Embed Size (px)
Citation preview

CARBONIC ANHYDRASE IN NORMAL AND NEOPLASTIC GASTROINTESTINAL TISSUESWith special emphasis on isoenzymes I, II, IX, XII, and XIV
ANTTIKIVELÄ
Department of Anatomyand Cell Biology,
University of Oulu
OULU 2003


ANTTI KIVELÄ
CARBONIC ANHYDRASE IN NORMAL AND NEOPLASTIC GASTROINTESTINAL TISSUESWith special emphasis on isoenzymes I, II, IX, XII, and XIV
Academic Dissertation to be presented with the assent ofthe Faculty of Medicine, University of Oulu, for publicdiscussion in the Auditorium of the Department ofAnatomy and Cell Biology, on June 13th, 2003, at 12noon.
OULUN YLIOPISTO, OULU 2003

Copyright © 2003University of Oulu, 2003
Supervised byProfessor Seppo ParkkilaProfessor Hannu Rajaniemi
Reviewed byProfessor Kai KailaDocent Anne Kallioniemi
ISBN 951-42-7001-0 (URL: http://herkules.oulu.fi/isbn9514270010/)
ALSO AVAILABLE IN PRINTED FORMATActa Univ. Oul. D 720, 2003ISBN 951-42-7000-2ISSN 0355-3221 (URL: http://herkules.oulu.fi/issn03553221/)
OULU UNIVERSITY PRESSOULU 2003

Kivelä, Antti, Carbonic anhydrase in normal and neoplastic gastrointestinal tissues.With special emphasis on isoenzymes I, II, IX, XII, and XIVDepartment of Anatomy and Cell Biology, University of Oulu, P.O.Box 5000, FIN-90014University of Oulu, Finland Oulu, Finland2003
Abstract
The carbonic anhydrases (CAs) catalyse the hydration of CO2 to bicarbonate at physiological pH.This chemical interconversion is crucial since HCO3
- is the substrate for several biosyntheticreactions. Carbonic anhydrases are involved in many physiological processes connected withrespiration and transport of CO2/bicarbonate between metabolising tissues and the lungs, pHhomeostasis and electrolyte secretion in a variety of tissues/organs. The present work was undertakento study the distribution and expression of CA isoenzymes in the normal and neoplasticgastrointestinal tissues.
The expression of CA I, II, IX and XII in the human intestine and colorectal tumours wasinvestigated by immunohistochemistry and western blotting. In the present study,immunohistochemical methods were also used to examine the location of CA IX and XII in thehuman pancreas and pancreatic tumours. The expression of CA XIV in the murine liver and intestinewas studied using immunostaining and northern blotting.
The present results suggest that transmembrane CA XII is absent from the small intestine, but isexpressed in all segments of the normal large intestine. The positive signal for CA XII was confinedto the basolateral plasma membranes of the epithelial cells of the surface epithelial cuff. In tumours,the signal for CA XII became stronger in the deep part of the lesion. The intensity of theimmunostaining for CA I and II was clearly found to decrease in benign lesions and became veryweak in malignant colorectal tumours. The reciprocal pattern of expression observed for membrane-associated (CA IX and XII) and cytoplasmic (CA I and II) isoenzymes in intestinal samples suggeststhat CA IX and XII may be functionally involved in tumour progression to malignancy and/or ininvasion. CA I and II, which are thought to play important physiological roles in the normal colorectalmucosa, may not be required for growth of colorectal cancers and their expression consistentlydiminishes with progression to malignancy.
In the human pancreas CA IX and XII appeared to be sporadically expressed in the basolateralplasma membrane of the normal acinar and ductal epithelium. The increased expression of CA IX inhyperplastic ductal epithelium may contribute to the pancreatic tumourigenesis.
CA XIV was expressed in the hepatocyte plasma membrane and its localization on both apical andbasolateral membrane domains suggests an important role for this isoenzyme in the regulation of ionand pH homeostasis in the liver.
Keywords: bicarbonate, carcinogenesis, colon, colorectal, epithelium, hepatocyte, ion,liver, pancreas, pH, plasma membrane


Dedicated to my family


Acknowledgements
This work was carried out at the Department of Anatomy and Cell Biology, University of Oulu during the years 1999-2003.
The deepest gratitude I owe to “the man at the helm”, my mentor and principal supervisor Professor Seppo Parkkila, M.D., Ph.D. He has with great dedication put himself into this project by inspiring and always friendly tutoring me at the decisive moments, especially when things were not going as originally planned. The door of his room has always been open and numerous are the hours he has spent teaching me scientific work, writing and reasoning. I thank him for sharing his vast knowledge with me, and most of all I am thankful for his trust and friendship.
I am truly grateful to my second supervisor, Professor Hannu Rajaniemi, M.D., Ph.D., Head of the Department of Anatomy and Cell Biology, for providing me with the opportunity to work with this subject in his department. His encouraging attitude and valuable advice, both on practical and theoretical matters have helped my work.
I thank the official examiners of this thesis, Professor Kai Kaila, Ph.D., and Docent Anne Kallioniemi, M.D., Ph.D., for their criticism and comments on this thesis.
Docent Tuomo J. Karttunen, M.D., Ph.D., deserves my warmest thanks for patience in guiding me in the world of pathology and his expertise in microscopy sessions. I express my respect and gratitude to Dr. Juha Saarnio, M.D., Ph.D., who took me to the operating theatre and linked our research to real patients. His great sense of humor and broad understanding of various aspects of human life have created a good fellowship. I would also like to thank Professor Abdul Waheed, Ph.D., for his contribution to the original papers and critical reading of the manuscript of this thesis. I am much indebted to Dr. Silvia Pastoreková, Ph.D., and Professor Jaromir Pastorek, Ph.D., for the opportunity to work under their guidance in their laboratory at the Department of Molecular Biology, Institute of Virology, Slovak Academy of Sciences, Bratislava, Slovak Republic in the summer of 2001. I regard it as a great privilege to have had an opportunity to work with all these people with such an enormous expertise.
I want to extend my thanks also to other co-authors, Dr. Anna-Kaisa Parkkila, M.D., Ph.D., Professor William S. Sly, M.D., Dr. Jeffrey H. Grubb, B.A., Professor Gul Shah, Ph.D., Dr. Özlem Türeci, M.D., Professor Kari Kaunisto, M.D., Ph.D., and Dr. Jukka Hakkola, M.D., Ph.D., for their invaluable contribution to the original papers.

I would like to thank all the staff of the Department of Anatomy and Cell Biology, especially Ms Lissu Hukkanen for her dedication to her work and her prompt and friendly collaboration. The language of this thesis and the original papers was reviewed by Mr Malcolm Hicks, M.A., for which I am thankful. I also wish to thank all my good friends outside the lab for many moments shared during the past years.
Financial support for this work was provided by the Häme Fund of the Finnish Cultural Foundation and the Finnish Medical Society Duodecim, which are gratefully acknowledged. Permission to reproduce the original articles was kindly granted by American Society for Investigative Pathology, Plenum Publishing Corporation, Springer-Verlag and BioMed Central Ltd.
Finally, I am deeply grateful to my father Dr. Jyrki Kivelä, D.D.S., Ph.D., for inspiring and encouraging me. You have always been available and you have given the most valuable criticism on all work I have done. I thank my mother Lilli Kivelä, and sisters Eeva and Sigrid. Your unfailing optimism, energy and love have given me great support.
Oulu, March 2003

Abbreviations
AE anion exchanger APC adenomatous polyposis coli AQP aquaporin BSA bovine serum albumin CA carbonic anhydrase CA carbonic anhydrase gene CA-RP carbonic anhydrase-related protein cDNA complementary deoxyribonucleic acid CFTR cystic fibrosis transmembrane conductance regulator CHO chinese hamster ovary COS simian fibroblast cells DAB 3,3'diaminobenzidine tetrahydrochloride DCC deleted in colon cancer DRA proteins downregulated in adenomas ENaC epithelial Na+ channel GAPDH glyceraldehyde-3-phosphate dehydrogenase GI Gastrointestinal GPI glycosylphosphatidylinositol HCA human carbonic anhydrase H&E hematoxylin-eosin HIF-1 hypoxia-inducible factor-1 HNSCC head and neck squamous cell carcinoma IgG immunoglobulin G KCC1 KCl cotransporter kDa kiloDalton MCA mouse carbonic anhydrase mRNA messenger ribonucleic acid NHE Na+/H+ exchanger NHERF Na+/H+ exchange regulatory factor nonO non-POU (Pit-Oct-Unc) domain-containing octamer-binding protein p54nrb nuclear RNA-binding protein, 54 kDa

PAGE polyacrylamide gel electrophoresis PBS phosphate-buffered saline PNGase F endoglycosidase F PVDF polyvinylidene fluoride pVHL von Hippel-Lindau protein RCA rat carbonic anhydrase RCC renal cell cancer RPTP receptor-type protein tyrosine phosphatase RT-PCR reverse transcriptase-polymerase chain reaction SD standard deviation SCFA short-chain fatty acids SDS sodium dodecyl sulphate SEM standard error of mean TBST Tris-buffered saline with Tween-20 VHL von Hippel-Lindau VHL von Hippel-Lindau gene

List of original publications
This thesis is based on the following original articles, which are referred to in the text by Roman numerals I-IV:
I Kivelä A, Parkkila S, Saarnio J, Karttunen TJ, Kivelä J, Parkkila A-K, Waheed A, Sly WS, Grubb JH, Shah G, Türeci Ö & Rajaniemi H (2000) Expression of a novel transmembrane carbonic anhydrase isozyme XII in normal human gut and colorectal tumours. Am J Pathol 156:577-584.
II Kivelä AJ, Saarnio J, Karttunen TJ, Kivelä J, Parkkila A-K, Pastoreková S, Pastorek J, Waheed A, Sly WS, Parkkila S & Rajaniemi H (2001) Differential expression of cytoplasmic carbonic anhydrases, CA I and II, and membrane-associated isozymes, CA IX and XII, in normal mucosa of large intestine and in colorectal tumors. Dig Dis Sci 46:2179-2186.
III Kivelä AJ, Parkkila S, Saarnio J, Karttunen TJ, Kivelä J, Parkkila A-K, Pastoreková S, Pastorek J, Waheed A, Sly WS & Rajaniemi H (2000) Expression of transmembrane carbonic anhydrase isozymes IX and XII in normal human pancreas and pancreatic tumors. Histochem Cell Biol 114:197-204.
IV Parkkila S, Kivelä AJ, Kaunisto K, Parkkila A-K, Hakkola J, Rajaniemi H, Waheed A & Sly WS (2002) The plasma membrane carbonic anhydrase in murine hepatocytes identified as isozyme XIV. BMC Gastroenterol 2:13.


Contents
Abstract Acknowledgements Abbreviations List of original publications 1 Introduction ...................................................................................................................15 2 Review of the literature .................................................................................................16
2.1 Carbonic anhydrases...............................................................................................16 2.1.1 General aspects ................................................................................................16 2.1.2 Membrane-associated carbonic anhydrases.....................................................19 2.1.3 Other carbonic anhydrases...............................................................................22 2.1.4 Carbonic anhydrase-related proteins ...............................................................25
2.2 Functions of carbonic anhydrases in the alimentary tract.......................................27 2.2.1 Carbonic anhydrases in the major digestive glands.........................................27 2.2.2 Carbonic anhydrases in the gastrointestinal canal ...........................................28
2.3 Carbonic anhydrases in neoplasia...........................................................................29 2.3.1 Carbonic anhydrases in neoplastic tissues .......................................................29 2.3.2 Regulation of CA IX and XII expression.........................................................31 2.3.3 CA IX and XII in tumour growth and spread ..................................................32
2.4 Colorectal carcinogenesis .......................................................................................33 2.4.1 General considerations ....................................................................................33 2.4.2 Morphological pathways of colorectal cancer .................................................34 2.4.3 Molecular biology of colorectal cancer ...........................................................35
3 Aims of the research ......................................................................................................37 4 Materials and methods...................................................................................................38
4.1 Tissue specimens (I-IV)..........................................................................................38 4.2 Preparation of tissues (I-IV) ...................................................................................39 4.3 Antibodies against CA isoenzymes (I-IV) ..............................................................40 4.4 Other antibodies (III) ..............................................................................................40 4.5 Immunoperoxidase and immunofluorescence staining methods (I-IV).................41 4.6 Evaluation of immunoreactivity (I-III) ...................................................................42

4.7 SDS-polyacrylamide gel electrophoresis (SDS-PAGE) and western blotting (I, IV) ...........................................................................................42 4.8 Extraction of RNA and reverse transcription-polymerase chain reaction (RT-PCR) (IV) ........................................................................................................43 4.9 Statistical methods (II, III)......................................................................................43
5 Results ...........................................................................................................................44 5.1 Expression of CA I, II, IX, XII, and XIV in normal gastrointestinal tract (I-IV) ...44 5.2 Expression of CA I, II, IX, and XII in gastrointestinal tumours (I-III)..................45 5.3 Expression of von Hippel-Lindau protein in normal gastrointestinal tract and colorectal tumours ..................................................................................................46
6 Discussion .....................................................................................................................50 6.1 Carbonic anhydrases in the gastrointestinal tract....................................................50 6.2 Carbonic anhydrases in neoplastic gastrointestinal tissues.....................................54
7 Conclusions ...................................................................................................................56 References

1 Introduction
The carbonic anhydrases (CAs) are an expanding family of zinc-containing metalloenzymes, which participate in the maintenance of pH homeostasis, ion transport and water and electrolyte balance in the human body by catalyzing the reversible reaction: CO2 + H2O HCO3
− + H+ (Maren 1967, Tashian 1992, Sly & Hu 1995a). There are three evolutionary unrelated CA gene families entitled alpha, beta and gamma. In the animal kingdom, eleven enzymatically active isoenzymes have been identified so far, and they are all of alpha type. At least some isoenzymes of the alpha-enzyme family are expressed in most organs of the mammalian body (Maren 1967, Hewett-Emmet & Tashian 1991).
CA is widely present in the gastrointestinal (GI) tract – from the mouth to the rectum. Different isoenzymes play important roles in several physiological functions such as production of saliva, gastric acid, bile, and pancreatic juice as well as in absorption of salt and water in intestine. Best characterized is the function of CA II which shows high enzymatic activity and is expressed in almost all epithelia of the GI tract. Discoveries of new CA isoenzymes has stimulated research work on this area, and especially the recent reports on plasma membrane-bound isoenzymes IX and XII have raised considerable interest since they have been reported to participate in cancer invasion and spread (Parkkila S et al. 2000a). CA IX and XII may also play a role in von Hippel-Lindau (VHL)-mediated carcinogenesis. This was indirectly supported by findings that normal von Hippel-Lindau protein down-regulates the expression of both CA IX and XII, while the mutations in the VHL gene found in renal cell carcinomas can lead to overexpression of the enzymes (Ivanov et al. 1998).
The purpose of the present study was to evaluate the distribution and expression of CA I, II, IX, XII, and XIV in the gastrointestinal tract. The expression levels of these isoenzymes were investigated in both the normal and neoplastic colorectal mucosa. New information on the expression of CAs would help not only in understanding the role of each isoenzyme but also in targeting certain cell types in forthcoming physiological and clinical investigations.

2 Review of the literature
2.1 Carbonic anhydrases
2.1.1 General aspects
Carbonic anhydrases are a group of zinc-containing metalloenzymes that catalyse the reversible hydration of carbon dioxide, CO2 + H2O <=> HCO3
− + H+. The existence of CAs was found in the late 1920’s, when a red blood cell substance catalyzing the interconversion of carbon dioxide and bicarbonate was recognized in studies on the rate of escape of carbon dioxide from haemolyzed blood (Henriques 1928). A few years later this substance was purified from red blood cells, shown to be an enzyme, and named carbonic anhydrase (Meldrum & Roughton 1932, 1933, Edsall 1968, Carter 1972). Up to now eleven enzymatically active alpha carbonic anhydrases have been identified and characterized in mammals including four cytoplasmic (CA I, CA II, CA III and CA VII), two mitochondrial (CA VA and CA VB), one secreted (CA VI), and four membrane-associated (CA IV, CA IX, CA XII and CA XIV) forms (Table 1, Fig. 2).
CAs are virtually ubiquitous in living systems, having varied functions in animal, plant and bacterial cells. The great diversity in both cellular distribution (Fig. 1) and biological functions is remarkable and the catalytic activity of CAs, found in almost all organisms, is extremely high, placing the high-activity isoenzymes among the most efficient enzymes known (Tashian 1989). In addition to its role in the regulation of pH homeostasis, CA activity facilitates biosynthetic processes which involve an early carboxylation step requiring bicarbonate. These processes include gluconeogenesis and the synthesis of certain amino acids (pyruvate carboxylase), lipogenesis (pyruvate carboxylase and acetyl CoA carboxylase), ureagenesis (carbamoyl phosphate synthetase I) and pyrimide synthetase (carbamoyl phosphate synthetase II) (Chegwidden et al. 2000). The possibility that carbonic anhydrase plays a role in substrate provision by facilitation of bicarbonate transport must also be considered, e.g. the facilitated diffusion of HCO3
−/CO2 through the cytosol and in the interstitial space (Longmuir et al. 1966).

17
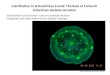
Fig. 1. Subcellular localization of the active CA isoenzymes and a novel nonclassical form of CA (nonO/p54nrb).
Fig. 2. Topography of membrane-associated CA isoenzymes. Carbonic anhydrase domain (CA), glycosylphosphatidylinositol link (GPI), transmembrane part (TM), cytoplasmic C-terminal tail (CT), proteoglycan domain (PG).
XIV
XIV
XII
XII
I
II
III
apical
lateral
basal
NonO/p54nrb
XIV
VBVA
IX
IX
VII
CA IV CA XIVCA IX CA XII
PG
CA
TM
CT CT CT
TM TM
CA CA
Plasma membrane
Cytoplasm
Extracellular spaceCA
GPI

18
Table 1. Immunohistochemical distribution of CAs in the human alimentary tract.
Isoenzyme Molecular weight (kDa)
Positive cell types Main references
CA I 30 Epithelial cells of colon
Subepithelial capillary endothelium
α-cells of Langerhans islets
Lönnerholm et al. 1985 Parkkila S et al. 1994
CA II 30 Epithelial cells of salivary glands oesophagus stomach duodenum jejunum colon biliary tract pancreatic duct
Hepatocytes
Lönnerholm et al. 1985 Parkkila S et al. 1994
CA III
30
CA IV 35 Epithelial cells of duodenum colon rectum biliary tract
Subepithelial capillary endothelium
Fleming et al. 1995 Parkkila S et al. 1996 Saarnio et al. 1998a
CA V 30 β-cells of Langerhans islets
Parkkila A-K et al. 1998
CA VI 42 Epithelial cells of salivary glands von Ebner’s glands
Parkkila S et al. 1990 Leinonen et al. 2001
CA VII
CA IX 54/58 Epithelial cells of salivary glands oesophagus stomach duodenum jejunum ileum colon rectum biliary tract pancreatic duct
Pastoreková et al. 1997 Turner et al. 1997 Saarnio et al. 1998a Ivanov et al. 2001
CA XII 44 Epithelial cells of salivary glands stomach colon pancreatic acini
Ivanov et al. 2001
CA XIV 37.5

19
2.1.2 Membrane-associated carbonic anhydrases
To date four membrane-associated carbonic anhydrase isoenzymes have been identified (Figs 1, 2). CA IV is a glycosylphosphatidylinositol-anchored CA being expressed in the apical plasma membrane of epithelial cells. The gene for CA IV is located on chromosome 17 (Okuyama et al. 1992, 1993) and the molecular weight of the human protein is 35 kDa (Zhu & Sly 1990). Human CA IV shows the most prominent immunoreaction on the apical surface of epithelial cells in the colon and is also present in the subepithelial capillary endothelium in all segments of the gastrointestinal canal (Fleming et al. 1995). It has been located in the luminal plasma membrane of the human gallbladder and bile duct epithelium (Parkkila S et al. 1996). In the liver, only the biliary epithelial cells of the intrahepatic bile ducts showed a positive signal for CA IV (Parkkila S et al. 1996). CA IV has been purified from bovine lung microsomes (Whitney & Briggle 1982), human kidney (Wistrand & Knuuttila 1989), and in a catalytically active form from human lung and kidney (Zhu & Sly 1990) and rat lung (Waheed et al. 1992a). CA IV has been located by immunohistochemistry in the plasma membranes of the proximal convoluted tubule and thick ascending limb of Henle in the rat kidney (Brown et al. 1990), endothelial cells of the choriocapillaries of the human eye (Hageman et al. 1991), endothelial cells of rat brain capillaries (Ghandour et al. 1992), and on the plasma face of endothelial cells of the pulmonary microvasculature (Fleming et al. 1993). CA IV is expressed in the skeletal muscle and also in the apical plasma membrane of the epithelial cells of the human epididymis, ductus deferens and ampulla of the ductus deferens (Waheed et al. 1992b, Parkkila S et al. 1993a). The expression of CA IV in the epididymis is regulated by androgens and oestrogen (Caflisch 1990, Caflisch & DuBose 1990, Kaunisto et al. 1998) and it is suggested to play a major role in the acidification of the epididymal fluid (Parkkila S et al. 1993a, Kaunisto et al. 1998).
CA IX was first recognized as the novel tumour-associated antigen, MN, in several human carcinomas and in normal gastric mucosa (Liao et al. 1994, Pastorek et al. 1994). When the full-length cDNA for MN protein was cloned, it was found to contain a central part with sequence homology to the CAs (Pastorek et al. 1994, Opavský et al. 1996), on which basis the MN protein was named CA IX (Hewett-Emmett and Tashian 1996). Human CA9 gene has been mapped to chromosome 17 (Ivanov et al. 1998). CA IX is a glycoprotein of 54 and 58 kDa mass expressed at the basolateral plasma membrane of the epithelial cells and, in some cases, also in the nucleus (Figs 1, 2) (Pastoreková et al. 1992). The molecule has been shown to consist of a signal peptide, proteoglycan-related sequence, CA domain, transmembrane segment, and a short intracellular tail (Pastorek et al. 1994).
Expression of CA9 gene is subject to complex regulation both via promoter/enhancer elements at the level of transcription initiation (Kaluz et al. 1999) and via VHL tumour suppressor protein, possibly at the level of transcription elongation (Chen et al. 1995, Ivanov et al. 1998). CA IX has been linked to oncogenesis, and its overexpression has been observed in malignant tumour cells. CA IX expression was found to be regulated by cell density in HeLa cells and to correlate with tumourigenicity in HeLa cell/fibroblast cell hybrids (Závada et al. 1993). Moreover, transfection of NIH3T3 fibroblasts with CA

