Embed Size (px)
Citation preview

INFECTION AND IMMUNITY, Aug. 1974, p. 356-360Copyright © 1974 American Society for Microbiology
Vol. 10, No. 2Printed in U.S.A.
Brain Polymorphonuclear Leukocyte Quantitation byPeroxidase Assay
A.. L. SMITH, I. ROSENBERG, D. R. AVERILL, E. R. MOXON, T. STOSSEL, AND D. H. SMITHDivisions of Infectious Disease and Hematology, Children's Hospital Medical Center, Boston, Massachusetts.
and The Laboratory of Pathology, Animal Research Center, Harvard Medical School, Boston,Massachusetts 02115
Received for publication 15 March 1974
Rat brain polymorphonuclear infiltration was quantitated by spectrophoto-metric assay of myeloperoxidase activity. Peroxidase was assayed by followingthe oxidation of o-dianisidine in the presence of a hydrogen peroxide generatingsystem (glucose-glucose oxidase). Enzyme activity was proportional to thenumber of polymorphonuclear leukocytes present at densities between 105 and106 cells. Brain tissue did not have detectable peroxidase activity and polymor-phonuclear leukocytes added to brain homogenates were quantitatively esti-mated. In infant rats with Haemophilus influenzae meningitis, the correlationbetween the number of polymorphonuclear leukocytes found in brain byperoxidase activity measurement and independent histological grading was notprecise, but there was an exact correlation in predicting presence or absence ofpolymorphonuclear leukocytes. Myeloperoxidase activity appears to afford arapid convenient tool for quantitative determination of tissue polymorphonu-clear leukocyte content.
The inflammatory response to an invadingorganism is one of the hallmarks of infection.From a mechanistic point of view, inflamma-tion can be defined as the host response tocellular injury involving changes in vascularpermeability and flow, and leukocytic infiltra-tion. These are elicited to a variable degree inresponse to a variety of stimuli. In the earlyphases of an acute inflammatory response, thepredominant cell type is the granulocyte.
Quantification of the numbers of granulo-cytes in lesions has been in general limited tothe study of anatomical areas where the cellscan be recovered with repeated washings, suchas the peritoneal cavity (12). The other ap-proach utilizes representative, or complete sec-tions of the area under study and microscopicenumeration of granulocytes. With the lattermethod, which is necessary for the study oftissue leukocytic infiltration, either calculationsare based on empirical assumptions, or tedious,time-consuming counting of many tissue sec-tions is necessary.Peroxidase catalyzes the reduction of H202
with the hydrogen donor substrate becomingoxidized. The activity of this enzyme variesmarkedly from tissue to tissue, being highest inspleen, the gastrointestinal tract, and the lungs,but with low, if any detectable activity in brain
(11). Myeloperoxidase is the name applied tothe peroxidase present in leukocytes of themyeloid series. These cells are particularly richin their content of this enzyme and it is esti-mated that 2 to 5% of the dry weight of thegranulocytes consists of myeloperoxidase (5,14). The enzyme is present in polymorphonu-clear leukocytes, to a lesser extent in monocytes(4), and a similar enzyme is present in eosino-philes (3). The presence of large amounts ofenzyme activity in these cells suggested thatperoxidase activity might serve as an indicatorof leukocytic infiltration, particularly in organswhich have low or absent peroxidase activity.This report describes the adaptation of a
peroxidase assay which will quantitate thenumber of polymorphonuclear leukocytesadded to, or present in, brain. We used ananimal model previously described (10) inwhich 80% of infant rats developed meningitisafter intranasal inoculation with Haemophilusinfluenzae E1.The quantitation of brain leukocyte content
is necessary when measuring metabolite con-centrations in brains of animals with pyogenicmeningitis. Without corrections for leukocytecontent, observed changes cannot be ascribed toresponse of the brain per se to the meningitisprocess.
