Embed Size (px)
Citation preview

Cell Tissue Res (1982) 227:153-160 Cell and Tissue Research �9 Springer-Verlag 1982
Brain stem innervation of the caudal neurosecretory system*
John P. O'Brien and Richard M. Kriebel**
Department of Anatomy and Neurobiology, The University of Vermont, College of Medicine, Burlington, Vermont, USA
Summary. The innervation of the caudal neurosecretory system of Poecilia sphenops (black molly) was studied by use of the retrograde horseradish peroxidase (HRP) method. The structure of the caudal neurosecretory system in this species was well suited for application of HRP procedures. Acrylamide/HRP gel implants were placed in the nucleus of the caudal neurosecretory system. Two neuronal groups which contained HRP filled cells were found in the brain stem. Bilateral projections originate from the dorsal tegmentum of the midbrain and the reticular nucleus of the medulla.
Key words: Caudal neurosecretory sys tem- Horseradish peroxidase me thods - Innervation
The purpose of this study was to determine the origin of the innervation of the caudal neurosecretory system of teleosts. Morphological studies have shown that neurosecretory cells in the caudal system receive synaptic contacts (Fridberg 1963; Bern et al. 1965; Fridberg et al. 1966; Sano et al. 1966; Baumgarten et al. 1970; Kobayashi et al. 1979; Kriebel 1980). Electrophysiological studies indicated that the innervation of these cells originates from supraspinal levels of the neuraxis (Morita et al. 1961; Bennett and Fox 1962; Yagi and Bern 1965). Yagi and Bern (1965) suggested that since transection of the spinal cord altered the physiological behavior of the caudal neurosecretory cells the descending synaptic input significantly influences the activity of these cells. Transection of the spinal cord rostral to the caudal neurosecretory nucleus resulted in degeneration of some of the axon terminals within the caudal nucleus further indicating a descending synaptic input (Kriebel and O'Brien 1980). Several investigators have shown that the caudal
Send offprint requests to: Dr. Richard M. Kriebel, Department of Anatomy and Neurobiology, The University of Vermont, College of Medicine, Burlington, Vermont 05405, USA
* Supported by PHS 5429-19-4 and BNS 8206452 ** The authors wish to thank Drs. R. Parsons, S. Freedman and J. Wells for reading this report and A.
Angel for photographic assistance
0302-766X/82/0227/0153/$01.60

154 J.P. O'Brien and R.M. Kriebel
n e u r o s e c r e t o r y cells rece ive a m o n o a m i n e r g i c i nne rva t i on . H o w e v e r , these
synapses are o f u n k n o w n or ig in ( B a u m g a r t e n et al. 1970; S w a n s o n et al. 1975;
K o b a y a s h i et al. 1979, 1980; A u d e t a n d C h e v a l i e r 1981).
O n e a p p r o a c h to d e t e r m i n e the o r ig in o f a f fe ren t i n n e r v a t i o n is to uti l ize
r e t r o g r a d e h o r s e r a d i s h - p e r o x i d a s e ( H R P ) t racer p rocedu re s (LaVa i l a n d LaVa i l
1972). A p p l i c a t i o n o f this t e c h n i q u e in m a n y te leosts is d i f f icul t because the cauda l
n e u r o s e c r e t o r y cells a re in t e r spe r sed a m o n g o t h e r sp inal c o r d cons t i tuen t s .
H o w e v e r , in PoeciIia sphenops (b lack mol ly) , the n e u r o s e c r e t o r y cells a re seg rega ted
f r o m o t h e r sp inal c o r d n e u r o n s and, since there are no neura l s t ruc tu res c a u d a l to
the n e u r o s e c r e t o r y cells, there are no fibers o f passage c o u r s i n g t h r o u g h the nuc leus
(Kr iebe l 1980). T h e o r ig in o f the sup ra sp ina l p r e s y n a p t i c i n n e r v a t i o n has n o t been
inves t iga ted unt i l the p resen t s tudy.
Materials and methods
Poecilia sphenops (black molly) were obtained from Maywood Fish Farms, Ruskin, Florida and held in 10 gallon filtered and aerated freshwater aquaria. A total of 16 fish, average length 6 cm, were used. All fish were anesthetized in MS222 prior to surgery and killing.
