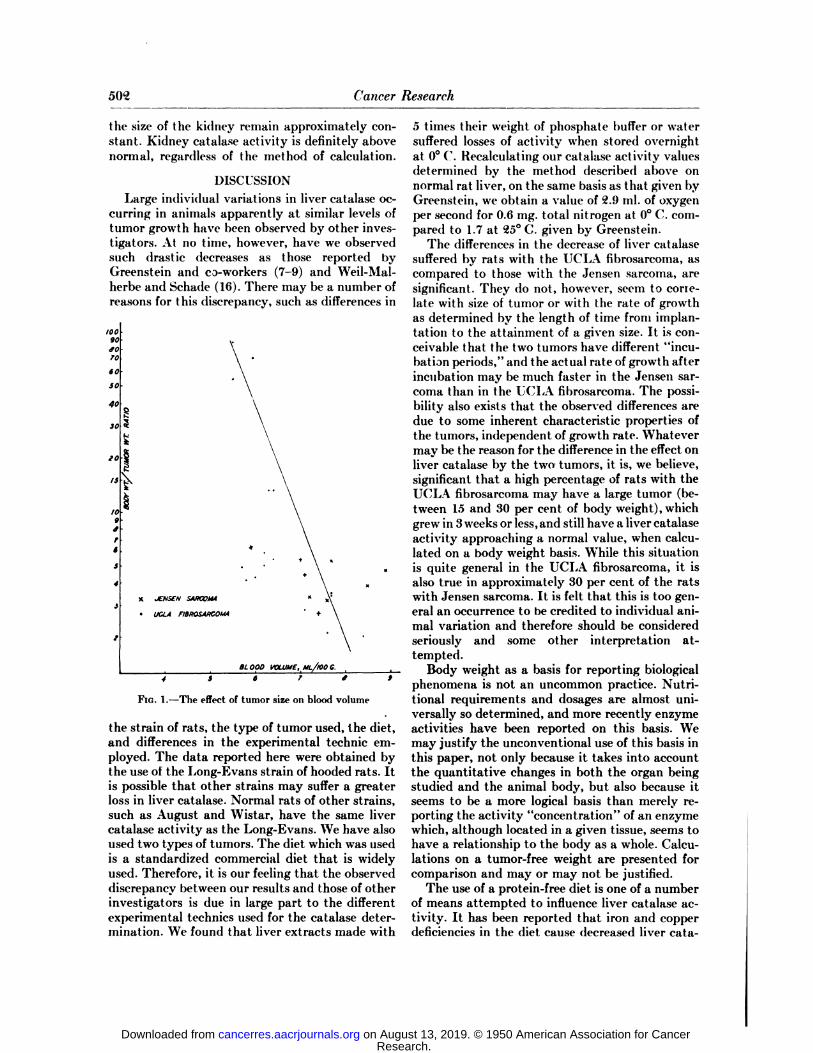
Figure 1 gives a graphic relationship betweentumor size as related to body weight and bloodvolume. It should be pointed out that the clusterof points which do not fit the graph represents ratsbearing the UCLA fibrosarcoma; although theseanimals developed large tumors, their liver catalase was within the normal range. It is regrettablethat data were not obtained with the Jensen sarcoma at early and intermediate stages of tumorgrowth.
Effects of a protein-free diet.—The results of this
experiment are summarized in Table 3. All animals placed on the diet suffer a fairly rapid loss ofliver catalase. After approximately 10 days of theprotein-free diet, the liver catalase stabilizes atapproximately one-half the normal value, and itremains there for the duration of the experiment—¿�53days. Returning the animal from the protein-free diet to a normal diet (21 per cent protein) for24-48 hours restores the liver catalase to a normalvalue (Table 3).
It is of interest to note that, although protein-depleted rats show a decrease in liver catalase ofthe same magnitude as rats with growing tumors,they do not show the other accompanying phenomena of the tumor-bearing rat. The size of theliver, the blood volume, erythrocyte volume, and
Research. on August 13, 2019. © 1950 American Association for Cancercancerres.aacrjournals.org Downloaded from

00 (V « —¿�. f.^t-wtcK^-sot-co etft •¿�*O *ÃŽ5ÃŽC¿�""
80 O O «5(N ~ —¿�f «5«5SO in :;
**5 ^â„¢Ã> ~o<x t- so o —¿�S 00
o»nt~ <x so so
•¿�*- O ^ OS XS ^««îoot- --»
« -t St
; x »o o *o
iOOiOO*-—'«-Ã^fWSÃCSOOO^-
S ^W1TO* «5
10- •¿�•¿�' »-—¿�»-.
