Embed Size (px)
Citation preview

This article was downloaded by: [Northeastern University]On: 05 October 2014, At: 00:46Publisher: Taylor & FrancisInforma Ltd Registered in England and Wales Registered Number: 1072954 Registered office: MortimerHouse, 37-41 Mortimer Street, London W1T 3JH, UK
Zoology and EcologyPublication details, including instructions for authors and subscription information:http://www.tandfonline.com/loi/tzec20
Breeding biology of the European Greenfinch Chlorischloris in the loquat orchards of Algeria (NorthAfrica)Taqiyeddine Bensouilahab, Hafid Brahmiab, Ali Zeraoulab, Zihad Bouslamaa & MoussaHouhamdiba Laboratory EcoSTAq: Ecology of Terrestrial and Aquatic Systems, Faculty of Sciences,Department of Biology, Badji Mokhtar University, BP 12, Annaba 23000, Algeriab Laboratory LBEE: Biology, Water and Environment, Faculty of Nature and Life Sciencesand Earth and Universe Sciences, Department of Nature and Life Sciences, 8 May 1945University, Guelma, AlgeriaPublished online: 11 Jul 2014.
To cite this article: Taqiyeddine Bensouilah, Hafid Brahmia, Ali Zeraoula, Zihad Bouslama & Moussa Houhamdi (2014)Breeding biology of the European Greenfinch Chloris chloris in the loquat orchards of Algeria (North Africa), Zoology andEcology, 24:3, 199-207, DOI: 10.1080/21658005.2014.934514
To link to this article: http://dx.doi.org/10.1080/21658005.2014.934514
PLEASE SCROLL DOWN FOR ARTICLE
Taylor & Francis makes every effort to ensure the accuracy of all the information (the “Content”) containedin the publications on our platform. However, Taylor & Francis, our agents, and our licensors make norepresentations or warranties whatsoever as to the accuracy, completeness, or suitability for any purpose ofthe Content. Any opinions and views expressed in this publication are the opinions and views of the authors,and are not the views of or endorsed by Taylor & Francis. The accuracy of the Content should not be reliedupon and should be independently verified with primary sources of information. Taylor and Francis shallnot be liable for any losses, actions, claims, proceedings, demands, costs, expenses, damages, and otherliabilities whatsoever or howsoever caused arising directly or indirectly in connection with, in relation to orarising out of the use of the Content.
This article may be used for research, teaching, and private study purposes. Any substantial or systematicreproduction, redistribution, reselling, loan, sub-licensing, systematic supply, or distribution in anyform to anyone is expressly forbidden. Terms & Conditions of access and use can be found at http://www.tandfonline.com/page/terms-and-conditions

Breeding biology of the European Greenfinch Chloris chloris in the loquat orchards of Algeria(North Africa)
Taqiyeddine Bensouilaha,b*, Hafid Brahmiab, Ali Zeraoulab, Zihad Bouslamaa and Moussa Houhamdib
aLaboratory EcoSTAq: Ecology of Terrestrial and Aquatic Systems, Faculty of Sciences, Department of Biology, Badji MokhtarUniversity, BP 12, Annaba 23000, Algeria; bLaboratory LBEE: Biology, Water and Environment, Faculty of Nature and Life Sciencesand Earth and Universe Sciences, Department of Nature and Life Sciences, 8 May 1945 University, Guelma, Algeria
(Received 30 January 2014; accepted 6 June 2014)
Few data are available on passerines’ breeding in farmlands in Algeria. The breeding biology of the Greenfinch wasstudied in the loquat orchards to describe the timing of breeding, clutch size, number of nestlings hatched and fledged,assess nesting success and the main causes of nesting attempt failure, and investigate nest placement in the loquat trees.Nests were monitored intensively to quantify the number of nesting attempts and territory densities. The breeding seasonextended from mid-March to late May. Mean clutch size was 4.37 ± 0.04, number of hatchlings was 2.62 ± 0.15, andnumber of fledglings per successful nest was 3.44 ± 0.16. Clutch size differed significantly across the breeding season.The density of breeding pairs was 12.83 ± 0.83 p/ha, and breeding success was lower than that reported in other studiesof finches and passerine birds. Predation was the cause of most nest failures. Loquat orchards seem to be the main suit-able breeding habitat for the Greenfinch, and differences in some breeding biology characteristics between Algeria andEuropean countries may be explained by climate and latitude.
Apie žvirblinių paukščių perėjimą dirbamos žemės laukuose Alžyre duomenų yra mažai. Buvo tiriama žaliukės(Chloris chloris) perėjimo biologija japoninės lokvos (Eriobotrya japonica) soduose: perėjimo laikas, dėties dydis, išsiritu-sių jauniklių skaičius, įvertinta perėjimo sėkmė, pagrindinės nepasisekusio perėjimo priežastys, nustatytos tinkamiausioslizdų sukimo vietos. Intensyviai ieškant lizdų nustatytas perėjimo bandymų skaičius ir teritorinis tankumas. Perėjimosezonas tęsėsi nuo kovo vidurio iki gegužės pabaigos. Vidutinis dėties dydis buvo 4,37 ± 0,04, išsiritusių jauniklių skaičius– 2,62 ± 0,15, lizdą palikusių jauniklių skaičius – 3,44 ± 0,16 kiekvienam sėkmingam lizdui. Dėties dydis perėjimo sezonometu labai skyrėsi. Vidutinis perinčių porų tankumas buvo 12,83 ± 0,83 porų/ha. Nustatyta žaliukių perėjimo sėkmė buvomažesnė nei aprašyta kituose tyrimuose apie kikilinius ir žvirblinius paukščius. Pagrindinė nesėkmingo perėjimo priežastisbuvo plėšrūnai. Nustatyta, kad japoninės lokvos sodai yra tinkamiausias biotopas žaliukės perėjimui. Pagrindinius žaliukiųperėjimo biologijos skirtumus Alžyre ir Europos šalyse lemia klimatas ir geografinė padėtis.