356
on May 29, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

VOL. 10, 1974 BRAIN POLYMORPHONUCLEAR LEUKOCYTE QUANTITATION
MATERIALS AND METHODS
Chemicals. Inorganic salts were obtained in thepurest form available from Mallinckrodt ChemicalCo., St. Louis, Mo. Glucose and tris(hydroxyme-thyl)aminomethane (Tris) were obtained fromSchwarz-Mann Inc., Orangeburg, N.Y.; orthodianisi-dine was obtained from Sigma Chemical Corp., St.Louis, Mo., whereas Triton X-100 was purchased fromAnalabs Inc.; glucose oxidase (yeast, catalogue#346385) was obtained from Calbiochem with a spe-
cific activity of 160 Am/mg. Horseradish peroxidasewas obtained from Calbiochem.
Bacteriological methods. The bacteriologicalmethods were modified from those previously re-
ported from this laboratory (1). H. influenzae b(Eagan) was isolated from a child with meningitis.The strain was originally isolated on chocolate agar,
but had been passed repeatedly on brain heart infu-sion (BHI) supplemented with heme and oxidizednicotinaminde adenine dinucleotide (NAD+). A strep-tomycin-resistant mutant (E-1) was selected on sup-
plemented BHI agar containing streptomycin at 500gg/ml. H. influenzae E-1 was inoculated into brainheart broth containing a source of heme (supernatantfluid of horse blood diluted 1: 3,000 with distilledwater, and the supematant fluid sterilized by filtra-tion through a 0.22 jAm pore size membrane filter)(Millipore Corp.), 0.1 ml/100 ml, and NAD+ (SigmaChemical Corp., St. Louis, Mo.) at a final concentra-tion of 1 g/ml. They were grown at 37 C to a densityof 1010 (an absorbance of 0.5 in Lumetron model #401using the 490-nm filter), harvested by centrifugationat 5,000 x g at 4 C, resuspended in skim milk, andfrozen in portions at -70 C.Animals used. Rats used in this study were derived
from the outbred strain COBS/CD, originally ob-tained from Charles River Breeding Laboratories.These animals were bred in the Children's HospitalMedical Center animal quarter, maintained at 70+5 F at 50% relative humidity on a 7 a.m. to 7 p.m.light schedule in a pathogen-free environment.
Inoculation of animals. H. influenzae b, E-1, was
grown in supplemented BHI broth to a density of 108organisms/ml, harvested by centrifugation at 5,000 x
g at 4 C, resuspended in phosphate-buffered salinecontaining 0.1% gelatin (PBS-G) in the same media,and inoculated intranasally in a volume of 0.01 mlbetween 11 a.m. and 1 p.m.
Histological methods. The animals were decapi-tated 48 h after inoculation. Tissue samples were
collected from each rat, fixed in 10% buffered neutralFormalin, embedded in paraffin, cut 1-MLm thick, andstained with hematoxylin and eosin, Giemsa, Good-pasture, and Brown and Brenn stains (for bacteria).Five coronal sections of a decalcified hemiskull were
examined with a microscope. The tissues were exam-
ined by one of us (D.R.A.) without knowledge of theinoculum. We defined meningitis as the presence ofacute inflammatory cells within the meninges or inthe subarachnoid space.
Preparation of leukocytes and their extracts.White blood cells were harvested from the peritonealcavity of rats 5 to 21 days old, 16 h after intraperito-
neal inoculation with 10% sodium caseinate (Difco) (1ml/10 g of body weight). Cells were aspirated underdirect vision, placed in normal saline, and centrifugedat 400 x g for 10 min at 4 C. The supernatant was
discarded, and the pellet was resuspended in an
equivalent volume of PBS (usually 0.5 ml per ani-mal). The number of white blood cells per milliliterwas determined by counting a portion on a Coultercounter. PBS was then added to the cell suspension tomake the final concentration 106 to 107 cells/ml in a
total volume of 4 ml. The resulting suspension was
sonically treated at 60 W for 2 min at -4 C. Thesupernatant was saved, refrigerated, and designatedas the white blood cell extract. This material served as
a source of peroxidase activity. We did not find any
significant variation in peroxidase activity of poly-morphonuclear leukocytes prepared from rats of vari-ous ages (e.g., 5 to 150 days of age), but this possibilityexists with other species.When tissues were assayed for peroxidase activity,
with or without the addition of leukocytes, they were
processed as follows. The organs were removed, blot-ted to remove excess blood, weighed, and immediatelyplaced in 10 ml of PBS at 4 C. They were homoge-nized in 5 ml of PBS per g (wet weight) with a
standard-clearance Potter-Elvehjem homogenizer.Then they were sonically treated as noted above,before further processing for the assay of myeloperoxi-dase. All further operations were performed in a coldroom maintained between 0 and 4 C.
