Embed Size (px)
Citation preview

Broad Bean Mottle Virus
National Diagnostic Protocol
Angela Freeman
Department of Primary Industries
Primary Industries Research Victoria, Horsham.
May 2006
ACKNOWLEDGMENTS
Plant Health Australia funded the project to develop this manual as part of their National Diagnostic Protocols Initiative. Dr Safaa Kumari (ICARDA) supplied many of the photographs and the protocol for TBIA, which was adapted for this manual.
DISCLAIMER
The scientific and technical content of this document is current to the date published and all efforts were made to obtain relevant and published information on the pest. New information will be included as it becomes available, or when the document is reviewed. The material contained in this publication is produced for general information only. It is not intended as professional advice on any particular matter. No person should act or fail to act on the basis of any material contained in this publication without first obtaining specific, independent professional advice. Plant Health Australia and all persons acting for Plant Health Australia in preparing this publication, expressly disclaim all and any liability to any persons in respect of anything done by any such person in reliance, whether in whole or in part, on this publication. The views expressed in this publication are not necessarily those of Plant Health Australia.

2
Contents
1.0 Introduction
2.0 National Diagnostic Protocol Procedure
2.1 Purpose and scope of diagnostic protocol
2.2 Responsibility
2.3 Procedure
2.4 Documentation
2.5 Records
3.0 Pest Risk Analysis
3.1 Background
3.2 Species name
3.3 Synonyms
3.4 Common names
3.5 Host Range
3.6 Distribution
3.6.1 Australian status
3.6.2 Current distribution
3.6.3 Potential distribution in Australia
3.7 Plant parts affected
3.7.1 Vegetative
3.7.2 Seedborne
3.8 Disease features
3.9 Biology
3.9.1 Identification
3.9.2 Virus strains
3.9.3 Serological relationships
3.9.4 Symptoms
3.9.5 Disease cycle
3.9.6 Dispersal
3.10 Assessment of likelihood
3.10.1 Entry potential
3.10.2 Host range potential

3
3.10.3 Establishment potential
3.10.4 Spread potential
3.11 Overall entry, establishment and spread potential
3.12 Assessment of consequences
3.12.1 Economic impact
3.12.2 Environmental impact
3.12.3 Social impact
3.13 Combination of likelihood and consequences to assess risks
3.14 Surveillance
3.15 Diagnostics
3.16 Training
3.17 References
4.0 Diagnostic protocol
4.1 The diagnostic test/s and diagnostic sequence
4.2 The initial samples
4.2.1 Sample handling and subsampling
4.2.2 Sample storage
4.2.3 Visual symptoms
4.2.4 Documentation
4.3 Further samples
4.3.1 Sample collection, transport and storage
4.3.2 Sample locations
4.4 Confirmation of diagnosis
5.0 Identification of pathogen (primary diagnostic test)
5.1 Enzyme-linked immunosorbent assay
5.1.1 Introduction
5.1.2 General items required
5.1.3 Specific items
5.1.4 Buffer recipes
5.1.4.1 Coating Buffer
5.1.4.2 Phosphate Buffered Saline
5.1.4.3 Wash Buffer
5.1.4.4 Extraction Buffer for DAS ELISA

4
5.1.4.5 Conjugate Buffer
5.1.4.6 Substrate Buffer
5.1.4.7 Extraction Buffer for indirect ELISA
5.1.5 Methods
5.1.5.1 Direct double antibody sandwich (DAS) ELISA
5.1.5.2 Indirect ELISA
5.2 Tissue blot immunoassay (TBIA)
5.2.1 Introduction
5.2.2 General items required
5.2.3 Specific items
5.2.4 Buffer recipes
5.2.5 Methods
5.2.6 Reagent suppliers
6.0 Confirmation of diagnosis
6.1 Electron microscopy
6.1.1 Introduction
6.1.2 General items required
6.1.3 Methods
6.1.3.1 Sap dip (negative staining) method
6.1.3.2 Immunosorbent electron microscopy
6.1.3.2.1 Trapping method
6.1.3.2.2 Decoration method
6.2 Light microscopy
6.2.1 Introduction
6.2.2 General items required
6.2.3 Method
6.3 Indicator plants
6.3.1 Introduction
6.3.2 General items required
6.3.3 Method
6.3.4 Buffer recipes
6.3.4.1 Inoculation buffer
6.3.5 Indicator plant species and reactions
7.0 Images

5
7.1 BBMV symptoms on host plants
Figure 3. BBMV symptoms on faba bean
Figure 4. BBMV symptoms on faba bean
Figure 5. BBMV symptoms on faba bean
Figure 6. BBMV symptoms on faba bean
Figure 7. BBMV symptoms on lentil
Figure 8. BBMV symptoms on french bean
7.2 BBMV symptoms on host seed
Figure 9. BBMV symptoms on faba bean seed
7.3 BBMV vectors and vector symptoms
Figure 10. The BBMV vector Sitona lineatus
Figure 11. The BBMV vector Sitona lineatus
8.0 References and websites
8.1 References
9.0 Appendices
Appendix 1. Preliminary Information Data Sheet (Plantplan, 2004)
Appendix 2. Hygiene

6
List of Figures
Figure 1. Flow chart of the basic procedure and responsibilities of the
relevant Departments if a suspect sample is received.
Figure 2. Flow chart of protocols for the diagnosis of suspect BBMV-
infected samples.

7
1.0 Introduction
Broad bean mottle virus (BBMV) (Bromovirus, Bromoviridae) is one of a number of viruses
which are seedborne in a range of temperate pulses and has been found in Africa, Asia,
Europe and the Middle East. The virus is seedborne in Cicer arietinum (chickpea), Pisum
sativum (field pea) and Vicia faba (faba bean, broad bean, tick bean) (Fortass M, Bos L
1992).
BBMV was first described by Bawden et al. (1951) from a severely infected broad bean
(Vicia faba) crop in Nottinghamshire, England. Three quarters of the plants displayed leaf
symptoms, many were dwarfed and produced few or no flowers. Infected plants were in
concentric patches, with the most severely infected in the centre, suggesting the virus was
spreading in the crop from randomly distributed initially infected plants. BBMV was then
found in a faba bean crop in Cambridge in 1957 but was not found elsewhere despite
surveys (Gibbs AJ, 1972). The virus was seen as being of minor importance until reports of
its widespread occurrence began to appear in the 1970s and 1980s. BBMV was reported in
faba bean crops in Portugal (Borges M, Louro D 1974), Sudan (Murant et al. 1974), Morocco
(Assou NM 1978), and Algeria (Ouffroukh A 1985). Makkouk et al. (1988a) then undertook a
regional survey and found BBMV in faba bean crops in Egypt, Morocco, Sudan, Syria and
Tunisia. Fortass and Bos (1991) surveyed faba bean crops in Morocco and found that
luteoviruses and BBMV were the most prevalent viruses. Fortass and Diallo (1993) surveyed
a range of legume crops in Morocco for BBMV and reported natural infection of chickpea,
lentil, pea and common bean. They considered that BBMV was an actual threat to a range of
food legume crops and improvement programs.
BBMV has now been established as having a wide host range among the legumes (Makkouk
et al. 1988a) and is spread by a range of beetle vectors. The symptoms of BBMV can be
confused with other legume viruses which cause mottle/mosaic symptoms eg broad bean
stain virus (BBSV), broad bean true mosaic virus (BBTMV) and bean yellow mosaic virus
(BYMV). Like BBSV, BBMV can affect seed quality by causing necrosis and shrivelling of the
seed. Seed transmission rates in legumes are low and range from 0.1% to 1.4%. Various
strains of BBMV have been distinguished on the basis of slight differences in host range and
symptomology ranging from almost symptomless to severe, but they have generally been
found to be serologically indistinguishable.
BBMV survives between growing seasons of the primary pulse host in an alternative host or
in infected seed. Due to the fact that the natural host range includes at least four families
(Brunt et al. 1997, Makkouk et al. 1988a) and includes many food legumes and non-legume

8
wild hosts, it may survive in a range of alternative hosts. Carryover in infected seed is a likely
method of survival of the virus and Makkouk et al. (1988a) and Fortass and Bos (1991)
suggest that seed transmission in faba beans and other legume crops would explain the
widespread occurrence of BBMV in West Asia and North Africa.

9
2.0 National Diagnostic Protocol Procedure
2.1 Purpose and scope of diagnostic protocol
The purpose of this manual is to provide a nationally accepted, standardised protocol for the
accurate detection of broad bean mottle virus (BBMV) in temperate pulses. BBMV is a
quarantinable pathogen in Australia and is routinely tested for in the post-entry quarantine
program at the DPI Temperate Pulse Quarantine Station, Horsham, Victoria, using ELISA.
The manual is designed for easy access to the relevant sections required to identify the
pathogen. The manual contains the Pest Risk Analysis for BBMV for Australia, the primary
diagnostic protocols (ELISA) and tissue blot immunoassay (TBIA) and secondary
confirmatory methods (light and electron microscopy, indicator plant tests), images of virus
symptoms on host plants and seeds, and references and appendices.
2.2 Responsibility
Figure 1 shows a flow diagram of the responsibilities and procedures required when a
suspect sample is received. The responsibilities are also listed quite clearly in the
following points:
A: State/territory agriculture departments receiving suspect plant sample:
Receiving scientists will record details of the sample so that a trace back can occur if
required.
Receiving scientists will examine the sample and provide diagnostic services (in this
case, conducting the serological tests ELISA and/or TBIA) to identify the pathogen.
Receiving scientists will notify the State Quarantine Authority (eg. DPI-Victoria Plant
Standards Branch) of the suspect sample.
The State Quarantine Authority will examine the evidence and inform the Office of the
Chief Plant Protection Officer (OCPPO) and AQIS and advise scientists of required action.
The State Quarantine Authority will participate in the Consultative Committee on Exotic
Plant Pests and Diseases (CCEPPD), chaired by the Chief Plant Protection Officer and
decisions made and actions required will be passed onto state scientists for action.
Scientists may be requested to provide expert advice to the CCEPPD.
Scientists will conduct a second type of diagnostic test (secondary confirmatory test) as
advised by the State Authority.
Scientists will send part of the sample to the interstate confirmatory laboratories for
repeat of the primary diagnostic test as advised by the State Authority.

10
Under direction from the State Authority, state scientists will undertake delimiting surveys
if required and undertake diagnostics on survey samples.
The State Authority will liaise with industry representatives.
The State Authority will develop communication strategies in conjunction with the
CCEPPD.
The State Authority will report to all interested parties (OCPPO, CCEPPD, AQIS, national
bodies and industry) as required.
The State Authority will keep up to date with the processing of the suspect sample and
will notify the clients of the final result and the corresponding decision for that result.
The State Authority will handle all correspondence with clients. This is very important
and is to be made clear to other personnel involved with handling the sample that they are
not to correspond with the client.
B: Interstate agriculture departments
Scientists will re-examine the suspect sample.
Scientists will repeat diagnostic tests and confirm diagnosis.
Scientists may be requested to provide expert advice to the CCEPPD.
State Quarantine Authority will inform the Chief Plant Protection Officer and the CCEPPD
and will implement their decisions.
C: Office of the Chief Plant Protection Officer (OCPPO)
OCPPO will convene the CCEPPD and all decisions regarding the steps involved in
handling and diagnosing the original sample will be made by the committee.
The CCEPPD will determine whether or not the incursion requires a national response or
involves only one state and will determine the need for delimiting surveys.
Information from each state will be provided to the CCEPPD to enable national decisions
to be made.
OCPPO will provide media releases to the public and interested parties.
OCPPO and the CCEPPD will determine whether or not the pathogen can be eradicated,
contained or will be declared endemic.
2.3 Procedure
Figure 2 shows the order of steps /procedures to be undertaken in the diagnostic process in
a flow diagram.

