Embed Size (px)
Citation preview

THE JOURNAL OF RIOLOGICAL CHEMISTRY Vol. 243, No. 8, Issue of April 25, pp. 1718-1724, 1963
Printed in U.S.A.
Bovine Pepsinogen and Pepsin
I. ISOLATION, PURIFICATION, AND SOME PROPERTIES OF THE PEPSINOGEN*
(Received for publication, December 6, 19G7)
ROSIE B. CHOW AA-D BEATRICE KASSELL
From the Department of Biochemistry, Marquette School of Medicine, Inc., Milwaukee, Wisconsin 53233
SUMMARY
As the first step in the investigation of the structure and action of little known gastric zymogens and enzymes, pep- sinogen has been isolated from the mucosa of the fourth stomach (abomasum) of the’ cow. The pepsinogen was purified by ammonium sulfate fractionation, batch absorption on diethylaminoethyl (DEAE) cellulose, recycling gel filtra- tion on Sephadex G-100, and finally chromatography on DEAE-cellulose. The preparation appears to be homoge- neous by chromatography, amino acid analysis, ultracen- trifugation, and amino-terminal analysis, and nearly homo- geneous by disc electrophoresis.
Amino acid analysis by ion exchange chromatography indicates the presence of 362 f 2 amino acid residues, which account quantitatively for the total nitrogen of the prepa- ration. Compared to swine pepsinogen (RAJAGOPALAN, T. G., MOORE, S., AND STEIN, W. H., J. BioZ. Chem., 241, 4940 (1966)), which has 363 amino acids, the bovine pep- sinogen shows considerable similarity in composition, es- pecially with respect to the large number of acidic and small number of basic residues. However, there are at least 22 amino acid substitutions. In both proteins there are five amino acids that occur in equal numbers of residues per molecule; these are 4 methionine, 6 half-cystine, 6 tryp- tophan, 15 phenylalanine, and 35 glycine. In addition, the sum of the residues of certain similar amino acids is constant for bovine and swine pepsinogens; this is true for the pairs lysine and arginine (14 residues), aspartic and glutamic acids (72 residues), and leucine and isoleucine (57 to 58 residues). Significant differences occur in some other amino acids, notably histidine (2 residues in bovine pepsinogen, 3 in swine) and proline (15 and 19, respectively).
* This investigation was supported by Public Health Service Research Grant. AM-09826 from the National Institute of Arthritis and Metabolic Diseases and by Public Health Service General Research Support Grant 5-Sol-FR-5434. An abstract of part of this work has been published (Abstracts Seventh International Congress of Biochemistry, 1967, IUB Vol. IV, p. 782, F-135). This work has been taken from a thesis to be submitted by Rosie B. Chow to Marquette University Graduate School in partial ful- fillment of the requirements for the degree of Doctor of Philos- ophy.
The molecular weight of the bovine pepsinogen is 38,900 by amino acid analysis and 37,500 by ultracentrifugation.
Bovine pepsin was isolated from gastric juice and crystallized by Northrop in 1933 (1). At that time, solubility studies showed that it was a different protein from swine pepsin (2)) although both enzymes had equal specific activities on several protein substrates. Other than the studies of Fruton and Bergmann on specificity (3) and on the pH dependence of the activity (4), which were both strikingly similar to swine pepsin, the bovine enzyme has received little attention. To our knowledge, bovine pepsinogen has never been isolated.
In contrast, a great deal of work has been done on the structure and properties of swine pepsinogen and pepsin. Sequence studies are approaching completion in several laboratories (e.g. Ref- erences 512).
A second well characterized zymogen and enzyme of this important group would aid in identifying the portions of the molecule essential for activity and for maintenance of conforma- tion. It would supplement chemical modification studies in elucidating the requirements for zymogen activation and for the action of the enzyme. We have therefore undertaken to study the primary structure of bovine pepsinogen and pepsin. The present paper describes the isolation, purification, and amino acid composition of the pepsinogen. It has proved to be sufficiently different from the swine zymogen to be of considerable interest.
EXPERIMENTAL PROCEDURE
Materials-The fundic portion of the fourth stomach of the cow (abomasum) was obtained partly through Wilson Labora- tories, Chicago, andpartly from theLiebmannPackingCompany, Green Bay, Wisconsin.’ Swine pepsin (twice crystallized, Lot PM 6HE) and swine pepsinogen (crystalline, Lot PG1117) were obtained from Worthington. Bovine hemoglobin (twice crystallized) was purchased from Pentex. Sephadex G-25 and
1 We wish to thank Mr. Leonard C. Liebmann for supplying part of the abomasum used in this investigation.
1718
by guest on April 11, 2020
http://ww
w.jbc.org/
Dow
nloaded from

Issue of April 25, 1968 R. B. Chow and B. Kassell 1719
G-100 (both bead form) were Pharmacia products. DEAE- cellulose was No. 71, type 20, from Schleicher and Schuell. Deionized distilled water was used throughout. Chemicals were reagent grade.
