Embed Size (px)
Citation preview

C- N.it~nndl L~brnry B~bl~othbque nat~onale , CANADIAN THESES TH&W CANADIENNES u
- - of Canxin - dt+ Canada - --- -- -- O W M~C3tOF1CHE- - - - SUR MICROFICHE - - - - -
on nitrogen nutrition in Zalerion m a r i t b u m .
, Y t l t i i 714 15 DtCflEE CONFERRED/ANN~!E DD'08TENTICW DE CE GRADE 1 Q 7 A I
M.:E. OF SUPERVISOR/NOM DU OIHECTEUR DE THZSE Dr. PI. McClaren
I 4 4
. , 0
Peisn~s's~on rs 'hereby grasted ta the NATIClNAL [email protected] OF L'eu€orisatiun est, par la pr4sente,-accord& B f a . 8 1 8 ~ 1 0 ~ ~ ~ - t
1
CANADA to mtcrofilm t h ~ s thesis and to lend or sell copies ' OUE AIATIONALE. DU CANADA de microfilmer cette thdse et 3
de prtfter w de vendre des exhnplaires du film.
Tlio nulltor reservos other pub1 icati on,rights, and neither the L'sutew se rdserve les eutres droits de publication; ni In -
ttw:,~s nor extensive extracts from i t may bipr inted or dher- thdseni de longs extraits de celle-ci ne doiwnt itre imprrrrrt!~
v;lsc reproduced without-the author's written permission. w autremsnt reprduits sans /'eutorisatitm Bcrire de I'arrteur. r

+ National .Library of Canada \
Cataloguing Branch Canadian Theses Division
Ottawa, Canada -
- KIA ON4 --
- 'NOTICE
Bibliotheque nationale du Canada ; \
Direction du cat logage Division des the t es canadiennes - - -- --
The quality of this microfiche is heavily dependent upon ' La qualite de cette microfiche d e p w grandement de la . the quality of the orignat thesis submittehtotpicrofi lh- quatite de la thesesoumiseau micrafilmage Nousavons--
ing. Every effort has been made to ensure the highest tout fait 'pour assurer une qualite superieure de rebo- quality of reproduction possible. , - duction. . I
€2 8 . \.
\ , If ;ages are missing, contact the universiiy which S'il manque des pages, veuillez &ornrnuniquer w e d
'granted the degree. I'universite gui 8 confere le grade.
Some pages may have indistinct print especially' if de certaines pages peut the original pages were typed with a poor typewriter pages originales ont ete ribbon'or if the university sent us a poor photocopy. a I'aided'bn ruban use ou si I'universite
nous a fait parvenirvlne photocopie de mauvaise qualite.
~reviously copyrighted materials (journal articles, ~ e s documkntsgui font dejh 1:objet d'un droit d'au- published tezts, etc.) are not filmed. teur (articles de revue, examens publies, etc.) ne sont pas
. I microf ilmes. I
N Reproduction in fult or in part of this film is governed . La reproduction, mbme partielle, de ce microfilm est by the Canqdian Copyright Act, R.S.C. 1970, c. C-30. soumise a la Loi canadienne sur !e droit d'quteyr, SRC +
Please read the authorization forms which accompany 1970,* c. C-30. Veuillez prendrs connaissance des for- this thesis.
j r mules d'autorisation qui accompagnent cette these.
* a
- THIS DISSERTATION LA THESE A ETE~ HAS BEEN MICROFILMED M ~ C R O F ~ L M ~ E TELLE QUE - EXACTLY AS RECEIVED NOUS L'AVONS RECUE.

WITH EMPHASIS ON NITROGEN lTUTRITION I N
Z A L E R ~ N MANTIMUM
V
* - by .
d - -- L 1
Lesl ie Marian Churchland
B.A. : University of Br i t i sh C o l T a , '1966 ( M.Sc., Simon Fraser University,
1
1 > -L . I e
1 '4. . @ . *
I - 3 - '
b ' 3 / p i n the ~ ) e ~ a > , k n t . \
* , I Biological S i e n c e s
0 Lesl ie Marian Churchland 1971 " , i
Simon Fraser University , September, 1976 '3
A l l r i gh t s reserved. This t hes i s may not be reproauced ?in whole o r in' part , by p otocophy o r other means, without p e d s s i o n of the author. % ' T h. . .

Name : , L e s l i e Marian Churchland
Degree : Doctor of Philosophy
T i t l e of Thesis :, N u t r i t i o n a l S tud ies of ~ a r i n e ~ u n $ Imperfect i , w i t h Emphasis on Nitrogen N u t r i t i o n i n Zaler ion mari timum
Examining Commi t r e e :
Chairperson : R . - -
/?
I w I -
M. McClaren Senior Supervisor
- L. D. Druehl
0 /-'- W Y T ' , - - " , , .
J. E. Rahe
- 1 / ' / - / I
G i l b e r t C. Hughes Ex te rna l Examiner . I i
Professor 0 2
Department of Botany and I n s t i t u t e df Oceanography --
m v e r s i t y o f ' B r i t i s h Columbia
Date Approved:. '7 - ,976

I h e r e b y g r a n t t o Simon F r a s e r U n i v e r s i t y the r i g h t t o lend '
my t h e s i s o r d i s s e r t a t i o n ( t h e t i t l e of which i s shown below) t o u s e r s 1 8
of t h e Simon F r a s e r U n i v e r s i t y L i b r a r y , and t o make p a r t i a l o r s i n g l e
c o p i e s o n l y f o r s u c h u s e r s o r i n r e s p o n s e t o a r e q u e s t ,from t h e l i b r a r y *
of a n y o t h e r u n i v e r s i t y , o r o t h e r e d u c a t i o n a l i n s t i t u t i o n , on i t s o w n - 3
-
b e h a l f o r f o r one of i t . s u s e r s . I \ f u r t h e r agree t h a t peymiss ion f o r
< m u l t i p l e copying of t h i s t h e s i s f o r s c h o l a r l y purposes may be g r a n t e d
- by me o r the Dean of Graduate S t u d i e s . Tt i s unders tood t h a t copy ing , s
o r p u b l i c a t i o n of t h i s t h e s i s L o r f i n a n c i a l g a i n s h a l l n o t be a l lowed *
I. w i t h o u t my w r i t t e n perrniss.ion. t
/-
T i t l e of ~ h e s i s / ~ i s s e r t a t i o n :
Nutritional studies of Marine Fungi Imperfecti with e m p s $
0 'd on nitrogen nutrition in Zalerlon m a r i t h u m
( s i g n a t u r e )
LESLIE MARIAN CHURCHLAND -
(da te ) i

N u t r i t i o n a l S t u d i e s Kf HaFine F u n g T I m p e r r e c t l w i t h Emphasis ' -
oh Ni t rogen N u t r i t i o n i n Za l e r ion Maritimugl 3,
ABSTRACT -
r
A con t inuous cu1tur.e. system was designed f o r t h e c u l t i v a t i o n o f
-- marine f u n g i . Growth of Huu&eola a l o p a l l o n e l 3 a -Meyers md-Moore, - - -
Monodic t y s pe l ag i ca ( ~ o h n s o n ) f ones , and Za le r ion mari timum ( ~ i n d e r ) I
Anas tas iou was h i g h e r i n con t inuous c u l t u r e than i n b a t c h c u l t u r e .
Experiments were c a r r i e d o u t t o e x p l a i n t h e poorer growth i n b a t c h
c u l t u r e . Experiments w i t h r e i n o c u l a t e d media showed t h a t n o n - v o l a t i l e
s t a l i n g p roduc t s were n o t p r e s e n t . Nu t r i en t exhaus t ion was a f a c t o r f
, i n Humicola a l o p a l l o n e l l a . I n a l l s p e c i e s , low pH c o n d i t i o n s i n
ammonium c o n t a i n i n g media l i m i t e d growth. Improved pH c o n t r o l was Y
ob ta ined by s u b s t i t u t i n g MES (2-[N-morpholino] e thanesu$ fon ic a c i d ) I
~ O ~ T R I S . ( t r i s [hydroxymethyl] aminomethane). Chemical a n a l y s e s of
n u t r i e n t s i n t h e n u t r i e n t s o l u t i o n i n d i c a t e d t h a t phosphate was -~~-rr,\
d e p l e t e d i n ba t ch c u l t u r e .
. \
Two i s o l a t e s o f - Z . maritimum, FMC and H 135, were used t o s t u d y
T i t r o g e b k t r ~ t i o n . Both i s o l a t e s g r w on each o f N H 4 C 1 , NaN03,
bnd NaN02. Of 21 amino a c i p s t e s t e d , b a l a n i n e and L-arg in ine . g-
- B - suppor ted b e s t growth o f t h e FMC i s o l a t e . L-valine , L-alanine ,
L-glutamic a c i d , L -gspa r t i c a c i d , L-asparagine, and L-arg in ine , 3
suppor ted good growth o f H 135. Both i s o l a t e s grew on peptone , u r e a , - iii
1 b

citruli l?i ,e, and a l l a n t o i n . -- R e s u l t s from t h e con t inuous sygtem - - -pp--p
d i f f e r e d both q u a l i t a t i v e l y and q u a n t i t a t i v e l y from ba t ch c u l t u r e . .
r e p u l t s . I n t h e FMC i s o l a t e , t h e r e was g r e a t e r growth i n b a t c h
c u l t u r e on L - ~ l ~ t a m i c a c i d t han on t h e t h r e e i n o r g a n i c n i t r o g e n 7
sou rces . I n con t iguous c u l t u r e , growth was equa l on a l l n i t r o g e n I I /
Nit rogen compounds were s u p p l i e d i n combination, and t h e i r
removal from t h e n u t r i e n t s o l u t i o n measured chemica l ly . Both i s o l a t e s - ,$
/ t ook up ammonium i n p r e f e r e n c e t o n i t r a t e . Although both I s o l a t e s
/ t ook up ammonium apd n i t r i t e , ammonium was removed a t a f a s t e r r a t e . 1
I s o l a t e FMC p r e f e r e n t i a l l y took up L -glutamic a c i d when s u p p l i e d w i t h
n i t r a t e . I s o l a t e H 135 took Up bo th compounds, t h e former a t a f a s t e r
rate than t h e l a t t e r . Both i s o l a t e s took up ammonium and a group of
e i g h t amino a c i d s , when s u p p l i e d i n combination.
- - Nei ther i s ~ l a t e demonstrated n i t r o g e n f i x a t i o n , d e n i t r i f i c a t i o ~ ,
o r n i t r i f i c a t i o n . Za l e r ion maritimum had a p a t t e r n o f n i t r o g e n 1 s ,
n u t r i t i o n s i m i l a r t o thaf? o f t h e t e r r e s t r i a l f u n g i .

ACKNOWLEDGEMENTS - ,
- - - --
> * I s i n c e r e l y thank my s u p e r v i s o r , D r . M. McClaren, f o r h i s -
encouragement and s u p p o r t d u r i n g my g radua t e s t u d i e s a t Simon F r a s e r
Un-ivers i ty . The members o f my s u p e r v i s o r y committee, D r s . L. D.
Druehl and L. A l b r i g h t , have a l s o been most h e l p f u l i n o f f e r i n g
s u g g e s t i o n s and c r i t i c i s m s d&ing t h i s r e s e a r c h . P a r t i c u l a r tKanks - - --
--
a r e due t o D r . ~ h a r l e s Wilson, D r . Melvin Goldstein-, and t h e Biology
Department of McGill U n i v e r s i t y , who k i n d l y provide8d me w i t h , .
l a b o r a t o r y space and equipment d u r i n g a year's l e a v e o f absence i n
Montreal .
Mrs. Ann Jenks g a v e ' v a l u a b l e t e c h n i c a l a s s i s t a n c e d u r i n g t h e r
m a j o r i t y o f t h i s investigation. I a i s o thank M r . Ron Long f o r h i s
photographic a s s i s t a n c e , Mr. M. Yang f o r amino a c i d a n a l y s e s , and i
-
t h e pe r sdnne l o f t h e g l a s sb lowing shop f o r t h e i r h e l p i n c o n s t r u c t i n g
t h e appa ra tu s . M r . B r i an Mauza, Mr. Conrad C l o u t i e r , and Miss M.
Cheveldayo f were ve ry h e l p f u l i , . i n i n t r o d u c i n g me t o t h e u se o f t h e
- computer. . <
. I p a r t i c u l a r l y thank my husband Dr/ &rlc ~ h u r c h l a n d f o r h i s . .
/ ' * - adv ice and good n a t u r e d s u p p o r t d u r i n g t h e 'cO,urse o f t h i s research: . '
i
I a l s o thank t h e Na t iona l Research Council of Canada f o r . t h e i E '
- - 1
f i n a n c i a l suppor t .


, - - - - . - -
. Paqe - - -
i I
C . < Growth i n con t inuous and ba t ch sys tems ............ : %
-- - - - -- - - - Z D
--- I
.................. D. Experiments w i t h r e i n o c u l a t e d media * 23 .
E. Buf fe r exper iments e ................................. 24 * -' n
.... F. N u t r i e n t up take exper iments p................... 27 I >
- . Discuss ion .............................................. 28 I
Chapter 11. Nit rogen n u t r i t i o n i n Z a l e r i o n mari timum 5-
t ate rials and Methods ........... : ......................... 39
A . Growth o f inoculum osn v a r i o u s , @
4 9 n i t r o g e n sources . . . . . . . . . . . . . . . . . . . . . . , . . . . . . . . . . . . . . . L
A
9
> ? 't . .
B. Growth on i n o r g a n i c n i t r o g e n s o u r c e s .....+.......... .. * *
t
C. Growth on o r g a n i c n i t r o g e n s o u r c e s ...,.............. 42 . - - --
- D. Removal o f n i t r o g e n s o u r c e combinat ions f ~ o m t h e n u t r i e n t s o l u t i o n .......... ;..............*...... . , 43
1) .' . E. Conversions o f n i t r o g e n ........................ ..-. 46
. * *
R e s u l t s . . . . . . . . . . . . . ..................................... X f 8
A . ' ~ r o w t h . o f inoculum on varpious n i t r o g e n ' . . ............................................ f s o u r c e s 4 8 Q
1 I 0 . B. Growth on i n o r g a n i c n i t r o g e n s o u r c e s :............... 48 \
- . .I C. Growth on o r g a n i t r o g e n s o u r c e s ................. Y- A'
54
D. Ilemoval o f n i t h g e n sou rce combinat ions from ' 64 . t h e n u t r i e n t s o l u t i o n ........... : .................... . ,
E. Conversions o f n i t r o g e n soui-ces..................... 72
. Discussion.. . . . . . . . , . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 74- - - .-- - ~ - -- -- pp-pppp---p-
...... ,i--. - c 1 +Summary .....$....... . . . . . . . . . . . . . . . . . . . . . . . . . . . . 92
Lp-~- t -
'a L i t e r a t u r e c i t e d ....................................... 9 4 1
Y Curriculum v i t a e ..................... ;........:...:....

Table I. Growth o f 2 . maritirnum FMC, M. p e l a g i c a , and H, a l o % l l o n e l l a i n b a t c r c u l t u r e -
' on r e i n o c u l a t e d media. .
Table 11. E f f e c t o f d i f f e r e n t c o n c e n t r a t i o n s o f , THAM and MES b u f f e r s on g r o w t f o f - -L-m&&wntFMC - i n -bat-ch cu lpu re . A-
Table 111. The effect. o f growing inoculufk i n v a r i o u s n i t r o g e n s o u r c e s on growth o f Z . maritimum FMC i n b a t c h - c u l t u r e . -
Table I V . The e f f e c t oaf i n o r g a n i c n i t r o g e n s o u r c e s and 1-glutamic a c i d on growth o f
,/ - Z'. maritimum FMC i n contin,uous and - b a t c h c u l t u r e .
Table V. ~ h b e f f e c t o f i n o r g a n i c n i t r o g e n s o u r c e s and 1-glutamic a c i d on growth o f Z . maritimum H 135 i n cont inuous and - ba tch c u l t u r e . \il 5 1
,
c 3
Table V I . The effkdt o f d i f f e r e n t a =-. o f n i t r a t e o a r o w t h o f
FMC i n cdn t inuous and 54
j . able V I I . The effect o f d i f f e r e n t c o n c e n t r a t i o n s
- , . of n i t r a t e on gr5wth of 2. .maritimum B 132 i n on t inuods and ba tch Cu l tu re . ,
J -\ 56 f \ Table V I I 1 . - The effect o f d i f f e r e n t c o n c e n t r a t i o n s
o f n i t r a t e and gluco'se on growth o f . . I
Z. maritimum FMC i n cont inuous and .z - \ 4 b a t c h c u l t u r e . r e 57
/ - - Table I X . The e f f e c t of d i f f e r e n t &oncen t r a t i ohs . o f n i t r a t e and g lucose on growth* o f
I -2: &rHAmi&-H +35-i*co~t+ m d - Y pppp- - \ - b a t c h c u l t u r e . 58
growth o f , 2. maritimum FMC and H 135 #
i n ba t ch c u l t u r e . . > - . I '60 ' <: L
/ - 3 n
0 LP " i viii -

- - -- - - page - - - - - - -
r
' d ~ a b l e XE.' The e f f e c t o f a combinat ion o f &in0 ~ i ; E t s ~ ~ e F Z , ~ i n cont inuotm and bz t ch c u l t u r e . 6 3
Table X I I . The ef?ect o f 3 combinat ion o f amino , %
' a c i d s on growth o f Z. m a r i t i r n u m H 135 i n continuous'and ba t& c u l t u r e .
Tab le X I I I . The e f f e c t o f o r g a n i c n i t r o g e n gources on growth o f Z . maritimum FMC i n b a t c h c u l t T r e .
Table, X I V . Th@ bffect 'of o r g a n i c n i t r o g e n s o w L e s an &mtb -of i~ m a c i t d m u m H 135 0
- --
/ i n ba tch c u l t T ~ e . 67 n
able\ XV. Changes i n c o n c e n t r a t i o n s o f amino. . . - L - a c i d s i n ' t h e n u t r i e n t s o l u t i o d du r ing a
A > -1
growth o f Z . maritimum FMCain ba tch Q - -
c u l t u r e . -
L 71 3
Table XVI. ' Changes i n c o n c e n t r a t i o n s o f amino ,. a c i d s i n t h e n h t r i e n t s o l h i o n
, dur ing growth .ofFZ. - marit3mum H 135 ba tch c u l t u r e .

t "
I . J s I86 laaws--- -- - -
&
I 3
Page 1 <
Fig . 1 . Diagram of a con t inuous c u l t u r e *
a p p a r a t u s for ' t h e growth o f - f i l amen tous marine f u n g i . 14
-.-4 a s
Fig . 2. , Photograph of a con t inuous c u l t u r e q. ' \ - - - appa ra tu s f o r t h e g rowth -o f * a
f i l amen tous --- m a r i ~ f w n g l , 15 - 4
Fig . 3. Growth of Z. maritimum H 135 and FMC ' - on seawater a g a r . 21 - .
. . Fig . 4. 'Growth of Z. marl- FMC, a. -, and u. w i c a i n con t inubus and ba tch c u l t u r e . , 22
Fig . 5 . Changes i n d r y we igh t , p ~ , and 4 c o n c e n t r a t i o n s of gAucose, phosphate , %
ammonium, and n i t r a t e d u r i n g growth i n b a t c h c u l t u r e o f Z. maritimum FMC and H 135. 28
- Fig . 6, Changes .in pH d u r i n g growth o f
z.' maritimum FMC i n con t inuous c u l t u r e . - 5 3 *
Fig . 7.' Changes i n d r y we igh t ,, pH, and c o n c e n t r a t i o n s of g l u c o s e , phosphate , - ammonium, and n i t r i t e d u r i n g grawth i n Gatch +
c u l t u r e * o f Z. ' maritimum FMC and H 17% 68 i,
1 3 I
'F
Fig . 8. Changes i n d r y we igh t , pH, and c o n c e n t r a t i o n s ; 4 i
of 1-glutamic a c i d and n i t r a t e du r ing .A
growth i n b a t c h c u l t u r e o f L. maritimum, . ';j FMC and H 1 35. , 70 4
- . a Fig . 9. Summary diagram of i t r o g e n n u t r i t i n i n . f 3 f
7 5 4 Z. mar i t im - -7 ... %3
., $
- - - --- -- L--+
Is - - - - -- - ---
P - - -
. -
4
7' 1
. L , $ X , ' .
t " >- * \ -'
, , .

The s tudy 'o f l i g n i c o l o u s marine fungi Gas begun by Eiargh*orn ,
J
(1944) and Linder ( 1944) * i n a combined s tudy o f . t h e taxonomy and 3 4 ;E
u * t
biology of marine fung i . T h i s work included ex tens ive taxonomic and ' 1 '
phys io log ica l informat ion , and was followed by t h e "descr ip t ive phasen i 3
i d
and l i s t e d s p e c i e s *dm v a r i o u s h a b i t a t s and &ographica l l o c a t i o n s . I
Taxonomic2 and f l o r i s t i c s t u d i e s a r e still an important p a r t o f marine \ --
? mycology, because, all t h e p o s s i b l e s u b s t r a t e s f o r marine fung i have C -
no t y e t been i n v e s t i g a t e d .
C
During th i s pe r iod , mycologists also i n v e s t i g a t e d t h e effects g f
c e r t a i n marine v a r i a b l e s on t h e gro,uth and d i s t r i b u t i o n o f s p e c i e s .
Temperature, s a l i n i t y , and t h e w b i n e d e f f e c t s the reof were s t u d i e d ,
both i n t h e l a b o r a t o r y ( R i t c h i e , 1957; Ri tchie and Jacobsohn, 1963)
and i n t h e f i e l d (Gold, 1959). g 'ubstrate preferencea observed i n t h e "4 '
f i e l d were t e s t e d i n t h e l a b o r a t o r y , and s e v e r a l s p e c i e s were shown t o P *
degrade c e l l u l o s e ' (Meyers and Reyndlds, 1959 ; Meyers , Priridle , and , b
Reynolds, 1960; Moore and Meyers, 1962).
Hughes (1975) h a s w r i t t e n a comprehensive review of s t u d i e s s i n c e
1961 on l i g n i c o l o u s , cau l i co lous , and f o l i i c o l o u s s~ec i e s . . I n r e c e n t
y e a r s , t h e emphasis has s h i f t e d from d e s c r i p t i v e t o n u t r i t i o n a l and
phys io log ica l s t u d i e s . The growth of marine fungi on var ious carbon

and- --so- tiTlxtemmnxi'i& Jones, 1 Y[ 4 ; Sguros , nodrigues , and
Simms, 1973) and t h e i o n i c c o n s t i t u e n t s o f seawater (Sguros and Simms,
1964; Jones and Jennings, 1965) have been inves t iga ted . The s a l i n i t y
requirements of both higher ( Tubaki , 19 66 ; ones , Byrne? and Alderman,
1971 ; Davidson, 1974a; Schaumann, 1974.) and lower (Aldermali and Jones,
1971 ; Harrison and Jones, 1974; Bremer, 1974) k r i n e fung i h a w been
skudie&.- -Wad&&o~&.f-y, e i ~ ~ r a b i a * ~ o w t ' h o r dry-wefgm- - - - - -
measurements have been used . to measure t h e growth o f marine fungi .
Spore production (Meyers and Hoyo, 1966; Byrne and 3ones, 1975b) and *
spore germination (Byrne and Jones, 1975a) a r e c u r r e n t l y being used a s
a d d i t i o n a l measures o f phys io log ica l response.
I n the 1 9 7 0 ' ~ ~ t h e r e has been increased i n t e r e s t i n marine fungal
biochemistry. The production of me tabo l i t e s such a s cho l ine s u l f a t e ,'
ester (Catalfomo e t a l , 1972-1973), e r g o s t e r o l , and etrczliae (Kirk and - . Catalfomo, 1970) have been documented. Pathways o f carbohydra te
metabolism such a s t h e Tr ica rboxy l i c Acid Cycle (Vembu and Sguros,
1972) and t h e Hexose Monophosphate Pathway (Holl igan and Jennings,
1972b; Lowe and Jennings, 1975) have been i n v e s t i g a t e d . The marine
fung i have been shown t o degrade such environmentally important
compounds a s cyclodiene p e s t i c i d e s (Quevedo, Sguros, and Thomann,
fung i , t h e i r role in c e r t a i n e s s e n t i a l marine processes is unknown. d

One of t h e most important cha l l anges i n marine mycology is t o c r e a t e
l abora to ry cond i t ions which appro'ximate t h e marine environment, and
thus a l low an a p p r a i s a l of t h e r o l e of thede-organisms i n t h e i r
n a t u r a l h a b i t a t . The g o a l s of t h i s t h e s i s were a s fol lows: . ?
I
1. To develop a continuous c u l t u r e system f o r t h e c u l t i v a t i o n of --
- - LA - - - A -
4 f i lamentous marine fung i (Chapter I) . 5
2. To compare growth i n c t inuous and batch c u l t u r e and t o ph 1
determine what f a c t o r o r f a c t o r s a f f e c t growth i n ba tch c u l t u r e
(Chapter I ) .
f 3. To use t h e cont ingous system and t h e batch system t o s tudy
n i t rogen n u t r i t i o n i n Zaler ion maritimum (Chapter 11).
4. To propose a hypo the t i ca l r o l e f o r &. maritimum i n t h e marine
n i t rogen c y c l e , t ak ing i n t o account t h e d i f f i c u l t i e s o f
e x t r a p o l a t i n g from i n v i t r o s t u d i e s t o t h e marine environment
(Chapter 11)'.