' CSO* SO« 50 *f !
w i—'-^ «S W^OÕÃOt-'ffOli—aO rt .
«5^p« «5
' « O Ci Õ- ,•¿�•¿�•¿�«o •¿�o oo <•¿�r-> O «5 O <— •¿�—'
_, ce x Ão50 i—o <s •¿�'fÃoS!<»coo o t-». <*8 i?8 S?°.
_ X *f t-
9f x »2co *f *i«
"»<cooot-~,-os-li$;
sHB1H7,ZaQ-O1In£r-^<
J~L-UXHiHE
EFFECTiHM
(>»00 O-<f^.eoîcossot*—a*o*wotSSm^OOt,9,
005§S?^2^Ofr-X-fWOS
X«^"fc** -X^iOO1^1•—«îcoooîo^
(VQO^rwKx^
os a 07 i—x*i— •¿�^sOOO»Ci-HOaCiO^Õ'0<N5'î2t*
«2CS «5O&•-fco^00*^ ^^^^csSoÃTfx-^oo^^o^^o»««*^C
50 "V ÕS •¿�*.u, CSX-OCSSOCSTfiGOO'f•**
50- •¿�•¿�- •¿�(W •¿�«5»O f» QrtCSt-5O®O«5»tOi—•¿�GO'f'^OiOtX§O
*~* CS Ot-öo»ooorjicoox*
—¿�, •¿�3 - - - •¿�•¿�-9*-OOÃDb-OÕatÃ5-f.oo»cw^-T—ao*cto(No*oiX
—¿�W00*SîO
•¿� •¿� •¿� •¿� 'Oà -Or-^ t1- t*"^OCXooOO^W^H^^t^t^iÃ^Q^
^(QJ«Î
—¿�O t-Å“gOTb-00!OCWOO*X^-t-xÅ“oooOw5<w>-HF2c^^2!5'5'ϥ
•¿�•H
"¿ "fe ^f—
—¿�>,-2*^1
lili-g c ococ«j
¿iSD|yM&O0-5-.—.^-^--OOAw 9 LI w I-H^M>•tÃ*^«*®feoo--^--^tf
> '•••^^ 5. ^â„¢ "-H O5 «"^*^ •¿�1*o* -\ 2 "-^~~^.*^ j-j«tjÃ^33 »•§«•f.'!'!'!iï-5_o-gccpcc-3o—
^2*-2^3^3 3^¿".
^ "*^ 2 T - ¿*¿'¿*¿i¿i*> '>CI^fllsl'il-SlITPil
t! !!&"•"¡i*C•¿�"Û. ^ C "^^4ÃCÃlÃ^ÃSS4)lìfS^iilllliiil"*
** S!*B f1 w o 5 o w ^* ^>S>,-5"g-§^ ~Å“Å“Å“ " ** ^-i— ^esE«;ç
iH»^ -J1ssso<!ÕI'S.H«îM2i;i~i;=^i,CgKÌB)m
g*'S.•¿.
co-
•¿�£»œ «5o*Aç-^C^t^WSOO*^AÃŽO
-«SOOOt--9,«CÔS§?£y
« <- 0» «5 —¿�t--»SOOO^-O^Wtxco^ooS•»f
t- ^H co •¿�*as•^.-t-t~t-'*'CXO3CC «5O••
- - - .«3.^.^H^^ÇO^eo-*oox —¿�oos-.o«co«»eoif5tO
•¿�*Ot i— «5<oi,K5'-'"<0'*tD>noSSOU5O^"cO'foot-i—
'OxoÃeó^aÃoox^Ã1
'f ^f «5 o «5[i. f^i ^ "îCD «if t—*O t~" f-^ MAi-oo«o'ot--~œ-§^,
****•
¿-t -f ^^"ti^cef aì2-è-'Sì|iiii||ill¿pggeMM-s
8S.
ufiSsfSSi^j .?^S5° ^22^>e S§~* S "•"3 M - ~ -2 Õ'5 "'S §T-5§
||i s fi"! fil ¿¿-s1]1*! 1 - * S l-t-t-l'l'-tl :^ S 1!*^fc fl "W ^'>-^^^^*J*JO O-O-ill!!§|fiifi|hîi|IlÃ^-i||
iiilljif|^^?ti-3rlïll5§g322
'8**&a±P888f5Sfei>.g11ï,-ë-s|îal^oufcsiëëfcr£8
o Z- a-2acae>>>3^>p»jM(gwfls«KaaaMMa
Research. on August 13, 2019. © 1950 American Association for Cancercancerres.aacrjournals.org Downloaded from
5()"2 Cancer Research
the size of the kidney remain approximately constant. Kidney catalase activity is definitely abovenormal, regardless of the method of calculation.