Myeloperoxidase assay: tissue preparation. Thesonically treated organs or leukocyte preparationswere centrifuged at 400 x g for 10 min, the pellet was
discarded, and the supernatant was centrifuged at39,000 x g for 20 min. The resulting pellet was thenhomogenized by hand in one-fourth of the originalvolume with 1.5 M NaCl until the solution was homo-geneous, and centrifuged at 39,000 x g for 15 min, andthe supematant was used as a source of peroxidaseactivity.
Peroxidase assay. The peroxidase activity of thetissue extracts was assayed by a modification of thespectrophotometric method of Lundquist and Josef-sson (9). This method uses glucose and glucoseoxidase to generate a continuous supply of H202, as
high concentrations of this substrate can inhibitperoxidase (2). Peroxidase then reduces the H202 towater with the concomitant oxidation of the hydrogendonor, o-dianisidine. The rate of oxidation of o-
dianisidine was followed by measuring the absorbanceat 440 nm in a Beckman Acta III double-beamspectrophotometer. We prepared 100 ml of glucosesolution by adding 3 g of glucose to a solution which is0.1 M NH4Cl and 0.1 M CaCl2. The 1% (wt/vol)o-dianisidine and 20% (wt/vol) Triton X-100 were
prepared in absolute ethanol. A standard peroxidasereagent was prepared by adding 1 ml of 1% o-dianisi-dine, 1 ml of 20% Triton, and 0.2 mg of glucoseoxidase to 100 ml of Tris-hydrochloride, 0.5 M (pH7.0) (20 C). The assay was run at 37 C with 0.2 ml ofglucose solution, 2.6 ml of peroxidase reagent, and 0.2ml of tissue extract. Routinely, the reference consistedof 1.5 M NaCl instead of a source of peroxidase, butneither substrate-free (glucose-glucose oxidase) nor
357
on May 29, 2018 by guest
http://iai.asm.org/
Dow
nloaded from
358 SMITH ET AL.
peroxidase-free samples showed detectable increasein the absorbance at 440 nm over the assay period(10 to 20 min).
RESULTSRelationship between polymorphonuclear
leukocyte number and peroxidase activity.Preliminary experiments with the standardassay system determined that the peroxidaseactivity was linear with nanogram quantities ofhorseradish peroxidase using a 2- to 20-minincubation period. In addition, peroxidase ac-tivity was demonstrated in leukocyte prepara-tions and brains from infected infant rats.However, there was poor agreement between theamount of tissue assayed and observed initialvelocity. Furthermore, only a variable percent-age of horseradish peroxidase activity could berecovered after addition to leukocytes, homog-enization, and assay. The introduction of the 1.5M NaCl extraction of enzyme activity fromcrude homogenate pellets permitted quantita-tive recovery of added horseradish peroxidase.Thus, all subsequent assays used the extractionprocedure as described in Materials andMethods.