20
IX caused changes in both morphology and growth parameters that were indicative of transformation (Pastorek et al. 1994).
CA IX was first found to be expressed in several human cancer cell lines, but not in their normal counterparts (Závada et al. 1993). Recent studies in rat and human tissues have revealed that CA IX is more widely expressed than initially recognized. High expression of CA IX has been reported in the normal epithelium of gastric, intestinal and gallbladder mucosa (Pastoreková et al. 1997; Saarnio et al. 1998a). In the intestine, CA IX is confined to the basolateral cell surface of enterocytes and the cellular distribution is restricted to the proliferative cryptal enterocytes, which is not seen with any other CA. Interestingly, its regional expression is distinctive compared with other CAs, being most intense in the duodenum and jejunum and decreasing distally to only weak and sporadic expression in the distal large intestine (Saarnio et al. 1998a). Immunohistochemical studies have also revealed a positive signal in basal layer of the oesophageal epithelium, and occasional cells in uterine cervix (Liao et al. 1994, Pastorek et al. 1994, Turner et al. 1997). In addition, CA IX has been found in the duct cells of liver and pancreas (Pastoreková et al. 1997), and in the epithelial cells of efferent ducts (Karhumaa et al. 2001). It has recently been shown that many colorectal tumours overexpress CA IX (Saarnio et al. 1998b). The co-occurrence of CA IX and Ki-67 at the site of rapid cell proliferation suggests that CA IX could be used as a biomarker of increased cell proliferation in the colorectal mucosa. Furthermore, its high expression in premalignant lesions such as adenomas suggested that it might be especially useful biomarker in early diagnosis of colorectal tumours. In hepatobiliary tumours the abnormal expression of CA IX may be linked to malignant transformation of hepatobiliary cells. This isoenzyme seems to be also a promising marker of biliary differentiation in hepatobiliary neoplasms (Saarnio et al. 2001). Some human renal cancer cell lines and renal clear cell adenocarcinomas have shown high expression of CA9 mRNA or CA IX protein (McKiernan et al. 1997, Parkkila S et al. 2000b). In addition, circulating renal carcinoma cells have been detected in the peripheral blood of patients by means of a RT-PCR assay for CA9 mRNA (McKiernan et al. 1999). Positive signal for CA IX has also been found in invasive non-small cell lung carcinoma, in situ carcinoma and microinvasive carcinoma, while pre-neoplastic lesions or normal bronchial epithelium remained negative (Vermylen et al. 1999).
Based on several independent observations, it has been suggested that CA IX may have a role in non-malignant cell proliferation and intercellular communication. First, the expression of CA IX in intestine is principally restricted to the rapidly proliferating area, crypts of Lieberkühn (Saarnio et al. 1998a). Second, its expression is regulated by cell density: CA IX is inducible in dense cultures of HeLa cells but not in rapidly growing sparse cultures (Pastoreková et al. 1992, Zavada et al. 1993, Pastoreková et al. 1997, Saarnio et al. 1998a). Third, it has been also shown to have cell adhesion properties via the proteoglycan-related domain (Zavada et al. 1997, 2000). Fourth, the recently published Car9-/- mice exhibited an interesting phenotype involving hyperplasia of gastric epithelial cells (Ortova Gut et al. 2002). CA IX was clearly required for the normal gastric morphogenesis and coordination of the dynamic homeostasis within the gastric epithelium.
Recently another active transmembrane CA isoenzyme, CA XII, which is over-expressed in some human renal cancer cells (Türeci et al. 1998) and human lung cancer

21
cells (Torczynski & Bollon 1996) has also been characterized and is broadly similar in overall structure to CA IX. The CA12 gene is mapped to chromosome 15 (Türeci et al. 1998). The cDNA sequence predicted a 354-amino acid polypeptide with a molecular mass of 39,448 Da (Ivanov et al. 1998, Türeci et al. 1998). The enzyme has features of type I membrane protein. Its sequence includes 29-amino acid signal sequence, 261-amino acid CA domain, an additional short extracellular segment, a 26-amino acid hydrophobic transmembrane domain, and a 29-amino acid C-terminal cytoplasmic tail containing two potential phosphorylation sites. The extracellular CA domain has three zinc-binding histidine residues found in active CAs and two potential sites for asparagine glycosylation (Türeci et al. 1998). It has a sequence identity of 30-42 % to other CAs. The reported molecular weight of CA XII produced in transfected COS cells is 43-44 kDa. It is reduced to 39 kDa by PNGase F digestion, which is consistent with the removal of two oligosaccharide chains (Türeci et al. 1998). Recombinant CA XII protein is an active isoenzyme and its catalytic properties are similar to those of the high-activity membrane-associated CA IV (Ulmasov et al. 2000).
CA12 mRNA has been demonstrated by northern blotting in various tissues including kidney, pancreas, colon, ovary, testis, prostate, lung, and brain (Ivanov et al. 1998, Türeci et al. 1998). Much wider tissue distribution pattern was detected using RT-PCR analysis (Türeci et al. 1998). Immunohistochemical studies have shown that CA XII is also expressed in the reproductive tissues. Positive signal for it has been detected in the surface and glandular epithelial cells of the human endometrium, occasional epithelial cells in the uterine cervix, nonciliated cells of the efferent ducts and apical mitochondria-rich cells of the epididymal duct (Karhumaa et al. 2000, 2001, Ivanov et al. 2001).
The exact function of CA XII in normal and malignant tissues is so far unknown. Türeci et al. (1998) have shown that in 10 % of patients with renal cell carcinoma the CA XII transcript was expressed at higher levels in the tumour than in the surrounding normal kidney tissue, suggesting that it is the second catalytically active membrane-associated CA isoenzyme that is overexpressed in certain cancers (Ivanov et al. 1998, 2001, Türeci et al. 1998, Parkkila S et al. 2000b). Ivanov et al. (1998) have demonstrated that, in certain renal cell carcinoma cell lines, both CA IX and CA XII are down-regulated by the product of the VHL tumour suppressor gene. Mutations in this gene are responsible for VHL disease and most sporadic renal cell carcinomas of the clear cell type (Latif et al. 1993). Indeed, the regulation of expression of CA IX and CA XII was subsequently cited as a major function of the VHL protein (Zbar et al. 1999). There is evidence that the extracellular pH of human tumours is generally more acidic than that of normal tissues (Griffiths 1991) and that this acidic pH may enhance both migratory and the invasive behaviour of some tumour types (Martinez-Zaguilan et al. 1996). Both CA IX and XII are transmembrane proteins and Ivanov et al. (1998) suggested that a high level of expression of these isoenzymes, which may be a consequence of inactivation of the VHL suppressor gene, could result in acidification of the extracellular milieu immediately surrounding certain cancer cells, creating a microenvironment conducive to tumour growth and invasion. Furthermore, the same authors mapped CA9 and CA12 gene loci to chromosomal regions (bands 17q21.2 and 15q22 respectively), which appear prone to amplification in a number of human cancers (Mitelman 1994). In addition, the expression of both CA XII and CA IX are induced under hypoxic conditions in tumours and cultured tumour cells (Ivanov et al. 2001).

22
CA XIV is a recently described transmembrane carbonic anhydrase (Mori et al. 1999), consisting of a putative amino-terminal signal sequence, a CA domain with high homology with other extracellular CAs, a transmembrane domain and a short intracellular C-terminal tail. A cDNA coding for human CA XIV has been reported and the CA14 gene has been mapped to chromosome 1q21 (Fujikawa-Adachi et al. 1999c). The northern blot method showed CA14 mRNA expression in human heart, brain, liver and skeletal muscle. RT-PCR analysis, a more sensitive technique, pointed to an intense signal in the normal human liver and spinal cord. RNA dot blot analysis showed weak signals in the small intestine, colon, kidney, and urinary bladder (Fujikawa-Adachi et al. 1999c). In situ hybridisation in mouse kidney showed that CA14 mRNA is expressed in the proximal convoluted tubule and in the outer border of the inner stripe of the outer medulla (Mori et al. 1999). Recently, Kaunisto et al. (2002) demonstrated by immunofluorescence abundant expression of CA XIV in the apical plasma membranes of the S1 and S2 segments of mouse and rat proximal tubules, and weaker staining in the basolateral membranes. Also, strong staining was observed in the initial portion of the thin descending limb of Henle. From these results they concluded that luminal CA XIV is strongly expressed in regions of the rodent nephron which have been thought to be important in urinary acidification. CA XIV is strongly expressed also in brain. Parkkila S et al. (2001) have demonstrated that CA XIV is present in the neuronal membranes and axons in different parts of the human and mouse brain. These observations suggested that CA XIV might represent the extracellular CA postulated to have an important role in modulating synaptic transmission in brain.
2.1.3 Other carbonic anhydrases
CA I is a well characterized cytoplasmic low-activity isoenzyme with a molecular weight of about 30 kDa (Bundy 1977, Lindskog et al. 1984). It is detected in erythrocytes, epithelial cells of gastrointestinal tract, both capillary and corneal endothelium, lens of the eye, islets of Langerhans, placenta and fetal membranes (Lönnerholm et al. 1985, Venta et al. 1987, Mühlhauser et al. 1994, Parkkila S et al. 1994). The CA1 gene has been cloned from a human genomic cDNA library (Tashian 1989, Butterworth et al. 1991). Interestingly, CA I is not expressed in the red cells of certain species, e.g., ruminants and felids, and no haematological abnormalities have emerged in its absence as the result of a mutation, though CA I is one of the most abundant proteins in mammalian red cells. Thus, the assignment of a physiological role in erythrocytes is problematic (Tashian et al. 1971, 1980, 1992).
The presence of cytoplasmic CA I in the human alimentary tract has been located in the surface epithelial cells of the oesophagus and colon (Lönnerholm et al. 1985, Sasaki et al. 1993, Parkkila S et al. 1994, Christie et al. 1997, Saarnio et al. 1998a), and Saarnio et al. (1998a) have demonstrated previously that it is also present in the cryptal enterocytes of the small intestine. A third major CA I-positive cell type is the capillary endothelium (Lönnerholm et al. 1985, Saarnio et al. 1998a). One previous study showed that CA I is expressed in the endocrine α-cells (Parkkila S et al. 1994). Interestingly, α-

23
cells are so far the only cells in the human alimentary tract that specifically express CA I but not CA II.
CA II, the most widely distributed member of the CA family, was originally found and purified from erythrocytes where it is involved in hydration of CO2 (Meldrum & Roughton 1932, 1933). It is expressed in epithelial cells involved in acid or alkaline secretion and in some non-epithelial cells, e.g. osteoclasts, being present in virtually every human tissue or organ (Tashian 1992). The gene for human CA II is 17 kb long and is located on chromosome 8, like those for CA I and III (Nakai et al. 1987, Tashian 1989). The high-activity, cytoplasmic isoenzyme CA II has a turnover number of 1.3-1.9 × 106 /s under physiological conditions, and thus it is one of the most efficient enzymes known (Khalifah 1971, Sanyal & Maren 1981, Wistrand 1981).
CA II has been found in several organs of the alimentary tract (Parkkila S & Parkkila A-K 1996). CA II has been suggested to participate in bicarbonate secretion in the exocrine glands of the alimentary tract. Immunohistochemical staining has shown CA II expression in the serous and ductal cells of the salivary glands (Parkkila S et al. 1990), as well as in the squamous epithelial cells of the oesophagus (Parkkila S et al. 1994). In the stomach, both the surface epithelial and parietal cells contain CA II and its main physiological function is to regulate the acidity of the gastric juice (Davenport & Fisher 1938, Davenport 1939, Kumpulainen 1984, O'Brien et al. 1977, Parkkila S et al. 1994, Parkkila S & Parkkila A-K 1996). CA II is expressed in the surface epithelial cells of the duodenum, jejunum, ileum, caecum, colon, and rectum and Brunner’s glands (Lönnerholm et al. 1985, Sasaki et al. 1993).
In the liver, results regarding the presence and physiological role of CA II have been contradictory. Spicer et al. (1982) reported that human hepatocytes are devoid of the isoenzyme, but Carter et al. (1989) and Parkkila S et al. (1994) later demonstrated its presence in rat and human hepatocytes and in the epithelium of the bile ducts. In the pancreas, immunohistochemical stainings have shown an intense positive signal for CA II in the epithelial duct cells (Kumpulainen & Jalovaara 1981, Spicer et al. 1982, Kumpulainen 1984). In renal tubular cells, the contribution of CA II to urinary acidification is well established (Wistrand 1980, Wåhlstrand & Wistrand 1980, Kumpulainen 1984, Sly & Hu 1995a,b, Lai et al. 1998). It contributes also to HCO3
− secretion by ciliary body epithelium and choroid plexus in the central nervous system (Kumpulainen et al. 1985, Parkkila A-K et al. 1994, 1995, 1997). In addition to the locations listed above, CA II is also expressed in some endocrine cells of the human pituitary and adrenal glands (Parkkila A-K et al. 1993, 1996, Sasano et al. 1994), the type II pneumocytes of the lung (Fleming et al. 1994), various cells in the tissues of the male reproductive tract (Kaunisto et al. 1990, 1995, Parkkila S et al. 1991a) and in the human placenta and foetal membranes (Mühlhauser et al. 1994). Furthermore, CA II has been suggested to take part in fatty acid and amino acid synthesis (Sly & Hu 1995a).
CA II participates in the control of bone resorption. Osteopetrosis, involving renal acidosis and cerebral calcification, is a human recessive inherited syndrome with a deficiency of CA II as a primary defect (Sly et al. 1983, 1985, Sly & Hu 1995a,b). To date, five mutations resulting in this severe but comparatively rare disease are known (Venta et al. 1991, Hu et al. 1992, Strisciuglio et al. 1998). Even though CA II is the most abundant isoenzyme in the alimentary tract, no gastrointestinal symptoms have been associated to CA II-deficient patients.

24
The cytoplasmic CA III has lowest activity of the isoenzymes and is present almost exclusively in the skeletal muscle type I fibres (Holmes 1976, Register et al. 1978), where its exact physiological function has remained undefined. Immunohistochemical staining and biochemical studies have shown that it is expressed to lesser amounts in the human and rodent liver (Jeffery et al. 1980, Carter et al. 1989, Kelly et al. 1991). Functional studies have linked it with the oxidative processes in the liver (Cabiscol & Levine 1995). Also the human uterus, urine bladder, lung, cardiac muscle (Jeffery et al. 1980), human myoepithelial cells (Väänänen & Autio-Harmainen 1987), equine thymus (Nishita & Matsushita 1989), guinea pig salivary glands and mouse colon (Spicer et al. 1990) contain CA III.
CA V is a low-activity isoenzyme located in the mitochondrial matrix (Nagao et al. 1993). The cDNA for human mitochondrial CA V has been cloned from a human liver cDNA library, and its gene has been localized to chromosome 16 (Nagao et al. 1993). Recently, two laboratories independently characterized another mitochondrial CA and thereafter the two isoenzymes have been termed CA VA and CA VB (Fujikawa-Adachi et al. 1999a, Shah et al. 2000). The expression of human CA5A mRNA has been demonstrated only in liver (Fujikawa-Adachi et al. 1999a). The CA5B mRNA has been shown in normal human heart and skeletal muscle by northern blotting and in pancreas, salivary glands, kidney, and spinal cord by reverse transcription-PCR (RT-PCR) (Fujikawa-Adachi et al. 1999a). Previous immunohistochemical and western blotting data indicated that the β-cells of Langerhans islets contain high levels of mitochondrial CA V (Parkkila A-K et al. 1998).
CA VI is to date the only known secretory isoenzyme of the CA gene family. It was initially found in the ovine parotid gland and saliva (Fernley et al. 1979), and it has recently been suggested that it may be present in serum (Kivelä et al. 1997) and secretory organs such as the lacrimal glands (Ogawa et al. 1995), pancreas (Fujikawa-Adachi et al. 1999b) and mammary gland (Karhumaa et al. 1998). CA VI was first purified from rat saliva, and its amino acid composition was determined by Feldstein and Silverman (1984). Later, it was also purified from human saliva (Kadoya et al. 1987, Murakami & Sly 1987, Leinonen et al. 2001). Fernley et al. (1988) reported the amino acid sequence of sheep CA VI, and the cDNA sequence of human CA VI has also been determined (Aldred et al. 1991). This isoenzyme with a molecular weight of 39-46 kDa has several properties that distinguish it from the well-characterized cytoplasmic isoenzymes (Feldstein & Silverman 1984, Kadoya et al. 1987, Murakami & Sly 1987, Fernley 1991a,b, Parkkila S et al. 1991b, Ogawa et al. 1992). On the other hand, the catalytic domain of CA VI is highly homologous to four other “extracellular” CAs (CA IV, CA IX, CA XII and CA XIV), which are in fact transmembrane proteins with an extracellularly exposed CA domain (Fujikawa-Adachi et al. 1999b, Mori et al. 1999).
A recent study by Leinonen et al. (1999) showed that salivary CA VI is associated with the enamel pellicle, a thin layer of proteins located between the enamel and the bacterial plaque. It has also been detected in the gastric mucus, but the failure to find any expression of it in the gastric epithelial cells (Parkkila S et al. 1994) implies that gastric CA VI must be of salivary origin. CA VI may contribute to the maintenance of the pH gradient on the surface epithelial cells. Recent studies have shown that it is also present in human and rat milk (Karhumaa et al. 2001). There were several lines of evidence to support this conclusion. The specific antibodies to salivary CA VI recognized

25
polypeptides of similar molecular mass (42 kDa) in human and rat saliva and milk, the polypeptide could be purified effectively from human milk by CA inhibitor affinity chromatography, and the partial amino acid sequence of the purified 42-kDa polypeptide was identical to that of salivary CA VI. The high concentration of CA VI in colostrum suggested that it may play an important role in the development of the gastrointestinal canal during the early postnatal period.
There are two other recent findings pointing to novel physiological roles for CA VI. First, gustin, a salivary factor involved in taste perception, was shown to be identical to CA VI (Thatcher et al. 1998), and second, identification of a novel stress-inducible intracellular form of CA VI (type B) suggested that the enzyme could also participate in intracellular pH changes induced by stress, including apoptosis (Sok et al. 1999).
CA VII is a cytoplasmic isoenzyme, the gene for which has been isolated from a human genomic library (Montgomery et al. 1991, Tashian 1992). The gene is about 10 kb long and located on chromosome 16, and the predicted amino acid sequence of CA VII showed that it is the most highly conserved isoenzyme in mammals (Lakkis et al. 1997). Recombinant CA VII has shown high enzyme activity, but the expression of the protein in tissues has not yet been described (Earnhardt et al. 1998). Its mRNA has been detected in the human salivary gland (Montgomery et al. 1991), rat and mouse lung (Ling et al. 1994) and mouse brain neurons (Lakkis et al. 1996, Lakkis et al. 1997).
2.1.4 Carbonic anhydrase-related proteins
Mammalian CA family contains three CA-related genes (CA-RP VIII, X, and XI) and two subtypes of receptor-type protein tyrosine phosphatases (RPTP β and γ). These proteins share CA-like domain but lack CA activity (Tashian et al. 2000). CA-related genes have been found from human and mouse cDNA libraries and named carbonic anhydrase-related proteins (CA-RPs) in order of their discovery. mRNA of CA-RP8 has been detected in mouse brain and by PCR from cDNA libraries of the human salivary gland, testis, placenta and lung (Skaggs et al. 1993). CA-RP X was initially identified by comparing overlapping, partial human cDNA sequences deposited in public databases of expressed sequence tag (EST) (Hewett-Emmett & Tashian 1996). Recently, Okamoto et al. (2001) sequenced the full-length human cDNA of CA-RP10, which appeared to encode a 328 amino acid polypeptide. CA-RP X lacks two out of three histidine residues, which are required for CA catalytic activity. RNA dot blotting showed positive signal for CA-RP10 mRNA in almost all parts of the human central nervous system. CA-RP10 gene is most probably located on chromosome 17 (Tashian et al. 2000). cDNA for CA-RP XI was first characterized from sheep, human and murine cDNA libraries. Sheep CA-RP11 cDNA appeared to encode a secreted 328-residue protein (Lovejoy et al. 1998). The absence of all three histidine residues, known to be essential for CA activity, indicates that CA-RP XI is not an active CA isoenzyme (Lovejoy et al. 1998). CA-RP XI has several potential phosphorylation sites and binding motifs, suggesting a role in signal transduction. A single copy of the CA-RP11 gene is located on human chromosome 19q13.2-3. Northern blot analysis has revealed a strong expression of CA-RP XI in the

26
human brain. Likewise, expression of its mRNA has been detected in the normal human pancreas, kidney, liver, salivary gland and spinal cord by using RT-PCR analysis. Interestingly, the signal for CA-RP XI in pancreatic cancer cell line was stronger that that in the normal pancreas (Fujikawa-Adachi et al. 1999b).
The CA-like domain has also been demonstrated in a subfamily of receptor-type protein tyrosine phosphatases (RPTP β and γ) (Krueger & Saito 1992, Barnea et al. 1993, Levy et al. 1993). The extracellular region of RPTPβ has been shown to be identical to a chondroitin sulphate proteoglycan called phosphacan (Maurel et al. 1994), which provides a binding site for a cell surface signal transducing molecule (Peles et al. 1995). The phosphorylation of tyrosine residues plays an important role in signalling cell growth, differentiation and transformation. However, the exact functions of RPTPs are still unknown. The CA-like domains of the RPTP β showed about 30-50% amino acid identity to CAs, but because of only one concerved histidine residue, it is unlikely that RPTPs have any CA activity (Lindskog 1982, Krueger & Saito 1992).
In addition to the members of the classical CA gene family, a 66-kDa polypeptide was recently purified from several rat tissues on CA inhibitor affinity chromatography (Karhumaa et al. 2000). Its amino acid sequence revealed that it represents the previously cloned and characterized nuclear protein, nonO/p54nrb, a non-POU (Pit-Oct-Unc) domain-containing octamer-binding protein, which is homologous to the nuclear 54 kDa RNA-binding protein (Dong et al. 1993, Yang et al. 1993). CA activity of nonO is interesting, because no other class of mammalian proteins except CAs have been shown to bind specifically to CA inhibitor affinity chromatography matrix and to contain CA catalytic activity. It is noteworthy that its catalytic activity is higher than determined for CA III (Jeffery & Carter 1980). There was significant immunological cross-reactivity between CA II and nonO. The predicted amino acid sequence of nonO shares no structural elements required for conventional CA activity. It lacks all conserved histidines, which are involved in zinc binding and have been considered essential for CA activity (Kannan et al. 1977, Eriksson & Liljas 1991). It neither showed structural similarity with β or γCAs (Dr. R.E. Tashian, personal communication). Since CA activity and zinc binding capacity of CA II is retained when His119 is substituted with glutamine (Lesburg et al. 1997), a polyglutamine stretch, Q29-Q38 in the nonO protein is a potential site for Zn binding, and could also be a potential site for the CA activity of nonO. However, tertiary structure of nonO could give more information of the CA catalytic site in nonO. CA catalytic activity in nonO explains at least part of the nuclear CA activity seen in histochemical CA stainings (Karhumaa et al. 2000). It also suggests that CA activity is important factor in transcriptional regulation, which would be a novel function for CA.