11
2.4 Documentation
An electronic and a hard copy of this manual are maintained by the Senior Virologist, Primary
Industries Research Victoria (PIRVic), Dept. of Primary Industries-Horsham, Victoria and
PHA.
2.5 Records
The Recording sheets contained in Appendix 1 must be copied and filled in as appropriate
for each sample received and kept together in a file marked “Suspect broad bean mottle
virus samples”.
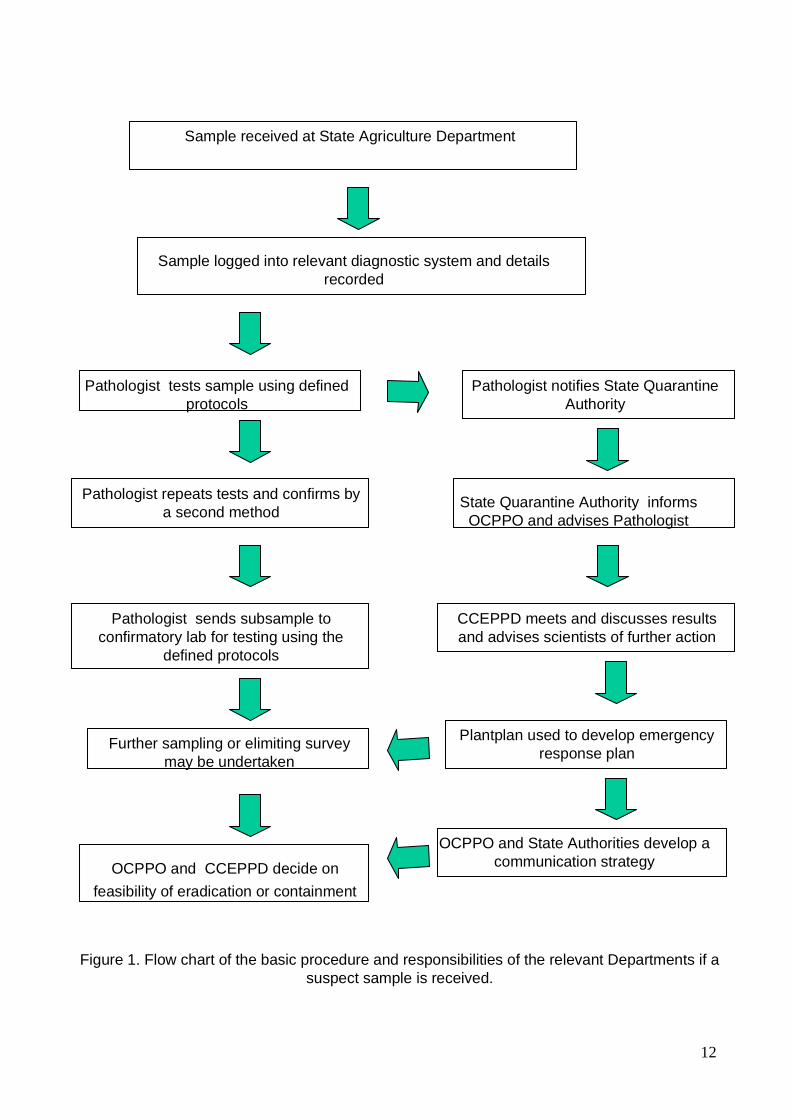
12
Sample received at State Agriculture Department
Figure 1. Flow chart of the basic procedure and responsibilities of the relevant Departments if a
suspect sample is received.
Sample logged into relevant diagnostic system and details
recorded
Pathologist tests sample using defined
protocols
Pathologist notifies State Quarantine
Authority
State Quarantine Authority informs
OCPPO and advises Pathologist
Pathologist repeats tests and confirms by
a second method
Pathologist sends subsample to
confirmatory lab for testing using the
defined protocols
CCEPPD meets and discusses results
and advises scientists of further action
Plantplan used to develop emergency
response planFurther sampling or elimiting survey
may be undertaken
OCPPO and State Authorities develop a
communication strategyOCPPO and CCEPPD decide on
feasibility of eradication or containment
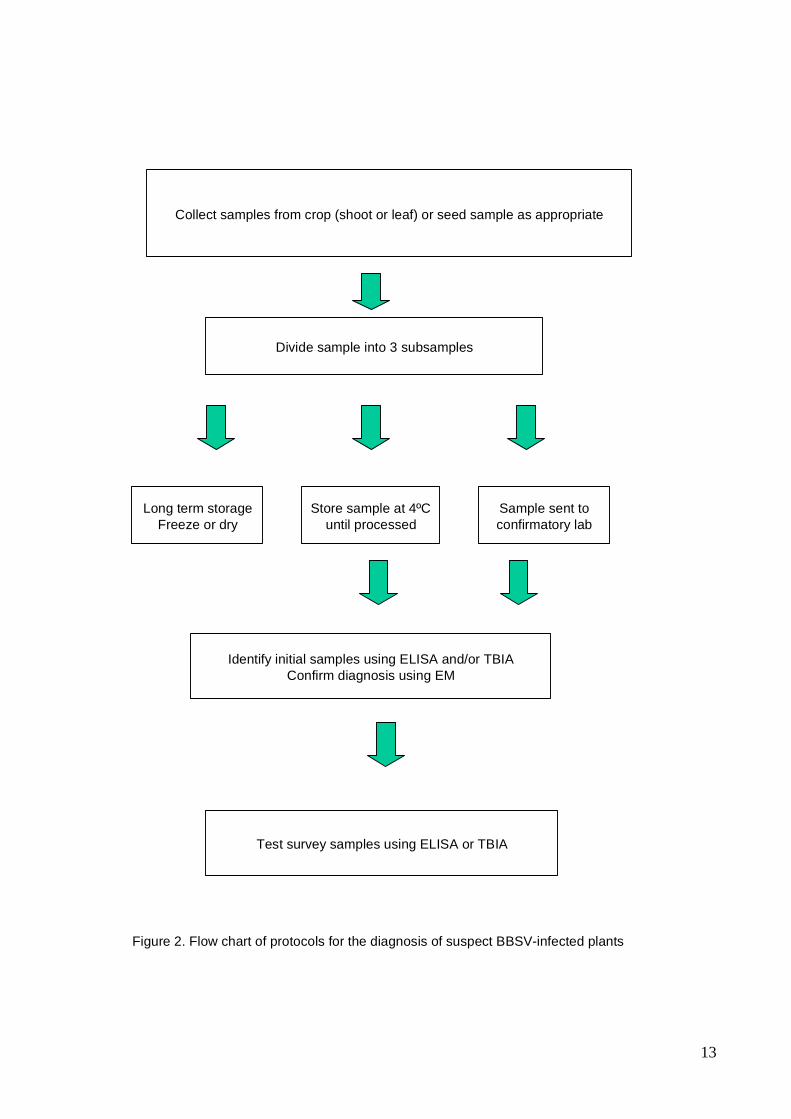
13
Collect samples from crop (shoot or leaf) or seed sample as appropriate
Divide sample into 3 subsamples
Long term storage
Freeze or dry
Store sample at 4ºC
until processed
Sample sent to
confirmatory lab
Identify initial samples using ELISA and/or TBIA
Confirm diagnosis using EM
Test survey samples using ELISA or TBIA
Figure 2. Flow chart of protocols for the diagnosis of suspect BBSV-infected plants

14
3.0 Pest Risk Analysis
3.1 Background
Broad bean mottle virus (BBMV) is listed on the Australian Quarantine and Inspection
Service (AQIS) ICON Import Conditions database as a quarantinable pathogen in Australia.
BBMV is tested for in post-entry quarantine in all hosts in which it is seedborne (Cicer, Pisum
and Vicia species) as required in the regulations listed on ICON.
3.2 Species name
Broad bean mottle virus.
3.3 Synonyms
None.
3.4 Common names
Broad bean mottle virus.
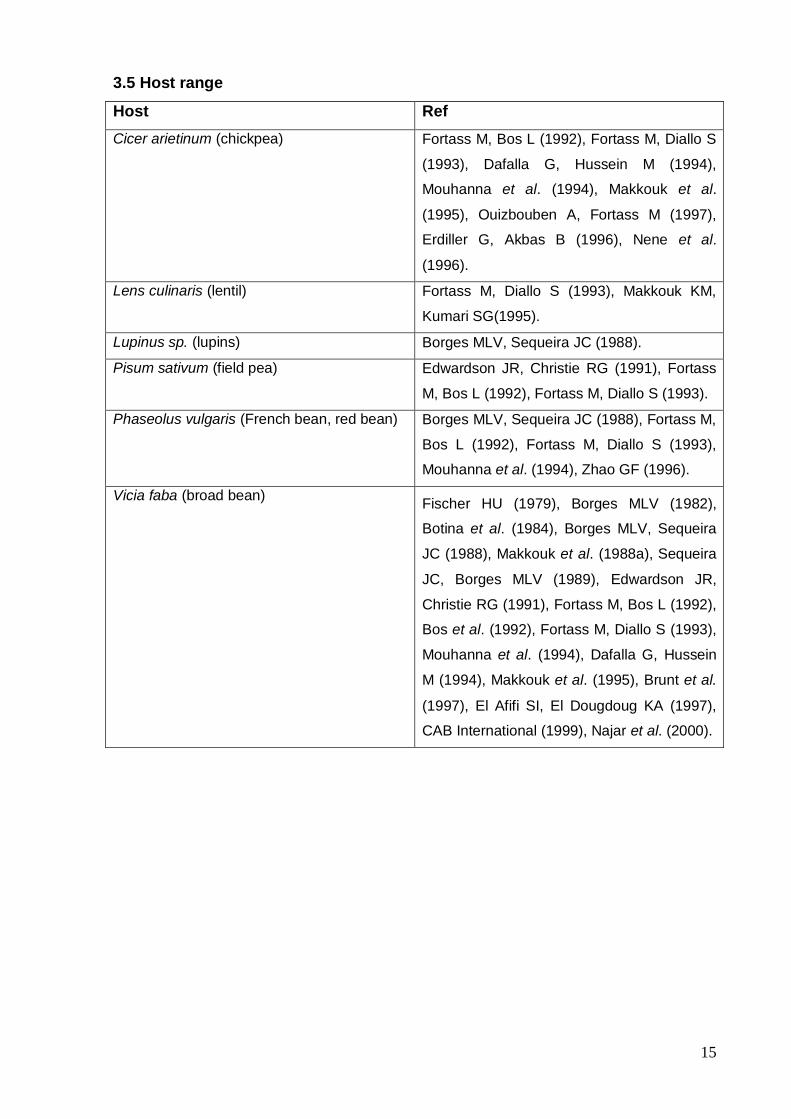
15
3.5 Host range
Host Ref
Cicer arietinum (chickpea) Fortass M, Bos L (1992), Fortass M, Diallo S
(1993), Dafalla G, Hussein M (1994),
Mouhanna et al. (1994), Makkouk et al.
(1995), Ouizbouben A, Fortass M (1997),
Erdiller G, Akbas B (1996), Nene et al.
(1996).
Lens culinaris (lentil) Fortass M, Diallo S (1993), Makkouk KM,
Kumari SG(1995).
Lupinus sp. (lupins) Borges MLV, Sequeira JC (1988).
Pisum sativum (field pea) Edwardson JR, Christie RG (1991), Fortass
M, Bos L (1992), Fortass M, Diallo S (1993).
Phaseolus vulgaris (French bean, red bean) Borges MLV, Sequeira JC (1988), Fortass M,
Bos L (1992), Fortass M, Diallo S (1993),
Mouhanna et al. (1994), Zhao GF (1996).
Vicia faba (broad bean) Fischer HU (1979), Borges MLV (1982),
Botina et al. (1984), Borges MLV, Sequeira
JC (1988), Makkouk et al. (1988a), Sequeira
JC, Borges MLV (1989), Edwardson JR,
Christie RG (1991), Fortass M, Bos L (1992),
Bos et al. (1992), Fortass M, Diallo S (1993),
Mouhanna et al. (1994), Dafalla G, Hussein
M (1994), Makkouk et al. (1995), Brunt et al.
(1997), El Afifi SI, El Dougdoug KA (1997),
CAB International (1999), Najar et al. (2000).
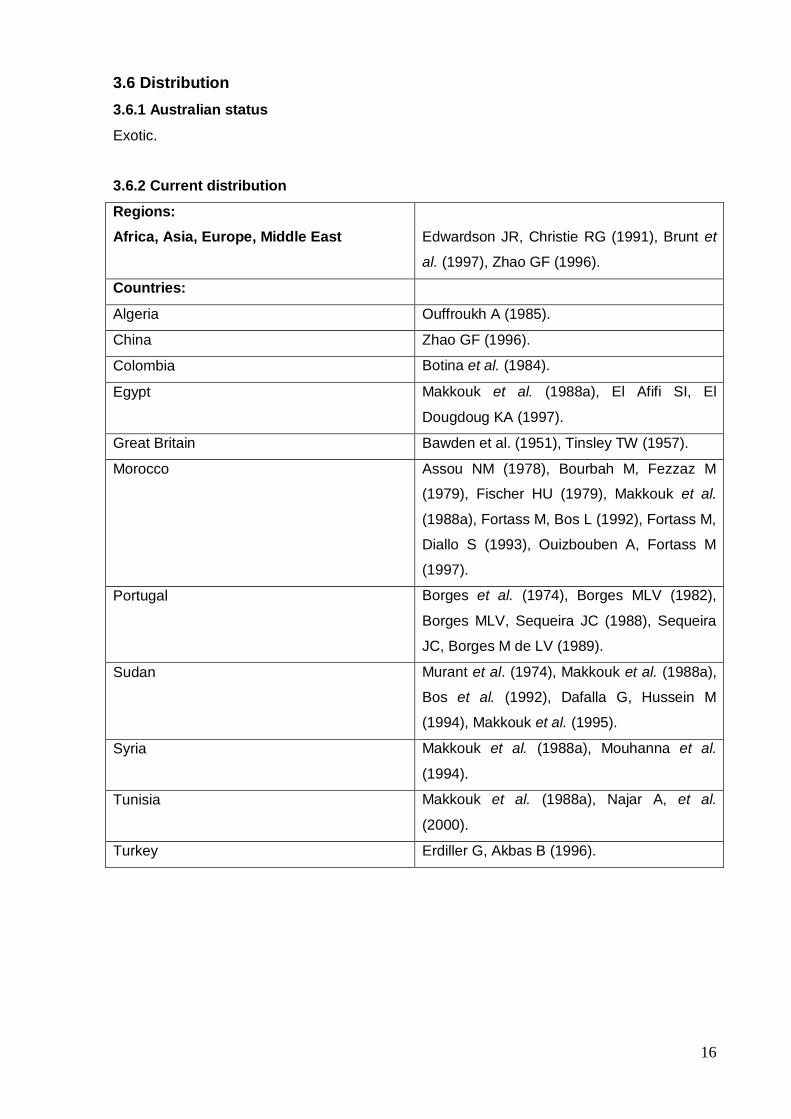
16
3.6 Distribution
3.6.1 Australian status
Exotic.
3.6.2 Current distribution
Regions:
Africa, Asia, Europe, Middle East
Edwardson JR, Christie RG (1991), Brunt et
al. (1997), Zhao GF (1996).
Countries:
Algeria Ouffroukh A (1985).
China Zhao GF (1996).
Colombia Botina et al. (1984).
Egypt Makkouk et al. (1988a), El Afifi SI, El
Dougdoug KA (1997).
Great Britain Bawden et al. (1951), Tinsley TW (1957).
Morocco Assou NM (1978), Bourbah M, Fezzaz M
(1979), Fischer HU (1979), Makkouk et al.
(1988a), Fortass M, Bos L (1992), Fortass M,
Diallo S (1993), Ouizbouben A, Fortass M
(1997).
Portugal Borges et al. (1974), Borges MLV (1982),
Borges MLV, Sequeira JC (1988), Sequeira
JC, Borges M de LV (1989).
Sudan Murant et al. (1974), Makkouk et al. (1988a),
Bos et al. (1992), Dafalla G, Hussein M
(1994), Makkouk et al. (1995).
Syria Makkouk et al. (1988a), Mouhanna et al.
(1994).
Tunisia Makkouk et al. (1988a), Najar A, et al.
(2000).
Turkey Erdiller G, Akbas B (1996).
17
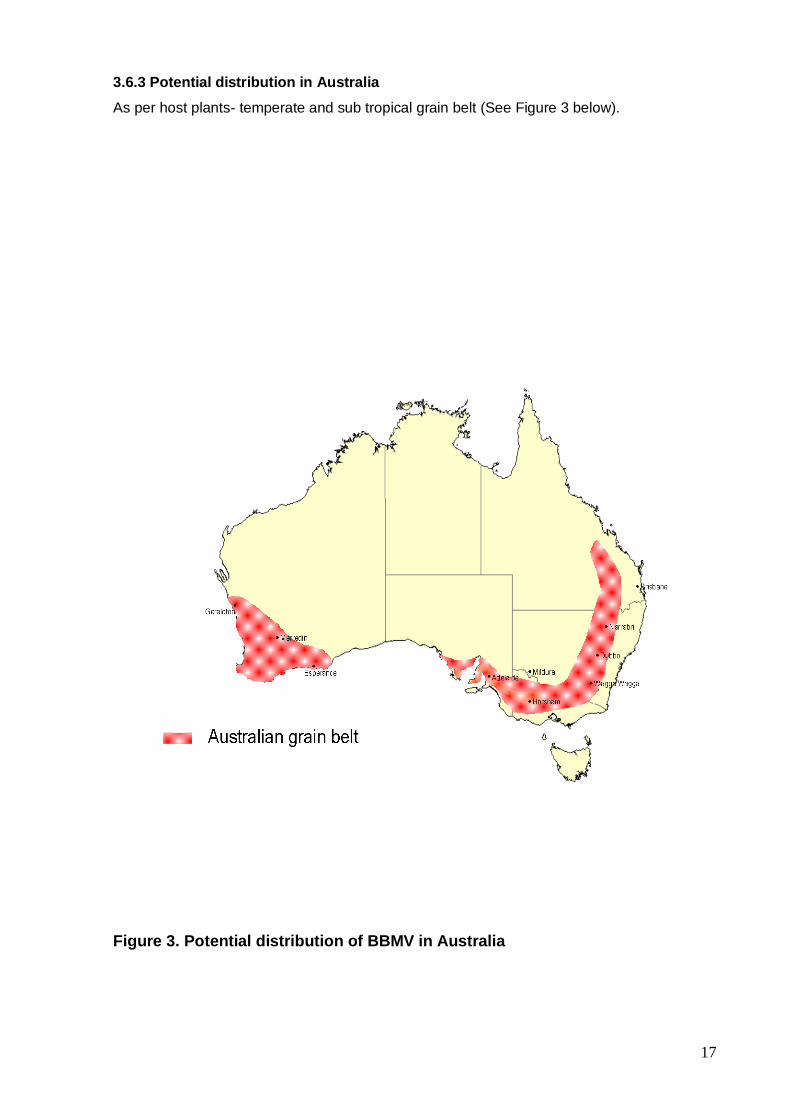
3.6.3 Potential distribution in Australia
As per host plants- temperate and sub tropical grain belt (See Figure 3 below).
Figure 3. Potential distribution of BBMV in Australia