Preparation of HRP implant. Microinjection and iontophoretic injection were unsuccessful in the caudal system preparation of the molly. However, the "slow release gel" method of Griffin et al. (1979) was suitable. With this method small localized implants of HRP could be made with minimal tissue damage and spread of label to undesirable areas. The control for HRP spread is discussed in the Results. The acrylamide/HRP gels in this study were prepared exactly as suggested by Griffin et al. (1979). The solution was drawn into a plastic pipet tip for polymerization and small wedges were cut from air-dried gels.
Placement of HRP implants. Anesthetized fish were placed on their side in a transparent petri dish. The fish were kept moist continually throughout surgery. The caudal peduncle of the fish tail was transilluminated with a fiber optic illuminator to visualize the vertebral column and to choose the exact spinal cord segment for the HRP implant. A small incision was made on the side of the fish, and skin and muscle retracted from the vertebral column. The spinal cord was approached from a dorsolateral direction between appropriate neural arches of the vertebrae. A single wedge of acrylamide/HRP gel was inserted into the dorsal aspect of the spinal cord of each fish between the ninth and third terminal vertebrae.
Processing of brain and spinal cord for HRP visualization. Forty-eight to 60 h following HRP implant the fish were anesthetized and the brain and spinal cord fixed in vivo in 2 % paraformaldehyde and 2.5 % glutaraldehyde in Millonig's buffer (pH 7.2; 0.06 M). The brain and spinal cord were dissected and flnmersed for no longer than 24 h in the same fixative. All tissue was frozen sectioned at 50 gm; brains were sectioned in either the transverse, horizontal, or sagittal planes; all spinal cords were sectioned horizontally. The sections were mounted on gelatinized slides and processed by the TMB procedure (Mesulam 1978). All sections were lightly counterstained with cresyl violet and examined and photographed with a Zeiss Universal photomicroscope. The location of HRP positive cells was plotted on camera lucida drawings of brain sections.
Results
Control for retrograde H R P transport. T h e p r inc ipa l c o n c e n t r a t i o n o f cauda l n e u r o s e c r e t o r y n e u r o n s in Poecilia sphenops is f o u n d wi th in the spinal c o r d at the
levels o f the f if th to s econd p r e t e r m i n a l ve r t eb ra l segments . In this species the
Innervation of caudal neurosecretory system 155
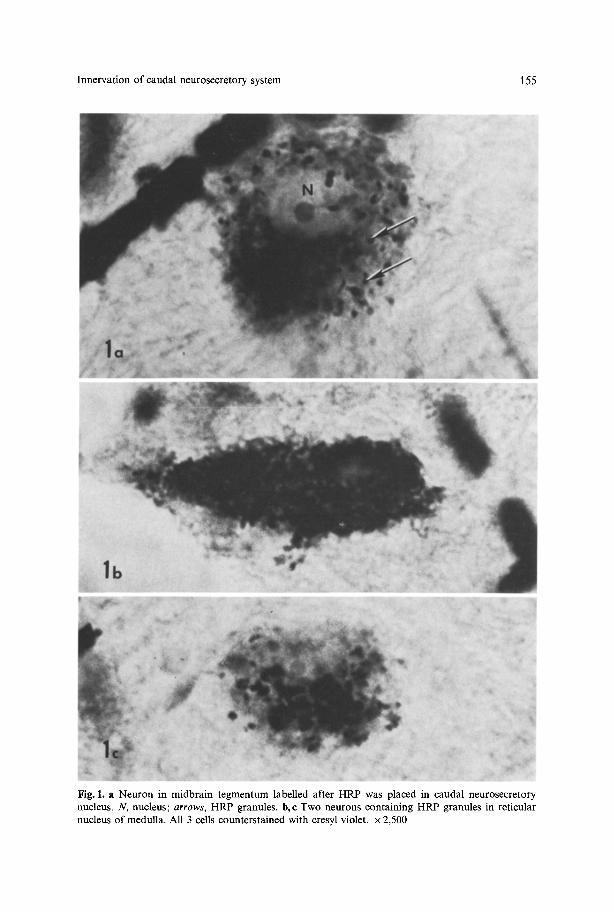
Fig. 1. a Neuron in midbrain tegrnentum labelled after HRP was placed in caudal neurosecretory nucleus. N, nucleus; arrows, HRP granules, b,e Two neurons containing HRP granules in reticular nucleus of medulla. All 3 cells counterstained with cresyl violet, x 2,500

156 J.P. O'Brien and R.M. Kriebel
. EQ Fig. 2A-D. Transverse sections through brain stem of P. sphenops showing midbrain and medulla projections. A Location of HRP-labelled neurons in midbrain (asterisks). B Transition between origin of midbrain and medullary projections; only labelled fibers in MLF. C, D Location of HRP-labelled cells (Q) in reticular nucleus of medulla (RNM). E Ventral funiculus and location of descending pathways to caudal neurosecretory system (arrow). OToptic tectum; TL torus longitudinalis; G nucleus glomerulosus; IL inferior lobe; MLF medial longitudinal fasciculus; TS torus semicircularis; VC valvulae cerebellae; CC corpus of cerebellum; VE vagal enlargement
