Embed Size (px)
Citation preview

This is an Open Access document downloaded from ORCA, Cardiff University's institutional
repository: http://orca.cf.ac.uk/124924/
This is the author’s version of a work that was submitted to / accepted for publication.
Citation for final published version:
Ratcliff, Michael, Rees, Daniel, McGrady, Scott, Buntwal, Luke, Hornsby, Amanda K. E., Bayliss,
Jaqueline, Kent, Brianne A., Bussey, Timothy, Saksida, Lisa, Beynon, Amy L., Howell, Owain W.,
Morgan, Alwena H., Sun, Yuxiang, Andrews, Zane B., Wells, Timothy and Davies, Jeffrey S. 2019.
Calorie restriction activates new adult born olfactory-bulb neurones in a ghrelin-dependent manner
but acyl-ghrelin does not enhance subventricular zone neurogenesis. Journal of Neuroendocrinology
31 (7) , -. 10.1111/jne.12755 file
Publishers page: http://dx.doi.org/10.1111/jne.12755 <http://dx.doi.org/10.1111/jne.12755>
Please note:
Changes made as a result of publishing processes such as copy-editing, formatting and page
numbers may not be reflected in this version. For the definitive version of this publication, please
refer to the published source. You are advised to consult the publisher’s version if you wish to cite
this paper.
This version is being made available in accordance with publisher policies. See
http://orca.cf.ac.uk/policies.html for usage policies. Copyright and moral rights for publications
made available in ORCA are retained by the copyright holders.

1
Calorierestrictionactivatesnewadultbornolfactory-bulbneuronesin1
a ghrelin-dependentmannerbut acyl-ghrelindoesnot enhance sub-2
ventricularzoneneurogenesis3
MichaelRatcliff1,DanielRees1*,ScottMcGrady1,LukeBuntwal1,AmandaK.E.Hornsby1,Jaqueline4
Bayliss2,BrianneA.Kent3,TimothyBussey4, LisaSaksida4,AmyLBeynon1,OwainWHowell1,5
AlwenaH.Morgan1,YSun5,ZaneB.Andrews2,TimothyWells6,JeffreyS.Davies1§6
7
1MolecularNeurobiology,InstituteofLifeSciences,SchoolofMedicine,SwanseaUniversity,UK.8
SA28PP.2DepartmentofPhysiology,BiomedicalDiscoveryUnit,MonashUniversity,Melbourne,9
Australia.3DepartmentofMedicine,Vancouver,UniversityofBritishColumbia,Canada.4Western10
University, London, Ontario, Canada. 5Department ofNutrition and Food Science, Texas A&M11
University,CollegeStation,USA.6SchoolofBiosciences,CardiffUniversity,UK.12
*Currentaddress:SchoolofManagement,SwanseaUniversity,BayCampus,Swansea,SA18EN,14
UK15
Keywords:Ghrelin,Neurogenesis,Olfactorybulb,Sub-ventricularzone,Calorierestriction16
17
Figures:418
Supplementaryfigures:519
Highlights20
• Acyl-ghrelinreceptor,GHSR,isnotexpressedintheSVZ21
• Acyl-ghrelindoesnotmodulateSVZcellproliferation22
• Acyl-ghrelindoesnotincreaseadultolfactorybulbneurogenesis23
• Geneticablationofghrelindoesnotaffectsurvivalofnewadultbornneurones24
• Acyl-ghrelinreceptor,GHSR,isexpressedintheolfactorybulb25
• Calorierestrictionactivatesnewadultbornneuronesinaghrelin-dependentmanner26
27
28
DrJeffreySDavies,29
MolecularNeurobiology,30
InstituteofLifeSciences,31
SchoolofMedicine,32
SwanseaUniversity,33
UK.SA28PP.34
35
Tel:+44(0)179260220936

2
Abstract38
Theageinganddegeneratingbrainshowdeficitsinneuralstem/progenitorcell(NSPC)plasticity39
thatareaccompaniedbyimpairmentsinolfactorydiscrimination.Emergingevidencesuggests40
thatthegut-hormoneghrelinplaysanimportantroleinprotectingneurones,promotingsynaptic41
plasticityandincreasinghippocampalneurogenesisintheadultbrain.Here,westudiedtherole42
ofghrelininmodulatingadultsub-ventricularzone(SVZ)NSPCsthatgiverisetonewolfactory43
bulb(OB)neurones.Wecharacterisedtheexpressionoftheghrelinreceptor,growthhormone44
secretagoguereceptor(GHSR),usinganimmuno-histochemicalapproachinGHSR-eGFPreporter45
micetoshowthatGHSRisexpressedinseveralregions,includingtheOB,butnotintheSVZof46
thelateralventricle.Thesedatasuggestthatacyl-ghrelindoesnotmediateadirecteffectonNSPC47
in the SVZ. Consistentwith these findings, treatmentwith acyl-ghrelin or genetic silencingof48
GHSR did not alter NSPC proliferation within the SVZ. Similarly, using a BrdU pulse-chase49
approachweshowthatperipheraltreatmentofadultratswithacyl-ghrelindidnotincreasethe50
number of new adult-born neurones in the granule cell layer (GCL) of the OB. These data51
demonstrate that acyl-ghrelin does not increase adult OB neurogenesis. Finally, we studied52
whetherelevatingghrelin indirectly,viacalorie restriction (CR), regulated theactivityofnew53
adult-borncellsintheOB.OvernightCRinducedc-Fosexpressioninnewadult-bornOBcells,but54
notindevelopmentallyborncells,whilstneuronalactivitywaslostfollowingre-feeding.These55
effectswereabsentinghrelin-/-mice,suggestingthatadult-borncellsareuniquelysensitiveto56
changes inghrelinmediatedby fastingandre-feeding. Insummary,ghrelindoesnotpromote57
neurogenesis in the SVZ and OB, however, new adult-born OB cells are activated by CR in a58
ghrelin-dependentmanner.59
60
61
62
63

3
Introduction64
Thegenerationofnewadult-bornneuronesintheolfactorybulb(OB)continuesthroughoutlife65
andcontributestoolfactorymemory.TheadultOBreceivesnewneuronesthatoriginatefrom66
divided neural stem / progenitor cells (NSPCs) residing in the sub-ventricular zone (SVZ)67
adjacenttothelateralventricles.FollowingNSPCdivision,thecellsdifferentiateintoimmature68
neuroblastsandmigratealongtherostralmigratorystream(RMS)priortointegrationwithlocal69
OBcircuitry1.ThisprocessofadultOBneurogenesis(AOBN)isregulatedbyseveralintrinsicand70
extrinsic factors including age, exercise, inflammation and glucocorticoids1. However, the71
underlyingmechanismsmediatingthisprocessarepoorlyunderstood.72
73
WithintheOB,newadult-bornneuronespromoteolfactorymemoryandenhancetheabilityto74
discriminatedistinctodours2,3.AOBNisalsoimportantforOBgranulecellreplacementandtissue75
maintenance4. Olfactory impairment has been reported as a prodromal indicator of several76
neurodegenerativediseases5.Forexample,deficitsinolfactorydiscrimination(i.etheabilityto77
distinguishodours)havebeendescribedinexperimentalneurodegenerativeanimalmodelsand78
humanParkinson’sdisease6.79
80
Ghrelin, an orexigenic gut hormone produced in response to calorie restriction, acts on the81
hypothalamustostimulatethereleaseofgrowthhormone(GH),andpromotemealinitiationand82
food intake. Emerging evidence suggests that acyl-ghrelin may also have important extra-83
hypothalamicfunctions7,suchasincreasingolfactorysensitivity8andregulatingactivityinbrain84
regionsinvolvedinolfactionandappetitivebehaviour9.85
In the neurogenic niche of the hippocampus, acyl-ghrelin has been shown to increase cell86
proliferation10 and the number of new adult-born neurones in adult rodents11. The ghrelin87
receptor,GHSR,whichisexpressedwithinthedentategyrusofthehippocampus,mediatesthe88
pro-neurogenic effect of calorie restriction (CR)12, as well as the increase in hippocampal89
neurogenesis and antidepressant-like effect following P7C3 treatment13. Moreover, ghrelin90
deficientmicearereportedtohaveimpairedcellproliferationintheSVZthatisnormalisedto91
wild-typelevelswithexogenousacyl-ghrelintreatment14.GHSRistheonlymolecularlyidentified92
receptorforghrelin,mediatingthecentraleffectsofthishormoneonappetite,bodyweightand93
energymetabolism15.However,itisnotknownwhetherGHSRisexpressedwithintheneurogenic94
nicheoftheSVZorwhetheracyl-ghrelinmodulatesadultolfactorybulbneurogenesis(AOBN).95
Here,weaimedtodeterminetheexpressionpatternofGHSRwithintheSVZandwhetherghrelin96
modulatesAOBN.97