Growth o f Zaler ion m a k t i m u m , ~ u m i c o l a a l o p a l l o n e l l a , and
Monodictys pe lagica i n continuous and batch cu lcure . ..
INTRODUCTION ki . In e a r l y n u t r i t i o n a l s t u d i e s o f marine fung i , growth was measured
a s i n c r e a s e s i n colony diameter on a g a r (Barghoorn, 1944; Johnson,
Ferchau, and Gold, 1959). This technique had t h e d isadvantages t h a t
t h e n u t r i e n t s o l u t i o n could not be chemically def ined dur ing growth,
pH c h a ~ g e s could n o t be monitored o r ad jus ted , and no c o r r e l a t i o n
could n e c e s s a r i l y be made between r a d i a l growth and fungal biomass
(Mandels, 1965), Radial growth measurements a r e no t g e n e r a l l y used
f o r n u t r i t i o n a l s t u d i e s today, al though they have been used t o s tudy
t h e e f f e c t s of temperature and s a l i n i t y on growth (Tubaki, 1966; 1
Schaumann, 1974). The r a d i a l growth technique was replaced by dry v
weight de terminat ion , i n which t h e mycelium was removed from a s t a t i c
l i q u i d c u l t u r e medium, d r i e d , and weighed (e.g. Gustafsson and F r i e s O
T
[1956]). This technique was then modified by incubat ing t h e c u l t u r e
f l a s k s on a shaker. This "batch c u l t u r e * technique is now g e n e r a l l y

In t h i s s tudy , I at tempted t o i n c r e a s e t h e growth r a t e s of marine
fungi grown i n a chemically def ined n u t r i e n t so lu t ion . I designed a
continuous flow system f o r t h e c u l t i v a t i o n of .filamentous marine
fung i , and compared t h e growth t h e r e i n with growth i n ba tch c u l t u r e .
Fungal may a f f e c t t h e chemical environment i n ba tch - -
-
4
c u l t u r e . F i r s t , one o r more n u t r i e n t s may be exhausted. Second,
depending upon t h e n i t rogen source pH changes may occur. Growth on
P d* ammonium lowers,pH, whereas growth on n i t r a t e and o rgan ic n i t rogen
raises pH (Sguros and Simms, 19164). These changes may occur d e s p i t e
t h e add i t ion o f b u f f e r , o r t h e buf fe r i t s e l f may be t o x i c t o t h e
fungus (Cochrane, 1958) i n concen t ra t ions high enough t o compensate
f o r t h e pH e f f e c t . Third, byproducts o f fungal metabolism may a f f e c t
growth i n a c losed system. Nitrite may accumulate and be t o x i c i n
a c i d i c media con ta in ing NH4N03. Rapid growth may be followed by
t h e production of s t a l i n g products , me tabo l i t e s which i n h i b i t fungal '
growth. ~ e c a d s e marine fung i a r e n a t u r a l l y bathed i n a renewable
n u t r i e n t s o l u t i o n , t h e use of a continffous c u l t u r e system may more - -
a c c u r a t e l y determine t h e i r n u t r i t i o n a l requirementa. A continuous rr
system should, t h e o r e t i c a l l y , supply f r e s h n u t r i e n t s , c o n t r o l pH
*changes, c o n t r o l osmotic changes, and prevent t h e accumulation o f
- - - - -- - - - - -
T e r r e s t r i a l f u n g i such as Coprinus ( F r i e s , 19561, Asperg i l lus 4
(Ng, Smith, and McIntosh, 19731, and usarium (Steensland, 1973) have T-

- ?,. - - - - - - --
been-grown i n cont inuous c u l t u r e . Johnson and Gold (1959) designed a
technique f o r t h e cont inuous recyc l ing o f seawater over marine fung i
growing on wood. However, a l l previous n u t r i t i o n a l s t u d i e s o f marine
fungi have been c 'arr ied out on a g a r o r i n batch c u l t u r e .
I n v e s t i g a t i o n - o f continuous c u l t u r e techniques l e d t o t h e --- - - --- -- - -
ques t ion of growth dynamics; why did growth s t o p i n t h e c losed system?
Was it due t o n u t r i e n t exhaust ion; i f so, of which n u t r i e n t o r
n u t r i e n t s ? Did s t a l i n g products i n h i b i t growth? Did e s e s p e c i e s a 2 grow determinate ly a s do high'er p l a n t s and some o t h e r fung i ' ( ~ o t t l i e b , . 1971)? To answer t h e s e ques t ions I compared growth i n ba tch and
continuous c u l t u r e , reinoculated used media w i t h fresh mycelium, and
analyzed spent medium f o r g lucose , ammonium, n i t r a t e , n i t r i t e , and
phosphate.
The t h r e e s p e c i e s used f o r t h e n u t r i t i o n a l experiments were -
Monodictys pe lagica (Johnson) Jones, Humicola a l o ~ a l l o n e l l a Meyers and
Moore, and Zalerion maritimum (Linder) Anastasiou. Each has-been
i s o l a t e d commonly from c e l l u l o s i c s u b s t r a t e s i n B. C." 'coastal waters
(Meyers and Reynolds, 1960; Hughes, 1969 ; Anastasiou and Churchland,
1969 ; Churchland and McClaren, 1973). Zalerion mapitimum and
M. p e d ~ i c a ha_e been i s o l a t e d from n o r t h a n d south temperate waters , . -- and occas iona l ly from t r o p i c a l and s u b t r o p i c a l s e a s ( ~ u g h e s , 1974).
-- -- - - - - - - - - - - - - - .---- -
U m i c o l a a l o ~ a l l o n ~ h a s been a s f requent ly i s o l a t e d *om t r o p i c a l
a s temperate waters (Hughes, 1974). A l l t h r e e s p e c i e s a r e *marinew

- - -
sensu Kohlmeyer (19741, i n t h a t they bo thgrow and reproduce i n t h e
marine environment: Monodictys pe lagica might be b e t t e r termed a
nsaltwatern fungus, a s it has been found i n a s a l i n e l a k e i n Wyoming
(Davidson , 1974b ) .
The major i ty of t h e work i n t h i s t h e s i s was done with bhe marine - - - -- - - - - - - - - - -
Imperfect spec ies Zalerion maritimum (Linder) Anastasiou. Linder
(1944) o r i g i n a l l y desc r ibed t h i s s p e c i e s a s Helicoma maritimum, and
separa ted i t from H. sal inum by t h e number of c o i l s and s e p t a t i o n s i n - b
t h e spores . In 1962 Moore and Meyers placed t h e s e two s p e c i e s i n ' t h e
genus Zaler ion , which d i f f e r e d from Helicoma by t h e presence o<
c o n s t r i c t e d s e p t a i n t h e s p o r e s and by t h e number of gyres i n t h e
spores . The genus was then d iv ided by Moore and Meyers i n t o four ,-
f ' . ,-
s p e c i e s ( Z . - e i s t l a -9 - Z. nappura, 2. x y l e s t r i r , and Z. r k p t o r ) , based . - 8 '
upon c o l o n i a l morphology, pigmentat ion, an? c e l l u l o s e u t i l i z a t i o n . , 4 . .
Subsequently, Anastasiou (1963) recombined t h e four s p e c i e s i n t o one 1 .
s p e c i e s , Zalerion maritimum, which included t h e 11. maritimum and H. I
C
salinum descr ibed by Barghoorn and Linder. Anastasiou considered
~ e y e r s ' four s p e c i e s of Zaler ion t o be physio logica l v a r i a n t s , and
t h i s i n t e r p r e t a t i o n i s now g e n e r a l l y .aorcepted (Tubaki, 1966; Hughes, 3
1975). Considering t h e phys io log ica l v a r i a b i l i t y of t h i s s p e c i e s I
have used twGf 901aCesj-drf ~ F i n g ~ ~ n c o l o i i T a l m O r p h o l o ~
pigmentation, fbr most w f t h e m x t r i t i o n a l studies.- - - - - - - - -- -- --

--- - - The ,go& of t h e experiments described i n t h i s chapter were' t o .
design a continuous c u l t u r e system f o r t h e growth o f marine f u n g i , t o
v' compare growth i n cont inuous and batch c u l t u r e , and t o determine what
f a c t o r o r f a c t o r s a f f e c t e d growth i n ba tch c u l t u r e . These experiments \
were designed t o i n c r e a s e our understanding o f t h e growth o f marine
.fungi, and al low t h e formulat ion of more balanced c u l t u r e medii f o r
-- - - - - - - +
f u t u r e n u t r i t i o n a l s t u d i e s .
MATERIALS AND METHODS
A . I s o l a t i o n o f t e s t s p e c i e s from t h e marine environment t I
i
Monodictys pe lagica was i s o l a t e d from a hemlock (Tsuga
heterophyl la [Raf.] Sarg . ) panel submerged.at 1 m f o r 2 months a t I
Horseshoe Bay, B r i t i s h
1969.
I
Humicola a l o p a l l o n e l l a
Columbia. The panel was c o l l e c t e d i n August, pi
Humicola a l o p a l l o n e l l a was i s o l a t e d from a cedar (Thus p l i c a t a - - - - - - -- - -- - -
DoM.) panel submerged a t 1 m f o r 4 months a t Cotton Po in t , Keats - -- -- - - - - -- - - - - - - - - - -
I s l a n d , B r i t i s h Columbia. The panel was c o l l e c t e d i n October, 1969.

Zalerion mari timum H 1 35
Zalerion maritimum H 135 was i s o l a t e d from a Douglas F i r
( P s e u d o t s u ~ a 'menziesi i [Mirb.] Franco) panel submerged at ' 1 m for 1 . .
6 \
' month a t Horseshoe Bay, B r i t i s h Columbia. The panel was c e i l e c t e d i n
Zaler ion maritimum FMC pS
-
Zalerion maritimum FMC was d s o l a t e d from a hemlock panel L
submerged a t 1 m f o r one month at. .Lions'Bay, B r i t i s h Columbia. The
panel was c o l l e c t e d i n J u l y , 1971. J
After removal from t h e s e a , t h e panels were incubated (Johnson
and Sparrow, 1961) 1-3 weeks a t 20 C. A hypha a s then removed from
t h e panel and placed on seawater a g a r (Johnson and Sparrow, 1961).
k. After autoclaving, 0.5 g / l of sir ptomycin s u l f a t e and 0.5 g / l of
P e n i c i l l i n G hag been added t o t h e seawater aga r . A s i n g l e hyphal t i p
was taken from t h e edge of t h e r e s u l t a n t colony and placed on seawater
a g a r without a n t i b i o t i c s . The c u l t u r e s were maintained on
seawater-yeast e x t r a c t s l a n t s a t 4 C. To demonstrate v a r i a t i o n i n -- -- - - - - - - - -- pp-pp-pp
morphology and pigmentat ion, t h e two i s o l a t e s o f Zaler ion were c
photographed a f t e r 3 w e e ~ s ' growth on seawater-yeast e x t r a c t a g a r a t - > - -
20 C.

. B. Media
The chemica l ly d e f i n e d l i q u i d c u l t u r e medium used .in t h e *
n u t r i t i o n a l s t u d i e s was t h a t o f Sguros and'simms (19631, and con ta ined \ I .
g l u c o s e , 9.0 g ; N H 4 N 0 3 , 0.4 g ; K2HP04 . 3H2@: 0.06.g; THAM 'e a
( t r i s [hydroxymethyl] aminomethane), 1.2 g ; v i t amins , trace e l emen t s , a n d 1 0 0 0 m l
f - - - -- - - -
Y of a r t i f i c i a l s eawa te r (Lyman and F m i q , 1940)- The v i t a m i n s were
added from a s t o c k s o l u t i o n t o g i v e c o n c e n t r a t i o n s o f 2 0 pg/l o f ,
f t h i amine , b i o t i n , p y r i d o x i n e , @ b o f l a v i n , pah to thdn ic a c i d , a s c o r b i c
' i % a c i d , i n o s i t o l , c h o l i n e , n i a c i n , p-aminobenzoic a c i d , and v i t amin
B-12. The t r a c e e l emen t s were added from a s t o c k s o l u t i o n t o g i v e a /---,
B c o n ~ e n t r a t i o n o f FeC13, 1400 p/l; CuS04, .ZnS04, Moo3, 10
pg/l; MnS04, 24 p g / l . The a r t i f i c i a l seawater was made u s i n g h
de ion ized water . The n u t r i e n t s o l u t i o n ( h e r e a f t e r
NSA) was a d j u s t e d t o pH 7.5 (Radiometer pH meter, model ,
22) by t h e a d d i t i o n o f IN H C 1 , f i l t e r . s t e r i l i z e d ( ~ i l l i p o r e GS, pore
s i z e 0 .22@, and added a s e p t i c a l l y toe s te r i le Erlenmeyer f l a s k s .
F i f t y m l of flSA were added t o each 250 m l f l a s k , except ,when o t h e r w i s e
s t a t e d .
C. Inoculum p r e p a r a t i o n t echn ique
Inoculum p r e p a r a t i o n techniques 'followed t h a t aP Sgurm, ,Meyers, -- - - -
, 5
4 and Simms (1962) . Inoculum was t a k e n from a seawater y e a s t e x t r a c t . 'i
4
s l a n t , p laced on t h e c e n t e r o f a p l a t e of F 1003 a g a r (Sguros et a l l -

t - -- - - - a
1962), and incuba ted a t 20 C f o r 3 weeks. A segment 0.5 mm i n
d iameter was then t a k e n from t h e edge o f t h e c o l o n y and p laced i n 125 P
25 m l o f NSA. The f l a s k s were t h e n incuba ted a t h 1
20 C f o r 43 days. Th i s d i f f e r e d from Sguros - e t a1 (1962) i n r p ~ ; " , e - r
of a chemica l ly de f ined medium f o r t h e stat% - c u l t ~ i r e phase. , The - d
inoculum was t h e n pmoved from t h e 125 m l f l a s k s , added t o 50 m l o f
a r t i f i c i a l seawatkr ( ASW) i n a S o r v a l l llOmnimixern, and macerated. t 4
The average d ry weight/ml of an inobulum sample was de terminee . , The
d r y weight o f t h e remaining my<elial suspension was then a d j u i t e d t o 4 *
mg/ml by d i l u t i o n w i t h ASW. One ml o f t h i s suspens ion was added t o 50 \ * 0
m l of NSA i n 250 m l f l a s k s , and incubated on a shake r r o t a t i n g a t 66 , 4
rpm a t room t empera tu re , 23 + 2C. Af t e r 7 days growth t h e mycelium
was aga in removed, added t o 5D m l Ah, and macerated f o r 20 seconds.
The myce l i a l suspens ion was then c e n t r i f u g e d a t 5,000 rw foi 30
minutes , washed, c e n t r i f u g e d a g a i n , rewashed, and t h e d r y weight o f
' t h e suspens ion a d j u s t e d t o 4 mg/ml. One ml of t h i s suspens ion was 0
used t o i n o c u l a t e each f l a s k i n t h e n u t r i t i o n a l exper iments .
, Inocu la t ed f l a s k s were incuba ted a t 23 C on a shake r r o t a t i n g a t 66 ---5 ,'*-a rpm f o r 7 and/or 14 days. Dry weight was determined by f i l t e r i n g t h e -
"elium through preweighed* g l a s s f i b r e f i l t e r s and washing thoroughly , -
wi th d i s t i l l e d wa te r . F i l t e r s were d r i e d f o r 24 hours i n a 60 C oven,
and t h e pH o f m e f i l t r a t e measured.
No, more than t h r e e consecu t ive t r a n s f e r s were made on l i q u i d
\ c u l t u r e medium. This r e s u l t e d from my obse rva t ion t h a t p e l l e t

--
morphology and/or p e n t a t i o n w of t r a n s f e r n i n l i q u i d c u l t u r e
l e s s a t 4 C.
i n Z . maritimum c h a n ~ e d a f t e r a s e r i e s
media. Inoculum was s t o r e d 4 days o r
Because of t h e q u a n t i t a t i v e v a r i a t i o n s be tween ' inocula t ions o f
- mycelia ( c f . Davidson, 1974) comparisons i n t h i s s tudy
wi th in experiments only. - - - --
r Throughout t h i s s tudy , and throughout o the r n u t r i t i o n a l and ,
physio logica l i n v e s t i g a t i o n s o f t h e f u n g i , t h e term " g r o d h w was \
f r e q u e n t l y used i n p lace 'of t h e term "dry weightw \ blthough dry b
weight I$ t h e most widely used and genera l ly appf icable , method o f
e s t i & t i n g growth (Mandels, 1965) ,. dry weight i s no t synonymous with
growth. When grown i n l i q u i d c u l t u r e Z. maritimum, H. a l o p a l l o n e l l a , * - -
and M. pe lagica formed from one t o s e v e r a l p e l l e t s which increased i n - volume. Whergas a d ry weight mzsurement represented t h e t o t a l
, '
p e l l e t , growth occurred only a t t h e hyphal t i p s . I n
o l d e r hyphae a dry weight measurement may have p a r t i a l l y r ep resen ted
t h e accumulation o f polysacchar ides , l i p i d s , o r wa l l m a t e r i a l s r a t h e r
than an i n c r e a s e i n l i v i n g protopham (Cochrane, 1958). Conversely,
p r o t e i n s y n t h e s i s may have continued a f t e r dry weight i n c r e a s e s had i
stopped (Cochrane, 1958). The term "growthw when used i n t h i s t h e s i s - -- -- - - - -- - - - - --
should be t r a n s l a t e d a s " the b e s t e s t ima te of growthw, ob ta inab le by -- - - - - -- - - - - -- -- - --
measuring t h e d r y , weight o f t h e bycelium.

The continuous c u l t u r e appara tus (Churchland and McClaren, 19-76)
was modified from ~ u b + t s ek (1970) and F r i e s 71956) ;- The s y i h
i l l u s t r a t e d i n Figs. 1-6: The 6L r e s e r v o i r f l a s k (B) flowed pt a , *
constant r a t e d e s p i t e c tanges i n t h e he igh t of t h e n u t r i e n t , s o l u t i o n . -
A s n u t r i h t 'a ~~ZTO?J ' e - e m i ~ - i r & f 1 ~ 9 k 3 t El air- w a s - & a ~ - - , .
i n t o A e r e s e r v o i r b o t t l e through a sterile. co t ton f i l t e r a t A . The - C
4 flow r a t e was c o n t r o l l e d by t h e h e i g h t o f t h e p ressure hgad ( t h e
-- - -
d i s t a n c e between t h e end of t h e a i r i n l e t tube and E) and t h e lengdh I . 6
V and-bore of t h e c a p i l l a r y tubing (D): A pressure head of 42 cm and i - 1
c a p i l l a r y tubing of 10 cm lePgth- and 0.2 mm bore provided a flow r a t e
of 4.0-5.0 ml/h. The c a p i l l a r y tubing was joined t o g l a s s tub ing 38 . - cm long and 5 mrn in . d iameter . TWO cm of high p u r i t y s u r g i c a l grade
l a t e x rubber tubing connected t h e g l a s s t u b i n g tcr t h e r e s e r v o i r f l a s k * L
a t C-. 3. 1 9.
li The 250 ml Erlenmeyer c u l t u r e f l a s k (F ) was stoppered wit;h.a i
co t ton plug through which g l a s s tubing was i n s e r t e d . The l i q u i d 4 %
medium dra ined through an overflow tube a t G , conta in ing a ground i g l a q s f i t t i n g &tached t o a s i n t e r e d g l a s s f i l t e r . The c u l t u r e f l a s k s 4
- 4 J - 'B were a g i t a t e d bn a shake; r o t a t i n g a t 66 rpm. Four r e s e r v o i r f l a s k s
supplying 16 c u l t u r e f l a s k s were used. Each c u l t u r e f l a s k overflowed - - - -
a t a volume o f approximately 75 m l ; JQ

Figure 1. Diagram of a continuous culture apparatus for the growth of
filamentous marine fungi. A, air inlet; B, reservoir flask,
C, rubber tubing; D, capillary tubing; E, glass tubing;
F, culture flask; G, exit tube with sintered-glass filter.


Figure 2. Photograph o f a continuous c u l t u r e appara tus
growth o f f i lamentous marine fungi .
f o r t h e


Flow r a t e s were monitored r e g u l a r l y by measuring t h i v o l u m F F T p -I
l i q u i d medium l e a v i n g the c u l t u r e f l a s k s . ( "
Ear ly des igns of t h e continuous c u l t u r e appara tus s u b s t i t u t e d
s u r g i c a l l a t e x rubber tubing f o r t h e g l a s s tub ing used i n t h e present
model. It was subsequent ly determined t h a t a l l types o f n a t u r a l and
/f s y n t h e t i c tubing t e s t e d were t o x i c t o one o r a l l o f - Z . maritimum,
H. a l o p a l l o n e l l a , and M . pe laqica . Lengths o f s u l f u r - f r e e , s u r g i c a l - - l a t e x , and s i l i c o n e rubber tubing were shaken i n n u t r i e n t s o l u t i o n f o r
2 weeks and removed; growth was assessed on t h e s e and c o n t r o l
s o l u t i o n s . The t o x i c ' p roper ty $bft i n t h e n u t r i e n t s o l u t i o n by these
tub ings i n h i b i t e d .ghwth. Subsequently, the amount o f tubing used was
reduced t o t h e minimum necessary f o r adequate f l e x i b i l i t y . Current ly ,
no more than 5 mm of tubing i s exposed t o t h e n u t r i e n t flow. ?
E. Comparativer growth i n cont inuous and batch c u l t u r e
The n u t r i e n t s o l u t i o n f o r t h e continuous c u l t u r e appara tus was
prepared a s fol lows. Five l i ters of a r t i f i c i a l seawater con ta in ing
7.2 g THAM were ad jus ted wi th 1N HC1 t o pH 7.0 and autoclaved; a l l
o the r NSA components were ad jus ted t o pH 7.5 and f i l t e r s t e r i l i z e d .
Autoclaving ve r sus f i l t e r s t e r i l i z a t i o n of a r t i f i c i a l seawater d id not - - - - -- - - -
a f f e c t growth of t h e t h r e e spec ies . Both s o l u t i o n s were then added - - - - &- - - -
a s e p t i c a l l y t o t h e autoclaved r e s e r v o i r f l a s k . The f i n a l pH of t h e
n u t r i e n t s o l u t i o n was approximately 7.7. The n u t r i e n t s i n t h e

r e s e r v o i r f l a s k s s tood f o r s e v e r a l days t o ensure l a c k o f
contamination. The c o n t e n t s o f t h e continuous c u l t u r e f l a s k s were
checked p e r i o d i c a l l y f o r contamination by inocu la t ion i n t o -22 1 6 ~
medium' (Zobel l , ' 1946). The n u t r i e n t soIut ion f o r t h e ba tch c u l t u r e
f l a s k s w prepared a s descr ibed i n (B) above. The batch c u l t u r e -
f l a s k s use i n t h e comparative growth curve experiments contained 75 -* t -
m l o f l i q h i d medium.
The continuous c u l t u r e f l a p k s were inocula ted w i t h 4 mg of <
inoculum 3 days before n u t r i e n t flow was begun, a l lowing a p e l l e t t o
be formed. For t h e comparative growbh curves , t h r e e f l a s k s fro'm batch , & )
j and continuous systems were removed every 4 days f o r dry weight -=-. . - r
. - determinat ion . Mycelium from each f l a s k was f i l t e r e d , weighed, and - sr
t h e pH of t h e f i l t r a te determined;
P. Expeiments w i t h reinoculat*edia
Zalerion maritimum, 8. a l o ~ a l l o n e l l a , and H. pe lan ica were grown
i n N S A f o r 2 weeks. The "oldn n u t r i e n t s o l u t i o n [was then f i l t e r e d and
divided i n t o two por t ions ; one por t ion was ad jus ted t o pH 7.5 wi th 1N
NaOH, t h e o the r por t ion was l e f t a t t h e low* value produced by t h e
then divided i n t o two p o r t i m s , and f r e s h ntzkriertts a d d e d t e - o n e -
por t ion o f each. 'Twenty-five m l of these four s o l u t i o n s , a r t i f i c i a l
seawater , and NSA were added t o 125 m l f l a s k s , inocu la ted wi th 2 mg
inoculum, incubated f o r 2 weeks, and t h e dF$ weight measured.