Keywords: Chloris chloris; laying date; clutch size; nest placement; breeding success; farmland
Introduction
The detailed breeding biology descriptions are importantto understanding the evolution of avian reproductivestrategies. Reproductive traits of small open nesting pas-serine species are well documented in Europe (Monk1954; Gil-Delgado and Catalá 1989; Kosinski 2001;Kelleher and O’Halloran 2006; Rodríguez and Moreno2008), North and South America (Martin and Roper1988; Martin and Li 1992; Martin 1993; Sockman 1997;Martin, Scott, and Menge 2000; Mezquida and Marone2001; Mezquida 2004), and Australia (Yom-Tov 1987;Lambert and Kleindorfer 2006; Barea 2008). However,the reproductive biology of most of the passerine speciesremains poorly studied in North Africa, especially inAlgeria.
Small open nesting passerines have long been knownto show certain specific life history features. For mostpasserine species, the selection of an appropriate nest siteis one of the most important factors influencing repro-ductive success (Martin and Roper 1988; Martin 1993;
Sockman 1997; Martin, Scott, and Menge 2000;Mezquida 2004), the majority of species conceal theirnests or build them in inaccessible sites (Nolan 1978;Filliater, Breitwisch, and Nealen 1994) to provide betterprotection against weather and predators given that pre-dation is often the main cause of nest failure for opennesting passerines (Martin and Roper 1988; Martin1993; Hatchwell et al. 1999; Mezquida and Marone2001; Lambert and Kleindorfer 2006; Rodríguez andMoreno 2008). Though some authors suggest that thereis no difference in predation risk between stages of thenesting cycle, Skutch (1949) suggests that increasedparental activity and the noisy begging of broods aroundnests increase the probability of nest predation duringthe nestling period.
The Greenfinch is very widespread in the WesternPalaearctic; it breeds throughout northwest coasts ofAfrica to Russia. It occurs in various kinds of woodlandformations (Cramp and Perrins 1994; Isenmann andMoali 2000).
*Corresponding author. Email: [email protected]
© 2014 Nature Research Centre
Zoology and Ecology, 2014Vol. 24, No. 3, 199–207, http://dx.doi.org/10.1080/21658005.2014.934514
Dow
nloa
ded
by [
Nor
thea
ster
n U
nive
rsity
] at
00:
46 0
5 O
ctob
er 2
014

In the current study, we try to increase informationof the reproductive biology of the European Greenfinchin agricultural landscapes in the south of its distributionarea. Specifically, we attempt to answer the followingquestions: (1) Do European Greenfinches select particu-lar placement to place their nests? (2) Does the timing ofbreeding affect clutch size? (3) Does predation rate differbetween nesting cycles?
Materials and methods
Study areas
The fieldwork was carried out on the loquat (Eriobotryajaponica) orchard (6 ha) of the Guelma region (36°46′N7°28′ E), northeast of Algeria, which is located 60 kmsouth of the Mediterranean Sea and has an elevation of279 m above sea level. The landscape of the Guelmaregion is dominated by forest and agricultural area,which occupy 27 and 65% of the total area, respectively.The climate of the region is sub-humid, mild and rainyin winter, and hot in summer, the annual average temper-ature is about 17.3 °C and ranges between 4 °C in winterand 35.4 °C in summer. The annual mean rainfall is654 mm.
Nest searching and monitoring
We located and monitored nests from early February tolate July in 2013. We found nests by systematicallysearching in the trees. We also located a small proportionof nests fortuitously by accidentally flushing an adult offthe nest while walking past and by using parental behav-iors as cues to location of nests (Martin and Geupel1993). We marked nests by using survey numbered flagsplaced on the trunk at least 1 m from the ground forrepeated location of nests.
Nests were found at different stages of development,including nest building, incubation, and brood-rearing.Once nests were located, we monitored them every2–4 days until they either fledged young or failed todetermine the laying dates, clutch size, brood size, incu-bation and nestling periods, hatching and fledging dates,and the number of fledglings. We waited for parents toleave nests of their own accord before checking nestcontents and took great care during successive nestchecks in an attempt to minimize time spent in thevicinity of active nests to reduce any effects of observeractivity on the probability of nest predation.
If the first-egg date was unknown, it was estimatedin conformance with observations of nests where theexact dates of laying, hatching, and/or fledging wereknown, assuming that incubation lasted 12 days plus oneday for each egg laid (the incubation period wascalculated from laying the last egg to hatching the firstone) and the average period of hatching to fledging was18 days, considering the information provided by thisstudy.
The maximum number of simultaneously active nestsduring a successive five-day period was used to calculatethe density of pairs.
Breeding success
A successful nest was one where at least one nestlingfledged based on direct observation of fledged nestlingsand/or observation of an empty nest with black andwhite droppings disposed around its periphery, whichcoincided with the post-fledging period. Hatching suc-cess was calculated as the total number of hatchlingsfrom the total number of eggs, fledging success as thetotal number of fledglings from the total number ofhatchlings, and breeding success as the total number offledglings from the total number of eggs laid per nest.
For failed nests, the date and, where possible, thecauses of failure were noted. The following cases wereconsidered a failure due to predation: observation of per-forated eggs or fragments of broken eggs in the nest, amessy nest with eggshell fragments off the nest and/oron the ground, and disappearance of eggs before hatch-ing or chicks before fledging under favorable climaticconditions.
Failures due to adverse weather conditions were rec-ognized as nests deserted with cold eggs or dead chicksafter rainfall.
Daily nest survival rate
Nest success was estimated by a second measure usingMayfield’s method (Mayfield 1961). The confidence lim-its were calculated from Johnson’s method (Johnson1979). Daily nest survival rates were calculated for: (1)egg period, which represents the period from when thefirst egg was laid to the hatch of the first one; (2) incu-bation period, which represents the period from whenthe last egg was laid to the hatch of the first one; (3)nestling period, which represents the period from whenthe first chick hatched to the fledge of the last one; and(4) overall period, which represents the whole nestingperiod from laying to fledging (see Kelleher andO’Halloran 2006). An average laying period of five daysand incubation and nestling periods of 12 and 18 days,respectively, were used (considering the informationprovided in this study).