4
Inaddition,asfastingandfeedingincrease16anddecreaseolfactorysensitivity17,respectively,we98
soughttodeterminewhetherghrelinmodulatesthefasting-inducedactivationofbothnewadult-99
bornanddevelopmentally-bornOBneurones. 100

5
MaterialsandMethods101
102
Animalsandprocedures103
All experiments involving animalswere performedwith appropriate ethical approval.Mouse104
studies were performed at Cardiff University (GHSR-null, Ghrelin-/-) and Monash University105
(GHSR-eGFP).StudiesinvolvingratswereperformedattheUniversityofCambridge.106
107
Mice108
GHSR-eGFP mice: Adult male GHSR-eGFP reporter mice were housed at room109
temperatureona12hlight,12hdarkcycle(0700-1900h)withfreeaccess(adlibitum)to110
foodandwater.GHSR-eGFPreportermice18(n=6)wereobtainedfromtheMouseMutant111
RegionalResourceCenterat theUniversityofCaliforniaDavis,andthehemizygousmice112
back-crossedtoC57BL/6Jmice.GHSR-eGFPreportermicewereterminallyanaesthetised113
andtrans-cardiallyperfusedwith4%paraformaldehyde(PFA)in0.1MPBS.Wholebrains114
wererapidlyremovedandpost-fixedinicecold4%PFAfor24hat4°Cbeforebeingsunk115
in 30% sucrose. Finally, brainswere transferred to PBS + 0.1% sodium azide (Sigma116
Aldrich,StLouis,USA)andstoredat4°Cpriortoanalysis.Brainswerefrozenusingafine117
powderofground-updryiceandmountedonaslidingsledgefreezingmicrotome(Zeiss,118
Microm HM 450) using Jung’s freezing medium. The thermostat was set to -30oC to119
ensurebrainsremainedfrozen.30µmthickcoronalsectionscutalongtheentirerostral-120
caudalaxis(bregma+5.345mmto-4.08)werecollectedina96-wellplate(Nunc,nunclon121
surface)filledwithPBS+0.1%sodiumazideandstoredat4oCuntilrequired.Ghsr-eGFP122
mousebrainswerealsocollectedinasagittalorientation(bregma+3.925to-0.20). 123
Immunofluorescence for GHSR-GFP: All experimentswere performed on free-floating tissue124
sectionsatroomtemperature,unlessstatedotherwise.A1in6seriesofcoronalorsagittalbrain125
sectionswereselected(minimumof10sectionspermouse),transferredintoa24-wellculture126
plate(Nunc,nunclonsurface)andwashedinPBS(SigmaAldrich,StLouis,USA)threetimesfor5127
minutes each. Tissue sections were then permeabilised in methanol (Fisher Scientific,128
Loughborough,UK)at-20°Cfor2minutesandwashed(asbefore)inPBS.Non-specificbinding129
siteswereblockedwith5%normalgoatserum(NGS)(SigmaAldrich,StLouis,USA) inPBS+130
0.1%tritonx-100(SigmaAldrich,Gillingham,UK)(PBS-T)for1h.Excessblockwasremovedand131
tissue sections incubated with chicken anti-GFP (Chicken polyclonal, Abcam, Cambridge, UK,132
Ab13970), diluted 1:1000 in PBS-T, for 24h at 4°C. Primary antibody was omitted from the133

6
negativecontrol.Sectionswerewashedandincubatedingoatanti-chickenAlexa-fluor488(Goat134
polyclonal,Lifetechnologies,USA,A11039),diluted1:500inPBS-T,for30minutesinthedark.135
Finally,sectionswerewashed,mountedontoSuperfrost+slides(Fisherbrand,Superfrost+slides)136
and cover-slippedwith vectashield (containing DAPI) (Vector Labs, Burlingame, USA) before137
being stored at 4°C. The slides were analysed by laser scanning confocal microscopy (Zeiss,138
LSM710) and Zen software (Zeiss, Zen 2010 edition) after 24h. Freely available GIMP v2.8139
softwarewasusedtopreparetiledimagesofcoronalandsagittalsections(www.gimp.org).140
GHSR-null mice: For assessing exogenous acyl-ghrelin regulation of SVZ cell proliferation,141
homozygousmaleloxTB-GHSRmice(GHSR-null)(agiftfromProfJeffreyZigman,Universityof142
TexasSouthwesternMedicalCenter,Dallas,TX)andW-T(C57BL/6J;W-T)controls(HarlanUK143
Ltd.)(14weeks-old,n=3/group)wereused19.Themethodologicalandmetabolicaspectsofthis144
studyhavepreviouslybeendescribed20.Briefly,micewerepreparedwithjugularveincannulae145
attached to osmoticmini-pumps (Alzetmodel 2001) under isofluorane anesthesia. Themini-146
pumpsdeliveredeithervehicleoracyl-ghrelin(48μg/day;PhoenixPharmaceuticals,USA)for7147
days.ThistreatmentprotocolwasshowntoincreaseabdominaladiposityviaGHSR,buthadno148
effectonbodyweight20.Micewereeuthanisedbycervicaldislocationandwholetrunkbloodwas149
collectedintoheparinizedtubesforplasmaseparationbycentrifugationat4,000gfor10minutes150
at4oC.Wholebrainwasremovedandimmediatelysnapfrozenondryiceandstoredat-80oC151
priortoanalysis.152
For analysis of Ki67, snap-frozen brains were sectioned at 10μm thickness using a cryostat 153
(Leica) and mounted directly onto superfrost+ coated slides (VWR). A one-in-fifteen series of 154
10μm sections (150μm apart) from each animal, a minimum of 8 sections per mouse, was 155
immunostained using rabbit anti-Ki67 (1:500, ab16667,Abcam)alongwithabiotinylatedgoat156
anti-rabbit forNi-DAB based detection, as previously described11. Cellswere imaged by light157
microscopy(Nikon50i)priortoquantificationusingImageJsoftware.158
Aseparatecohortof19-weekoldmaleGHSR-nullmice,derivedfromcrossesbetweenanimals159
thatwereheterozygousfortheGHSR-nullalleleandthathadbeenbackcrossed>10generations160
onto a C57BL/6J genetic background, and WT littermate mice were housed under normal161
laboratoryconditions (12h light:12hdark, lightsonat06.00h) (n=3/genotype).Micewere162
killedbycervicaldislocationunderterminalanaesthesia,wholebrainwasremoved,immersedin163
4%PFAfor24hat4◦Candcryoprotectedin30%sucrosepriortopreparationofcoronalsections164
(30µm)cutintoa1:12seriesalongtheentirerostro-caudalextentofthebrainusingafreezing-165
stage microtome (MicroM, ThermoScientific) and collected for IHC. For DAB-166

7
immunohistochemical analysis of GHSR labelling, a minimum of 6 sections per mouse were167
washed in 0.1M PBS (2× 10min) and 0.1M PBS-T (1× 10min). Subsequently, endogenous168
peroxidaseswerequenchedbywashing inaPBSplus1.5%H2O2 solutionfor20min.Sections169
werewashedagain(asabove)andincubatedin5%NDSinPBS-Tfor1h.Sectionswereincubated170
overnightat4◦Cwithrabbitanti-GHSR1a(PhoenixPharmaceuticals,H-001-62),diluted1:2000171
inPBS-Tand2%NGSsolution.Anotherwashstepfollowedpriortoincubationwithbiotinylated172
goat anti-rabbit (1:400; Vectorlabs, USA) in PBS-T for 70min. The sectionswerewashed and173
incubatedinABC(Vector-labs,USA)solutionfor90mininthedarkpriortoanothertwowashes174
in PBS, and incubation with 0.1 M sodium acetate pH6 for 10min. Immunoreactivity was175
developed in Nickel enhanced DAB solution followed by two washes in PBS. Sections were176
mountedontosuperfrost+slides(VWR,France)andallowedtodryovernightbeforebeingde-177
hydratedandde-lipifiedinincreasingconcentrationsofethanol.Finally,sectionswereincubated178
in histoclear (2× 3min;National Diagnostics, USA) and coverslipped using entellanmounting179
medium(Merck,USA).Slideswereallowedtodryovernightpriortoimaging. 180
Calorie restriction inGhrelin-/-mice:Adult femalehomozygous ghrelinknockout (ghrelin-/-)181
mice21 and theirwild type (WT) littermateswerederived fromcrossesbetweenanimals that182
wereheterozygousfortheghrelin-nullallele.Thesemicewerebackcrossed>10generationson183
aC57BL/6Jgeneticbackgroundandacclimatizedtobeingindividuallyhousedfor7daysunder184
normallaboratoryconditions(12hlight,12hdarkcycle;0700-1900h)priortotheonsetofthe185
study.Miceweredividedintosixgroups(n=5-8/group)thatincludedad-libitumfedWT,calorie186
restricted(CR)WT,calorierestricted/re-fed(CR/RF)WT,ad-libitumfedghrelin−/−,CRghrelin−/−187
andCR/RFghrelin−/−.Forthefirst28daysofthestudy,micewerefedonanad-libitumdietwith188
dailyinjections(fromdays1-4)ofthethymidineanalogue,BrdU(50mg/kg;i.p),tolabelactively189
dividingcells.Onday28, foodwaswithdrawnat17.30hfromtheCRandCR/RFmice.Onthe190
subsequent day, CR/RF mice were allowed to feed ad-libitum for 1h prior to all animals191
undergoing cervical dislocationwhilst under terminal anaesthesia (~18h CR). Ghrelin-/-mice192
havegrowthratesandappetitesimilartoWTlittermates,withnoimpairmentinhyperphagia193
afterfasting21,22.Similarly,adultablationofghrelininmicedoesnotimpairgrowthnorappetite23.194
Wholebrainswereremoved, immersedinicecold4%PFAfor24handcryoprotectedin30%195
sucrose.Coronalsections(30µm)werecutina1:12seriesalongtheentirerostral-caudalaxisof196
theolfactorybulb(bregma+5.345mmto+2.445mm)usingafreezingstagemicrotome(MicroM,197
ThermoScientific)andcollectedforIHC.198
199
Quantification of BrdU+/c-Fos+: All IHC was performed on free-floating sections at room200
temperature,unlessotherwisestated.A1:6seriesof30μmsections(180μmapart)werewashed201