DISCUSSIONLarge individual variations in liver catalase oc
curring in animals apparently at similar levels oftumor growth have been observed by other investigators. At no time, however, have we observedsuch drastic decreases as those reported byGreenstein and co-workers (7-9) and Weil-Mal-herbe and Schade (16). There may be a number ofreasons for this discrepancy, such as differences in
10090fOTO
tofO
40
to
IS
IO
s
4
.•:>.i : SA/KOMA
UCLA FIBROSARCOMA
BLOOD VtJLUMC,ML/100 C.
4 l S 7 t i
FIG. 1.—The effect of tumor size on blood volume
the strain of rats, the type of tumor used, the diet,and differences in the experimental technic employed. The data reported here were obtained bythe use of the Long-Evans strain of hooded rats. Itis possible that other strains may suffer a greaterloss in liver catalase. Normal rats of other strains,such as August and Wistar, have the same livercatalase activity as the Long-Evans. We have alsoused two types of tumors. The diet which was usedis a standardized commercial diet that is widelyused. Therefore, it is our feeling that the observeddiscrepancy between our results and those of otherinvestigators is due in large part to the differentexperimental technics used for the catalase determination. We found that liver extracts made with
5 times their weight of phosphate buffer or watersuffered losses of activity when stored overnightat 0°('. Recalculating our catalase activity values
determined by the method described above onnormal rat liver, on the same basis as that given byGreenstein, we obtain a value of 2.9 ml. of oxygenper second for 0.6 mg. total nitrogen at 0°C. compared to 1.7 at 25°C. given by Greenstein.
The differences in the decrease of liver catalasesuffered by rats with the UCLA fibrosarcoma, ascompared to those with the Jensen sarcoma, aresignificant. They do not, however, seem to cone-late with size of tumor or with the rate of growthas determined by the length of time from implantation to the attainment of a given size. It is conceivable that the two tumors have different "incubation periods," and the actual rate of growth after
incubation may be much faster in the Jensen sarcoma than in the UCLA fibrosarcoma. The possibility also exists that the observed differences aredue to some inherent characteristic properties ofthe tumors, independent of growth rate. Whatevermay be the reason for the difference in the effect onliver catalase by the two tumors, it is, we believe,significant that a high percentage of rats with theUCLA fibrosarcoma may have a large tumor (between 1.5and 30 per cent of body weight), whichgrew in 3weeks or less, and still have a liver catalaseactivity approaching a normal value, when calculated on a body weight basis. While this situationis quite general in the UCLA fibrosarcoma, it isalso true in approximately 30 per cent of the ratswith Jensen sarcoma. It is felt that this is too general an occurrence to be credited to individual animal variation and therefore should be consideredseriously and some other interpretation attempted.
Body weight as a basis for reporting biologicalphenomena is not an uncommon practice. Nutritional requirements and dosages are almost universally so determined, and more recently enzymeactivities have been reported on this basis. Wemay justify the unconventional use of this basis inthis paper, not only because it takes into accountthe quantitative changes in both the organ beingstudied and the animal body, but also because itseems to be a more logical basis than merely reporting the activity "concentration" of an enzyme
which, although located in a given tissue, seems tohave a relationship to the body as a whole. Calculations on a tumor-free weight are presented forcomparison and may or may not be justified.
The use of a protein-free diet is one of a numberof means attempted to influence liver catalase activity. It has been reported that iron and copperdeficiencies in the diet cause decreased liver cata-
Research. on August 13, 2019. © 1950 American Association for Cancercancerres.aacrjournals.org Downloaded from

00„i3
nHSSEJ1i•<u¡M-gL¿oX~sEfa7.5~Oa5yOgeCd
Cd£¿ROTEIP-i3CdSH^
oscoi-iCO O «O9! 9<<NX
t 9»t- O 00 "B •¿�*•¿�*St*co9>
CO O»ftX 9* O* îOl—St-- •¿�-9<^--f O !O 9* f*9*O!