Estimating productivity
The number of fledglings produced per breeding attempt(see Peach, Siriwardena, and Gregory 1999; Siriwardenaet al. 2000) was calculated as follows:
FPA ¼ CS� CER� ð1� EFRÞEP � ð1� NFRÞNP
where FPA is the number of fledglings produced perbreeding attempt, CS is clutch size, CER is chick eggratio, EFR and NFR are daily nest failure rates of the
200 T. Bensouilah et al.
Dow
nloa
ded
by [
Nor
thea
ster
n U
nive
rsity
] at
00:
46 0
5 O
ctob
er 2
014

egg and nestling period, respectively, and EP and NP arethe lengths of the egg and nestling period in days,respectively.
The number of hatchlings produced per breedingattempt (see Paradis et al. 2000; Kelleher and O’Halloran2006) was calculated as follows:
ROA ¼ BS� ð1� IFRÞIP � ð1� NFRÞNP
where the reproductive output per attempt (ROA) is thenumber of hatchlings produced per breeding attempt, BSis the maximum brood size at hatching, IFR and NFRare daily nest failure rates of the incubation and nestlingperiod, respectively, and IP and NP are the lengths ofthe incubation and nestling period in days, respectively.
Nest site characteristics
Nest site characteristics were recorded on the last visit toa nest after the nest failed or nestlings fledged. We col-lected the following variables for each nest tree: nestheight above ground (NHG), distance from the nest to thetrunk (DNT), distance from the nest to the external partof the canopy (DNEC), distance from the nest to thelowest part of the canopy (DNLC), nest tree height(NTH), nest branch length (NBL), nest relative verticalposition in the canopy (NRVPC = [DNLC/[(NTH –NHG) + DNLC]] × 100), and nest position index (NPIC= [DNT/(DNT + DNEC)] × 100) (Mezquida 2004;Rodríguez and Moreno 2008; Hanane and Baamal 2011).We also recorded with a digital calliper (0.01 mm accu-racy) external and internal nest cup diameter and cupdepth.
Egg size
We also measured egg length (L) and breadth (B) usinga digital calliper, and all eggs were weighted with a digi-tal balance (0.1 g accuracy). Egg volume was calculatedusing Hoyt’s (1979) method: V = 0.51 × L × B2. The coef-ficient of variation was calculated using equation CV =(SD/mean) × 100.
Weather
The influence of weather on incubation and nestlingperiods, nest survival and outcome was examined.Daily rainfall (mm) and temperature (°C) were availablefrom the National Office of Weather (Guelma station)for the breeding seasons (March–June) of 2013. Thetwo climatic factors were strongly correlated and onlytemperature was significantly related to the seasonalday number. Thus, temperature increased significantlywith season (r = 0.53, n = 122, p < 0.0005), and therewas a negative correlation between temperature andrainfall (r = –0.41, n = 122, p < 0.0005). These correla-tions are based on climatic data from 122 days duringthe breeding season.
Statistical analysis
All variables were tested for homoscedasticity usingLevene’s test and normality using a one-sampleKolmogorov–Smirnov test at a 95% confidence level.Where possible, non-normally distributed variables werelog10 transformed to improve the normality of the data.We tested for correlation between weather variables andboth egg laying dates and incubation and nestling peri-ods using Pearson’s correlation coefficients. Also, weused Pearson’s correlation to examine the relationshipsbetween nest site characteristics. We used quadraticregression to test for seasonal change in clutch size,number of fledglings, and fledging success. In addition,we used linear regression to test for seasonal variation inboth incubation and nestling period, the number ofhatchlings and hatching success. Results are presented asmean ± SE. All tests are two tailed, and results were con-sidered significant at p < 0.05. Statistical analyses weremade using SPSS Version 19.
Results
Territory densities
A total of 185 nests were found, three were deserted atthe building stage, and 70 were predated. The meandensity of breeding pairs was 12.83 ± 0.83 p/ha.
Nest placement
Nest heights ranged between 1.51 and 3.46 m (mean =2.34 ± 0.02 m; n = 177), and nest height was positivelycorrelated with tree height (r = 0.62, n = 177, p < 0.0005).Distance between nest place and the trunk varied from0.27 to 2.03 m (mean = 1.08 ± 0.03 m, n = 177), and dis-tance from the nest to the trunk was positively correlatedwith tree height (r = 0.15, n = 177, p < 0.003); these dis-tances were longer as the length of the branchesincreased (r = 0.53, n = 177, p < 0.0005). Nest site char-acteristics are shown in Table 1.
The majority of nests (79.1%) had a height between2 and 3 m, and 48.5% were located between 1 and 1.5 mfrom the trunk. Both nest relative vertical position in thecanopy and nest position index significantly differedfrom a uniform distribution (Kolmogorov–Smirnovtest, Z = 0.082, n = 177, p < 0.006; Z = 0.092, n = 177,p < 0.001, respectively). The relative vertical positionwas located in the upper part of the canopy (median =59.9%, range 26.5–84.2%). Half of the nests had the rel-ative vertical position in the canopy between 51.1 and69.1%. However, the relative position in the brancheswas located in the midway point between the trunk anddistal part of the branches (median = 51.4%, range13.0–79.9%), and half of the nests had the relative posi-tion in the branches between 36.1 and 59.6% (Figure 1).There was a positive correlation between canopy diame-ter (length of branches) and the relative position index(r = 0.18, n = 177, p = 0.016), as nests were closer to the
Zoology and Ecology 201
Dow
nloa
ded
by [
Nor
thea
ster
n U
nive
rsity
] at
00:
46 0
5 O
ctob
er 2
014

center of canopy diameter. These results showed thatGreenfinches placed their nests in the upper half of thecanopy; this finch usually selected vertical and medianparts of the branches for nesting, but not the canopyperiphery.
Three nests were reused after egg predation; we alsoobserved once a female adapting a nest of the SerinSerinus serinus and adding nesting material a few daysafter the fledging of chicks of the Serin.