27
2.2 Functions of carbonic anhydrases in the alimentary tract
2.2.1 Carbonic anhydrases in the major digestive glands
Two CA isoenzymes, CA II and VI, are known to be expressed in the mammalian salivary glands (Kivelä et al. 1997). In the serous acinar cells of the parotid and submandibular glands CA II has been suggested to participate in the production of bicarbonate into the saliva (Parkkila S et al. 1990, 1991b, 1994, Ogawa et al. 1993). CA VI is a secretory isoenzyme produced by the serous acinar and demilune cells of the parotid and submandibular glands (Parkkila S et al. 1990). CA VI and II may together form a complementary system regulating the acid-base balance in the mouth and upper alimentary tract (Parkkila S et al. 1990, 1996, Kivelä et al. 1997). CA II in the salivary glands may supply the saliva with HCO3
− and the CA VI secreted into the saliva would then accelerate the removal of bacterially produced acid in the mouth in the form of CO2. CA VI may protect teeth by catalyzing the bicarbonate-based buffering system in the oral cavity and thus accelerate the removal of acid from the local microenvironment of the tooth surface. It is located at the optimal site on dental surfaces for catalyzing the conversion of salivary bicarbonate and microbe-delivered hydrogen ions to carbon dioxide and water (Leinonen et al. 1999). This hypothetical model of CA VI function was supported by Kivelä et al. (1999), who showed that low salivary CA VI concentrations are associated with increased caries prevalence, particularly in subjects with neglected oral hygiene.
Compared with other secretory organs, the mammalian liver contains relatively low levels of total CA activity. A basic physiological function of CA II in the liver is to produce HCO3
− for the alkalization of the bile (Swenson 1991). The mammalian liver expresses high levels of mitochondrial CA V. Physiologically,
CA VA has been implicated in two metabolic processes in the mitochondria of hepatocytes: ureagenesis and gluconeogenesis, supplying bicarbonate for the first urea cycle enzyme, carbamyl phosphate synthetase I in ureagenesis and for pyruvate carboxylase in gluconeogenesis (Dodgson 1991). CA inhibitors have been observed to retard both of these processes in the livers of guinea pigs and rats (Dodgson et al. 1983, Metcalfe et al. 1985, Dodgson 1991). CA V is the first CA isoenzyme to be found to participate in intermediary metabolism, but it is conceivable that it may also have other functions, as it is contained in both periportal and perivenous hepatocytes, while urea and glucose synthesis occur only in the periportal region.
The presence of low activity, hormonally regulated CA III in hepatocytes has aroused interest in its specific function. Cabiscol & Levine (1995) have demonstrated that it functions in an oxidizing environment and that it is the most oxidatively modified protein in the liver known so far. These and other recent results have suggested that CA III may provide protection from oxidative damage and CA III may serve as a useful marker protein to investigate in vivo the mechanisms, which contribute to oxidative damage in the liver. It has also been suggested that lower levels of free radicals in cells overexpressing CA III may affect growth signalling pathways (Räisänen et al. 1999).

28
Earlier studies have indicated that both CA IV and CA IX are expressed in the biliary epithelial cells, whereas hepatocytes are devoid of immunoreactivity for these isoenzymes (Parkkila S et al. 1996, Pastoreková et al. 1997). Interestingly, previous histochemical studies have indicated CA enzymatic activity also in the hepatocyte plasma membrane (Ridderstråle et al. 2000). CA XIV could be the most promising candidate protein to represent the hepatocyte isoenzyme, since its mRNA has recently been reported in both mouse and human liver (Mori et al. 1999, Fujikawa-Adachi et al. 1999c).
In pancreas, the role of CA II in the secretion of bicarbonate into the pancreatic juice by the epithelial duct cells is well documented (Swenson 1991). CA I is expressed in the α-cells of the endocrine Langerhans islets (Parkkila S et al. 1994). However, the physiological role of CA I in α-cell function has remained unclear. CA V is the second isoenzyme described in the endocrine pancreas where its expression is solely confined to the β-cells (Parkkila A-K et al. 1998). The suggestion that CA V may have a role in the regulation of insulin secretion was based on its cellular distribution and the observation that the CA inhibitor acetazolamide inhibited glucose-stimulated insulin secretion (Parkkila A-K et al. 1998).
2.2.2 Carbonic anhydrases in the gastrointestinal canal
CA II has been located in the squamous epithelial cells of the oesophagus, where it may contribute to HCO3
− secretion (Meyers & Orlando 1992, Parkkila S et al. 1994, Christie et al. 1997). The presence of this isoenzyme in the oesophagus is physiologically important, because endogenous HCO3
− secretion is capable of raising the pH of the gastro-oesophageal reflux-derived residual acid from 2.5 almost to neutrality. The immunohistochemical evidence for the presence of CA II in the human oesophagus is thus in accordance with the biochemical evidence that the oesophagus disposes of an endogenous mechanism for protecting the mucosa against acidity, but suggests that it is the stratified oesophageal epithelium rather than the submucous glands that is responsible for HCO3
− secretion. Immunohistochemical techniques have revealed cytosolic CA II in the parietal cells of
the gastric glands, where it regulates the acidity of the gastric juice by proton secretion (Davenport & Fisher 1938, Davenport 1939, O'Brien et al. 1977, Sato et al. 1980, Lönnerholm et al. 1985, Parkkila S et al. 1994, Parkkila S & Parkkila A-K 1996). On the other hand, in the gastric surface epithelial cells CA II is involved in the secretion of mucus and HCO3
− to form a bicarbonate containing mucous gel layer covering the epithelium and protecting it from digestion. This gastroduodenal HCO3
− secreted by the surface epithelial cells neutralizes the gastric acid (Richardson 1985).
Membrane-associated CA IX is another major CA isoenzyme expressed in the gastric epithelium. Both parietal and surface epithelial cells contain CA IX at the basolateral plasma membrane (Pastoreková et al. 1997). Evolutionary conservation in vertebrates and the abundant expression of CA IX in the normal human gastric mucosa have indicated its physiological importance. CA IX may participate in physiological processes via the activity of its CA-like domain. On the other hand, basolateral localization of CA

29
IX suggests its possible involvement in intercellular communication and/or cell proliferation.
It is widely known that both CA I and II are expressed in the non-goblet epithelial cells of the mammalian colon (Lönnerholm et al. 1985, Parkkila S et al. 1994), in which these isoenzymes have been implicated in the regulation of electroneutral NaCl reabsorption via the synchronous operation of apical Na+-H+ and Cl−-HCO3
− exchange (Charney & Egnor 1989). In addition to cytosolic CA I and II, the intestinal enterocytes express at least two membrane-associated isoenzymes, CA IV (Fleming et al. 1995) and IX (Pastoreková et al. 1997, Saarnio et al. 1998a). The location of CA IV in the apical brush border of the colonic epithelium has suggested a functional role in the regulation of colonic ion homeostasis (Fleming et al. 1995). Recent studies have demonstrated that the distribution of CA IX in the gut has several unique features when compared to the other CA isoenzymes (Pastoreková et al. 1997, Saarnio et al. 1998a). One of them was its localization on the basolateral surfaces of proliferating cryptal enterocytes, suggesting that it may serve as a ligand or receptor for another protein that regulates intercellular communication or cell proliferation. Furthermore, CA IX has a completely conserved active site domain of CAs (Opavský et al. 1996), suggesting that it could also participate in CO2/ HCO3
− homeostasis in the colon. The most novel CA isoenzymes, CA XII and XIV, may also be expressed in the gut.
Positive signals have been obtained in the human colon using mRNA blots (Ivanov et al. 1998, Türeci et al. 1998, Fujikawa-Adachi et al. 1999c). However, their cellular localisation was not reported in the gastrointestinal tract prior to the completion of the present study.
2.3 Carbonic anhydrases in neoplasia
2.3.1 Carbonic anhydrases in neoplastic tissues
In view of the experimental data available, it seems that tumour-associated CA isoenzymes may play a role, as yet undefined, in cell proliferation and oncogenesis. Studies show an important causal link between hypoxia, extracellular acidification, and induction or enhanced expression of these enzymes in human tumours (Ivanov et al. 2001).
CA IX is a predominant isoenzyme in various tumours including those arising from the GI tract. It has previously been shown that many colorectal tumours overexpress CA IX (Saarnio et al. 1998b). The co-occurrence of CA IX and Ki-67 at the site of rapid cell proliferation indicates that CA IX could be used as a biomarker of increased cell proliferation in the colorectal mucosa. Furthermore, its high expression in premalignant lesions such as adenomas suggests that it might be especially useful biomarker in early diagnosis of colorectal tumours.
The expression and localisation of CA IX has been examined also in head and neck squamous cell carcinoma (HNSCC) (Beasley et al. 2001). The enzyme was related to the

30
location of tumour microvessels, angiogenesis, necrosis and stage, and was considered a potential target for future therapy in HNSCC (Beasley et al. 2001). In another study, Turner et al. (1997) suggested that the tumour-associated CA IX may play a role in proliferation and regeneration in esophageal squamous epithelium, and loss of its expression may be related to cancer progression in Barrett's-associated adenocarcinomas.
Recently, Saarnio et al. (2001) showed that in the biliary epithelial tumours, immunostaining for CA IX was mainly localised at the basolateral surface of the epithelial cells, as in the normal mucosa (Pastoreková et al. 1997). The presence of CA IX in neoplastic hepatobiliary cells and its absence in hepatocellular carcinomas suggested that this isoenzyme could be used as a marker for biliary differentiation in hepatobiliary neoplasms (Saarnio et al. 2001).
Some human renal cancer cell lines and RCCs have shown high expression of CA9 mRNA or CA IX protein (McKiernan et al. 1997, Parkkila S et al. 2000b). The enhanced RT-PCR assay for CA9 may prove useful in the diagnosis and monitoring of RCC (McKiernan et al. 1999). Furthermore, Li et al. (2001) showed that flow cytometry might serve as a fast tool of immunocytochemical detection of renal cancer cells. These results confirmed that CA9 antigen would be an ideal marker for RCC.
The positive signal for CA IX has also been found in invasive non-small cell lung carcinoma, in situ carcinoma and microinvasive carcinoma, while pre-neoplastic lesions or normal bronchial epithelium remained negative (Vermylen et al. 1999). It has been concluded that CA IX is an important molecule in non-small cell lung cancer, the upregulation of which occurs in highly hypoxic/necrotic regions of the tumours (Giatromanolaki et al. 2001). The expression of CA IX is linked to the expression of a constellation of proteins involved in angiogenesis, apoptosis inhibition and cell-cell adhesion disruption, which explains the strong association of this enzyme with poor clinical outcome (Giatromanolaki et al. 2001).
In invasive breast carcinoma, the expression of CA IX has been associated with worse relapse-free survival and overall survival (Chia et al. 2001). A significant association with necrosis also indicated the potential role of CA IX as a marker of hypoxia within breast carcinomas. A recent study indicated that ectopic activation of CA9 gene may be implicated in breast carcinogenesis and also suggested potential use of CA IX as a breast tumour marker (Bartošová et al. 2002).
There is evidence to suggest that hypoxia-regulated gene expression influences tumour aggressiveness, contributing to the poorer outcome of patients with hypoxic tumours. Loncaster et al. (2001) showed clinical evidence that CA IX expression is up-regulated in hypoxic human cervical tumours and is associated with a poor prognosis. It may have a role as an intrinsic marker of tumour hypoxia and poor outcome after radiation therapy and the level of CA IX expression may be used to aid in the selection of patients who would benefit most from hypoxia-modification therapies or bio-reductive drugs (Loncaster et al. 2001).
Irrespective of its function, CA IX attracts a significant interest due to the strong association with neoplasms and absence in corresponding normal tissues, suggesting a potential to serve as a tumour marker. CA IX has also been considered as an excellent target for immunotherapy in renal cell carcinoma (Steffens et al. 1997, Uemura et al. 1999). The discovery of this tumour-associated protein with a central CA domain

31
(Pastoreková et al. 1992) suggests the possibility of an additional or alternative mechanism by which specific CA inhibitors may inhibit tumour cell growth.
Carbonic anhydrase XII, broadly similar in overall structure to CA IX (excluding the proteoglycan like domain of CA IX), is also over-expressed in some human renal cancer cells (Ivanov et al. 1998, Türeci et al. 1998, Parkkila S et al. 2000b) and human lung cancer cells (Torczynski & Bollon 1996). The high expression of CA XII in some selected cancer cell lines suggested that it could serve as a useful biomarker of some malignant tumours and could also be considered as a potential target for novel therapeutic applications (Ivanov et al. 1998).
In the previous literature, there are only two reports focused on the immunohistochemical distribution of CA I or II in colorectal tumours. Gramlich et al. (1990) demonstrated that adenomas and adenocarcinomas failed to stain for carbonic anhydrase I and II. Later, Mori et al. (1993) showed that only minority of colorectal cancers express CA I, and those tumours may have a more favorable outcome than CA I-negative tumours.
CA I has been localised in the cells expressing glucagon in malignant endocrine tissue (Parkkila S et al. 1995). In addition, CA II has been detected in the neoplastic ductal epithelium in the exocrine pancreatic tissue. The expression rate of CA II in the ductal cells, sustained after malignant transformation, did not correlate with the malignancy of the tumours, suggesting a limited value in diagnostic evaluation of pancreatic carcinoma (Parkkila S et al. 1995). CA II is also expressed in astrocytic tumours, oligodendrogliomas, ependymal and choroid plexus tumours and tumours of nerve sheath cell origin (Parkkila A-K et al. 1995). It has been suggested that some tumours contain abundant CA II, which might leak into the CSF (Parkkila A-K et al. 1995).
Recently, Leppilampi et al. (2002) investigated the presence of CA isoenzymes in malignant hematopoietic cell lines and malignant blast cells of bone marrow samples. Three out of six malignant hematopoietic cell lines expressed CA II, whereas no expression was detected for CA I, IX or XII. Positive reactions were found in 62% of acute myeloid leukaemia samples, 73% of acute lymphoblastic leukemia, and 50% of chronic myelomonocytic leukemia. The results indicated that CA II expression is not restricted to one cell lineage only but may result from a genetic aberration that occurs in both myeloid and lymphatic lineages or in their progenitor cell.
2.3.2 Regulation of CA IX and XII expression
In 1998, Ivanov and his group (Ivanov et al. 1998) used renal cell carcinoma cell lines stably transfected with wild-type VHL-expressing transgenes to discover genes involved in VHL-mediated carcinogenesis. The involvement of CA IX and XII was indirectly supported by findings that normal von Hippel-Lindau protein down-regulates the expression of both CA IX and XII, while the mutations in the VHL gene found in renal cell carcinomas can lead to overexpression of the isoenzymes.
The VHL gene is a tumour suppressor gene. This means that its role in a normal cell is to stop uncontrolled growth and proliferation. If the gene is deleted or mutated, its

32
inhibitory effect on cell growth is consequently lost or diminished, which, in combination with defects in other regulatory proteins, can lead to cancerous growth. VHL seems to act as a 'gatekeeper' to the multistep process of tumourigenesis. The molecular mechanisms by which product of VHL (pVHL) modulates the expression of target genes is not well understood. The original hypothesis based on the discovery of elongin B binding by pVHL assumed that VHL could negatively regulate transcription elongation of target genes, CA9 and CA12, by inhibiting the elonging function. Although this expectation has been confirmed in vitro, there is no compelling evidence to date that pVHL can exert the same effect in vivo (Duan et al. 1995, Ivanov et al. 1998, Zbar et al. 1999).
The transcriptional complex hypoxia-inducible factor-1 (HIF-1) has emerged as an important mediator of gene expression patterns in tumours. This transcription factor regulates gene induction in response to hypoxia (Wenger & Gassmann 1997). HIF-1 is regulated by ubiquitin-mediated proteolysis and is targeted for destruction by the pVHL in normoxia and stabilized under hypoxia (Maxwell et al. 1999).
Both CA9 and CA12 are strongly induced by hypoxia in a range of tumour cell lines. Wykoff et al. (2000) showed that in renal carcinoma cells, defective for the VHL tumour suppressor, up-regulation of these CAs is associated with loss of regulation by hypoxia, consistent with the critical function of pVHL in the regulation of HIF-1. Furthermore, Wykoff et al. (2000) defined a HIF-1-dependent hypoxia response element in the minimal promoter of CA9 and demonstrated that tight regulation by the HIF/pVHL system was reflected in the pattern of CA IX expression within tumours. In comparison with vascular endothelial growth factor (VEGF) mRNA, expression of CA IX demonstrated a similar pattern of expression around regions of necrosis (Wykoff et al. 2000). CA9 and CA12 define a new class of HIF-1-responsive gene, the activation of which has implications for the understanding of hypoxic tumour metabolism and which may provide endogenous markers for tumour hypoxia (Wykoff et al. 2000).
2.3.3 CA IX and XII in tumour growth and spread
Both CA IX and CA XII are transmembrane proteins with catalytic domain on the cell exterior, suggesting that they might participate in acid-base regulation of the extracellular space. There is substantial evidence that extracellular pH of human tumours is generally more acidic than that of normal tissues (Griffiths 1991) and that this acidic pH may enhance both the migration and the invasive behaviour of some tumour types (Martinez-Zaguilan et al. 1996). Acidification of the extracellular milieu of malignant tumours is reported to increase the invasive behavior of cancer cells. Parkkila S et al. (2000a) investigated the functional role of CA activity in cancer cells by analyzing the effect of acetazolamide, a potent CA inhibitor, on the invasive capacity of renal carcinoma cell lines. The results clearly showed that acetazolamide alone reduced invasiveness of these cancer cells in vitro and suggests that the CAs expressed in these renal cancer cells contribute to invasiveness. CA inhibitors may also reduce invasiveness in other tumours that overexpress one or more CAs (Parkkila S et al. 2000a).

33
The mechanism by which the CA inhibitors inhibit tumour growth is not known at present, but several hypotheses have been proposed. These compounds may reduce the provision of bicarbonate for the synthesis of nucleotides and other cell components. On the other hand, CA inhibitors could prevent the acidification of intracellular milieu. A combination of several mechanisms is also possible. E7070, a member of recently reported class of antitumour sulfonamides, blocks cell cycle progression in the G1 phase. It has been suggested that E7070, possessing a free SO2NH2 moiety, probably acts as a strong CA inhibitor. This compound demonstrates significant antitumour activity both in vitro and in vivo against different human tumours, e.g. human colon carcinoma. E7070 produces not only growth suppression but also reduction in tumour size. Presently, E7070 is in Phase II clinical trials (Casini et al. 2002.)
2.4 Colorectal carcinogenesis
2.4.1 General considerations
Colorectal malignancies continue to be one of the most frequent and life-threatening diseases throughout the world, especially in well-developed and industrialized countries (Weisburger & Wynder 1987), representing one of the main causes of mortality in all Western countries (Levi et al. 1994). Adenomas and carcinomas of the large intestine show basically the same risk profile. The development of colorectal adenomas has been directly related to the consumption of meat and animal fat, and to low physical activity. In contrast, vegetables, fruit, fibre and several micronutrients (such as calcium, folate and antioxidant vitamins) seem to have protective effect (Kampman et al. 1994, Potter et al. 1999). Other risk factors are age, prior colorectal cancer, ulcerative colitis and Crohn disease, heriditary polyposis and colorectal cancer syndromes, and genetic factor. The importance of environmental factors in the pathogenesis of colorectal cancer is emphasized by the high incidence of the disease in industrialized countries and among emigrants from low-risk to high-risk regions (Hamilton et al. 1998).
Most of malignant colorectal tumours are adenocarcinomas (96%) arising from the columnar surface epithelium, and, in many cases (10-20%), may show a mucinous component. These tumours encompass a large variety of precancerous and preneoplastic lesions, many of which can easily be removed at endoscopy. Other malignant colorectal tumours are carcinoid tumours and large bowel lymphoma. The colorectal neoplasms might be prevented by interfering with the various steps of carcinogenesis, which begins with uncontrolled cell replication, continues with the formation of adenomas of various dimensions, and eventually evolves into malignancy (Ponz de Leon & Di Gregorio 2001).