1 - -- -
1 G. Buf fe r experiments
f
The b a s a l n u t r i e n t s o l u t i o n (NSA) was made up wi th .005 M , . O 1 M ,
.02 M , .03 M , ,011 M , and .05 H THAM and MES (2-[N-morpholinol
e t h a n e s u l f o n i c a c i d ) r e s p e c t i v e l y . F l a s k s c o n t a i n i n g t h e s e b u f f e r s
were i n o c u l a t e d , d r y weight measured after 1 and 2 weeks, and t h e pH
o f $he f i l t r a t e measured.
H. Nut r ie ,I./' Uptake Experiments
Two i s o l a t e s o f 2. maritimum - were grown i n NSA c o n t a i n i n g 0.02 M i
MES i n s t e a d of THAM. T k p medium was analyzed eve ry 2 days f o r
g lucose , ammonium, phosphate , n i t r a t e , and n i t r i t e . S i x f l a s k s o f
each i s o l a t e were removed from t h e shake r ; t h r e e f l a s k s were used f o r
d r y weight and pH measurement. Three f l a s k s were f i l t e r e d and t h e
f i l t r a t e a n a l y z e d . f o r t h e abovementioned n u t r i e n t s .
Analyses of a l l n u t r i e n t s except ammonium were preceded by .a 9
II d i l u t i o n o f t h e medium w i t h seawater o r d i s t i l l e d water. To o b t a i n
c a l i b r a t i o n cu rves NSA was made up a t a range o f n u t r i e n t ,
c o n c e n t r a t i o n s and d i l u t e d aco rd ing ly .
- -- - - - - - - - --
4
After a 1:3 d i l u t i o n w i t h ASW, g lucose was measured us ing - --- - ---- -
o r t h o t o l u i d i n e (Hultman, 1959). The abso rp t ion o f t h e sa&les was
measured a t 610 nm w i t h a Bausch & Lomb Spec t ron ic 20

spec t rophotometer . Nitrate and n i t r i t e were measured acco rd ing t o
S t r i c k l a n d and Parsons (19x2) . The samples were a i i u t e d 1:200 w i t h
ASH, and a b s o r p t i o n S p e c t r o n i c 20 a t 543 nm. Phosphate 1
was determined u s i n g t h e t echn ique o f S t r i c k l a n d and Parsons (1972) . v
Samples were d i l u t e d 1:200 w i t h d i s t i l l e d and de ion ized water, and
a b s o r p t i o n measured w i t h a 10 CRI oel1 at 885 nrn on a Beckman DU - -
h spec t rophotometer .
Ammonium was measured usng a gas-sens ing ammonium e l e c t r o d e
(Orion Model 95-10) ,coupled t o a F i s h e r Accumet model 520 d i g i t a l I
pH/ion meter. After a l k a l i f i c a t i o n o f t h e n u t r i e n t s o l u t i o n , ammonium
i o n was conve r t ed t o ammonia gas which d i f f u s e d through t h e
d i f f e r e n t i a l l y permeable probe membrane.
2 The terms "morew o r "lessn, when used i n t h e t e x t t o modify
growth o r i o n up take , mean s i g n i f i c a n t l y more o r s i g n i f i c a n t l y less a t
t h e 95% p r o b a b i l i t y l e v e l . The stat istxcal test used- was e i t h e r t h e
s t u d e n t T - t e s t
dependent upon
me a n s .
o r t h e S tuden t Newman Keul m u l t i p l e range tes t ,
9 . whether comparisons were made between two o r s e v e r a l

RESULTS
A . Growth of two i s o l a t e s o f Zalerion
Figures 3 A and 3B show photographs of t h e two i s o l a t e s a f t e r
growth f o r 3'weeks a t 20 C on seawater agar . Notable d i f f e r e n c e s
were: t h e o l i v e pigmentation i n i s o l a t e FMC vs . grey ~ i g m e n t a t i o n i n
I s o l a t e H 135; and t h e more d i s t i n c t formation of r a d i a l zones by t h e
FMC i s o l a t e .
i
B. The continuous c u l t u r e system
The apparatus ( F k s . 1-2).was operated f o r 20 days o r longer n.
without contamination o r s i g n i f i c a n t change i n flow r a t e s . The
removable s i n t e r e d g l a s s e x i t tube did not c l o g wi th mycelium dur ing i \ t h e 21 day incubat ion per iod , and we e not replaced.
\ ' 4 C. Growth i n continuous and batch systems
The growth r a t e s o f each s p e c i e s (Fig . 4 ) were higher i n
continuous than batch c u l t u r e . Tn t h e case of Z. maritimum t h e r e was - e a f ive - fo ld d i f f e r e n c e i n d r y weight a f t e r 21 days. In ba tch c u l t u r e .
1
-
p ieces of m@elium which became separa ted from t h e main - - - -
-- - - - - - -
grow. I n the continuous system mycelium continued growing 04 t h e
/'
Z bottom and s i d e s o f t h e f l a s k s . Occasionally more than o n i l a r g e
i t

Figure 3. Growth af 2. maritimum H 135 and FMC on seawater agar. - _II


Figure 4 . Growth. of M. pelagica (A) , H. a lopa l lone l la (B) , and - - Z . maritimum (C) i n continuous and batch cu l tu r e . -
Cl - growth, batch cu l tu r e .-growth, continuous cu l tu r e
0 - p H , batch cu l tu r e @-pH, continuous cu l tu r e
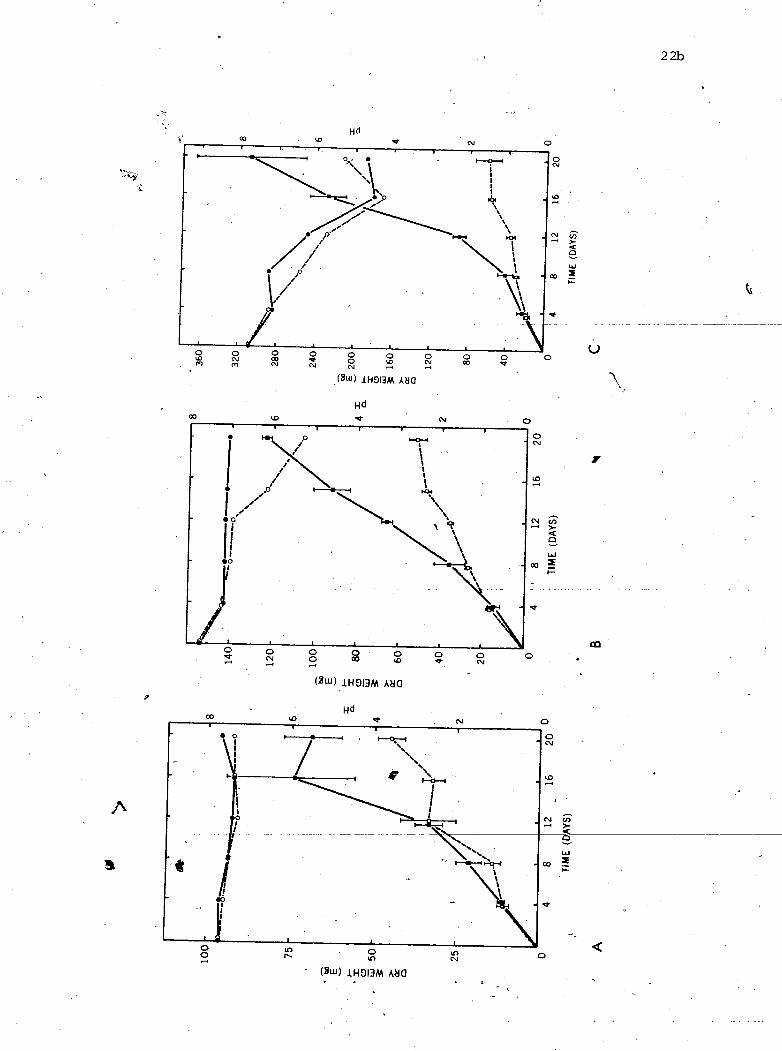

.-' - - - - - pp --
3 I-
p e l l e t would b e formed. I n 2. maritimum t h e appearance o f myoelium * $ d f
grown i n t h e two systems d i f f e r e d . After 8 day$ i n ba tch c u l t u r e , t h e 7
+ *
mycel ia l p e l l e t was dark o l i v e to -b lack and sporu la t ing heav i ly . -
After 8 days i n cont inuous c u l t u r e , t h e mycelial p e l l e t was l i g h t , 1 -7
'- '-
d i f f u s e , and produced very few spores . Af ter 21 days, t h e mycelium
near ly f i l l e d t h e cont inuous c u l t u r e f l a s k , and had darkened i n A - - - - - - - A 7
c e r t a i n arpas . f
When a mycel ia l p e l l e t of - Z . maritimum FMC was t r a n s f e r r e d t o a -
continuous f l a s k
halo o f a c t i v e l y
p e l l e t wi th in 48
e
a f t e r s e v e r a l weeks' incubat ion i n a c losed f l a s k , a
growing, l i g h t coloured mycelium developed around t h e
hours.
The n u t r i e n t s o l u t i o n was 'not
FMC was grown i n e i t h e r continuous
buffered by THAM when g. maritimum
o r ba tch c u l t u r e (Fig . 4). Growth
o f - H. a l o p a l l o n e l l a and - M. pe lagica i n continuous c u l t u r e was lower
than growth o f - Z .
o r sporu la t ion of
H. a l o p a l l o n e l l a , - whereas t h e pH of
of both s o l u t i o n s
maritimum. There was no d i f f e r e n c e i n pigmentation
the'
t h e
t h e
mycelium i n e i t h e r spec ies . During growth o f
pH of t h e ba tch c u l t u r e s o l u t i o n f e l l t o 5.6, 4 5 4
cont inuous c u l t u r e s o l u t i o n f e l l t o 7.0. The pH i 4
remained cons tan t dur ing growth o f - M. pelagica'.
d
D. ~ x ~ e r i m e n t $ with r e i n o u l a t e d media -
Y After 2 weeks' growth in NSA, 2. m a r i t i m u m FMC had lowered the pH ,-

- A - - -
2 4 - - --- - - - -
5 . I
of t h e n u t r i e n t s o l u t i o n t o 4.2. Growth was poor on reinoculat 'ed, low ,
pH media (Table I ) . More growth occurred on low pH m3dia wi th than
without added n u t r i e n t s . Growth on pH ad jus ted , r e inocu la ted media
was a s high a s
ad jus ted media
growing i n low
growing i n a l l I
Growth of
%
on NSA. ~ h d w a s a g r e a t e r pH drop after growth on pH L
with than without added n u t r i e n t s . The mycelium -
pH f l a s k s was be ige i n co lour , whereas t h e mycelium7 - *
-- --- --- - *' %..
--- -- o t h e r s o l u t i o n s was black.
without added
equa l , except
n u t r i e n t s .
n u t r i e n t s (Table I ) . Growth on a l l o t h e r s o l u t i o n s was t
f o r t h e h igher growth on p ~ * a d j u s t e d media wi th added +
I I
M. pe lag ica (Table I ) was poor i n r e inocu la ted , low pH - media (pH 5.5). Growth was s l i g h t l y increased by t h e a d d i t i o n of
n u t r i e n t s . Growth of - M . p e l a g i c a i n pH ad jus ted , r e inocu la ted ' media
B was g r e a t e r than i n NSA. There was a g r e a t e r pH drop in pH adjusGed
media with than without added n u t r i e n t s .
Growth o f - H. a l o p a l l o n e l l a was lowest bn low pH (5.43 medium
E. Buffer experiments
A t r - a ~ l - - c o n c e ~ ~ i o m ME%bufferehmore ef f ec t b e l y than THAH
growth on media con ta in ing TEIAM. Each compound buffered more
e f f e c t i v e l y a t h igher co rken t ra t ions . Th= higher concen t ra t ions of / .
each buf fe r d i d no t reduce growth. tk

L'
of - Z;
mak
itim
um
FMC
, k_.
p
ela
g"b
a and g
. a
lcs
pa
llo
n~
lla
in b
atc
h c
ult
ure
on
rein
oc
ula
ted
".
I ~ 2
. m
arit
imu
m
- -r
M.
pe
lag
ica
1 - H.
alo
pa
llo
ne
lla
I
So
luti
on
D
ry w
eig
ht
(mgj
~
in
al
p
H
Dry
wei
gh
t- (
mg)
F
ina
l p
&
~r
$
we
igh
t (m
g)
Fin
al pH.
I I I
5.9
*
8.2
,
7.8
. 3
.8'
8.1
i.
sea
wa
ter
~ ~
I -0.3
*#
1
I -0
.1
1 I
No pH
ad
jus
b
d 15.9
3.3
1
4.8
3.
X
l$. 9
3.4
len
r/ 1
-0
.5
ko
ad
ded
nu
kri
e t
s
. 1 t
0.6
1
-0.7
.
+
Y'
No
pH
ad ju
sb
en
2
.9
18
.2
' 3
.1
3.2
N
utr
ien
ts a
qd
ed
1 -0.4
I
- I
I I
,
NSA
i
pH
ad
jus
ted
to
7.5;
no
ad
d&
n
utr
ien
ts
~ I
pH
ad
jus
ted
to
7
.5;
nu
trie
ts
add
ed
l' 1
No
te:
~e
an
/i jo
+n
ed b
y
a c
omm
on
lin
e a
re n
ot
sig
nif
ica
ntl
y d
iffe
ren
t (~
ew
ma
k4
eu
ls te
st
t p
< 0
.05
) ~
* =
mea
n,
** =
st
an
da
rd e
rro
r sa
mp
le s
ize =
4
I I
I

Table 11.
Eff4ct of different concentrations of
THA
M and MES buffers on growth of Z. mardtimum FMC in
-
ba
tah
cylture.
I
*,
~ Incubated 7 days
Concentration
(hi)
I -
, Note : x =
mea
n
Dry Weight (mg)
Final pH
TRIS
- ms
- -
.
TRI S
- -
MES
- -
X
SX
X
SX
Incubated 14 days
: standard error, sample size =
3
*-
\
,t

F. . Nutr ient uptake experiments
i
When MEShwas s u b s t i t u t e d f o r THAMI, t he pH d id n o t f a l l below 6.0
dur ing growth o f Z . maritimum FMC (Fig . 5 ) . Af ter 2 weeks' - i ncuba t ion , 50% of t h e glucose remained i n t h e n u t r i e n t s o l u t i o n .
Ammonium bu t not n i t r a t e was removed r a p i d l y dur ing growth. - - -
Evaporation from t h e shake c u l t u r e f l a s k s was approximately 11% i n t h e
2 week incubation per iod , and may account f o r t h e apparent i n c r e a s e i n -
n i t r a t e concentra t ion . Phosphate was r a p i d l y removed and was p ~ e s e n t
i n t r a c e amounts a f t e r 8 days. A s i m i l a r p a t t e r n of n u t r i e n t uptake
was shown by I s o l a t e H 135. Af ter 14 days, 85Z.of t h e glucose $
remaihed i n t h e n u t r i e n t s o l u t i o n . Ammonium was r a p i d l y removed up t o n
bu t not beyond day 6. N i t r a t e was no t removed from t h e medium and
apparen t ly increased i n concen t ra t ion over t h e 14 day incuba t ion
per iod . A s i n t h e FMC i s o l a t e , t r a c e amounts of phosphate were . de tec ted a f t e r 8 days. During growth o f each i s o l a t e t h e medium was
analyzed f o r t h e presence o f n i t r i t e , and none was found.
DISCUSSION f
Many f i lamentous marine fung i can e a s i l y be grown i n axenic
mltur e -on seawater y e a s t extracty-TmmT T e a s t e x t r a c t i s f r e g u e X 1 y
i ne luded in - b ~ ~ ~ ~ ~ d -so%f Brnedb- asT-a-source- o f -phosphorus and -- --- - - +J
vi tamins (Sguros and Simms, 1963). The growth o f marine fung i i n
chemically defined media is poor i n comparison t o media con ta in ing
i Z

Figure 5. Changes i n dry weight, pH, and concentrations of glucose. -
phosphate, ammonium, and n i t r a t e during growth i n batch
, cul ture of Z. malitimum FMC and H 135. -

PO, mg PM
-? 9 I C\J T I
313 '6 asom16 PO, mg PI/

y e a s t e x t r a c t . Some i n v e s t i g a t o r s have s u b s t i t u t e d yeas t e x t r a c t ash
f o r yeas t e x t r a c t (Sguros and Simms, 1964; Sguros e t a1,1973). The
chemically defined n u t r i e n t s o l u t i o n used i n t h e s e experiments was
t h a t o f Sguros and Simms
minera ls was s u b s t i t u t e d
The moderate growth
H. a l o p a l l o n e l l a , and M. - -
(19631, i n which a mixture o f v i tamins a v
f o r yeas t e x t r a c t .
- -
r a t e s shown by L.
p l a ~ i c a on t h e chemically def ined medium
suggested a d i f f e r e n t system of c u l t i v a t i o n . In o rde r t o i n c r e a s e
growth r a t e s and approximate t h e cond i t ions o f t h e marine environment,
a continuous c u l t u r e system was designed f o r t h e c u l t i v a t i o n of marine
fungi .
I n the .con t inuous c u l t u r e system, n u t r i e n t s .flowed continuously
over a mycelial pelle: which was r e t a i n e d i n a c u l t u r e v e s s e l . 4
Zaler ion maritimum, H. a l o p a l l o n e l l a , and M. pe lagica were s u i t e d t o - - c u l t i v a t i o n i n t h i s appara tus because t h e mycelial fragments u s u a l l y
formed a s i n g l e submerged p e l l e t soon a f t e r inocu la t ion . The
continuous c u l t u r e system was no t a chemostat, i n which s t e a d y s t a t e
growth cond i t ions wi th in t h e c u l t u r e f l a s k a r e maintained by a
cons tan t e f f l u x of homogeneous mate r i a l . Although y e a s t s and some
growth shown by t h e s e g a r i n e s p e c i e s wouldmake s t ~ a d y state graklt3- - - - - -
cond i t ions d i f f i c u l t t o achieve.

c u l t u r e r e s u l t s . - However, i f a h ighe r biomass were r e q u i r e d ( i . e . ,
should t h e marine f u n g i produce compounds o f economic o r b iomedica l
impor t ance ) , a l a r g e r c u l t u r e v e s s e l could be des igned . iF
The f low r a t e o f aproximate ly 4.5 ml/h was chbsen t o minimize -
media a d d i t i o n s t o t h e r e s e r v o i r f l a s k s t o me p e r 14 day period.. In - - t ' a d d i t i o n t o t h e inconvenience o f making l a r g e volumes o f media,
rl f r e q u e n t opening o f t h e r e s e r v o i r f l a s k i n c r e a s e d t h e r i s k of
contamina t ion . The f low r a t e c o u l d e a s i l y be l e s sened o r i n c r e a s e d by 1
r e s p e c t i v e l y l eng then ing o r s h o r t e n i n g t h e c a p i l l a r y t ub ing .
It was imposs ib le t o ana lyze t h e growth cu rves ob t a ined i n e i t h e r
b a t c h o r con t inuous c u l t u r e acco rd ing t o *any g iven mathemat ica l -4
f u n c t i o n . Terms borrowed from b a c t e r i a l growth k i n e t i c s do n o t $
n e c e s s a r i l y d e s c r i b e f u n g a l growth. Because growth o f a f i l a m e n t o u s
fungus is n o t ' i u t o c a t a l y t i c , t h e "exponent ia l growth phasen o f t h e
b a c t e r i a l growth cu rve i s b e s t termed t h e "phase o f r a p i d growthn b
(Mandels, 1965) o r t h e " l i n e a r growth phasen i n t h e f u n g a l growth \
c u r v e . Mandels (1965) h a s sugges t ed t h a t a be r o o t f u n c t i o n + 1 d e s c r i b e s mathemat ica l ly t h e growth o f a f u n g a j p e l l e t i n shake
c u l t u r e . However, t h e p e l l e t s o f Z . maritimum, H. a l o p a l l o n e l l a , and - - M. p e l a g i c a were t o o v a r i a b l e f o r t h e u se o f a c u b i c a l f u n c t i o n . -
- - -- - -- - - - - - - --
Moreover, p i e c e s o f mycelium c o u l d s e p a r a t e from t h e main p e l l e t and
a t t a c h t o t h e s i d e s o r bottom o f t h e c u l t u r e v e s s e l , forming new
c e n t e r s f o r growth.

I n cont inuous
i n c u b a t i o n pe r iod .
con t inuous c u l t u r e
because t h e fungus
c u l t u r e , growth cont inued throughout t h e 21 day
The growth ra te o f each s p e c i e s d e c l i n e d i n
after day 16, p o s s i b l y because of pH changes o r a
had expanded t o f i l l t h e c u l t u r e v e s s e l . I n b a t c h
c u l t u r e , t h e d u r a t i o n o f l i n e a r growth was dependent upon c u l t u r a l
c o n d i t i o n s . I n maritimum, t h e moat r a p i d growth was g e n e r a l l y
between 4 and 8 days (see growth cu rves i n Chapter I I ) , fo l lowed by a
pe r iod o f f l u c t u a t i o n i n d r y weight . Sguros e t a 1 (1962) p a r t i a l l y
a t t r i b u t e d d r y weight f l u c t u a t i o n i n ~ r i c h o c l a d i u m achrasporum (Meyers*
and Moore) Dixon t o a u t o l y s i s , and s t a t e d t h a t d r y we igh t s beyond 8
days probably r e f l e c t e d t h e p re sence o f dead and p a r t i a l l y a u t o l y s e d
c e l l s . I n L. maritimum l i n e a r growth cou ld con t inue from 10 t o -12
days o r l o n g e r . I n most exper iments , growth measurements i n b a t c h
c u l t u r e were t a k e n after 7 and 14 days. The 7 day measurements should
r e p r e s e n t t h e d r y we igh t o f a c t i v e l y growing mycelium, whereas t h e 14
day measurements ma p a r t i a l l y r e p r e s e n t t h e accumulat ion o f Y po lysaccha r ides and o t h e r s t o r a g e p roduc t s i n t h e a l d e r hyphae. I n
t h e con t inuous sys tem, a l l exper iments were h a r v e s t e d after 14 days ,
d u r i n g t h e per iod o f most r a p i d growth o f each s p e c i e s t e s t e d . 1
The use o f a c o n t i n u o u s c u l t u r e system showed a r e l a t i o n s h i p ppLpp -- -- -
d
between p igment p roduc t ion , s p o r u l a t i o n , and c e s s a t i o n o f growth i n - - -- pp - -pp -
2. A pH changq i n t h e n u t r i e n t s o l u t i o n d i d n o t e x p l a i n
t h e o n s e t o f s p o r u l a t i o n ; mycelium i n
produced few spo re s . Cochrane ( 1958 )
1
+ , r e i n o c u l a t e d , low pH media
s t a t e d t h a t t h e major

s t i m u l a t i o n t o r e p r o d u c t i o n i n t h e f u n g i was n u t r i e n t exhaus t ion .
Perhaps af ter s e v e r a l days i n b a t c h c u l t u r e and 16 days i n con t inuous
c u l t u r e s p o r u l a t i o n was s t i m u l a t e d by a l a c k o f one o r more n u t r i e n t s .
I
Experiments were, c a r r i e d o u t t o exp la in t h e d i f f e r e n t growth
r a t e s i n con t inuous and ba t ch c u l t u r e . P e l l e t s which-had stopped -
growing i n a c l o s e d f l a s k grew a g a i n when t r a n s f e r r e d t o t h e
con t inuous c u l t u r e system. Thus growth d id n o t slow down i n b a t c h
c u l t u r e due t o a d e t e r m i n a t e s i z e f a c t o r , a s may occu r i n c e r t a i n \ o t h e r f u n g i ( G o t t l i e b , 1971) . T h i s r e s u l t a l s o showed t h a t growth was
no t i n h i b i t e d by an i n t r a m y c e l i a l s t a l i n g f a c t o r , which c o u l d be
t r a n s p o r t e d from t h e o l d e r hyphae t o t h e pe r iphe ry o f t h e . c o l o n y
( M o l i t o r i s , 1974). Experiments w i th r e i n o c u l a t e d media demonstrated
t h a t t h e r e were no non v o l a t i l e s t a l i n g p roduc t s p r e s e n t in t h e
n u t r i e n t s o l u t i o n . Only i n g. a l o p a l l o n e l l a d i d n u t r i e n t exhaus t ion
appea r t o be a growth ' i n h i b i t i n g f a c t o r .
4
A pH effect may e x p l a i n t h e poorer growth shown by Z. maritimum, - H. a l o p a l l o n e l l a , and & p e l a ~ i c a i n ba t ch c u l t u r e . When ammonium is
used a s a n i t r o g e n s o u r c e , t h e up take o f t h i s i o n and t h e accompanying
exchange o f hydrogen i o n s from t h e c e l l g e n e r a l l y results i n a pH drop - - -- - -- - - - - - - -
i n t h e n u t r i e n t s o l u t i o n (Sguros and:Simms, 1963) . - -- - --- -
t h e s e t h r e e s p e c i e s i n a c i d i c media was improved by
n u t r i e n t s , which probably r a i s e d t h e pH o r improved
The poor growth o f - ~ - ~-
t h e a d d i t i o n o f
t h e b u f f e r i n g
c a p a c i t y o f t h e media.