Timing of breeding
Egg-laying took place from mid-March to late May(Figure 2). Generally, the breeding season spread over aperiod of 11 weeks (77 days), and the number of clutcheslaid during successive five-day periods showed three peaks(Figure 2): during the first week of April (n = 34, 18.7%),the third week of April (n = 20, 11.0%), and the secondweek of May (n = 18, 9.9%).
Clutch size
Eggs were laid on consecutive days (approximately 24 hinterval) in the first hours following sunrise. The meanclutch size was 4.37 ± 0.04 (n = 182), the highest in May
(4.54 ± 0.07, n = 50), and the lowest in March (4.14 ±0.07, n = 58). Clutch size varied from three (6.6%) tofive (44.0%) eggs, and most clutches had four eggs(49.4%).
The seasonal pattern shows seasonal increase with apeak in the mid-season and then a decline (Figure 3).The quadratic equation was significant (r2 = 0.154;F = 16.353; p < 0.0005).
Egg size
The average weight for 348 eggs was 2.003 ± 0.007 g.The average length was 2.001 ± 0.004 cm and width was1.426 ± 0.002 cm. The mean volume of eggs was 2.080± 0.009 cm3 (Table 2). Egg measurements usuallyshowed little variation. However, volume varied morethan mass (Table 2).
Incubation and nestling periods
Incubation was undertaken by females. The incubationperiod varied from 10 to 14 days, with a mode of
Table 1. Greenfinch nest site characteristics.
Variables Mean ± SE (n = 177) Range (n = 177) Successful (n = 91) Unsuccessful (n = 86)
ENCD (cm) 13.09 ± 0.13 9.87–16.78 12.89 ± 0.19 13.29 ± 0.17INCD (cm) 7.49 ± 0.05 6.04–8.89 7.40 ± 0.08 7.58 ± 0.07CD (cm) 4.87 ± 0.04 3.79–6.26 4.86 ± 0.06 4.89 ± 0.07NHG (m) 2.34 ± 0.02 1.51–3.46 2.35 ± 0.04 2.33 ± 0.04DNT (m) 1.08 ± 0.03 0.27–2.03 1.04 ± 0.04 1.12 ± 0.04DNEC (m) 1.16 ± 0.02 0.46–1.96 1.17 ± 0.04 1.14 ± 0.04DNLC (m) 1.11 ± 0.02 0.39–1.73 1.11 ± 0.03 1.11 ± 0.02NTH (m) 3.10 ± 0.02 2.16–3.91 3.12 ± 0.03 3.09 ± 0.03NBL (m) 2.24 ± 0.02 1.18–3.17 2.21 ± 0.03 2.26 ± 0.04NRVPC (%) 59.73 ± 0.96 26.48–84,15 59.35 ± 1.36 60.13 ± 1.36NPIC (%) 47.74 ± 1.29 12.98–79.91 46.59 ± 1.90 48.95 ± 1.73
Note: ENCD, external nest cup diameter; INCD, internal nest cup diameter; CD, cup depth; NHG, nest height above ground; DNT, distance from thenest to the trunk; DNEC, distance from the nest to the external part of the canopy; DNLC, distance from the nest to the lowest part of the canopy;NTH, nest tree height; NBL, nest branch length; NRVPC, nest relative vertical position in the canopy; NPIC, nest position index.
Figure 1. Vertical and horizontal relative positions of nests.
Figure 2. Distribution of Greenfinch egg-laying dates insuccessive five-day periods in a loquat orchard.
202 T. Bensouilah et al.
Dow
nloa
ded
by [
Nor
thea
ster
n U
nive
rsity
] at
00:
46 0
5 O
ctob
er 2
014
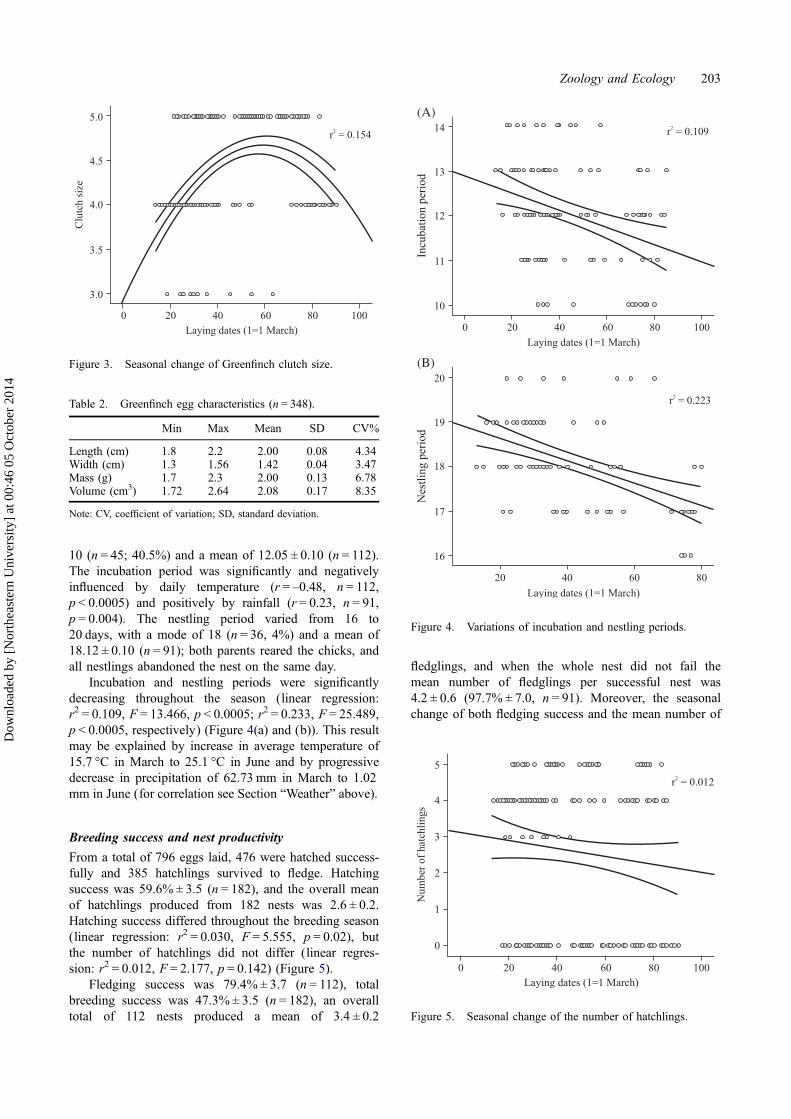
10 (n = 45; 40.5%) and a mean of 12.05 ± 0.10 (n = 112).The incubation period was significantly and negativelyinfluenced by daily temperature (r = –0.48, n = 112,p < 0.0005) and positively by rainfall (r = 0.23, n = 91,p = 0.004). The nestling period varied from 16 to20 days, with a mode of 18 (n = 36, 4%) and a mean of18.12 ± 0.10 (n = 91); both parents reared the chicks, andall nestlings abandoned the nest on the same day.