M a u t h n e r axons te rmina te at app rox ima te ly the level o f the seventh pre te rmina l ver tebral segment (Kriebel , unpub l i shed observat ion) . The descending pro jec t ion axons o f the M a u t h n e r cells are well known in the teleost spinal cord and can be visual ized by H R P re t rograde t racer me thods (Kimmel and Powell 1980). In the present s tudy this system was uti l ized as an internal cont ro l for spread o f H R P into the neuropi l ros t ra l to the cauda l neurosecre tory system. Implan t s o f H R P rostra l to the t e rmina t ion o f M a u t h n e r axons filled these large cells within the medul la as well as o ther b ra in stem cell groups in a pa t t e rn s imilar to that r epor ted in zebraf ish (Kimmel et al. 1982). The M a u t h n e r system was examined in each bra in to de termine if imp lan ted H R P had been taken up by non-cauda l neurosecre tory fibers. We s tudied only those cases in which the M a u t h n e r system was unlabel led. In addi t ion , several fish that did no t receive exogenous HRP were examined to de termine if any cells were endogenous ly reactive to the peroxidase v isual iza t ion procedure . There were no endogenous ly HRP-pos i t ive neurons in any regions o f the fish brain.

Innervation of caudal neurosecretory system 157
Fig. 3. Horizontal section of spinal cord; rostral to left. Note rootlet leaving spinal cord, fibers containing HRP within ventral funiculus (arrows). x 1,050
Projections to the caudal neurosecretory system. Neurons which contained retrogradely transported HRP were observed at midbrain and medullary levels of the brain stem.
Midbrain projection. Figure 1 a shows a neuron containing HRP granules of the midbrain projection to the caudal neurosecretory system. In relation to other neurons in the molly brain these cells are large (25-30 gm diameter) and polygonal. The cells within the midbrain are bilaterally distributed around the midline (Fig. 2). They lie just beneath the subependymal layer, which is wide in this part of the brain stem, in the nucleus of the medial longitudinal fascicle. The medial longitudinal fascicle contains numerous HRP-labelled fibers presumably from these midbrain cells.
Medulla projection. A bilateral projection was also observed originating from the medullary region of the brain stem. The HRP-filled cells in the medulla are fusiform and smaller in comparison to the midbrain projection cells (Figs. 1 b, c). The cells are located in the ventrolateral aspect of the medullary tegmentum (Fig. 2). Labelled fibers in the spinal cord course in the ventral funiculus (Fig. 3) but do not form a discrete fiber tract. The ventral funiculus contains predominantly myelinated fibers; however, it is not possible to determine if the labelled fibers are myelinated in these preparations.

158 J.P. O'Brien and R.M. Kriebel
Discussion
The caudal neurosecretory system of Poecilia sphenops is a relatively discrete grouping of neurosecretory neurons within terminal spinal cord segments (Kriebel 1980). In this species we have taken advantage of the structure of this system to examine the origin of the descending synaptic input to the caudal system by using HRP retrograde tracer methods. Conclusions from this method depend on the control of spread of HRP and on possible transport of HRP within fibers of passage. The origin of the innervation has been difficult to study in most caudal neurosecretory systems of teleosts because in most fishes it is difficult to lesion or inject into a homogeneous population ofneurosecretory cells (Bern 1969; Kriebel et al. 1979).
The results from this study support the electrophysiological and morphological evidence of a descending synaptic input to the caudal neurosecretory system (Morita et al. 1961 ; Bennett and Fox 1962; Yagi and Bern 1965; Baumgarten et al. 1970; Kobayashi et al. 1979). Yagi and Bern (1965) showed in extracellular recording studies that electrical activity of caudal neurosecretory units was altered following transection of the spinal cord. These authors concluded that caudal neurosecretory ceils are "synaptically controlled by a center located probably in the brain" (Yagi and Bern 1965).