8
threetimesinPBSfor5min,permeabilisedinmethanolat-20°Cfor2min,andwashedasbefore.202
DNAwasdenaturedwith2MHCLfor30minat37°Cpriortowashingsectionsin0.1Mborate203
buffer(pH8.5)for10minutes.Sectionswerewashed,blockedwith5%normalgoatserum(NGS)204
plus5%bovineserumalbumin(BSA)inPBSplus0.1%Triton(PBS-T)for60minandincubated205
inacocktailofprimaryantibodiesthatincludedratanti-BrdU(1:400;MCA2060,ABDSerotec)206
andrabbitanti-c-Fos(1:1000;SC-52,SantaCruz)inPBS-Tovernightat4°C.Theprimaryantibody207
wasomittedfromthenegativecontrol.208
Followingprimaryantibodytreatment,sectionswerewashed,incubatedwithbiotinylatedgoat209
anti-rat(1:400;BA-9400,VectorLabs)inPBS-Tfor60mininthedarkandthenwashedasbefore.210
Similarly, secondary antibodieswere also applied as a cocktail that included goat anti-rabbit211
(1:400;BA-1000,VectorLabs)andstreptavidinAF-594(1:500;S11227,LifeTechnologies) in212
PBS-T for 30 min. Following another wash, including one containing Hoechst nuclear stain,213
sectionsweremountedontosuperfrost+ slides(VWR,France)andcover-slippedwithprolong214
goldanti-fadesolution(LifeTechnologies,USA).215
216
Quantificationofimmunolabelledcells:A1:6seriesof30μmsections(180μmapart)fromeach217
animalwasanalysedfor immunoreactivityusinganepi-fluorescentmicroscopesystem(Zeiss,218
ImagerM1withAxiocamMRm).Immunolabelledcellsweremanuallycountedbilaterallyusinga219
´40objectivethroughthez-axisoftheentirerostral-caudalextentofthedorsalgranulecelllayer220
(GCL),glomerularlayer(GL),subependymalzone(SEZ)andthelateralolfactorytractbody(LOT).221
Resultingnumbersweredividedbythetotalareameasurementtogiveacountperpixel,which222
wasconvertedintomm2andaveragedforeachbrain.Allanalyseswereperformedblindtoboth223
genotypeandtreatment.224
225
Rats226
Adultmale lister hooded rats (n=10/11per group,weighing 250-300g;Harlan, Bicester, UK)227
werehousedingroupsoffourandmaintainedatroomtemperatureona12hlight,12hdarkcycle228
(0700-1900h).Theseexperimentalprocedureshavepreviouslybeendescribed11.Briefly,from229
days 0-14, rats received daily intra-peritoneal injections of acyl-ghrelin (Phoenix230
Pharmaceuticals,031-31)orsaline(10µg/kgbodyweight)withBrdUinjections(50mg/kg)on231
days5-8.Onday29,ratswereterminallyanaesthetised,trans-cardiallyperfusedwith4%PFA232
andbrainswereremovedforimmersionfixationandcryoprotection(asbefore).Analysisofadult233
hippocampal neurogenesis (AHN) in these rats demonstrated that acyl-ghrelin significantly234
increasedthenumberofnewadultbornneurones11.235
236

9
Double immunofluorescence for BrdU/NeuN: A 1 in 6 series of coronal OB brain sections237
(bregma+5.345mmto+2.445)weretransferredintoa24-wellcultureplateandwashedinPBS,238
permeabilised in methanol at -20°C for 2 minutes and washed in PBS as before. DNA was239
denaturedusing2Mhydrochloricacid(HCL)(FisherScientific,Loughborough,UK)andincubated240
at37oCfor30minutes.ExcessHCLwasremovedandthesectionswashedin0.1Mboratebuffer,241
pH 8.5, for 10 minutes to neutralise the remaining HCL. Tissue sections were then washed,242
blockedwith5%NGSdilutedinPBS-Tfor1handincubatedwithratanti-BrdU(Ratmonoclonal,243
ABDSerotec,Oxfordshire,UK,MCA2060),diluted1:3000inPBS-Tfor24hat4°C.Theprimary244
antibody was omitted from the negative control. Sections were washed and incubated with245
biotinylatedgoatanti-rat(Goatpolyclonal,Vectorlabs,Burlingham,USA,BA-9400),diluted1:400246
inPBS-Tfor1hinthedark.Tissuesectionsweresubsequentlywashed,incubatedinstreptavidin247
AF594 (Life technologies, Eugene, USA, S11227), diluted 1:500 in PBS-T for 30minutes and248
washedasbefore.Sectionswerethenincubatedinmouseanti-NeuN(Mousemonoclonal,EMD249
Millipore,Massachusetts,USA,MAB377),diluted1:1000inPBS-Tfor1h.Thenegativecontrol250
containedPBS-T.Tissuesectionswerethenwashedandincubatedingoatanti-mouseAF488251
diluted1:500inPBS-Tfor30minutes,priortobeingwashedwithHoescht,diluted1:10000in252
PBS,for5minutes.Finally,sectionswerewashed,mountedontoSuperfrost+slidesandcover-253
slippedwithprolonggoldanti-fadereagent,priortostorageat4°C.254
255
QuantificationofBrdU+:ImageJsoftware(version1.47)wasusedtoquantifythenumberofnew256
adult-borncellsinthedorsalandventralgranularcelllayer(GCL)oftheOB.Imagestakenbythe257
fluorescentmicroscopewereinvertedandunsharp-masked,usingaradiusof10.0pixelsanda258
maskweightof0.60.Thepolygontoolwasthenusedtodrawaroundthegranularcelllayerand259
the total areameasured.Each image’s thresholdwas individuallyoptimised, typically ranging260
from0.100–0.180.Theparticlesizewassetto20-300pixel2andcircularityat0.0-1.0.Resulting261
numbersweredividedbythetotalareameasurementtogiveacountperpixel,whichwasthen262
averagedforeachbrain.263
264
QuantificationofBrdU+/NeuN+:Toquantifythenumberofnewadult-bornneuronsinthedorsal265
and ventral granular cell layer of the OB, BrdU+/NeuN+ immunoreactive cellsweremanually266
countedthroughthez-axisoftheentirerostral-caudalextentoftheOB.Resultingnumberswere267
dividedbytheareaofthez-stacktogiveacountperpixel,whichwasthenaveragedforeach268
brain.269
270