18
3.7 Plant parts affected
3.7.1 Vegetative
All.
3.7.2 Seedborne
Cicer arietinum (chickpea) Fortass M, Bos L (1992), Erdiller G, Akbas B
(1996).
Pisum sativum (field pea) Fortass M, Bos L (1992).
Vicia faba (broad bean) Fortass M, Bos L (1992), Makkouk et al.
(1988a).
3.8 Disease features
BBMV is one of a number of seedborne viruses, which infect temperate pulse crops and
cause mottle/mosaic symptoms. Symptoms of BBMV may be difficult to distinguish from
those of broad bean stain virus (BBSV) or broad bean true mosaic virus (BBTMV) on faba
bean. All three viruses can be spread by weevils and a distinguishing feature of infection by
all of them may be chewed leaf margins as a result of the weevil vector feeding (Cockbain et
al., 1975). BBMV and BBSV (but not BBTMV) may cause staining, necrosis or crinkling of
Vicia faba seed. The three viruses are difficult to distinguish on particle morphology as they
all have isometric particles of similar diameter (Makkouk et al. 1988a). BBMV particles are 26
nm diameter and rounded, whilst BBSV and BBTMV have angular, isometric particles of
about 25 nm diameter (Gibbs AJ, Smith HG, 1970). BBMV can be distinguished from BBSV
and BBTMV by its host range, which includes most temperate pulses and extends outside
the Fabaceae (Makkouk et al. 1988a, Fortass M, Diallo S, 1993, Mouhanna et al. 1994, Brunt
et al. 1997). BBSV and BBTMV are characterised by very narrow host ranges, restricted to a
few temperate pulses.
3.9 Biology
3.9.1 Identification
Identification of the virus is based on serological reactions. BBMV can be confused with two
exotic pulse comoviruses, BBSV and BBTMV, which it is difficult to distinguish from by
symptoms and particle morphology, but to which it is serologically unrelated. Sequence data
for BBMV are listed on the Genebank database although there do not appear to be any
published PCR tests for BBMV diagnosis. Indicator plant tests can also be used to identify
the virus and the diagnosis may be confirmed by electron microscopy (Makkouk et al. 1988,
Fischer 1979). BBMV also causes the formation of large characteristic inclusions consisting

19
of isometric virus particles in infected plant cells, which are considered diagnostic and are
visible by light microscopy (Makkouk et al. 1988, El-Afifi and El-Dougdoug 1997).
(NB. We are currently developing a PCR test for BBMV detection and this method will be
added to the manual after completion and validation.)
3.9.2 Virus strains
BBMV was first found in England in Nottinghamshire in 1951 in a severely infected faba bean
crop and then in a Cambridge crop in 1957 but was not found elsewhere despite surveys
(Gibbs AJ, 1972). The first report of BBMV outside Great Britain was from the Sudan (Murant
AF, Abu Salih HS, Goold RA (1974) and the Sudan strain was reported as being slightly less
virulent and slightly different antigenically to the English type strain. Botina et al. (1984)
described a strain of BBMV from Columbia, which differed in symptoms and host range from
those described in the literature, but they did not compare strains by serology. Makkouk et al.
(1988a) undertook detailed studies on four isolates of BBMV, one each from Morocco,
Tunisia, Sudan and Syria. They found that although they were indistinguishable serologically
when tested by ELISA and gel diffusion tests, they differed slightly in host range and
symptoms and concluded that they were different strains of BBMV. Sequeira and Borges
(1989) identified isolates of BBMV from faba beans in Portugal which gave very mild
symptoms but found that they could be lethal in peas. The Portugese isolates were found to
be serologically identical to English isolates. Bos et al. (1992) described a new strain of
BBMV from Sudan, which produced a neally symptomless infection in faba beans despite a
high concentration of the virus in infected plants. They reported that it did not differ from
other strains in gel diffusion and ELISA tests, light and electron microscopy, host range and
symptoms on other major hosts. Fortass and Bos (1992) studied ten isolates of BBMV from
Morocco, Algeria, Sudan and Tunisia and were able to group them into three groups based
on pathogenicity (mild, intermediate and severe) towards a number of grain legume
genotypes and Gomphrena globosa but found that they all gave similar reactions to antisera
to a Syrian and and a Moroccan isolate. From their studies, within and between host species,
they concluded that virus isolates differed in virulence rather than aggressiveness and that
hosts differed in sensitivity rather than susceptibility. El-Afifi and El-Dougdoug (1997)
identified three strains of BBMV in Egypt, which differed in their host range and the
localisation of virus particles in infected cells, but found that all strains caused characteristic
inclusion bodies.

20
3.9.3 Serological relationships
Rybicki and von Wechmar (1981) studied the serological interrelatedness of the
bromoviruses by determining the relatedness of three group members, BBMV, brome mosaic
virus (BMV) and cowpea chlorotic mottle virus (CCMV). They compared homologous and
heterologous relationships using gel precipitin, "rocket" immunoelectrophoresis and direct,
indirect and double antibody sandwich (DAS) ELISA techniques with native virus and coat
protein under a range of pHs, after fixation with formaldehyde and under capsid swelling
conditions. They concluded that the bromoviruses were serologically related but found that
the relationship depended on antigenic determinants more easily recognised in swollen virus
or free protein than in compact, low pH virus. BMV and CCMV were more closely related to
each other and only distantly related to BBMV. Although the relationship between BMV or
CCMV and BBMV could be demonstrated by manipulation of test conditions, they concluded
that there was no reaction between them under any conditions in the DAS ELISA, which is a
strain-specific means of differentiating viruses. This work confirmed the earlier findings of
Scott and Slack (1971) who found that BMV and CCMV were distantly serologically related to
each other but not to BBMV and Hull (1972), who found that BBMV was less closely related
to BMV and CCMV than they were to each other. Valverde (1985) described a new member
of the bromovirus group, spring beauty latent virus (SBLV), which infected Spring beauty
(Claytonia virginica). Using Ouchterlony double diffusion tests, he concluded that SBLV was
distantly serologically related to BMV and CCMV but not to BBMV. Recent sequence
comparisons between SBLV, BMV, CCMV and BBMV (Fujisaki et al. 2003) demonstrated
that SBLV is closely related to BMV and CCMV but not to BBMV.
3.9.4 Symptoms
Early reports of the symptoms of BBMV on faba beans in England described them as
"characteristic vein-clearing of the youngest leaves, which subsequently faded and was
replaced by a bright interveinal mottle" (Bawden et al. 1951, Walters HJ, Surin P 1973).
Makkouk et al. (1988a) tested 14 faba bean cultivars and nine breeding lines for their
reaction to a Moroccan and a Tunisian strain of BBMV and reported that all genotypes
reacted in about seven days with severe systemic vein chlorosis to chlorotic vein banding,
developing into overall chlorosis in a couple of leaves following the inoculated leaves,
sometimes with remaining interveinal islands of green tissue and in some genotypes with
necrosis along leaf edges (Figs. 3, 4, 5, 6). Plants thereafter more or less recovered
temporarily, with a persistent mottling, marbling or diffuse mosaic recurring in leaves that
developed later. There were some additional or slightly different symptoms in some
genotypes, such as irregular vein chlorosis or vein mosaic and additional necrotic streaks on

21
stems. Fortass and Bos (1992) found that faba bean breeding lines all reacted with mottling
symptoms typical of BBMV when inoculated, but varied in the presence or absence of local
symptoms and the degree of stunting that occurred. A different strain of BBMV has been
reported in Sudan, which despite high concentrations of virus in infected plants is neally
symptomless in faba beans, although its host range and symptoms on other major hosts
does not differ from other isolates (Bos et al. 1992). In a yield loss experiment, Makkouk et
al. (1988b) found that infection of faba bean with BBMV 11 weeks (pre-flowering), 15 weeks
(flowering) and 20 weeks (pod setting) after sowing lead to 54%, 84% and 37% yield loss
respectively. BBMV can also cause symptoms on faba bean seed, which resemble those
caused by BBSV and include necrotic patches, staining and shrivelling (Fig. 9).
BBMV has a wide host range and Makkouk et al. (1988a) found that one or more isolate of
BBMV systemically infected 21 species (12 legume and 9 non-legume) out of 27 tested.
They found that infection in several species was symptomless but major legumes such as
chickpea, lentil and especially pea, suffered severely from infection. Fortass and Diallo
(1993) then reported the first natural occurrence of BBMV in chickpea, lentil, pea and
common bean in Morocco. They found the field symptoms on these species difficult to
define, probably due to mixed infections. The symptoms on peas and lentils consisted of a
necrosis of the lower leaves and a mild mosaic on the tip leaves of peas and vein clearing
and mottling on lentils. Chickpeas developed a striking wilt of the lower leaves and a yellow
mosaic of the upper leaves. Common bean leaves developed chlorotic spots. The authors
were unable to determine whether or not these field symptoms were solely caused by BBMV
infection but the symptoms are consistent with those described by other authors and
summarised below.
BBMV infects all pea cultivars and generally causes desiccating, rapidly expanding local
lesions leading to withering of inoculated leaves, followed by systemic stem and apical
necrosis (Bawdwn et al. 1951, Waltters HJ, Surin P 1973, Makkouk et al. 1988a, Bos et al.
1992, Fortass M, Bos L 1992). Fortass and Bos (1992) found that ten pea accessions tested
were vulnerable to BBMV and that some highly vulnerable lines gave additional symptoms of
systemic wilting or necrosis and a few lines gave atypical symptoms of an unusual yellow
mosaic.
Makkouk et al. (1988a) reported differing reactions of two chickpea lines to a Moroccan
isolate of BBMV, one reacting with vein chlorosis followed by mosaic and the other reacting