34
2.4.2 Morphological pathways of colorectal cancer
Adenomas are well demarcated lumps of epithelial dysplasia which can be classified into three major histological types: tubular, villous and tubulovillous. An adenoma can be considered “malignant” when there is evidence that neoplastic cells pass through the muscularis mucosae and infiltrate the submucosa (Morson et al. 1983); in this case, the definition of “carcinoma developed in adenoma” is appropriate. After crossing the submucosa, the local invasion of neoplastic cells and potential for metastasis is possible. Adenomas show different grades of dysplasia, degree of severity, and these abnormalities can be graded into “mild”, moderate” and “severe” dysplasia (Morson & Dawson 1990). Colorectal carcinoma can be graded into well-differentiated, moderately-differentiated and poorly-differentiated lesions (Bosman 1995).
As already emphasized in the previous chapter, carcinogenesis is known to be a multistep process (Faeron & Volgenstein 1990) with both genetic (Faeron & Volgenstein 1990, Kuismanen 1999) and environmental components (Ames et al. 1995, Ames & Gold 1998, de Kok et al. 1999). The adenoma-carcinoma sequence refers to a traditional view that colorectal malignancies develop from adenomatous polyps and, indeed, a large amount of evidence has been accumulated indicating that carcinomas of the large intestine arise – in most if not all cases – from pre-existing, premalignant lesions, especially when associated with dysplastic changes (Morson et al. 1983).
First, histological aspects of adenomas are frequently observed in specimens of colorectal carcinoma (Kyzer et al. 1992, Pollard et al. 1992). Second, the accurate surveillance with systematic removal of all newly detected polyps led to a significant reduction in the expected number of invasive carcinomas (Winawer et al. 1993). Third, there is a cumulative risk of cancer at the polyp site (Stryker et al. 1987). Fourth, adenomas and malignant tumours tend to have a similar distribution in the various segments of large intestine, with a maximum frequency in the rectosigmoid region (Morson et al. 1983). Fifth, the age-specific incidence rate of colorectal polyps shows an earlier rise than cancer, with a peak of incidence that anticipates by five years that of carcinomas (Ponz de Leon et al. 1987). This pattern of incidence speaks in favour of a sequence between adenomatous polyps and cancer, since the malignant transformation of a benign adenoma usually takes years (Stryker et al. 1987). Sixth, patients with with colorectal cancer and individuals with adenomas both share a similar increased frequency of site-specific tumours among first-degree relatives (Ponz de Leon et al. 1989). Seventh, recent observations showed that the risk of developing colorectal adenomas is associated with the same environmental factors (diet and lifestyle) as those related to the risk of colorectal malignancies (Kampman et al. 1994). Finally, adenomas and carcinomas of the large intestine may show similar molecular alterations (Vogelstein et al. 1988, Boland et al. 1995). This, however, is an oversimplification, since most polyps do not acquire malignant features during their natural history, and might also regress or disappear (Waddell et al. 1995). In addition, the earliest phases of colorectal tumourigenesis presumably initiate in normal mucosa, with a disorder of cell replication and renewal (Lipkin 1974), and with the appearance of clusters of enlarged crypts (Siu et al. 1999) showing proliferative, biochemical and biomolecular abnormalities. Although several lines of evidence indicate that carcinomas usually originate from pre-existing adenomas,

35
this does not imply that all polyps undergo malignant changes, and does not exclude “de novo” carcinogenesisis – i.e., the development of cancer from flat mucosa.
Recently, distinct entity of premalignant colorectal lesions has been described and the traditional assumption of benign hyperplastic polyps and the adenoma-carcinoma sequence has been challenged. K-ras mutations and microsatellite instability have been described in hyperplastic polyps (Otori et al. 1997, Iino et al. 1999), and histogenetic continuum to serrated adenomas has been proposed (Yao et al. 1999). The malignant potential of serrated adenomas has been documented and progression to frank carcinoma has been suggested in individual cases (Mäkinen et al. 2001). In addition, recognition of polyps called flat adenomas is important, since the malignant potential seems to be considerably higher than that of common sessile or pedunculated polyps of the same size (Adachi et al. 1991).
2.4.3 Molecular biology of colorectal cancer
Many molecular abnormalities have been reported in colorectal carcinoma and it is now believed that accumulated alterations of suppressor genes and protooncogenes are required for the development of colorectal cancers (Vogelstein et al. 1988, Fearon & Vogelstein 1990). It has been estimated that a minimum of eight to ten mutational events must accumulate during multistep carcinogenesis to produce an invasive colorectal cancer (Hamilton et al. 1998).
The loss of heterozygosity in a tumour suppressor gene by deletion or somatic mutation of the remaining normal allele predisposes to tumour development. Tumour suppressor genes are increasingly being incriminated in the pathogenesis of both hereditary and spontaneous cancers in humans (Hamilton et al. 1998). Germline mutations in the adenomatous polyposis coli (APC) gene, a putative tumour suppressor gene, are responsible for familial adenomatous polyposis. Moreover, the majority of sporadic colorectal cancers contain a mutation in the same gene. It has been suggested that APC has an important role in the early development of most colorectal neoplasms. DCC gene, labelled “deleted in colon cancer”, is another putative tumour suppressor gene. It is located on chromosome 18, and is often missing in colorectal cancers. In addition, p53 tumour suppressor gene is mutated or deleted in many cases of colorectal neoplasms. Point mutations activating the ras protooncogene are an example of mutations in oncogenes. It has been suggested that chromosome 3p at the von Hippel-Lindau disease gene locus may be implicated in the development or progression of sporadic colorectal carcinoma. Zhuang et al. (1996) showed that VHL disease gene deletion frequently occurs in sporadic colon carcinoma. Because this deletion was not present in adenomas, VHL gene may play a role in colonic carcinogenesis and represent a relatively late event in colonic neoplasia progression. However, the association of pVHL in colorectal carcinogenesis has not been studied extensively. The inactivation of the von Hippel-Lindau gene causes the VHL syndrome, associated with renal cell carcinoma and hemangioblastoma of the brain. The normal VHL protein complexes with and inhibits

36
elongins B and C, two factors that activate the transcription factor elongin A (Hamilton et al. 1998).
Another major pathway of carcinogenesis involves defects in DNA mismatch repair, being present in 15%. Mutation in the group of these genes (such as MLH1 and MSH2) involved in the early steps of DNA nucleotide mismatch repair lead to DNA replication errors, including microsatellite instability and ubiquitous somatic mutations. (Vogelstein et al. 1988, Boland et al. 1995). Heriditary nonpolyposis colorectal cancer syndrome reflects a germline mutation in one of four genes involved in DNA nucleotide mismatch repair.
The aspect that adenomas and carcinomas of the large intestine may show similar molecular alterations underscores the “evolution” of the concept of adenoma-carcinoma sequence, from a morphological phenomenon to a complex pathway of biomolecular events presumably induced by (or related to) exogenous and metabolic factors (Potter 1999). Many of these genetic events may target the transition from normal mucosa to small adenomas, and from these to large adenomas and infiltrating carcinoma (Potter 1999, Lairmore & Norton 1997, Redston 2001).

3 Aims of the research
The overall aim of this research was to elucidate the distribution and expression of CA I, II, IX, XII, and XIV in the gastrointestinal tract. The expression levels of these isoenzymes except CA XIV were investigated in both normal and neoplastic tissues. The specific goals were:
− to investigate the expression of CA I, II, IX and XII in the human intestine and colorectal tumours
− to investigate the location of CA IX and XII in the human pancreas and pancreatic tumours
− to examine the cellular localization of CA XIV in the murine liver and intestine.

4 Materials and methods
4.1 Tissue specimens (I-IV)
The procedures had the approval of the ethics committee of Oulu University Hospital and the research was carried out according to the provisions of the Declaration of Helsinki.
The human material studied for paper I consisted of samples taken together with routine histopathological specimens during surgical operations carried out at Oulu University Hospital for gastric carcinoma (n=2), rectal carcinoma (n=1), colon polyposis (n=2), or colon inertia (n=1). In addition to the samples of normal human gut, 67 (I) or 69 (II) colonic lesions from 59 (I) or 48 (II) patients were studied. Most of these tumour specimens contained adjacent normal tissue, which were studied for paper II. Seven metastases of colorectal adenocarcinomas were also analyzed, including 6 mesenteric lymph node metastases and 1 liver metastasis (I and II).
The human tissue samples from pancreas (n=39) used in paper III were obtained during surgical operations for pancreatic tumours. Most tumour specimens contained adjacent normal tissue. In addition, one specimen of normal pancreas was obtained from a patient with traumatic laceration of the pancreas. Two ductal carcinomas showed divergent differentiation in separate areas, which were considered separate lesions in immunohistochemical analyses. The only acinar carcinoma was well differentiated. Some specimens showed both normal and neoplastic histology in the same section.
The rodent tissue samples studied for paper IV consisted of samples obtained from three adult mice, which were anaesthetised and sacrificed by decapitation.
Details of the tissue specimens studied for papers I-III, as shown in Table 2, are provided in the original papers.

39
Table 2. Specimen types and number of samples studied in papers I-III.
Specimen type I II III Normal gut and tumours of intestinal origin
normal duodenum 1 1 normal jejunum 2 2 normal ileum 3 3 normal cecum 3 3 normal ascending colon 3 3 normal transverse colon 3 3 normal descending colon 3 3 normal sigmoid colon 2 2 normal rectum 4 4 hyperplastic polyp 8 8 juvenile polyp 1 adenoma, low dysplasia 12 13 adenoma, moderate dysplasia 18 18 adenoma, high dysplasia 8 9 adenocarcinoma, well differentiated 6 7 adenocarcinoma, moderately differentiated 9 9 adenocarcinoma, poorly differentiated 5 5 mesenteric lymph node metastasis 6 1 liver metastasis 6 1
Pancreatic tumours
microcystic adenoma 3 mucinous cystadenoma 3 acinar carcinoma 1 mucinous cystadenocarcinoma 4 ductal carcinoma, well differentiated 9 ductal carcinoma, moderately differentiated 10 ductal carcinoma, poorly differentiated 4 anaplastic carcinoma 1 endocrine tumour 6
4.2 Preparation of tissues (I-IV)
Tissue samples were divided into several small pieces about 5 mm thick. The specimens were fixed either in Carnoy’s fluid (absolute ethanol + chloroform + glacial acetic acid 6:3:1) for 6 hr (samples of the normal human gut (I) and pancreas (III)) or in 4% neutral-buffered formaldehyde for 24-48 hr (normal gut (II), tumour samples (I-III)). The

40
samples were then dehydrated, embedded in paraffin in a vacuum oven at 58°C, and sections of 5 µm were placed on gelatin-coated microscope slides.
The rodent liver for paper IV was perfused in situ through the abdominal aorta with 3% paraformaldehyde in phosphate-buffered saline (PBS), removed, and cut into slices. The slices were further immersion-fixed in 3% paraformaldehyde for 2 h at room temperature, cryoprotected overnight in 20% sucrose-PBS, and rapidly frozen in liquid nitrogen-cooled isopentane. Sections were cut at 5 µm using a Microm Cryo-Star microtome (Microm, Walldorf, Germany) amd dried onto Superfrost Plus microscope slides (Menzel, Braunschweig, Germany).
4.3 Antibodies against CA isoenzymes (I-IV)
Details of the production and characterisation of the antisera for the CA isoenzymes, as shown in Table 2, are provided in the original papers.
Table 3. CA isoenzyme antisera used in papers I-IV.
Antiserum Paper (I-IV) Reference Anti-HCA I II Parkkila A-K et al. 1993 Anti-HCA II II Parkkila A-K et al. 1993 Anti-RCA II IV Fleming et al. 1994 Anti-MCA IV IV Brion et al. 1992 Anti-HCA IX II, III Pastorekovà et al. 1992 Anti-HCA XII I, II, III Karhumaa et al. 2000 Anti-MCA XIV IV Parkkila S et al. 2001
4.4 Other antibodies (III)
In paper III, immunostaining for Ki-67 was performed to demonstrate the proliferative activity of the epithelial cells expressing CA IX. The rabbit antibody for human Ki-67 was purchased from Zymed Laboratories, San Francisco, CA (III). Monoclonal antibodies against human pVHL have been characterized previously by Sakashita et al. (1999). Some sections were immunostained using monoclonal mouse anti-human p53 antibody (DO-7) purchased from Novocastra Laboratories (Newcastle upon Tyne, United
Kingdom).

41
4.5 Immunoperoxidase and immunofluorescence staining methods (I-IV)
The CA isoenzymes were immunostained by the biotin-streptavidin complex method, employing the following steps:
I Pre-treatment of the sections with undiluted cow colostral whey (Biotop Oy, Oulu, Finland) for 30 min and rinsing in PBS
II Incubation for 1 hr with the primary anti-HCA XII serum diluted 1:100 in 1% bovine serum albumin (BSA-PBS), the M75 hybridoma medium (anti-HCA IX) diluted 1:10, anti-VHL antibodies diluted 50 µg IgG/ml or normal rabbit serum diluted 1:100 in 1% BSA-PBS. In paper II, anti-HCA I and II sera and normal rabbit serum were diluted 1:100 in 1% BSA-PBS and the samples were incubated for 12 hr at +4ºC.
III Incubation for 1 hr with biotinylated swine anti-rabbit IgG (Dakopatts, Glostrup, Denmark) or goat anti-mouse IgG (Dakopatts) diluted 1:300 in 1% BSA-PBS.
IV Incubation for 30 min with peroxidase-conjugated streptavidin (Dakopatts) V Incubation for 2 min in DAB solution containing 9 mg 3,3’-diaminobenzidine
tetrahydrochloride (Fluka, Buchs, Switzerland) in 15 ml PBS + 5 µl 30% H2O2. The sections were washed three times for 10 min in PBS after incubation steps 2 and 3, and four times for 5 min in PBS after step 4.
VI All the incubations and washings, except the first incubation (step 2) with anti-HCA I or II, were carried out at room temperature. After the immunostaining, the tumour sections were counterstained with hematoxylin. The stained sections were examined and photographed with Leitz Aristoplan (Wetzlar, Germany), Olympus BH (Tokyo, Japan) or Nikon Eclipse E600 (Tokyo, Japan) microscope.
To examine the subcellular localization of CA XII in the human colonic epithelium, tissue sections were stained by an immunofluorescence method and analyzed by confocal laser scanning microscopy in paper I. The steps in the immunofluorescence were as follows: (a) pre-treatment of the sections with 1% BSA-PBS for 40 min, (b) incubation for 1 hr with 1:100 diluted anti-HCA XII serum in 1% BSA-PBS, (c) washing three times for 5 min in PBS, (d) incubation for 1 hr with 1:30 diluted fluorescein-conjugated goat anti-rabbit IgG (Dakopatts) in 1% BSA-PBS, and (e) washing three times for 5 min in PBS. The sections were examined with a confocal laser scanning microscope (Leitz CLSM; Leica Microsystems, Heidelberg, Germany).
In paper IV, sections on Superfrost Plus microscope slides (Menzel, Braunschweig, Germany) were incubated with PBS containing 20% cow colostral whey for 40 min. The sections were then incubated for 2 h with polyclonal anti-MCA XIV or preimmune serum, diluted 1:200 in 1% BSA-PBS, washed three times for 5 min in PBS and incubated for 1 h with Alexa 568-coupled goat anti-rabbit IgG, diluted 1:200 in BSA-PBS. After four 5-min washes in PBS, slides were mounted in Immu-mount (Shandon, Pittsburgh, PA). The sections were examined with a conventional epifluorescence microscope (Nikon Corporation, Tokyo, Japan) or a confocal laser-scanning microscope (Zeiss, Göttingen, Germany).

42
4.6 Evaluation of immunoreactivity (I-III)
For papers I and II the intensity of the immunoreactions were scored on semiquantitative scales of 0 to 4 as follows: 0, no reaction; 1, occasional positive cells; 2, weak reaction; 3, moderate reaction; 4, strong reaction. The distribution of immunoreactivity between the superficial and deep parts of the mucosa was recorded separately.
In paper III, the extent and intensity of the staining reaction for CA IX and CA XII were scored in the central area of the tumour on a scale of 0 to 4. In terms of the staining intensity, the scores were assessed as follows: 0, no reaction; +/-, very weak reaction; +, weak reaction; ++, moderate reaction; +++, strong reaction. The extent of the positive signal was scored as follows: no positive cells, <25% positive cells, 25-<50% positive cells, 50-<75% positive cells, and 75-100% positive cells.
4.7 SDS-polyacrylamide gel electrophoresis (SDS-PAGE) and western blotting (I, IV)
For paper I, samples containing 25 µg of protein from homogenized human transverse colon were analyzed by SDS-PAGE under reducing conditions according to Laemmli (1970). All the reagents for SDS-PAGE were from Bio-Rad Laboratories (Richmond, CA) or Sigma (St. Louis, MO). The electrophoreses were performed in a Mini-Protean electrophoresis unit (Bio-Rad Laboratories) using a 10% acrylamide separating gel and a 4% acrylamide stacking gel. The proteins were transferred electrophoretically from the gel to a PVDF membrane (Millipore; Bedford, MA) in a Novex Blot Module (Novex; San Diego, CA). After the transblotting, the sample lanes were first incubated with TBST buffer (10 mM Tris-HCl, pH 7.5, 150 mM NaCl, 0.05% Tween-20) containing 10% cow colostral whey (Biotop Oy) for 30 min and then with the first antibody diluted 1:2000 in TBST buffer for 1 hr. The membranes were washed five times for 5 min with TBST buffer and incubated for 30 min with alkaline phosphatase-conjugated goat anti-rabbit IgG (Bio-Rad Laboratories) diluted 1:3000 in TBST buffer. After washing four times for 5 min in TBST buffer, the polypeptides were visualized by means of a chemiluminescence substrate (Bio-Rad Laboratories). All the steps were carried out at room temperature.
For paper IV, mice were sacrificed by decapitation, and the liver and colon were removed. 20 µg of total cell protein per lane from homogenized tissue samples or stably transfected CHO cells expressing wild-type mouse CA XIV (Parkkila S et al. 2001) was subjected to SDS-PAGE under reducing conditions. Protein standards for SDS-PAGE were from Bio-Rad Laboratories (Richmond, CA). The electrophoreses were performed in a Novex XCell SureLock electrophoresis unit (Invitrogen Corp/NOVEX, Carlsbad, CA) using a Novex NuPAGE 10% Bis-Tris gels. The proteins were transferred electrophoretically from the gel to a nylon membrane (Millipore; Bedford, MA) in a Novex Blot Module. The further procedures were the same as described above.

43
4.8 Extraction of RNA and reverse transcription-polymerase chain reaction (RT-PCR) (IV)
In paper IV, the liver, colon, jejunum, and kidneys were removed from mice and the samples were used for RT-PCR analysis. Epithelial cells of the intestinal mucosa were obtained by using a scraping technique as described (Waheed et al. 1999). Total cellular RNA was extracted from the cell or tissue samples using TRIzol reagent (Gibco-BRL, Gaithersburg, MD), according to the manufacturer’s instructions. The concentration and purity of RNA was determined by spectrophotometer at 260 and 280 nm. Reverse transcription (RT) and polymerase chain reaction (PCR) amplification were performed using a single-step method (Aatsinki et al. 1994). Two primers for amplifying CA XIV were chosen based on the published mouse CA XIV sequence (Mori et al. 1999). Primers for rat glyceraldehyde-3-phosphate dehydrogenase (GAPDH) (Holappa et al. 1999) were used to monitor the quality of the RNA samples.
Ten micrograms of total RNA was used as a template for subsequent RT and PCR reactions. Reverse transcription was performed at 42°C for 1 hr followed by denaturation at 95°C for 2 min. The PCR cycling protocol consisted of 30 cycles of denaturation at 95°C for 1 min, annealing at 55°C for 1 min, and extension at 72°C for 1 min, followed by final extension at 72°C for 5 min. The PCR products were analyzed by electrophoresis on 1.5% agarose gels containing ethidium bromide.
4.9 Statistical methods (II, III)
In paper II, the statistical evaluations were performed by one-way analysis of variance and Bonferroni’s t-test. The correlation between staining intensity and extent was analysed using Spearman’s correlation analysis (III).