10
Microscopy271
Tissuesectionswereanalysedusingafluorescentmicroscope(Zeiss,ImagerM1withAxiocam272
MRm)withAxiovisionsoftware(version4.6)andalaserscanningconfocalmicroscope(Zeiss,273
LSM710)withZensoftware(Zen2010edition).Imageswerecollectedusing´4,´10and´40274
objectives.BrdU+/NeuN+immunoreactivenewbornadultneurones,inthedorsalandventralGCL,275
wereimagedusinga´40oilimmersionobjective.Az-stackconsistingof21-25tissueslicesat276
0.7µmintervals(14.0-16.8µmrange)weretakenthroughouttherostral-caudalextentoftheOB.277
Allexperimentsandanalyseswereperformedblindtogenotypeandtreatment.278
279
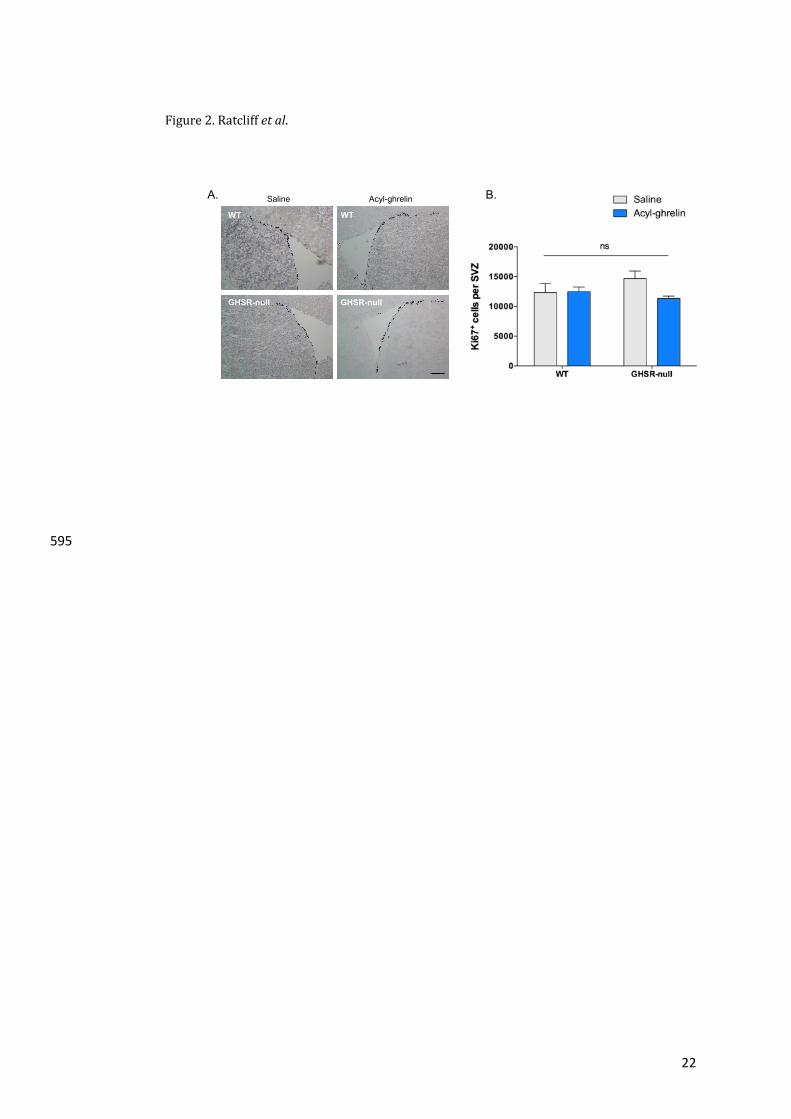
StatisticalAnalysis280
StatisticalanalyseswereperformedusingGraphPadPrism6.0forMac(GraphPadSoftware,San281
Diego,CA).Statisticalsignificancewasassessedbyanunpairedtwo-tailedStudent’st-testorone-282
wayANOVAwithBonferroni’spost-hoctest.Wheretherewasmorethanonevariableatwo-way283
ANOVAwithTukey’smultiplecomparisonstestwasusedoraKruskal-Wallistestfollowedbya284
Dunn’smultiple comparisons testwas usedwhen a normal distribution of data could not be285
assumed. Data are presented as a mean ±SEM. *, P<0.05; **, P<0.01; ***, P<0.001 and ****,286
P<0.0001wereconsideredstatisticallysignificant.287
288
289

11
Results290
GHSRisexpressedintheadultOBbutnotintheSVZ291
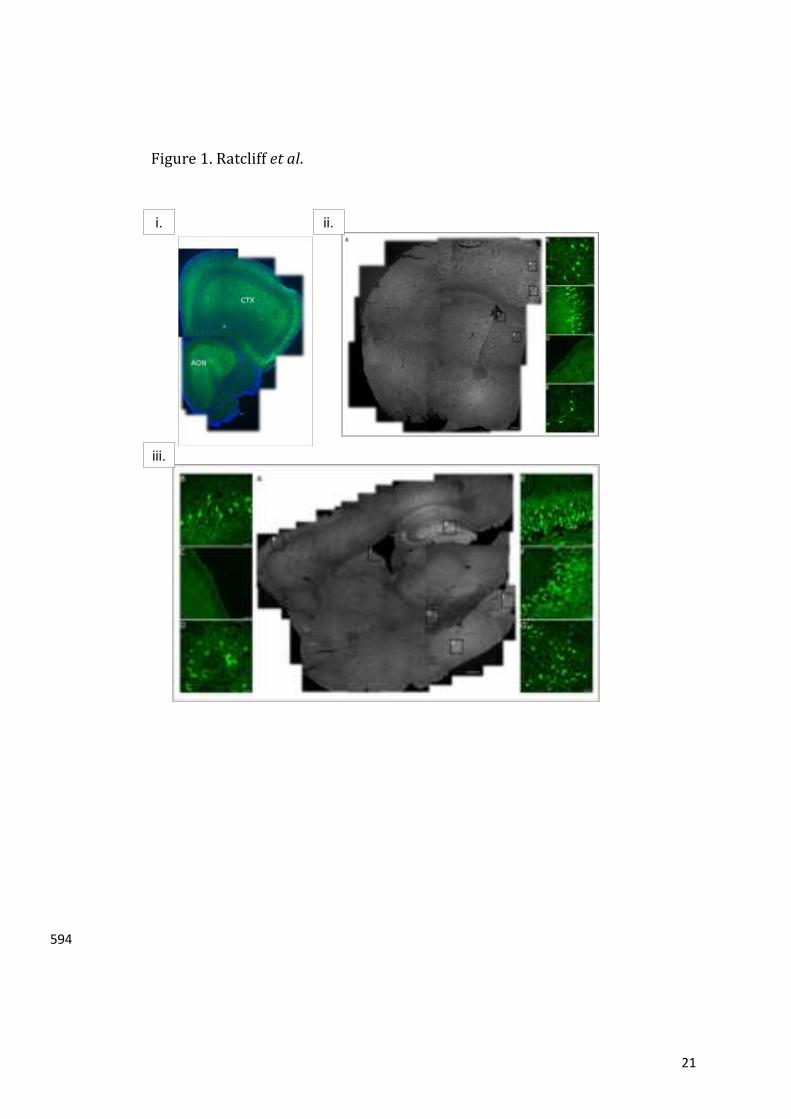
TheexpressionofGHSRwasassessedtodeterminewhetherghrelincoulddirectlyinfluencethe292
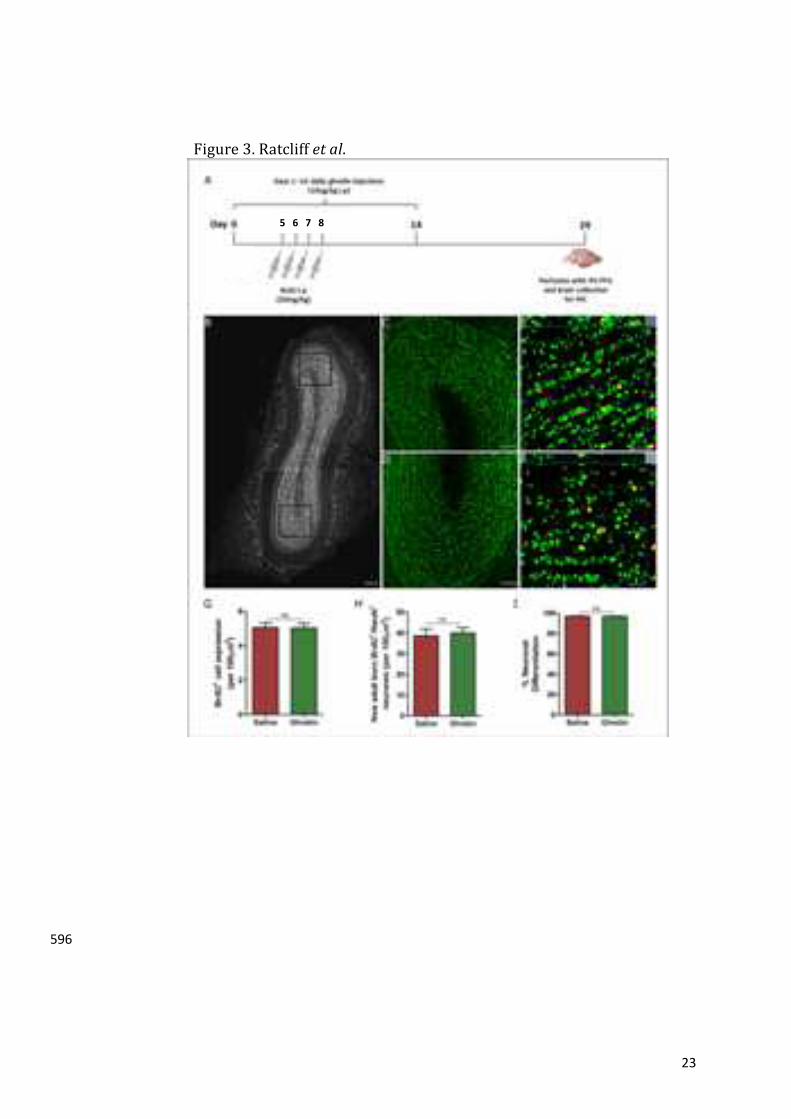
proliferationofNSPCsintheSVZ.ToachievethisaimweusedGHSR-eGFPreportermicetoshow293
thateGFPimmunoreactivitywaspresentwithintheanteriorolfactorynucleus(AON)andorbital294
andmotororbitalcortexinthecaudalOB(figure1i).Immunoreactivitywasobservedwithinthe295
anteriorcingulatecortex,motorcortexand lateralseptalnucleus(figure1ii).Sagittalsections296
revealed strong immunoreactivity in the anterior amygdala area, granule cell layer of the297
hippocampaldentategyrus(DG)andthemedialamygdalanucleus(figure1iii).Notably,staining298
wasabsentwithinthelateralliningoftheSVZintissuesectionedinbothacoronalandsagittal299
orientation(figure1iiDandiiiC).TodeterminewhethertheGHSR-eGFPimmunoreactivitywas300
similartothatobservedwithGHSR1aantisera,weperformedIHCusingarabbitanti-GHSR1a301
antibody on adultWT and GHSR-null mouse brain tissue. These analyses revealed a similar302
patternof immunoreactivityonbothWTandGHSR-null tissues, includingtheSVZ(figureS1),303
suggestingalackofbindingspecificityfortheGHSR1aantigen.Thesedatasuggestthatghrelin304
maybeinvolvedinolfactoryfunctionbutnotthroughdirectmodulationofNSC’sliningthelateral305
walloftheSVZ.306
307
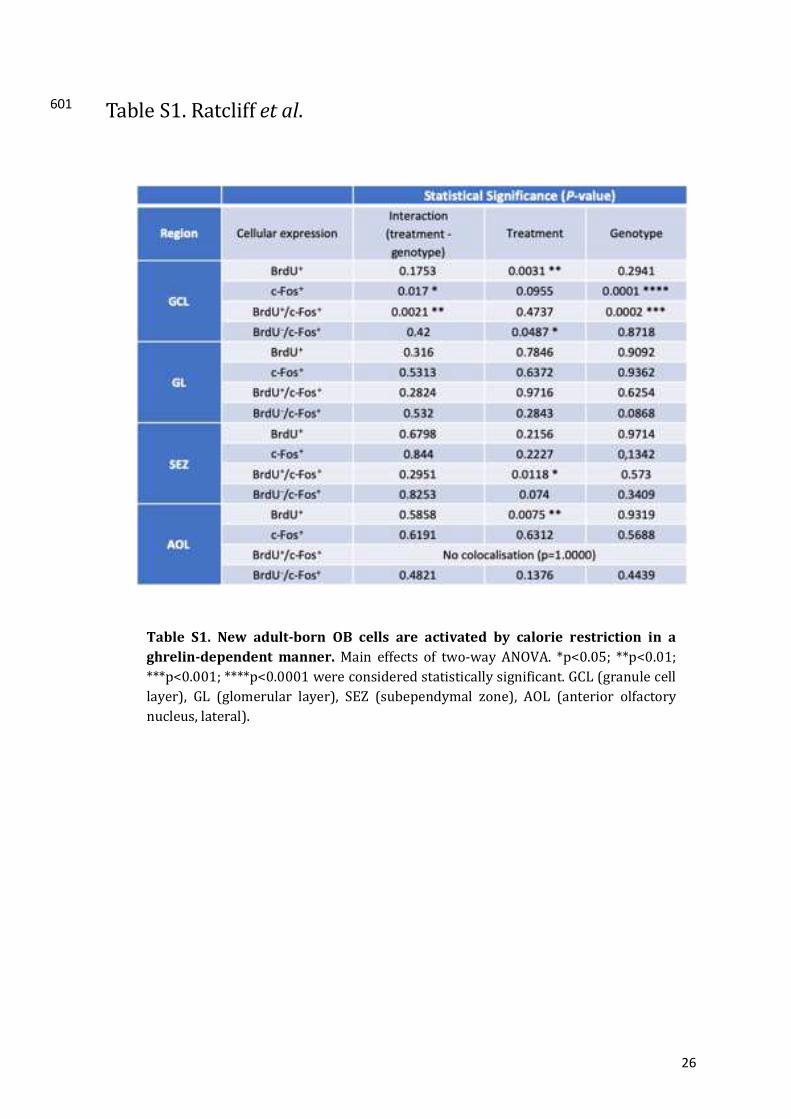
Acyl-ghrelindoesnotincreasecellproliferationintheadultSVZ308
TheproliferativeeffectofghrelinandGHSR-agonistshavebeenwidelyreportedwithinCNSand309
peripheraltissues24–26.Arecentstudyreportedthatghrelinpromotedproliferationofcellswithin310
the SVZ14.Here,we tookadvantageof geneticallymodifiedmice to analyse the effect of acyl-311
ghrelin treatment on SVZ cell proliferation in adultWT and GHSR-nullmice, where GHSR is312
transcriptionallysilenced.Usingthemitoticmarker,Ki67,wereportthatacyl-ghrelintreatment313
hadnoeffectonthenumberofproliferatingcellswithintheSVZnicheinWTmice(figure2,P=314
>0.99).Similarly,transcriptionalsilencingofGHSRdidnotaffecttherateofSVZcelldivisionin315
vehicle(WTvehvsGHSR-nullveh,P=0.6388)oracyl-ghrelintreatedmice(GHSR-nullvehvs316
GHSR-null acyl-ghrelin,P = 0.0944). The low number of replicatesmeans that the statistical317
analysis is of low power, however, the data suggest that acyl-ghrelin does not regulate cell318
proliferationintheadultmouseSVZandthatgeneticsilencingofGHSRdoesnotdecreasecell319
divisioninthisniche.320
321