Pepsin Assay-The potential pepsin content of the pep- sinogen was routinely determined by a modification* of the hemoglobin method of Anson (13). The substrate solution was prepared by dissolving 1 g of hemoglobin in 32 ml of water, then adding 8 ml of 0.3 M HCI; the solution was filtered through a coarse, sintered glass funnel and kept in the refrigerator for up to 1 week. The amount of 0.3 M HCI required to bring 0.1 ml of each unknown solution to pH 2.0 was predetermined. Poly- carbonate centrifuge tubes (100 x 16 mm) were placed in a 37.6” water bath. The required amount of 0.3 M HCl plus a solution of 0.01 M HCl in 0.1 M NaCl to bring the volume to 0.9 ml were placed in the tubes; 0.1 ml of the solutions to be assayed was added to each. With timing, 1 ml of previously equilibrated hemoglobin solution was added, the solution was mixed, and after exactly 5 min the reaction was terminated by adding 2 ml of 5% trichloracetic acid. The mixture was cooled in ice for 10 min and centrifuged in a Sorvall SS-4 centrifuge at 28,000 x g for 30 min. Cellulose nitrate tubes, 17 x 95 mm, were used as adapters for the polycarbonate tubes in the SM.24 rotor. The absorbance of the clear supernatant solution was measured at 280 rnp, usually against a blank which contained all the reagents but the solution to be assayed. With the crude, dark solutions of the early purification steps, blanks were prepared (12) by using the same amount of assay solution, and reversing the order of addition of trichloracetic acid and hemoglobin solutions.
Standard solutions of swine pepsin, containing 3 to 10 pg/O.l ml, were prepared in 0.1 M sodium acetate buffer of pH 5.3. A standard curve was prepared by assaying these as described above. The potential specific activity of the bovine zymogen was expressed as microgram equivalents of swine pepsin per AZW of the bovine pepsinogen solution.
Milk-clotting Test-Milk-clotting activity was used to estimate the amount of active pepsin (if any) present in the pepsinogen solution. The method was a modification of the procedure of Seijffers, Segal, and Miller (14). The stock milk solution was prepared by dissolving 16.8 g of nonfat dry milk (Carnation) in 100 ml of deionized water. The diluted milk solution contained 0.1 M sodium acetate buffer of pH 5.3, milk stock solution, and 0.1 M CaClz solution in the proportions 5:5: 1. The solution for testing consisted of 2 ml of the acetate buffer and 0.5 ml of the diluted milk solution in a glass test tube (15 X 125 mm) in a water bath at 37.6”. The solution was mixed by means of an air-driven magnetic stirrer immersed in the water bath. The clot was detected more readily by observing the solution as it moved in a siphon (15), made by inserting a small piece of tubing (0.5- mm inside diameter) into one end of a 35-mm length of trans- parent Tygon tubing of 1.5-mm inside diameter. This siphon tube was filled with buffer and clamped. After 0.1 ml of the enzyme solution, containing about 5 pg of pepsin, had been added to the milk-buffer mixture, the siphon was inserted and the clamp was released. The time required to show the first flock in the tubing was taken as the milk-clotting time, and was about 30 set for 5 pg of pepsin. For unknown solutions, up to 0.5 ml could be used, and the volume of acetate buffer was adjusted
2 We are grateful to Dr. Robert J. Peanasky for a part of the modification described.
accordingly. A solution was considered to be free of active pepsin if it did not clot in 2 min when it contained at least 25 pg of pepsinogen.
Analytical Methods-Amino acid analysis was carried out essentially as described by iMoore and Stein (16). Duplicate samples of about 15 mg were dissolved and diluted to 10 ml with water. From these solutions, l-ml samples were taken for hydrolysis in 5.7 N HCl for 16, 24, 48, and 72 hours at 110” in boiling toluene, and 1.5-ml samples were removed for total nitrogen determination by the Kjeldahl method (17). A blank with HCl alone was used to correct for ammonia in the reagents. A Beckman/Spinco model 120B amino acid analyzer was used for ion exchange chromatography (18, 19). The absorbance of the protein at 280 rnp was determined on appropriate dilutions of the same solutions.
A moisture determination (100” in vacuum) was made on a sample taken at the same time. Inorganic phosphate was determined (20) after removal of the protein from solution by precipitation with trichloracetic acid. The content of phosphate in the sample, which had been dialyzed at pH 7.0, was calculated as the sum of NaH2P04 and NazHPOI, determined from the pK, value of 7.28 at 5”. The weights of the samples were then corrected for moisture and salts.
Cystine was determined as cysteic acid (21), and tryptophan was measured spectrophotometrically (22) on separate samples. Methionine sulfoxide was determined after alkaline hydrolysis (23). The Sanger method as described by Fraenkel-Conrat, Harris, and Levy (24) was used for amino-terminal analysis.
Disc electrophoresis on 3 to 5 pg of the purified protein was carried out with a Canalco instrument, model 66. Standard procedures were followed except for buffer modifications (25). A diethylbarbituric acid Tris buffer of pH 7.0 was used during electrophoresis.
Conductivity of chromatographic effluents was measured on a Yellow Springs Instrument Company conductivity bridge, model 31.
RITSULTS
PuriJicatim of Bovine Pepsinogen
Bovine abomasa were removed immediately after slaughtering and were washed thoroughly to prevent activation of the zymogen by the highly acidic gastric juice (26) The fundic portion was cut out; the mucosa was separated, trimmed and frozen for storage. All subsequent operations were carried out at about 5”.
Step 1: E&rc&i&-The hashed mucosa (9.1 kg) was ex- tracted with 18 liters of 0.1 M sodium phosphate buffer of pH 7.3 for 15 min. Solca Floe BW-20, 4.5 kg, was added, and the mixture was filtered on three al-cm-diameter Buchner funnels. The filtered cakes were re-extracted with two 18-liter portions of the pH 7.3 buffer.
Step z?: Ammonium Sulfate Fractionation-The extracts were combined. Solid ammonium sulfate was added to 23% satura- tion, with NaOH introduced as needed to keep the pH close to 7.3. Again 4.5 kg of BW-20 were added, and the mixture was filtered. The cakes were washed with 10 to 15 liters of buffer solution, 23% saturated with ammonium sulfate. The filtrate
3 We are grateful to Wilson Laboratories, especially to Dr. Conrad de Fiebre and Mr. Edward P. Smith, for carrying out the extraction and ammonium sulfate fractionation, and also for supplying part of the abomasum.
by guest on April 11, 2020
http://ww
w.jbc.org/
Dow
nloaded from

1720 Bovine Pepsinogen and Pepsin,. I Vol. 243, No. 8
TABLE I Purification steps and yield of bovine pepsinogen
step
Solution of ammonium sulfate cake.