5 Results
5.1 Expression of CA I, II, IX, XII, and XIV in normal gastrointestinal tract (I-IV)
The immunoperoxidase technique was used to study the distribution of CAs in the human alimentary tract. Cellular and subcellular localisation of CA XII, in human intestine and CA XIV in murine hepatocytes were demonstrated by immunofluorescence technique.
CA XII showed a characteristic staining pattern in human gut (I). First, it was highly expressed in the cecum, ascending colon, transverse colon, descending colon, sigmoid colon, and rectum, but absent from the small intestine. Second, the staining for CA XII was most intense in the surface epithelial cuff region of the large intestine. Third, the positive signal for CA XII was clearly confined to the basolateral plasma membrane in the enterocytes. In large intestine, both CA I and II were abundant in the normal epithelium of the surface epithelial cuff region as described previously by Parkkila S et al. 1994. In contrast, CA IX was confined to the crypt enterocytes while it was absent in the superficial part of the mucosa. This finding was originally reported by Pastoreková et al. (1997). Our results have further indicated that CA XIV is another CA isoenzyme expressed in the mouse colonic enterocytes (IV). The positive signal was mainly located in the apical plasma membrane and it was found both in the crypts and the surface epithelial cuff region. CA XIV was also expressed in the smooth muscle cells of the muscularis propria locating beneath the mucosa and submucosa layers of the gastrointestinal canal. The positive signal showed a somewhat patchy staining pattern, which is consistent with the typical caveolar structure of the plasma membrane in those cells.
Isoenzymes CA IX and XII appeared to be expressed in the acinar and ductal compartments of the human pancreas, but not in the endocrine glands (III). Epithelial cells of some intra- and interlobular ducts in the human pancreas showed positive staining for CA IX. In addition, sporadic expression of CA IX was observed in some acini of the exocrine pancreas. The positive signal was clearly confined to the basolateral plasma membranes in the acinar and ductal cells. The staining for CA XII was most

45
intense in the acinar cells. A faint but distinct staining was also found in the ductal epithelium. In both places, the positive signal was confined to the basolateral plasma membrane.
In paper IV, the mouse liver was analyzed for CA XIV protein expression using a specific antibody raised against the recombinant mouse CA XIV. The isoenzyme was expressed at the plasma membrane of all hepatocytes. The staining intensity showed no apparent zonal variation within a hepatic lobule. Both the apical (canalicular) and basolateral (sinusoidal) plasma membrane domains were positive. In western blotting, anti-MCA XIV serum recognized a faint 44/46-kDa doublet band in the liver. It is notable that the western blotting reaction was stronger in the liver than in the colon, which was also in line with the immunostaining results. The mouse colon, jejunum, and liver were also analyzed for CA XIV mRNA expression. Strong 434-kb bands were observed in the liver and kidney. Weak CA XIV mRNA expression was detected in the colon, while the jejunum remained totally negative.
5.2 Expression of CA I, II, IX, and XII in gastrointestinal tumours (I-III)
In most specimens of the large intestine, the positive staining for CA I decreased in hyperplastic areas compared to the normal epithelium. The hyperplastic mucosa usually showed a weak reaction for CA I in the enterocytes of the surface epithelial cuff region, whereas the signal was barely apparent in the deep parts of the cryptal epithelium. The intensity of the immunoreaction for CA I was mostly similar in the adenomatous surface epithelium and hyperplastic polyps. In malignant colorectal tumours, the positive immunoreaction for CA I was restricted to occasional epithelial cells, being present in both the superficial and deep parts of the lesions. The mean intensity of the immunostaining was significantly decreased in the superficial malignant mucosa when compared to adenomas (II).
A moderate reaction for CA II was observed in the enterocytes of the normal surface epithelial cuff region, while the deep parts of the mucosa remained negative. CA II-positive staining was further decreased from moderate reaction in hyperplastic polyps to weak in adenomas. The positive immunoreaction for CA II in the deep parts of the adenomatous mucosa was restricted to a few occasional epithelial cells. The staining intensity for CA II in the upper parts of the lesions became weaker with a more severe grade of dysplasia, while the immunoreaction for CA I in the same region remained weak in all adenoma dysplasia categories. In the deep parts of the lesions, faint positive immunostaining appeared only in a few epithelial cells for both isoenzymes as is true for normal mucosa. Both CA I- and II-positive adenomas were evenly distributed along the cranial-caudal axis of the large intestine. Compared to adenomas, superficial malignant mucosa showed significantly weaker immunostaining for CA II (II).
The data related to CA IX was extracted from studies replicating previous results (Saarnio et al. 1998a,b), in which CA IX expression was evaluated in the same tumour samples. In striking contrast, the staining intensity for CA IX was clearly higher in

46
adenomas and carcinomas (more so in the superficial part) compared to the normal mucosa and hyperplastic lesions as described previously by Saarnio et al. 1998b.
CA XII was highly expressed in the surface epithelial cuff region of the normal large intestinal mucosa, but expressed extremely weakly in the lower parts of the mucosa (I). Also the hyperplastic polyps generally showed a moderate reaction in the enterocytes of the surface epithelial cuff region, whereas the signal for CA XII was barely apparent in the deep parts of the cryptal epithelium. The CA XII-positive adenomas were evenly distributed along the cranial-caudal axis of the large intestine and the reaction became more diffusely spread in the lower parts of the lesion in the adenomas with a more severe grade of dysplasia. In the colorectal carcinomas the immunoreaction was generally diffuse, being present in both the superficial and deep parts of the mucosa (I).
The staining reaction for CA IX and XII in the hyperplastic epithelium in pancreatic tumour specimens generally showed a stronger basolateral immunoreaction for CA IX compared to the normal epithelial cells (III). Immunostaining for Ki-67 was performed in parallel sections to demonstrate the proliferative activity of the epithelial cells expressing CA IX. 10 out of 29 malignant tumours of exocrine pancreas showed moderate or strong immunoreaction for CA IX. The staining extent or intensity did not correlate with the grade of differentiation. Interestingly, desmoplastic connective tissue in the malignant lesions occasionally showed prominent immunostaining for CA IX. All mucinous cystadenocarcinomas showed weak to strong immunoreaction that was present in <75% of malignant cells and all mucinous cystadenomas showed strong immunostaining, suggesting that CA IX expression is characteristic for these tumours. The benign microcystic adenomas showed weak to strong immunoreaction, whereas all endocrine tumours were negative for CA IX. The expression of CA XII was very weak or weak in most pancreatic carcinomas. In the ductal carcinomas, the positive signal for CA XII was present in 0-75% of the epithelial cells. From the benign tumours of exocrine pancreas, only one mucinous cystadenoma and one microcystic adenoma showed strong positive signal for CA XII. Three endocrine tumours remained unstained and one glucagonoma showed a weak positive reaction.
5.3 Expression of von Hippel-Lindau protein in normal gastrointestinal tract and colorectal tumours
Expression of pVHL was examined in series of 18 samples of histologically normal human colon or rectum and 23 colorectal tumours. These included 7 adenomas and 16 adenocarcinomas. The adenomatous lesions included 6 tubular and 1 tubulovillous tumours and the group of 16 malignant colorectal tumours consisted of 6 well differentiated, 9 moderately differentiated, and 1 poorly differentiated adenocarcinomas. Immunohistochemical staining for pVHL showed positive epithelial staining in all normal and adenomatous tissue specimens. All malignant tumours except one rectum carcinoma were positive. Figure 3B demonstrates a representative example of immunostaining of pVHL in normal colonic mucosa. Almost all enterocytes showed positive cytoplasmic signal. Similarly, adenomatous and malignant lesions were positive

47
for pVHL (Figs 4B, 5B, F). Figure 4F shows pVHL immunostaining results in normal and adjacent transitional epithelium. There was no distinct difference in the staining intensity or extent between these two regions.
Fig. 3. Immunohistochemical staining for pVHL, CA IX, and CA XII in normal colon. Arrows in panel C indicate CA IX positive staining in the enterocytes. Hematoxylin-eosin (H&E) counterstaining in panel A.

48
Fig. 4. Examples of immunohistochemical staining for pVHL, CA IX, and XII in serial sections of moderate (lower part in panels A-D) and grave (upper part in panels A-D) adenoma and transitional epithelium (adenoca gr I) (small arrows) (E-H) adjacent to normal tissue (arrowhead). Hematoxylin-eosin (H&E) counterstaining in panels A and E.

49
Fig. 5. Immunohistochemical staining for pVHL, CA IX, and XII in serial sections of adenocarcinoma (gr II) with (A-D) or without (E-H) mucinous component. Hematoxylin-eosin (H&E) counterstaining in panels A and E.

6 Discussion
6.1 Carbonic anhydrases in the gastrointestinal tract
Carbonic anhydrases are evolutionarily ancient enzymes, which are expressed in all mammalian tissues and they are found along the entire GI tract, from the mouth and salivary glands to the rectum (Parkkila et al. 1994, Saarnio et al. 1998a, Leinonen et al. 2001). However, there is a considerable heterogeneity between different organs with respect to CA enzymatic activity, isoenzyme content, and their cellular localization. It is also notable that CAs of the GI tract can serve several functions such as regulation of acid-base homeostasis, neutral salt absorption and cell volume (Parkkila S & Parkkila A-K 1996).
The new evidence that at least seven enzymatically active CA isoenzymes (CA I, II, IV, V, IX, XII, and XIV) are expressed in the mammalian large intestine indicates the complexity of the physiological processes occurring in the GI tract. In addition, the CA isoenzymes are differentially expressed along the intestinal segments and also between cryptal and villal enterocytes. Finally, it is a matter of debate whether the regulatory mechanisms could be different depending on the species studied. Even though some of these isoenzymes have heretofore been demonstrated only in rodents or humans, it is conceivable that general transport and pH regulation mechanisms are qualitatively similar in various species (Wills 1985, Dawson 1991). Based on the previous and the present data, there are four membrane-bound isoenzymes in the colonic enterocytes (Fleming et al. 1995, Saarnio et al. 1998a, I, IV). The transmembrane forms, CA IX and XII are located on the basolateral surfaces and CA XIV predominantly in the apical brush border membrane. The fourth membrane-associated isoenzyme is GPI-linked CA IV which is restricted to the brush border membrane.

51
Fig. 6. Schematic model of the cellular absorption and secretion of electrolytes and water in the mammalian colon. Examples of ion transport mechanisms are demonstrated in a single enterocyte. Na+/H+ exchangers type 1, 2, and 3 (NHE1/2/3), proteins downregulated in adenoma (DRA), anion exchangers type 1, 2, and 3 (AE1/2/3), aquaporin 3, 4, and 8 (AQP3/4/8), cystic fibrosis transmembrane conductance regulator (CFTR), short-chain fatty acids (SCFA-), KCl cotransporter (KCC1), carbonic anhydrase (CA). (Dawson 1991, Alrefai et al. 2001, Flemström & Isenberg 2001, Seidler et al. 2001, Kunzelmann & Mall 2002.)
apical
DRA
DRA
NHE2/3
CFTR
AE1?
AQP8
basolateral
KCC1
AE21/3?
NHE1
AQP4
AQP3
+
CA
+
+
CA
Na+
H+
OH-
HCO3-
HCO3-
Cl-
Cl-
Cl-
Cl-
HCO3-
K+
HCO3-
H2O
Cl-
Na+
SCFA-
2 K+
HCO3-
HCO3-
Na+
Na+
3 Na+
K+
K+
Cl-
Cl-
Cl-
H+
H2O
H2O
H+
HCO3-
H2O
CO2
H2O
H+
OH-
CO2
HCO3-

52
CA function is ultimately linked to certain transport proteins in both apical and basolateral cell membranes. In a recent article Kunzelmann and Mall (2002) have thoroughly reviewed these ion transport mechanisms in both colonic absorption and secretion. Identification of various ion transport proteins in colon had made it possible to build schematic models for ion movement in different segments of the colon. Figure 6 represents a model for colonic ion and water transport that incorporates current knowledge regarding membrane transport processes (Dawson 1991, Alrefai et al. 2001, Flemström & Isenberg 2001, Seidler et al. 2001, Kunzelmann & Mall 2002). It is important to emphasize that it is still a matter of discussion whether some of the proposed ion transport proteins are functional in human colon. The absorption of NaCl can be electrogenic or electroneutral depending on the site of the process. In the proximal colon the absorption is primarily due to an electroneutral process via parallel luminal Na+/H+ and Cl−/HCO3
− exchange (Rajendran & Binder 2000). The electroneutral luminal Na+ uptake is driven by the basolateral Na+-K+-ATPase lowering intracellular Na+ concentration. Na+ and Cl− transport is coupled via changes in intracellular pH and Cl− concentration (Geibel et al. 2001). Electroneutral NaCl absorption is also stimulated by short-chain fatty acids (SCFA), which are produced by colonic bacteria. The absorbed SCFA can stimulate both apical Na+/H+ and Cl−/HCO3
− exchangers, and thus, have a significant impact also on the regulation of colonic fluid balance and luminal as well as cytosolic pH (Kunzelmann & Mall 2002). The electrogenic absorption of NaCl mainly occurs in the distal colon via the luminal epithelial Na+ channels (ENaC) and transcellular/paracellular absorption of Cl− (Greger et al. 1997).
The participation of the colonic CA in the electroneutral NaCl reabsorption is well established. Studies with acetazolamide suggest that CA is involved in the absorption of NaCl via the synchronous operation of apical Na+/H+ and Cl−/HCO3
− exchange processes. CAs are also shown to participate in the alkalinization of the luminal contens by generating HCO3
− for apical Cl−/HCO3− exchange (Feldman 1994). Recent results from
Casey’s laboratory have indicated that CAs can physically and functionally associate with Cl−/HCO3
− (AE1) and Na+/H+ exchangers (Sterling et al. 2001, 2002, Li et al. 2002). So far, CA IV is the only known membrane-bound CA isoenzyme which interacts with AE1 protein (Sterling et al. 2002). It would be tempting to hypothesize that CA IX, XII and XIV could also drive ion exchange across the plasma membrane via a direct link with transport protein. The location of the membrane-associated isoenzymes CA IV, IX, XII and XIV in the surface epithelium of the colon place them in a strategic position for the regulation of luminal and extracellular acid-base homeostasis and microclimate (Fleming et al. 1995, Saarnio et al. 1998a, I, IV).
The net movement of water in colon is driven osmotically. Water is transported via both paracellular and transepithelial routes (Spring 1998). Although the proteins involved in NaCl absorption have been studied for decades, the exact molecular mechanisms of water absorption have remained largely unresolved. Recently, new information on water channel proteins, aquaporins (AQP), has opened new avenues to explore this understudied area of physiology. From different isoforms of AQPs, at least AQP3, AQP4, and AQP8 are expressed in the colonic epithelial cells (Hasegawa et al. 1994, Ishibashi et al. 1995, Frigeri et al. 1995, Koyama et al. 1997). AQP3 and AQP4 are expressed at the basolateral plasma membrane like CA XII. AQP8 is confined to the apical plasma membrane like CA IV and CA XIV. Interestingly, Nakhoul et al. (1998) have shown in

53
Xenopus oocytes that CO2 flux can be mediated by AQP1, and therefore, AQPs could act as a gas channel even though CO2 has ealier been believed to diffuse freely across the cell membranes. Even though there is no experimental evidence yet available, one could speculate that AQPs would be an interesting target when searching protein interactions of membrane-bound CAs. By analogy to ion exchangers, AQPs are typically expressed in tissues expressing high levels of CAs such as colon, liver, pancreas, kidney, salivary gland, and stomach (Ma & Verkman 1999).
Ion transport mechanisms similar to those described above for the colonic epithelium are common in epithelial cells of other GI organs. The best characterized are probably those transport proteins and CAs that are expressed in the gastric mucosa. In addition to the colonic mucosa, our studies were focused on the two major digestive organs, pancreas and liver, in which the role of different CAs has remained a matter of debate. For example, expression of CA III is well documented in the rat hepatocytes where it has been linked to protection against oxidative damage (Cabiscol & Levine 1995, Mallis et al. 2000, 2002). The oxidative modification of CA III was earlier associated to loss of phosphatase activity demonstrated in CA III enzyme preparation under the oxidative stress (Cabiscol & Levine 1995, 1996). However, more detailed studies showed evidence that the phosphatase activity of CA III was due to a contamination of the enzyme (Kim et al. 2000).
In addition to CA III, hepatocytes are known to contain three other CAs, namely CA II, VA and XIV (Parkkila S et al. 1994, Dodgson 1991, IV). From these isoenzymes, CA XIV is the only membrane-bound form which most probably regulates pH and ion transport between the hepatocytes, bile canaliculi and hepatic sinusoids. Even though CA XIV showed positive signal in both domains of the plasma membrane, i.e. basolateral and apical (canalicular), the apical membrane was more strongly labelled (IV). It is well known that bile acids are effectively transported across the apical plasma membrane of the hepatocyte into the bile canaliculi (Erlinger 1996). Even though the previous studies have shown that Na+/H+ exchanger, Na+-HCO3
− cotransporter and Cl−/HCO3− exchanger
are expressed in hepatocytes (Gleeson et al. 1989, Renner et al. 1989, Muller & Jansen 1997, Strazzabosco et al. 1997, Furimsky et al. 2000), there is no precise information on the subcellular localization of these transport proteins. Interestingly, AQP0 and AQP8 have been demonstrated in the canalicular plasma membrane, suggesting a role for these AQPs in regulation of hepatocyte volume and bile secretion (Calamita et al. 2001, Tani et al. 2001, Huebert et al. 2002). However, it has been proposed that the major fraction of water and electrolyte tranportation into bile takes place at a more distal (ductular) level in part under the control of secretin (Erlinger 1996). In either case, we will need to reconsider the localization and function of CAs in the entire biliary system using novel approaches such as analysis of different knockout animal models.
The third major area of the present investigation was pancreas, which contains two morphologically and functionally different elements: endocrine and exocrine compartments. Our studies did not reveal any novel findings related to the endocrine part. In the exocrine pancreas, however, both CA IX and XII were found to be expressed in the ductal and acinar cells (III). Functionally, the pancreatic epithelial cells have several similar features with colonic epithelium in terms of the maintenance of ion and pH homeostasis. In this context, it is important to recall that the physiological outcome of the pancreatic and colonic function are very different: secretion of bicarbonate-rich fluid in

54
pancreas and absorption of water and electrolytes in colon. The mechanisms involved in the pancreatic duct cell ion transport have been recently reviewed by Ishiguro et al. (2001). They presented a model where the basolateral plasma membrane contained Na+-HCO3
− cotransporter and Cl−/HCO3− exchanger. The bicarbonate secretion across the
apical plasma membrane was proposed to occur by exchange for Cl− ions on Cl−/HCO3−
exchanger working in parallel with the cystic fibrosis transmembrane conductance regulator (CFTR) Cl− channel. Solving the question of how different CAs can function in pancreatic duct and acinar cells will be a challenging task, not least technically. The knockout mouse models will hopefully provide invaluable information on the role of each isoenzyme. It is noteworthy, however, that recent studies on CA IX-deficient mice did not reveal any pancreatic phenotype (Ortova Gut et al. 2002).
6.2 Carbonic anhydrases in neoplastic gastrointestinal tissues
Besides the role of different CA isoenzymes in the normal physiology of the colon, liver and pancreas, we also studied the distribution and expression of CAs in GI tumours. During the last decade the CA research field gained a lot of attraction as CA IX and XII were found to be associated with neoplastic processes. A direct link was obtained by the discovery that both isoenzymes are down-regulated by wild-type VHL tumour suppressor protein (Ivanov et al. 1998).
It has recently been demonstrated that CA IX is expressed in the proliferating enterocytes of the human colon and it showed a distinct increase in the expression level in most adenomas and primary colorectal carcinomas (Saarnio et al. 1998a,b). Expression of CA XII was up-regulated in the deep parts of the tumours where it was normally weakly expressed (I). In contrast, CA I and II showed maximal immunoreactions in the normal colorectal mucosa declining in adenomas and carcinomas (II). Based on the immunostaining patterns, these four CA isoenzymes expressed in colon can be considered promising tumour markers, which might be utilized as diagnostic tools to assess tumour progression to malignancy.
The reciprocal changes in the expression of the soluble cytoplasmic isoenzymes (CA I and II) and the membrane-associated forms (CA IX and XII) observed in colorectal tumours are probably due to different mechanisms. Loss of expression of the closely linked CA1 and 2 genes could result from loss of alleles specifying these genes. As a second option, the down-regulation of CA I and II could also result from reduced levels of a common transcription factor. The latter is strongly supported by a recent finding that a reduced expression of CA II in colorectal cancer is due to disruption of β-catenin/TCF-4 activity (van de Wetering et al. 2002). Up-regulation of CA IX and XII could also be explained by two different mechanisms. First, contribution of microenvironmental stress to development of tumor phenotype has become an increasingly accepted explanation. Indeed, Wykoff et al. (2000) have demonstrated a close connection between tumour hypoxia and the expression of CA IX and XII. In the same study they showed that up-regulation of CA IX occurs at the level of transcriptional activation via HIF-1 transcriptional complex, the critical mediator of hypoxic responses. Second,

55
overexpression of CA IX and XII has been associated with loss of expression of the VHL tumour suppressor gene. Our preliminary studies indicated that pVHL is, indeed, expressed in the normal and neoplastic colorectal mucosa (Figs 3, 4, 5). Therefore, future studies should be aimed to investigate VHL mutations or deletions in the colorectal tumours. This would strengthen the hypothesis that loss of this gene product underlies the observed up-regulation of CA IX and XII in colorectal tumours.
Ivanov et al. (1998) hypothesized that CA XII may be involved in tumour invasion by acidifying the extracellular milieu surrounding the cancer cells. This acidification would, in turn, contribute to activation of enzymes involved in extracellular matrix degradation. This hypothesis has been supported by more recent findings that the invasive capacity of renal cancer cells is markedly reduced by treating them with acetazolamide, a potent inhibitor of CA activity (Parkkila S et al. 2000a). Even more interesting from our point of view is the finding that a similar effect was described in cells derived from colorectal cancer (Supuran et al. 2001). It is of considerable interest that some other CA inhibitors also show substantial antitumour activity and are in phase II clinical trials for treatment of different types of cancer (Casini et al. 2002). These trials may turn out to be successful, because Teicher et al. (1993) reported that acetazolamide is useful in anticancer therapies in combination with cytotoxic agents. Based on the previous and present findings, CA II seems to be the major isoenzyme expressed in pancreatic tumours (Parkkila S et al. 1995, III). As a high activity isoenzyme it may play a key role in regulating the pH in the vicinity of cancer cells, and therefore, it would be the major target in pharmacological studies on pancreatic CAs.

7 Conclusions
− CA XII is absent from the small intestine, but is expressed in all segments of the normal large intestine. The positive signal for CA XII is confined to the basolateral plasma membranes of the epithelial cells of the surface epithelial cuff.
− The reciprocal pattern of expression observed for membrane-associated (CA IX and XII) and cytoplasmic (CA I and II) isoenzymes in intestinal samples suggests that CA IX and XII may be functionally involved in tumour progression to malignancy and/or in invasion.
− CA I and II, which are felt to play important physiological roles in the normal colorectal mucosa, may not be required for growth of colorectal cancers and their expression consistently diminishes with progression to malignancy.
− In the human pancreas CA IX and XII appear to be sporadically expressed in the basolateral plasma membrane of the normal acinar and ductal epithelium.
− The increased expression of CA IX in hyperplastic ductal epithelium may contribute to the pancreatic tumourigenesis.
− CA XIV is expressed in the hepatocyte plasma membrane and its localization on both apical and basolateral membrane domains suggests an important role for this isoenzyme in the regulation of ion and pH homeostasis in the liver.