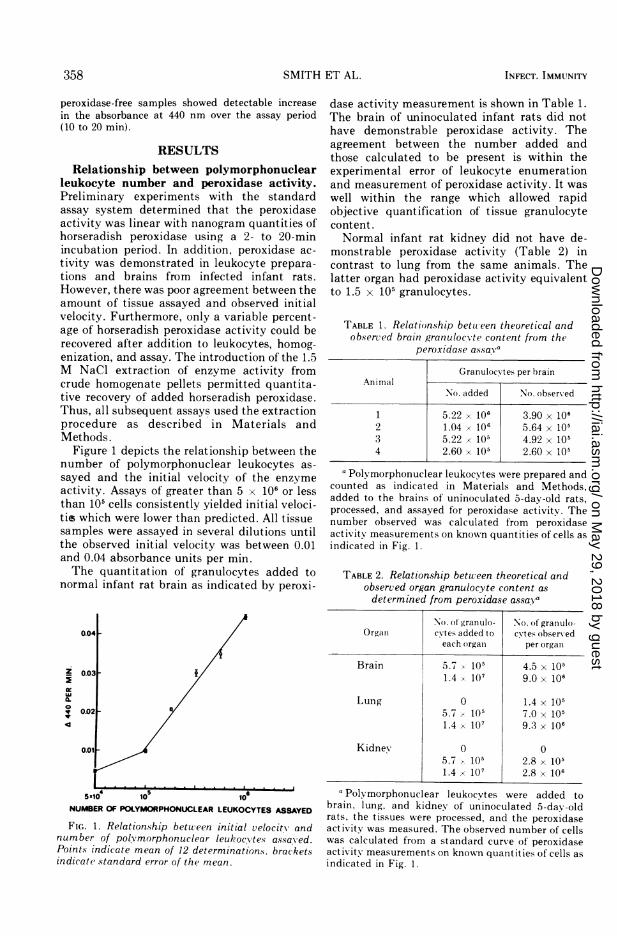
Figure 1 depicts the relationship between thenumber of polymorphonuclear leukocytes as-sayed and the initial velocity of the enzymeactivity. Assays of greater than 5 x 106 or lessthan 105 cells consistently yielded initial veloci-tis which were lower than predicted. All tissuesamples were assayed in several dilutions untilthe observed initial velocity was between 0.01and 0.04 absorbance units per min.The quantitation of granulocytes added to
normal infant rat brain as indicated by peroxi-
0.04-
X 0.03
0.02[
NUMBER OF POLYMORPHONUCLEAR LEUKOCYTES ASSAYED
FIG. 1. Relationship between initial velocity andnumber of poly morphonuclear leukoc'vtes assayved.Points indicate mean of 12 determinations. bracketsindicate standard error of the mean.
INFECT. IMMUNITY
dase activity measurement is shown in Table 1.The brain of uninoculated infant rats did nothave demonstrable peroxidase activity. Theagreement between the number added andthose calculated to be present is within theexperimental error of leukocyte enumerationand measurement of peroxidase activity. It waswell within the range which allowed rapidobjective quantification of tissue granulocytecontent.Normal infant rat kidney did not have de-
monstrable peroxidase activity (Table 2) incontrast to lung from the same animals. Thelatter organ had peroxidase activity equivalentto 1.5 x 105 granulocytes.
TABLE 1. Relationship between theoretical andobserved brain granulocvte content from the
peroxidase assava
Granulocvtes per brainAnimal
No. added No. obserxved
1 5.22 x 106 3.90 x 1062 1.04 x 106 5.64 x 1053 5.22 x 105 4.92 x 1054 2.60 x 105 2.60 x 105
aPolymorphonuclear leukocytes were prepared andcounted as indicated in Materials and Methods,added to the brains of uninoculated 5-day-old rats,processed, and assayed for peroxidase activity. Thenumber observed was calculated from peroxidaseactivity measurements on known quantities of cells asindicated in Fig. 1.