3. First batch absorption on DEAE-cellulose.
5. Second batch absorp- tion on DEAE-cellu- lose
6. Sephadex G-100, re- cycling.
7. Chromatography on DEAE-cellulose
‘otal pepsinogen a each step
At start
- 1
w
1433
910
114
95
68
18
63 1
38 228 2
83 430 4
72 514 5B
pecific poten. Elution dia- tial activity* gram in Fig.
a Recovery at each step is based on the center portion of the peak actually pooled. The yield could be improved by combining and rechromatographing the edges of the peaks from several runs. At Step 5, the yield based on the three potentially active peaks was 59%.
b Contamination with active pepsin, which amounted to 5 to 15% of the total potential activity of the ammonium sulfate cakes, decreased to zero by Step 6.
and washings were combined. Solid ammonium sulfate was added to 63% saturation. Perlite 4106 (500 g) was added, and the mixture was allowed to settle overnight. The supernatant was siphoned off and discarded. The precipitate was collected on three Buchner funnels and was shipped frozen to the labo- ratory.
Step 3: First Batch Absorption on DEAE-cellulose-The ammonium sulfate cake was divided into 600-g batches. Each batch was suspended in 500 ml of 0.01 M Tris-phosphate buffer of pH 7.0 and dialyzed against two changes of the same buffer. (The molarity of the Tris-phosphate buffers was determined by the concentration of Tris; all were adjusted to pH 7.0 f 0.05.) Centrifugation of the suspension for 3 hours at 13,000 X g in a Sorvall refrigerated centrifuge sedimented the Perlite, which was resuspended in an equivalent volume of buffer and centrifuged again. The combined supernatant solutions were then centri- fuged at 36,000 X g in a Spinco L-2 ultracentrifuge for 3 to 4 hours. The sediment was washed as before. An almost clear solution was obtained, which was filtered through Schleicher and Schuell No. 588 folded paper.
The DEAE-cellulose was precycled with 0.5 M NaOH and 0.5 M HCl (27) and equilibrated with 0.05 M Tris-phosphate buffer. After removal of fine particles, it was allowed to settle overnight or longer.
An amount of the crude pepsinogen solution, equivalent to 80,000 Azso units and containing 1.0 to 1.5 g of pepsinogen (see Table I for a typical experiment), was diluted to 7,900 ml with 0.05 M Tris-phosphate buffer and mixed with 2,100 ml of settled DEAE-cellulose (equivalent to about 190 g). The suspension was gently stirred mechanically for 1 hour and filtered through cloth on a 24-cm Buchner funnel. The moist cake was resus- pended in fresh 0.05 M Tris-phosphate buffer and transferred to a chromatographic column 7 X 60 cm. After settling, the height of the resin was 30 to 35 cm. The column was washed with
several liters of the same buffer until the absorbance of the effluent at 280 rnp fell to about 0.3. The pepsinogen was then eluted with 0.45 M NaCl in the same buffer. The elution profile is shown in Fig. 1.
The portion of the peak containing potential pepsin activity over 40 I.cg per A283 unit was pooled and concentrated to about 750 ml by ultrafiltration in a DiaFlo model 400 ultrafiltration cell with reservoir (Amicon Corporation, Cambridge, Massachusetts). The membrane used was No. UM-1. The solution was stirred very slowly during ultrafiltration to avoid denaturation. The ultrafiltrate contained a small amount of material which absorbed at 280 rnp but did not have potential peptic activity.
Step 4: Desalting on Sephadex G-%5-A mixture of coarse and fine Sephadex beads in the proportion of 9: 1 was used to give a flow rate of about 180 ml per hour. The, column, 5 X 100 cm, prepared as recommended by the manufacturer (28), was equilibrated with 0.01 M Tris-phosphate buffer. Up to 250 ml of the concentrated solution from Step 3 could be placed on the column at one time. The protein was eluted with 0.01 M
Tris-phosphate buffer. The column was used repeatedly after washing out the salt. The desalted protein was pooled.
Step 5: Second Batch Absorption on DEAE-cellulose, with Gradient Elution-The desalted solution, containing 15,180 Azso units and 910 mg of pepsinogen (Table I), was treated as in Step 3 with 100 g of DEAE-cellulose in a total volume of 5 liters. The suspension was filtered as before; the filtrate contained 2,100 AQgO units, but no pepsinogen. The filter cake was suspended in 1,200 ml of 0.05 M Tris-phosphate buffer, and the slurry was transferred to a column (5 x 60 cm) containing a 15-cm packed bed of fresh DEAE-cellulose, equilibrated with the same buffer. The gradient described by Arnon and Perlmann (5) for the purification of crystalline swine pepsinogen was adjusted to the size of the column used. The first constant volume bottle contained 2,250 ml of equilibrating buffer. The gradient solu- tion was 0.45 M NaCl in the same buffer. The elution profile is shown in Fig. 2. Fractions with potential peptic activity of over 100 pg per Azso unit were pooled in three portions, as indicated by the arrows. The center portion, used for the subsequent steps of purification, contained 348 mg.