22
with necrotic stem streaking and tip necrosis. Fortass and Bos (1992) found that nine
chickpea breeding lines tested were all vulnerable to all isolates of BBMV tested.
Makkouk et al. (1988a) found that four lentil genotypes inoculated with the same Moroccan
isolate of BBMV reacted with systemic interveinal chlorosis, mottling or mosaic and
sometimes local necrosis. Fortass and Bos (1992) found that ten lentil lines tested were
mostly but not always susceptible to most BBMV isolates. Symptoms were usually a local
necrotic reaction followed by systemic yellow mosaic and sometimes necrosis (Fig. 7).
Makkouk et al. (1988a) also found that the Moroccan isolate could infect most cultivars of
common bean, usually causing small chlorotic local lesions, usually followed by systemic
infection which may involve yellow stippling or blotching in some lines or interveinal mottle
(Brunt et al. (1997) (Fig. 8).
3.9.5 Disease cycle
BBMV is an obligate plant pathogen. It survives between growing seasons of the primary
pulse host in an alternative host or in infected seed. Due to the fact that the natural host
range includes at least four families (Brunt et al. 1997, Makkouk et al. 1988a) and includes
many food legumes and non-legume wild hosts it may survive in a range of alternative hosts.
Carryover in infected seed is a likely method of survival of the virus and Makkouk et al.
(1988a) and Fortass and Bos (1991) suggest that seed transmission in faba beans and other
legume crops would explain the widespread occurrence of BBMV in West Asia and North
Africa. Initial infection of a crop occurs when infected seed is sown or it is spread to the crop
host plants from the alternative host by its beetle vectors. Secondary spread occurs within
the crop during insect vector feeding.
A number of beetle species have been reported as vectors of BBMV, including Acalymma
trivittata (striped cucumber beetle), Diabrotica undecimpunctata (spotted cucumber beetle)
and Colaspis flavida (grape colaspis) (Walters HJ, Surin P 1973). A number of weevil
species are also vectors of BBMV and include Sitona lineatus and Apion arrogans which
have also been reported to transmit BBSV and BBTMV (Cockbain AJ 1971, Rothamstead
Experimental Station Report 1982, Borges M, de LV, Louro D 1974, Edwardson JR, Christie
RG 1991, Makkouk KM, Kumari S 1989,). Ahmed and Eisa (1991) reported that the larvae of
the lesser armyworm (Spodoptera exigua) transmitted BBMV with an efficiency of up to 31%.

23
3.9.6 Dispersal
A number of beetle and weevil species have been reported as vectors of BBMV, including
Acalymma trivittata (striped cucumber beetle), Diabrotica undecimpunctata (spotted
cucumber beetle) and Colaspis flavida (grape colaspis), Sitona lineatus and Apion arrogans
(Rothamstead Experimental Station Report 1982, Borges M, de LV, Louro D 1974,
Edwardson JR, Christie RG 1991, Makkouk KK, Kumari S 1989).
Walters and Surin (1973) undertook transmission experiments with five beetle species and
found that Acalymma trivittata (striped cucumber beetle), Diabrotica undecimpunctata
(spotted cucumber beetle) and Colaspis flavida (grape colaspis) all transmitted BBMV at a
transmission rate of 6-10%. Fortass and Diallo (1993) undertook transmission experiments
with curculionid weevils in Morocco and found that Apion radiolus, Hypera variablis (H.
postica), Pachytychius strumarius, Smicronyx cyaneus and Sitona lineatus were all vectors
of BBMV and that S. lineatus was an efficient vector with aquisition and inoculation occurring
at the first bite, 41% transmission rate and virus retention of at least seven days. Makkouk
and Kumari (1995) assessed the ability of four weevil species, naturally occurring on
legumes in Syria, to transmit BBMV and found that Apion arrogans, Sitona limosus and S.
lineatus were able to transmit BBMV but that S. crinita (S. macularius) failed to transmit
BBMV. These findings confirmed their earlier finding of Apion arrogans as a vector of BBMV
(Makkouk KM, Kumari SG, 1989). Borges and de Louro (1974) found that Sitona lineatus
var. viridifrons was abundant in faba bean fields in Portugal and confirmed it as a vector of
BBMV, with a transmission efficiency of about 6%, in glasshouse feeding experiments.
Borges and Sequeira (1988) confirmed it as the vector of BBMV in the field. Ahmed and Eisa
(1991) reported significant crop loss due to BBMV in the Sudan but were unable find any of
the reported vectors infesting affected fields. They observed a relationship between
infestation of the fields with larvae of the lesser armyworm (Spodoptera exigua) and in
glasshouse transmission studies found that the larvae transmitted BBMV with an efficiency of
up to 31%.
3.10 Assessment of likelihood
3.10.1 Entry potential
HIGH BBMV is seedborne in Cicer arietinum (chickpea), Pisum sativum (field pea) and
Vicia faba (broad bean) (Makkouk et al. 1988a, Fortass M, Bos L (1992).

24
A mild strain of BBMV in faba bean in the Sudan was reported to be seed transmitted by
Murant et al. (1974) in conjunction with bean yellow mosaic virus (BYMV). Makkouk et al.
(1988a) then reported experimental seed transmission of BBMV and BYMV in faba bean of
1.37% after inoculating plants in the field. Fortass and Bos (1992) reported the first natural
infections of pea and chickpea with BBMV in Morocco. They undertook host range and seed
transmission experiments in a range of food legumes by inoculating with BBMV. They found
seed transmission in faba bean, pea and chickpea at rates of 1.2%, 0.1% and 1.0%
respectively. They were unable to determine whether seed transmission occurred in lentils,
as infected plants failed to set seed. Erdiller and Akbas (1996) confirmed that BBMV was
seed transmitted in chickpea by growing out and testing farmers' seed in Turkey.
Seed is the only commodity imported for these species, therefore importation of infected
seed may occur. BBMV occurs in most areas where the host species are grown except North
America. Seed of all the above genera are imported for breeding purposes from countries
where BBMV is endemic. The virus would survive intact in seed shipments.
3.10.2 Host range potential
MEDIUM: The natural host range is listed in 3.5 and includes most temperate pulses (faba
bean, field pea, chickpea, lentil, lupin and French or red bean). Experimental transmissions
have shown that many other species in the Fabaceae are susceptible eg. Lathyrus oderatus,
Trifolium incarnatum, T. pratense, T subterraneum, Glycine max (Bawden et al. 1951); L.
oderatus, Lupinus albus, Melilotus officinalis, T. pratense, T. repens (Walters HJ, Surin P
1973); M. albus, Vigna unguiculata (Makkouk et al. 1988a). There are no data on seed
transmission of BBMV in these hosts. BBMV has a wide host range and Makkouk et al.
(1988a) found that one or more isolate of BBMV systemically infected 21 species (12 legume
and 9 non-legume) out of 27 tested. These data suggest that there is potential for BBMV to
establish in species in the Fabaceae beyond its recorded natural host range. Clover pastures
in Australia would be likely potential hosts of BBMV. Susceptible hosts were also identified
by experimental inoculation in the Chenopodiaceae, Cucurbitaceae, Solanaceae and the
Amaranthaceae (Walters HJ, Surin P 1973, Makkouk et al. 1988a, Bos et al. 1992, Brunt et
al. 1997).
3.10.3 Establishment potential
HIGH: BBMV has the potential to survive and become established throughout most or all of
the range of hosts. Distribution is not limited by environmental conditions that prevail in
Australia. Based on its current world distribution and known conditions of survival, it is likely
to survive wherever major hosts are grown.

25
The host crops are well established in Australia and these cropping areas are suitable for
BBMV to establish. The natural host range is listed in Table 3.5, however experimental
transmissions have shown that other species in the Fabaceae (Lathyrus oderatus, Trifolium
incarnatum, T. pratense T. repens, T subterraneum, Melilotus albus, M. officinalis, Lupinus
albus, Glycine max, Vigna unguiculata) are susceptible (Bawden et al. 1951, Walters HJ,
Surin P 1973, Makkouk et al. 1988a). These data suggest that there is potential for BBSV to
establish in species in the Fabaceae beyond its recorded natural host range. Clover pastures
in Australia would be likely potential hosts of BBMV. Susceptible hosts were also identified
by experimental inoculation in the Chenopodiaceae, Cucurbitaceae, Solanaceae and the
Amaranthaceae (Walters HJ, Surin P 1973, Makkouk et al. 1988a, Bos et al. 1992, Brunt et
al. 1997).
Seed to seedling transmission rates are relatively low: faba beans < 1.37%; peas < 0.1%;
chickpeas < 0.9% (Makkouk et al. 1988a, Fortass M, Bos L 1992).
3.10.4 Spread potential
LOW: The pest has potential for natural spread locally. The known vectors are not present in
Australia. The only known way of introducing this virus into disease-free areas is through
infected seed (Naumann, 1993). It is expected that only localised infection would occur
where infected seed is sown, with no secondary spread in the field. Further spread would
depend on the rate of seed transmission and distribution of the harvested seed. However, if
the beetle vectors were also brought into Australia, the spread potential would be much
higher. There are no data on the only endemic Sitona species, Sitona discoideus, as a
potential vector of BBMV. The preferred hosts of S. discoideus are lucerne (Medicago sativa)
and burr medic (M. polymorpha). M. sativa is listed as a susceptible experimental host of
BBMV (Brunt et al. 1997). There are no other insect species recorded in Australia of the
same genera as the known BBMV vectors (Naumann 1993).
The symptoms of the virus on the host are not diagnostic and are similar to those caused by
a range of seedborne viruses, therefore it is unlikely that the virus will be detected
immediately. However, unless a vector was found to co-exist with the virus, widespread
establishment could not occur, as the virus will not spread in the field. The rates of seed
transmission are low, suggesting that in the absence of field spread, the proportion of
infected seed will decrease each year.

26
3.12 Overall entry, establishment and spread potential
The overall pest rating is MEDIUM (ratings based on PHA Industry Biosecurity Planning
Guide) or LOW based on Biosecurity Australia ratings.
3.13 Assessment of consequences
3.13.1 Economic impact
LOW: The economic impact is likely to be low due to the absence of the known vectors in
Australia. If the vector also entered Australia with the virus and both became established,
then the economic impact would be greatly increased. Crop yields can be severely affected
in areas where the virus and its vector co-exist. Faba bean yield losses have been reported
to range from 37 to 84% (Makkouk et al. 1988b). BBMV is widespread in West Asia and
North Africa and is considered a threat to production (Fortass M, Diallo S 1993) and has
been reported at within crop virus incidences of around 20% (Ouizbouben A, Fortass M
1997, Najar et al. 2000).
Elsheikh and Osman (1995) found that faba bean plants which were inoculated with BBMV
four weeks after sowing showed a 52% reduction in shoot weight and similar reduction in
root weight 12 weeks later. They found that BBMV caused a decrease in the number of pods
(62%), the dry weight of pods (33%), the number of seeds per plant (43%) and the 100-seed
weight (15%). They found that seedborne BBMV caused a decrease in nodulation of 24%
and a decrease in shoot and root dry weight, and the number of nodules and flowers per
plant. Makkouk et al. (1988b) conducted a yield experiment by inoculating faba beans with
BBMV 11 (pre-flowering), 15 (flowering) and 20 (pod setting) weeks after sowing and
recorded 54%, 84% and 37% yield losses respectively.
BBMV can also cause symptoms on faba bean seed, which resemble those caused by BBSV
and include necrotic patches, staining and shrivelling (Fig. 9) which would affect the
marketability of the affected grain.
3.13.2 Environmental impact
NEGLIGIBLE: there is no potential to degrade the environment or otherwise alter the
ecosystems by affecting species composition or reducing the longevity or competitiveness of
wild hosts. It has no effect on human or animal health.

27
3.13.3 Social impact
NEGLIGIBLE: there is no potential to affect the social environment.
3.14 Combination of likelihood and consequences to assess risks
The pest risk is MEDIUM, the economic impact is LOW, the environmental and social
impacts are NEGLIGIBLE. Therefore the economic risk rating is MEDIUM, the
environmental risk rating is LOW and the social risk rating is LOW (Risk ratings based on
PHA Industry Biosecurity Planning Guide).
3.15 Surveillance
BBMV is a quarantinable pathogen and is actively tested for in post-entry quarantine.
Regular surveys of pulse crops for endemic viruses could easily be extended to include
screening of samples for BBSV using ELISA or TBIA.
3.16 Diagnostics
Samples suspected of being infected with BBMV would need to be identified quickly and
accurately. The accompanying report describes methods for sampling and diagnosing BBMV
using ELISA and TBIA and other confirmatory methods. The ELISA procedure for BBSV
detection is used on a regular basis at the DPI Post-entry Quarantine Station for temperate
Pulses at DPI-Horsham, Victoria, and could be undertaken by any trained virologist. TBIA is
used at the Quarantine Station to confirm positive ELISA results. The initial diagnosis would
need to be confirmed by another virologist and by a second method. BBMV may be been
confused with two exotic pulse comoviruses, BBSV and BBTMV, which are difficult to
distinguish by symptoms and particle morphology but which are serologically distinct
(Cockbain et al., 1975). BBMV is also spread by some of the same vectors as BBSV and
BBTMV. Mixed infections may occur and careful diagnosis is required.
NB. A PCR test for detection of BBMV is currently being developed at DPI-Horsham and will
be added to this manual when completed and validated.
3.17 Training
There is a general need for industry training in biosecurity and awareness of the potential
impact of exotic diseases. Due to the similarity of symptoms of a range of pulse viruses,
including BBMV, on pulse hosts, training and education need to be of a general nature.