t-OS-COO«««
9»i—
«ÎX^
2 ^ - —¿�-<vo
cox-^i-. «5 «5•¿�**^
coo o*o*o
<**o^-co »o»oos x* <S* - •¿� -co"-
CO O «ÎO»O*Ö*OS
p00OS
Q»*0 «5«NCOfi*MNco•*
*oogÃ’> Ã> OS O COt>OS
CO O «5CO COCOg
os «oco o»o*x—
-—¿�t-iCO 0* «ÎOS X
t- ' 0 «0« 0»S<«
'-4ao*K«.-222i*.»*¿2S«-iO*
t-00gOS *O OS «CXX^^Uo
«1g
S 35i-i«s38g9Õ91 O *O 9( ^*^fÕ5
6MOE
2^22£
s'E'S+*
§°¿?¿"lÃlja
HlCJïXeQOi^^»n
3St
9)9,2?
2SS•
«Î <5Ît-_°
00 l-V.9»
"»S•
os ocoIOoO*
CO ^r-<91»•
X «QÕ§§»-¡S
SSco»—i
CO «C«§91
CO «5f-t-•
O CO^ CO ^P«Îo-a
3«7Ε
œ^i—O*!OXt>-i
i*XÌ0E
^^ a«.s«60g ~^-|>
^1•5-5
E:«.3s og'?
'> B*jo
o is 2J5112
1*XX^3!S<
äa 9* cot-cooo< z+1u0
EuJU<a>hgas
SSr -d•<9»9(g
OSt-""**
°a
«^s»ocoo00
OSg^"CO^"00
OÜ-
QQ *>SO*f
0*O0
XEt, ^ COCO"*
COO00
^^o*co5cooot
co«aœ«*CO9(Ow
O •¿�"
CO 00C++
9»1Cco
cooOS
Xa,^ »œCO
9<oOS
Os -,1-^9«»tort
U5a «atœ9*COO00
XSos00œ"9«
Os
151Ì&£"°*!*C
^>ili
1OSXas»O
«5+1t-«5COIOX«Jo•0o•ro»•o00«ice«5e»
•¿�o00co«5
«S«5CM"O1oo"i
I"e1S9»
CO'
CÌ9ÌÌ+19(
CMet^i»eCMIQI-H00CMA,_«CM0
CMaaCM
CMCMCMiO—ì¿
_sCs0^+1*CMo00sCM00
00s00g00ciCM'e00s00Q
O00gsCM500<oitg.1'33>>italase
activil0|B"5
O_-' S°"+l+1«Sce9t^SAi'Ng«OCMO£T1gist^
»oIQKi§CMa§5°|>.1li3%>j_>-US2P^
oco+1§0091
•¿�*SO*
«3"500o*
-^i?9»
-•-t9¡
5$9(
Si
~??9(
COCMS
sIQt-X
9*COXs
-,i-*00¡^
00tiO1
1'E'E33£
£>•?•¿�>catalaseacti catalaseacti**>
>>t««7-i
eci^-sJXX5-1
+1$*«OS
•¿�*«5^.552S
So•O
00jo
»—gg>O
001«5
91ss$8sss-t§_il
..COD Õ-c'e Z 2t8'S I12RI
gMvS Õ, S,i.«fe
-S •¿�§s-Ã'Sft V *f-Qrrt
to' * SO15^
g WCGC:" Z—¿�- -~ass
Research. on August 13, 2019. © 1950 American Association for Cancercancerres.aacrjournals.org Downloaded from

504 Cancer Research
lase activity (14). In some preliminary experiments weproduced marked liver catalase reductionsby producing anemia with a folie acid antagonist,but observed no reduction of erythrocyte catalase. The production of polycythemia by subcutaneous injections of cobalt has also resulted in a decrease of liver catalase activity. Our work on inanition confirms the results reported by Miller(10, 11). Animals starved for 5-7 days suffer a lossof liver catalase of approximately the same magnitude as rats on a protein-free diet. The resultsare, however, more variable. The most significantresult of this experiment is the remarkable constancy of the new level of liver catalase, which develops in 7-10 days and persists as long as the animals are kept on a protein-free diet. It would seemas if this lower limit of liver catalase is a minimum,essential for the survival of the animal. When livercatalase drops below this level, the animal is evidently unable to carry on normal metabolic processes for any significant time. The production ofsuch a situation by growth of a tumor indicates aterminal condition leading to death in a shorttime.