Step 6: Recycling Gel Filtration on Sephodex G-100--Initially, a single gravity flow filtration was tried, but imperfect separation
EFFLUENT, L
FIG. 1. Stepwise elution of pepsinogen with 0.1 and 0.45 M NaCl in 0.05 M Tris-phosphate buffer of pH 7.0, after the first batch absorption on DEAE-cellulose (see the text, Step 3). The effluent was collected in large fractions, indicated by boxes, during elution with 0.1 M NaCl. The column was 7 X 31 cm; sample weight, 1433 mg; flow rate, 120 ml per hour. O--O, Az~o; A- - -A, specific activity.
by guest on April 11, 2020
http://ww
w.jbc.org/
Dow
nloaded from
Issue of April 25, 1968 R. B. Chow and B. Kassell 1721
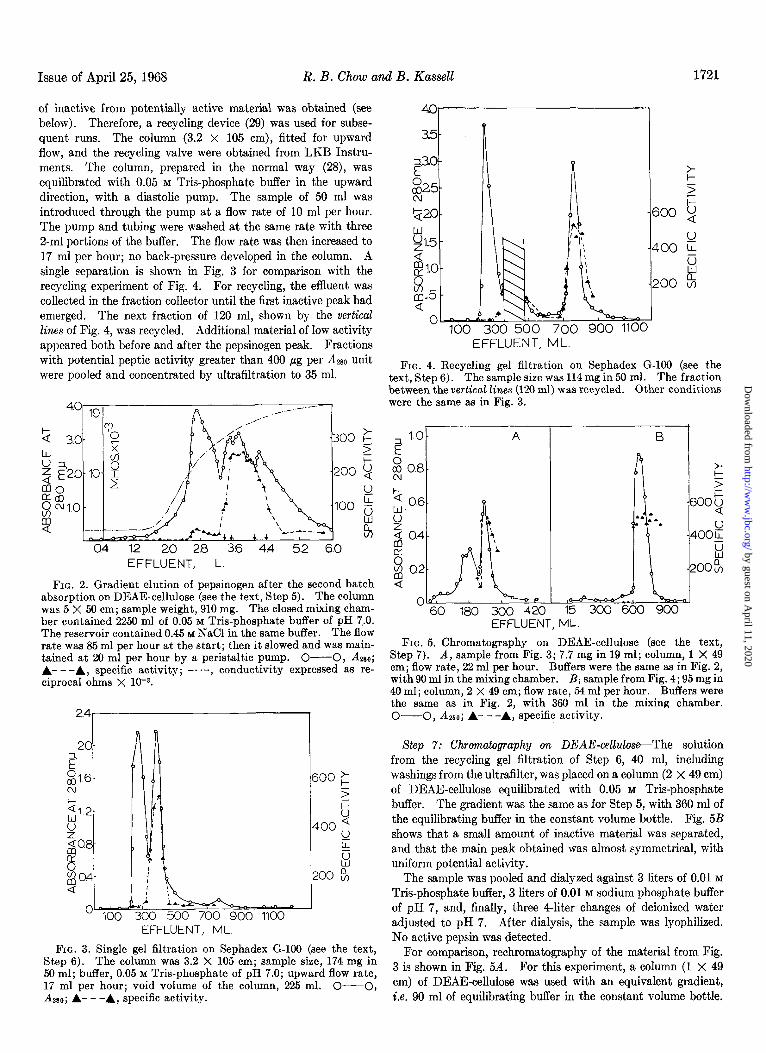
of inactive from potentially active material was obtained (see below). Therefore, a recycling device (29) was used for subse- quent runs. The column (3.2 x 105 cm), fitted for upward flow, and the recycling valve were obtained from LKB Instru- ments. The column, prepared in the normal way (28), was equilibrated with 0.05 M Tris-phosphate buffer in the upward direction, with a diastolic pump. The sample of 50 ml was introduced through the pump at a flow rate of 10 ml per hour. The pump and tubing were washed at the same rate with three 2-ml portions of the buffer. The flow rate was then increased to 17 ml per hour; no back-pressure developed in the column. A single separation is shown in Fig. 3 for comparison with the recycling experiment of Fig. 4. For recycling, the effluent was collected in the fraction collector until the first inactive peak had emerged. The next fraction of 120 ml, shown by the vertical Zincs of Fig. 4, was recycled. Additional material of low activity appeared both before and after the pepsinogen peak. Fractions with potential peptic activity greater than 400 pg per A280 unit were pooled and concentrated by ultrafiltration to 35 ml.
cI4 12 2.0 2.8 3.6 44 5.2 6.0 EFFLUENT, L
FIG. 2. Gradient elution of pepsinogen after the second batch absorption on DEAE-cellulose (see the text, Step 5). The column was 5 X 50 cm; sample weight, 910 mg. The closed mixing cham- ber contained 2250 ml of 0.05 M T&phosphate buffer of pH 7,.0. The reservoir contained 0.45 M NaCl in the same buffer. The flow rate was 85 ml per hour at the start; then it slowed and was main- tained at 20 ml per hour by a peristaltic pump. O-O, ⅈ A- - -A, specific activity; -. .-, conductivity expressed as re- ciprocal ohms X 1c3.
O 100 300 500 700 900 1100 EFFLUENT, ML.
FIG. 3. Single gel filtration on Sephadex G-100 (see the text, Step 6). The column was 3.2 X 105 cm; sample size, 174 mg in 50 ml; buffer, 0.05 M Tris-phosphate of pH 7.0; upward flow rate, 17 ml per hour; void volume of the column, 225 ml. O-O,
3.5-
$X0-
%5-
Q20-
g1.5 -
&.O-
8 s-5 -
200 %
0 100 300 500 700 900 1100
1
EFFLUENT, ML
FIG. 4. Recycling gel filtration on Sephadex G-100 (see the text, Step 6). The sample size was 114 mg in 50 ml. The fraction between the vertical lines (120 ml) was recycled. Other conditions were the same as in Fig. 3.
if 1.0
z 0.8 z 3
‘- 0.6
P
6005
$ 0.4
8
400:
g a2 aw
2OOcn Q
0 60 180 300 420 15 300 600 900
EFFLUENT, ML.