L - . - -
Growth o f 2. ar i t imum and M. p e l a q i c a caused a g r e a t e r pH drop
in r e i n o c u l a t e d , pH g d j u s t e d media wi th added n u t r i e n t s , t h a n i n pH
a d j u s t e d media wi thout added n u t r i e n t s . There was no growth
d i f f e r e n c e t o cause t h i s r e s u l t . A p o s s i b l e e x p l a n a t i o n i s t h a t i n
t h e s o l u t i o n wi thou t added n u t r i e n t s , t h e ammonium was l a r g e l y
u t i l i z e d d u r i n g t h e f irst growth pe r iod . When t h e medium was * r e i n o c u l a t e d , t h e fungus used n i t r a t e i n a d d i t i o n t o ammoniwn. The pH
t h e r e f o r e d i d n o t f a l l as d r a s t i c a l l y a s i n t h e media which had
r ece ived a f r e s h supp ly o f NH4N03. A pH drop i n a Q u t r i e n t
s o l u t i o n cont%ininq N H 4 N 0 3 i s g e n e r a l l y cons ide red presumptive
ev idence f o r t h e p r e f e r e n t i a l u t i l i z a t i o n o f ammonium (Nicho la s ,
It is c l e a r t h a t THAM d i d n o t b u f f e r t h e n u t r i e n t s o l u t i o n
e f f e c t i v e l y . F o r t h i s r e a s o n MES, de sc r ibed as a b i o l o g i c a l b u f f e r by
Good e t a1 (1966) and used by Ch i ld , Knapp, and Evele igh (1973) t o C
b u f f e r agar media ' du r ing t h e growth o f s e v e r a l funga l s p e c i e s , was
s u b s t i t u t e d f o r THAM i n c e r t a i n experiments . Growth was s l i g h t l y
lower on MES t h a n on THAM; however, t h e former uompound bu f f e r ed more
e f f e c t i v e l y a t a l l c o n c e n t r a t i o n s t e s t e d .
limit growth i n b a t c h c u l t u r e i f &he pH_ o f t h e n u t r i e n L ~ s o l u t i ~ ~ ~ w _ e r _ e - -
c o n t r o l l e d . Glucose was n o t exhaus ted , and a l t hough a large p a r t ( H
135) o r a l l (FMC) of t h e ammonium was removed, a l a r g e supp ly o f ,

n i p a t e remained i n t h e medium. S t u d i e s wi th S c o p u l a r i o p s i s -. 4
b r e v i c a u l i s (Sacc . ) B a i n i e r (Morton and MacMillan, 1954) i n d i c a t e t h a t
af'ter exhaus t ion o f ammonium, n i t r a t e can be used. Both i s o l a t e s had
removed a l l o f t h e phosphate a f t e r 8 days growth, a t which time t h e r e
. was a marked d e c r e a s e i n t h e growth rate. A similar test w i t h
H. a l o p a l l o n e l l a and M . p e l a g i c a showed t h a t t hey a136 removed t h e - - major p o r t i o n o f t h e phosphate from t h e n u t r i e n t s o l u t i o n . '
Growth d i d n o t s t o p in b a t c h c u l t u r e s imply because o f phosphorus
l i m i t a t i o n . I n c r e a s i n g t h e amount o f phosphorus i n t h e medium d i d n o t
markedly i n c r e a s e growth o f e i t h e r i s o l a t e . P o s s i b l y phosphate was
removed from t h e n u t r i e n t s o l u t i o n and s t o r e d i n an i n t e r n a l p o o l , as
is t h e c a s e wi th amino a c i d s (N icho la s , 1965) and ammonia (Hunter and
S e g e l , 1971). S i e g e n t h a l e r e t a1 (1967) demonstrated a n i n t r a c e l l u l a r
phosphate pool i n t h e marine Phycomycete Thrauatochytr ium roseum
Golds t e in . If t h i s were t h e c a s e i n Z. maritimum, phosphorus would
n o t n e c e s s a r i l y l i m i t growth d e s p i t e i t s absence from t h e n u t r i e n t
s o l u t i o n . I n conc lus ion , t h e lower growth i n b a t c h e u l t u r e was due t o
changes i n pH o f the n u t r i e n t s o l u t i o n . When t h e pH was a o n t r o l l e d ,
t h e r e s u l t s s u g g e s t t h a t l a c k o f phosphorus, p o s s i b l y i n combinat ion
w i t h o t h e r n u t r i e n t s , l i m i t e d growth i n ba t ch c u l t u r e .
S ince marine f u n g i i n n a t u r e do n o t affectT the-pH or-chemicaL - - - -
composi t ion o f t h e su r round ing n u t r i e n t s o l u t i o n , t h e con t inuous
system may more c l o s e l y approximate t h e marine environment t h a n t h e

--
ba tch system. The c o n t i n u o u s system may a l l ow t h e u se o f d i l h t e ,
media, t h u s more a c c u r a t e l y r e c r e a t i n g marine n u t r i e n t , concen t r a t i ons .
Despi te t h e s e advantages , t h e con t inuous system shoul'd n o t r e p l a c e t h e
ba t ch system i n a l l f u t u r e n u t r i t i o n a l s t u d i e s . Problems of making '$
large volumes o f media, l i m i t e d sample s i z e , and s i z e o f a p p a r a t u s
make t h i s l o g i s t i c a l l y imposs ib l e . A d d i t i o n a l l y , growth o f each \
m y c e l i a l fragment i n o o n t i n u o u s c u l t u r e r e s u l t s i n I a r g e - d r y weimt'
v a r i a t i o n s . I sugges t t h a t t h e con t inuous system be used i n a d d i t i o n
t o t h e b a t c h system, when i n t e r f e r e ~ c e i n growth from n u t r i e n t
exhaus t ion , pH changes , o r s t a l i n g p roduc t s i s suspec t ed .
I n summary, t h e moderate growth o f Z . maritimum, - H. a l o p a l l o n e l l ~ , and M. p e l a g i c a i n chemica l ly d e f i n e d media - - sugges ted t h e u s e o f a con t inuous c u l t u r e system. The s i g n i f i c a n t l y
h ighe r growth o f a l l s p e c i e s i n t h e cont inuous system l e d t o an
i n v e s t i g a t i o n o f growth dynamics i n t h e ba t ch system. Analyses o f
n u t r i e n t s i n t h e c u l t u r e medium and experiments w i t h r e i n o c u l a t e d
media sugges ted t h a t pH changes and d e p l e t i o n o f phosphorus and /o r B
o t h e r n u t r i e n t s l i m i t e d growth i n b a t c h c u l t u r e . These r e s u l t s may
a l l o w , t h e fopuu la t i on o f more ba lanced c u l t u r e media f o r f u t u r e
n u t r i t i o n a l s t u d i e s .
- - -
The in fo rma t ion on c u l t d v a t i o n and growth which is p re sen t ed i n - - - - -- -- - - - - --- -- -- -- - - - - - -
Chapter I p rov ides t h e neces sa ry background f o r an unde r s t and ing o f
Chapter 11, a s t u d y of n i t r o g e n n u t r i t i o n i n Za l e r ion maritimum.

CHAPTER I1
Nitrogen N u t r i t i o n i n Zalerion maritimum
\
INTRODUCTION .
- - -
Various s t u d i e s have been c a r r i e d out on t h e n i t rogen
. requirements of t e r r e s t r i a l fung i . Many spec ies (Sarbhoy, 1965; Said
and Harhash, 1966; Agnihotr i and Vaar ta ja , 1967; Mitra and Tandon,
1970) use both inoeganic and organic n i t rogen. Some f u n g i (Grover and
~ i d h u , 1965; Jones, 1965; Yusef and Rllam, 1967; Stephen and Chan,
1970) grow bes t on o rgan ic n i t rogen. Certan Phycomycetes cannot
metabolize e i t h e r n i t r a t e o r n i t r i t e (Bhargava, 1945; Dayal, 1961.;
Barr, 1969, Nolan, 1970). Most fung i , when suppl ied wi th ammonium and
n i t r a t e , w i l l p r e f e r e n t i a l l y u t i l i z e t h e former (Morton and MacMillan,
1954; Nicholas, 1965; Cove and Pateman, 1969). Ammonium and n i t r i t e
may b e taken up simultaneously (Morton and MacMillan, 1954). The
optimal n i t rogen source and carbon t o n i t rogen r a t i o is n o t
n e c e s s a r i l y t h e same f o r growth and f o r sporu la t ion (Yusef and Allam,
1967; Aube,and Gagnon, 1969; Singh and Tandon, 1970).
The - - - - fungi - - - 'do-not - p lay t h e same rooleasthe_bcLeria2ILkhe
t e r r e s t r i a l n i t rogen cyc le . There is no evidence that-the f u n g i - - - - -- - -- -- -- - - ---
, e i t h e r f i x atmospheric n i t rogen (Benemann and Valent ine , 1972) o r
d e n i t r i f y inorganic compounds t o n i t rogen gas (Nicholas, 1965). Eylar

- - --- -
and Schmidt (1959) and Hatcher and Schmidt (1971) have shown t h a t s o i l
fung i such a s Aspergi l lus f l a v u s Link e x F r . convert o rgan ic n i t rogen
compounds t o n i t r a t e . The main r o l e of the fung i i n t h e t e r r e s t r i a l
n i t rogen cycle is t o break down organ ic molecules, and conver t thein t o -- .- -
s impler and more u t i l i z a b l e chemical forms.
- -- - -pip -- -
L i t t l e work has been done on t h e n i t rogen requirements o f marine
fungi . Gustafsson and Fries (1956) s tud ied growth o f Pleospora
purpurascens R. Sant . Zaler ion maritimum and Diplodia orae- mar is
Linder on aspa r / i n e , (NH4)2HP04, and KN03 Johnson e t a 1
(1959) s tud ied r a d i a l growth o f s e v e r a l higher marine fung i on aga r .
made up with v a r i o u s n i t r o g e n sources and a t va r ious carbon t o
n i t r o g e n r a t i o s . Sguros and Simms (1963) s tud ied t h e e f f e c t o f
v a r i o u s concen t ra t ions of carbon and n i t rogen on growth o f c
Trichocladium achrasporum, Humicola a l o p a l l o n e l l a , Orbimvces
s p e c t a b i l i s Linder, and H a l o s ~ h a e r i a m e d i o a e t b ~ e r a (Cribb & Cribb).
Holl igan and-Jennings (1972b) i n v e s t i g a t e d t h e e f f e c t o f n i t rogen
sources on carbohydrate metabolism i n D e n d r v ~ h i e l l a s a l i n a (Suth.)
Pugh and Hicot. Sguros st a1 (1973) c a r r i e d o u t t h e most
comprehensive s tudy t o d a t e o f n i t rogen n u t r i t i o n i n t h e f i lamentous
marine fungi . They grew Ijalosphaeria m e d i o s e t i ~ e r a , Hwnicola
a c i d s , amines, and n u c l e i c a c i d s . Each was supp l i ed s i n g l y as t h e
n i t r o g e n source and growth measured by d r y weight determination.

concen t ra t ions of NH4N03 on growth o f these t h r e e spec ies . P e r r s b
et. - a1 (1975) determined t h e mycelial f r e e amin9 a c i d cod ten t o f t e n . s p e c i e s o f h igher marine -fungi . ' I
S tud ies c on , the n i t r o g e d requirements of marine Phycomycetes were . -
c a r r i e d b w t - by. Vishniac L 19551, _Go_ld_ste_in ( 1963 a Lb,cL)l Gpldstein and 1
' Belsky ( 1 9 6 4 ) ~ and Alderman and Jones (1971). 0
A 1 1 o f the abovementioned s t u d i e s were c a r r i e d out ~ e i t h s r on
s o l i d aga r 'or i n l i q u i d , ba tch c u l t u r e . In t h i s s tudy a continuous
c u l t u r e s y s t e m was used i n a d d i t i o n t o t h e ba tch . ' cu l tu re technique.
It was hoped that a cont inuous c u l t u r e , system - would more c l o s e l y
aproximate t h e c o n d i t i o n s o f t h e marine knvi'ronment . Addi t iona l ly , it
- was hoped that 'a comparison between t h e two systems would show whether \ .
c u l t i v a t i o n techniques could a f f e c t n u t r i t i o n a l r e s u l t s . '
Previous inves t ig&ors have 'supplied n i t r o g e n sources s i n g l y i n ,
t h e growth medium. In t h e i r n a t u r a l environment marine f fungi are
s u p p l i e d wi th v a r i o u s n i t rogen sources , a b i l a b l e f r b m b o t i t h e - c e l l ~ l ~ ~ i ~ s u b s t r a t e and t h e surrounding seawater. 1x1 t h i s s tudy t h e . .
r e l a t i v e removal from t h e n u t r i e n t s o l u t i o n o f copb ina t ions o f
n u t r i e n t s w a s determined by chearical analyses . '

A l l o f t h e nutritional e x p e r i m e n t s i n this c r a p t e r were c a r r i e d
out with Zaler ion m a k t i m u m , chosen because of i t s common occurrence a.
i n temperate marine wa te r s and i ts p o s s i b l e importance .as a wood i
1 decomposer (Jones , 1957-58). Because of t h e phys io log ica l v a r i a b i l i t y . - of t h i s fungus, two i s p l a t e s were used f o r t h e mhjor i ty of t h e
experiments. - - - - - - - - - A - --
/
The g o a l s o f t h e experiments descr ibed i n t h i s chap te r were as -
fol lows: - t o determine t h e e f f e c t of growing inoculum on va r ious , -
n i t rogen sources oh subsequent f i a t t e rns o f n i t rogen n u t r i t i o n ; t o , -
determine what inorgan ic and o rgan ic n i t rogen sources were u t i l i z e d ,
and t o compare t h e s e r e s u l t s with t h e a v a i l a b i l i t y of n i t rogen i n t h e n
sea ; t o determine which, i f any, n i t rogen sources were p r e f e r e n t i a l l y
removed from t h e n u t r i e n t s o l u t i o n ; ( t o determine how much n i t rogen was > --
necessary f o r growth, and how changes i n C:N r a t i o affected growth. 4 - r 3,
Addi t ional ly , an a t tempt was made t o determine whether &. maritimum
could c a r r y ou t any o f t h e conversions o f t h e marine n i t r o g e n cycle . -
I
MATERIALS AND METHODS 4 &
Techniques o f inoculum prepara t ion , methods o f ou l t i - t ion , and
u n l e s s otherwise s p e c i f i e d . A l l n i t rogen sources were A p p l i e d a t a - - - - pp - - - - - - p- - -- -- - - - - -- - --
.concentraton of 100 mg N / 1 u n l e s s otherwise s t a t e d . Sample s i z e s f o r
each experiment were i n d i c a t e d on t h e corresponding f i g u r e s and
graphs.

- - - - - -
..A. Growth o f inoculum on v a r i o u s n i t rogen s o u r c e s
Por t ions o f mycelium 5 mm i n diameter o f z. maritimum
removed from F 1003 a g a r p l a t e s and placed i n NSA. N H 4 W 3
FMC were '
was
replaced by NH4C1, NaNO N a N 0 3 , o r I.-alanine. K + t e r 43 days 2 ' growth t h e four ba tches o f mycelium were macerated, q u a n t i f i e d , and
- - - -- t h e inoculum a d j u s t e d t o 4 m g / m l . Ten f l a s k s c o n t a i n i n g each Bf,*heL - - - - - - - / ,
four n i t rogen sources were inocu la ted with myceLium grown on t h e
corresponding n i t r o g e n source. After 7 days i n shake c u l t u r e , t h e
mycelium was macerated, centr$fuged, and washed. Inoculum grown on
N H 4 C l was used t o i n o c u l a t e 10 f l a s k s containing-NH4C1, 10 f l a s k s
con ta in ing NaN02, 10 f l a s k s con ta in ing NaN03, and 10 f l a s k s Ll
conta in ing 1-alanine. After 7 and 14 days i n shake c u l t u r e , t h e
mycelium was weighed and t h e pH of t h e n u t r i e n t s o l u t i o n measured.
Inoculum grown-on H a N 0 2 , NaN03, and I-alanine was t r e a t e d i n t h e f 7
same manner.
B. Growth on inorgan ic n i t rogen sources
Growbh o f two i s o l a t e s o f Zaler ion maritimum on va r ious n i t rogen /
sources was compared i n cont inuous and batch c u l t u r e . The NH4N03
i n NSA was s u b s t i t u t e d by NaN03, NaN02, NH4C1, o r 1-glutamic - - - - - - - --
ac id . Each of t h e - f o u r r e s e r v o i r f l a s k s o f t h e continuous system - - - -- p p -- -- - -- - - pp -
- -- -
contained one o f the. n i t rogen sources . The c u l t u r e f l a s k s were ,
i nocu la ted 3 days be fo re n u t r i e n t flow was begun. The c u l t u r e f l a s k s

from t h e continuous system w9re removed f o r d ry
days a f t e r i n o c u l a t i o n . The pH of the e f f l u e n t
system was measured every 2 days. Mycelial dry
batch c u l t u r e f l a s k s was measured a f t e r 1 and 2
from t h e continuous
weight and pH i n t h e
weeks' growth.
The e f f e c t o f d i f f e r e n t n i t rogen concen t ra t ions on growth o f H.
maritimum i n batch and continuous c u l t u r e was t e s t e d . NH4N03-in
NSA was s u b s t i t u t e d by 100 mg N / 1 , 10 mg N / 1 , 1 mg N / 1 , and 0.1 mg N / 1 I
of NaN03. Each of t h e r e s e r v o i r f l a s k s contained one o f t h e s e
s o l u t i o n s , and a f i f t h r e s e r v o i r f l a s k contained no n i t rogen . The
c u l t u r e f l a s k s were inocu la ted 3 days before n u t r i e n t fl6w was begun,
and d r y weightdneasured 14 days a f t e r incubat ion . The pH of t h e
e f f l u e n t was measured every 2 days. The dry weight and pH of t h e
batch c u l t u r e f l a s k s was measured a f t e r 7 -and 1-4- days.
The e f f e c t o f va r ious carbon and n i t rogen concen t ra t ions on
growth i n continuous and batch c u l t u r e was t e s t e d . NSA was prepared
with t h e fo l lowing c o n c e n t r a t i i o n s o f glucose and NaN03: glucose
3.6 g C / 1 , NaN03 100 mg N / 1 ; g lucose 0.36 g C / 1 , NaN03 10 mg N / 1 ;
g lucose 0.036 g C I 1 , NaN03 1 mg N / 1 ; g lucose 0.0036 g C / 1 , NaN03 a
0.1 mg/l. The experiment was c a r r i e d out i n ba tch and continuous

-
C. Growth on o rgan ic n i t rogen sources .
1. S i n g l e amino a c i d s J
Growth of - Z . maritimum FMC and H 135 was tested i n ba tch c u l t u r e
i n NSA t o which 21 amino a c i d s and NaNO were added s i n g l y as sole nitrogen 3
souraes. Two mg of inoculum were added t o 125 m l f l a s k s o c o n t a i n i n g 25
m l o f l i q u i d medium. After 7 days i n shake c u l t u r e , t h e mycelium was
weighed and t h e f i n a l pH of t h e n u t r i e n t s o l u t i o n measured.
2. A combination o f amino a c i d s
L-arginine-HC1, 1-a lanine , and 1-glutamic a c i d were added s i n g l y
and i n combination t o NSA i n continuous and batch c u l t u r e . The s i n g l e
amino a c i d s were
amino a c i d s were
N / 1 . Dry weight
added a t a concentra t ion of 100 mg N / 1 . The t h r e e b
added e q u a l l y t o g ive a t o t a l concen t ra t ion o f 100 mg
was measured a f t e r 14 days i n cont inuous c u l t u r e , and
t h e pH of the e f f l u e n t was measured every 2 days. Dry weight and pH
of t h e n u t r i e n t s o l u t i o n was measured i n batch c u l t u r e a f t e r 7 and 14
days.
3 . Organic compounds o the r than amino aaids
Both i s o l a t e s
peptone, &anbic
e-itfulLizucas s o l e
comparison wi th an
were grown i n batoh c u l t u r e on NSA con ta in ing u rea ,
m i d , zil2anboi.a ,vFtamiUraecasarnino a c i d s , and s
s o u r c e s nf n i t - r o g e ~ . N i t r a t e was - included - for -- - - - -- - - -
i no rgan ic n i t rogen source. Bacto-peptone and
vi tamin-free casamino a c i d s were added a t concen t ra t ions of 0.6173 g/ l

and 1.4285 g / l r e s p e c t i v e l y . These amounts &orresponded t o . -
appraximate ly 100 mg N / 1 (Di fco manual ) . Dry weight of mycelium and
pH of t h e n u t r i e n t s o l u t i o n was measured a f t e r 7 and 14 days
incuba t ion .
D. Removal of n i t r o g e n sou rce combinat ions from t h e n u t r i e n t
s o l u t i o n
1. Ammonium and n i t r i t e
I s o l a t e s FMC and H 135 were grown i n N S A c o n t a i n i n g 50 mg N / 1
each o f NaM02 and NH4C1. I n t h i s experiment , two times t h e usua l
phosphate c o n c e n t r a t i o n o f .06 g / l was added. MES was s u b s t i t u t e d f o r
TRIS a t a concen t r a ton o f .02 M. A t 2 day i n t e r v a l s , three f l a s k s
were removed f o r d r y weight and pH measurements. Three a d d i t i o n a l
flasks were removed f o r de t e rmina t ion o f g lucose , ammonium, n i t r i t e ,
r n i t r a t e , and p ospha te . The t echn iques f o r t h e a n a l y s e s o f t h e s e i o n s
were desc r ibed i n Chapter I.
B
2. L-glutamic a c i d and n i t r a t e
Both i s o l a t e s were grown i n NSA c o n t a i n i p g 50 mg N / 1 .each of
1-glutamic a c i d and NaNO . PiES was s u b s t i t u t e d f o r TRIS a t a -- -- - -3
c o n c e n t r a t i o n of .02 M. Every 2 days t h r e e f l a s k s were removed f o r - -
-- - pp - -- -- - -
d r y weight and pH measurements. Three a d d i t i o n a l f l a s k s were removed
f o r measurement o f n i t r a t e and g lu tamic a c i d . Nitrate was measured
accord ing t o S t r i c k l a n d and Parsons (1972). To measure g lu t amic a c i d ,

-- - - -- - - -- - ---
t h e n u t r i e n t s o l u t i o n was d i l u t e d 2: 1 wi th ASW. One m l of ninhydrin
s o l u t i o n (100 m l 4 N sodium a c e t a t e b u f f e r , 300 m l f i l t e r e d methyl
c e l l o s o l v e , 8 g ninhydr in , 0.16 g SnC12) was added t o 5 m l of d i l u t e J
n u t r i e n t s o l u t i o n , mixkd on a vor tex mixer, and boi led f o r 30 seconhs
( S h r i n e r , Fuson, and Cur t in , 1964). After coo l ing , t h e absorbance of
t h e samples'was measured a g a i n s t an ASW blank on a Bausch and Lomb -& - A - -
Spectronic-20 'spectrophotometer a t 570 rim. The r e s u l t s were compared
wi th a c a l i b r a t i o n curve obta ined by adding ninhydrin t o va r ious
concen t ra t ions o f g lu tamic a c i d i n NSA. A c o n t r o l of uninoculated NSA
j wa-s included i n each a n a l y s i s .
3. Ammonium and e i g h t amino a c i d s
I s o l a t e s FMC and H 135 were grown, i n batch c u l t u r e , i n NSA which
contained NH4C1 and ' e igh t amino'acids (1-qspart ic a c i d , 1- threonine ,
1 - se r ine , 1-glutamic a c i d , 1-a lanine , 1-val ine, 1 - i so leuc ine , and
1- leucine) . TRIs was replaced by .02 M PIES. The e i g h t amino a c i d s
were each added a t .05 mM/1, Equimolar concen t ra t ions of t h e amino
a c i d s were used i n o r d e r t o o b t a i n peaks o f a s i m i l a r he igh t from t h e
amino a c i d ana lyze r , For each amino a c i d , .05 m M / 1 corresponded t o
approximately 14 mg N / 1 . Nine f l a s k s were removed from t h e shaker
a f t e r 3 ,6 ,9 , and 12 days. Three f l a s k s were used f o r d r y weight and S
pH measurements. The c o n t e n t s o f t h r e e flasks were f i l t e r e d -
'J Y
(Mil l ipore HA, 0 . 4 5 ~ 1 ) and ammonium measured i n t h e f i l t rate . - - - - - - - -- - -- - - -- -
Ammonium was measured according t o S t r i ck land and Parsons (1972).
Absorbance was measured on a Beckman DU spectrophotometer a t 640 nm.

