Embed Size (px)
Citation preview

Blocking dephosphorylation at Serine 120 residue in t-SNARESNAP-23 leads to massive inhibition in exocytosis from mast cells
PIEU NASKAR, NILOFER NAQVI and NITI PURI*
Cellular and Molecular Immunology Laboratory, School of Life Sciences, Jawaharlal Nehru University,New Delhi 110 067, India
*Corresponding author (Email, [email protected], [email protected])
MS received 3 June 2017; accepted 29 January 2018; published online 17 February 2018
Mast cells (MCs) respond to allergen challenge by release of pre-stored inflammatory mediators from their secretorygranules, on cross-linking of Fce receptor I (FceRI) receptors. The target-SNARE (t-SNARE) SNAP-23 has been shown toplay an important role in MC exocytosis and undergoes transient phosphorylation at Serine 95 (S95) and Serine 120 (S120),concomitant with mediator release. During current study we explored the importance of transient nature of phosphorylationat S120 in MC exocytosis. A phosphomimetic SNAP-23-S120D mutant of rodent SNAP-23 was cloned into EGFP vectorand its effect on the exocytosis and the mechanisms involved was studied in RBL-2H3 MC line. Secretion reporter assaywith SNAP-23-S120D transfected MCs revealed a very significant inhibition of exocytosis, and reduced ruffling inresponse to FceRI cross-linking. Further, the effect of this mutation on localization of SNAP-23 in MCs was studied.Immunofluorescence microscopy studies and membrane-cytosol fractionation of green fluorescent protein-tagged SNAP-23-S120D (GFP-SNAP-23-S120D) transfected MCs showed that a large proportion of GFP-SNAP-23-S120D was residingin cytosol unlike wild-type SNAP-23, in resting and activated MCs and even the membrane associated portion was oninternal lysosomal membranes than plasma membrane. These studies imply that dephosphorylation of S120 is important forSNAP-23 membrane association dynamics and subsequently MC degranulation.
Keywords. Allergic response; dephosphorylation; inflammation; mast cell; SNAP-23
Abbreviations: DNP-BSA, Dinitrophenyl-bovine serum albumin; ECL, Enhanced chemiluminescence; EDTA, Ethylenediamine tetra acetic acid; EGFP, Enhanced green fluorescent protein; EGTA, Ethylene glycol bis (2-aminoethyl ether) tetraacetic acid; FceRI, Fce receptor I; HEPES, N-2-hydroxyethylpiperazine-N0-2-ethanesulfonic acid; IgE, Immunoglobulin E;LAMP, Lysosomal associated membrane protein; mAb, Monoclonal antibody; MC, Mast cell; P. coefficient, Pearson’scoefficient; PBS, Phosphate buffered saline; PFA, Paraformaldehyde; PVDF, Polyvinylidene fluoride; RBL, Rat basophilicleukemia; SG, Secretory granule; SNAP-23, Synaptosomal associated protein of 23 kDa; SNARE, Soluble N-ethylmaleimide sensitive factor (NSF) attachment protein receptor; t-SNAREs, Target-SNARE; VAMP, Vesicleassociated membrane protein; v-SNAREs, Vesicle-SNARE; Syt, Synaptotagmin
1. Introduction
Mast cells are specialized secretory cells that play a crucialrole in inflammation and allergic responses, and also inprotection against pathogens, in wound healing, angiogen-esis and cancer metastasis (Kalesnikoff and Galli 2008; Urband Sheppard 2012). In all these roles, they sense antigens,allergens or pathogens with the help of various receptorspresent on their surface and release pre-stored mediators byregulated exocytosis. During hypersensitivity reactions indifferent diseases like asthma, allergic rhinitis, atopic der-matitis, food and drug allergies, they are mainly activated bycross-linking of Fce receptor I (FceRI) boundImmunoglobulin E (IgE) by a multivalent allergen (Kraftand Kinet 2007). This physiological trigger initiates a
cascade of events that results in the translocation, docking,and fusion of secretory granule with plasma membrane (Urband Sheppard 2012) leading to release of inflammatorymediators stored in the secretory granules (Puri and Roche2008; Galli et al. 2008). This process proceeds throughcompound exocytosis where either the vesicles fuse witheach other prior to plasma membrane fusion (multivesicularexocytosis) or in a sequential manner, i.e. one after anotherunderneath the plasma membrane (sequential exocytosis)(Lorentz et al. 2012).