When the animal is returned to a normal diet,or for 24 hours on a high casein diet, liver catalasereturns to a normal value in approximately thesame time interval as in the case of a cancerousanimal when the tumor is excised. It should also benoted that, although the liver catalase is reducedto approximately 50 per cent of normal, which isfairly close to the average reductions suffered bythe tumor-bearing rats, this is not accompanied bythe same phenomena that were observed in thecase of the tumor-bearing rats. The size of theliver and the blood volume remains essentially normal; the erythrocyte volume suffers a very smalldecrease, while the kidney catalase appears to increase with increased time on a protein-free diet.These changes are relatively minor as compared tothose that occur in the tumor-bearing rat.
The results reported in this paper, as well asmany observations on cancerous animals whereonly one or two factors (blood volume or livercatalase, etc.) were determined and are thereforenot included here, prompt us to suggest some possible explanations of the observed phenomena. Ifwe assume that in a normal rat liver catalase is ina steady state, i.e., the rate of its destruction isequal to the rate of synthesis, then it follows thatany deviation from the normal is due to a disturbance in this dynamic equilibrium. Thus, in acancerous rat, or a rat on a protein-free diet, theobserved decrease in catalase activity of the livermay be brought about by either an increase in therate of destruction or a decrease in the rate of syn
thesis. It is also possible that, in the case of tumor-bearing rats, the rate of catalase synthesis may remain normal or even increase, but the rate of destruction, due to tumor growth, is increased tosuch an extent that catalase activity continues todrop. In the case of the rats on a protein-free dietboth the rates of synthesis and ot destruction decrease approximately to the same extent, resultingin a new steady state at a lower catalase activity.
That some catalase synthesis continues in thetumor-bearing rat, even at fairly advanced stagesof cancer growth, is evidenced by the fact that inthe UCLA fibrosarcoma the erythrocyte volumeper unit of total weight is maintained at a fairlyconstant level with a constant catalase activity.This indicates that new erythrocytes must beformed, since the total weight of the rat continuesto increase. It follows, therefore, that both hemoglobin and catalase must be synthesized, because,as it has been pointed out earlier, there does notseem to be a decrease of either hemoglobin orcatalase in the erythrocytes of tumor-bearing animals.
One may speculate that there is no relationshipbetween the catalase synthesis in the liver andthat in the erythrocytes. These may be quite independent processes. The data obtained with ratson a protein-free diet support this supposition.Here the liver catalase drops to 50 per cent of normal, while the erythrocyte volume and catalaseactivity remain constant. Since the average life ofrat erythrocytes is approximately 30 days, it isevident that those animals in the experimentwhich were kept longer than 30 days had a complete replacement of their erythrocytes. Thus, thetotal erythrocyte catalase was either conserved orresynthesized, or the average life period of theerythrocytes was increased.
The suggestion of Greenstein that the tumor"by abstracting from the circulation some materi
al essential to the normal maintenance of the livercatalase" decreases the liver catalase, is not out of
harmony with our observations. It is only necessary to add the additional supposition that thesubstance abstracted is specific for the synthesisof liver catalase and is not essential for the synthesis of catalase in the erythrocytes or the kidney,thus accounting for the numerous instances whereonly liver catalase is decreased and not the catalase in the erythrocytes or kidney.
SUMMARY
Comparative studies of catalase activity in theliver, kidney, and erythrocytes of normal, cancerous, and protein-depleted rats were made. The results indicate:
Research. on August 13, 2019. © 1950 American Association for Cancercancerres.aacrjournals.org Downloaded from

APPLBMANet al.—Catatase in Normal and Cancerous Rats 505
1. Liver catalase activity decrease is moresevere in the case of Jensen rat sarcoma than in theUCLA fibrosarcoma. In many cases the UCLAfibrosarcoma grew to the same size as the Jensensarcoma in the same time interval, with the livercatalase activity and erythrocyte volume closelyapproaching that of normal rats.
2. In our experience with either tumor, livercatalase has not been below 20 per cent of the normal value. This is a less drastic effect than hasbeen described in the literature.
8. Kidney and erythrocyte catalase activityshow small decreases, which may very well bewithin the experimental errors of the methods.
4. Rats with growing tumors which suffer a lossin liver catalase also show an enlargement of theliver and the spleen and an increase in blood plasma. Cancerous rats with normal liver catalase activity show normal blood volumes and normalliver and spleen size.
5. Rats kept on a protein-free diet suffer a dropin liver eatalase activity to approximately 50 percent of normal within 7-10 days from the timethey were placed on the diet, and they remain atthat, level for 53 days or longer. When these animals are returned to a normal diet, the liver catalase activity returns to a normal value within 1-3days, which is similar to the time required for recovery of tumor-bearing rats after extirpation ofthe tumor. Kidney catalase activity is slightlyhigher than normal, while erythrocyte catalaseremains normal. Rats on a protein-free diet showno changes in blood volume or liver and spleensize.