- - - - - - -- - - -
Each rnample w a s diluted 1 :2QO with d b t i l h i ~ e s u l t a
compared wi th a c a l i b r a t i o n curve . The c a l i b r a t i o n c u r v e was o b t a i d by p r e p a r i n g a range of c o n c e n t r a t i o n s o f N H 4 C 1 i n NSA, fol lowed by
r a 1:200 d i l u t i o n wi th d i s t i l l e d water .
The f i n a l t h r e e f l a s k s were removed every 3 days f o r e x t r a c t i o n
and a n a l y s e s o f t h e e i g h t amino a c i d s . Fo r ty m l from each f l were
f i l t e r e d and f r o z e n a t -20 C. A t t h e end of t h e expe r imen ta l pe r iod ,
a l l t e s t f l a s k s p l u s t h r e e uninocula ted c o n t r o l f l a s k s were f r e e z e - - -
d r i e d i n a Virtis Uni t rap f r eeze -d rye r , Model No. 10-100. The f l a s k s
were then d e s a l t e d u s i n g t h e s o l v e n t e x t r a c t i o n t echn ique of Smith 8'
(1960). Samples were e x t r a c t e d t h r e e t imes wi th 10 m l o f ace tone
c o n t a i n i n g 5 % 6N HC1. One m l o f d i s t i l l e d water was added, t h e
sample placed i n i c e , and t h e ace tone blown o f f w i th f o r c e d a i r . Ten
mI of d i s t i l l e d water were added and t h e samples r e f r i g e r a t e d a t 4 C.
Amino a c i d c o n c e n t r a t i o n s were determined by adding 0.1 m l of sample B
t o a Beckman Model 119 au tomat ic amino a c i d ana lyze r . Those samples
c o n t a i n i n g low c o n c e n t r a t i o n s o f amino a c i d s were added i n q u a n t i t i e s a
>0.1 m l . Three f l a s k s o f un inocula ted medium were f r o z e n , e x t r a c t e d , L
- and added t o t h e amino a c i d a n a l y z e r a s s t anda rds . The accuracy o f
t h e column d a t a was 210-15%. The a n a l y s e s took 3 weeks, a f t e r which
t h e amino a c i d c o n c e n t r a t i o n s o f t h e c o n t r o l f l a s k s were remeasured.

E. Conversions o f n i t r o g e n
I n o r d e r t o de te rmine whether - Z. maritimum FMC and H 135 grew
a n a e r o b i c a l l y , mycelium was t r a n s f e r r e d on to e i g h t seawater a g a r
p l a t e s . Four were p l aced i n a Brewer anaerobic j a r c o n t a i n i n g a BBL
GasPak, and f o u r were incuba ted i n a i r . The GasPak genera ted
s u f f i c i e n t H2 and C02 t o c r e a t e an anae rob ic environment. Both
i s o l a t e s were a l s o grown i n a cand le j a r .
$
1. Nitrogen f i x a t i o n
The n i t r o g e n f i x a t i o n test was adapted from P o s t g a t e (1972).
Both i s o l a t e s were grown i n test t u b e s c o n t a i n i n g NSA wi thout
n i t r o g e n , w i th 15 gil o f aga r . The h l a n t s were incuba ted i n a i r and - i n a cand le j a r . After 2 and 6 weeks, t h e c o t t o n p lugs were pushed A
0 L
i n s i d e and serum s t o p p e r s p laced on t h e end of each tube . F ive c c o f i 4
a c e t y l e n e were i n j e c t e d i n t o each t u b e , and i n t o an un inocu la t ed :"
c o n t r o l . Gas samples were i rqjected i n t o a Carlo Erba Frac tovap Model * : 7
u 3 ;i
GV as Chromatograph, and t h e r e s u l t a n t peaks compared w i t h s t a n d a r d 7 l - i
a c e t y l e n e and e t h y l e n e peaks. I n case t h e a d d i t i o n o f 5 c c of i
a c e t i l e n e was t o x i c ( a c e t y l e n e would then c o n s t i t u t e 25% o f t h e gas d
volume), - t h e experiment was r e p e a t e d us ing 1 c c of ace ty l ene . -- - - - -- - - pp & -
Two methods were used t o t e s t f o r d e n i t r i f i c a t i o n by Za le r ion
maritimum. T e s t t u b e s c o n t a i n i n g n i t r a t e b r o t h (Difco) were made up

with a r t i f i c i a l s eawa te r . I n v e r t e d Durham tubes were added and t h e
t e s t t ubes were au toc l aved . The t u b e s were i n o c u l a t e d wi th a heavy
suspension o f FMC o r 135, t i g h t l y s e a l e d wi th screw c a p s , and shaken
such t h a t t h e mycelium went i n s i b t h e Durham tube . The t u b e s were
observed f o r 4 weeks f o r format ion o f g a s i n s i d e t h e Durham tube.
A d d i t i o n a l l y , n i t r a t e a g a r (Difco) was made up wi th a r t i f i c i a l
seawater and poured i n t o t h e bottom o f test tubes . S t a b c u l t u r e s were
then made of I s o l a t e s FMC and H 135. Cooled n i t r a t e agar was poured
ove r top of t h e f i r s t a g a r l a y e r . These c u l t u r e s were observed f o r 4
weeks f o r any ev idence o f g a s format ion .
To t e s t f o r format ion t3T n i t r i t e and a&onia from n i t r a t e , FMC
and H 135 were grown on N S A i n which N a N O was t h e s o l e n i t r o g e n 3 source . h f t e r 7 and 14 days i n ba t ch c u l t u r e , t h r e e f l a s k s o f each
2 - J
i s o l a t e were remov f i l t e r e d , and t h e f i l t ra te analyzed f o r n i t r i t e
and ammonium. N i t r i t e was measured accord ing t o S t r i c k l a n d and
Parsons (1972) and ammonium was measured wi th t h e ammonium e l e c t r o d e .
3 . N i t r i f i c a t i o n
FMC-and- t35 were ~ ~ w n - ~ ~ f - l p c o n t t m m u ~ b a h z I m x ~ - N S k -
c o n t a i n i n g NH@l a s the s o h - n i t r o g e n sourcs , -TRZS w a s w e e d - by-- --
.02 M MES. Batch c u l t u r e flasks were removed a f t e r 7 and 14 days , .
f i l t e r e d , and analyzed f o r n i t r i t e and n i t r a t e .

RESULTS & A . Growth o f inoculum on va r ious n i t r o g e q sources
Table 111 shows t h e r e s u l t s o f growing inoculum on a v a r i e t y of
n i t rogen s ~ u r c e s . - Emculum grown on 1 -a lan ine grew b e s t on 1 - a h n i n e
a f t e r both 7 and 14 days incubat ion . Growth was l e s s and equal on
NaN03, NaN02, and NH4C1. Inoculum grown on N H 4 C l grew e q u a l l y
on a l l four n i t r o g e n sources a f t e r 7 and 14 days. Inoculum grown qn b ~
NaN02 grew bes t on NaN02 and 1-alanine a f t e r 7 days. Af ter 14
days, growth was higher on 1-a lanine than on t h e o t h e r t h r e e n i t rogen
sources . Inoculum grown on NaN03 grew bes t on 1-a lanine and N a N 0 3
a f t e r one weeks' incubat ion . After 2-weeks' i ncuba t ion , t h e r e was no
d i f f e r e n c e , i n between t h e f o u r n i t rogen sources . '
B. Growth on inorgan ic n i t rogen sources. .
1. N H 4 C 1 , NaN03, NaN02, and 1-glutamic ac id r, * B 5 .
Both i s o l a t e s o f &. maritimum u t i l i z e d each o f 1-glutamic a c i d ,
I NH4C1, N a N 0 2 , and NaNO i n continuous and batch c u l t u r e (Tables 3 3 4 I V - V ) . Growth of both i s o l a t e s was h ighes t on t h e organic n i t rogen 3
- - -- - - - - - - --- - - - - - ".1 2 3
source a f t e r 1 weeks' incubat ion i n ba tch c u l t u r e . After 2 weeks i n %z $ - - - - - - -- - - - - - - - - - - - - p- -- - -
P batch c u l t u r e , growth o f I s o l a t e FMC was h ighes t on t h e o rgan ic
n i t r o g e n source , and growth o f I s o l a t e H 135 was equal on a l l n i t rogen
sources . Growth of i s o l a t e FMC was higher i n ba tch c u l t u r e a f t e r 1


Table IV.
The effect of inorganic nitrogen and 1-glutamic acid on growth of Z. maritimum F
MC
in continuous
-.
and bdtch culture.
I I CONTINUOUS
BATCH
Nitrogen source
No nitrogen
Note:
Means
* =
mean, *'
One week
Wo weeks
Dry weight (mg)
Final pH
Dry weight
Final pH
Dry weight
Final pH
d by a common line are not significantly different (Newnan-Kepls test, p <
0.0
5)
dard error sample size =
4


or n i t r i t e . After 2 weeks' incubat ion, growth was equ iva len t on t h e f'
three inorganic n i t rogen sources . Growth o f both i s o l a t e s lowered pH
i n batch c u l t u r e medium con ta in ing ammonium. In batch c u l t u r e , t h e pH d
* J
i
of media conta in ing n i t r a t e , n i t r i t e , o r 1-glutamic a c i d was r a i s e d by 4
growth o f I s o l a t e FMC. 1
7 --- - A - - A A - - - - - -- - L- -- - Y ,
I n continuous c u l t u r e , t h e r e was no d i f f e r e n c e i n growth on t h e I
1, .3
var ious n i t rogen sources . I s o l a t e H 135 grqw less i n 2 weeks than d id 4
-a - FRC and caused l i t t l e pH change i n t h e continuous c u l t u r e medium. d
There was a s t eady pH drop ( F i g . 6 ) when FMC was grown i n continuous
c u l t u r e on ammonium. When i s o l a t e FHC was grown without n i t rogen i n
continuous c u l t u r e t h e mycelium became very black and sporu la ted
heavily. This c o n t r a s t e d wi th t h e l i g h t coloured , asporogenous 2 3
myceliudP i n a l l o t h e r f l a s k s .
\
2. Various concen t ra t ions o f n i t r a t e , /
I n both continuous and batch c u l t u r e , growth-af I s o l a t e FUC
(Table VI) w a s equal a t 100 I& N / 1 and 10 mg N / 1 . There was no
d-ifference a n growth, i n continuous and batch c u l t u r e , between f l a s k s \
with 1 m g N / 1 , 0.1 mg, # / l , and no ni trogen. Growt-h i n continuous
c u l t p r e was h igher than growth i n batch c u l t u r e a t a l l concentra t ions . - - - -- - --- - -- -
t e s t e d . Growth uac;also & d e d ~ n p ~ b l i ~ h e d r e s u l t s ) i n NSA
-- i
-
conta-2UD-Ptg A A. ~ r o ~ i c o n t i n u o u i a ~ 6 a & F ~ i ~ 8 ~ ~ a ~ ----- - equal a t 200 rug N / 1 and 100 mg W1,. The lower t h e concen t ra t ion o f
- - - - - - -- --- - - -A -, -- -- - -

0 TI
f Figure 6. Changes i n p H during growth of Z. m a r i t i m u m FMC i n continuous f
-

. \ *
\ i
, .* a
\ =
o...... NO2- *\., m
o- L - GLUTAM ATE \ e--- NO3- *\*
6 8
TIME (DAYS)


n i t r o g e n , t h e d a r k e r t h e pigmentaton AT mycelium in. bo th c u l t u r e
systems.
Za le r ion maritimum H 135 (Table f $ X ) greu e q u a l l y i n con t inuous .- c u l t u r e a t 100 mg N / 1 and 10 mg N / 1 . $h b a ~ c h o c u l t u r e , growth was
_I.
lower at 10 mg N / 1 t h a n a t 100 mg N / l gFter one weeks' i ncuba t ion .
Growth a t 1 mg N / 1 , 0.1 mg N / 1 , and Ob &SA wi thou t n i t r o g e n was equa l
i n b a t c h c u l t u r e andsequal i n cont inuobh c u l t u r e . The pigmentat ion o f
t h e mycelium was l i g h t a t a l l concent r$ t ions both i n b a t c h and
cont inuous c u l t u r e .
3. Various c o n c e n t r a t i o n s o f n i t r a t e qhd glucose
There was p r o g r e s s i v e l y lower g r o w n o f 2. maritimum FMC i n ba t ch
and cont inuous c u l t u r e as concent ra t iof lp of n i t r a t e and g lucose were
lowered (Table VIII). I n ba t ch c u l t u b 4 , growth was equa l a t 1 mg N / l , ,
0.036 g C / 1 and 0.1 mg N / 1 , 0.0036g Cfk, A t t h e two lowes t 4
c o n c e n t r a t i o n s , t h e p igmenta t ion of t h P tttyceliurn was v e r y d a r k i n bo th
cont inuous and ba t ch c u l t u r e .
Za le r ion maritimum H 135 (Table I$] grew as w e l l i n con t inuous
c u l t u r e a t 100 mg N/1, 3.6 g C / l as a t 10 mg N / l , 0.36 g C / 1 . Growth p--p- ---- -
A
J'in con t inuous c u l t u r e a t 1 mg N / 1 , 0.69 & C / l was h ighe r than growth 3
- - - --- - - - - - - - - 4
a t 0.1 mg N / 1 , 0.0036 g C / 1 . I n ba tcb ~ L l t u r e , growth was g r e a t e s t a t
t h e h i g h e s t c o n c e n t r a t i o n of carbon aad 4Atrogen. Growth was h ighe r

m a O W Q C O rfC(! . . . . . . a m d o O D 0 coo m + I Wl + I *I

r l m COO . . I .
cn-4' O d * I *I
I- m . . d'o
+ I
rl \rl v'.
Z
m~ 'is'- * 0
0 rl


---- - -
0.0036 g C / l . Growth of both i s o l a t e s was higher i n continuo'us than
batch c u l t u r e a t a l l concen t ra t ions t e s t e d .
.C. Growth on organic n i t rogen sources
L .
1. S i n g l e amino a c i d s - - - ---
1 Although H 735 and FRC 1 grew t o 'a l i m i t e d e x t e n t on nea r ly a l l , -
o f t h e 21 amino a c i d s t e s t e d (Table XI, only a few supported good
v growth. I s o l a t e H 135 grew b e s t on 1-val ine , 1-a lanine , 1-glutamic
a c i d , 1 -aspa r t i c a c i d , 1-glutamine, 1-asparagine and 1-arginine. \
I I s o l a t e ~kk grew b e s t on 1-a lanine and 1-arginine. Both i s o l a t e s grew . 3 .[ ,
poorly on 1-glycine , 1-methionine, 1-cys t ine , 1 - lys ine , and r
1 - h i s t i d i n e . A yellow green d i f f u s i b l e pigment was produced by
I s o l a t e FMC i n those f l a s k s which contained r ing-conta in ing amino
- a c i d s .
2. A combination o f amino a c i d s i
In continuous c u l t u r e , both i s o l a t e $ grew a s wel l on s i n g l e amino I
a c i d s as on t h e combination o f &in0 ac ids . Growth o f I s o l a t e FMC e+'
able X I ) after 7 days . in ba tch c u l t u r e was higher ona 1 -a lan ins and
t h e t h r e e amino a c i d mixture than on 1-arginine and 1-glutamic ac id .
After 14 days, gr0ul.h was equal .on a l l four n i t r o a e n sources . IKI --
-- - - -- -- - -- - - --
batch c u l t u r e , pigments had formed+af ter 7 days i n t h e f l a s k s --
-- -- ----- -
con ta in ing 1-arginine-HC1, 1-glutamic 'acid, and t h e t h r e e amino a c i d
mixture.
* <
- - - -
- r

Table X. The effect of single amino acids on owth of Z. maritimum P" - FMC and H 135 in batch culture. I
Isolate FMC Isolate H 135
Amino acid Dry weight Final pH Dry weight Final pH
Monoamino monocarboxylic t
Monoamino dicarboxylic
L-glutamic acid $4.3 7.7 $1.4 7.5, -1.2 -0.5
L-aspartic acid
f Diamino dicarboxylic

Table X (continued)
- - Isolate FMC I s o l a t e H 135
&no a c i d Dry weight F i n a l p H D r y weight F i n a l p H
- Ring-containing
-
L- t ryptophan $3.8 7.2 $3.4 7.4 1 -0.7 -2.5
With hydroxyl & L-s e r i n e
L-threonine
O t h e r -

Table X (continued)
I so l a t e FMC I s o l a t e H 135
Amino Acid Dry weight Final p H Dr>y weight Final p H
No nitrogen
NaNO 3
Note: - * = mean ** = standard e r r o r sample s i z e = 3


Gm&B ef &We H -5 WZe :=I+--w- 7 days f il
batch c u l t u r e on a l l four n i t r o g e n sources. Af te r 14 days, growth was
h igher pn 1 -a rg in ine and 1-a lanine than on 1-glutamic a c i d and t h e
amino a c i d combination. . ,
3. Growth on o rgan ic n i t rogen sources o t h e r than amino a c i d s
Zaler ion mari t imum F1C- l T a b l e i X I I I l grew on u c ~ - p e & o n e - i
vi tamin f r e e casamino a c i d s , a l l a n t o i n , a l l a n t o i c a c i d , and , . ',
c i t r u l l i n e . Growth was poorest on a l l a n t o i c a c i d , b e s t on urea . C
Zaler ion maritimum H 135 (Table XIV) grew on u r e a , peptone, v i tamin
f r e e casamino a c i d s , a l l a n t o i n W f a n d c i t r u l l i n e . Growth on c i t r u l l i n e
occurred dur ing t h e second, week b u t ndt t h e f i r s t
I s o l a t e H 135' grew b e s t on urea and beptone.
D. Growth on combinations o f n i t r o g e n sowces
1. Ammonium and n i t r a t e
week o f incubat ion .
The r e s u l t s i n Chapter I showed t h a t both i s o l a t e s o f q
\ ' *"
Z. maritimum took up ammonium in preference t o n i t r a t e . Nitrate was, - ,. *
no t used by e i t h e r i s o l a t e dur ing t h e 14 day incuba t ion period. .
.? t '* - i
2. Ammopium and n i t r i t e * 6 - - - - - -- - -- - - - -
C
Zalerion maritimum FMC took up n i t r i t e and ammonium , . - - - - -- - - --- ----- -
simultaneously from the n u t r i e n t s o l u t i o n (Fig. 7 ) . Ammonip was ' ; -: 5 3 .
removed a t a f a s t e r r a t e than n i t r i t e . Phosphate was n o t e x h h s t e d


\
\ p
ab
le. xIII.
~h
d
;f fe
ct
df o
rga
nic
niJ
zrog
en
sou
rce
s o
n g
row
th o
f Z
. m
ari
thu
m >FMC i
n b
atc
h c
ult
ure
. 1
-
I
One
wee
k Tw
o w
eek
s :4
, d
i
,-
t '
~r
y
wei
gh
t (m
g)
- F
ina
l D
ry w
eigh
t/ (mg)
Fin
al
pH
*
.
- -
X'
. -
SX
-
X
I
I
SX
I
I 17
.8
1.5
7.7
L '$
$ &)
h4
( 25
.5
, 1
0.4
, 7.5
1 4
. N
itro
ge
h s
c rc
e -
1 '
No
nit
rog
en
'I
* '
I
' A
lla
nto
ic. a
.
e
I P
epto
ne
,
I I p i
t ra
te' .
I ~ a
All
an
toin
I I
Urea
1' I,
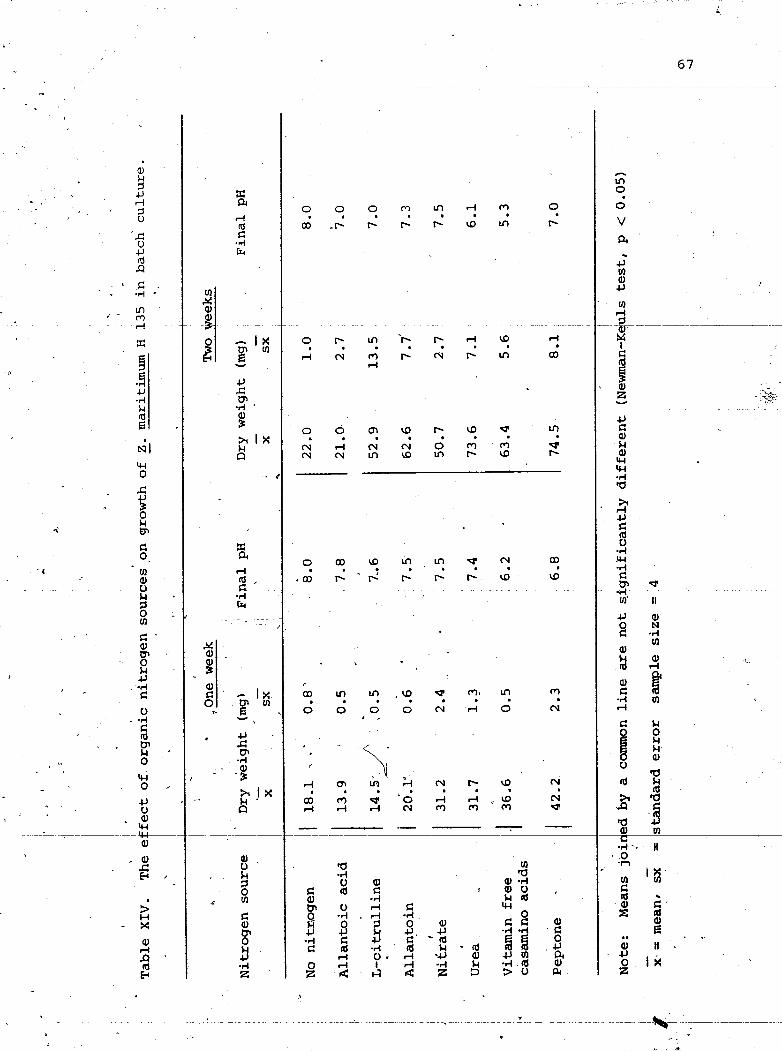
fec
t o
f o
rga
nic
nit
rog
en
so
urc
es.
on
gro
wth
of - 2.
m
arit
imu
m H
1
35
in
batch
cu
ltu
re.
Ta
ble
XIV.
Tpe
e
I I
A
I "
,One
wee
k Tw
o yv
eeks
I
Nit
rog
en
SO
U~
C~
' D
ry w
eig
ht
(mg)
,
Fin
al
pH
.
Dry
we
igh
t (mg)
Fin
al
pH
.-
-
-
-
SX
X
S
X
4
I I
I
( 1
8.1
.
+-
0
.8'
No
nit
rog
en
I
I
All
an
toic
ac
id
1 1
3.9
I I I
if
ra
te
I
Ure
a I
,
I I
* ;
Vit
amin
fr
ee
1 1.
cis
min
o a
cid
s
I
I N
ote
:
Mea
ns
ed by
a c
omm
on
lin
e a
re n
ot
sig
nif
ica
ntl
y d
iffe
ren
t (~
ew
ma
n-~
+ls
tes
t, p <
0.0
5)
I .'
t I
- '
d
sta
nd
ard
err
or
sam
ple
s
ize
= 4
C1
1

Figure 7'. Changes i n ' d r y weight, p H and cyncentra t ions o f g lucose ,
phosphate,
- c u l t u r e of
i amm6inium, aiid n i t r i t e d y i n g
Z. m a r i t i m u m FMC and - H 135. -
-- -
g r o w 6 i n batch

3.6 +PI-B
1 8
I/
I
I 8
I -..
GLU
CO
SE -
k,/+
'.
%>
'.
0 0
'0..
t
.O 62
.4 -
'. 4 -. . . . ,.
p.
A*..+
,.... " .-
....
42
z ../
PH
OS
PH
ATE
k... ---...,
*. i
-4
s
-0
0
I ----.-%
.+
*,*. 1 T
6 3
1.2
- G
(3
,/'
I I %
..
/
k.
TIM
E -D
AY
S

dur ing the 14 day incuba t ion period. I s o l a t e H 135 (Fig . 7 ) removed
ammoniurg r a p i d l y from the ' n u t r i e n t s o l u t i o n dur ing t h e first 4 days o f
growth. N i t r i t e was note&&n up u n t i l hay 4 , and was then slowly
taken up f o r t h e d u r a t i o n of t h e experiment. Af ter 14 days, 11% of
t h e phosphate remained i n t h e n u t r i e n t s o l u t i o n . Neither i s o l a t e
excre ted n i t r a t e a t any time during t h e experiment. - - - - A -
3. N i t r a t e and 1-glutamic a c i d L.
The two i s o l a t e s responded d i f f e r e n t l y t o t h e presence o f t h e s e
two n i t rogen sources i n t h e n u t r i e n t s o l u t i o n (Fig. 8) . I s o l a t e FMC 4
p r e f e r e n t i a l l y took up t h e amino a c i d , exhaust ing i t from t h e n u t r i e n t
s o l u t i o n a f t e r 9 days. N i t r a t e was no t removed from t h e n u t r i e n t
s o l u t i o n dur ing t h e 14 day incuba t ion period. I s o l a t e H 135 took up
1-glutamic ac id e x c l u s i v e l y u n t i l day 6, whereupon t h e n i t r a t e was
removed, i n smal l e r q u a n t i t i e s , along wi th t h e 1-glutamic ac id .
C
4. Eight amino a c i d s and ammonium
Zalerion maritimum FHC (Table XV) r a p i d l y removed 1-threonine,
1-ser ine , 1-a lanine , 1 -va l ine , 1- iso leucine , and 1-leucine from t h e
n u t r i e n t so lu t ion . The l e v e l s o f 1-glutamic a c i d and 1 -aspar t i c a c i d 0
were comparat ively h igh and v a r i a b l e throughout t h e experiment. *
t h r e e days o f incubat ion . The ammonium l e v e l then rose and s tayed
r e l a t i v e l y cons tan t for t h e dura t ion of t h e experiment. Despi te t h e

Figure 8. Changes i n dry weight, p H , and concentrations of 1-glutamic
acid and n i t r a t e during growth i n batch cu l ture of Z. - inari t i m u m FMC and H 135.
- FMC; . - H 135.