TABLE 2. Relationship between theoretical andobserved organ granulocyte content asdetermined from peroxidase assaya
No. of granulo- No. ot granulo-Organ cNtes added to cvtes observed
each orgall per orgala
Brain 5.7 105 4.5 x 1051.4 10( 9.0 X 106
Lung 0 1.4 x 1055.7 / 105 7.0 x 1051.4 x 107 9.3 x 106
Kidney 0 05.7 105 2.8 x 1051.4 x 107 2.8 x 106
a Polvmorphonuclear leukocytes were added tobrain, lung. and kidney of uninoculated 5-day-oldrats, the tissues were processed, and the peroxidaseactivitv was measured. The observed number of cellswas calculated from a standard curve of peroxidaseactivity measurements on known quantities of cells asindicated in Fig. 1.
on May 29, 2018 by guest
http://iai.asm.org/
Dow
nloaded from
VOL. 10, 1974 BRAIN POLYMORPHONUCLEAR LEUKOCYTE QUANTITATION
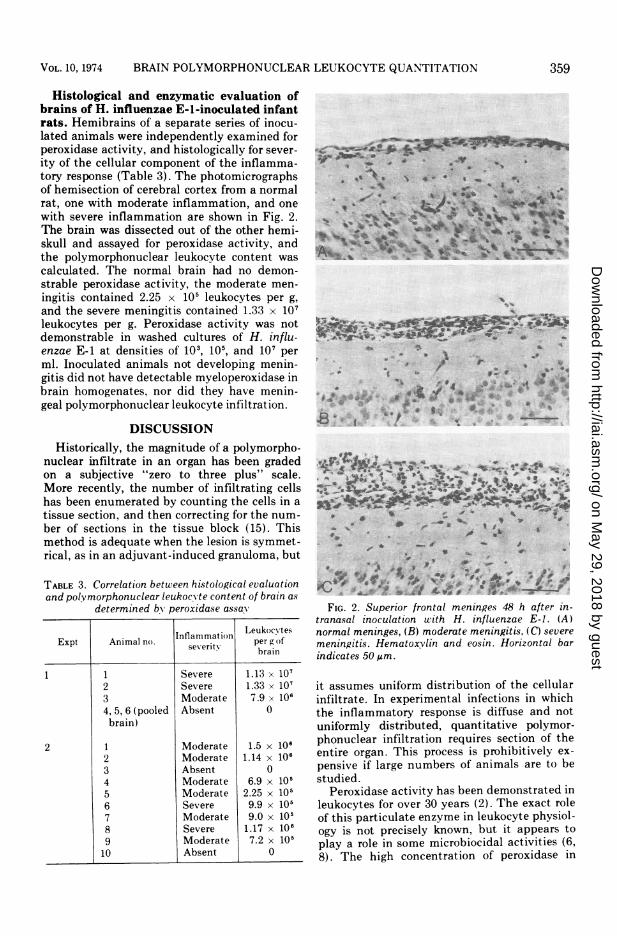
Histological and enzymatic evaluation ofbrains of H. influenzae E-l-inoculated infantrats. Hemibrains of a separate series of inocu-lated animals were independently examined forperoxidase activity, and histologically for sever-ity of the cellular component of the inflamma-tory response (Table 3). The photomicrographsof hemisection of cerebral cortex from a normalrat, one with moderate inflammation, and onewith severe inflammation are shown in Fig. 2.The brain was dissected out of the other hemi-skull and assayed for peroxidase activity, andthe polymorphonuclear leukocyte content wascalculated. The normal brain had no demon-strable peroxidase activity, the moderate men-ingitis contained 2.25 x 105 leukocytes per g,and the severe meningitis contained 1.33 x 107leukocytes per g. Peroxidase activity was notdemonstrable in washed cultures of H. influ-enzae E-1 at densities of 103, 105, and 107 perml. Inoculated animals not developing menin-gitis did not have detectable myeloperoxidase inbrain homogenates, nor did they have menin-geal polymorphonuclear leukocyte infiltration.