6. Evidence for the synthesis of catalase in tumor-bearing rats is shown by the UCLA fibrosarcoma by the constant erythrocyte volume maintained, with increasing total weight of the animal.The suggestion is made that the synthetic mechanism for liver catalase is probably not disturbed.The reduction in the catalase activity may be dueto either a more rapid destruction of liver catalase
or the abstraction from the blood stream of somenecessary constituent by the tumor.
REFERENCES1. BRAHN,B. Further Researches on the Enzymes in the
Liver of the Cancerous. Sitzungsb. Prcuss. Akad. Wis-sensch. 20:478-81, 1916.
2. DUNN, M. S.; FEAVER,E. R.; and MURPHY,E. A. TheAmino Acid Composition of a Fibrosarcoma and ItsNormal Homologous Tissue in the Rat. Cancer Research,9:306-13, 1949.
3. GHEENSTEIN,J. P. Titration of the Liver Catalase Activityof Normal and Tumor-bearing Rats and mice. J. Nat.Cancer Inst., 2:525-30, 1942.
4. —¿�—¿�.Further Studies of Liver Catalase Activity ofTumor-bearing Animals. Ibid., 3:397-404, 194,'i.
5. GREENSTEIN,J. P., and ANDERVONT,H. B. Liver Catalase Activity of Tumor-bearing Mice and the Effectof Spontaneous Regression and of Removal of CertainTumors. J. Nat. Cancer Inst., 2:345-55, 1942.
6. —¿�—¿�.The Liver Catalase Activity of Pregnant Miceand of Mice Bearing Growing Embryonic Implants. J. Nat.Cancer Inst., 4:283-89, 1943.
7. GREENSTEIN,J. P.; ANDERVONT,H. B.; and THOMPSON,J. W. Kidney and Blood Catalase Activity of Tumor-bearing Animals. J. Nat. Cancer Inst., 2:589-94, 1942.
8. GREENSTEI.V,J. P.; JENRETTE,VV.V.; and WHITE,J. TheLiver Catalase Activity of Tumor-bearing Rats and theEffect of Extirpation of the Tumors. J. Biol. Chem., 141:327-28, 1941.
9. —¿�—¿�.The Liver Catalase Activity of Tumor-bearingRats and the Effect of Extirpation of the Tumor. J. Nat.Cancer Inst., 2:283-91, 1941.
10. MILLER,L. L. Changes in Rat Liver Enzyme Activity withInanition. Fed. Proc., 6:279, 1947.
11. . Changes in Rat Liver Enzyme Activity withAcute Inanition. J. Biol. Chem., 172:113-21, 1948.
12. NAKAHARA,W., and FUKUOKA,F. Toxohormone: ACharacteristic Toxic Substance Produced by CancerTissue. Gann, 40:45-71, 1949.
13. ROSENTHAL,E. Investigations on the Catalase Content ofthe Liver and Blood of Cancer Mice. Deutsch, med.Wchnschr., 38:2270-72, 1912.
14. SCHULTZE,M. D., and KUIKEN, K. A. The Effect ofDeficiencies in Copper and Iron on the Catalase Activityof Rat Tissues. J. Biol. Chem., 137:727-34, 1941.
15. TAYLOR,A., and POLLACK,M. S. Hemoglobin I^evel andTumor Growth. Cancer Research, 2:223-27, 1942.
16. WEIL-MALIIERBE,H., and SCHAUE,R. Studies on theLiver Catalase of Normal and Cancerous Rats. Biochem.J., 43:118-25, 1948.
Research. on August 13, 2019. © 1950 American Association for Cancercancerres.aacrjournals.org Downloaded from

1950;10:498-505. Cancer Res David Appleman, Edwin R. Skavinski and Abraham M. Stein Catalase Studies on Normal and Cancerous Rats
Updated version
http://cancerres.aacrjournals.org/content/10/8/498
Access the most recent version of this article at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
Subscriptions
Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications
Permissions
Rightslink site. Click on "Request Permissions" which will take you to the Copyright Clearance Center's (CCC)
.http://cancerres.aacrjournals.org/content/10/8/498To request permission to re-use all or part of this article, use this link
Research. on August 13, 2019. © 1950 American Association for Cancercancerres.aacrjournals.org Downloaded from