References
Aatsinki JT, Lakkakorpi JT, Pietilä EM, Rajaniemi H. A coupled one-step reverse transcription PCR procedure for generation of full-length open reading frames. Biotechniques 16:282-288, 1994.
Adachi M, Muto T, Okinaga K & Morioka Y (1991) Clinicopathologic features of the flat adenoma. Dis Colon Rectum 34:981-986.
Aldred P, Fu P, Barret G, Penschow JD, Wright RD, Coghlan JP & Fernley RT (1991) Human secreted carbonic anhydrase: cDNA cloning, nucleotide sequence, and hybridization histochemistry. Biochemistry 30:569-575.
Alrefai WA, Tyagi S, Nazir TM, Barakat J, Anwar SS, Hadjiagapiou C, Bavishi D, Sahi J, Malik P, Goldstein J, Layden TJ, Ramaswamy K & Dudeja PK (2001) Human intestinal anion exchanger isoforms: expression, distribution, and membrane localization. Biochim Biophys Acta 1511:17-27.
Ames BN & Gold LS (1998) The causes and prevention of cancer: the role of environment. Biotherapy 11:205-220.
Ames BN, Gold LS & Willet WC (1995) The causes and prevention of cancer. Proc Natl Acad Sci USA 92:258-265.
Balboni E & Lehninger AL (1986) Entry and exit pathways of CO2 in rat liver mitochondria respiring in a bicarbonate buffer system. J Biol Chem 261:3563-3570.
Barnea G, Silvennoinen O, Shaanan B, Honegger AM, Canoll PD, D’Eustachio P, Morse B, Levy JB, Laforgia S, Huebner K, Musacchio JM, Sap J & Schlessinger J (1993) Identification of a carbonic anhydrase-like domain in the extracellular region of RPTP gamma defines a new subfamily of receptor tyrosine phosphatases. Mol Cell Biol 13:1497-1506.
Bartošová M, Parkkila S, Pohlodek K, Karttunen TJ, Galbavý S, Mucha1 V, Harris AL, Pastorek J & Pastoreková S (2002) Expression of carbonic anhydrase IX in breast is associated with malignant tissues and is related to overexpression of c-erbB2. J Pathol 197:314-321.
Beasley NJ, Wykoff CC, Watson PH, Leek R, Turley H, Gatter K, Pastorek J, Cox GJ, Ratcliffe P & Harris AL (2001) Carbonic anhydrase IX, an endogenous hypoxia marker, expression in head and neck squamous cell carcinoma and its relationship to hypoxia, necrosis, and microvessel density. Cancer Res 61:5262-5267.
Boland CR, Sato J, Appelman HD, Bresalier RS & Feinberg AP (1995) Microallelotyping defines the sequence and tempo of allelic losses at tumour suppressor gene loci during colorectal cancer progression. Nat Med 1:902-909.
Bosman FT (1995) Prognostic value of pathological characteristics of colorectal cancer. Eur J Cancer 31:1216-1221.
Brion LP, Cammer W, Satlin LM, Suarez C, Zavilowitz BJ & Schuster VL (1997) Expression of carbonic anhydrase IV in carbonic anhydrase II-deficient mice. Am J Physiol 273:F234-F245.

58Brown D, Zhu XL & Sly WS (1990) Localization of membrane-associated carbonic anhydrase type
IV in kidney epithelial cells. Proc Natl Acad Sci USA 87:7457-7461. Bundy HF (1977) Carbonic anhydrase. Comp Biochem Physiol 57B:1-7. Butterworth PHW, Barlow JH, Brady HJM, Edwards M, Lowe N & Sowden JC (1991) The
structure and regulation of the human carbonic anhydrase I gene. In: Dodgson SJ, Tashian RE, Gros G & Carter ND (eds) The Carbonic Anhydrases. Cellular Physiology and Molecular Genetics. Plenum Press, New York, p 197-207.
Cabiscol E & Levine RL. Carbonic anhydrase III (1995) Oxidative modification in vivo and loss of phosphatase activity during aging. J Biol Chem 270:14742-14747.
Cabiscol E & Levine RL (1996) The phosphatase activity of carbonic anhydrase III is reversibly regulated by glutathiolation. Proc Natl Acad Sci USA 93:4170-4174.
Caflisch CR (1990) Effect of a nonsteroidal antiandrogen, flutamide on intraluminal acidification in rat testis and epididymis. Andrologia 25:363-367.
Caflisch CR & DuBose TD (1990) The effect of estrogen on in situ pH, pCO2 and bicarbonate concentration in rat testis and epididymis. Mol Androl 2:341-351.
Calamita G, Mazzone A, Bizzoca A, Cavalier A, Cassano G, Thomas D & Svelto M (2001) Expression and immunolocalization of the aquaporin-8 water channel in rat gastrointestinal tract. Eur J Cell Biol 80:711-719.
Carter MJ (1972) Carbonic anhydrase: isoenzymes, properties, distribution, and functional significance. Biol Rev 47:465-513.
Carter N, Jeffery S, Shiels A, Edwards Y, Tipler T & Hopkinson DA (1979) Characterization of human carbonic anhydrase III from skeletal muscle. Biochem Genet 17:837-854.
Carter N, Wistrand PJ & Lönnerholm G (1989) Carbonic anhydrase localization to perivenous hepatocytes. Acta Physiol Scand 135:163-167.
Casini A, Scozzafava A, Mastrolorenzo A & Supuran CT (2002) Sulfonamides and sulfonylated derivatives as anticancer agents. Current Cancer Drug Targets 2:55-75.
Charney AN & Egnor RW (1989) Membrane site of action of CO2 on colonic sodium absorption. Am J Physiol 256:C584-C590.
Chegwidden WR, Dodgson SJ & Spencer IM (2000) The roles of carbonic anhydrase in metabolism, cell growth and cancer in animals. In: Chegwidden WR, Carter ND & Edwards YH (eds) The Carbonic Anhydrases New Horizons. 1st ed. Basel: Birkhäuser Verlag, p 343-363.
Chen F, Kishida T, Duh FM, Renbaum P, Orcutt ML, Schmidt L & Zbar B (1995) Suppression of growth of renal carcinoma cells by the von Hippel-Lindau tumor suppressor gene. Cancer Res 55:4804-4807.
Chia SK, Wykoff CC, Watson PH, Han C, Leek RD, Pastorek J, Gatter KC, Ratcliffe P & Harris AL (2001) Prognostic significance of a novel hypoxia-regulated marker, carbonic anhydrase IX, in invasive breast carcinoma. J Clin Oncol 19:3660-3668.
Christie KN, Thomson C, Xue L, Lucocq JM & Hopwood D (1997) Carbonic anhydrase isoenzymes I, II, III, and IV are present in human oesophageal epithelium. J Histochem Cytochem 45:35-40.
Davenport HW (1939) Gastric carbonic anhydrase. J Physiol (London) 97:32-43. Davenport HW & Fisher RB (1938) Carbonic anhydrase in the gastrointestinal mucosa. J Physiol
(London) 94:16P-17P. Dawson DC (1991) Ion channels and colonic salt transport. Annu Rev Physiol 53:321-339. de Kok TM, van Faassen A, Glinghammer B, Pachen DM, Eng M, Rafter JJ, Baeten CG, Engels
LG & Kleinjans JC (1999) Bile acid concentrations, cytotoxicity, and pH of fecal water from patients with colorectal adenomas. Dig Dis Sci 44:2218-2225.
Dodgson SJ (1991) Liver mitochondrial carbonic anhydrase (CA V), gluconeogenesis, and ureagenesis in the hepatocyte. In: Dodgson SJ, Tashian RE, Gros G & Carter ND (eds) The Carbonic Anhydrases. Cellular Physiology and Molecular Genetics. Plenum Press, New York, p 297-306.
Dodgson SJ, Forster RE, Schwed DA & Storey BT (1983) Contribution of matrix carbonic anhydrase to citrulline synthesis in isolated guinea pig liver mitochondria. J Biol Chem 258:7696-7701.

59Dodgson SJ, Forster RE & Storey BT (1984) The role of carbonic anhydrase in hepatocyte
metabolism. Ann NY Acad Sci USA 429:516-524. Dong B, Horowitz DS, Kobayashi R & Krainer AR (1993) Purification and cDNA cloning of HeLa
cell p54nrb, a nuclear protein with two RNA recognition motifs and extensive homology to human splicing factor PSF and Drosophila NONA/BJ6. Nucleic Acids Res 21:4085-4092.
Duan DR, Pause A, Burgess WH, Aso T, Chen DY, Garrett KP, Conaway RC, Conaway JW, Linehan WM & Klausner RD (1995) Inhibition of transcription elongation by the VHL tumor suppressor protein. Science 269:1402-1406.
Earnhardt JN, Qian M, Tu C, Lakkis MM, Bergenhem NC, Laipis PJ, Tashian RE & Silverman DN (1998) The catalytic properties of murine carbonic anhydrase VII. Biochemistry 37:10837-10845.
Edsall JT (1968) The carbonic anhydrases of erythrocytes. Harvey Lect 62:191-230. Elder JA & Lehninger AL (1973) Respiration-dependent transport of carbon dioxide into rat liver
mitochondria. Biochemistry 12:976-982. Eriksson AE & Liljas A (1991) X-ray crystallographic studies of carbonic anhydrase isoenzymes I,
II, and III. In: Dodgson SJ, Tashian RE & Carter ND (eds) The Carbonic Anhydrases. Cellular physiology and molecular genetics. Plenum Press, New York, p. 33-48.
Erlinger S (1996) Reviw article: new insights into the mechanisms of hepatic transport and bile secretion. J Gastroenterol Hepatol 11:575-579.
Faeron ER & Volgenstein B (1990) A genetic model for colorectal tumorigenesis. Cell 61:759-767. Feldman GM (1994) HCO3
- secretion by rat distal colon: effects of inhibitors and extracellular Na+. Gastroenterology 107:329-338.
Feldstein JB & Silverman DN (1984) Purification and characterization of carbonic anhydrase from the saliva of the rat. J Biol Chem 259:5447-5453.
Fernley RT (1991a) Carbonic anhydrases secreted in the saliva. In: Dodgson SJ, Tashian RE, Gros G & Carter ND (eds) The Carbonic Anhydrases. Cellular physiology and molecular genetics. Plenum Press, New York, p 365-373.
Fernley RT (1991b) Secreted carbonic anhydrases. In: Botré F, Gros G & Storey BT (eds) Carbonic anhydrase. From biochemistry and genetics to physiology and clinical medicine. VCH Verlagsgesellschaft, Weinheim, p 178-185.
Fernley RT, Wright RD & Coghlan JP (1979) A novel carbonic anhydrase from the ovine parotid gland. FEBS Lett 105:299-302.
Fernley RT, Wright RD & Coghlan JP (1988) Complete aminoacid sequence of ovine salivary carbonic anhydrase. Biochemistry 27:2815-2820.
Fleming RE, Crouch EC, Ruzicka CA & Sly WS (1993) Pulmonary carbonic anhydrase IV: developmental regulation and cell-specific expression in the alveolar capillary endothelium. Am J Physiol 265:L627-L635.
Fleming RE, Moxley MA, Waheed A, Crouch EC, Sly WS & Longmore WJ (1994) Carbonic anhydrase II expression in rat type II pneumocytes. Am J Respir Cell Mol Biol 10:499-505.
Fleming RE, Parkkila S, Parkkila A-K, Rajaniemi H, Waheed A & Sly WS (1995) Carbonic anhydrase IV in rat and human gastrointestinal tract: Regional, cellular, and subcellular localization. J Clin Invest 96:2907-2913.
Flemström G & Isenberg JI (2001) Gastroduodenal mucosal alkaline secretion and mucosal protection. News Physiol Sci 16:23-28.
Frigeri A, Gropper M, Turck CW & Verkman AS (1995) Immunolocalization of the mercurial-intensitive water channel and glycerol intrinsic protein in epithelial cell plasma membranes. Proc Natl Acad Sci USA 92:4328-4331.
Fujikawa-Adachi K, Nishimori I, Sakamoto S, Morita M, Onishi S, Yonezava S & Hollingsworth MA (1999b) Identification of carbonic anhydrase IV and VI mRNA expression in human pancreas and salivary glands. Pancreas 18:329-335.
Fujikawa-Adachi K, Nishimori I, Taguchi T & Onishi S (1999a) Human mitochondrial carbonic anhydrase VB. cDNA cloning, mRNA expression, subcellular localization, and mapping to chromosome X. J Biol Chem 274:21228-21233.

60Fujikawa-Adachi K, Nishimori I, Taguchi T & Onishi S (1999c) Human carbonic anhydrase XIV
(CA 14): cDNA cloning, mRNA expression, and mapping to chromosome 1. Genomics 61:74-81.
Furimsky M, Moon TW & Perry SF (2000) Evidence for the role of a Na+/HCO3- cotransporter in
trout hepatocyte pHi regulation. J Exp Biol 14:2201-2208. Geibel JP, Rajendran VM & Binder HJ (2001) Na+ -dependent fluid absorption in intact perfused
rat colonic crypts. Gastroenterology 120:144-150. Ghandour MS, Langley OK, Zhu XL, Waheed A & Sly WS (1992) Carbonic anhydrase IV on brain
capillary endothelial cells: a marker associated with the blood-brain barrier. Proc Natl Acad Sci USA 89:6823-6827.
Giatromanolaki A, Koukourakis MI, Sivridis E, Pastorek J, Wykoff CC, Gatter KC & Harris AL (2001) Expression of hypoxia-inducible carbonic anhydrase-9 relates to angiogenic pathways and independently to poor outcome in non-small cell lung cancer. Cancer Res 61:7992-7998.
Gleeson D, Smith ND & Boyer JL (1989) Bicarbonate-dependent and –independent intracellular pH regulatory mechanisms in rat hepatocytes. Evidence for Na+-HCO3
- cotransport. J Clin Invest 84:312-321.
Gramlich TL, Hennigar RA, Spicer SS & Schulte BA (1990) Immunohistochemical localization of sodium-potassium-stimulated adenosine triphosphatase and carbonic anhydrase in human colon and colonic neoplasms. Arch Pathol Lab Med 114:415-419.
Greger R, Bleich M, Leipziger J, Ecke D, Mall M & Kunzelmann K (1997) Regulation of ion transport in colonic crypts. News Physiol Sci 12:62-66.
Griffiths JR (1991) Are cancer cells acidic? Br J Cancer 64:425-427. Hageman GS, Zhu XL, Waheed A & Sly WS (1991) Localization of carbonic anhydrase IV in a
specific capillary bed of the human eye. Proc Natl Acad Sci USA 88:2716-2720. Hamilton SR, Farber JL & Rubin E (1998) The gastrointestinal tract. The large intestine. Malignant
tumors. In: Rubin E & Farber JL (eds) Pathology. Third edition. Philadelphia: Lippincott, p 742-747.
Hasegawa H, Lian SC, Finkbeiner WE & Verkman AS (1994) Extrarenal tissue distribution of CHIP28 water channels by in situ hybridisation and antibody staining. Am J Physiol 266:C893-C903.
Hazen SA, Waheed A, Sly WS, LaNoue KF & Lynch CJ (1996) Differentiation-dependent expression of CA V and the role of carbonic anhydrase isozymes in pyruvate carboxylation in adipocytes. FASEB J 10:481-490.
Helm JF (1989) Role of saliva in esophageal function and disease. Dysphagia 4:76-84. Helm JF, Dodds WJ, Hogan WJ, Soergel KH, Egide MS & Wood CM (1982) Acid neutralizing
capacity of human saliva. Gastroenterology 83:69-74. Helm JF, Dodds WJ, Pelc LR, Palmer DW, Hogan WJ & Teeter BC (1984) Effect of esophageal
emptying and saliva on clearance of acid from the esophagus. N Engl J Med 310:284-288. Henriques OM (1928) Die Bindungsweise des Kohlendioxyds im Blute. Biochem Z 200:1-24. Hewett-Emmet D & Tashian RE (1991) Structure and evolutionary origins of the carbonic
anhydrase multigene family. In: Dodgson SJ, Tashian RE, Gros G & Carter ND (eds) The Carbonic Anhydrases. Cellular Physiology and Molecular Genetics. Plenum Press, New York, p 15-32.
Hewett-Emmett D & Tashian RE (1996) Functional diversity, conservation, and convergence in the evolution of the alpha-, beta-, and gamma-carbonic anhydrase gene families. Mol Phylogenet Evol 5:50-77.
Holappa K, Mustonen M, Parvinen M, Vihko P, Rajaniemi H & Kellokumpu S (1999) Primary structure of a sperm cell anion exchanger and its messenger ribonucleic acid expression during spermatogenesis. Biol Reprod 61:981-986.
Holmes RS (1976) Mammalian carbonic anhydrase isozymes: Evidence for a third locus. J Exp Zool 197:289-295.
Hu PY, Roth DE, Skaggs LA, Venta PJ, Tashian RE, Guibaud P & Sly WS (1992) A splice junction mutation in intron 2 of the carbonic anhydrase II gene of osteopetrosis patients from Arabic countries. Hum Mutat 1:288-292.

61Huebert RC, Splinter PL, Garcia F, Marinelli RA & LaRusso NF (2002) Expression and
localization of aquaporin water channels in rat hepatocytes. J Biol Chem 277:22710-22717. Iino H, Jass JR, Simms LA, Young J, Leggett B, Ajioka Y & Watanabe H (1999) DNA
microsatellite instability in hyperplastic polyps, serrated adenomas, and mixed polyps: a mild mutator pathway for colorectal cancer? J Clin Pathol 52:5-9.
Ishibashi K, Sasaki S, Saito F, Ikeuchi T & Marumo F (1995) Structure and chromosomal localization of human water channel (AQP3) gene. Genomics 27:352-354.
Ishiguro H, Naruse S, San Román JI, Case M & Steward MC (2001) Pancreatic ductal bicarbonate secretion: past, present and future. J Pancreas 2:192-197.
Ivanov SV, Kuzmin I, Wei M-H, Pack S, Geil L, Johnson BE, Stanbridge EJ & Lerman MI (1998) Down-regulation of transmembrane carbonic anhydrases in renal cell carcinoma cell lines by wild-type von Hippel-Lindau transgenes. Proc Natl Acad Sci USA 95:12596-12601.
Ivanov S, Liao SY, Ivanova A, Danilkovitch-Miagkova A, Tarasova N, Weirich G, Merrill MJ, Proescholdt MA, Oldfield EH, Lee J, Zavada J, Waheed A, Sly W, Lerman MI & Stanbridge EJ (2001) Expression of hypoxia-inducible cell-surface transmembrane carbonic anhydrases in human cancer. Am J Pathol 158:905-919.
Jeffery S & Carter N (1980) Isoenzymes of human muscle carbonic anhydrase III generated by thiol interaction. Biochem Soc Trans 8:643.
Jeffery S, Edwards Y & Carter N (1980) Distribution of CA III in fetal and adult human tissue. Biochem Genet 18:843-849.
Juvonen T, Parkkila S, Parkkila A-K, Niemelä O, Lajunen LHJ, Kairaluoma MI, Perämäki P & Rajaniemi H (1994) High activity carbonic anhydrase isoenzyme (CA II) in human gallbladder epithelium. J Histochem Cytochem 42:1393-1397.
Kadoya Y, Kuwahara H, Shimazaki M, Ogawa Y & Yagi T (1987) Isolation of a novel carbonic anhydrase from human saliva and immunohistochemical demonstration of its related isoenzymes in salivary gland. Osaka City Med J 33:99-109.
Kaluz S, Kaluzove M, Opavský R, Pastoreková S, Gibadulinová A, Dequiedt F, Kettman R & Pastorek J (1999) Transcriptional regulation of the MN/CA 9 gene coding for the tumor-associated carbonic anhydrase IX. Identification and characterization of a proximal silencer element. J Biol Chem 274:32588-32595.
Kampman E, Giovannucci E, van 't Veer P, Rimm E, Stampfer MJ, Colditz GA, Kok FJ & Willett WC (1994) Calcium, vitamin D, dairy foods, and the occurrence of colorectal adenomas among men and women in two prospective studies. Am J Epidemiol 139:16-29.
Kannan KK, Petef M, Fridborg K, Cid-Dresidner H & Lövgren S (1977) Structure and reaction of carbonic anhydrases: Imidatsole binding to human carbonic anhydrase B and the mechanism of action of carbonic anhydrases. FEBS Lett. 73:115-119.
Karhumaa P, Kaunisto K, Parkkila S, Waheed A, Pastorekova S, Pastorek J, Sly WS & Rajaniemi H (2001) Expression of the transmembrane carbonic anhydrases, CA IX and CA XII, in the human male excurrent ducts. Mol Hum Reprod 7:611-616.
Karhumaa P, Leinonen J, Parkkila S, Kaunisto K, Tapanainen J & Rajaniemi H (2001) The identification of secreted carbonic anhydrase VI as a constitutive glycoprotein of human and rat milk. Proc Natl Acad Sci USA 98:11604-11608.
Kaunisto K (1998) Ion transport proteins in the male reproductive tract [dissertation]. Acta Univ Oul D 444, Univ. of Oulu, Oulu (Finland).
Kaunisto K, Parkkila S, Parkkila A-K, Waheed A, Sly WS & Rajaniemi H (1995) Expression of carbonic anhydrase isoenzymes IV and II in rat epididymal duct. Biol Reprod 52:1350-1357.
Kaunisto K, Parkkila S, Rajaniemi H, Waheed A, Grubb J & Sly WS (2002) Carbonic anhydrase XIV: luminal expression suggests key role in renal acidification. Kidney Int 61:2111-2118.
Kaunisto K, Parkkila S, Tammela T, Rönnberg L & Rajaniemi H (1990) Immunohistochemical localization of carbonic anhydrase isoenzymes in the human male reproductive tract. Histochemistry 94:381-386.
Kelly CD, Carter ND, de Boer P, Jeffery S, Moorman AF & Smith A (1991) Detection of CA III mRNA in rat skeletal muscle and liver by in situ hybridisation. J Histochem Cytochem 39:1243-1247.