DISCUSSIONHistorically, the magnitude of a polymorpho-
nuclear infiltrate in an organ has been gradedon a subjective "zero to three plus" scale.More recently, the number of infiltrating cellshas been enumerated by counting the cells in atissue section, and then correcting for the num-ber of sections in the tissue block (15). Thismethod is adequate when the lesion is symmet-rical, as in an adjuvant-induced granuloma, but
TABLE 3. Correlation between histological evaluationand polvmorphonuclear leukocx te content of brain as
determined by peroxidase assaY
LeukocN tesIntlammat ion rExpt Animal no. severit pergao
brain
1 1 Severe 1.13 x 1072 Severe 1.33 X 1073 Moderate 7.9 x 1064, 5, 6 (pooled Absent 0brain)
2 1 Moderate 1.5 x 1062 Moderate 1.14 x 1063 Absent 04 Moderate 6.9 x 10'5 Moderate 2.25 x 10'6 Severe 9.9 x 10'7 Moderate 9.0 x 10'8 Severe 1.17 x 1069 Moderate 7.2 x 10'10 Absent 0
S '*\ 4, # 9 v.
-o. '1I,
' '~~9 _
, * r~~~~~iVIAV
t E, s
.Q. ...
9:I
*4d; 9- 9is* *i--:; *t A 'p e - o ::
**9} *4,p.
* ~* ,,, 9
4. OF,. Ij
-9N
FIG. 2. Superior frontal meninges 48 h after in-tranasal inoculation with H. influenzae E-I. (A)normal meninges, (B) moderate meningitis, (C) severemeningitis. Hematoxvlin and eosin. Horizontal barindicates 50 Am.
it assumes uniform distribution of the cellularinfiltrate. In experimental infections in whichthe inflammatory response is diffuse and notuniformly distributed, quantitative polymor-phonuclear infiltration requires section of theentire organ. This process is prohibitively ex-pensive if large numbers of animals are to bestudied.
Peroxidase activity has been demonstrated inleukocytes for over 30 years (2). The exact roleof this particulate enzyme in leukocyte physiol-ogy is not precisely known, but it appears toplay a role in some microbiocidal activities (6,8). The high concentration of peroxidase in
359
on May 29, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

360 SMITH
these cells suggested the possibility that en-zyme activity might be used as an index ofleukocyte number. This would be possible ifassay conditions were used in which peroxidaseactivity was proportional to the amount ofenzyme assayed. This was readily achievablewith crystalline horseradish peroxidase, but, asmentioned, linearity with known quantities ofleukocytes was not obtained until the extractionstep was used. We found, as did Himmelhoch etal. (7), that the total amount of peroxidaseactivity found in the solubilized extract ofleukocytes was several times greater than theamount assayable in the whole homogenate orsonic extract.Our failure to detect peroxidase activity in rat
brain differs from data reported by Neufeld etal. (11). These investigators found that adult ratbrain had barely detectable peroxidase activ-ity, and was the lowest of any tissue surveyed.This difference might be due to the age at whichthe rats were studied, but it is of note thatthe tissues found to have the highest peroxidaseactivity (e.g., uterus, gastrointestinal tract, andspleen) are the same tissues which physiologi-cally contain large numbers of leukocytes. Thisraises the possibility that the peroxidase activ-ity in these tissues is due to the containedleukocytes. This explanation is also compatiblewith our observation of demonstrable peroxi-dase activity in infant rat lung, but none inbrain or kidney. Histological examination ofthese organs between one and two weeks of ageconsistently demonstrates a leukocyte infiltratein lung (primarily mononuclear) but not inbrain or kidney. This hypothesis could be testedby measurements of peroxidase activity in tis-sues of gnotobiotic rats. A similar hypothesishas been proposed relating organ eosinophilecontent and peroxidase activity (13).
It should be emphasized that quantitatingbrain leukocytic infiltration by measurementsof peroxidase activity uses polymorphonuclearleukocytes as the reference standard. Peroxi-dase activity is then equated with the number ofpolymorphonuclear leukocytes needed to yieldgiven peroxidase activity. If the brain is infil-trated by leukocytes other than polymorphonu-clear leukocytes, which may contain more orless peroxidase per cell, then the peroxidaseactivity measurements would yield falsely highor low leukocyte numbers. Eosinophiles, whichhave more peroxidase activity per cell thanpolymorphonuclear leukocytes, would greatlyincrease the peroxidase activity observed inbrain and lead to an overestimation of thenumber of leukocytes present. Conversely, infil-tration by monocytes, which have low peroxi-dase activity, would lead to an underestimation
INFECT. IMMUNITY
of the numbers of leukocytes present. Thesespecial circumstances, which occur rarely dur-ing the acute inflammatory response, would beapparent on histological evaluation of the proc-ess under study. In situations in which thepredominant cell type is the eosinophile ormonocyte, standard curves prepared using thesecells would allow quantitation of leukocyticinfiltration.