28
Training in the recognition of seed symptoms is likely to maximise the likelihood of early
detection.
3.18 References
See Section 8.

29
4.0 Diagnostic protocol
4.1 The diagnostic test/s and diagnostic sequence
Double antibody sandwich enzyme-linked immunosorbent assay (DAS-ELISA) is the
recommended primary test for detection of BBMV (Clark MF, Adams AN, 1977). There are
published sequence data for BBMV and a PCR test is currently being developed at DPI-
Horsham but is not yet available for routine diagnostics. The method will be added to this
manual following completion and validation. There are extensive published data on
serological detection of BBMV, including the serological relationship between BBMV and
other bromoviruses (Section 3.9.3) and differences between strains of the virus (Section
3.9.2). Tissue-blot immunoassay (TBIA) is included as an alternative primary diagnostic test,
because although less widely used than ELISA it has proved to be the test of choice under
some circumstances (Makkouk KM, Comeau A, 1994). Both ELISA (eg. Makkouk et al.
1988a, Fortass M, Bos L 1992, Bos et al. 1992, Fortass M, Diallo S 1993, Ouizbouben A,
Fortass M 1997, Makkouk et al. 1994) and TBIA (eg. Tadesse et al. 1999, Najar et al. 2000,
El-Muadhidi et al. 2001, Makkouk et al. 1992) have been used widely for the identification of
BBMV from field samples and in surveys.
ELISA is the most commonly used serological method for BBMV detection and its use and
comparisons of various ELISA methods are widely reported in the literature. Rybicki and Von
Wechmar (1981) compared direct, indirect and sandwich ELISA methods for the detection of
three bromoviruses, including BBMV, and found that they were each best suited to a different
application. They concluded that direct ELISA is a sensitive if limited means of showing virus
relationships and that indirect ELISA is an ideal sensitive method for screening large
numbers of antisera for heterospecific reactivity. They found that the addition of one specific
reaction step in the DAS-ELISA, that of adsorbed specific antibody, greatly increased the
specificity and sensitivity of the test, and they recommended DAS-ELISA for specific virus
detection. Salama et al. (2003) compared three ELISA methods for detecting BBMV in faba
bean and found that the most sensitive methods were ELISA with enzyme amplification (EA-
ELISA) and double antibody sandwich ELISA (DAS-ELISA) which both enabled detection of
BBMV in sap diluted to 1/2500. Penicillinase ELISA (PNC-ELISA) was found to be much less
sensitive for BBMV detection.
4.2 The initial samples
4.2.1 Sample handling and subsampling
It is important that the samples are entered onto sample reference sheets (Appendix 1)
which contain sufficient information to enable revisiting of the site, describe symptoms and

30
other relevant information and recording of diagnostic test results. It is vital that information is
provided here to ensure that samples are handled correctly, that sub-samples are taken as
reference samples and so that material can be sent to other experts for confirmation.
4.2.2 Sample storage
As soon as the diagnostician becomes aware that the sample submitted for diagnosis may
be an exotic or emergency pathogen, the diagnostician has the responsibility to seek expert
advice from State Plant Standards or equivalent or AQIS or the Office of the Chief Plant
protection officer (OCPPO) on the appropriate manner/location in which the sample should
be stored and appropriate further testing/action. It is not appropriate for the diagnostician to
continue tests without informing the proper authorities. In Victoria, suspected BBMV-infected
plants can be stored in the DPI pulse quarantine station's AQIS registered storage area for
quarantine samples. Reference material from the original sample should always be kept: for
virus samples, material should be dried and/or frozen, and if possible nucleic acid extractions
conducted.
4.2.3 Visual symptoms
Visual symptoms should be recorded and photos taken where possible.
4.2.4 Documentation
It is important to note that proper documentation of samples and diagnostic procedures and
results is initiated at this stage.
4.3 Further samples
It is important to note that proper documentation of samples and diagnostic procedures and
results is initiated at this stage.
4.3.1 Sample collection, transport and storage
It is important that samples are collected and stored correctly as deteriorating plant samples
may be unsuitable for diagnostic tests. Leaf samples should be placed in labelled sealed
plastic bags and stored in the field in a cooled, insulated container (Esky). Samples should
then be transferred to a refrigerator if they are to be tested within a week of collection. If
there are to be delays in testing then samples for use in ELISA tests should be frozen. This
practice is not recommended as freezing is likely to reduce the sensitivity of the test and
should only be resorted to if there is some impediment to rapid receicval and processing of a

31
batch of samples. If samples are to be used in TBIA tests then samples should be blotted
onto multiple nitrocellulose membranes and the remaining tissue frozen or dried. If a survey
is being conducted and a team of people is assembled to assist, pictures of plants with virus
symptoms will help in sample selection. Advice on phytosanitary measures required to
prevent disease spread in the field should be provided (Appendix 2).
4.3.2 Sample locations
It is important to record the precise location of all samples collected, preferably using GPS,
or if this is not available, map references including longitude and latitude and road names
should be recorded.
4.4 Confirmation of diagnosis
It is important that all diagnoses of suspected exotic and emergency pathogens are
undertaken according to the following parameters: the diagnostician has expertise in this
form of diagnosis, the test is undertaken as described in this manual, the results are
confirmed by diagnosis in another recognised laboratory or another diagnostician and where
possible diagnosis is confirmed by a second method. Methods suitable for confirming the
primary diagnosis are described in Section 6 (eg electron microscopy to confirm presence of
the correct size virus particle).

32
5.0 Identification of pathogen (primary diagnostic test)
5.1 Enzyme-linked immunosorbent assay (ELISA)
5.1.1 Introduction
Direct double antibody sandwich ELISA (DAS-ELISA) (Clark, MF, Adams, AN, 1977) is used
with polyclonal antisera and indirect ELISA (Voller A et al. 1976, Torrance L, Pead MT 1986)
is normally employed with monoclonal antibodies. Both procedures are described below.
5.1.2 General items required
1. Samples - leaves, shoots, seedlings (germinated seeds).
2. Microtitre plates (e.g. Nunc maxisorp plate) and lids.
3. Mortars and pestles (sterile and kept cool), or sap extractor machine or plastic sample
bags and rolling pins.
4. 200 L pipettes, 0.5-200 L pipettes, 100-200 L multistepper pipettes and sterile tips.
5. Balance (that weighs to at least two decimal places) and weighboats.
6. pH meter and magnetic stirrer.
7. Plate reader and computer.
8. Incubator kept at 37°C.
9. 1 litre graduated glass bottles with lids, volumetric flasks, flasks, beakers, tube racks
and screw cap 5 ml plastic specimen storage bottles, plastic wash bottles.
10. Disposable gloves, paper towels, plastic reservoir.
5.1.3 Specific items
1. BBMV antisera and conjugate.
BBMV antisera are not available commercially. The antiserum recommended for this protocol
was produced at the International Centre for Agricultural Research in the Dry Areas
(ICARDA), Aleppo, Syria. It has been used extensively by Dr Khaled Makkouk and Dr Safaa
Kumari (previous and current Manager of the Virology Laboratory, ICARDA, respectively) in
survey and research work throughout the Middle East and North Africa (as cited in this
protocol). It has been used at the DPI pulse quarantine station, DPI, Horsham, Victoria, for
post-entry quarantine testing for BBMV for ten years. It is used successfully in both DAS-
ELISA and TBIA. The BBMV antiserum is conjugated with alkaline phosphatase for use in
DAS-ELISA. 1ml of the ICARDA BBMV antiserum has been purchased and is stored at DPI,
Horsham, for use in an incursion. Dried and frozen BBMV- infected leaf tissue is also stored
in the locked quarantine -18 C storage facility at the quarantine station. A stock of BBMV
antiserum is held at the station for use in post-entry quarantine testing and additional
33
antiserum could be made available for use in an incursion. Additional BBMV- alkaline
phosphatase conjugate would be prepared when required.
NB. If further antisera has to be purchased from overseas an AQIS permit is required. Dr
Angela Freeman, manager DPI pulse quarantine station holds a current permit to import the
antiserum and virus infected tissue.
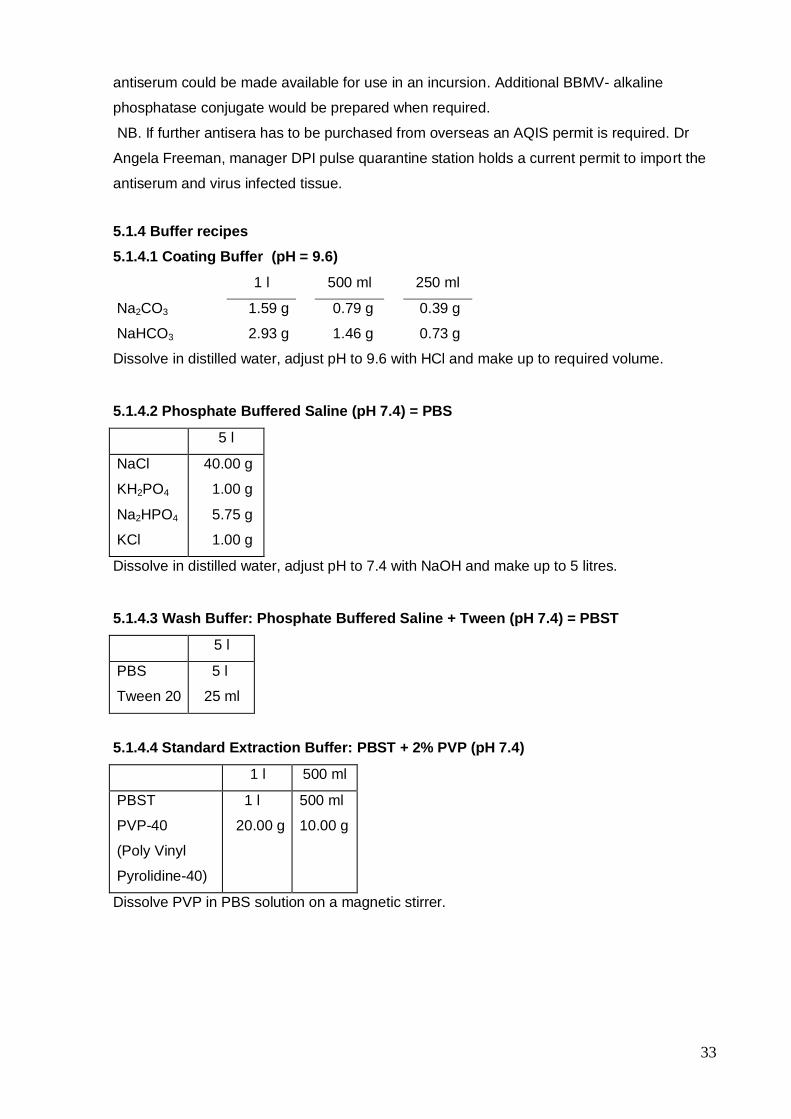
5.1.4 Buffer recipes
5.1.4.1 Coating Buffer (pH = 9.6)
1 l 500 ml 250 ml
Na2CO3
NaHCO3
1.59 g
2.93 g
0.79 g
1.46 g
0.39 g
0.73 g
Dissolve in distilled water, adjust pH to 9.6 with HCl and make up to required volume.
5.1.4.2 Phosphate Buffered Saline (pH 7.4) = PBS
5 l
NaCl
KH2PO4
Na2HPO4
KCl
40.00 g
1.00 g
5.75 g
1.00 g
Dissolve in distilled water, adjust pH to 7.4 with NaOH and make up to 5 litres.
5.1.4.3 Wash Buffer: Phosphate Buffered Saline + Tween (pH 7.4) = PBST
5 l
PBS
Tween 20
5 l
25 ml
5.1.4.4 Standard Extraction Buffer: PBST + 2% PVP (pH 7.4)
1 l 500 ml
PBST
PVP-40
(Poly Vinyl
Pyrolidine-40)
1 l
20.00 g
500 ml
10.00 g
Dissolve PVP in PBS solution on a magnetic stirrer.

34
5.1.4.5 Conjugate Buffer: PBST + 2% PVP + 0.2% ovalbumin (pH 7.4)
1 l 500 ml
PBST +2% PVP
Ovalbumin
1 l
2.0 g
500 ml
1.0 g
Dissolve ovalbumin in PBST-PVP solution on a magnetic stirrer.
5.1.4.6 Substrate Buffer (pH = 9.8)
1 l 500 mL 250 mL
Diethanolamine 106.7 g 53.35 g 26.65 g
Dissolve in distilled water, adjust pH to 9.8 with HCl and make up to required volume.
5.1.4.7 Indirect ELISA Extraction Buffer (pH = 9.6)
1 L 500 mL 250 mL
Na2CO3
NaHCO3
PVP
1.59
2.93
20.00
0.79
1.46
10.00
0.39
0.73
5.00
Dissolve in distilled water, adjust pH to 9.6 with HCl and make up to required volume.
5.1.5 Methods
5.1.5.1 Direct double antibody sandwich (DAS) ELISA
1. Coat microtitre plate with polyclonal BBMV antibody diluted at recommended rate in
coating buffer using 200 l/well. Coat 60 internal wells only. Put 200 l distilled water in
each of the outside wells.
2. Cover microtitre plate and incubate at 37oC for 4 hours.
3. Extract samples individually in extraction buffer (1:5 or 1:10 w/v) using a fresh mortar and
pestle for each sample, or a sap extractor with crusher rollers that are washed thoroughly
in between each sample or a plastic bag and rolling pin. Collect extract and store in
labelled 5ml plastic specimen bottles.
4. Wash coated plate with wash buffer (PBST) at least three times (3-5 times), using a wash
bottle, allowing a three minutes soak for each wash.