A class of proteins termed SNAREs (Soluble NSFattachment protein receptor) [including the ubiquitouslyexpressed SNAP-23 and its neuronal counterpart SNAP-25(Synaptosomal associated protein of 23 kDa and 25 kDarespectively), Syntaxins and VAMPs (Vesicle associated
http://www.ias.ac.in/jbiosci 127
J Biosci Vol. 43, No. 1, March 2018, pp. 127–138 � Indian Academy of SciencesDOI: 10.1007/s12038-018-9740-y

membrane proteins)] are known regulators of plasma mem-brane-secretory granule fusion events in various secretorycells (Jahn and Sudhof 1999; Lin and Scheller 2000).Through hydrophobic interactions in their coiled-coildomains t-SNARE and its cognet v-SNARE form a fourhelical bundle (Poirier et al. 1998), so-called trans complexthat is minimally required for membrane fusion (Weber et al.1998).
Given that SNAREs play a central role in membrane fusionprocess, it is imperative that their function is also modulated.Post-translational modifications of SNARE proteins and thebinding of regulatory proteins are two of the mechanisms thatcan generate temporal and spatial regulation of trans-SNAREcomplex formation. Some of these regulatory mechanismshave been studied in mast cells, which are packed withSNARE decorated granules that must remain inactive inresting cells, but on cell activation, must function rapidly andon a large scale to allow for membrane fusion and degranu-lation. In particular, protein kinases and phosphatases, whichhave been extensively associated with the regulation ofexocytosis, could participate in SNARE function by phos-phorylating residues essential in SNARE complex assemblyor the binding of SNARE regulatory proteins. Many serine/threonine kinases are shown to be involved in cell differen-tiation, proliferation, apoptosis, and autophagy. Even in thecontext of regulation of exocytosis many phosphorylationevents are known to occur. In resting mast cells phosphory-lation by Rab3D-associated kinase renders the t-SNARESyntaxin-4 inactive (Blank et al. 2002). In bone marrow mastcells (BMMCs) Src homology 2-containing inositol 50-phosphatase (SHIP) was found, that negatively regulatesmast cell degranulation (Huber et al. 1998).
Our previous study on the molecular mechanisms ofregulated exocytosis in mast cells revealed that t-SNARESNAP-23 is transiently phosphorylated at S95 and Ser120during mast cells degranulation (Hepp et al. 2005). Thekinetics of SNAP-23 phosphorylation also mirrors thekinetics of exocytosis. Other researchers have explored andspeculated on the kinase involved in this phosphorylation(Suzuki and Verma 2008; Karim et al. 2013). Recently, wehave explored the importance of joint phosphorylation anddephosphorylation of SNAP-23 at these two residues S95and S120 for mast cell exocytosis and SNAP-23 membranedynamics (Naskar and Puri 2017). In the present study, wedecided to explore the importance of a single residue Ser120in mast cell exocytosis. That too, specifically we have triedto illustrate the importance of dephosphorylation of S120after transient phosphorylation on degranulation from mastcells. For this, RBL-2H3 cell line, the established model forstudying degranulation from mast cells in response to FceRIreceptor by multivalent allergen and IgE, was used (Heppet al. 2005; Naskar and Puri 2017). MCs were activated bycross-linking of DNP-specific IgE sensitized mast cells withthe multivalent antigen dinitrophenyl-bovine serum albumin
(DNP-BSA), exactly as on mast cells of atopic individualsduring an allergen challenge (Gould et al. 2003; Naskar andPuri 2017). SNAP-23 phosphomimetic mutant was made byreplacing the serine 120 residue to aspartate, and calledSNAP-23 S120D, to study the importance of dephosphory-lation on mast cell exocytosis. For better visualization andquantitation of intracellular localization of this mutant indifferent physiological states in biochemical as well asmicroscopic studies over expressed enhanced green fluo-rescent protein (EGFP)-tagged SNAP-23 S120D was used.Our studies reveal the utmost importance of dephosphory-lation following phosphorylation at a single S120 in SNAP-23 during various events culminating in efficient mast cellmediator release.
2. Materials and methods
2.1 Cell culture
Rat basophilic leukemia (RBL)-2H3 cells are taken as mastcell model, (kind gift from Dr. Paul A Roche, NationalInstitutes of Health, Bethesda, MD, USA). Cell culture wasdone in equal parts minimal essential medium (Sigma, MO,USA) and Iscove’s medium (Gibco, life technologies, GrandIsland, NY, USA) containing 20% fetal bovine serum (FBS)(Gibco, life technologies, Grand Island, NY, USA), 25 mMN-2-Hydroxyethylpiperazine-N0-2-ethanesulfonic acid(HEPES) (Sigma, MO, USA), and 120 lg/ml gentamicin(RBL complete medium). Cells were maintained as sub-confluent monolayers in a humidified atmosphere containing5% CO2 at 37�C and passaged with trypsin.