The quantitation of leukocytic infiltrationinto brain by measurement of peroxidase activ-ity appears to be rapid, convenient, and eco-nomical. The construction of a standard curveof polymorphonuclear leukocytes and peroxi-dase activity should be explained for eachanimal species.
ACKNOWLEDGMENTS
This work was supported in part by Public Health Servicegrants NS 09048, HD 06276 (A.L.S.) from the National Insti-tute of Child Health and Human Development, RR 00401(D.R.A.), and Al 08362 (I.R. and D.H.S.) from the NationalInstitute of Allergy and Infectious Diseases.
LITERATURE CITED
1. Anderson, P., R. B. Johnston, and D. H. Smith. 1972.Human serum activities against Haemophilusinfluenzae type b. J. Clin. Invest. 51:31-38.
2. Agner, K. 1943. Verdoperoxidase. Advan. Enzymol.3:137-148.
3. Archer, G. T. 1962. Release of peroxidase from eosino-phile granules in vitro. Nature (London) 194:973-974.
4. Archer, G. T., and J. G. Hirsch. 1963. Isolation ofgranules from eosinophile leukocytes and study of theirenzyme content. J. Exp. Med. 118:277-286.
5. Chance, B. and B. Hagihara. 1960. Initiation of succinateoxidation in aged pidgeon heart mitochondria. Bio-chem. Biophys. Res. Commun. 3:1-5.
6. Hamon, C. B., and S. J. Klebanoff. 1973. A peroxidasemediated Streptococcus mitus dependent antimi-crobial system in saliva. J. Exp. Med. 137:438-450.
7. Himmelhoch, S. R., W. H. Evans, M. G. Mage, and E. A.Peterson. 1969. Purification of myeloperoxidase fromthe bone marrow of the guinea pig. Biochemistrv8:914-921.
8. Klebanoff, S. J. 1971. Intraleukocytic microbicidal de-fects. Annu. Rev. Med. 22:39-62.
9. Lundquist, I., and J. 0. Josefsson. 1971. Sensitivemethod for determination of peroxidase activity intissue by means of coupled oxidation reaction. Anal.Biochem. 41:567-577.
10. Moxon, E. R., A. L. Smith, D. R. Averill, and D. H.Smith. 1974. Haemophilus influenzae meningitis ininfant rats after intranasal inoculation. J. Infect. Dis.129:154-162.
11. Neufeld, H. A., A. N. Levay, F. V. Lucas, A. P. Martini,and E. Stotz. 1958. Peroxidase and cytochrome oxidasein rat tissues. J. Biol. Chem. 233:209-211.
12. Paz, R. A., and W. G. Spector. 1972. The mononuclear-cell response to injury. J. Pathol. Bacteriol. 84:85-103.
13. Rytomaa, T., and H. Teir. 1961. Relationship betweentissue eosinophiles and peroxidase activity. Nature(London) 192:271-272.
14. Schultz, J., and K. Kaminker. 1962. Myeloperoxidase ofthe leukocyte of normal human blood: content andlocalization. Arch. Biochem. Biophys. 96:465-467.
15. Spector, W. G., A. W. J. Lykke, and D. A. Willoughby.1967. A quantitative study of leukocyte emigration inchronic inflammatory granulomata. J. Pathol. Bacte-riol. 93:101-107.
[ET AL.
on May 29, 2018 by guest
http://iai.asm.org/
Dow
nloaded from





