12
Acyl-ghrelindoesnotincreasethenumberofnewadult-bornolfactorybulbneurones322
Werecentlyshowedthattreatmentwithacyl-ghrelinincreasedadulthippocampalneurogenesis323
(AHN)inadultrats11.Todeterminewhetheracyl-ghrelintreatmentmodulatesAOBNinasimilar324
waytoAHN,wequantifiedthenumberofnewadult-bornneuronesintheOBofadultratsfrom325
thesamestudy.UsingOBtissuefromthesameratswherebyacyl-ghrelinincreasedAHNprovides326
us with valuable experimental controls. We show there was no significant difference in the327
number of new adult-born cells (BrdU+/NeuN-) in the GCL of the OB following acyl-ghrelin328
treatmentcomparedtosalinetreatment(figure3G,P=0.8482).Similarly,nodifferenceswere329
observedinthenumberofnewadult-bornneurones(BrdU+/NeuN+)(figure3H,P=0.7388)orin330
therateofneuronedifferentiation(figure3I,P=0.6870).331
332
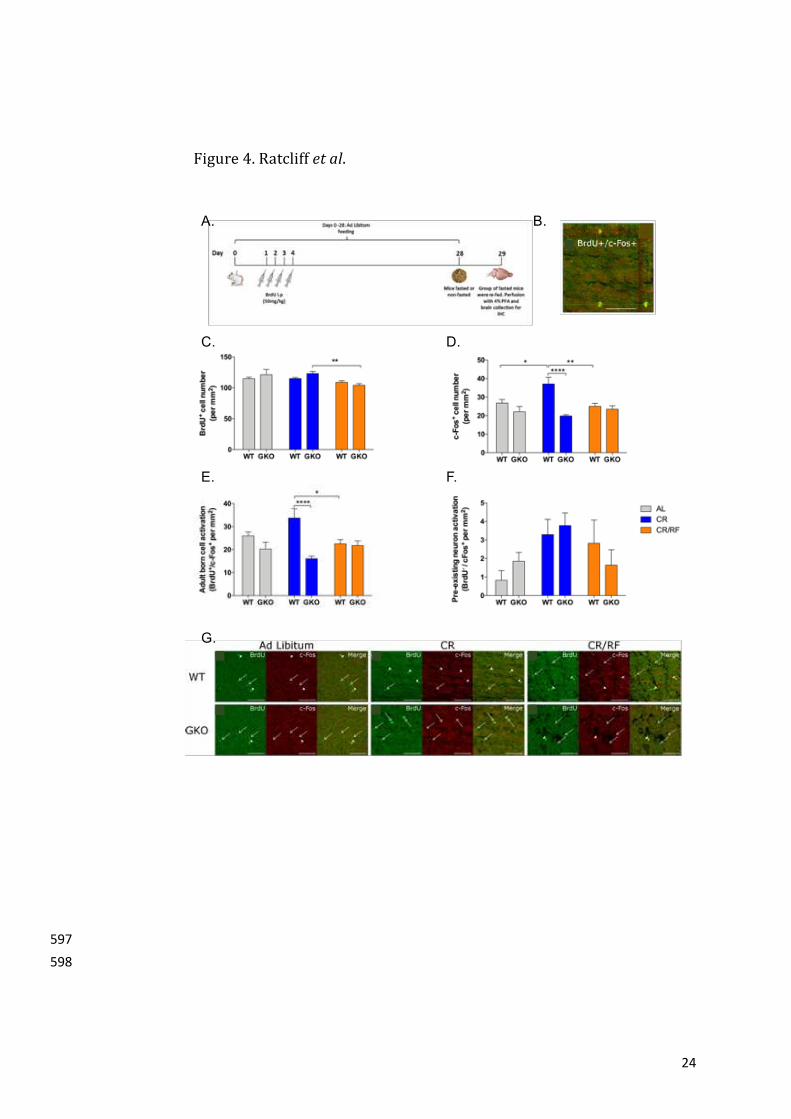
Calorierestrictioninducesactivationofnewadult-bornOBcellsinaghrelin-dependent333
manner334
TodeterminewhetheraCR-mediatedincreaseinendogenousacyl-ghrelinwasabletoincrease335
theexpressionoftheproto-oncogene,c-Fos,innewadult-bornOBcells,weanalysedthenumber336
ofactivec-Fos+cellswithintheGCL,GL,SEZandLOTinWTandghrelin-/-mice.Atwo-wayANOVA337
revealedastatisticallysignificantmaineffectoftreatmentonBrdU+(p=0.0031)andBrdU-/c-Fos+338
(p=0.0487) cellswithin theGCL. Comparatively, genotype and the interaction (treatment and339
genotype)showedasignificanteffectonc-Fos+(p=0.0001and0.017,respectively)andBrdU+/c-340
Fos+(p=0.0002and0.00021,respectively)cellswithintheGCL.OutsideoftheGCL,asignificant341
maineffectoftreatmentwasreportedinBrdU+/c-Fos+cellsoftheSEZ(p=0.0118)andBrdU+cells342
oftheLOT(p=0.0075).NoincreasewasobservedwithintheGL(TableS1).343
ATukeyposthoc test revealeda reduction in thenumberofnewadult-born cells (BrdU+) in344
CR/re-fed(CR/RF)ghrelin-/-mice,comparedwithCRghrelin-/-mice(figure4C,P=0.0086)within345
theGCL.Furthermore,therewasanincreasednumberofactivatedcells(c-Fos+)inCRWTmice,346
comparedwithadlibitumWT(P=0.0258)andCR/RFWT(P=0.0043)mice.Notably,CRalso347
increasedactivatedcellsinWTrelativetoCRghrelin-/-mice(P<0.0001)withintheGCL(figure348
4D).Furtheranalysisrevealedthatthenumberofactivenewadult-borncells(BrdU+/c-Fos+)was349
increased in CR WT mice compared with CR/RF WT (P = 0.0169) and CR ghrelin-/- mice350
(p<0.0001)within theGCL (figure4E).Whereas, therewerevery fewactivedevelopmentally351
borncells(BrdU-/c-Fos+)andthesecellswerenotsignificantlyaffectedbytreatmentorgenotype352
(figure4F).Outsideof theGCL, thenumberofactivenewadult-borncells(BrdU+/c-Fos+)was353
reducedinCRghrelin-/-mice,comparedwithad-libitumfedghrelin-/-mice(P=0.0169)within354
theSEZ.Nosignificantdifferenceswerereportedintheotherregionstested(TableS1).There355
wasnosignificantdifferenceinbodyweightchangeinghrelin-/-micerelativetoWTmiceineither356
of the groups (Two-way ANOVA;main effect of genotype,P = 0.9335;main effect of feeding357