', -. ,
1
-
. - - --- v- - - - -
', 8 *
*
Table XV. Changes i n concentrations of m i n o ac ids &n khe' nut r ien t ' so lu t ion during growth of 2. maritimum FMC i n batch cul tu re .
Concentration UM Bef o r e Inoculation Day 3 , Day 6 Day, 9 Day 12 Amino ac id
spart tic ac id 50
Tbeonine 50
Serine 50
- Glutamic ac id . 50
Alanine 50
Valine 50 t
Isoleucine A 5 0
Leucine
Dry Weight (mg) a $4.5 23.9 41.2 $5.2 -0.. 1 -4.8 -4.3 -5.3 , -
Fina l p H 7.5 7 . 3 6 ;4 6 . 3 5.9 A
. i + Note: Numbers a r e means o f . t h r ee samples - one standard e r ro r .

a d d i t i o n of MES, t h e pH f e l l t o s l i g h t l y below 6.0 dur ing t h e 3
experiment. I s o l a t e H 135 (Table X V I ) took up, t o some extent, ' a l l o f .,* I
% L
t h e amino a c i d s suppl ied i n t h e n u t r i e n t s o l u t i o n . . L-aspartici a c i d ,
1-alanine, 1-val ine , 1 - i so leuc ine , and 1-leucine were taken up a t a -
f a s t e r r a t e than were 1- threonine , 1-ser ine , .and l - g l u t a k q ac id .
There w a p more v a r i a t i o n i n amino a c i d concen t ra t ion megsucements - -
P
a f t e r 9 and 12 days incuba t ion than t h e r e was a f t e r 3 and 6 days'
\I incubat ion . I s o l a t e H '35 removed apvnonium from t& n u t r i e n t s o l u t i o n
throughout t h e @-owt'h per iod .
E. Conversions o f n i t r o g e n sources ' \
C
1. Nitrogen f i x a t i o n
~ 5 t h i s o l a t e s grew on a g a r without n i t rogen . Acetylene was n o t - * -
~. , I .
reduced by any of t h e samples t e s t e d . ; ,
2. D e n i t r i f i c a t i o n
When grown on n i t r a t e - aga r and n i t r a t e b r o t h , t h e r e was no . .
measureable N2 production by e i t h e r i s o l a t e . Neither i p o l a t e grew
i n t h e s t a b c u l t u r e where t h e cond i t ions were anaerobic . *
. >
When i s o l a t e FHC was grown i n NSA conta in ing n i t r a t e , n k t r i t e was - - - - - - - ep- A- --- ---- -
de tec ted i n t h e n u t r i e n t s o l u t i o n after 1 and 2 weeks' growth. . -
N i t r i t e was de tec ted a f t e r 2 weeks' growth o f I s o I a t e 135. There was >
no measureable ammonium produced by e i t h e r i s o l a t e , h e ammonilln t

Table XVI. Changes i n concentrations of amino ac ids i n t h e nu t r i en t so lu t ion during growth of Z. maritimum H 135 i n batch - cu l tu re .
Concentration p M Before
Amino ac id Inoculation Day 3 Day 6 Day 9 g a y 12 - -
Aspartic ac id 5 0 39 -1.1
36 -0.9
z7 ,- -6.6
3 -5.4
Threonine 50 . 39 ? 37 j7 ,? . . -2.5 -1.6 -6.6 -1.4
Serine -
.Glutamic ac id 5 0 42 -3.5
3l -1.0
34 -5.4
32 -4.0
Valine i -
50 3 O -2.7
P -5.1
Isoleucine 5 0 ?2 -2.8 -2.3
3 -8.3
L e u c i ~ e . .
50 ?3 -2.7 -2.6 -7.4
3' . -5.7
~~21' ( m ~ ) . 3.56 ), 78 J.17 q . 9 1 9.85 -0.84 - 0 . 0 1 ' -0.17 -0.1
Dry Weight (mg)
Fina l p H 7.5 7.3 6.8
+ Note: umbers are means of three samples - one standard error.

- - - - -
concen t ra t ions in t h e inocu la ted s o l u t i o n s a f t e r 1 and 2 weeks'
incubat ion were lower than i n t h e uninoculated c o n t r o l .
3. N i t r i f i c a t i o n
Neither i s o l a t e exc re ted measureable amounts of n i t r i t e o r
n i t r a t e a f t e r 1 weeks' growth on NSA with NH4C1 a s a n i t rogen
source.
grow
b
The observat ion was made t h a t al though Z. m a r i t i m u m FMC d id n o t
anae rob ica l ly , growth on an a g a r p l a t e incubated i n a candle j a r /'
was g r e d tei. than growth i n a i r . After 3 weeks' incuba t ion , t h e /
diameter of c o l o n i e s grown i n candle ja rs .was approximately 15 mm
greater than c o l o n i e s grown i n a i r .
F igure 9 sumlrragizes t h e h y p o t h e t i c a l r o l e of 5. m a r i t i m u m i n t h e
marine n i t r o g d y c 1 l a .
DISCUSSION /
Nitrogen s n u t r i t i o n i n Z. maritisurn w i l l be dis&ussed, followed by
a comparison of t h e s e r e s u l t s with t h e a v a i l a b i l i t y o f n i t rogen i n t h e
-
sea .
- - - - I
The e f f e c t o f growing inoculum on var ious n i t r o g e n sources on
p a t t e r n s o f n i t r o g e n n u t r i t i o n has n o t been i n v e s t i g a t e d i n marine

Figme 9. Summary diagram of nitrogen n u t r i t i o n i n 2. m a r i t h u m . -

- Nitrogen sources used for growth. "-. - Preferred nitrogen sources for growth.
---I---+ Nitrogen sources excreted into* solution. -
- \

l-- week o f incubat ion , before t h e mycelium had s'acclimatized" t o the - . C . 4
J -. - - 3 bl - ' n i t r o g e i source. Despite t h e h i s t o r y of t h e inoculum, t h e o rgan ic
i 4 4 A
n i t rogen 'source ( i n t h i s case a l a n i n e ) genera l ly supported t h e h ighes t 4 'ii
erowth. Inocwlum grown on NaN02 grew b e t t e r on NaN02 than on t h e - 3
- - - 3 n - -pa Il?i Other inorgan ic n i trogeq* sources ; inoculum o n NaNO3 grew
0
t .+ g e t t e r on NaNO than on t h e o t h e r inorganic n i t rogen sources; 3
d
1 i p o s s i b l e t h a t the enzymes necessary for t h e reduct3on of n i t r a t e and -
- I < - %
n i t r i t e were induced when t h e inoculum was grown o n t h e corresponding 2
7 .
n i t r g g e n source. 'l'here would t h e r e f o r e be no l a g period necessary f o r G i
-.! t h e s y n t h e s i s o f n i t r i t e and n i t r a t e r e d u c t a s e w$pn t h e 'i
d - '2 +s 3
t r a n s f e r r e d ' t o f r e s h medium. Inoculum grown i n ammonium d i d n o t +.
4 - .c
p r e f e r am&nirna t o o t h e r inorgan ic n i t r o p n 'sources: Poss ib ly t h e 4 - -- + - - - - -ii 7*
inoculum was a f f e c t e d I$ t h e law pH cond i t ions produced by the uptake + f'
71
i % =>
- of t h i s ion . . :7" * - a
a % G , ilr L I 4
. - 2 'x: I 3
-The r e s u l t s from t d e n i t rogen u t i l i z a t i o n . s t u d i e s i n d i c a t e t h a t f -
a41 t h r e e of t h e inorgan ic 'n i t rogen sources commonly f o w d i n th; sea - f r
apparen t ly u s e any one o f n i t r a t e , zii e, o r ammdnia t o synthesLze * ----
a 2 - l n o c T d s ~ e s s a r y f o r myceXal_protein. &her i n v e s t i g a t o r s

*, Yie a ur. < grew on each o f NH4C1, KN03, KN02, and NH4N03. Johnson kt
* e'=
@ (1.959) found t h a t 21 s p e c i e s o f higher marine fungi grew on agar
"containing NaN03, W03, and (NH4 ) 3P04. Tubaki ( I966 ) showed
/c that one marine fungus could u t i l i z e t h e end product o f n i t rogen B a
r educ t ion by another marine fungus. He demonstrated t h a t
Linder could grow on- n i t r i t e c o n t a i n i m m e e a a ,+
- - - - , UP----
when c u l t i v a t e d wi th ~ o r o l l o s ~ o r a maritimk Werdermann, t h e former \
spec ies u t i l i z i n g t h e ammonim produced a f t e r n i t r i t & reduct ion by t h e Y
- A
l a t t e r species: -
In c o n t r a s t t o t h e h igher marine fung i , marine Phycomycetes grow . . . 2 >: 4
0
poorly on inorganic n i t rogen sources . Alderman and Jones (19719 1 f C ,5 d
demongtrated t h a t Al thornia c r o u c h i i Jones and Alderman d id not use 6 P1
n i t r a t e o r ammonium. Thraus tochytr i& mult i rudimenta le Goldstein and .*
. - T. motivum Goldstein (Golds te in , 1963a) d id not use KN03 o r
(NH4 )$04* Thraustochytrium roseum (Goldstein, 1 9 6 3 ~ ) and
Schiz0chytriu.m a m r e ~ a t u n Golds te in ( G o l d ~ t e i n and g el sky, 1964) grew . 2 '
I.
poor ly on ( N H ~ ) ~ s o ~ and d id no t grow on WO The majo r i ty o f . 3 ' both higher and lower marine fung i have been shown t o grow b e s t on
o rgan ic sources o f nitFogen. . P P
- >---- -7 -
%
A 1 1 of t h e abovementioned s t u d i e s were c a r r i e d ou t e i t h e r on
4
e ~ p e r i m e n t s ~ r e p o r t e d he re show that continuous cu l tu r@ r e s u l t s 0 '
d i f f e r e d q u a l i t a t i v e l y a s well a s q u a n t i t a t i v e l y from batch c u l t u r e

*$
- - ---- - -, ,- r e s u i t s . In batch c u l t u r e , i s o l a t e s o f Z. maritimum grew i
s i g n i f i c a n t l y b e t t e r on 1-gl'utamic 'acid than on inorgan ic n i t rogen I
sources . However, i n continuous' c u l t u r e t h e r e was no preference shown
. by e i t h e r t s o l a t e f o r t h e o rgan ic n i t rogen source.
- .
r. P
The poorer growth on t h e inorgan ic n i t rogen sources i n ba tch - ! c u l t u r e may be explained by t h e presence of growth l im5t ing f a c t o r s i n
L,
t h e n u t r i e n t s o l u t i p n . It was demonstrated i n Chapter I t h a t t h e pH
ilrop which accompanies g roy th on ammonium can limit growth. When
i - ammonium was s u b s t i t u t e d by - n i t r a t e o r - n i t r i t e , t h e pronounqed f a l l i n
pH d i d n o t occur. Grbwtli4was nonetheless lower on inorgan ic than
o rgan ic n i t rogen s o u r c e s , poss ib ly due t o t h e exhaust ionrof phosphate, . C
which is necessary f o r n i t r a t e r educ tase a c t i v i t y (Nicholas and
Scawin, 1956). It is a l s o p o s s i b e t h a t exhaust ion of molybdenum ( t h e 3 - - -
a
metal3 ic c o f a c t o r i n n i t r a t e r educ tase ) o r some o t h e r e s s e n t i a l 7- *
. - micro'element i n h i b i t e d growth in batch c u l t u r e on inorgan ic n i t r o g e n .
sources.
S t u d i e s wi th low concen t ra t ions o f n i t r a t e showed t h a t both . I '1
i s o l a t e s grew wel l i n cont inuous c u l t u r e a t one t e n t h t h e n i t rogen
con ten t of NSA. Thus a ten times chdage i n t h e C:N r a t i o , f'rom 30: 1
P '
( 1963 ) demonstrated a requiremrtat f o r a -h ighALmt50- in - - -: ?
- - -
T . achrasporum, H a l o ~ a l l o n e l l a , 2. s p e d t a b i l i s , and medioset ikera. '
Sguros e t (1973) showed t h a t t h e l a t t e r t h r e e species. couid grow a t
T - low n i t rogen l e v e l s and a t a h igh C:N r a t i o . 6
- - - - - - -
I -

-- - - -
In the continuous c u l t u * e ~ s ~ s t e m , *both i s o l a t e s g r e a e q u a l l y on % - \
t h e two Lowest n i t r o g e n concentratiorrk'and on medium without n i t rogen . A
A c e r t a i n n th resho ldn l e v e l of ni trogen. may be necesstii-y be fo re good
growth can opcur. A p o s s i b l e explanati-on could be t h a t the very high
C:N r a t i o (3,000: 1 ; 30,000: 1 ) i n these experiment3 produced a growth
i n h i b i t i o n e f f e c t . Gowth a-t low concen t ra t ions of n i t r a t e and i n NSA - - - - -L A - - - - - - - - - - -
ui=nrogen was h igher in ' continuous than i n ba tch c u l t u r e . This
sugges ts t h a t growth can occur i n t h e continuous system a t very low
n i t r o g e n l e v e l s , poss ib ly on the n i t rogen suppl ied a s contaminants i n
reagent ,grade chemicals.
\
Qecent s t u d i e s (Meyers 1966; Meyers and Hoyo, 1966; Byrne and
Jones, 1 9 7 5 2 , b ) have i n d i c a t e d t h a t spore p r o d u c t i c m m d spore 4
/
germination may be a s important a s d ry weight i n measuring t h e
. physio logica l response o f a .mar ine fungus. Although t h e c o n t e n t s of
each f l a s k were not s y s t e m a t i c a l l y examined f o r e r u l a t i o n , s e v e r a l
observat ions were made i n t h e course of e n t s . There was
h a d i r e c t r e l a t i o n s h i p between sporula't ion of 2. mariti um FHC and l a c k 4 4
of n i t rogen i n t h e n u t r i e n t i iolut ion. Holl igan and J e ~ i n g S ( 1972a)
observed t h a t c o n i d i a t i o n o f t h e marine fungus Dendryphiella s a l i n a
(Suth.) Pugh and Nicot was heavy a t low n i t r a t e l e v e l s . Mg e t a&
continuous c u l t u r e was r e l a t e d t o t h e concentrat ion-of carbon a n d - - -- -- lp - - - - ---- - -- - - -- - - -
n i t r o g e n i n t h e c u l t u r e medium. They s t a t e d t h a t the/most g e n e r a l l y . . . .
e s s e n t i a l 9 0 d i t i o n f o r con id ia t ion was t h e absence o r nea r absence o 3

a W i W l e n i t r ~ a h t h e p e c e of a s s imi lab le carbon. Zalerion
maritimum FMC, on t h e o t h e r hand, sporu la ted heav i ly a t low
, concen t ra t ions o f n i t rogen and i n t h e absence o f a s s i m i l a b l e carbon.
4 When coneen t ra t ions o f cartion and n i t rogen were both lowered,
thus kepping t h e carbon t o n i t rogen r a t i o cons tan t , r e s u l t s ' d i f f e r e d a
-- - betweerr i s o l a t e s am--between batch andLcGXIiiu6iiGs yst%iiz lnnbatc h
c u l t u r e l a c k o f g lucose red&ed growth o f both i s o l a t e s when glucose
concen t ra t ions were reduced t en fo ld . I n continuous c u l t u r e , good -
. I
growth ~ c c u r r e d a t 1 mg N / 1 and a C:N r a t i o o f 30: 1. However? a t 1
N / 1 and a C:N r a t i o o f 3,000:1, growth was no higher than on n u t r i e n t
s o l u t i o n without n i t rogen . - This would i n d i c a t e t h a t t h e C:N r a t i o may
a f f e c t growth i n cont inuous c u l t u r e of 2. maritimum H 135.
P I n summary, t h e r e s u ~ t s i n d i c a t e t h a t both i s o l a t e s o f Zaler ion
o'an grow on low concen t ra t ions o f n i t rogen , and a t high C:N r a t i o s .
However, r e s u l t s from t h e continuous c u l t u r e system demonstrate t h a t a
C:N r a t i d o f 3,000: 1 may i n h i b i t growth o f i s o l a t e H 135. Sporula t ion
was encouraged by low concen t ra t ions o f n i t rogen i n t h e n u t r i e n t "
s o l u t i o n . 9 I
I
\
Both i s o l a t e s o f Z. ~ r i t i m ~ grew t o a c e r t a i n e x t e n t on nea r ly - -- - - --- - - -- --
a l l of t h e & i n ~ a c i d s t e s t e d . Small growt inc reased may ha re been, - 5 2 -
due t o contaminants such a s v i tamins , which ban adhere t o o rgan ic s
r eagen t s i ~ o c h r a n e , 1958). Both i s o l a t e s ' grew on each o f t h e amino . .v
d < - .
.

a c i d s most commonly found i n t h e s e a (Hood, 1963): th reon ine , s e r i n e ,
\ i s o l e u c i n e , l e u c i n e , glutamic a c i d , ' a s p a r t i c a c i d , a l a n i n e , and *
vali#e. Growth o f bo th i s o l a t e s was p a r t i c u l a r l y good on a l a n i n e , I
poss ib ly because t h i s amino a c i d p a r t i c i p a t e s i n t ransaminat ion I
r e a c t i o n s . Growth was a l s o good on a r g i n i n e , an &no a c i d r e h t e d t o L
- - n i t r o g e n con ta in ing s t o r a g e products i n wood. The r i n g con ta in ing
- - - - -
amino a c i d s appeared t o func t ion i n pigment formation i n I s o l a t e FMC. I
I s o l a t e H 135 grew well on t h e monoaminodicarboxylic a c i d s and
' t h e i r amides, a s d i d Hqlosphaeria meckfoset i~era, Cu lc f t a lna
achraspora , a+Humicola a l o p a l l o n e l l a l (Sguros e t a l ,
Zalerion maritimum grew wel l on most o f t h e o rgan ic n i t rogen L
sources o the r than amino a c i d s which were t e s t e d . Urea was a good
source of n i t rogen f o r 3. maritimum, a s it was f o r H. m e d i o s e t i ~ e r a ,
H. a l o p a l l o n e l l a , and C. achraspora (Sguros eLa, 1973). On t h e - o t h e r hand, Johnson s t a 1 (1959) found t h a t most o f t h e marine s p e c i e s I
I
3 1
t e s t e d grew poorly on urea . The wood s to rage products a l l a n t o i n and .
c i t r u l l i n e (Kremers, 1963) supported 'good growth o f both i s o l a t e s .
This sugges t s t h a t - Z. maritimum, which grows on c e l l u l o s i c s u b s t r a t e s
i n t h e marine environment, may be a b l e t o u t i l i z e t h e n i t rogeh i n
wood.. a
a \
- I > - -- - - - - - p-pp \ ,--- - --
-p--pp-
\
b - - - - - To axte ,-marinee m ~ - o ~ o ~ i s t stmve-*stuclte& thmf f e e t ef- nit rew- -- - --
I
sources suppl ied s i n g l y i n t h e n u t r i e n t so lu t ion . I n n a t u r e ,
2. maritimum grows i n an environment con ta in ing v a r i o u s inorgan ic a+ -

n i t rogen sources were t aken up when a combination o f forms was, ' ,
suppl ied .
Chemical a n a l y s i s o f t h e n u t r i e n t s o l u t i o n demonstrated t h a t -
Z. maritimum took up ammonium t o t h e complete exclus ion o f n i t r a t e .
It can t h-=exfore h s l a s s m e L t _the_pmLae,n9eeo ff_ar_0_111I~lnh1b_i5ed _ - 1
n i t r a t e r educ t ion , by r e p r e s s i n g t h e s y n t h e s i s o f n i t r i t e
r educ tase (Morton and MacMillan, 1954; Kinsky, 1961 ; Cove and Pateman, -
1969). Sguros and Simms (1963) suggested t h a t H. mediobetigera;
T. achrasporum, g. a l o p a l l o n e l l a , and Q. s p e c t a b i l i s p r e f e r e n t i a l l y - u t i l i z e d ammonium t o n i t r a t e , because of t h e r ap id pH drop in media
con ta in ing ammonium. P r e f e r e n t i a l u t i l i z a t i o n o f ammonium over -.-.
n i t r a t e i s genera l ly acknowledged (Morton and MacMillan, 1954;
Nicholas, 1965) t o be t h e p a t t e r n i n t e r r e s t r i a l f w i . - - T h e usual
evidence presented f o r tLis theory is a sharp drop i n pH d y i n g e a r l y
growth.
Analyses o f t h e n u t r i e n t s o l u t i o n suggested t h a t 2. maritimum - < *
took up n i t r i t e a t a slower r a t e than ammonium. There was no removal
/ o f n i t r i t e dur ing t h e first two days ~ f ' ~ r o w t h , a l a g time which may -
b have been necessary f o r t h e induct ion o f n i t r i t e reductase . Morton
-and MacMillan (1954) showeci t h a t ammonium and n i t r i t e were. e-

The f a c t t h a t a n u t r i e n t has been removed from t h e c u l t u r e medium
does n o t n e c e s s a r i l y mean t h a t t h e n u t r i e n t has been u t i l i z e d . 1t- is
p o s s i b l e t h a t a c e r t a i n propor t ion of t h e n u t r i e n t s may have been - A /
adsorbed t o t h e c e l l wal l . Detect ion i n s i d e t h e c e l l by mass
.spectroscopy o r t r a c e r methodology does not prove t h a t a n u t r i e n t has
been u t i l i z e d . Both ammonium and amino a c i d s form pools wi th in t h e A
-- --A - ---- -
-
mycelium, from which t h e y may o r may n o t be removed f o r amination, ' - - !
4 t ransaminat ion , o r t h e s y n t h e s i s o f c e l l u l a r p r o t e i n . There is
evidence t h a t Zaler ion r e q u i r e s a ni t rogen source f o r good growth. ' -
Thus we can assume t h a t a por t ion o f t h e n i t rogen which was removed I
from t h e n u t r i e n t s o l u t i o n was used i n c e l l u l a r metabolism.
Glutamic a c i d was t aken up e i t h e r a t a f a s t e r r a t e than n i t r a t e
( I s o l a t e H 135) o r t o t h e complete exclus ion of n i t r a t e ( I s o l a t e FMC).
Amino a c i d s a r e . thought t o be incorporated d i r e c t l y i n t o p r o t e i n
without p r i o r degradat ion t o ammonia (Nicholas, 1965; Sguros e t a l , - 1973). It is t h e r e f o r e l o g i c a l t h a t glutamic a c i d would be more
r e a d i l y taken up than n i t r a t e , which must be ' reduced t o n i t r i t e and b 0
ammonia before p r o t e i n s y n t h e s i s can occur. I
* - ll , .
. . Y
Both i s o l a t e s s imul taneously removed both amino a c i d s and
f
p a r t i c i p a t e s i n t ransaminaton r e a c t i o n s , were taken up by I s o l a t e FMC
i n smal l q u a n t i t i e s . This is i n apparent a t r a d i c t i o n t o t h e r e s u l t s pr , .