Incubation and nestling periods were significantlydecreasing throughout the season (linear regression:r2 = 0.109, F = 13.466, p < 0.0005; r2 = 0.233, F = 25.489,p < 0.0005, respectively) (Figure 4(a) and (b)). This resultmay be explained by increase in average temperature of15.7 °C in March to 25.1 °C in June and by progressivedecrease in precipitation of 62.73 mm in March to 1.02mm in June (for correlation see Section “Weather” above).
Breeding success and nest productivity
From a total of 796 eggs laid, 476 were hatched success-fully and 385 hatchlings survived to fledge. Hatchingsuccess was 59.6% ± 3.5 (n = 182), and the overall meanof hatchlings produced from 182 nests was 2.6 ± 0.2.Hatching success differed throughout the breeding season(linear regression: r2 = 0.030, F = 5.555, p = 0.02), butthe number of hatchlings did not differ (linear regres-sion: r2 = 0.012, F = 2.177, p = 0.142) (Figure 5).
Fledging success was 79.4% ± 3.7 (n = 112), totalbreeding success was 47.3% ± 3.5 (n = 182), an overalltotal of 112 nests produced a mean of 3.4 ± 0.2
fledglings, and when the whole nest did not fail themean number of fledglings per successful nest was4.2 ± 0.6 (97.7% ± 7.0, n = 91). Moreover, the seasonalchange of both fledging success and the mean number of
Figure 3. Seasonal change of Greenfinch clutch size.
Table 2. Greenfinch egg characteristics (n = 348).
Min Max Mean SD CV%
Length (cm) 1.8 2.2 2.00 0.08 4.34Width (cm) 1.3 1.56 1.42 0.04 3.47Mass (g) 1.7 2.3 2.00 0.13 6.78Volume (cm3) 1.72 2.64 2.08 0.17 8.35
Note: CV, coefficient of variation; SD, standard deviation.
(A)
(B)
Figure 4. Variations of incubation and nestling periods.
Figure 5. Seasonal change of the number of hatchlings.
Zoology and Ecology 203
Dow
nloa
ded
by [
Nor
thea
ster
n U
nive
rsity
] at
00:
46 0
5 O
ctob
er 2
014

fledglings shows a seasonal increase and then a decline(Figure 6). The quadratic equation was significant(quadratic regression: r2 = 0.108; F = 6.585; p = 0.002;r2 = 0.081; F = 4.828; p = 0.01, respectively). The overallnest productivity estimation was 2.32 fledglings pro-duced per nesting attempt and 2.46 hatchlings producedper nesting attempt.
Daily nest survival rates
The overall daily nest survival rate was 0.969 ± 0.0012for the egg period, 0.959 ± 0.0025 for the incubation per-iod, and 0.988 ± 0.0006 for the nestling period. Theoverall nest survival rate over the whole nesting periodwas 49.3%. No significant difference was observedbetween nest outcome and daily temperature or rainfall.
Cause of nest failure
Ninety-four nesting attempts failed due to the followingreasons: (1) nest deserted at the building stage (n = 3),(2) eggs eaten by predators (n = 60), (3) nestlingpredation (n = 10), (4) nestling died as a result of badweather or for unknown reasons (n = 7), (5) eggs cold-ness as a result of bad weather (n = 4), (6) nest disap-peared (n = 6), and (7) nest dropped (n = 4).
Most nest failures were caused by predators (74.5%)at the egg (63.9%) or nestling (10.6%) stage. Althoughpredator identity could not be determined reliably but45% (n = 27) of egg predation were due to the Europeangreen lizard Lacerta viridis. However, in 10 of predationevents, predators were directly recorded; for the othercases, the identity of the predator was inferred from nestsand egg appearances. We did not observe any potentialmammalian predators; however, we did not make spe-cific searches for them nor visited the study site at night;also avian predators were virtually absent but snakeswere seen at the study site.
Discussion
The results of this study provide basic information onthe breeding biology of the European Greenfinch occu-pying farmland in Algeria. Though some authors referredto their breeding at the Orange Grove (Gil-Delgado andCatalá 1989; Isenmann and Moali 2000) and olive orch-ard (Isenmann and Moali 2000), to our knowledge, thisis the first study realized in the loquat orchard. The terri-tory density of the European Greenfinch at Guelma’sloquat orchards (12.83 ± 0.83 p/ha) is much higher thanthat in urban habitats (5.4 nests per 10 ha) as reported byKosinski (2001) and in forest (2.73 nests per 10 ha) asrevealed by Kouidri (2013). Certainly, the presence ofhedgerows (Stoate, Moreby, and Szczur 1998; Macleod,Parish, and Hubbard 2004), herbaceous vegetation(Stoate and Szczur 1994) and a large extent of cerealcrops (Hanane and Baamal 2011) are known to influencethe breeding density of several passerine species.
The breeding season started two weeks earlier thanthat reported for Greenfinch by Kouidri (2013) in Algeriaand about four to five weeks earlier than in Europeancountries (Monk 1954; Kosinski 2001; Gil-Delgado et al.2005), but it seems that these results are quite similar tothose in North Africa (Heim de Balsac and Mayaud1962; Isenmann and Moali 2000; Isenmann et al. 2005).In addition, the beginning of the breeding season varieswith latitude and altitude. It starts early in southern lati-tudes (Heim de Basac and Mayaud 1962; Gnielka 1986;Gil-Delgado and Catalá 1989; Cramp and Perrins 1994)and late in high altitudes (Chabi and Isenmann 1997). Itis also due to annual variations because it is mainlyinfluenced by meteorological factors and food availabil-ity (Meijer et al. 1999; Durant et al. 2007).