62Kennedy ED, Rizzuto R, Theler J-M, Pralong W-F, Bastianutto C, Pozzan T & Wollheim CB
(1996) Glucose-stimulated insulin secretion correlates with changes in mitochondrial and cytosolic Ca2+ in Aequorin-expressing INS-1 cells. J Clin Invest 98:2524-2538.
Khalifah RG (1971) The carbon dioxide hydration activity of carbonic anhydrase. I. Stop-flow kinetic studies on the native human isoenzymes B and C. J Biol Chem 246:2561-2573.
Kim G, Selengut J & Levine RL (2000) Carbonic anhydrase III: the phosphatase activity is extrinsic. Arch Biochem Biophys 377:334-340.
Kivelä J, Parkkila S, Parkkila A-K & Rajaniemi H (1999) A low concentration of carbonic anhydrase isoenzyme VI in whole saliva is associated with caries prevalence. Caries Res 33:178-184.
Kivelä J, Parkkila S, Waheed A, Parkkila A-K, Sly WS & Rajaniemi H (1997) Secretory carbonic anhydrase isoenzyme (CA VI) in human serum. Clin. Chem 43:2318-2322.
Koyama Y, Yamamoto T, Kondo D, Funaki H, Yaoita E, Kawasaki K, Sato N, Hatakeyama K & Kihara I (1997) Molecular cloning of a new aquaporin from rat pancreas and liver. J Biol Chem 272:30329-30333.
Krueger NX & Saito H (1992) A human transmembrane protein-tyrosine.phosphatase, PTP zeta, is expressed in brain and has an N-terminal receptor domain homologous to carbonic anhydrases. Proc Natl Acad Sci USA 89:7417-7421.
Kuismanen SA, Holmberg MT, Salovaara R, Schweizer P, Aaltonen LA, de La Chapelle A, Nyström-Lahti M & Peltomäki P (1999) Epigenetic phenotypes distinguish microsatellite-stable and -unstable colorectal cancers. Proc Natl Acad Sci USA 96:12661-12666.
Kumpulainen T (1984) Immunohistochemical localization of human carbonic anhydrase isozymes. Ann NY Acad Sci 429:359-368.
Kumpulainen T & Jalovaara P (1981) Immunohistochemical localization of carbonic anhydrase isoenzymes in the human pancreas. Gastroenterology 80:796-799.
Kumpulainen T, Rajaniemi H, Myllylä VV & Korhonen LK (1985) A single-step solid phase radioimmunoassay for quantifying human carbonic anhydrase I and II in cerebrospinal fluid. Clin Chim Acta 150:205-212.
Kunzelmann K & Mall M (2002) Electrolyte transport in the mammalian colon: mechanisms and implications for disease. Physiol Rev 82:245-289.
Kyzer S, Begin LR, Gordon PH & Mitmaker B (1992) The care of patients with colorectal polyps that contain invasive adenocarcinoma. Endoscopic polypectomy or colectomy? Cancer 70:2044-2050.
Laemmli UK (1970) Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 227:680-685.
Lai LW, Chan DM, Erickson RP, Hsu SJ & Lien YH (1998) Correction of renal tubular acidosis in carbonic anhydrase deficient mice with gene therapy. J Clin Invest 101:1320-1325.
Lairmore TC & Norton JA (1997) Advances in molecular genetics. Am J Surg 173:37-43. Lakkis MM, Bergenhem NC & Tashian RE (1996) Expression of mouse carbonic anhydrase VII in
E. Coli and demonstration of its CO2 hydrase activity. Biochem Biophys Res Commun 226:268-272.
Lakkis MM, O’Shea KS & Tashian RE (1997) Differential expression of the carbonic anhydrase genes for CA VII (Car7) and CA-RP VIII (Car8) in mouse brain. J Histochem Cytochem 45:657-662.
Latif F, Tory K, Gnarra J, Yao M, Duh F-M, Orcutt ML, Stackhouse T, Kuzmin I, Modi W, Geil L, Schmidt L, Zhou F, Li H, Wei MH, Chen F, Glenn G, Choyke P, Walther MM, Weng Y, Duan D-SR, Dean M, Glavač D, Richards FM, Crossey PA, Ferguson-Smith MA, Le Paslier D, Chumakov I, Cohen D, Chinault AC, Maher ER, Linehan WM, Zbar B & Lerman MI (1993) Identification of the von Hippel-Lindau disease tumor suppressor gene. Science 260:1317-1320.
Leinonen J, Kivelä J, Parkkila S, Parkkila A-K & Rajaniemi H (1999) Salivary carbonic anhydrase isoenzyme VI is located in the human enamel pellicle. Caries Res. 33:185-190.
Leinonen J, Parkkila S, Kaunisto K, Koivunen P & Rajaniemi H (2001) Secretion of carbonic anhydrase isoenzyme VI (CA VI) from human and rat lingual serous von Ebner's glands. J Histochem Cytochem 49:657-662.

63Leppilampi M, Koistinen P, Savolainen ER, Hannuksela J, Parkkila AK, Niemela O, Pastorekova
S, Pastorek J, Waheed A, Sly WS, Parkkila S & Rajaniemi H (2002) The expression of carbonic anhydrase II in hematological malignancies. Clin Cancer Res 8:2240-2245.
Lesburg CA, Huang C, Christianson DW & Fierke CA (1997) Histidine --> carboxamide ligand substitutions in the zinc binding site of carbonic anhydrase II alter metal coordination geometry but retain catalytic activity. Biochemistry 36:15780-15791.
Levi F, Lucchini F & La Vecchia C (1994) Worldwide patterns of cancer mortality, 1985-89. Eur J Cancer Prev 3:109-143.
Levy JB, Canoll PD, Silvennoinen O, Barnea G, Morse B, Honegger AM, Huang JT, Cannizzaro LA, Park SH, Druck T, Huebner K, Sap J, Ehrlich M, Masacchio JM & Schlessinger J (1993) The cloning of a receptor-type protein tyrosine phosphatase expressed in the central nervous system. J Biol Chem 268:10573-10581.
Li X, Alvarez B, Casey JR, Reithmeier RA & Fliegel L (2002) Carbonic anhydrase II binds to and enhances activity of the Na+/H+ exchanger. J Biol Chem 277:36085-36091.
Li G, Passebosc-Faure K, Lambert C, Gentil-Perret A, Blanc F, Oosterwijk E, Mosnier J-F, Genin C & Tostain J (2001) The expression of G250/MN/CA9 antigen by flow cytometry: its possible implication for detection of micrometastatic renal cancer cells. Clin Cancer Res 7:89–92.
Liao SY, Brewer C, Závada J, Pastorek J, Pastoreková S, Manetta A, Berman ML, DiSaia PJ & Stanbridge EJ (1994) Identification of the MN antigen as a diagnostic biomarker of cervical intraepithelial squamous and glandular neoplasia and cervical carcinomas. Am J Pathol 145:598-609.
Lindskog S, Engberg P, Forsman C, Ibrahim SA, Jonsson B-H, Simonsson I & Tibell L (1984) Kinetics and mechanism of carbonic anhydrase isoenzymes. Ann NY Acad Sci 429:61-75.
Lindskog S (1982) Carbonic anhydrase. In: Eichhorn GL & Marzilli L (eds) Advances in Inorganic biochemistry, Vol.4. Elsevier/North Holland, New York: 115-170.
Ling B, Bergenhem NCH, Dodgson SJ, Forster RE & Tashian RE (1994) Determination of mRNA levels for six carbonic anhydrase isozymes in rat lung. Isozyme Bull 28:32.
Lipkin M (1974) Phase 1 and phase 2 proliferative lesions of colonic epithelial cells in diseases leading to colonic cancer. Cancer 34:suppl:878-888.
Loncaster JA, Harris AL, Davidson SE, Logue JP, Hunter RD, Wycoff CC, Pastorek J, Ratcliffe PJ, Stratford IJ & West CM (2001) Carbonic anhydrase (CA IX) expression, a potential new intrinsic marker of hypoxia: correlations with tumor oxygen measurements and prognosis in locally advanced carcinoma of the cervix. Cancer Res 61:6394-6399.
Longmuir IS, Forster RE II & Woo CY (1966) Diffusion of carbon dioxide through thin layers of solution. Nature (London) 209:393-394.
Lovejoy DA, Hewett-Emmett D, Porter CA, Cepoi D, Sheffield A, Vale WW & Tashian RE (1998) Evolutionary concerved, “acatalytic” carbonic anhydrase-related protein XI contains a sequence motif present in the neuropeptide sauvagine: the human CA-RP XI gene (CA11) is embedded between the secretor gene cluster and the DBP gene at 19q13.3. Genomics 54:484-493.
Lönnerholm G, Selking Ö & Wistrand PJ (1985) Amount and distribution of carbonic anhydrases CA I and CA II in the gastrointestinal tract. Gastroenterology 88:1151-1161.
Ma T & Verkman AS (1999) Topical review: Aquaporin water channels in gastrointestinal physiology. J Physiol 517:317-326.
MacDonald MJ (1995) Feasibility of a mitochondrial pyruvate malate shuttle in pancreatic islets. Further implication of cytosolic NADPH in insulin secretion. J Biol Chem 270:20051-20058.
MacDonald MJ & Chang CM (1985) Do pancreatic islets contain significant amounts of phosphoenolpyruvate carboxykinase or ferroactivator activity? Diabetes 34:246-250.
Mallis RJ, Hamann MJ, Zhao W, Zhang T, Hendrich S & Thomas JA (2002) Irreversible thiol oxidation in carbonic anhydrase III: protection by S-glutathiolation and detection in aging rats. Biol Chem 383:649-662.
Mallis RJ, Poland BW, Chatterjee TK, Fisher RA, Darmawan S, Honzatko RB & Thomas JA (2000) Crystal structure of S-glutathiolated carbonic anhydrase III. FEBS Lett 482:237-241.
Maren TH (1967) Carbonic anhydrase: chemistry, physiology, and inhibition. Physiol Rev 47:595-781.

64Martinez-Zaguilan R, Seftor EA, Seftor RE, Chu YW, Gillies RJ & Hendrix MJ (1996) Acidic pH
enhances the invasive behavior of human melanoma cells. Clin Exp Metastasis 14:176-186. Maurel P, Rauch U, Flad M, Margolis RK & Margolis RU (1994) Phosphacan, a chondroitin
sulfate proteoglycan of brain that interacts with neurons and neural cell-adhesion molecules, is an extracellular variant of receptor-type protein tyrosine phosphatase. Proc Natl Acad Sci USA 91:2512-2516.
Maxwell PH, Wiesener MS, Chang G-W, Clifford SC, Vaux EC, Cockman ME, Wykoff CC, Pugh CW, Maher ER & RatcliffePJ (1999) The tumour suppressor protein VHL targets hypoxia-inducible factors for oxygen-dependent proteolysis. Nature 399:271-275.
McKiernan JM, Buttyan R, Bander NH, de la Taille A, Stifelman MD, Emanuel ER, Bagiella E, Rubin MA, Katz AE, Olsson CA & Sawczuk IS (1999) The detection of renal carcinoma cells in the peripheral blood with an enhanced reverse transcriptate-polymerase chain reaction assay for MN/CA9. Cancer 86:492-497.
McKiernan JM, Buttyan R, Bander NH, Stifelman MD, Katz AE, Chen MW, Olsson CA & Sawczuk IS (1997) Expression of the tumor-associated gene MN: a potential biomarker for human renal cell carcinoma. Cancer Res 57:2362-2365.
Meldrum NU & Roughton FJW (1932) Some properties of carbonic anhydrase, the carbon dioxide enzyme present in blood. J Physiol (London) 75:15-16.
Meldrum NU & Roughton FJW (1933) Carbonic anhydrase. Its preparation and properties. J Physiol (London) 80:113-142.
Metcalfe HK, Monson JP, Drew PJ, Iles RA, Carter ND & Cohen RD (1985) Inhibition of gluconeogenesis and urea synthesis in isolated rat hepatocytes by acetazolamide. Biochem Soc Trans 13:255.
Meyers RL & Orlando RC (1992) In vivo bicarbonate secretion by human esophagus. Gastroenterology 103:1174-1178.
Mitelman F (1994) Chromosomes, genes, and cancer. CA Cancer J Clin 44:133-135. Montgomery JC, Venta PJ, Eddy RL, Fukushima Y-S, Shows TB & Tashian RE (1991)
Characterization of the human gene for a newly discovered carbonic anhydrase, CA VII, and its localization to chromosome 16. Genomics 11:835-848.
Mori K, Ogawa Y, Ebihara K, Tamura N, Tashiro K, Kuwahara T, Mukoyama M, Sugawara A, Ozaki S, Tanaka I & Nakao K (1999) Isolation and characterization of CA XIV, a novel membrane-bound carbonic anhydrase from mouse kidney. J Biol Chem 274:15701-15705.
Mori M, Staniunas RJ, Barnard GF, Jessup JM, Steele GD Jr. & Chen LB (1993) The significance of carbonic anhydrase expression in human colorectal cancer. Gastroenterology 105:820-826.
Morson BC, Bussey HJR & Day DW (1983) Adenomas of large bowel. Cancer Surveys 3:451-477. Morson BC, Dawson IMP, Day DW, Jass JR, Price AB & Williams GT (1990) Morson's and
Dawson's Gastrointestinal Pathology, 3rd ed. Blackwell Scientific, Oxford, p 563-596. Muller M & Jansen PL (1997) Molecular aspects of hepatobiliary transport. Am J Physiol
272:G1285-G1303. Murakami H & Sly WS (1987) Purification and characterization of human salivary carbonic
anhydrase. J Biol Chem 262:1382-1388. Mühlhauser J, Crescimanno C, Rajaniemi H, Parkkila S, Milovanov AP, Castellucci M &
Kaufmann P (1994) Immunohistochemistry of carbonic anhydrase in human placenta and fetal membranes. Histochemistry 101:91-98.
Mäkinen MJ, George SM, Jernvall P, Mäkelä J, Vihko P & Karttunen TJ (2001) Colorectal carcinoma associated with serrated adenoma - prevalence, histological features, and prognosis. J Pathol 193:286-294.
Nagao Y, Platero JS, Waheed A & Sly WS (1993) Human mitochondrial carbonic anhydrase: cDNA cloning, expression, subcellular localization, and mapping to chromosome 16. Proc Natl Acad Sci USA 90:7623-7627.
Nakai H, Byers MG, Venta PJ, Tashian RE & Shows TB (1987) The gene for human carbonic anhydrase II (CA2) is located at chromosome 8q22. Cytogenet Cell Genet 44:234-235.
Nakhoul NL, Davis BA, Romero MF & Boron WF (1998) Effect of expressing the water channel aquaporin-1 on the CO2 permeability of Xenopus oocytes. Am J Physiol 274:C543-C548.

65Nishita T & Matsushita H (1989) Immunocytochemical localization of carbonic anhydrase isozyme
III in equine thymus. Histochemistry 91:39-42. O'Brien P, Rosen S, Trencis-Buck L & Silen W (1977) Distribution of carbonic anhydrase within
the gastric mucosa. Gastroenterology 72:870-874. Ogawa Y, Chang CK, Kuwahara H, Hong SS, Toyosawa S & Yagi T (1992) Immunoelectron
microscopy of carbonic anhydrase isozyme VI in rat submandibular gland: comparison with isozymes I and II. J Histochem Cytochem 40:807-817.
Ogawa Y, Hong SS, Toyosawa S, Kuwahara H, Shimazaki M & Yagi T J (1993) Immunoelectron microscopy of carbonic anhydrase isozyme VI in human submandibular gland: comparison with isozymes I and II. Histochem Cytochem 41:343-351. Ogawa Y, Toyosawa S, Inagaki T, Hong SS & Ijuhin N (1995) Carbonic anhydrase isozyme VI in
rat lacrimal gland. Histochem Cell Biol 103:387-394. Okamoto N, Fujikawa-Adachi K, Nishimori I, Taniuchi K & Onishi S (2001) cDNA sequence of
human carbonic anhydrase-related protein, CA-RP X: mRNA expressions of CA-RP X and XI in human brain. Biochim Biophys Acta 1518:311-316.
Okuyama T, Batanian JR & Sly WS (1993) Genomic organization and localization of gene for human carbonic anhydrase IV to chromosome 17q. Genomics 16:678-684.
Okuyama T, Sato S, Zhu XL, Waheed A & Sly WS (1992) Human carbonic anhydrase IV: cDNA cloning, sequence comparison, and expression in COS cell membranes. Proc Natl Acad Sci USA 89:1315-1319.
Opavský R, Pastoreková S, Zelník V, Gibadulinová A, Stanbridge EJ, Závada J, Kettman R & Pastorek J (1996) Human MN/CA9 gene, a novel member of the carbonic anhydrase family: structure and exon to protein domain relationships. Genomics 33:480-487.
Ortova Gut M, Parkkila S, Vernerová Z, Rohde E, Závada J, Höcker M, Pastorek J, Karttunen T, Gibadulinová A, Závadová Z, Knobeloch K-P, Wiedenmann B, Svoboda J, Horak I & Pastoreková S (2002) Gastric hyperplasia in mice with targeted disruption of the carbonic anhydrase gene Car9. Gastroenterology 123:1889-1903.
Otori K, Oda Y, Sugiyama K, Hasebe T, Mukai K, Fujii T, Tajiri H, Yoshida S, Fukushima S & Esumi H (1997) High frequency of K-ras mutations in human colorectal hyperplastic polyps. Gut 40:660-663.
Parkkila A-K, Herva R, Parkkila S & Rajaniemi H (1995) Immunohistochemical demonstration of human carbonic anhydrase isoenzyme II in brain tumours. Histochem J 27:974-982.
Parkkila A-K, Parkkila S, Juvonen T & Rajaniemi H (1993) Carbonic anhydrase isoenzymes II and I are present in the zona glomerulosa cells of the human adrenal gland. Histochemistry 99:37-41.
Parkkila A-K, Parkkila S & Rajaniemi H (1996) Carbonic anhydrase isoenzyme II is located in corticotrophs of the human pituitary gland. J Histochem Cytochem 44:245-250.
Parkkila A-K, Parkkila S, Reunanen M, Niemelä O, Tuisku S, Rautakorpi I & Rajaniemi H (1997) Carbonic anhydrase II in the cerebrospinal fluid: its value as a disease marker. Eur J Clin Invest 27:392-397.
Parkkila A-K, Parkkila S, Serlo W, Reunanen M, Vierjoki T & Rajaniemi H (1994) A competitive dual-label time-resolved immunofluorometric assay for simultaneous detection of carbonic anhydrase I and II in cerebrospinal fluid. Clin Chim Acta 230:81-89.
Parkkila A-K, Scarim AL, Parkkila S, Waheed A, Corbett JA & Sly WS (1998) Expression of carbonic anhydrase V in pancreatic β-cells suggests role for mitochondrial carbonic anhydrase in insulin secretion. J Biol Chem 273:24620-24623.
Parkkila S (2000c) Roles of carbonic anhydrases in the alimentary tract. In: Chegwidden WR, Carter ND & Edwards YH (eds) The Carbonic Anhydrases New Horizons. 1st ed. Basel: Birkhäuser Verlag, p 461-474.
Parkkila S, Halsted CH, Väänänen HK & Niemelä O (1999) Expression of testosterone-dependent enzyme, carbonic anhydrase III, and oxidative stress in experimental alcoholic liver disease. Digest Dis Sci 44:2205-2213.
Parkkila S, Kaunisto K, Kellokumpu S & Rajaniemi H (1991a) A high activity carbonic anhydrase isoenzyme (CA II) is present in mammalian spermatozoa. Histochemistry 95:477-482.