Two centers within the brain stem send direct projections to the caudal neurosecretory nucleus. The counterstained sections of the brains examined in this study clearly indicate cells of origin at the midbrain and medullary levels of the brain stem. The cells of the midbrain and medullary projections appear to be components of other brain stem nuclei. The midbrain cells are included in the nucleus of the medial longitudinal fascicle (Eaton and Farley 1973) and in the medulla the cells are located in the rostral part of the medullary reticular nucleus (Bartelmez 1915; Stefanelli and Camposano 1946; Smeets and Nieuwenhuys 1976; Nieuwenhuys 1977; Kimmel et al. 1982). In view of the known monoaminergic innervation of caudal neurosecretory cells (Baumgarten et al. 1970; Lederis et al. 1974; Swanson et al. 1975; Kobayashi et al. 1979, 1980; Audet and Chevalier 1981) it is interesting to note that the reticular nucleus of the medulla is intensely fluorescent in some species (Parent et al. 1978). We are presently studying the medullary reticular nucleus ofP. sphenops to determine if the HRP-labelled neurons are monoaminergic.
Observations have been made recently of direct projections of magnocellular neurons to spinal cord levels in other vertebrates (e.g., Nilaver et al. 1980; Swanson and Sawchenko 1980). In the present study no HRP-labelled cells were seen at the levels of the preoptic or lateral tuberal nuclei of the diencephalon. Therefore, we do not believe that the magnocellular neurosecretory nuclei project directly to the caudal neurosecretory system.
Current reports associate the caudal neurosecretory system with the mainte- nance of ionic balance in fishes (Fryer et al. 1978; Marshall and Bern 1979, 1981; Loretz and Bern 1980, 1981). Indeed, Yagi and Bern (1965) suggested that changes in ionic composition are the stimulus for the caudal neurosecretory cells but that the receptor for this response is within cranial regions. The descending systems identified in these studies may be the coordinating link between the caudal

Innervation of caudal neurosecretory system 159
neurosecretory system and cranial centers. The present study is the first step in understanding how the caudal neurosecretory system is integrated into the neuroendocrine circuitry of the teleost central nervous system.
References
Audet C, Chevalier G (1981) Monoaminergic innervation of the caudal neurosecretory system of the brook trout Salvelinusfontinalis in relation to osmotic stimulation. Gen Comp Endocrino145:189- 203
Bartelmez GW (1915) Mauthner's cell and the nucleus motorius tegmenti. J Comp Neurol 25:87-128 Baumgarten HG, Falck B, Wartenberg H (1970) Adrenergic neurons in the spinal cord of the pike (Esox
lucius) and their relation to the caudal neurosecretory system. Z Zellforsch 107:479-498 Bennett MVL, Fox S (1962) Electrophysiology of caudal neurosecretory cells in the skate and fluke. Gen
Comp Endocrinol 2:77-95 Bern HA (1969) Urophysis and caudal neurosecretory system. In: Hoar WS, Randall DJ (eds) Fish
physiology, Vol II. Academic Press, New York, p 399 Bern HA, Yagi K, Nishioka RS (1965) Structure and function of the caudal neurosecretory system of
fishes. Arch Anat Microsc Morphol Exp 54:217-238 Eaton RC, Farley RD (1973) Development of the Mauthner neurons in embryos and larvae of the
zebrafish, Brachydanio retio. Copeia 1973:673-682 Fridberg G (1963) Electron microscopy of the caudal neurosecretory system in Leuciscus rutilus and
Phoxinus phoxinus. Acta Zool (Stockh) 44:245-267 Fridberg G, Bern HA, Nishioka R (1966) The caudal neurosecretory system of the isospondylous teleost,
Albula vulpes, from different habitats. Gen Comp Endocrinol 6:195-212 Fryer JN, Woo NYS, Gunther RL, Bern HA (1978) Effect of urophysial homogenates on plasma ion
levels in Gillichthys mirabilis (Teleostei: Gobiidae). Gen Comp Endocrinol 35:238-244 Griffin G, Watkins LR, Mayer DJ (1979) HRP pellets and slow-release gels: two new techniques for
greater localization and sensitivity. Br Res 168:596-601 Kimmel CB, Powell SL (1980) Brain neurons that project to the spinal cord in young larvae of the