FIG. 5. Chromatography on DEAE-cellulose (see the text, Step 7). A, sample from Fig. 3; 7.7 mg in 19 ml; column, 1 X 49 cm; flow rate, 22 ml per hour. Buffers were the same as in Fig. 2, with 90 ml in the mixing chamber. B, sample from Fig. 4; 95 mg in 40 ml; column, 2 X 49 cm; flow rate, 54 ml per hour. Buffers were the same as in Fig. 2, with 360 ml in the mixing chamber. O-O, A2so; A- ---A, specific activity.
Step 7: Chromatography 072 DEAE-cellulose-The solution from the recycling gel filtration of Step 6, 40 ml, including washings from the ultrafilter, was placed on a column (2 X 49 cm) of DEAE-cellulose equilibrated with 0.05 M Tris-phosphate buffer. The gradient was the same as for Step 5, with 360 ml of the equilibrating buffer in the constant volume bottle. Fig. 5B shows that a small amount of inactive material was separated, and that the main peak obtained was almost symmetrical, with uniform potential activity.
The sample was pooled and dialyzed against 3 liters of 0.01 M
Tris-phosphate buffer, 3 liters of 0.01 M sodium phosphate buffer of pH 7, and, finally, three 4-liter changes of deionized water adjusted to pH 7. After dialysis, the sample was lyophiliaed. No active pepsin was detected.
For comparison, rechromatography of the material from Fig. 3 is shown in Fig. 5A. For this experiment, a column (1 X 49 cm) of DEAE-cellulose was used with an equivalent gradient, i.e. 90 ml of eauilibrating buffer in the constant volume bottle. A 280; A- - -A, specific activity. A
by guest on April 11, 2020
http://ww
w.jbc.org/
Dow
nloaded from
1722 Bovine Pepsinogen and Pepsin. I Vol. 243, No. 8
TABLE II Amino acid composition of bovine pepsinogen
Values are based on a molecular weight of 38,943 and a total nitrogen content of 14.560/,.
Amino acid
Lysine Histidine Arginine Aspartic acid . . Threonine Ferine Glutamic acid... Proline. Glycine Alanine. Half-cystine. Valine Methionine . . Isoleucine. Leucine. Tyrosine Phenylalanine. Tryptophan.. NH,... .:‘::‘.:
Total.
I- i-
Amino acid residues per molecule
16 hrs
- 24 hrs 48 hrs 12 hrs No. per molecule
8.44 7.94 7.87 8.07 1.98 1.98 2.01 2.02 5.92 5.83 5.82 5.88
42.24 39.21 38.47 40.65 25.83 25.87 24.22 23.67 46.49 46.49 40.12 37.19 35.17 29.78 30.31 33.03 15.42 15.83 15.29 14.95 3G.12 34.01 34.14 35.09 18.75 15.50 15.50 15.96
22.90 22.96 24.47 24.91 4.20 3.78 3.52 3.67
29.39 30.46 30.78 31.53 25.33 24.93 24.73 25.12 17.59 17.04 16.74 16.64 15.26 14.84 14.88 15.04
41.76 42.93 50.73 56.79
a Based on sample weights. 1 Seventy-two-hour hydrolysate. b Molecular weight 38,944 (F). 0 Includes alloisoleucine. c Average of all four hydrolysates. h Phenylalanine was taken as 15 residues to calculate the resi-
Calculated residues
8.03c f 0.16 1.99 f 0.03 5.85” f 0.05
40.14c f 1.67 2G.G2d f 0.28 49.84d f 0.90 32.0’ic f 2.51 15.37c 3z 0.38 34.84c f 0.98 15.920 f 0.59 5.86e
24.911 3.79” f 0.29
31.53/* 0 25.03” f 0.26 17.59d f 0.21 15.0C*hf 0.20 5.75”
36.93d f 0.53
-
--
-
Per cent0
2.43 0.64 2.16
10.89 6.34
10.23 9.75 3.44 4.89 2.67 1.44 5.81 1.17 8.41 8.68 G.76 5.20 2.75
% of iOld
3.64 1.35 5.31 9.09 G.03
11.29 7.27 3.48 7.90 3.GO 1.36 5.64 0.80 7.14 5.07 3.98 3.40 2.84
10.17
8 10 2 3 6 4
40 44 27 26 50 46 32 28 15 19 35 35 16 19 B 6
25 23 4 4
32 25 25 33 18 17 15 15
(3;) (4
91.46 100.0 362 f 2 363
Nitrogena
Residues per molecule to nearest integer
Bovine Swine (6Jb
d Intercept of a straight line calculated by the method of least dues of amino acids per mole. squares. i Tryptophan was determined spectrophotometrically (22).
* Determined as cysteic acid (21).