- -- - - --- - A - - a - ------- - ---- -- -
which -- show -- t h a t - -- -- glutamic a c i d was completely removed from t h e n u t r i e n t
s o l u t i o n ;hen supp l i ed i n combination with n i t r a t e . The poor uptake
o f glutamic a c i d and a s p a r t i c a c i d may be explained by t h e presence o f
ammonium i n t h e n u t r t e n t s o l u t i o n . Robinson, Anthony, and Drabble i
(1974) found t h a t ammonium was used before glutamic a c i d dur ing
cond ia l germination and growth o f Aspergi l lus c l a v a t u s . They
suggested that ammonium ac ted from wi th in t h e c e l l , poss ib ly - - - --- --La -La--- - A- -- - --
L /
r e p r e s s i n g t h e s y n t h e s i s o f t h e permease necessary f o r t h e t r a n s p o r t '
of glutamic ac id . An a d d i t i o n a l explanation may be t h a t t h e r e were
- --
i n h i b i t o r y i n t e r a c t i o n s i n t h e mixture o f amino a c i d s (Nolan, 1970),
o r t h a t t h e r e was competi t ion f o r uptake between t h e amino aci,dS
(Jones , 1963). . The opt imal pH f o r the a c i d i c amino a c i d t r a n s p o r t 1
system i s approximately 5.5 (Hunter and Segel , 1971), and t h e n e u t r a l
amino a c i d s may have been more r e a d i l y taken up a t t h e pH range
(6.0-7.5 of t h e n u t r i e h t s o l u t i o n dur ing t h e s e experiments. - -
0 -
The r a p i d removal of t h e o t h e r amino a c i d s was n o t r e l a t e d t o
t h e i r a b i l i t y t o suppor t growth. Rlanine, an e x c e l l e n t n i t rogen -
b a
source ' f o r both i s o l a t e s , was removed. a t t h e same r a t e as i s o l e u c i n e
o r l euc ine . Previous work wi th t e r r e s t r i a l fungi (Zalokar , 1961 ; f
G o t t l i e b , Moi tor is , and E t t en , 1968; Hunter and Segel , 1971) showed i
t h a t amino a c i d s are removed from t h e n u t r i e n t b l u t i o n and &red i n in ternzlp --
pools , rom which t h e y may o r may no t b e withdrawn far
incar po-mti 01-into c ~ r 3 € e ~ r ~ o t h e r ~ c ~ a T r S a ~ i o n S ~ $e rate
o f uptake i s n o t d i r e c t l y dependent on subsequent metabolism (Jones,

1963) ,\and amino a c i d s may be taken up by t h e c e l l even though they do -- - - - - - - - --
b
n o t support growth when suppl ied s i n g l y a s n i t rogen sources ( ~ o l a n ,
Pe te r s e t a 1 (1975) s t u d i e d t h e composition o f t h e f r e e amino
a c i d pool i n 1 0 s p e c i e s o f h i g h e r m a r i n e f u n g i w h i c h h a d b e e n g r o w n o n
yeas t e x t r a c t a s a n i t rogen s o q c e . They found t h e amino a c i d
composition o f a l l s p e c i e s t o be q u a l i t a t i v e l y similar, each '+
con ta in ing a l a n i n e , g l y c i n e , v a l i n e , p r o l i n e , l e u c i n e , i s o l e u c i n e ,
- + s e r i n e , threonine , h y d r a x y p ~ o l i n e , a s p a r t i c a c i d ,
a c i d , phenylalanine, o r n i t h i n e , l y s i n e , t y r o s i n e , t ryptophan, c y s t l n e ,
c y s t e i n e , and h i s t i d i n e . They specula ted t h a t m a r g e fung i may, supply, J
i;
hydroxyproline , a componbnt o f c o l
d e t r i t u s feeders i n t h e marine environment.
-, - '3
? 0 . ' The r e s u l t s sugges t t h a t 3. mariitimum did not
f i x a t i o n , d e n i t r i f i c a t i o n t o atmospheric nitrogen1, o r n i t r i f i c a t i o n . I.
N i t r i f i c a t i o n us ing an organic n i t rogen source was n o t i n v e s t i g a t e d , '-c
/
'. and i t i s the re fo re p o s s i b l e t h a t Zalerion cou ld+conver t a subs tance
such a s peptone t o amm&um, n i t r i t e , o r n i t r a t e . These
i n v e s t i g a t i o n s showed t h a t Z&&&Q did not grow anaePobica l ly , J
although t h e low oxygen c o n d i t i o n s c h a r a c t e r i s t i c o f a cand le jar d i d
oxygen irr-&Re-"exygwtllf R f t l t u ~ - h y e ~ r t n d + t m ~ i n e s e d f m r & q + b - -
9 C
e f f e c t o f v a r i o u s concen t ra t ions o f d issolved oxygen on growth o f
marine fungi would be a worthwhile a r e a o f f u t u r e i n v e s t i g a t i o n .

- -
The r e s u l t s from &g v i t r o studies do not n e c e s s a r i l y r e p r e s e n t
t h e phys io log ica l response of an organism i n its n a t u r a l environment.
Wood (1965, p 8) summarizes t h e problem i n t h e fo l lowing statement:
8'
"The s tudy of t h e physiology and biochemistry o f microorganisms i n pure c u l t u r e s 'is f raugh t with'many dangers, f o r microorganisms, l i k e t h e i r macro-counterparts, can be t r a i n e d even by t h e very * a r t i f i c i a l i t y o f t h e i r l a b o r a t o r y environmerit t o perform s t r a n g e t r i c k s which t h e y would scorn t o do i n t h e i r na t i r ra l s t a t e e n __ - - -- Lp-L--- --- --
Marine mycologists d v e n o t gt developed t h e techniques f o r in \
\ - s i t u i n v e s t i g a t i o n s o f n u t r i t i o n a l problem3 such a s n f t rogen -
u t i l i z a t i o n . If bne a t t e m p t s t o grow t h e organism o n ' i t s n a t u r a l - s u b s t r a t e , one is confronted wi th t h e problem o f n o t knowing t h e
chemTca1 co?posi t ion o f t h e growth medium. It is then impossible t o
. e
t e l l what n u t r i e n t o r combination o f n u t r i e n t s has produced t h e 9 1
observed growth response. , It is t h e r e f o r e necessary t o use a .
chemically de f ined n h t r i e n t s o l u t i o n fo&mparative s t u d i e s on
s p e c i f i c n u t r i e n t s . Such s t u d i e s i n d i c a t e whether t h e fungus b l e "
t o c a r r y o u t a given r e a c t i o n o r u t i l i z e a c e r t a i n n u t r i e n t . The
accur-acy with which such r e s u l t s can be ex t rapo la ted t o t h e marine /- #
environment depends upon how c i o s e l y cond i t ions o f c u l t i v a t i o n such. as
n u t r i e n t composition and n u t r i e n t concentra t ion parallel those o f t h e t C t
n a t u r a l s u b s t r a t e . It a l s o depends on whether t h e system under
i n v e s t i g a t i o n i s v a r i a b l e wi th r eapec t t o t h e ~ o n d i t i o n s of ppp- --- - -- --
by 2. maritimun i s - <-- 1
7--- - v a r i a b l e and dependent upon t h e composition of t h e growth medium
1

- a n d M e v e r s . - ~ a t t e r h s o f Qn the
o t h e r hand, can b e r e l a t i b e l y s t a b l e . Morton and MacMillan (1954) .
i
found t h a t q i t r o g e n n u t r i t i o n i f l S c o p u l a r i o p s i s b r e v i c a u l i s was no t
a f f e c t e d ' by presence o r absence ,- of agar , type of s t e r , l l i za&on o f
n u t r i e n t s , and presence o r absence o f vi tamins. They concluded t h a t . I
t h e i r n u t r i t i o n a l r e s u l t s r e f l e c t e d t h e t r u e phys io log ica l s t a t e o f i
The use of a chemical ly de f ined n u t r i e n t s o l u t i o n was necessary . J- 3.
, i n t h i s s tudy g i v k h e n u t r i t i o n a l n a t u r e of t h e problem. It is
hoped t h a t t h e u s e o f a r e l a t i v e l y d i l u t e n u t r i e n t s o l u t i o n , t h e use
- o f a continuous flow c u l t u r e system, and t* use of ion uptake 0 .
measurements gave a s u f f i c i e n t l y t r u e
a c t i v i t y t o al low a comparison between l abora to ry r e s u l t s and t h e + \
\ marine environment. - - - - -
i - f . L . , I . I
Zaler ion maritimum and o t h e r h igher marine fung i i n h a b i t ' /
cellulosic s u b s t r a t e s such as .wood and l eaves whi6h are submerged,
cont inuously o r i n t e r m i t t e n t l y , i n t h e sea . _These s u b s t r a t e s may be
found on t h e ocean bottom, i n thk i n t e r t i d a l zone, o r a s driftwood,.
deadheads, o r p i l i n g s . &J,erio~ has also' been i s o l a t e d from
marine sediments ( ~ G h n k , 1956). S t u d i e s to d a t e i n d i c a t e t h a t Z
2. W i r n u ~ p has 1
- - - &--- Campion-APsurnaY
\
3 - -- <
a l i m i t e d v e r t i c a l d i s t ~ i b u t i o n . Jones and Le 1
-- - -- --- -
( 1970) d id not i s o l a t e Z. below 126 m. s

and produces reproduct ive s t r u c t u r e s o n 6 s u r f a c i . Thus both . i %
seawater and t h e s u b s t r a t e a r e a v a i l a b l e t o t h e fungus-as a source qf
n u t r i e n t s .
i- d
The most,abundant rm'of n i t rogen i n seawater is molecular
- --- ni t rogen , which amounts t 20 tAe& - e -amouat&-ccmhine~itrl.ogeri 7-- - .
(Vaccaro, 1965). Combined n i t rogen can be present i n s e v e r a l forms,
t h e a v a i l a b i l i t y o f which i s d e ~ e n d e n t ~ u p o n b i o l o g i c a l a c t i v i t y . . In 4 1 - - -
'the open ~ c e a n , ~ c o m b i n e d n i t rogen i s p r i n c i p a l l y found as n i t r a t e 3- -
( 62%) , organic nitpogen (37%) , and ammonium and n i t r i t e ( 1%) (Vaccaro , 3
1965). I n coast* a r e a s , this r a t i o can be q t e r e d by planktonic 1, . blooms and o the r b i o l o g i c a l a c t i v i t y . I n su r face waters dur ing t h e
summer months, t h e ammonium concentra t ion may exceed t h e combined
- concentra t ions of n i t r a t e and n i t r L t e , An approximate f i g u r e for the P
amount o f n i t rogen a v a i l a b l e i n seawater is 450 p g / l (&t in , 1970) .'
, Concentrat ions o f combined n i t rogenAncrease with depth, due t o i
b i o l o g i c a l a c t i % i t y a t t h e sediment&mter i n t e r f a c e (Vaccaro, 1965).
*,
Nitrogen i n undecayed wood i s pr imar i ly p resen t as p ro te in .
Asparagine, glutamf ne , a l l a n t o f n , a l l a n t o i c a c i d , and c i t r u l l i n e w-4
( c l o s e l y r e l a t e d t o a r g i n i n e ) are a l l important i n n i t r o e e n s t o r a g e
(Kremers, 1963). Various p r o t e i n s are found i n t h e bark and, s i e v e - -
tube l i q u i d may c o n t a i n v a r i o u s amino ac ids . The ni t rogen c o n b n t - o f
mature wood is low; t h e propor t ion o f p&in v a r i e s from a s high as 1
20% i n young t i s s u e s t o less than-1% i n mature wood (Kremers; 1963).

** 'J. blocks i n d i c a t e s t h a t previous "digestl;;onn of wood n u t r i e n t s i s no t a
P
4
necessary p recursor t o fungal growth (Johnson and Sparrow, 1961).- '
However, i n t h e sea Z. maritimum is growing i n combination with o t h e r
decay--c*msing organisms such a s Urnnoria, Bankia, and marine b a c t e r i a , / t
It is poss ib le t h a t these o r g k i s m s could e x c r e t e n i t r agen c o n t a i n i n g L
I .
produc t s which .could be uwd - l t r y - ~ . - ~ l ~ a r i t i n u q - a n d _ e t h e c ~ funni- -_-- - + - . . - ~
According t o Ray (1959) been no nLtrogen-containing
excre to ry products -, which h a s no recognizable - -
excre tory organ. She suggested t h a t ~ i m n b r i a may u t i l i z e e f f i c i e n t l y
t h e very low n i t r o & concan t ra t ions i n wood.
It d s ' p o s s i b l e t h a t n a r i n e b a c t e r i a , which a r e numerous i n and *
above t h e sediments, may y o d d c e ammonim from t h e decay of wood and
theorganismsassocatedwith submerged woo&(Le, d i a t a m s , -
8/ protozoans, bladed a l g a e , Limnoria, .and Bankia). Concentrat ions o f -
ammonium a r e higher above t h e sediments than i n t h e rest o f t h e water
column, due t o t h e decompostion o f organic h a t e r i a l (Kriss, 1963). .
Both i s o l a t e s of 2. marit- grew oh n i t r a t e , n i t r i t e , ammonium,
and organic n i t rogen, ;hen suppl ied as s o l e n i t rogen sources. O f '?
greater i n t e r e s t was- tbe' preference shown f o r w i n o f t h e s e
most reduced form, amoniua , was preferred . Anmionitmi-was probably
used first because it can be d i r e c t l y incorporated i n t o cellular

. v o t e i n , .whereS n i t r a t e and n i t r i t e must first be enzymat ica l ly -.
.converted t@ ammokum. * 4 Severa l of t h e amino a c i d s t e s t e d served a s good n i t r o g e n sources
4
f o r Z, maritimum. b ike ammonium, glutamic a c i d was taken up from t h e
n u t r i e n t solut&n i n g r e a t e r q u a n t i t i e s than n i t r a t e . When a mixture
taken up simultaneously from t h e n u t r i e n t s o l u t i o n . We can conclude i i
t h a t t h e forms of n i t r o g e n most r e a d i l q u t i l i z e d by Z. maritirnuq were
ammonium and o rgan ic n i t rogen. I n t e r p r e t i n g t h e s e r e s u l t s i n terms of
t h e hab iQt o f z . maritimum and o t h e r marine f u n g i , I would sugges t
t h a t t h e fungus u t i l d z e d the, organic n i t rogen i n wood and t h e ammonium
produced a s an o rgan ic decay product. ~ m o n i u m would be t h e form o f
n i t rogen m q t a v a i l a b l e t o f u n g i i n sediments, because o f t h e reducing
* 3- \ I envi.ronment c r e a t e d by t h e low dissolved oxygen concen t ra t ion . fi
J I
would also ass&@ t h a t ammonium and d i s so lved organic n i t rogen , i f f . '
presentiJn seawater i n s u f f i c i e n t q u ? n t i t i e s , -would be absorbed by t h e
spores and mycelium on t h e s u r f a c e of t h e wood. 4 i
&
Zaler ion maritinurn can grow on low concentra t ion9 o f n i t rogen;
and a t low C:N r a t i o s . This f e a t u r e may i n d i c a t e an adap ta t ion t o u
growth on wood, a s C:N r a t i o s i n heartwood may va ry from 500:l t o
g r e a t e r than 1,250: 1 ( G r i f f i n , 1972). -Wood r o t t i n g terrestrial fung i
such a s folvoorus s r d c o l o r can adapt t h e i r mycel ia l n i t r o g e n c o n t e n t 2
* when grown on media low i n n i t r o g e n ( H e r r i l l a n d "Cowling, 1966). *
1

__ - - - - - __ - -- - o t h e r i n v e s t i g a t o r s have demonstrated t h a t marine fung i can
produce compounds which may be used a s food by marine animals. Kirk ---..
and Catalfomo ( 1970) demonstrated t h e presence o f e r g o s t e r o l add 4
cho l ine i n Z. maritimum and other .marine fungi , and suggested t h a t
marine fungi may d i r e c t l y supply v i t a m i n s f t o marine nematodes. Block .a
e t a 1 (1973) showed t h a t t h e mycelium of marine fung i contained h igh - _ - _ __ -_ -_ - _- - -- I-------
yieTds of f a t t y a c i d s , which could be important sources o f energy f o r
f i l t e r feeding beach animals. P e t e r s e t a; (-1975) suggested t h a t
hydroxyproline and o t h e r amino a c i d s found i n marine f u n g i may provide
n i t r o g e n t o d e t r i t u s feeding animals. 1
This stud* i n d i c a t e d t h a t Z. e i t h e r converted o r
exc re ted ni trogenous products which could poss ib ly be u t i l i z e d by
o t h e r organisms. Growth of Z. maritimum on n i t r a t e produced
measureable q u a n t i t i e s o f n i t r i t e ; growth on a mixture of amino a c i d s
r e s u l t e d i n t h e e x c r e t i o n o f two new amino a c i d s i n t o t h e n u t r i e n t
s o l u t i o n . Growth o f H. maritimum FMC on amino a c i d s and ammonium ,
i n d i c a t e d t h a t ammonium can l e a v e as w e l l a s e n t e r t h e fungal cell . tp
?
I suggest t h e fo l lowing hypo the t i ca l r o l e of Zalarion . i n the marine n i t r o g e n cycle : an a b i l i t y t o u t i l i z e n i t r a t e , n i t r i t e ,
which are a v a i l a b l e e i t h e r d i s so lved i n t h e ocean o r ino t he_ s u b s t r a W F -__-- - -- - - -- __-- --- -
an i n a b i l i t y t o n i t r i f y , d e n i t r i f y , o r f i x atmospheric n i t rogen; an
a b i l i t y t o exc re te o r conver t n i t r o g e n con ta in ing compounds which may

1 92
- - - - -
- *-- - --- -
3
possmy be Wed-by o t h e r m x n e o r g a n i s m s , an a b i l i t y t o s e r v e a s L
d i r e c t sources o f o rgan ic n i t rogen f o r animals feeding on fungal
myc elium .
SUMMARY
marine fung i , i n ord,er t o more c l o s e l y approximate t h e marine
environment than t h e batch c u l t u r e system. Growth o f
5. a l o p a l l o n e l l a , g. pe lag ica , and Z. maritimum was h igher i n t h i s
appara tus than i n ba tch c u l t u r e .
Development o f t h e continuous c u l t u r e system l e d t o an
' i n v e s t i g a t i o n o f growth dynamics i n ba tch c u l t u r e . I
I n ammonium
con ta in ing medium, a pH drop in batch c u l t u r e i n h i b i t e d growth o f a l l -
s p e c i e s t e s t e d . Improved pH contFol was obtained with MJ3S
(2- [N-~orphol inol e thanesu l fon ic a c i d ) . Chemical ana lyses o f
n u t r i e n t s showed t h a t phosphate but not glucose was exhausted from t h e
n u t r i e n t s o l u t i o n . A l l o f t h e ammonium and none of-.Me n i t r a t e w a s d
0
removed dur ing growth.
These i n v e s t i g a t i o n s should permit two improvements b f u t l l r a - i
al low the use of' a more d i l u t e c u l t u r e medium, and t h e n u t r i e n t uptake %
informat ion should a l low t h e formulat ion o f a more balanced n u t r i e n t
s o l u t i o n . I

source o r t h e or$snic n i t rogen source. It was concluded t h a t Z. L."
maritimum pre fe r red t h e n i t rogen sources a v a i l a b l e i n wood o r decaying. . organ ic m a t e r i a l (ammonium, o rgan ic n i t rogen) t o t h e n i t rogen sources
most commonly a v a i l a b l e ' in seaua ter (molecular n i t rogen , n i t r a t e ) .
1 Continuous and batch c u l t u r e systems were used t 0 s tudy n i t rogen
n u t r i t i o n i n Z . maritimum. Batch c u l t u r e r e s u l t s d i fSered - b
q u a l i t a t i v e l y and q u a n t i t a t i v e l p from continuous c u l t u r e r e s u l t s . The
preference shon) by L. maritimum f o r t h e organic n i t rogen source i n - r b
batch c u l t u r e was n o t sh6wn i n continuous c u l t u r e .
Zaler ion maritimum used .various inorganic and o rgan ic n i t rogen .
sources. When n i t rogen sources were added i n combination,
Z. maritimum p t h e most reduced inorgan ic n i t rogen -
The p a t t e r n o f n i t rogen n u t r i t i o n shown by this marine fungus was . s i m i l a r t o t h a t shown by many t e r r e s t r i a l fungi .
The experiments descr ibed i n t h i s t h e s i s i n d i c a t e d t h a t \ t
Z . maritimum did not f i x atmospheric n i t rogen, n i t r i f y , o r d e n i t r i f y - i no rgan ic compounds t o n i t rogen gas. Zalerion maritimum e x c r e t e d ;
n i t rogenous products such as n i t r i t e and amino' a c i d s , which could
conceivably be used a s n u t r i e n t s by o t h e r marine organisms.

LITERATURE - CITED - . -, .
1
.?
h n
Agniho t r i , V. P. and 0. V a a r t a j a . 1967. The i n l uence of n i t rogenous compounds on growth o f Pythium s p e c i e s . Can. J. Microbio l . 13: 1509-1519. -
Alderman, D . J. and E. B. G . Jones . 1971 . P h y s i o l o g i c a l recquirkments of two maf-ine Phycomycetes, A l t h o r n i a c r o u c h i t and
.Os t racoblabe implexa. Trans. Br. mycol. Soo. z: 213-225.
Anas tas iou , C. J. 1963. The genus Za l e r ion Moore & Meyers. Can. - - - s
s J. Bat. 4f: ?t3!7-1139.
Anas tas iou , C. J. and L. H. Churchland. 1969. Fungi on decaying L
l e a v e s i n marine h a b i t a t s . Can. J. Bot. a '251-257.
c Aube, C. and C. Gagqon, 7969. Effect o f carbon and o i t r o g e n n u t r i t i o n on growth and s p o r u l a t i o n o f Trichoderma v i r i d e Pe r s . ,
% . ", ex F r i e s . Can. J. Microbial. 13: 703-706.
Barghoorn, E. S. 1944. 11. B i o l o ~ i c a l Aspects , p. 434-467. I,n E. S. Barghoorn and D. H . L inder . Marine f u n ~ i : Their a& B ~ o ~ o Q ~ . Fa r lowia 1: 395-467.
+ Barr, D. J . S . 1969. S t u d i e s o'n Rhizophydium and Phlvc tochvt r ium ( C h y t r i d i a l e s ) . 11. Comparative physiology. Can. J. Bot. g: 999-1005.
Benemann, J. R . and R. C. Va l en t ine . 1972. The pathways o f \ n i t r o g e n f i x a t i o n , p. 590 t o 606. A. H. Rose and D. W. Tempest ( e d . ) Advances & MicrobiaA E b w i o l o a y , W. 8. Academic P r e s s , London and New York.
Bhargava, K . S. 1945. P h y s i o l o g i c a l s t u d i e s on some members o f , t h e f ami ly sgp ro l egn iaceae . 111. Nitrogen requi rements . J. I n d i a n Botan. Soc. a: 68-72. 5
\
t Block, J. H . , P. Catalfomo, G . H. Cons tan t ine , Jr., and P. W. , Kirk , Jr. 1973. T r i g l y c e r i d e f a t t y a c i d s o f s e l e c t e d h i g h e r marine f u n g i . Mycologia 43: 488-491.
Bremer, G. B. 1974. P h y s i o l o g i c a l responses o f some t h r a u s t o c h y t r i d furig;i, p. 237-~-t-o025 -0----- - . Q A. Gaer tne r (ed . ) Marine Mvkoloaie
, 1912- ~ ---
Byrne, P . J. and E. B. G. Jones . 1975a. E f f e c t o f s a l i n i t y on spo re germina t ion o f terrestr ia l and marine f u n g i . Trans. Br.
e: 497-503.'

Byrne, P . J. and E. B, G. Jmes, 1 9 - T 5 b 2 e & - o f s a l l n l t . . a- t h e r ep roduc t ion of terrestrial and marine f u n g i . Trans. B r . mycol. Soc. 65: 185-200.
,*
Catalfomo., P . , J . H. B lock , G . H. Cons tan t ine , Jr. , and P. W , r
. Kirk , J r . 1972-1973. C h ~ l i n e s u l f a t e (es ter) i n marine h ighe r f u n g l . Mar, Chem. 1: 157-162.
Ch i ld , J. J . , e. h a p p ; a n d D. I?. EveJeigh. 1973. Improved pH c o n t r o l o f f u n g a l c u l t u r e media.. Mycologia 65: 1078-1086.
' =
Churchland, L. M. and M. McClaren. 1973. Marine f u n g i i s o l a t e d from a Kraft pu lp m i l l o u t f a l l a r e a . Can. J; Bot. z>___ - - --
1703-1710.
Churchland, L. M. and M. McClaren. Growth o f f i l amen tous 'marine f u n g i i n a c o n t i n u o u s c u l t u r e system. Can. J. Bot. 54: - 893-8990 \
Cochrane, V. W. 1958. P h y s i o l o a o_f Fungi. John Wiley & Sons, New York, London, Sydney. 524 p.
Cove, D . J. and J. A. 9. Autoregula t ion o f t h e s y n t h e s i s o f i n A S p e r ~ i l l u S n i d u l a n s . J. Bact. 97: 1374-1378. - L
Davidson, D, E. 1974a. The e f f e c t o f s a l i n i t y on a h a r i n e and a f reshwater Ascomycete. Can. J. Bot. 52: 553-563.
Davidson, D . E. 1974b. Wood-inhabiting and marine f u n g i from a s a l i n e l a k e ' i n Wyoming. Trans. B r . mycol. Soc. a: 143-1 49.
Dayal, R . 1961. Ni t rogen requi rements o f some members o f t h e fami ly Sap ro l egn iaceae . Proc . Nat . Acad . Sci . I n d i a S e c t . B . ' 21: 332-336
Difco Manual o f Dehydrated C u l t u r e -Media and Reagents f o r $
Microb io log ica l and C l i n i c a l a t o r y procedures . Ninth ed i t i on , Di fco L a b o r a t o r i e s , i t . 350 p.
E y l a r , 0. R . , and E. I. ~ c h m i d t . 1959. A survey o f h e t e r o t r o p h i c microorganisms from s o i l f o r a b i l i t y t o form n i t r i t e and n i t r a t e . 6
J. gen. Pl icrobiol . 20: 473-481. - _i
F r i e s , L. 1956. S t u d i e s i n t h e physiology of Coprinus. 111. C u l t i v a t i o n exper iments w i t h r u n n i n g media. Bot. Notis= 109:
- -
12-20.