Moreover, the laying period (77 days) is longer thanthat reported by Kouidri (2013) (47 days) and relativelyshorter than previously reported in Europe (88.2 ± 10.8days, Kosinski 2001). Egg-laying activity showed threepeaks in contrast to other studies where only the firstpeak was observed (Gnielka 1986; Kouidri 2013), and ifa second peak was found, it was not clearly marked(Monk 1954; Kosinski 2001).
Compared with other studies, the mean clutch size inthis study was similar to that reported by Gil-Delgadoet al. (2005) in Spain (4.4 ± 0.83, n = 370), higher thanthat reported in Algeria (4.26 ± 0.63, n = 50, Isenmannand Moali 2000; 4.09 ± 0.75, n = 37, Kouidri 2013), andsignificantly smaller to that reported in European coun-tries (4.83 ± 0.03, n = 593, Monk 1954; 5.07 ± 0.74,n = 215, Kosinski 2001). Differences between our resultsand the study of Kouidri (2013), which was conductedat 1593 m above sea level, may be due to a slightdecrease of clutch size with increasing altitude. Further-more, these results can be explained by the decrease ofclutch sizes with latitudes (Perrins and Birkhead 1983).In this study, clutch size increased progressively to amid-season maximum peak and then decreased until theend of the breeding season which is consistent withFigure 6. Seasonal change of the number of fledglings.
204 T. Bensouilah et al.
Dow
nloa
ded
by [
Nor
thea
ster
n U
nive
rsity
] at
00:
46 0
5 O
ctob
er 2
014

multi-brooded species (Monk 1954; Gnielka 1986;Gil-Delgado and Catalá 1989; Crick, Gibbons, andMagrath 1993; Kosinski 2001; Gil-Delgado et al. 2005).
Sharing the incubation duties by both sexes is recog-nized in some passerine species, but in the Greenfinchonly females incubated eggs, which is consistent withother finch species, and the mean incubation periodfound by this study can be compared to that given byMonk (1954), Cramp and Perrins (1994), and Kouidri(2013).
The average nest height in this study was similar tothat observed in others studies (Hudec 1983; Gnielka1986; Kouidri 2013). It is often stressed that Greenfinchnests are usually placed between 2 and 3 m aboveground in urban environment, forest habitat, and agricul-tural landscape despite the presence of tall trees availablefor nesting (Kosinski 2001; Kouidri 2013; this study).The preference for nest placement in the upper half ofthe canopy of the nest tree was also found in otherpasserines (Barea 2008). We found that nests were morethan thrice likely to be placed in the upper half of thecanopy than in the lower half; we suggest that this pref-erence could increase nest concealment. Microclimateselection may also affect nest placement.
In fact, the Greenfinches do not place their nests atrandom. Nest sites may be selected with reference to pre-vious experience of unsuccessful nesting due to coldweather or predation as nest placement influences theowner’s fitness (Snow 1978; Hatchwell et al. 1999), andseveral studies suggest that variation in nest sites mayaffect the vulnerability to predation (Martin and Roper1988; Martin and Li 1992; Sockman 1997). Furthermore,several studies have shown that nest height affects nest-ing success because high off-ground nests are inaccessi-ble to most ground-dwelling predators (Martin 1993;Lambert and Kleindorfer 2006). In our case, there wasno difference in nest height and all nest site characteris-tics between successful and unsuccessful nests (Table 2).Other studies have also failed to find differences in nestsuccess related to nest placement (Filliater, Breitwisch,and Nealen 1994; Howlett and Stutchbury 1996;Rodríguez and Moreno 2008; Hanane and Baamal 2011).
Our data show that the overall mean of hatchlingsproduced was lower than that reported by Monk (1954),Kosinski (2001), and Kouidri (2013). However, this lowvalue is mainly influenced by the high rate of egg preda-tion, but not by low fertility of eggs as previous studiesreferred (Heim de Balsac and Mayaud 1962; Kouidri2013). With regard to the number of fledglings, theirmean number per successful nest was similar to thatfound in European countries such as Czechoslovakia –4.26 (Hudec 1983), Eastern Germany – 4.2 (Blumel1983), and Poland – 4.29 (Kosinski 2001); breeding suc-cess was also quite similar to that reported in Europeand varied between 40.7 and 52.9% (Monk 1954;Blumel 1983; Kosinski 2001).
The number of fledglings varied seasonally like inother multi-brooded species (Kosinski 2001; Kelleher
and O’Halloran 2006), and it was significantly higher inMay than in April and June. Furthermore, the number offledglings produced per nesting attempt in this study waslower as a result of a higher predation rate; the valuesobserved in Europe varied from 2.04 (Kosinski 2001) to2.21 (Blumel 1983) and 2.55 (Monk 1954). Breedingsuccess was slightly lower when compared with themean reported for other canopy-nesting species (Martin1993; Rodríguez and Moreno 2008; Kouidri 2013).
Nest survival was closer to that reported in Europe(Snow and Mayer-Gross 1967; Gnielka 1986; Kosinski2001). Daily egg failure was higher than that reported inEurope for some finch species breeding in farmland, butnestling failure was slightly lower (Siriwardena et al.2000).