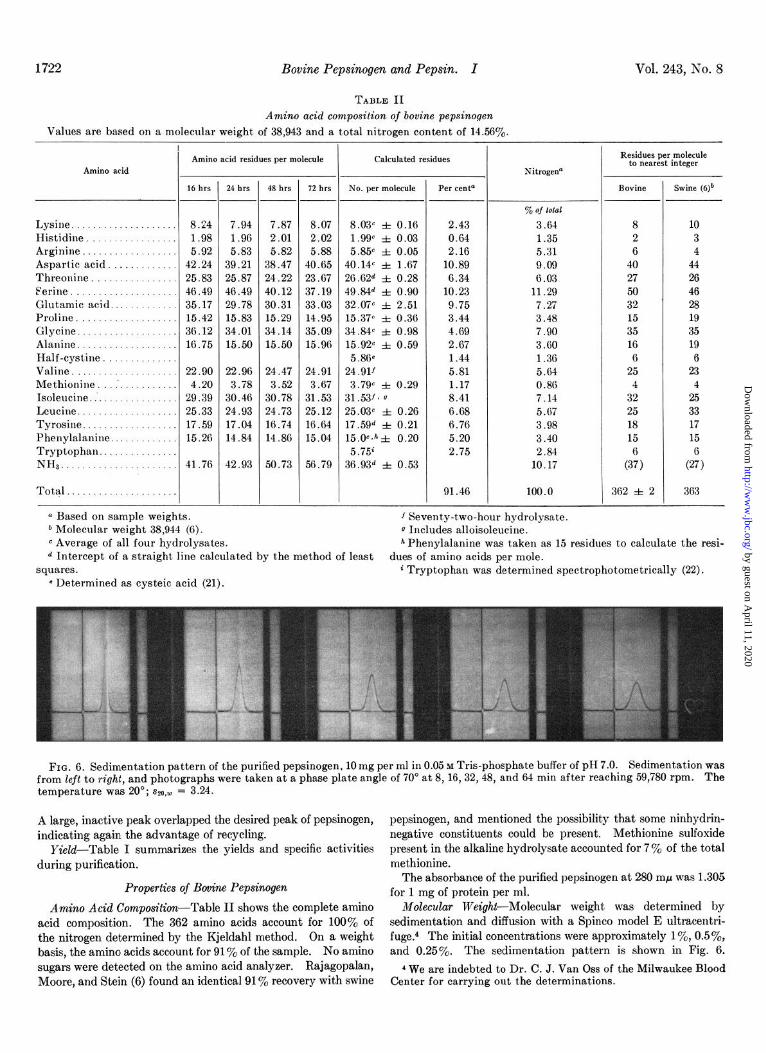
FIG. 6. Sedimentation pattern of the purified pepsinogen, 10 mg per ml in 0.05 w Tris-phosphate buffer of pH 7.0. Sedimentation was from left to right, and photographs were taken at a phase plate angle of 70’ at 8, 16,32,48, and 64 min after reaching 59,780 rpm. The temperature was 20”; ~~0,~ = 3.24.
A large, inactive peak overlapped the desired peak of pepsinogen, pepsinogen, and mentioned the possibility that some ninhydrin- indicating again the advantage of recycling. negative constituents could be present. Methionine sulfoxide
Yield-Table I summarizes the yields and specific activities present in the alkaline hydrolysate accounted for 7 y0 of the total
during purification. methionine.
Properties of Bovine Pepsinogen The absorbance of the purified pepsinogen at 280 rnp was 1.305
for 1 mg of protein per ml. Amino Acid Composition-Table II shows the complete amino Molecular Weight-Molecular weight was determined by
acid composition. The 362 amino acids account for 100% of sedimentation and diffusion with a Spinco model E ultracentri-
the nitrogen determined by the Kjeldahl method. On a weight fuge.4 The initial concentrations were approximately lye, 0.5%, basis, the amino acids account for 91% of the sample. No amino and 0.25%. The sedimentation pattern is shown in Fig. 6. sugars were detected on the amino acid analyzer, Kajagopalan, 4 We are indebted to Dr. C. J. Van Oss of the Milwaukee Blood Moore, and Stein (6) found an identical 91% recovery with swine Center for carrying out the determinations.
by guest on April 11, 2020
http://ww
w.jbc.org/
Dow
nloaded from
Issue of April 25, 1968 R. B. Chow and B. Kassell 1723
3.1 a
$ a.1 4 t
2 w .I o-
Y $ .06-
5 $ .02-
EFFLUENT, ML
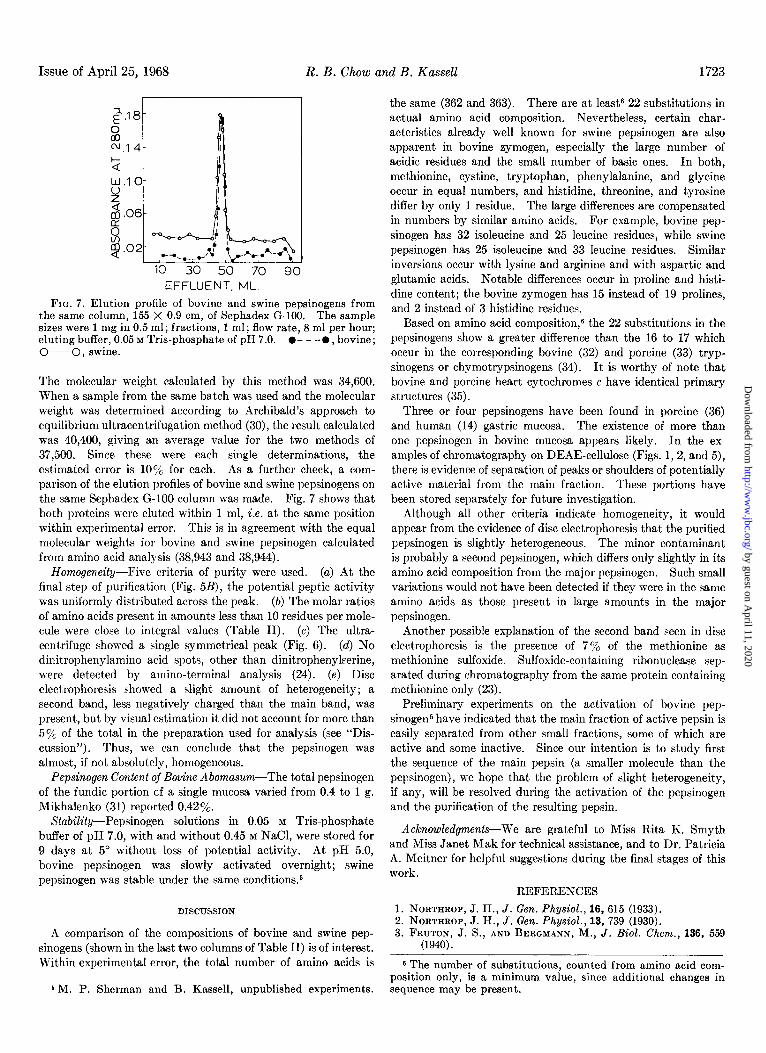
FIG. 7. Elution profile of bovine and swine pepsinogens from the same column, 155 X 0.9 cm, of Sephadex G-100. The sample sizes were 1 mg in 0.5 ml; fractions, 1 ml; flow rate, 8 ml per hour; eluting buffer, 0.05 M Tris-phosphate of pH 7.0. l - - -0, bovine; O-O, swine.