35
5. Pipette sample extracts and control extracts into paired wells (200 l/well) on the
washed, coated plate. Controls should consist of four wells each of extraction buffer
(blank), healthy plant extract (negative) and extracts of at least one known BBMV-
infected plant (positive).
6. Cover microtitre plate and incubate in refrigerator at 4°C overnight. Although overnight
incubation is recommended, 4 hours at 37°C is satisfactory.
7. Remove microtitre plate contents and rinse with PBST, using pressure with the wash
bottle, to remove traces of plant material.
8. Wash plate with wash buffer (PBST) at least three times (3-5 times), using a wash bottle,
allowing a three minutes soak for each wash and ensuring no plant material remains.
9. Dilute conjugate (BBMV antibody conjugated with alkaline phosphatase enzyme) to
recommended rate in conjugate buffer, and add to the 60 inner wells of the plate at 200
l/well. Put 200 l distilled water in each of the outside wells of the plate.
10. Cover microtitre plate and incubate at 37°C for 4 hours.
11. Remove microtitre plate contents. Wash plate with wash buffer (PBST) at least three
times (3-5 times), using a wash bottle, allowing a three minute soak for each wash.
12. Just prior to usage, prepare p-nitrophenyl phosphate (PNP) substrate by dissolving PNP
tablets (Sigma 104-105) in substrate buffer to make a 1 mg/ml solution.
13. Add 200 l/well of freshly prepared PNP substrate to all the wells of the plate. Incubate at
room temperature for 1-2hr or until unambiguous reactions are obtained.
14. Read the absorbance values (OD values) of each well at a wavelength of 405 nm (A405)
using a plate reader after 30-60 minutes and again after 2 hours.

36
5.1.5.2 Indirect ELISA
1. Extract samples individually in indirect-ELISA extraction buffer (1:5 or 1:10 w/v) using a
fresh mortar and pestle for each sample, or a sap extractor with crusher rollers that are
washed thoroughly in between each sample or a plastic bag and rolling pin. Collect
extract and store in labelled 5ml plastic specimen bottles.
2. Pipette sample extracts and control extracts into paired wells (200 l/well) of the plate. Fill
the 60 internal wells only. Put 200 l distilled water in each of the outside wells. Controls
should consist of four wells each of extraction buffer (blank), healthy plant extract
(negative) and extracts of at least one known BBMV-infected plant (positive).
3. Cover microtitre plate and incubate in refrigerator at 4°C overnight.
4. Remove microtitre plate contents. Wash plate with wash buffer (PBST) at least three
times (3-5 times), using a wash bottle, allowing a three minute soak for each wash.
5. Dilute monoclonal antibody to recommended rate in conjugate buffer, and add to the 60
inner wells of the plate at 200 l/well. Put 200 l distilled water in each of the outside
wells.
6. Cover microtitre plate and incubate at 37°C for 4 hours.
7. Remove plate contents. Wash plate with wash buffer (PBST) at least three times (3-5
times), using a wash bottle, allowing a three minute soak for each wash.
8. Dilute anti-species conjugated antibody (eg goat anti-mouse) to the recommended rate in
conjugate buffer and add 200 l/well to the 60 inner wells. Put 200 l distilled water into
each of the outside wells.
9. Cover microtitre plate and incubate at 37°C for 4 hours.
10. Remove microtitre plate contents. Wash plate with wash buffer (PBST) at least three
times (3-5 times), using a wash bottle, allowing a three minute soak for each wash.

37
11. Just prior to usage, prepare p-nitrophenyl phosphate (PNP) substrate by dissolving PNP
tablets (Sigma 104-105) in substrate buffer to make a 1 mg/ml solution.
12. Add 200 l/well PNP substrate to all the wells of the plate.
13. Read the absorbance values (OD values) of each well at a wavelength of 405 nm (A405)
using a plate reader after 30-60 minutes and again after 2 hours.
5.2 Tissue-blot immunoassay (TBIA)
5.2.1 Introduction
Lin et al. (1990) found that TBIA was suitable for detection of viruses in the cucumovirus,
luteovirus, potexvirus, potyvirus and tospovirus groups and from a range of tissue types. Hsu
and Lawson (1991) compared both direct blotting of tomato spotted wilt virus plant tissue
with dot-blot immunoassay and ELISA, both of which involve extracting the virus into buffer.
TBIA was found to be sensitive, reliable and rapid and had the added advantages of
simplicity and convenience. Makkouk et al. (1994) found that TBIA was a simple, sensitive
and quick method for barley yellow dwarf virus detection. Makkouk and Comeau (1994)
looked at a range of modifications to this method and their modifications form the basis of the
method described in this manual.
5.2.2 General items required
1. Nitrocellulose membrane (NCM) (CN. 0.45 um) from Schleicher & Schuell, Cat No.
401.196 BA85
2. Goat Anti-Mouse-AP labelled (from Bioreba Cat No. 1031-04 or Sigma Cat No. A-5153)
3. Goat Anti-Rabbit-AP labelled (from Bioreba Cat No. 4050-04 or Sigma Cat No. A-8025)
4. BCIP (from Sigma Cat No. B-8503 or ROCHE Cat no. 1017373)
5. NBT (from Sigma Cat No. N-6876 or ROCHE Cat No. 1087479)
6. Poly Vinyl Alcohol (PVA)
7. Small plastic, flat-bottomed containers with lids (eg empty pipette tip boxes or food storage
boxes).
8. Shaker
5.2.3 Specific items
1. BBMV polyclonal antiserum form ICARDA (See ELISA section)

38
5.2.4 Buffer recipes
5.2.4.1 Phosphate Buffered Saline (pH 7.4) = PBS
5 l
NaCl
KH2PO4
Na2HPO4
KCl
40.00 g
1.00 g
5.75 g
1.00 g
Dissolve in distilled water, adjust pH to 7.4 with NaOH and make up to 5 litres.
1.0g NaN3, a carcinogen used as preservative, may be added per 5 litres. As an alternative,
store at – 4 C instead.)
5.2.4.2 Wash Buffer: Phosphate Buffered Saline + Tween (pH 7.4) = PBST
5 l
PBS
Tween 20
5 l
2.5 ml
5.2.4.3 Standard Extraction Buffer: PBST + 2% PVP (pH 7.4)
1 l 500 ml
PBST
PVP-40
(PolyVinyl
Pyrolidine-40)
1 l
20.00 g
500 ml
10.00 g
Dissolve PVP in PBS solution on a magnetic stirrer.
5.2.4.4 Conjugate Buffer: PBST + 2% PVP + 0.2% ovalbumin (pH 7.4)
1 l 500 ml
PBST
Ovalbumin
(Sigma A-5253)
1 l
2.0 g
500 ml
1.0 g
Dissolve ovalbumin in PBST-PVP solution on a magnetic stirrer.

39
5.2.4.5 Substrate Buffer: Tris 0.1 M, NaCl 0.1 M (pH 9.5)
1 l
Tris (0.1 M) 12.1g
NaCl (0.1 M) 5.8g
MgCl2 (5 mM) 0.4g
5.2.4.6 Substrate stock solutions: NBT and BCIP
Tube 1. Prepare a p-Nitro Blue Tetrazolium (NBT) stock solution of 75 mg/ml in 70%
diemethylformamide. Store at -20 C.
Tube 2. Prepare a 5-Bromo-4-Chloro-3-Indolyl Phosphate (BCIP) stock solution of 50
mg/ml in undiluted diemethylformamide. Store at -20 C.
5.2.5 Method for tissue-blot immunoassay (TBIA) on nitrocellulose membranes
1. It is important to select a type of nitrocellolose membrane which will optimise the result.
The preferred membrane is the Schleiger and Schultz Protran 55 m membrane. Do not
use nylon membranes as they give high backgrounds. Prepare nitrocellulose membrane
by stamping a 10 x 10 grid or use a standard 5 x 10 cm sheet and underline every fifth
row of blots to keep straight. Mark top left corner with an asterisk (*). Handle the
membranes with blunt ended forceps. N.B. If blotting faba bean samples, which tend to
oxidise and go brown/ purple on the membrane, pre-treat membrane by first dipping in
1% sodium sulphite.
2. It is much easier to blot petioles than leaves, which have to be rolled into a cigar shape
before blotting, therefore collect leaves with petioles intact if possible. If thin tissues such
as leaves are used, roll them into a tight core or cigar and secure with masking tape.
3. Wearing gloves, hold tissues in one hand and cut petiole or cigar with a scalpel blade
with the other hand to obtain a single plane cut surface. It has been found that it is not
necessary to wash the blade between the cutting of each sample.

40
4. Press the newly cut surface with a firm but gentle force onto one of the cells of the grid on
the nitrocellulose membrane (NCM).
5. Always include blotted positive and negative controls in the test. These can be
blotted at the same time as the samples or a large number of controls can be blotted in
advance and small pieces of NCM with controls can be added to the dish for processing.
6. If TBIA tests are not used regularly it is advisable to run through the procedure outlined
below using only the positive and negative controls to ensure that the test is working well.
If results are not clear, optimise the test by increasing the concentration of antiserum and
checking the viability of the conjugate and possibly increasing its concentration. Some
monoclonal reactions can be improved by incubating overnight in the fridge. This does
not work for polyclonals and gives a high background. The colour development varies
with different conjugates. It may be worth comparing the results with a couple of different
brands of conjugate.
7. Prior to commencing the processing, prepare polyclonal antisera by reacting out
antibodies to healthy plant proteins. First grind 1g fresh healthy host tissue in 20mls PBS-
Tween (or equivalent 1:20 extraction if larger volume needed). Strain to remove
particulate matter. Dilute polyclonal antiserum 1/2000 or as required in the extract and
incubate 2 hrs in the oven at 37 C.
8. Place all membrane/s in one dish by carefully lowering them, making sure the back of the
membranes are wet first. Wash 3 times with PBS-Tween at 5 minute intervals on a small
shaker.
9. Block NCM in 1 µg/ml Polyvinyl Alcohol (PVA) in PBST and incubate for 1 minute at room
temperature. Save PVA as it is reused many times. Mark date on record sheet on back of
bottle
10. Wash 3 times with PBS-Tween at 5 minute intervals.
11. Place membranes into separate, labelled dishes.
12. Remove prepared polyclonal antisera from oven and prepare monoclonal antisera by
diluting 1/2000 or as required in conjugate buffer (PBST +2% PVP + 0.2% ovalbumin).

41
Add diluted antiserum to each dish and incubate for 1 hour at room temperature on a
small shaker. Save antiserum to be reused and mark record sheet on bottle (may be
used up to 10 times).
13. Wash 3 times with PBS-Tween at 5 minute intervals.
14. Add anti-rabbit conjugate (dilution 1/2000 in conjugate buffer) to membranes processed
with polyclonal antisera or anti-mouse conjugate to membranes processed with
monoclonal antibodies and incubate for 1 hour at room temperature on shaker. N.B.
Membranes can be processed together at this stage. Save conjugate to be reused, mark
date on record sheet on back of bottle (may be used up to 10 times).
15. In preparation for Step 16 remove frozen substrate buffer and substrate solutions tube1
and tube 2 from freezer and thaw.
16. Wash as instep 8.
17. Prepare substrate solution. First check pH of substrate buffer- it is essential that the pH is
exactly 9.5. The reaction will be much fainter if the pH is not optimum. For each 5 ml
substrate buffer add 20 µl tube 1 (NBT) and 20 µl tube 2 (BCIP).
18. N.B. The substrate solution must be prepared fresh just prior to usage. Add substate
solution to the dish (all membranes may be processed together as long as there is
sufficient movement between sheets and they do not stick together thus impeding
development). Incubate for 5 minutes on shaker.
19. To stop reaction, wash with deionised H20.
20. Dry membranes and view under a dissecting microscope. Phloem restricted viruses will
show as a clear dark purple staining of the phloem. Viruses which invade the whole plant
will be seen as clear dark purple staining of the whole petiole cross section.