zebrafish. Soc Neurosci Abstr 6:628 Kimmel CB, Powell SL, Metcalfe WK (1982) Brain neurons which project to the spinal cord in young
larvae of the zebrafish. J Comp Neurol 205:112-127 Kobayashi Y, Ichikawa T, Kobayashi J (1979) Innervation of the caudal neurosecretory system of the
teleost. Gunma Sym Endocrinol 16:81-86 KobayashiY, Kobayashi H, Ohshiro S, OsumiY, Fujiwara M (1980) Monoaminergic innervation of the
caudal neurosecretory system of the carp, Cyprinus carpio. Zb Vet Med C Anat Histol Embryol 9: 65- 72
Kriebel RM (1980) The caudal neurosecretory system of Mollienesia sphenops: Light and electron microscopy. J Morphol 165:157-165
Kriebel RM, O'Brien JP (1980) Afferent projections to the caudal neurosecretory cells in P. sphenops. Soc Neurosci Abstr 6:456
Kriebel RM, Burke JD, Meetz GD (1979) Morphological features of the caudal neurosecretory system in the blueback herring, Pomolobus aestivales. Anat Rec 195:553-572
LaVail JH, LaVail MM (1972) Retrograde axonal transport in the central nervous system. Science 176:1416
Lederis K, Bern HA, Medakovic M, Chan DKO, Nishioka RS, Letter A, Swanson D, Gunther R, Resanovic M, Horne B (1974) Recent functional studies on the caudal neurosecretory system of teleost fishes. In: Knowles F, Vollrath L (eds) Neurosecretion: The final neuroendocrine pathway. Springer, New York, p 94
Loretz CA, Bern HA (1980) Ion transport by the urinary bladder of the gobiid teleost, Gillichthys mirabilis. Am J Physiol 239 (Regulatory Integrative Comp Physiol 8): R415-R423
Loretz CA, Bern HA (1981) Stimulation of sodium transport across the teleost urinary bladder by urotensin II. Gen Comp Endocrinol 43:325-331
Marshall WS, Bern HA (1979) Teleostean urophysis: urotensin II and ion transport across the isolated skin of a marine teleost. Science 204:519-521

160 J.P. O'Brien and R.M. Kriebel
Marshall WS, Bern HA (1981) Active chloride transport by the skin of a marine teleost is stimulated by urotensin I and inhibited by urotensin II. Gen Comp Endocrinol 43:484~491
Mesulam MM (1978) Tetramethyl benzidine for horseradish peroxidase neurochemistry: A non- carcinogenic blue reaction product with superior sensitivity for visualizing neural afferents and efferents. J Histochem Cytochem 26:106 117
Morita H, Ishibashi T, Yamashita S (1961) Synaptic transmission in neurosecretory cells. Nature (Lond) 191:183
Nieuwenhuys R (1977) The brain of the lamprey in a comparative perspective. Ann NY Acad Sci 299: 97 145
Nilaver G, Zimmerman EA, Wilkins J, Michaels J, Hoffman D, Silverman A (1980) Magnocellular hypothalamic projections to lower brain stem and spinal cord of the rat. Neuroendocrinol 30:150 ~ 158
Parent A, Dube L, Braford MR, Northcutt RG (1978) The organization of monoamine-containing neurons in the brain of the sunfish (L. gibbosus) as revealed by fluorescence microscopy. J Comp Neurol 182:495-516
Sano Y, Iida T, Taketomo S (1966) Weitere elektronenmikroskopische Untersuchungen am kaudalen neurosekretorischen System von Fischen. Z Zellforsch 75:328-338
Smeets WJAJ, Nieuwenhuys R (1976) Topological analysis of the brain stem of the lungfish Lepidosiren paradoxa. J Comp Neurol 187:589-612
Stefanelli A, Camposano A (1946) I Centri tegmentali dell'anguilla e la relazioni degli elementi giganti del tegmento dei ciclostomi, dei pesci e degli anfibi; recerche sul sistema mauthneriano. Pub Staz Zool Napoli 20:19-45
Swanson DD, Nishioka RS, Bern HA (1975) Aminergic innervation of the cranial and caudal neurosecretory systems in the teleost Gillichthys mirabilis. Acta Zool (Stockh) 56:225-237
Swanson LW, Sawchenko PE (1980) Paraventricular nucleus: A site for the integration of neuroendocrine and autonomic mechanisms. Neuroendocrinol 31:410~417
Yagi K, Bern HA (1965) Electrophysiologic analysis of the response of the caudal neurosecretory system of Tilapia mossambica to osmotic manipulations. Gen Comp Endocrinol 5:509-526
Accepted May 3, 1982



























![Muscle Innervation Chart II[1]](https://img.pdfslide.us/doc/110x75/55241db64a7959da488b45f0/muscle-innervation-chart-ii1.jpg)