66Parkkila S, Kaunisto K & Rajaniemi H (1991b) Location of the carbonic anhydrase isoenzymes VI
and II in human salivary glands by immunohistochemistry. In: Botré F, Gros G & Storey BT (eds) Carbonic anhydrase. From Biochemistry and Genetics to Physiology and Clinical Medicine. VCH Verlagsgesellschaft, Weinheim, p 254-257.
Parkkila S, Kaunisto K, Rajaniemi L, Kumpulainen T, Jokinen K & Rajaniemi H (1990) Immunohistochemical localization of carbonic anhydrase isoenzymes VI, II and I in human parotid and submandibular glands. J Histochem Cytochem 38:941-947.
Parkkila S & Parkkila A-K (1996) Carbonic anhydrase in the alimentary tract. Roles of the different isozymes and salivary factors in the maintenance of optimal conditions in the gastrointestinal canal. Scand J Gastroenterol 31:305-317.
Parkkila S, Parkkila AK, Juvonen T, Lehto VP & Rajaniemi H (1995) Immunohistochemical demonstration of the carbonic anhydrase isoenzymes I and II in pancreatic tumours. Histochem J 27:133-138.
Parkkila S, Parkkila A-K, Juvonen T & Rajaniemi H (1994) Distribution of the carbonic anhydrase isoenzymes I, II, and VI in the human alimentary tract. Gut 35:646-650.
Parkkila S, Parkkila A-K, Juvonen T, Waheed A, Sly WS, Saarnio J, Kaunisto K, Kellokumpu S & Rajaniemi H (1996) Membrane-bound carbonic anhydrase IV is expressed in the luminal plasma membrane of the human gallbladder epithelium. Hepatology 24:1104-1108.
Parkkila S, Parkkila A-K, Kaunisto K, Waheed A, Sly WS & Rajaniemi H (1993a) Location of a membrane-bound carbonic anhydrase isoenzyme (CA IV) in the human male reproductive tract. J Histochem Cytochem 41:751-757.
Parkkila S, Parkkila A-K, Lehtola J, Reinilä A, Södervik H-J, Rannisto M & Rajaniemi H (1997) Salivary carbonic anhydrase protects gastroesophageal mucosa from acid injury. Dig Dis Sci 42:1013-1019.
Parkkila S, Parkkila AK, Rajaniemi H, Shah GN, Grubb JH, Waheed A & Sly WS (2001) Expression of membrane-associated carbonic anhydrase XIV on neurons and axons in mouse and human brain. Proc Natl Acad Sci USA 98:1918-1923.
Parkkila S, Parkkila AK, Saarnio J, Kivela J, Karttunen TJ, Kaunisto K, Waheed A, Sly WS, Tureci O, Virtanen I & Rajaniemi H (2000b) Expression of the membrane-associated carbonic anhydrase isozyme XII in the human kidney and renal tumors. J Histochem Cytochem 48:1601-1608.
Parkkila S, Parkkila A-K, Vierjoki T, Ståhlberg T & Rajaniemi H (1993b) Competitive time-resolved immunofluorometric assay for quantifying carbonic anhydrase VI in saliva. Clin Chem 39:2154-2157.
Parkkila S, Rajaniemi H, Parkkila A-K, Kivelä J, Waheed A, Pastoreková S, Pastorek J & Sly WS (2000a) Carbonic anhydrase inhibitor suppresses invasion of renal cancer cells in vitro. Proc Natl Acad Sci USA 97:2220-2224.
Pastorek J, Pastoreková S, Callebaut I, Mornon JP, Zelník V, Opavský R, Zat’ovicová M, Liao S, Portetelle D, Stanbridge EJ, Závada J, Burny A & Kettmann R (1994) Cloning and characterization of MN, a human tumor-associated protein with a domain homologous to carbonic anhydrase and a putative helix-loop-helix DNA binding segment. Oncogene 9:2788-2888.
Pastoreková S, Parkkila S, Parkkila A-K, Opavský R, Zelnik V, Saarnio J & Pastorek J (1997) Carbonic anhydrase IX, MN/CA IX: Analysis of stomach complementary DNA sequence and expression in human and rat alimentary tracts. Gastroenterology 112:398-408.
Pastoreková S, Zavadova Z, Kostal M, Babusikova O & Zàvada J (1992) A novel quasi-viral agent, MaTu is a two-component system. Virology 187:620-626.
Peles E, Nativ M, Campbell PL, Sakurai T, Martinez R, Lev S, Clary DO, Schilling J, Barnea G, Plowman GD, Grumet M & Schlessinger J (1995) The carbonic anhydrase domain of receptor tyrosine phosphatase β is a functional ligand for the axonal cell recognition molecule contaction. Cell 82:251-260.
Pollard CW, Nivatvongs S, Rojanasakul A, Reiman HM & Dozois RR (1992) The fate of patients following polypectomy alone for polyps containing invasive carcinoma. Dis Colon Rectum 35:933-937.

67Ponz de Leon M, Antonioli A, Ascari A, Zanghieri G & Sacchetti C (1987) Incidence and familial
occurrence of colorectal cancer and polyps in a health-care district of northern Italy. Cancer 60:2848-2859.
Ponz de Leon M & Di Gregorio C (2001) Pathology of colorectal cancer. Dig Liver Dis 33:372-388.
Ponz de Leon M, Sassatelli R, Sacchetti C, Zanghieri G, Scalmati A & Roncucci L (1989) Familial aggregation of tumors in the three-year experience of a population-based colorectal cancer registry. Cancer Res 49:4344-4348.
Potter JD (1999) Colorectal cancer: molecules and populations. J Natl Cancer Inst 91:916-32. Rajendran VM & Binder HJ (2000) Characterization and molecular localization of anion
transporters in colonic epithelial cells. Ann NY Acad Sci 915:15-29. Redston M (2001) Carcinogenesis in the GI tract: from morphology to genetics and back again.
Mod Pathol 14:236-245. Register AM, Koester MK & Noltmann EA (1978) Discovery of carbonic anhydrase in rabbit
skeletal muscle and evidence for its identity with “basic muscle protein”. J Biol Chem 253:4143-4152.
Renner EL, Lake JR, Persico M & Scharschmidt BF (1989) Na+-H+ exchange activity in rat hepatocytes: role in regulation of intracellular pH. Am J Physiol 256:G44-G52.
Richardson CT (1985) Pathogenic factors in peptic ulcer disease. Am J Med 79:1-7. Ridderstråle Y, Wistrand PJ, Holm L & Carter ND (2000) Use of carbonic anhydrase II-deficient
mice in uncovering the cellular location of membrane-associated isoforms. In: Chegwidden WR, Carter ND & Edwards YH (eds) The carbonic anhydrases. New horizons. 1st ed. Basel: Birkhäuser Verlag, p 143-155.
Räisänen SR, Lehenkari P, Tasanen M, Rahkila P, Härkönen PL & Väänänen HK (1999) Carbonic anhydrase III protects cells from hydrogen peroxide-induced apoptosis. FASEB J 13:513-522.
Saarnio J, Parkkila S, Parkkila A-K, Haukipuro K, Pastoreková S, Pastorek J, Kairaluoma MI & Karttunen TJ (1998b) Immunohistochemical study of colorectal tumors for expression of a novel transmembrane carbonic anhydrase, MN/CA IX, with potential value as a marker of cell proliferation. Am J Pathol 153:279-285.
Saarnio J, Parkkila S, Parkkila AK, Pastorekova S, Haukipuro K, Pastorek J, Juvonen T & Karttunen TJ (2001) Transmembrane carbonic anhydrase, MN/CA IX, is a potential biomarker for biliary tumours. J Hepatol 35:643-649.
Saarnio J, Parkkila S, Parkkila AK, Waheed A, Casey MC, Zhou XY, Pastorekova S, Pastorek J, Karttunen T, Haukipuro K, Kairaluoma MI & Sly WS (1998a) Immunohistochemistry of carbonic anhydrase isozyme IX (MN/CA IX) in human gut reveals polarized expression in the epithelial cells with the highest proliferative capacity. J Histochem Cytochem 46:497-504.
Sakashita N, Takeya M, Kishida T, Stackhouse TM, Zbar B & Takahashi K (1999) Expression of von Hippel-Lindau protein in normal and pathological human tissues. Histochem J 31:133-144.
Sanyal G & Maren TH (1981) Thermodynamics of carbonic anhydrase catalysis. A comparison between human isoenzymes B and C. J Biol Chem 256:608-612.
Sasaki K, Igarashi S-I, Amasaki T, Amasaki H, Nishita T, Kano Y & Asari M (1993) Comparative immunohistolocalization of carbonic anhydrase isozymes I, II and III in the equine and bovine digestive tract. Histochem J 25:304-311.
Sasano H, Kato K, Nagura H, Parkkila S, Parkkila A-K, Rajaniemi H & Sugai N (1994) Carbonic anhydrases in the human adrenal gland and its disorders: Immunohistochemical and biochemical studies of the enzymes. Endocr Pathol 5:100-106.
Sato A, Spicer SS & Tashian RE (1980) Ultrastructural localization of carbonic anhydrase in gastric parietal cells with the immunoglobulin-enzyme bridge method. Histochem J 12:651-659.
Seidler U, Bachmann O, Jacob P, Christiani S, Blumenstein I & Rossmann H (2001) Na+/HCO3-
cotransport in normal and cystic fibrosis intestine. J Pancreas 2:247-256. Shah GN, Hewett-Emmett D, Grubb JH, Migas MC, Fleming RE, Waheed A & Sly WS (2000)
Mitochondrial carbonic anhydrase VB: differences in tissue distribution and pattern of evolution from those of CA VA suggest distinct physiologiacal roles Proc Natl Acad Sci USA 97:1677-1682.

68Siu IM, Robinson DR, Schwartz S, Kung HJ, Pretlow TG, Petersen RB & Pretlow TP (1999) The
identification of monoclonality in human aberrant crypt foci. Cancer Res 59:63-66. Skaggs LA, Bergenhem NC, Venta PJ & Tashian RE (1993) The deduced amino acid sequence of
human carbonic anhydrase-related protein (CARP) is 98% identical to the mouse homologue. Gene 126:291-292.
Sly WS, Hewett-Emmett D, Whyte MP, Yu Y-S L & Tashian RE (1983) Carbonic anhydrase II deficiency identified as the primary defect in the autosomal recessive syndrome of osteopetrosis with renal tubular acidosis and cerebral calcification. Proc Natl Acad Sci USA 80:2752-2756.
Sly WS & Hu PY (1995a) Human carbonic anhydrases and carbonic anhydrase deficiencies. Annu Rev Biochem 64:375-401.
Sly WS & Hu PY (1995b) The carbonic anhydrase II deficiency syndrome: osteopetrosis with renal tubular acidosis and cerebral calcification. In: Scriver CR, Beaudet AL, Sly WS & Valle D (eds) The Molecular and Metabolic Bases of Inherited Disease, 7th ed. McGraw-Hill, New York, p 4113-4124.
Sly WS, Whyte MP, Sundaram V, Tashian RE, Hewett-Emmett D, Guibaud P, Vainsel M, Baluarte J, Gruskin A, Al-Mosavi M, Sakati N & Ohlsson A (1985) Carbonic anhydrase II deficiency in 12 families with the autosomal recessive syndrome of osteopetrosis with renal tubular acidosis and cerebral calcification. N Engl J Med 313:139-145.
Sok J, Wang XZ, Batchvarova N, Kuroda M, Harding H & Ron D (1999) CHOP-Dependent stress-inducible expression of a novel form of carbonic anhydrase VI. Mol Cell Biol 19:495-504.
Spicer SS, Ge Z-H, Tashian RE, Hazen-Martin DJ & Schulte BA (1990) Comparative distribution of carbonic anhydrase isozymes III and II in rodent tissues. Am J Anat 187:55-64.
Spicer SS, Sens MA, Tashian RE (1982) Immunocytochemical demonstration of carbonic anhydrase in human epithelial cells. J Histochem Cytochem 30:864-873.
Spring KR (1998) Routes and mechanism of fluid transport by epithelia. Annu Rev Physiol 60:105-119.
Steffens MG, Boerman OC, Oosterwijk-Wakka JC, Oosterhof GO, Witjes JA, Koenders EB, Oyen WJ, Buijs WC, Debruyne FM, Corstens FH & Oosterwijk EJ (1997) Targeting of renal cell carcinoma with iodine-131-labeled chimeric monoclonal antibody G250. Clin Oncol 15:1529-1537.
Sterling D, Alvarez BV & Casey JR (2002) The extracellular component of a transport metabolon. Extracellular loop 4 of the human AE1 Cl-/HCO3
- exchanger binds carbonic anhydrase IV. J Biol Chem 277:25239-25246.
Sterling D, Reithmeier RA & Casey JR (2001) A transport metabolon. Functional interaction of carbonic anhydrase II and chloride/bicarbonate exchangers. J Biol Chem 276:47886-47894.
Strazzabosco M, Joplin R, Zsembery A, Wallace L, Spirli C, Fabris L, Granato A, Rossanese A, Poci C, Neuberger JM, Okolicsanyi L & Crepaldi G (1997) Na+-dependent and –independent Cl-/HCO3
- exchange mediate cellular HCO3- transport in cultured human intrahepatic bile duct
cells. Hepatology 25:976-985. Strisciuglio P, Hu PY, Lim EJ, Ciccolella J & Sly WS (1998) Clinical and molecular heterogeneity
in carbonic anhydrase II deficiency and prenatal diagnosis in an Italian family. J Pediatr 132:717-720.
Stryker SJ, Wolff BG, Culp CE, Libbe SD, Ilstrup DM, MacCarty RL (1987) Natural history of untreated colonic polyps. Gastroenterology 93:1009-1013.
Supuran CT, Briganti F, Tilli S, Chegwidden WR & Scozzafava A (2001) Carbonic anhydrase inhibitors: sulfonamides as antitumor agents? Bioorg Med Chem 9:703-714.
Supuran CT & Scozzafava A (2002) Applications of carbonic anhydrase inhibitors and activators in therapy. Expert Opin Ther Patents 12:217-242.
Swenson ER (1991) Distribution and functions of carbonic anhydrase in the gastrointestinal tract. In: Dodgson SJ, Tashian RE, Gros G & Carter ND (eds) The Carbonic Anhydrases. Cellular Physiology and Molecular Genetics. Plenum Press, New York, p 265-287.
Tani T, Koyama Y, Nihei K, Hatakeyama S, Ohshiro K, Yoshida Y, Yaoita E, Sakai Y, Hatakeyama K & Yamamoto T (2001) Immunolocalization of aquaporin-8 in rat digestive organs and testis. Arch Histol Cytol 64:159-168.

69Tashian RE (1989) The carbonic anhydrases: Widening perspectives on their evolution, expression
and function. BioEssays 10:186-192. Tashian RE (1992) Genetics of the mammalian carbonic anhydrases. Adv Genet 30:321-356. Tashian RE, Goodman M, Headings VE, DeSimone J & Ward RE (1971) Genetic variation and
evolution in the red cell carbonic anhydrase isoenzymes of macaque monkeys. Biochem Genet 5:183-200.
Tashian RE, Kendall AG & Carter ND (1980) Inherited variants of human red cell carbonic anhydrases. Hemoglobin 4:635-651.
Tashian RE, Hewett-Emmett D, Carter ND & Bergenhem NCH (2000) Carbonic anhydrase (CA)-related proteins (CA-RPs), and transmembrane proteins with CA or CA-RP domains. In: Chegwidden WR, Carter ND & Edwards YH (eds) The Carbonic Anhydrases New Horizons. 1st ed. Basel: Birkhäuser Verlag, p 105-120.
Teicher BA, Liu SD, Liu LT, Holden SA & Herman TS (1993) A carbonic anhydrase inhibitor as a potential modulator of cancer therapies. Anticancer Res 13:1549-56.
Thatcher BJ, Doherty AE, Orvisky E, Martin BM & Henkin RI (1998) Gustin from human parotid saliva is carbonic anhydrase VI. Biochem Biophys Res Commun 250:635-641.
Torczynski RM & Bollon AP (1996) Gene sequence and probe for a marker of non-small cell lung carinoma. U.S. Patent 5,589,579.
Turner JR, Odze RF, Crum CP & Resnick MB (1997) NM antigen expression in normal, preneoplastic esophagus. Human Pathol 28:740-744.
Türeci Ö, Sahin U, Vollmar E, Siemer S, Göttert E, Seitz G, Pfreundschuh M, Parkkila A-K, Shah G, Grubb JH & Sly WS (1998) Human carbonic anhydrase XII: cDNA cloning, expression and chromosomal localization of a novel carbonic anhydrase gene that is overexpressed in some renal cell cancers. Proc Natl Acad Sci USA 95:7608-7613.
Uemura H, Nakagawa Y, Yoshida K, Saga S, Yoshikawa K, Hirao Y & Oosterwijk E (1999) MN/CA IX/G250 as a potential target for immunotherapy of renal cell carcinomas. Br J Cancer 1999 81:741-746.
Ulmasov B, Waheed A, Shah GN, Grubb JH, Sly WS, Tu C & Silverman DN (2000) Purification and kinetic analysis of recombinant CA XII, a membrane carbonic anhydrase overexpressed in certain cancers. Proc Natl Acad Sci USA 97:14212-14217.
van de Wetering M, Sancho E, Verweij C, de Lau W, Oving I, Huristone A, van der Horn K, Batlle E, Coudreuse D, Haramis A-P, Menno T-P-F, Moerer P, van den Born M, Soete G, Pals S, Eilers M, Medema R & Clevers H (2002) The β-catenin/TCF-4 complex imposes a crypt progenitor phenotype on colorectal cancer cells. Cell 111:241-250.
Venta PJ, Montgomery JC & Tashian RE (1987) Molecular genetics of carbonic anhydrase isozymes. Isozymes: Curr Top Biol Med Res 14:59-72.
Venta PJ, Welty RJ, Johnson TM, Sly WS & Tashian RE (1991) Carbonic anhydrase II deficiency syndrome in a Belgian family is caused by a point mutation at an invariant histidine residue (107 His-Tyr): complete structure of the normal human CA II gene. Am J Hum Genet 49:1082-1090.
Vermylen P, Roufosse C, Burny A, Verhest A, Bosscherst T, Pastorekovà S, Ninane V & Sculier JP (1999) Carbonic anhydrase IX antigen differentiates between preneoplastic malignang lesions in non-small cell lung carcinoma. Eur Respir J 14:806-811.
Vogelstein B, Fearon ER, Hamilton SR, Kern SE, Preisinger AC, Leppert M, Nakamura Y, White R, Smits AM & Bos JL (1988) Genetic alterations during colorectal-tumor development. N Engl J Med 319:525-532.
Väänänen HK & Autio-Harmainen H (1987) Carbonic anhydrase III: a new histochemical marker for myoepithelial cells. J Histochem Cytochem 35:683-686.
Waddell WR & Miesfeld RL (1995) Adenomatous polyposis coli, protein kinases, protein tyrosine phosphatase: the effect of sulindac. J Surg Oncol 58:252-256.
Waheed A, Parkkila S, Saarnio J, Fleming RE, Zhou XY, Tomatsu S, Britton RS, Bacon BR & Sly WS (1999) Association of HFE protein with transferrin receptor in crypt enterocytes of human duodenum. Proc Natl Acad Sci USA 96:1579-1584.

70Waheed A, Zhu XL & Sly WS (1992a) Membrane-associated carbonic anhydrase from rat lung.
Purification, characterization, tissue distribution, and comparison with carbonic anhydrase IVs of other mammals. J Biol Chem 267:3308-3311.
Waheed A, Zhu XL, Sly WS, Wetzel P & Gros G (1992b) Rat skeletal muscle membrane-associated carbonic anhydrase is 39-kDa, glycosylated, GPI-anchored CA IV. Arch Biochem Biophys 294:550-556.
Weisburger JH & Wynder EL (1987) Etiology of colorectal cancer with emphasis on mechanism of action and prevention. In: Devita VT, Hellman S & Rosenberg SA (eds) Important advances in oncology. Philadelphia: Lippincott, p 197-220.
Wenger RH & Gassmann M (1997) Oxygen(es) and the hypoxia-inducible factor-1. Biol Chem 378:609-616.
Whitney PL & Briggle TV (1982) Membrane-associated carbonic anhydrase purified from bovine lung. J Biol Chem 257:12056-12059.
Wills NK (1985) Apical membrane potassium and chloride permeabilities in surface cells of rabbit descending colon epithelium. J Physiol (London) 358:433-445.
Winawer SJ, Zauber AG, O'Brien MJ, Ho MN, Gottlieb L, Sternberg SS, Waye JD, Bond J, Schapiro M, Stewart ET, Panish J, Ackroyd F, Kurtz RC & Shike M, for The National Polyp Study Workgroup (1993) Randomized comparison of surveillance intervals after colonoscopic removal of newly diagnosed adenomatous polyps. The National Polyp Study Workgroup. N Engl J Med 328:901-906.
Wistrand PJ (1980) Human renal cytoplasmic carbonic anhydrase. Tissue levels and kinetic properties under near physiological conditions. Acta Physiol Scand 109:239-248.
Wistrand PJ (1981) The importance of carbonic anhydrase B and C for the unloading of CO2 by the human erythrocyte. Acta Physiol Scand 113:417-426.
Wistrand PJ & Knuuttila K-G (1989) Renal membrane-bound carbonic anhydrase. Purification and properties. Kidney Int 35:851-859.
Wykoff CC, Beasley N, Watson PH, Campo L, Chia SK, English R, Pastorek J, Sly WS, Ratcliffe P & Harris AL (2001) Expression of the hypoxia-inducible and tumor-associated carbonic anhydrases in ductal carcinoma in situ of the breast. Am J Pathol 158:1011-1019.
Wykoff CC, Beasley NJP, Watson PH, Turner KJ, Pastorek J, Sibtain A, Wilson GD, Turley H, Talks KL, Maxwell PH, Pugh CW, Ratcliffe PJ & Harris AL (2000) Hypoxia-inducible expression of tumor-associated carbonic anhydrases. Cancer Res 60:7075–7083.
Wåhlstrand T & Wistrand PJ (1980) Carbonic anhydrase C in the human renal medulla. Ups J Med Sci 85:7-17.
Yao T, Kouzuki T, Kajiwara M, Matsui N, Oya M & Tsuneyoshi M (1999) 'Serrated' adenoma of the colorectum, with reference to its gastric differentiation and its malignant potential. J Pathol 187:511-517.
Yang Y-S, Hanke JH, Carayannopoulos L, Craft CM, Capra JD & Tucker PW (1993) NonO, a non-POU-domain-containing, octamer-binding protein, is the mammalian homolog of Drosophila nonAdiss. Mol Cell Biol 13:5593–5603.
Zàvada J, Zàvadovà Z, Pastorek J, Biesova Z, Jezek J & Velek J (2000) Human tumour-associated cell adhesion protein MN/CA IX: identification of M75 epitope and of the region mediating cell adhesion. Br J Cancer 82:1808-1813.
Zàvada J, Zàvadovà Z, Pastorekovà S, Chiampor F, Pastorek J & Zelník V (1993) Expression of MaTu-MN protein in human tumor cultures and in clinical specimens. Int J Cancer 54:268-274.
Zbar B, Kaelin W, Maher E & Richard S (1999) Third International Meeting on von Hippel-Lindau disease. Cancer Res 59:2251-2253.
Zhu XL & Sly WS (1990) Carbonic anhydrase IV from human lung. Purification, characterization, and comparison with membrane carbonic anhydrase from human kidney. J Biol Chem 265:8795-8801.
Zhuang Z, Emmert-Buck MR, Roth MJ, Gnarra J, Linehan WM, Liotta LA & Lubensky IA (1996) von Hippel-Lindau disease gene deletion detected in microdissected sporadic human colon carcinoma specimens. Hum Pathol 27:152-156.