42
5.2.6 Suppliers
Schleicher & Schell
BioScience GmbH,
P.O. Box 4
Hahnestrasse 3
Dassel 1, D-3354
Germany
e-mail: [email protected]
Internet: http//www.s-und-s.de
ROCHE Diagnostics GmbH
Dept. GD-F
68298 Mannheim
Germany
Bioreba AG
Chr. Merian-Ring 7
CH-4153 Reinach BL1
Schweiz
e-mail: [email protected]

43
6.0 Confirmation of diagnosis
6.1 Electron microscopy
6.1.1 Introduction
BBMV has isometric particles of about 25-26 nm diameter, rounded in profile without
conspicuous capsomere arrangement (Bawden et al. 1951, Gibbs AJ 1972, Brunt et al. 1997,
Makkouk et al. 1988a). Makkouk et al. (1988a) reported that BBMV isometric particles were
readily seen with the electron microscope in crude sap preparations from faba bean field
samples, either fresh or dried, or from faba bean plants inoculated and maintained in the
glasshouse. Virus particles mostly occurred in very high concentrations and a high proportion
of them were penetrated by phosphotungstic acid (PTA) stain.
BBSV and BBTMV also have isometric particles of 25 nm diameter (Gibbs AJ, Smith HG,
1970), therefore direct examination of plant sap using the electron microscope (sap dip) is
only suitable as a confirmatory test for the presence of isometric particles. Immunosorbent
electron microscopy (trapping or decoration) with virus-specific antibodies enables trapping
of particles of the target virus and offers a more definitive test result. However, like ELISA,
this is a serological test, which means that both these forms of diagnosis/virus detection
depend on the quality of the antiserum. It is preferable to use a confirmatory test, which
depends on other properties of the virus.
Key references: Noordham (1973), Ball (1974), Milne (1986), Roberts (1986).
6.1.2 General items required
1. Samples - leaves, shoots or washed roots.
2. Electron microscope grids: copper 400 mesh, Formvar coated, then carbon coated.
3. Glass microscope slides, waxed glass microscope slides, plastic wells, pasteur pipettes,
filter papers, fine forceps.
4. Distilled water, 0.1M sodium phosphate buffer, pH 7.0.
5. Freshly prepared stains: 2% phosphotungstic acid (PTA) and 2% uranyl acetate (UA)
dissolved in distilled water, adjusted to pH 7.0 with NH3.
6. Freshly prepared dilutions of appropriate virus antibodies in 0.07 M sodium phosphate
buffer, pH 6.5.

44
6.1.3 Method
6.1.3.1 Sap dip (negative staining) method
1. Using the scalpel blade, cut approximately 3 mm2 of the test plant material and place it
on a clean microscope slide (if the test material has any suspicious virus symptoms, take
the tissue from this area).
2. Place a 3 mm diameter drop of PTA next to the piece of plant material and thoroughly
crush the plant material into the PTA. If necessary add an extra drop of PTA.
3. Pick up a coated grid with forceps and touch it, coated-side down, onto the drop of PTA
and plant sap mixture.
4. After 2-3 seconds, drain the excess droplet of the grid by touching its edge with a piece of
torn filter paper.
5. Allow the grid to dry for approximately 2 minutes then observe grids for virus particles
using an electron microscope.
6.1.3.2 Immunosorbent electron microscopy
6.1.3.2.1 Trapping method
1. Pipette 30 l drops of antiserum diluted 1:100, 1:1,000 and 1:10,000 in normal saline
onto waxed glass slides.
2. Float a carbon-coated grid, film-side down, on each drop and incubate for 2-3 hours at
37°C.
3. Wash grids five times in normal saline or place grids in 0.1 M sodium phosphate buffer,
pH 7.0, in plastic wells, agitate at intervals for 10 minutes, transfer to a second plastic
well containing buffer and leave for a further 10 minutes and then drain.
4. Extract plant material in normal saline at 1:5 wt/vol and then centrifuge at 8,000g for 10
minutes.
5. Float grids on 30 l drops of sample sap extract and incubate for 2 hours at room
temperature or for 3-36 hours at 4°C.
6. Wash grids five times in normal saline or place grids in plastic wells, containing normal
saline, agitate at intervals for 10 minutes, transfer to a second plastic well containing
normal saline and leave for a further 10 minutes and then drain.
7. Stain grids with 2% PTA and/or UA by floating grids on the stain for 10 minutes then drain
grids by touching the edge with torn filter paper.

45
8. Observe grids for virus particles using an electron microscope.
6.1.3.2.2 Decoration method
1. As an additional step, just prior to examination of the prepared grid using the electron
microscope, in either of the above procedures, add a drop of suitably diluted antiserum
to the prepared grids, incubate for 3 hours at 37°C and drain.
2. Observe grids using an electron microscope for antibody halos surrounding virus
particles. Such halos indicate the specific binding of the virus-specific antibody to the
trapped virus particles on the grid and therefore provide evidence of the true identity of
the virus, based on the specificity of the antiserum used.
6.2 Light Microscopy
6.2.1 Introduction
Rubio and van Slogteren (1956) found granular inclusion bodies (X-bodies) associated with
cellular infection of BBMV. The X-bodies were visible by light microscopy and electron
microscopy showed that the X-bodies consisted of spherical particles that were identical to
those of BBMV. De Zoeten and Schlegel (1967) studied the ultrastructure of BBMV infected
faba bean leaf cells and found that the mesophyll cells containing virus appeared to be
saturated with virus to the point of crystallisation. In some infected cells, amorphous vesicular
bodies, containing osmiophilic areas and sometimes crystalline virus, could be identified.
They concluded that there was an unusually large amount of virus in the cytoplasm of BBMV
infected cells. More recent authors have used this as a diagnostic characteristic, which
differentiates BBMV from other small, isometric legume viruses (eg. Makkouk et al. 1988a,
Bos et al. 1992, Brunt et al. 1997). However, it should be noted that Comoviruses, such as
BBSV and BBTMV also cause inclusion bodies in infected cells (Brunt et al. 1997). Christie
(1967) developed a rapid staining procedure for differentiating plant virus inclusions in
epidermal strips. This method was used, with minor modifications, by Bos (1969) to observe
inclusion bodies caused by a number of legume viruses and by Makkouk et al. (1988a) to
observe inclusion bodies caused by BBMV.
6.2.2 General items required
1. Epidermal strips from stems, petioles or the underside of leaves.
2. Glass microscope slides, petri dishes, forceps.
3. Staining solution made up of 2 volumes ethylene glycol monomethyl ether, 1 volume 95%
ethanol, and 1 volume distilled water.
4. 1% phloxine made up in the above staining solution.

46
5. 1% methylene blue made up in the above staining solution.
6. Light microscope.
6.2.3 Method
1. Place epidermal strips in equal volumes of 1% phloxine solution and 1% methylene blue
solution (made up as described above) in a Petri dish for 15 mins.
2. Rinse strips in water for a few minutes and mount on slides in water.
3. View under light microscope at magnifications of about 500-750X.
Makkouk et al. (1988a) found that infected cells contained enlarged nucleoli and the
cytoplasm contained granular and often vesicular material which was initially widely
distributed but later in the infection often condensed to form granular, partly vacuolated or
vesiculated inclusion bodies, one or more per cell. They were of varying, sometimes globular,
shape and often exceeded the size of the nucleus. They persisted throughout a 30 day
observation period of after inoculation.
6.3 Indicator plant tests
6.3.1 Introduction
Although BBMV has similar particle morphology to BBSV and BBTMV and they all can cause
similar mottle/mosaic symptoms in legume hosts, BBMV differs from BBSV and BBTMV in
that it has a much wider host range and causes local lesion symptoms on a number of
common indicator plants. The host ranges of BBSV and BBTMV are restricted to the
Fabaceae apart from a few hosts experimental hosts of BBSV in the Chenopodiaceae and
Solanaceae. However, these BBSV hosts do not include the common indicator plants
Chenopodium amaranticolor and Nicotiana clevlandii, which are both hosts of BBMV. A
selection of suitable indicator plant species and their reactions to BBMV are given in 6.3.5. A
comprehensive list of indicator plants are given in Walters and Surin (1973) and Makkouk et
al. (1988a).
6.3.2 General Items required
1. Samples from BBMV-suspect plants, normally fully expanded young leaves or dried or
frozen leaf tissue.
2. BBMV-infected samples and healthy host samples as positive and negative controls.
3. Selected indicator plant species (See 6.3.5), 3-6 plants of each species for each test
sample. (NB. It is probably simpler to grow the indicator plants with 3-6 plants per large pot).
4. Mortars and pestles.

47
5. Sponges, tags for pots, marker pen, wash bottle.
6. Phosphate buffer (See 6.2.4.1), and fine carborundum powder (Grade??).
6.3.3 Method
1. Collect control samples and test samples (young fully expanded leaves, dried leaves,
frozen leaves). Keep fresh material on ice in an Esky.
2. On each indicator plant, sprinkle carborundum powder onto the four youngest fully
expanded leaves.
3. In clean mortar, grind up a small quantity of test plant material with a pestle, using about 5-
10 volumes of 0.05 M phosphate buffer, depending on type of plant material.
4. Using a clean foam square, dip into ground up sap mixture and gently wipe onto leaves of
a set of indicator plants that have been sprinkled with carborundum powder.
5. Leave for a few minutes then wash off powder and buffer mixture from the leaves using a
wash bottle.
6. Label tag with sample identity and date and place in pot.
7. Using new foam square each time, repeat above steps for each sample and controls, as
required.
8. Soak mortar and pestle in bucket with bleach overnight and wash hands with soap.
9. Observe and record indicator plant symptoms regularly over a 4-6 week period.
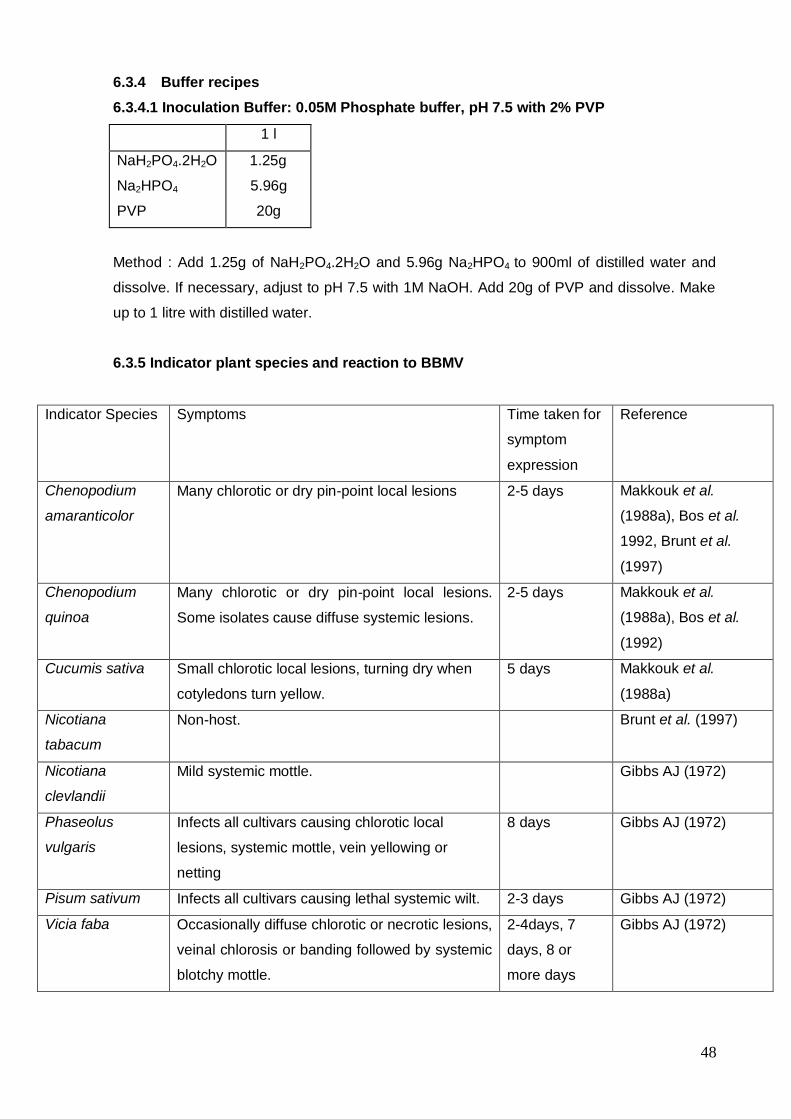
48
6.3.4 Buffer recipes
6.3.4.1 Inoculation Buffer: 0.05M Phosphate buffer, pH 7.5 with 2% PVP
1 l
NaH2PO4.2H2O
Na2HPO4
PVP
1.25g
5.96g
20g
Method : Add 1.25g of NaH2PO4.2H2O and 5.96g Na2HPO4 to 900ml of distilled water and
dissolve. If necessary, adjust to pH 7.5 with 1M NaOH. Add 20g of PVP and dissolve. Make
up to 1 litre with distilled water.
6.3.5 Indicator plant species and reaction to BBMV
Indicator Species Symptoms Time taken for
symptom
expression
Reference
Chenopodium
amaranticolor
Many chlorotic or dry pin-point local lesions 2-5 days Makkouk et al.
(1988a), Bos et al.
1992, Brunt et al.
(1997)
Chenopodium
quinoa
Many chlorotic or dry pin-point local lesions.
Some isolates cause diffuse systemic lesions.
2-5 days Makkouk et al.
(1988a), Bos et al.
(1992)
Cucumis sativa Small chlorotic local lesions, turning dry when
cotyledons turn yellow.
5 days Makkouk et al.
(1988a)
Nicotiana
tabacum
Non-host. Brunt et al. (1997)
Nicotiana
clevlandii
Mild systemic mottle. Gibbs AJ (1972)
Phaseolus
vulgaris
Infects all cultivars causing chlorotic local
lesions, systemic mottle, vein yellowing or
netting
8 days Gibbs AJ (1972)
Pisum sativum Infects all cultivars causing lethal systemic wilt. 2-3 days Gibbs AJ (1972)
Vicia faba Occasionally diffuse chlorotic or necrotic lesions,
veinal chlorosis or banding followed by systemic
blotchy mottle.
2-4days, 7
days, 8 or
more days
Gibbs AJ (1972)

49
7.0 Images
7.1 BBMV symptoms on host plants
Figure 3. BBMV symptoms on faba bean include
mottle/mosaic and veinal chlorosis of the leaves.