The molecular weight calculated by this method was 34,600. W7hen a sample from the same batch was used and the molecular weight was determined according to Archibald’s approach to equilibrium ultracentrifugation method (30), the result calculated was 40,400, giving an average value for the two methods of 37,500. Since these were each single determinations, the e&mated error is 10% for each. As a further check, a com- parison of the elution profiles of bovine and swine pepsinogens on the same Sephadex G-100 column was made. Fig. 7 shows that both proteins were eluted within 1 ml, i.e. at the same position within experimental error. This is in agreement with the equal molecular weights for bovine and swine pepsinogen calculated from amino acid analysis (38,943 and 38,944).
liTomogeneity--Five criteria of purity were used. (a) At the final step of purification (Fig. 5B), the potential peptic activity was uniformly distributed across the peak. (b) The molar ratios of amino acids present in amounts less than 10 residues per mole- cule were close to integral values (Table II). (c) The ultra- centrifuge showed a single symmetrical peak (Fig. 6). (d) No dinitrophenylamino acid spots, other than dinitrophenylserine, were detected by amino-terminal analysis (24). (e) Disc electrophoresis showed a slight amount of heterogeneity; a second band, less negatively charged than the main band, was present, but by visual estimation it did not account for more than 5y0 of the total in the preparation used for analysis (see “Dis- cussion”). Thus, we can conclude that the pepsinogen was almost, if not absolutely, homogeneous.
Pepsinogen Content of Bovine Abomasum-The total pepsinogen of the fundic portion of a single mucosa varied from 0.4 to 1 g. Mikhalenko (31) reported 0.42%.
Stability-Pepsinogen solutions in 0.05 M Tris-phosphate buffer of pH 7.0, with and without 0.45 M NaCl, were stored for 9 days at 5” without loss of potential activity. At pH 5.0, bovine pepsinogen was slowly activated overnight; swine pepsinogen was stable under the same conditions5
DISCUSSION
A comparison of the compositions of bovine and swine pep- sinogens (shown in the last two columns of Table II) is of interest. Within experimental error, the total number of amino acids is
5 M. P. Sherman and B. Kassell, unpublished experiments.
the same (362 and 363). There are at least! 22 substitutions in actual amino acid composition. Nevertheless, certain char- acteristics already well known for swine pepsinogen are also apparent in bovine zymogen, especially the large number of acidic residues and the small number of basic ones. In both, methionine, cystine, tryptophan, phenylalanine, and glycine occur in equal numbers, and histidine, threonine, and tyrosine differ by only 1 residue. The large differences are compensated in numbers by similar amino acids. For example, bovine pep- sinogen has 32 isoleucine and 25 leucine residues, while swine pepsinogen has 25 isoleucine and 33 leucine residues. Similar inversions occur with lysine and arginine and with aspartic and glutamic acids. Notable differences occur in proline and histi- dine content; the bovine zymogen has 15 instead of 19 prolines, and 2 instead of 3 histidine residues.
Based on amino acid composition,6 the 22 substitutions in the pepsinogens show a greater difference than the 16 to 17 which occur in the corresponding bovine (32) and porcine (33) tryp- sinogens or chymotrypsinogens (34). It is worthy of note that bovine and porcine heart cytochromes c have identical primary structures (35).
Three or four pepsinogens have been found in porcine (36) and human (14) gastric mucosa. The existence of more than one pepsinogen in bovine mucosa appears likely. In the ex- amples of chromatography on DEAE-cellulose (Figs. 1,2, and 5), there is evidence of separation of peaks or shoulders of potentially active material from the main fraction. These portions have been stored separately for future investigation.
Although all other criteria indicate homogeneity, it would appear from the evidence of disc electrophoresis that the purified pepsinogen is slightly heterogeneous. The minor contaminant is probably a second pepsinogen, which differs only slightly in its amino acid composition from the major pepsinogen. Such small variations would not have been detected if they were in the same amino acids as those present in large amounts in the major pepsinogen.
Another possible explanation of the second band seen in disc electrophoresis is the presence of 7% of the methionine as methionine sulfoxide. Sulfoxide-containing ribonuclease sep- arated during chromatography from the same protein containing methionine only (23).
Preliminary experiments on the activation of bovine pep- sinogens have indicated that the main fraction of active pepsin is easily separated from other small fractions, some of which are active and some inact,ive. Since our intention is to study first the sequence of the main pepsin (a smaller molecule than the pepsinogen), we hope that the problem of slight heterogeneit,y, if any, will be resolved during the activation of the pepsinogen and the purification of the resulting pepsin.
Acknowledgments-We are grateful to Miss Rita K. Smyth and Miss Janet Mak for technical assistance, and to Dr. Pat,ricia A. Meitner for helpful suggestions during the final stages of this work.