13
pattern, P = 0.0049; main effect of interaction (feeding pattern vs genotype), P = 0.3469).358
Collectively, these data suggest that CR increases the activation of new adult-born cells in a359
ghrelin-dependentmanner.360
361
Discussion362
ThegenerationofnewOBneuronesintheadultbrainisimportantforolfactorydiscrimination,a363
processthatisimpairedinageingandseveralneurodegenerativedisorders.Here,wetestedthe364
hypothesisthatghrelinisanimportantregulatorofAOBN.First,wecharacterisedexpressionof365
GHSRintheadultmousebrain.Numerousstudieshaveattemptedtocharacterisetheexpression366
patternofGHSRinseveralspecies,includingmouse,ratandlemur27–30,thoughthelackofreliable367
anti-GHSRantibodieshavelimitedprogress.Morerecently,areportusingtheGHSR-eGFPmouse368
and in situ hybridisation histochemistry demonstrated GHSR expression within the OB,369
hippocampusandhypothalamicnuclei15.Furthermore,Cre-activity inGhsr-IRES-Cre/ROSA26-370
ZsGreenreportermicewasalsoreportedinthemainandaccessoryOB31.371
Here, using adult GHSR-eGFPmice, we report GHSR immunoreactivity in theMCL, AON and372
orbitalandmotororbitalcorticesoftheOB,aswellaswithintheanteriorcingulatecortex,motor373
cortex,lateralseptalnucleus,entopeduncularnucleus,hippocampusandthemedialamygdaloid374
nucleus.However,GHSRwasnotexpressedwithintheneurogenicnicheoftheSVZinGHSR-eGFP375
reportermice.Indeed,thisfindingisconsistentwithpreviousstudiesthatdonotreportGHSR376
immunoreactivitywithintheSVZniche.Together, thesefindingssuggestthatghrelindoesnot377
mediatedirecteffectsonNSPCproliferation.378
AseGFPimmunoreactivityinthistransgenicmodelmaycorrespondtotwostructurallydifferent379
receptors,GHSR1a,whichencodesthefunctionalreceptor,andthetruncatedGHSR1b,generated380
fromalternativesplicingofGHSR,wesoughttoidentifyGHSR1aexpressingcellsusingantisera381
raisedagainstGHSR1a.ThespecificityofpolyclonalantibodiesusedtocharacteriseGHSRwithin382
the adult brain remainsunclear. Liet al.14 reportedGHSRexpressionwithin the adultmouse383
neurogenicnicheoftheSVZusingimmuno-fluorescencewiththerabbitanti-GHSR1aantibody384
(PhoenixPharmaceuticals,H-001-62),diluted1:500.Inourstudy,usingthesameantibody,IHC385
inbraintissuefromadultGHSR-nullandWTmicerevealeddetectableimmunoreactivityintissue386
frombothgenotypes.Ourdatasuggestthattherabbitanti-GHSRantibodyresultedinnon-specific387
stainingwithintheSVZandcingulatecortex(figureS1),preventingit’susetodetermineGHSR1a388
expression in this context. Combined, these studies suggest that the ghrelin receptor is not389
expressedintheSVZ,andthusdoesnotdirectlymodulateNSPCproliferation.390
TodeterminewhetherghrelininducescellproliferationwithintheSVZwetreatedGHSR-nulland391
wild-typemicefor7-dayswithacyl-ghrelin.Subsequentanalysisrevealednoeffectofgenotype392

14
or treatment on the number of dividing Ki67+ cells in the SVZ. In contrast, a previous study393
reportedthatghrelin-/-micehadareducednumberofproliferatingNSPCs,migratingneuroblasts394
andOBinterneurones,thatcouldberestoredtoWTlevelsbyintraperitonealadministrationof395
acyl-ghrelin14.Severaldifferencesbetweenthetwoexperimentalproceduresmightaccountfor396
the contrasting results. For example, Li et al. used 8-9week oldWT and ghrelin-/-mice that397
receivedacyl-ghrelin(80μg/kg)viaintraperitonealinjection,oncedailyfor8consecutivedays.398
Whereas,inourstudy14-weekoldWTandGHSR-nullmiceweregivenacyl-ghrelin(48μg/day)399
viaintravenousmini-pump.Therefore, inconsistenciesbetweenstudiesmaybeattributableto400
geneticbackground,thephysiologicaldoseortherouteofadministrationofacyl-ghrelin.401
402
Next, using aBrdUpulse-chase approachwedetermined the effect of exogenous acyl-ghrelin403
treatmentonthematurationandsurvivalofnewadult-bornneuronesintheratOB.Consistent404
withourpreviouscellproliferationanalysisinmice,acyl-ghrelindidnotincreaseinthenumber405
ofnewadult-bornBrdU+cellsorBrdu+/NeuN+neuronesintheGCLoftheOB.Furthermore,no406
differenceswereobservedintherateofneuronaldifferentiation.Notably,wehavepreviously407
reported that adult hippocampal neurogenesis was significantly increased by acyl-ghrelin in408
theserats11.ThehighlevelofGHSRexpressionwithinhippocampalneurogenicniche12andit’s409
absence in the SVZ niche is likely responsible for this effect. These data provide compelling410
evidencethatacyl-ghrelindoesnotpromoteAOBN.411
Numerous studies have suggested that ghrelin plays an important role in olfactory-related412
behavioursincludingodourdiscriminationandsensitivity8,32,33.Lochetal.reportedanincreased413
responsiveness of themouse olfactory epithelium following nasal application of ghrelin. This414
resulted in a higher reactivity of olfactory sensory neuroneswithin the olfactory epithelium,415
which in turn, increased the activity of receptor-specific glomeruli. GHSR expression on the416
surfaceofolfactorysensoryneuronessuggestthatghrelinandGHSRmayplayanimportantrole417
in enhancing neuronal responsiveness and olfaction. However, the underlyingmechanism by418
which acyl-ghrelin enhances olfaction remains elusive and it is unclear if new adult-bornOB419
neuronesareinvolvedinthisphysiology.420
Asacyl-ghrelinisknowntoregulatebotholfactionandappetitewesoughttodeterminewhether421
new adult-born OB neurones are activated by CR in a ghrelin-dependent manner. Our data422
demonstratethatovernightCRactivatednewadult-borncellsintheOB.Re-feedingforonehour423
returnedthenumberofc-Fospositivecellstobaseline,suggestingthatthenewadult-borncells424
aresensitivetofeedingstatus.Notably,thisCReffectwasabsentinghrelin-/-micedemonstrating425
that the activation of new adult-born cells was dependent upon intact ghrelin signalling.426
Furthermore,therewasnoCR-mediatedactivationofdevelopmentally-borncells(BrdU-/c-Fos+)427