We did not observe any case of nest abandonment ornestling starvation, which indicates that food resourceswere abundant in our site. Gil-Delgado and Catalá(1989) reported that predation accounted for 24% of theloss; this value is low compared with the finding fromPoland – 38.7% (Kosinski 2001) and from this study –38.5%. The average value of predation rate was similarto other canopy-nesting birds (Martin 1993; Rodríguezand Moreno 2008). It should be stressed that small open-cup nesting passerines in general suffer high rates of nestpredation (Martin and Roper 1988; Martin 1993;Hatchwell et al. 1999; Mezquida and Marone 2001;Lambert and Kleindorfer 2006). In addition, a higherlevel of predation during the incubation phase was alsofound in other studies of open-cup nesting passerines.Indeed, uniparental incubators at exposed nests incur agreater nest predation during the incubation stage(Martin, Scott, and Menge 2000; Mezquida and Marone2001; Kelleher and O’Halloran 2006; Lambert andKleindorfer 2006), and open nesting species in generalhad a higher failure during the incubation period thancavity-nesting species had (Martin and Li 1992); thisresult may be explained by visual conspicuousness ofnests. Under a high predation pressure the nests that aremost vulnerable to predation are likely to be destroyedby predators early in the nesting cycle, whereas thosenests that are difficult for predators to detect survive tolater stages (Nolan 1978; Filliater, Breitwisch, andNealen 1994; Martin, Scott, and Menge 2000; Lambertand Kleindorfer 2006). Moreover, in this study, nest pre-dation did not increase in the nestling phase despiteincrease of parental activity and chick calls. Combinedthese results argue against Skutch’s (1949) hypothesis.
AcknowledgmentsWe are especially grateful to Mr J.A. Gil-Delgado (ValenciaUniversity, Spain) for provided valuable suggestions whichgreatly improved this paper. We would like to thank MissF. Abdaoui (8 May 1945 University, Guelma, Algeria) and MrS. Hanane (Forestry Research Centre, Rabat, Morocco) for theircomments on an earlier version of this manuscript. Also, wethank the national weather office of Guelma station and thedirection of agriculture for their help.
Zoology and Ecology 205
Dow
nloa
ded
by [
Nor
thea
ster
n U
nive
rsity
] at
00:
46 0
5 O
ctob
er 2
014

ReferencesBarea, L. P. 2008. “Nest-site Selection by the Painted Honey-
eater (Grantiella picta), a Mistletoe Specialist.” Emu 108:213–220.
Blumel, H. 1983. Der Grünling. Die Neue Brehm-Bücherei [TheGreen Compact]. Wittenberg Lutherstadt: A. ZiemsenVerlag.
Chabi, Y., and P. Isenmann. 1997. “La reproduction de lamésange bleue Parus caeruleus ultramarinus dans dessubéraies quercus suber á trois différentes altitudes en Alg-érie [The Breeding Biology of Blue Tit Parus CaeruleusUltramarinus in Coak Oaks Quercus Suber at Three Differ-ent Altitudes in Algeria].” Alauda 55: 13–18.
Cramp, S., and C. M. Perrins. 1994. Handbook of the Birds ofEurope, the Middle East and North Africa. The Birds ofthe Western Paleartic. Vol. 8. Crows to Finches. London:Oxford University Press.
Crick, H. Q. P., D. W. Gibbons, and R. D. Magrath. 1993.“Seasonal Changes in Clutch Size in British Birds.” TheJournal of Animal Ecology 62: 263–273.
Durant, J. M., D. Ø. Hjermann, G. Ottersen, and N. C. Stenseth.2007. “Climate and the Match or Mismatch between Preda-tor Requirements and Resource Availability.” ClimateResearch 33: 271–283.
Filliater, T. S., R. Breitwisch, and P. M. Nealen. 1994. “Preda-tion on Northern Cardinal Nests: Does Choice of Nest SiteMatter?” The Condor 96: 761–768.
Gil-Delgado, J. A., and M. C. Catalá. 1989. “El verderon-común (Carduelis chloris): población nidificante, tamańode la puesta y éxitore productor. Un ejemplode estudios alargo plazo [The European Greenfinch (Carduelis Chloris):Breeding Population, Size of the Set and Reproductive Suc-cess. One Example of Long-term Studies].” MediterráneaSer. Biol. 11: 121–132.
Gil-Delgado, J. A., E. Marco, M. Paredes, and C. Vives-Ferrandiz.2005. “Seasonal Clutch Size Variation of Multi-brooded BirdSpecies: Comparisons between Breeding Season andLatitudes.” Ibis 147: 206–212.
Gnielka, R. 1986. “Daten zur Brutbiologie des Grünfinken(Carduelis chloris) nach Nestkarten aus dem Bezirk Halle[Data on the Breeding Biology of the Greenfinch (CarduelisChloris) to Nest Cards from the Hall County].” Beitr.Vogelkd 4: 235–244.
Hanane, S., and L. Baamal. 2011. “Are Moroccan Fruit orChards Suitable Breeding Habitats for Turtle Doves Strep-topeliaturtur?” Bird Study 58: 57–67.
Hatchwell, B. J., A. F. Russell, M. K. Fowlie, and D. J. Ross.1999. “Reproductive Success and Nest-site Selection in aCooperative Breeder: Effect of Experience and a DirectBenefit of Helping.” The Auk 116 (2): 355–363.
Heim de Balsac, H., and N. Mayaud. 1962. Les Oiseaux duNord-Ouest de l’Afrique: Distribution géographique, écolo-gie, migration, reproduction [The Birds of North-WestAfrica Geographical Distribution, Ecology, Migration,Reproduction]. Paris: Le chevalier.
Howlett, J. S., and B. J. Stutchbury. 1996. “Nest Concealmentand Predation in Hooded Warblers: Experimental Removalof Nest Cover.” The Auk 113: 1–9.
Hoyt, D. F. 1979. “Practical Methods of Estimating Volumeand Fresh Weight of Bird Eggs.” The Auk 96: 73–77.
Hudec, K. 1983. Fauna CSSR. Ptáci [Aves] 3/1. Praha:Academia.
Isenmann, P., and A. Moali. 2000. Oiseaux D’Algérie [Birds ofAlgeria]. Paris: SEOF.
Isenmann, P., T. Gaultier, A. El-Hili, H. Azafzaf, H. Dlensi,and M. Smart. 2005. Oiseaux De Tunisie [Birds of Tuni-sia]. Paris: SEOF.
Johnson, D. H. 1979. “Estimating Nest Success: The MayfieldMethod and an Alternative.” Auk 96: 651–661.