REFERENCES
1. NORTHROP, J. H., J. Gen. Physiol., 16, 615 (1933). 2. NORTHROP. J. H.. J. Gen. Phusiol.. 13. 739 (1930). 3. FRUTON, J: S., A& BERG&W, Ik, 2. B&l. -&em., 136, 559
(1940).
6 The number of substitutions, counted from amino acid com- position only, is a minimum value, since additional changes in sequence may be present.
by guest on April 11, 2020
http://ww
w.jbc.org/
Dow
nloaded from

1724 Bovine Pepsinogen and Pepsin. I Vol. 243, No. 8
4. BERGMANN, M., AND FRUTON, J. S., Advance. Enzymol., 1, 63 (1941).
5. ARNON, R., AND PERLMANN, G. E., J. Biol. Chem., 233, 653 (1963).
6. RAJAGOPALAN, T. G., MOORE, S., AND STEIN, W. H., J. Biol. Chem., 241, 4940 (1966).
7. DOPHEIDE, T. A. A., MOORE, S., AND STEIN, W. H., J. Biol. Chem., 242, 1833 (1967).
8. PERHAM. R. N., AND JONES, G. M. T., Europe. J. Biochem., 2, 84 (1967). ’
_
9. STEPANOV, V. M., VAKHITOVA, E. A., EGOROV, C. A., AND AVAEVA. S. M., Biochim. Biovhvs. Acta. 110. 632 (1965).
10. K~ZNET&V, Y. S:, KOVALEVA, G. G., AND STEP~NOV, ?. Ml, Biochim. Biophys. Acta, 118, 219 (1966); Biokhimiya, 31, 781 (1966).
11. KEIL, B., MORAVEK, L., AND FORM, F., Collect. Czech. Chem. Cohmuh., 32, 196s (1967).
12. OESTREICHER, A. B., AND PERLMANN, G. E., Biochim. Biophys. Acta, 140, 319 (1967).
13. ANSON, M. L., J. Gen. Physiol., 22, 79 (1938). 14. SEIJFFERS. M. J.. SEGAL. H. L.. AND MILLER. L. L.. Amer.
J. Physibl., 206; 1099 (1963). ’ 15. CLAESSON, O., Lantbrukshogskol Ann., 31, 237 (1965). 16. MOORE, S., AND STEIN, W. H., in S. P. COLOWICK AND N. 0.
KAPLAN (Editors), Methods in enzymology, Vol. VI, Aca- demic Press, New York, 1963, p. 819.
17. MILLER, L., AND HOUGHTON, J. A., J. Biol. Chem., 169, 373 (1945).
18. SPACKMAN, D. H., STEIN, W. H., AND MOORE, S., Anal. Chem., 30, 1190’(1958)..
19. SPACKMAN. D. H.. Fed. Proc.. 22. 244 (1963). 20. ROCKSTEI~, M., END HERRON, P. W., ‘Anai. Chem., 23, 1500
(1951).
21. MOORE, S., J. Biol. Chem., 233, 235 (1963). 22. BEAVEN, G. H., AND HOLIDAY, E. R., Advance. Protein Chem.,
7, 369 (1952). 23. NEUMANN, N. P., MOORE, S., AND STEIN, W. H., Biochemistry,
1, 68 (1962). 24. FRAENKEL-CONRAT, H., HARRIS, J. I., AND LEVY, A. L., Method.
Biochem. Anal., 2, 360 (1955). 26. WILLIAMS. D. E.. AND REISFELD. R. A.. Ann. N. Y. Acad. Sci..
121,373’ (1964): 26. HERRIOTT, R. M., J. Gen. Physiol., 21, 501 (1938). 27. Whatman Data Manual No. 6000, H. Reeve Angel, Clifton
New Jersey. 28. Sephadex 8, theory and experimental technique, Pharmacia,
Uppsala, Sweden. 29. PORATH, J., AND BENNICH, H., Arch. Biochem. Biophys.,
Suppl. 1, 152 (1962). 30. CLAESSON, S., AND MORING-CLAESSON, I., in P. ALEXANDER
AND R. J. BLOCK (Editors), A laboratory manual of analytical chemistry, Vol. III, Pergamon Press, New York, 1961, p. 114.
31. MIKHALENKO, B. S., Prikladnaya Biokhim. Mikrobiol., 3, 126 (1967).
32. WALSH, K. A., AND NEURATH, H., Proc. Nat. Acad. Sci. U. S. A., 62, 884 (1964).
33. CHARLES, M., ROVERY, M., GUIDONI, A., AND DESNUELLE, P., Biochim. Biophys. Acta, 69, 115 (1963).
34. CHARLES, M., GRATECOS, D., ROVERY, M., AND DESNUELLE, P., Biochim. Biophys. Acta, 140, 395 (1967).
35. SMITH, E. L., AND MARGOLIASH, E., Fed. Proc., 23,1243 (1964). 36. RYLE, A. P., in H. N. MUNRO (Editor), The role of the gastro-
intestinal tract in protein metabolism; a symposium of the Council of International Organizations of Medical Science, Blackwell Scientific Publications, Oxford, 1964, p. 25.
by guest on April 11, 2020
http://ww
w.jbc.org/
Dow
nloaded from

Rosie B. Chow and Beatrice KassellPROPERTIES OF THE PEPSINOGEN
Bovine Pepsinogen and Pepsin: I. ISOLATION, PURIFICATION, AND SOME
1968, 243:1718-1724.J. Biol. Chem.
http://www.jbc.org/content/243/8/1718Access the most updated version of this article at
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
http://www.jbc.org/content/243/8/1718.full.html#ref-list-1
This article cites 0 references, 0 of which can be accessed free at
by guest on April 11, 2020
http://ww
w.jbc.org/
Dow
nloaded from





