15
intheGCLoftheOBindicatingthatadult-bornneuronesareuniquelyresponsivetoacutechanges428
in food intake. Therefore,we confirm that CR activates newadult-bornOB cells in a ghrelin-429
dependentmanner.ThisfindingprovidesfurthersupportforghrelinactingasamediatorofCR-430
associated physiology, including, neuroprotection34, anti-anxiety35, hippocampal neurogenesis431
andcognitiveenhancement12,andglycemicregulation36.432
Althoughtherelationshipbetweenhungerstimulationandolfactionhasbeenlongrecognised,a433
molecularmechanismrelatingthetwoprocesseshasnotbeendetermined37.Soria-Gomezetal.434
observed that cortical feedback projections to theOB crucially regulate food intake, possibly435
through cannabinoid type-1 receptor (CB1R) signalling. The endocannabinoid system, in436
particularCB-1Rs,promotedfoodintakeinfastedmicebyincreasingodourdetection.Notably,437
ghrelin’sorexigeniceffectislostinCB-1Rknock-outmice38.Althoughtherelationshipbetween438
ghrelinandtheendocannabinoidsystemintheOBisunknown,bothGHSRandCB-1RareGPCRs439
known to formhomo- and heterodimers (or higher-order oligomers) as part of their normal440
traffickingandfunction39,40.Therefore,heterodimerisationofCB-1RandGHSRmaybeimportant441
inlinkingghrelintoadult-bornOBneuronesandolfaction.442
Severalquestionsremainunanswered,includingwhetherghrelinalterstheelectrophysiological443
propertiesand/ordirectlyactivatesGCsintheOBtoenhanceodourdiscrimination.Asnewadult-444
born OB cells enhance the odour-reward association3, further work is needed to determine445
whethertheghrelin-inducedintakeofrewardingfoods41requiressignallingvianewneuronesin446
theOB.Similarly,itisnotknownwhetherghrelincanincreaseappetiteandimproveolfactionin447
theabsenceofnewadult-bornOBcells.448
Insummary,thesedatademonstratethatwhileghrelindoesnotincreaseSVZ-OBneurogenesis,449
itdoesmediatetheCR-inducedactivationofnewadult-bornOBcells.Wespeculatethatghrelin450
modulatesnewOBneuroneactivitytointegrateolfactoryresponseswithnutritionalstatus. 451
452

16
Figurelegends453
454
Figure1.CharacterisationofGHSR1aintheadultGHSR1a-eGFPmousebrain.455
(i).GHSR1a-eGFPimmunoreactivityispresentwithintheorbitalandmotororbitalcortexandthe456
anteriorolfactorynucleus.CTX,cortex;AON,anteriorolfactorynucleus.457
(ii).Collageofcoronalmousesections(A). Inset imagesofGHSR1a-eGFP immunoreactivity in458
anteriorcingulatecortexdorsal(B),anteriorcingulatecortex(C)andlateralseptalnucleus(E).459
GHSR1a-eGFPimmunoreactivityisabsentinthelateralliningoftheSVZ(D).460
(iii).Collageofsagittalmousesections(A). Inset imagesofGHSR1a-eGFPimmunoreactivity in461
primarymotorcortex(B),anterioramygdalaarea(D),dorsalgranulecell layerof thedentate462
gyrus(E),ventraldentategyrus(F)andmedialamygdalarnucleus(posterodorsal)(G).GHSR1a-463
eGFPimmunoreactivityisabsentwithinthelateralliningoftheSVZ(C).464
Montageimagescalebar=200µm.Insetimagescalebar=50µm.465
466
Figure2.Acyl-ghrelin treatmentdoesnot increasecellproliferation in theSVZofadult467
wild-typeorGHSR-nullmice.(A)GHSR-nullandWTlittermatemiceweretreatedfor7-days468
with either saline or acyl-ghrelin (48ug/day i.v) via osmotic mini-pump before brains were469
collectedandKi67immunoreactivityquantifiedthroughouttherostro-caudalextentoftheSVZ.470
(B)TotalnumberofKi67+cellsdidnotchangefollowingacyl-ghrelintreatmentineitherWTor471
GHSR-nullmice.Dataaremean+/-SEM,n=3micepergroup.Statisticalanalysisperformedby472
Kruskal-Wallistest(P=0.2087)followedbyapost-hocDunn’smultiplecomparisontest.473
474
Figure3.Exogenousacyl-ghrelindoesnotincreasethenumberofnewadultbornneurones475
in thegranule cell layerof the ratolfactorybulb. (A)Experimental paradigm. (B)Collage476
imageof theratolfactorybulb.Representative imagesofBrdU(red)andNeuN(green) in (C)477
dorsalgranulecelllayer(GCL)and(D)ventralGCLoftheOB.Scalebar=200µm.Representative478
imagesofnewadult-bornneuronesco-expressingNeuN+andBrdU+(yellow)in(E)dorsalGCL479
and(F)ventralGCL.Scalebar=50µm.Quantificationofnewadult-bornOBcells(G)(P=0.8482),480
newadult-bornneurones(H)(P=0.7388)and%neuronaldifferentiation(I)(P=0.6870)after481
acyl-ghrelinorsalinetreatment.Dataaremean+/-SEM.Statisticalanalysiswasperformedby482
two-tailedunpairedStudent’st-test.P<0.05consideredsignificant,ns=notsignificant.n=11483
ratspergroup.484
485
Figure 4. New adult-born OB cells are activated by calorie restriction in a ghrelin-486
dependent manner. (A) Schematic of experimental paradigm. (B) New adult-born active487

17
neurone(yellow;scalebar=25µm)coexpressingBrdU(green)andc-Fos(red)intheGCLofthe488
OB.Scalebar=50µm.Quantificationof(C)newadult-borncells(BrdU+),(D)activecells(c-Fos+),489
(E)activenewadult-borncells(BrdU+/c-Fos+)and(F)activedevelopmentallyborncells(BrdU-490
/c-Fos+)intheGCLoftheOB.(G)Representativeimagesofnewadult-borncells(BrdU+;green),491
activecells(c-Fos+;red)andactivenewadult-borncells(BrdU+/CFos+;yellowinmergedimage).492
Arrowscorrespond tonewBrdU+/c-Fos- cells,whilst arrowheads representactivenewadult-493
bornBrdU+/c-Fos+cells.Scalebar=50µm.Statisticalanalysiswasperformedbytwo-wayANOVA494
withTukeyposthoctest.*P≤0.05,**P≤0.01,****P≤0.0001.Alldatashownaremean+/-SEM;495
n=5-8ratspergroup.AL(ad-libitum),CR(calorierestriction),CR/RF(calorierestriction/re-496
fed),WT(wild-type),GKO(ghrelin-/-).497
498