2.2 Plasmids
pCMV-FLAG-Rat SNAP-23 and pCDNA3-hGH constructwere obtained from Dr. Paul A Roche (National Institutes ofHealth, Bethesda, MD, USA) (described previously in (Heppet al. 2005, Puri et al. 2003)). The cDNA encoding full lengthwild-type SNAP-23 was subcloned into EGFP-C2 plasmid(Clontech, CA, USA) by using EcoRI and ApaI restriction sitesto generate amino terminal GFP-tagged protein (called GFP-SNAP-23). Point mutation at Serine 120 site to aspartate(D) was introduced by Quick Change Multi-Mutagenesis kit(Stratagene) as per the manufacture’s instructions. The integrityof subcloned plasmid was confirmed by sequencing from GCCBiotech and SciGenom Labs Pvt Ltd., India.
2.3 Antibodies
Anti–DNP IgE (clone TIB 142) was obtained from theAmerican Type Culture Collection (Manassas, VA, USA).
128 P Naskar, N Naqvi and N Puri

Rabbit anti-serum recognizing the SNAP-23 carboxyl ter-minus and SNAP-23 phosphorylated at S120 were a giftfrom Paul A. Roche (National Institutes of Health, Bethesda,MD, USA). As this antibody is specific for phosphorylatedSNAP-23, and does not bind SNAP-23 that is not phos-phorylated at S120, it detects phosphorylation of SNAP-23at S120 (Hepp et al. 2005; JBC). Anti-CD63/lysosomeassociated membrane protein (LAMP) 3 monoclonal anti-body (mAb) AD1 (BD Biosciences, San Diego, CA, USA)was used for immunofluorescence microscopy. Alexa dye-conjugated secondary antibodies were obtained fromMolecular Probes (Eugene, OR, USA). Anti-Rabbit ProteinA-HRP conjugated antibody (Southern Biotech, Birming-ham, AL, USA) and anti-mouse IgG-HRP conjugated anti-body (Southern Biotech, Birmingham, AL, USA) were used.
2.4 Stimulation of RBL cell exocytosis
Exocytosis of RBL cells was monitored by measuring theb-hexosaminidase activity released from cells as describedpreviously (Vaidyanathan et al. 2001; Hepp et al. 2005).Briefly, Sub-confluent RBL cells were sensitized with anti-DNP IgE (TIB 142 tissue culture supernatant at requireddilution) in RBL complete medium overnight at 37�C.After a washing step, IgE sensitized cells were either mock-stimulated or stimulated with 100 ng/ml DNP-BSA inphenol red free RPMI medium (Sigma, MO, USA) for 10min. Cell supernatants were collected and the cells werelysed for 1 h in ice-cold phenol red-free RPMI mediumcontaining 0.2% Triton X-100 (Sigma, MO, USA). A col-orimetric assay with p-nitrophenyl-N-acetyl-b-D-glu-cosaminide (Sigma Aldrich, MO, USA) as the substratewas used to measure the b-hexosaminidase content inmedium and remaining in the cells. The b-hexosaminidaseactivity released was expressed as a percentage of theactivity released into the medium relative to the totalactivity (released plus cell-associated). Each independentexperiment was repeated at least three times. RBL cellswere transfected with pCDNA3 hGH (2 lg) by electropo-ration together with GFP-SNAP-23 wild-type or phosphor-mutant (20 lg). After 4–5 h the cells were sensitized withanti DNP IgE. After 16–18 h they were mock-stimulated orstimulated with DNP-BSA as described before (Hepp et al.2005). The human growth hormone enzyme-linkedimmunosorbent assay (Roche Diagnostics Corp.) was usedto determine the amount of human growth hormonereleased into the medium or left in the cell, as describedpreviously (Puri et al. 2003). For quantitative experiments,statistical analyses were carried out by using a Student’s t-test. Results were considered significant when a p-value ofless than 0.05 was obtained. Statistical calculations weremade using a one-tailed distribution in a two-sample equalvariance Student’s t-test.
2.5 Confocal microscopy
RBL cells were seeded on 10 mm diameter coverslips withor without transfection, 18 h before analysis. They werestimulated or mock stimulated as described above. Forimmunofluorescence analysis, the cells were either fixedwith 4% paraformaldehyde (PFA) in phosphate bufferedsaline (PBS) for 30 min and excess paraformaldehydequenched with 50 mM NH4Cl in PBS. After washing, thefixed cells were permeabilized with 1% IGEPAL (Sigma,MO, USA) in a buffer containing PBS, 3% normal goatserum (Sigma, MO, USA) and 0.05% saponin (SD finechem. Limited, Boisar, India). To prevent nonspecific pro-tein binding the cells were then incubated with 3% normalgoat serum and 0.05% saponin in PBS for 1 h. Primary Absdiluted in the same buffer were added to the cells, andincubation was conducted for 1–2 h. After washing, the cellswere incubated for 30 min in the presence of secondary goatAbs conjugated to Alexa Fluor 546 (red) (Molecular Probes,Eugene, OR, USA). As a control, samples were stained withan irrelevant antibody and no staining was observed in therespective channel for all confocal fluorescence microscopyexperiments.