50
Figure 4. BBMV symptoms on faba bean include a strong mosaic
resulting in islands of dark and light tissue on the leaves.
(Photo courtesy: Dr S Kumari ICARDA).
Figure 5. BBMV symptoms on faba bean including veinal chlorosis and
mottling of the leaves.
(Photo courtesy: Dr S Kumari, ICARDA).

51
Figure 6. BBMV symptoms on faba bean include interveinal mottle.
(Photo courtesy: Dr S Kumari, ICARDA).
Figure 7. BBMV symptoms on lentil (left) include chlorosis, mottling/mosaic and
sometimes local necrosis. Healthy plant (right).
(Photo courtesy: Dr S Kumari, ICARDA).

52
Figure 8. BBMV symptoms on french bean include systemic yellow
stippling or blotching or interveinal mottle.
(Photo courtesy: Dr S Kumari, ICARDA).

53
7.2 BBMV symptoms on host seed
Figure 9. BBMV symptoms on faba bean seed (right) include necrosis and staining. Healthy
seed (left).
(Photo courtesy: Dr S Kumari, ICARDA)

54
7.3 BBMV vectors and vector symptoms
Figure 10. The BBMV vector, Sitona lineatus, feeding on leaf (note characteristic leaf notching). NB S. lineatus is also a vector of BBSV and BBTMV.
(Photo courtesy: Dr S Kumari, ICARDA, from Agrevo).

55
Figure 11. BBMV vector Sitona lineatus.
(Photo courtesy: Dr S Kumari, ICARDA, from Agrevo).

56
8.0 References and websites
8.1 References
Assou NM (1978). Contribution a l'etude de la feve. Memoire de fin d'etude, Ecole Nationale d'Agriculture de Meknes (Maroc): 34 pp.
Bawden FC, Chauduri RP, Kassanis B (1951). Some properties of broad bean mottle virus. Annals of Applied Biology 38: 774-784.
Borges MLV, Sequeira JC (1988). Protection of legumes in relation to virus infection. Comunicacoes, Jornadas Portuguesas de Proteaginosas: 281-304.
Borges M de LV (1982). Mosaic of faba bean in Portugal. Aetiology and virus-host relations. Garcia de Orta, Estudos Agrononicos 9: 279-284.
Borges M de LV, Louro D (1974). A biting insect as vector of broad bean mottle virus. Agronomia Lusitana 36: 215-216.
Bos L (1969). Inclusion bodies of bean yellow mosaic virus, some less known closely related viruses and beet mosaic virus. . Netherlands Journal of Plant Pathology 75: 137-143.
Bos L, Mahir MAM, Fortass M, Makkouk KM (1992). A mild strain of broad bean mottle virus from faba bean (Vicia faba L.) in the Sudan. Netherlands Journal of Plant Pathology 98: 253-
256.
Bourbah M, Fezzaz M (1979). Contribution a l'etude des viroses de la feve dans la region de Meknes. Memoire de fin d'etude, Ecole Nationale d'Agriculture de Meknes (Maroc): 48 pp.
Botina R, Munoz M de, Guerrero GO (1984). Identification of host plants of broad bean (Vicia faba L.) mottle virus in the Department of Narino. Revista ICA 19: 395-401
Brunt AA, Crabtree K, Dallwitz MJ, Gibbs AJ, Watson L, Zurcher EJ (Eds) Version: 16 th January 1997. Plant viruses online: Descriptions and lists from the VIDE database (BBMV). http://biology.anu.edu.au/Groups/MES/vide/(1997).
CAB International (1999). Crop Protection Compendium. Global Module. CAB International.
Christie RG (1967). Rapid staining procedures for differentiating plant virus inclusions in
epidermal strips. Virology 31:268-271.

57
Cockbain AJ (1971). Weevils, aphids and virus diseases of field beans. Report of the Rothamsted Experimental Station for 1970, pp184-186, pub. Harpenden, Herts., UK. Cockbain AJ, Cook SM, Bowen R (1975). Transmission of broad bean stain virus and Echtes Ackerbohnenmosaik virus to field beans (Vicia faba) by weevils. Annals of Applied Biology
81: 331-339.
Dafalla G, Hussein M (1994). Disease and pest outbreaks. Sudan. Identification of a new virus disease affecting legumes in the Sudan. Arab and Near East Plant Protection Newsletter 18: 30.
Edwardson JR, Christie RG (1991). Handbook of viruses infecting legumes. (CRC Press, Inc.: Florida).
El Afifi SI, El Dougdoug KA (1997). Identification of some mechanically transmitted viruses affecting faba bean plants in Egypt. Annals of Agricultural Science (Cairo) 42: 29-49.
El-Muadhidi MA, Makkouk KM, Kumari SG, Jerjess M, Murad SS, Mustafa RR, Tarik F (2001). Survey for legume and cereal viruses in Iraq. Phytopathologia Mediterranea 40: 224-233.
Erdiller G, Akbas B (1996). Seed transmission of some viruses in chickpea and lentil. Journal of Turkish Phytopathology 25: 93-101.
Fischer HU (1979). The identification and differentiation of virus infections of broad bean. Awamia 57: 41-72.
Fortass M, Bos L (1992). Broad bean mottle virus in Morocco; variability in interaction with food legume species and seed transmission in faba bean, pea and chickpea. Netherlands Journal of Plant Pathology 98: 329-342.
Fortass M, Diallo S (1993). Broad bean mottle virus in Morocco; curculionid vectors, and natural occurence in food legumes other than faba bean (Vicia faba).
Fujisaki K, Hagihara F, Kaido M, Mise K, Okuno T (2003). Complete nucleotide sequence of spring beauty latent virus, a bromovirus infectious to Arabidopsis thaliana. Archives of
Virology: 148: 165-175. Gibbs AJ (1972). Broad bean mottle bromovirus. CMI/AAB Descriptions of Plant Viruses.
No.101. Journal. Gibbs AJ, Smith HG (1970). Broad bean stain virus. CMI/AAB Descriptions of Plant Viruses. No.29. Journal.
Makkouk KM, Dafalla G, Hussein M, Kumari SG (1995). The natural occurrence of chickpea chlorotic dwarf geminivirus in chickpea and faba bean in the Sudan. Journal of Phytopathology 143: 465-466.

58
Makkouk KM, Kumari S (1989). Apion arrogans, a weevil vector of broad bean mottle virus. FABIS Newsletter 24: 26-27.
Makkouk KM, Kumari SG, Al Daoud R (1992). Survey of viruses affecting lentil (Lens culinaris Med.) in Syria. Phytopathologia Meditterranea 31:188-190)
Makkouk KM, Kumari SG (1995). Transmission of broad bean stain comovirus and broad bean mottle bromovirus by weevils in Syria. Zeitschrift fur Pflanzenkrankheiten und Pflanzenschutz 102 136-139.
Makkouk KM, Bos L, Rizkallah A, Azzam OI, Katul L (1988a). Broad bean mottle virus: identification, host range, serology, and occurrence on faba bean (Vicia faba) in West Asia
and North Africa. Netherlands Journal of Plant Pathology 94: 95-212.
Makkouk KM, Bos L, Azzam O, Koumari S, Rizkallah A (1988b). Survey of viruses affecting faba bean in six Arab countries. Arab Journal of Plant Protection 6: 61-63. Makkouk KM, Rizkallah L, Madkour M, El-Sherbeeny M, Kumari SG, Amriti AW, Solh MB (1994). Survey of faba bean (Vicia faba L.) for viruses in Egypt. Phytopathologia Meditterranea 33: 207-211.
Mouhanna AM, Makkouk KM, Ismail ID (1994). Survey of virus diseases of wild and cultivated legumes in the coastal region of Syria. Arab Journal of Plant Protection 12: 12-19.
Murant AF, Abu Salih HS, Goold RA (1974). Viruses from broad bean in the Sudan. Annual Report of the Scottish Horticultural Research Institute, 1973: 67.
Najar A, Makkouk KM, Boudhir H, Kumari SG, Zarouk R, Bessai U, Othman FB, (2000). Viral diseases of cultivated legume and cereal crops in Tunisia. Phytopathologia Mediterranea 39: 423-432.
Naumann I (1993). CSIRO Handbook of Australian Insect Names (6th edn). CSIRO Division of Entomology.
Nene YL, Sheila VK, Sharma SB (1996). A world list of chickpea and pigeonpea pathogens. (ICRISAT: India).
Oufffroukh A (1985). Contribution a la connaissance des viroses des plantes en Algerie: inventaire des virus present chez des legumineuses a longue cosse. Etude approfondie de deux maladies isolees de feve et haricot. These de Docteur de 3e cycle, Universite Pierre et Marie Curie, Paris VI: 101 pp.
Ouizbouben A, Fortass M (1997). Survey of chickpea for viruses in Morocco. Bulletin OEPP 27: 249-254.

59
Rothamsted Experimental Station (1983). Report for 1982, Part 1:200-202 pub. Harpenden, Herts., UK. Rybicki EP, Von Wechmar MB (1981). The serology of the Bromoviruses. 1. Serological interrelationships of the Bromoviruses. Virology 109: 391-402.
Scott HA, Slack SA (1971). Serological relationship of brome mosaic and cowpea chlorotic mottle viruses. Virology 46: 490-492.
Sequeira JC, Borges M de LV (1989). Broad bean mottle virus in Portugal, host range, transmission, serology and virion characterisation. Boletim da Sociedade Broteriana 62: 291-303.
Tadesse N, Ali K, Gorfu D, Yusuf A, Abraham A, Ayalew M, Lencho A, Makkouk KM, Kumari SG (1999). Survey for chickpea and lentil virus diseases in Ethiopia. Phytopathologia Meditterranea 38:149-158. Tinsley TW (1957). UK, Rothamsted Experimental Station: Report for 1956, Part 1:98. Valverde RA (1985). Spring beauty latent virus: A new member of the bromovirus group. Phytopathology 75: 395-398.
Voller A, Bartlett A, Bidwell DE, Clark MF, Adams AN (1976). The detection of viruses by enzyme-linked immunosorbent assay (ELISA). Journal of General Virology 33: 165-167.
Walters HJ, Surin P (1973). Transmission and host range studies of broad bean mottle virus. Plant Disease Reporter 57: 833-836.
Zhao GF (1996). Integrated management of rust and virus disease of red bean. Progress of research on plant protection in China. Proceedings of the third national conference of integrated pest management, Beijing, China, 12-15 November 1996: 140-143.
8.2 Websites
AAB-CMI Descriptions of plant viruses http://www.dpvweb.net/index.php
All the virology on the WWW http://www.virology.net/garryfavwebplant.html
CABI Crop Protection Compendium http://www.cabicompendium.org/cpc/home.asp
Plant viruses online: Descriptions and lists from the VIDE database
http://biology.anu.edu.au/Groups/MES/vide/(1997)

60
Appendix 1. Preliminary Information Data Sheet (Plantplan, 2004)
Date: / /
SUBJECT
Site details:
Ownership:
Location:
Map (lat. & long.):
GPS identifier:
Host plant location (clearly mark plant if necessary):
HOST DETAILS
Species and variety:
Age:
Developmental stage:
DAMAGE
Description of symptoms:
Part of host affected:
Percent incidence:
Percent severity:
DETAILS OF WHEN AND WHERE THE PEST WAS FIRST NOTICED:
RECORDS OF PRODUCT MOVEMENT ON AND OFF DETECTION SITE:
SYMPTOMS / PHOTOGRAPHS:
FURTHER DETAILS OR COMMENTS:

61
Appendix 2. Hygiene
Personnel Hygiene
On entering the paddock, personnel must:
wear protective overalls and rubber boots
prepare footbath of bleach, and spray bottles of methylated spirits brew (95% metho, 5%
water) for use following completion of the inspection
conduct inspections by foot, refer to Machinery for vehicle access
On leaving the paddock, personnel must :
wash boots in foot bath of disinfectant (solution of household bleach 10%) and
remove adhering material, ie soil, with a suitable brush (ie domestic scrubbing brush)
spray boots with methylated sprits brew until soaked
remove overalls and place into a bag and seal
exterior of sample bags to be sprayed/swabbed with methylated spirits brew
spray hands with methylated spirits brew irrespective of whether disposable gloves have
been worn
You must decontaminate before leaving the paddock always.
Overalls must be washed and allowed to completely dry before being used again. If
disposable overalls are used, they can be either washed, or if disposed, sent to land fill or
burnt.
Machinery
No machinery, including vehicles, are to enter paddock without prior approval from the
applicant. Approval to use vehicles in paddock must be included with the application for
access
Decontamination procedures must be followed immediately before leaving the site at
the area identified for decontamination
Decontaminate the machinery by removing all visible lupin trash and wash down with a
high pressure spray using detergent, paying particular attention to the underside, axles,
wheels and tyres. This also includes all hand held tools such as hoes and shovels

62
Personal decontamination procedures must follow the decontamination of machinery
It is recommended that any machinery or vehicle that has entered the paddock is not to be
taken into another green lupin crop this season
Harvest Machinery
In addition to the above requirements, machinery will be cleaned of all seed and
trash remaining. This material will be destroyed in a manner approved by NSW
Agriculture (ie, landfill within quarantine boundary or similar)
It is requested that the harvest machinery not be used to harvest lupins again this
season. If this is not possible, it must be used to harvest two crops, other than
lupins, before it can harvest lupins again.





