18
References499
1. Lledo,P.-M.,Alonso,M.&Grubb,M.S.Adultneurogenesisandfunctionalplasticityin500
neuronalcircuits.Nat.Rev.Neurosci.7,179–93(2006).501
2. Alonso,M.etal.Activationofadult-bornneuronsfacilitateslearningandmemory.Nat.502
Neurosci.(2012).doi:10.1038/nn.3108503
3. Grelat,A.etal.Adult-bornneuronsboostodor–rewardassociation.Proc.Natl.Acad.Sci.504
201716400(2018).doi:10.1073/pnas.1716400115505
4. Imayoshi,I.etal.Rolesofcontinuousneurogenesisinthestructuralandfunctional506
integrityoftheadultforebrain.11,1153–1161(2008).507
5. Mesholam,R.I.,Moberg,P.J.,Mahr,R.N.&Doty,R.L.OlfactioninNeurodegenerative508
Disease.Arch.Neurol.55,84(1998).509
6. Boesveldt,S.etal.Acomparativestudyofodoridentificationandodordiscrimination510
deficitsinParkinson’sdisease.Mov.Disord.23,1984–1990(2008).511
7. Andrews,Z.B.Theextra-hypothalamicactionsofghrelinonneuronalfunction.Trends512
Neurosci.34,31–40(2011).513
8. Tong,J.etal.GhrelinEnhancesOlfactorySensitivityandExploratorySniffinginRodents514
andHumans.J.Neurosci.31,5841LP-5846(2011).515
9. Malik,S.,McGlone,F.,Bedrossian,D.&Dagher,A.GhrelinModulatesBrainActivityin516
AreasthatControlAppetitiveBehavior.CellMetab.7,400–409(2008).517
10. Zhao,Z.etal.Ghrelinadministrationenhancesneurogenesisbutimpairsspatiallearning518
andmemoryinadultmice.Neuroscience257,175–85(2014).519
11. Kent,B.A.etal.Theorexigenichormoneacyl-ghrelinincreasesadulthippocampal520
neurogenesisandenhancespatternseparation.Psychoneuroendocrinology51,431–439521
(2015).522
12. Hornsby,A.K.E.etal.Short-termcalorierestrictionenhancesadulthippocampal523
neurogenesisandremotefearmemoryinaGhsr-dependentmanner.524
Psychoneuroendocrinology63,198–207(2016).525
13. Walker,aK.etal.TheP7C3classofneuroprotectivecompoundsexertsantidepressant526
efficacyinmicebyincreasinghippocampalneurogenesis.Mol.Psychiatry1–9(2014).527
doi:10.1038/mp.2014.34528
14. Li,E.etal.Ghrelinstimulatesproliferation,migrationanddifferentiationofneural529
progenitorsfromthesubventricularzoneintheadultmice.Exp.Neurol.252,75–84530
(2014).531
15. Mani,B.K.etal.Neuroanatomicalcharacterizationofagrowthhormonesecretagogue532
receptor-greenfluorescentproteinreportermouse.J.Comp.Neurol.3666,3644–3666533
(2014).534

19
16. Goetzl,F.&Stone,F.Diurnalvariationsinacuityofolfactionandfoodintake.535
Gastroenterology9,444–453(1947).536
17. Aimé,P.etal.Fastingincreasesandsatiationdecreasesolfactorydetectionforaneutral537
odorinrats.Behav.BrainRes.179,258–264(2007).538
18. Reichenbach,A.,Steyn,F.J.,Sleeman,M.W.&Andrews,Z.B.Ghrelinreceptorexpression539
andcolocalizationwithanteriorpituitaryhormonesusingaGHSR-GFPmouseline.540
Endocrinology153,5452–5466(2012).541
19. Zigman,J.M.etal.Micelackingghrelinreceptorsresistthedevelopmentofdiet-induced542
obesity.J.Clin.Invest.115,3564–3572(2005).543
20. Davies,J.S.etal.GhrelininducesabdominalobesityviaGHS-R-dependentlipidretention.544
Mol.Endocrinol.23,914–24(2009).545
21. Sun,Y.,Ahmed,S.&Smith,R.G.Deletionofghrelinimpairsneithergrowthnorappetite.546
Mol.Cell.Biol.23,7973–7981(2003).547
22. Wortley,K.E.etal.Geneticdeletionofghrelindoesnotdecreasefoodintakebut548
influencesmetabolicfuelpreference.Proc.Natl.Acad.Sci.101,8227–8232(2004).549
23. McFarlane,M.R.,Brown,M.S.,Goldstein,J.L.&Zhao,T.J.Inducedablationofghrelincells550
inadultmicedoesnotdecreasefoodintake,bodyweight,orresponsetohigh-fatdiet.Cell551
Metab.20,54–60(2014).552
24. Zhang,W.,Hu,Y.,Lin,T.R.,Fan,Y.&Mulholland,M.W.Stimulationofneurogenesisinrat553
nucleusofthesolitarytractbyghrelin.26,2280–2288(2005).554
25. Zhang,W.etal.Ghrelinstimulatesneurogenesisinthedorsalmotornucleusofthevagus.555
3,729–737(2004).556
26. Moon,M.,Hwang,L.&Park,S.GhrelinRegulatesHippocampalNeurogenesisinAdult557
Mice.Endocr.J.56,525–531(2009).558
27. Guan,X.etal.DistributionofmRNAencodingthegrowthhormonesecretagoguereceptor559
inbrainandperipheraltissues.Mol.BrainRes.48,23–29(1997).560
28. Mitchell,V.etal.ComparativedistributionofmRNAencodingthegrowthhormone561
secretagogue-receptor(GHS-R)inMicrocebusmurinus(Primate,Lemurian)andrat562
forebrainandpituitary.J.Comp.Neurol.429,469–489(2000).563
29. Sun,Y.,Garcia,J.M.&Smith,R.G.GhrelinandGrowthHormoneSecretagogueReceptor564
ExpressioninMiceduringAging.Endocrinology148,1323–1329(2007).565
30. Zigman,J.M.,Jones,J.E.,Lee,C.E.,Saper,C.B.&Elmquist,J.K.Expressionofghrelin566
receptormRNAintheratandthemousebrain.J.Comp.Neurol.494,528–548(2006).567
31. Mani,B.K.etal.Theroleofghrelin-responsivemediobasalhypothalamicneuronsin568
mediatingfeedingresponsestofasting.Mol.Metab.6,882–896(2017).569
32. Loch,D.,Breer,H.&Strotmann,J.EndocrineModulationofOlfactoryResponsiveness:570

20
EffectsoftheOrexigenicHormoneGhrelin.Chem.Senses40,469–479(2015).571
33. Prud’homme,M.J.etal.NutritionalstatusmodulatesbehaviouralandolfactorybulbFos572
responsestoisoamylacetateorfoododourinrats:rolesoforexinsandleptin.573
Neuroscience162,1287–1298(2009).574
34. Bayliss,J.etal.Ghrelin-AMPKsignallingmediatestheneuroprotectiveeffectsofCalorie575
RestrictioninParkinson’sDisease.J.Neurosci.36,3049–3063(2016).576
35. Lutter,M.etal.Theorexigenichormoneghrelindefendsagainstdepressivesymptomsof577
chronicstress.Nat.Neurosci.11,10–11(2008).578
36. Zhao,T.-J.etal.GhrelinO-acyltransferase(GOAT)isessentialforgrowthhormone-579
mediatedsurvivalofcalorie-restrictedmice.Proc.Natl.Acad.Sci.U.S.A.107,7467–72580
(2010).581
37. Soria-Gómez,E.etal.Theendocannabinoidsystemcontrolsfoodintakeviaolfactory582
processes.Nat.Neurosci.(2014).doi:10.1038/nn.3647583
38. Kola,B.etal.Theorexigeniceffectofghrelinismediatedthroughcentralactivationofthe584
endogenouscannabinoidsystem.PLoSOne3,e1797(2008).585
39. Prinster,S.C.,Hague,C.&Hall,R.A.HeterodimerizationofGProtein-CoupledReceptors :586
SpecificityandFunctionalSignificance.57,289–298(2005).587
40. Wellman,M.&Abizaid,A.GrowthHormoneSecretagogueReceptorDimers:ANew588
PharmacologicalTarget.eneuro(2015).589
41. Egecioglu,E.etal.Ghrelinincreasesintakeofrewardingfoodinrodents.Addict.Biol.15,590
304–311(2010).591
592
593
21
594
Figure1.Ratcliffetal.
ii.
iii.
i.
22
595
Figure2.Ratcliffetal.
WT
GHSR-nullGHSR-null
WT
Saline Acyl-ghrelinA. B.
23
596
Figure3.Ratcliffetal.
5 6 7 8
24
597
598
Figure4.Ratcliffetal.
A. B.
F.E.
D.C.
G.

25
599
600
Subventr
icula
r zone (
SV
Z)
Cin
gula
te c
ort
ex
GHSR1a
WT GHSR-null
Figure S1. Immunoreactivity (IR, white arrowheads) for anti-
GHSR1a was observed in the
adult SVZ of both WT (A) and
GHSR-null (B) mice. Similar IR
was observed in the cingulate cortex of WT (C) and GHSR-
null (D) mice. These data
suggest that the rabbit anti-
GHSR1a antisera cross-reacted
with nonspecific antigen in the adult mouse brain. Scale bar =
50µm.
A
D
B
C
26
601 TableS1.Ratcliffetal.
Table S1. New adult-born OB cells are activated by calorie restriction in a
ghrelin-dependentmanner.Main effects of two-way ANOVA. *p<0.05; **p<0.01;
***p<0.001;****p<0.0001wereconsideredstatisticallysignificant.GCL(granulecell
layer), GL (glomerular layer), SEZ (subependymal zone), AOL (anterior olfactory
nucleus,lateral).





