Kelleher, K. M., and J. O’Halloran. 2006. “Breeding Biologyof the Song Thrush Turdus Philomelos in an Island Popula-tion: Capsule Breeding Characteristics were Explained byAgricultural Intensification, Climate, Latitude and InsularSyndrome.” Bird Study 53: 142–155.
Kosiński, Z. 2001. “The Breeding Ecology of the GreenfinchCarduelis chloris in Urban Conditions (Study in Krotoszyn,W Poland).” Acta Ornithologica 36: 111–121.
Kouidri, M. 2013. Contribution à l’étude de l’avifaune niche-use de la région de l’Atlas saharien [Contribution to theStudy of the Breeding Birds of the Saharian Atlas], ThèseDoct: University Annaba.
Lambert, S., and S. Kleindorfer. 2006. “Nest Concealment butNot Human Visitation Predicts Predation of New HollandHoneyeater Nests.” Emu 106: 63–68.
Macleod, C. J., D. M. B. Parish, and S. F. Hubbard. 2004.“Habitat Associations and Breeding Success of the Chaf-finch Fringilla Coelebs: Capsule Population Trends forChaffinch on Farmland are Unlikely to be Explained bytheir Preference for Non-crop Habitats Alone.” Bird Study51: 239–247.
Martin, T. E. 1993. “Nest Predation among Vegetation Layersand Habitat Types: Revising the Dogmas.” The AmericanNaturalist 141: 897–913.
Martin, T. E., and J. J. Roper. 1988. “Nest Predation and Nest-site Selection of a Western Population of the HermitThrush.” The Condor 90: 51–57.
Martin, T. E., and P. Li. 1992. “New Perspectives on Life his-tory Traits of Open–versus Cavity-nesting Birds.” Ecology73: 579–592.
Martin, T. E., and G. R. Geupel. 1993. “Nest-monitoring Plots:Methods for Locating Nests and Monitoring Success.”Journal of Field Ornithology 64: 507–519.
Martin, T. E., J. Scott, and C. Menge. 2000. “Nest PredationIncreases with Parental Activity: Separating Nest Site andParental Activity Effects.” Proceedings of the Royal SocietyB: Biological Sciences 267: 2287–2293.
Mayfield, H. 1961. “Nesting Success Calculated from Expo-sure.” Wilson Bulletin 73: 255–261.
Meijer, T., U. Nienaber, U. Langer, and F. Trillmich. 1999.“Temperature and Timing of Egg-laying of European Star-lings.” The Condor 101: 124–132.
Mezquida, E. T. 2004. “Nest Site Selection and Nesting Suc-cess of Five Species of Passerines in a South AmericanOpen Prosopis Woodland.” Journal of Ornithology 145:16–22.
Mezquida, E. T., and L. Marone. 2001. “Factors AffectingNesting Success of a Bird Assembly in the CentralMonte Desert, Argentina.” Journal of Avian Biology 32:287–296.
Monk, J. F. 1954. “The Breeding Biology of the Greenfinch.”Bird Study 1: 2–14.
Nolan, V. 1978. “The Ecology and Behavior of the PrairieWarbler Dendroica Discolor.” American Ornithologists’Union Ornithological Monographs 26: 1–595.
Paradis, E., S. R. Baillie, W. J. Sutherland, C. Dudley, H. Q. P.Crick, and R. D. Gregory. 2000. “Large-scale Spatial Varia-tion in the Breeding Performance of Song Thrushes Turdusphilomelos and Blackbirds T. merula in Britain.” Journal ofApplied Ecology 37: 73–87.
Peach, W. J., G. M. Siriwardena, and R. D. Gregory. 1999.“Long-term Changes in Over-winter Survival Rates Explainthe Decline of Reed Buntings Emberiza schoeniclus inBritain.” Journal of Applied Ecology 36: 798–811.
Perrins, C. M., and T. R. Birkhead. 1983. Avian Ecology.Glasgow: Blackie.
Rodríguez, F., and Ángel C. Moreno. 2008. “Breeding Biologyof the Endangered Blue Chaffinch Fringilla Teydea Polatzeki
206 T. Bensouilah et al.
Dow
nloa
ded
by [
Nor
thea
ster
n U
nive
rsity
] at
00:
46 0
5 O
ctob
er 2
014

in Gran Canaria (Canary Islands).” Acta Ornithologica 43:207–215.
Siriwardena, G. M., S. R. Baillie, H. Q. P. Crick, and J. D.Wilson. 2000. “The Importance of Variation in the Breed-ing Performance of Seed-eating Birds in Determining TheirPopulation Trends on Farmland.” Journal of AppliedEcology 37: 128–148.
Skutch, A. F. 1949. “Do Tropical Birds Rear as Many Youngas They can Nourish?” Ibis 91: 430–455.
Snow, D. W. 1978. “The Nest as a Factor Determining Clutch-sizein Tropical Birds.” Journal of Ornithology 119: 227–230.
Snow, D. W., and H. Mayer-Gross. 1967. “Farmland as aNesting Habitat.” Bird Study 14: 43–52.
Sockman, K. W. 1997. “Variation in Life-history Traits andNest-site Selection Affects Risk of Nest Predation in theCalifornia Gnatcatcher.” The Auk 114: 324–332.
Stoate, C., and J. Szczur. 1994. “Nest Site Selection and Terri-tory Distribution of Yellowhammer Emberiza citrinella andWhitethroat Sylvia Communis in Field Margins.” In FieldMargins: Integrating Agriculture and Conservation,129–131. Monograph 58, British Crop Protection Council.
Stoate, C., S. J. Moreby, and J. Szczur. 1998. “Breeding Ecol-ogy of Farmland Yellowhammers Emberiza citrinella.” BirdStudy 45: 109–121.
Yomtov, Y. 1987. “The Reproductive Rates of AustralianPasserines.” Wildlife Research 14: 319–330.
Zoology and Ecology 207
Dow
nloa
ded
by [
Nor
thea
ster
n U
nive
rsity
] at
00:
46 0
5 O
ctob
er 2
014





