- - - - - - -
Gold, H. S. 1959. D i s t r i b u t i o n o f some l i g n i c o l o u s Ascomycetes and Fmg5 fn ipsr fee krs arr e s t u m . 5. Elf sha Mitchell s c i e n t .
Go lds t e in , S. 1963a. Development and n u t r i t i o n o f new s p e c i e s of Thraustochytr ium. Am. J. Bot . 50: 271 -279.
Go lds t e in , S. 1963b. Morphological v a r i a t i o n and n u t r i t i o n of a new monocentr ic marine7fungus. Arch. d i k r o b i o l . 45: - 101-1 10.
Go lds t e in , S. 1 9 6 3 ~ . S t u d i e s of a new s p e c i e s of Th*raustrochytrium tha t d i s p l a y s l i g h t s t i m u l a t e d growth. Mycologia 55: 799-81 1. -
' Golds te in , S c a ~ d H. BelskyT f964. >Axenic c 6 l t u p e s t u d i e s of a- new -
marine Phycomycete pos se s s ing a n unusual t y p e of a s e x u a l r ep roduc t ion . Am. J. Bot. - 51: 72-78.
Good, N. E . , G. D. Winget, W . Winter , T. N. Connolly, S. Izawa, , j,
', and R. M. M. S ingh. 1966. Hydrogen i o n b u f f e r s f o r b i o l o g i c a l r e s e a r c h . Biochemis t ry - 5: 467-477.
G o t t l i e b , D. 1971. Limited growth i n fungi* Mycologia Q: 619-629.
G o t t l i e b , D . , H. P . M o l i t o r i s , . a n d J. L. van E t t e n . 1968. V * Changes i n f u n g i w i t h age . 111. I n c o r p o r a t i o n o f amino a c i d s
>.a '2 i n t o c e l l s o f Rhizoctonia solani and Sclerotlum _ - ". + bataticola.
'I Arch. Mikrobio l . - 61: 394-398.
G r i f f i n , D. M. 197%. E c o l o q of S o i l F u n g i . - & & u s e U n i v e r s i t y P r e s s , L ib ra ry o f , ~ o n ~ r e c s ' . T 3 p. -
pfl > . ?
Grover, R. K . and J. S. S idhu . 1965. E f f e c t of n i t 9ogen s~&c&ls 'on t h e growth o f Pythium a p h a n i d e r m a t w YEdsors) F i t z . Sydowia 19: 231-237. I - -
Gustafsson , U. and N. E r i e s . 1956. ~ u t r i t i o n a l r equ i r emen t s of some marine f u n g i . Phys io log ia plantarum 2: 462-46,5.
Ha r r i son , J. L. and E. B. G . Jones . 1974. P a t t e r n s of s a l i n i t y t o l e r a n c e d i s p l a y e d b y z h e lower f u n g i , p. 197-220. In' A . Gaerbqer ( ed . ) Marine M y k o l o ~ i e 11, I n t e r n a t i o n a l S-.
e
'4
Mycoloaie, Bremerhaven, 1412. Ver'o'ff .. I n s t . Meeresforsch. +
Bremerh. Suppl . 5. i - 3
Hb'hnk, W. 1956. S t u d i e n z u r Brack-und Seewassermykologie V I . ijber d i e p i l z l i c h e Bes ied lung ve r sch i eden s a l z i g e r submerser S t a n d o r t e . ~ e r b . f f . I n s t . Meeresforsch. Bremerhaven 3: 195-213. C i t ed in: Johnson, T. W . , Jr. and F. K . Sparrow, Jr. Fungi % Oceans and E s t u a r i e s . J. Cramer, Weinheim. 668 P.

H o l l i m n , P . X - ~ n d DL L dennings, 1 3 - 7 7 n ~ a a ~ ~ - metabolism i n t h e fungus D e n d r v ~ h i e l l a s a l i n a . I. Changes i n t h e l e v e l s of s o l u b l e ca rbohydra t e s dur ing growth. New Phyto l .
: 569-582.
Hol l igan , P. M. and D. H. Jennings . 1972b. Carbohydrate metabolism i n t h e fungus D e n d r v ~ h i e l l a s a l i n a . 11. The i n f l u e n c e of d i f f e r e n t carbon and n i t r o g e n s o u r c e s on t h e accumulat ion o f manni to l and a r a b i t o l . New Phyto l . z: 583-594
Hood, D. W. 1963. Chemical Oceanography, p. 129-155. In H. Barnes ( e d . ) Oceanography and Marine Biologv, ~JI Annual re vie^,^ -
Vol. - '1. - Geor8e Al len & Unwin Ltd. , London. '
Hughes, G . C. 1969. Marine f u n g i from B r i t i s h Columbia: occu r rence and d i s t r i b u t i o n o f l i g n i c o l o u s s p e c i e s . S y e s i s 2: 121-1 40. d
Hughes, G. C. 1974. Geographical d i s t r i b u t i o n o f t h e h i g h e r marine f u n g i , p. 419-441. In - A. Gaer tne r ( ed . ) Marine Myk010~ie II,
k I n t e r n a t i o n a l Symp. Marine Mykoloaie, Bremerhaven, 1%. ~er'o'f f . I n s t . Meeresfor'sch . Bremerh. Suppl . 5. -
Hughes, G. C. 1975. S t u d i e s o f f u n g i i n oceans and e s t u a r i e s s i n c e .1961. 1 . L ign ico lous , c a u l i c o l o u s and f o l i i c o l o u s s p e c i e s . Oceanogr. Mar,. B i o l . Ann. Rev. 12: 69-180. .
t Hultman, E. 1959. Rapid s p e c i f i c method f o r de t e rmina t ion o f
a ldosaccha r ides i n body f lu idas . Nature 183: - 108-109.
Hunter , D . R . and I . - H . Sege1.+971. Acidic and b a s i c amino a c i d t r a n s p o r t systems o f P e n i c i l l i u m chryso~enum. Arch. Biochem. Biophys. - 144: 168-183.
Johnson, T. W . Jr. and H. S. Gold. 1959. A system f o r - cont inua l - f low sea-water c u l t u r e s . Mycologia 51: 89-94.
Johnson, T. W. Jr . , H.' A . Ferchau, and H. S. old. 1959. I s o l a t i o n , c u l t u r e , growth, and n u t r i t i o n o f some l i g n i c o l o & marine fung i . Phyton g: 65-80.
Johnson, T. W . Jr. and F . K . Sparrow, Jr. 1961. FUU& PEeBa8 E s t u a r i e s . J. Cramer, Weinheim. 668 p. - -- - --
Jones , D. H. t965. The e f f e c t o f n i t r o g e n and phosphorus compounds . on one o f t h e microorganisms r e s p o n s i b l e -- - f o r - s ludge bulking.- -- A -
PToc. Ind. Waste-conf. ~ u r d u e Univ. 2_0: 297-315. 9

Jones , E. B. G . 1957-58. A p r e l i h n a r y s tudy , l e a d i n g t o an inu_est+gatiO~~of t h ~ & Y % & of ~~P&&MBES&RS k t f f g i , - o f f - W t h e s e a . Bachelor o f Sc ience Thesis : Univ. College o f Wales, Aberystwyth. 70 pp. c i t e d in: Johnson, T. W. J r . and F. K. Sparrow, Jr. 1961. Fungi &I Oceans E s t u a r i e s . J. Cramer, Weinheim. 668 p.
Jones, E. B. G . , P. Byrne, and D. J. Alderman. 1971. The , , . .
response o f f u n g i t o s a l i n i t y . Vie Y l i e u , Suppl . 22 : 265-280. I - 1
J o n e s , E. B. G. and D. H. Jennings . 1965. The e f f e c t o f c a t i o n s on t h e growth o f f u n g i . New Phytol . Q: 86-100.
I
/ Jones, 0 . T. G. 1963. The accumulat ion o f am_inp a i d s by f u n g i , . - - - -
. w i t h p a r t i c u l a r r e f e r e n c e t o t h e p l a n t p a r a s i t i c f h g u s Wtr- faba.e. J. Exp. Bot. 14: 399-411.
Kinsky, S. C. 1961. Induc t ion and r e p r e s s i o n o f n i t r a t e r e d u c t a s e i n Neurospora c r a s s a . J. Bact. 82: 898-904. <
A K i r k , P. W. Jr . and P, Catalfomo. 1970. Marine fung i : t h e occurrence o f e r g o s t e r o l and .chol ine . Phytochemistry 4: 595-8597.
Kohlmeyer, 'J. 1974. On t h e d e f i n i t i o n and taxonomy of h ighe r marine f u n g i , p. 263-286. I n - A . Gaer tner ( ed . ) Marine Mykologie 11, I n t e r n a t i o n a l .Symp. Marine Mykologie , Bremerhaven , 1972.. Verb'ff. I n s t . Meeresforsch. Bremerh. Suppl. 5. 8.
Kremers, R . E . 1963. The o h e m i s t r v ' o f developing whod, p. 369-404. L Browning, B. L*.- (ed . m j ~ of W d . I n t e r s c i e n c e .
Publ ishers , New York and London.
Kriss, A. E. 1963. k r i n e Mic rob io lom (Deep Sea ) . T r s c i e h c e P u b l i s h e r s , John Wiley and Sons, New York. 536
Kubitschek, H. E. 1970. I n t r o d u c t i o n t o Research w i t h Continuous Cu l tu re s . P ren t i ce -Ha l l , Englewood C l i f f s , New Je r sey . 195 p.
L inder , D. H. 1944. 1. C l a s s i f i c a t i o n of t h e marine f u n g i , p. 401-433. &I E. S. Bapghoorn and D. H. Linder . Marine f ' u n ~ i : Thei r Taxonomy a& Bioloav. Far lowia 1: 395-467.
Lowe, D. A . and D. H. Jennings . 1975. Carbohydrate metab t h e fungus g e n d r v ~ h i e l l a s a l i n a . ~ ' p h e p a t t e r n o f . i z - - a r a b i t o l and po lysaccha r ide after growth i n t h e presence o f s p e c i f i c a l l y l a b e l l e d carbon sources . - --- New Phytol: a: 6 E 2 9 ,

Lyman, J. and R. H. Fleming. 1940. Composition o f s e a water . J. Mar. Res. A 3: 134-146.
Mandels, G. R . 1965.. K ine t i c s o f fungal growth, p. 599-612. I_n G . C. Ainsworth and A . S. Sussman (ed . ) T& Fungi: an Advanced Trea t i se . V J . L, Tle u. Academic Press , New York and London.
Martin, D. F . 1970. Marine Chemistry, V T l . 2. Theory a> ADDlicationS. Marcel Drekker Inc . , New York. 451 p.
P
M e r r i l l , W. and E. B. Cowling. 1966. Role of n i t rogen i n wood
Phytopathology 56: 1083-1090. d e t e r i o r a t i o n : amount
Meyers, S. P. 1966. V a r i a b i l i t y i n growth and reproduct ion o f t h e marine fungus Lulworthia f l o r i d a n a . ~ e l g o l ' a n d e r w i s s . meersunters., 12: 436-443.
\
Meyers, S. P. and L . Hoyo. 1966. Observations on t h e growth o f t h e marine Hyphomycete Varicosporina r a m u l o s ~ . Can. J. Bot. 44: 1133-1 140. - h r
Meyers, S. P . , B. P r i n d l e , and E! S. Reynolds. 1960. C e l l u l o l y t i c a c t i v i t y of marine fungi . Degradation of l igno-ce l lu lose m a t e r i a l . Tappi u: 534-538.
Meyers, S. P. and E. S. Reynolds. 1959. Growth and c e l l u l o l y t i c a c t i v i t y of l i g n i c o l o u s Deuteromycetes from marine l o c a l i t i e s . Can. J. Microbiol. 2: 493-503.
.Meyers, S. P. and E. S. Reynolds. 1960. Occurrence o f l i g d i c o l o u s fung i i n northerfi A t l a n t i c and P ~ c i f i c marine l o c a l i t i e s . Can. J. Bot. - 38: 217-226.
Mitra, S. K . and R. N. Tandon. 1970. Nitrogen requirements of t h r e e s p e c i e s o f P e s t a l o t i a . Myc-opath. Mycol . appl . 42: 9-1 6. - J
M o l i t o r i s , H. Pi 1974. AlterungsvorgXnge bei P i l z e n , p. 475-508. In A. G a e r b e r (ed . ) Marine Mykologie 11, I n t e r n a t i o n a l $ 3 .
Marine Mykologie , Bremerhaven, I=. Ver'df f . I n s t . Meeresforsch. Bremerh. Suppl. 5.
. , Morton, A. G. and A. MacMillan. 1954,- The a s s i m i l a t i o n o f
n i t rogen from ammonium s a l t s and n i t r a t e by fung i . J. Exp. Bot. 2: 232-252.
-

- 100 ,
4
J - - - - - - - - 2
4 'L Ng, A . M. L . , J. E. Smith, and A . 3 . McIntosh. 1975. \
Conidiat i6i i 07 A s p e r g i T I u S n i a e F i n c5ntinuous c u l t u r e . Arch. t Mikrobiol. @: 119-126. s .
Nicholas, D. J. D. 1965. U t i l i z a t i o n o f inorganic n i t rogen compounds and amino a c i d s by f u n g i , p. 349-376. I& G. C. Ainsworth and A . S. Sussman ( e d ) . The F u n ~ i : an Advanced T r e a t i s e , V J l 1. The Fungal Q&. Academic Press , New York and London. +9
I ,-
Nicholas, D. J , D,and J. H. Scawin. 1956. A phosphate b r e q u i r e m e n t - f o r - n i t r a t e r educ tase from Neurospora c r a s s a . Nature 178: 1474-1475. -
(2 J Nolan, R. A. 1970. Th Phycomycete C a t e n a r b . Growth - requljrements. J. gen. Microbiol. a: 16
r
peter^,^ J. E . , P. Catalfomo, G . H. Co t t i n e , Jr., v . w,, Kirk, Jr. 1975. Free amino a c i & p higher marine f'ungi. J. Pharm. S c i . 6A: 176-177; > -
Postgate , J. R . 1972. The ace ty lene reduct ion t e s t f o r .ni trogen f i x a t i o n , p. 343-356. In J; R . 'Norris and D. W. Robbins (ed . ) Methods i n - Microbiology, VA. @. Acadzmic Press , London and New York.
Quevedo, R. A . , P. L . Sguros, and W . R. ~homann. 1973. Removal of cy 1 lodiene p e s t i c i d e s from seawater medium by a f i lamentous I
marine fungus. Abst. Am. Soc. Hicrobiol . Q: 184 only. - Ray, D. L. 1959. N u t r i t i o n a l physiology i n Limnoria, p. 46-60. &
D . L. Ray (ed . ) Marine Boring a d Fouling Or~an i sms . S e a t t l e , Univers i ty of Washington Press . 1
\
Ri tch ie , D. 1957. S a l i n i t y optima f o r marine fungi a f f e c t e d by temperature. Am. J. Bot. 44: 870-874.
R i t ch ie , D . and M. K . Jacobsohn. 1963. The e f f e c t s o f osmotic and n u t r i t i o n a l v a r i a t i o n on growth o f a s a l t - t o l e r a n t fungus,
- Zalerion e i s t l a , p. 286-299. C. H. Oppenheimer (ed. ) Symposium Marine ~ r o b i o l o p v . C. C. Thomas, S p r i n g f i e l d ,
I I l l i n o i s .
Robinson, J. H. , & Anthony, and W . T. Drabble. 1974. The u t k l i z a Won o f nitogenToiiFmsTyAsierni1lus cIavatus . 3 . Gen. Microbioq B: 23-28.
Sa id , H. and A. W. Harhash. 1966. Comparative s tudy of t h e e . f fec t o f n i t r a t e and amino ac id-ni t rogen on t h e r e s p i r a t i o n , ke toacid production alnd n i t r o g e n metabolism of F u s a r i q g x v a ~ o r u . Acta. b i o l . med. german 12: 135-144.

Mucorales. V. 1. U t i l i z a t i o n o f . d i f f e r e n t n i t rogen compounds.
- + . Path. Microbial. 28: 816-829. '
Schaumann, K . 1974. Experimentel le Untersuchungen zum ~ i n f l u h des S a l g e h a l t e s und deh,Teqpera tur .auf das Mycelwachstum hb'herer P i l z e aus dem ~ e e r - & h . ~ a c k w a s s e r , p. 443-474. A. Gaertner ( ed . ) Marine Mycologie 12, .In te rna t iona l Symp. Marine M y c o l o ~ i e , Bremerhaven, 1972. ~ e r b ' f f . I n s t . Meeresforsch. Breqerh.
. Suppl. 2: 443-474. 42 4
t
Sguros, P. L . and J. Simms. 1 9 6 3 . x o l e of marine fungi i n the
n i t rogen , and . TRIS( hydroxyme t h y l ) aminomethane on growth and pH changes i n s y n t h e t i c media. Mycologia 55: 728-741.
a f
Sguros, P. L. and J. $ i m m s . 1964. Role o f marine fung i i n tkie biochemistry o f t h e oceans. I V . Growth responses t o seawater /I inorganic macroconst i tuents . J. Bact. @: 346-355.
Sguros, P. L . , S. P.'Meyers, and J. Simms. 1962. Role of marine fungi i n the biochemistry of t h e oceans. I. Establishment of q u a n t i t a t i v e t echn iq f o r c u l t i v ~ t i o n , growth measurement and
4
! Y production of inocula . Mycologia 2: 52 1-535.
Sguros, P. L . , J. Rodrigues, and J. Simms. 1973. Ro.le of marine - - - f fungi i n t h e biochemistry o f t h e oceans. V. ~ a < t e r n s oC c o n s t i t u t i v e n u t r i t i o n a l growth responses. Mycologia 62: 161-1747--
S h r i n e r , R. L . , R . C. Fuson, and D. Y. Curtin. 1964. T h e Systematic I d e n t i f i c a t i o n of O r a a n i ~ m, A LaboratnrP
# w. John Wiley & Sons, Inc . , New York, London, Sydney.*458 P . -
Siegen tha le r , P. A., M. M. e el sky, S. Goldstein, and M;. Menna. 1967. Phosphate uptake i n an o b l i g a t e l y marine fungus. 11. Role of c u l t u r e c o n d i t i o n s , energy sources , and i n h i b i t o r s . J. Bact. s: 1281-1288.
i
Singh, B. P. and R. N. Tandon. 1970. Nitrogen requirements o f c e r t a i n i s o l a t e s o f A l t e r n a r i a tenuis,Auct. Mycopath. Mycol. appl . Q: 33-38.
- - - - -- -- - - - - -- -
Smith, I. 1960. C h r o m a t o ~ r a ~ h i q & ElectroDhoretic m. In t e r s c i e n c e Pllblishers, N ~ K ~ Q & . _ ~ L ' L p - - - - - - - - - -
Steens land, H. 1973. Continuous c u l t u r e of a sewage fungus Fusaflum aquaeduc tum. Arch. Mikrobiol . 93 : 28.7-294. -

s t ephen , R. C. and C. Chan. 1970. Ni t rogenwequi rements o f ' t h e . g e m s L i n d e r i m r i 3 k r . J. b t . ' 4 8 : - 6% -oyo. '^"
J , S t r i c k l a n d , J. D.. H. Snd T. R . Parsdins. 1972. A Y r a c t i c a l B
Handbook of Seawater Analys is . F i s h e r i e s e s e a r c h Board o f Canada B u l l e t i n #167, Ottawa. 310 /
i - t u b a k i , K . 1966. Marine f u n g i from Japan. Lignico lous I. Trans.
mycol. Soc. Jap. 7: 73-87. - .. , + %
cca ro , R. F. 1965 .'+-inorganic n i t r i g k h n seawater , p. 365-408. J. Pi Riley. and G . Skinrow (ed. ) Chemical Oceanography, V A .
1. Academic P r e s s , London and ~ e w York. t
- - ' *
'vembu, D. and P. L. Scprps . 1972. C i t r i c a c i d c y c l e and -. glyoxyl ,a te '~ypass %n glucose-grown f i l amen tous marine fung i .
i Bact . Proc'. 72: 166 only.
- ' Vishniao, H. S i 1955. The morphology and n u t r i k i o n o f a new s p e c i e s
of S i ro lv id ium. Mycologia 4 4 633-645. . t '.
Wood, E. J. F. 1965. Marine Micrpgia l Ecolbgy. ~ h a p a a n and Hall, London.; Reinhold , . ~ e w York, 243 p , ? I .
I * V
~ u $ e f , H. M. and H. E. Allam.)967. The carbon and, n i t r o g e n D n u t r i t i o n of c e r t a i n f u n g i . Can. J. M i c r o b i d . Q:
1097-1 106. 1 2
Zalokar , M. 1961. Kinet.icS o f amino a c i d uptake and rote in s y h t h e s i s i n ' ~ s u r o s m r a . Biochim. ~ i o & & . keta: 44: 423-4 32. C
Zobe l l , C . E. 1946. b r i n e u c r o b i o l o m . Chronica Botanica Co., I n t . P l a n t Sc i epce P u b l i s h e r s , Waltham, kss. \

Name L e s l i e Marian Churchland \
Place and year l x x Vancouver, B r i t i s h Columbia, 1945. of b i r t h
Education Univers i ty of B r i t i s h Columbia, B. A. ( F i r s t Class Honours), Botany, 1966.
Experience
. - Simon F r a s e r Univers i ty , M. 5 c . (Biology) ,
- - A - - - - - ,- A - - - A - - - - A A A A - - 1971. Graduate %;dies, PhD program, 1971-1976. Y
McGill Univers i tv . Research a t McGill Un ive r s i ty , 1973. On leave from Simon F r a s e r Universi ty. r
Univers i ty of B r i t i s h Columbia, Botany , Department. 1963-1966. P a r t - L h e
r e s e a r c h a s s i s t a n t i n Mycology, Marine Mycology . 1966-1969. Research a s s i s t a n t ( f u l l t ime) i n ' ~ a r i n e Mycology, Mycology, Protozoology.
Simon Frase r Univers i tv , Biology Department. 1969-1975. Graduate s t u d i e s i n Marine Mycology. Teaching a s s i s t a n t s h i p s i n In t roduc to ry Biology,.Biology of t h e Populat ion ' ~ x p l o s i o n , Non-vascular P l a n t s , Microbiology, Marine Biology. "
u n i v e r s i t y Qf m, Friday Harbor Labora tor ies . Summer, 1972. Teaching a s s o c i a t e , Botany 565b (Marine Mycology).
McGill -. 1973. On l eave from Simon Frase r Universi ty. Independent reseaPch i n Mycology labora tory . Fall,. 1973. Laboratory a s s i s t a n t ,
- - - - - - - - -- - -- - - - - - -- - - --- - - -- - -- ~ n a s g a s i o u , C. J. and L. M. Churchland. 1968. An O l ~ i d i o ~ s i a
. . p a r a s i t i c on a marine fungus. Syes i s 1: 81-85.
- An-stasi6~1, C: J.. and I;. M.' churchland. 1969'. Fungi on decaying l e a v e s , i n marine hab i t a t s . . Can. J. Bot. . . : 2517257. '

Churchland, L L K and M. MhClaren. 1972. The e f f e c t of - - Kraft pulp m i l l e f f l u e n t s on t h e growth o f Zaler ion maritimum. Can. J. Bot. 50: 1269-1273.
Churchland, L. M. and M. E c ~ l a r e n . '1973. Marine ?u&i ,
i s o l a t e d from a Kraft pulp m i l l a r ea . Can. J-. /
Bet. 3: 1703-1710.
Churchland, L. M. and. M. ' M c ~ l a r e n . 1976. Growth o f f i lamentous marine fung i i n a continuous c u l t u r e
,system. Can. J. Bot. 59: 893-899. - LA--
- --- -- Presented p a w s "Groi?€h and Nut r i t ion of - ~ i l % e n t o u s
Marine Fungi." Presented to : P l a i n s \
~ n i v e r s i ' t i e s Biologica l Seminars.
1 Regina, February 19-22, 1976.
Membership i n MycologicaI Socie ty o f America Profess ional S o c i e t i e s 1971-1976.
Scholarships and Awards
B. C. Government Scholarship , 1962,1966. *
S . I t a l i a n Government Book P r i z e , 1962-1966.
National Research Council of Canada Postgraduate Scholarship , 1969;1973.
5. Simon Frase r Univers i ty Graduate Scholarship,, 1975.
t





