Olympus Fluoview FV1000 microscope and AndorSpinning Disk Confocal microscope (Nikon Eclipse TiE,Software-Andor iQ 2.7) were used to collect the images at100X magnification (sometimes with 2X zoom). an opticalslice thickness of 1.0 lm. Image Z-stacks were collectedthrough the depth of the cell using 0.4 lm step size. Co-localization analysis was done for each plane of the indi-vidual image stacks using the co-localization analysis featureof the Fluoview software Ver.1.7a (Olympus) and organizedinto figures using Microsoft Power Point 2007.
2.6 Transfection of RBL cells
Transfection of RBL cells was performed as described ear-lier (Puri et al. 2003) with some modifications. Briefly,exponentially growing RBL cells (10 9 106/ 0.5 ml serumfree media) were transfected by electroporation (320 mV,960 lF) with 20 lg DNA. Immediately after electroporation,the cells were plated in complete medium and analysed 24 hlater. For microscopy, transfected cells were plated on cov-erslips and then treated as per requirement. For Flowcytometry the untransfected/transfected cells were washedwith PBS followed by fixation with 4% PFA. Then theywere analysed by flow cytometry. To prepare lysate theuntransfected/transfected cells were washed with PBS andlysed for 1h in ice-cold lysis buffer [10 mM Tris (pH 7.4),150 mM NaCl containing 1% Triton X-100 as well as pro-tease inhibitors (5 mM iodoacetamide, 50 mM phenyl-methylsulfonyl fluoride, and 0.1 mM Na-p-tosyl-L-lysinechloromethyl ketone, from Sigma, MO, USA) and
SNAP-23 dephosphorylation in mast cell secretion 129

phosphatase inhibitors (5 mM Ethylene diamine tetra aceticacid (EDTA), 5 mM Ethylene glycol bis (2-aminoethylether) tetra acetic acid (EGTA), 50 mM NaF, 10 mM Na4-P2O7, and 1mM Na3VO4) from Sigma, MO, USA]. Thenlysates were analysed by SDS-PAGE and immunoblotting.
2.7 Flow cytometry
10 9 106 RBL mast cells were transfected with EGFP-C2vector and EGFP-SNAP-23 plasmids separately. After 24 hthe cells were harvested by trypsinization and counted bytrypan blue exclusion method. They were then washed twotimes with phosphate buffered saline and then immediatelyanalysed by flow cytometer (BD FACS Calibur, CA, USA)(Cell Questpro software). In each case cell count was 10x103
during acquisition. Statistical analysis was done by using aStudent’s t test.
2.8 Membrane-cytosol fractionation
Membrane-cytosol fractionation was done as described earlier(Hepp et al. 2005). Briefly, transfected RBL mast cells wereharvested by trypsinization after 24 h and washed in phosphatebuffered saline. The cells were resuspended in hypotonic buffer(10 mM Tris, 1 mM KCl, 1 mM EGTA, 0.5 mM MgCl2, pH7.4) containing protease inhibitors (5mM iodoacetamide,50mM phenylmethylsulfonyl fluoride, and 0.1 mM Na-p-tosyl-L-lysine chloromethyl ketone) and phosphatase inhibitors (5mM EDTA, 5 mM EGTA, 50 mM NaF, 10 mM Na4P2O7, and1 mM Na3VO4). They were then disrupted by repeated passageof cells through a 25-gauge syringe. Nuclei and unbroken cellswere removed by centrifugation at 1000g and the post-nuclearsupernatant was subjected to centrifugation at 100,000g for 1hat 4�C to isolate membrane (pellet) and cytosol (supernatant).The membrane pellet and cytosolic supernatant were brought tothe same volume in hypotonic buffer and each was adjusted toa final concentration of 1% Triton X-100. Equal portions ofeach fraction were analysed by SDS-PAGE andimmunoblotting.
2.9 SDS PAGE and immunoblotting
RBL cell lysates and membrane-cytosol fractions wereboiled in b-mercaptoethanol containing sample buffer andthen proteins were separated in 12.5% SDS-polyacrylamidegel. Immunoblotting was performed with SNAP-23 C-ter-minus antibody or GFP antibody clone A.v. (Clontech, CA,USA) as previously described (Puri et al. 2003). ForImmunoblotting separated proteins were transferred to 0.2 lPVDF membrane (Bio-Rad, USA) and visualized byenhanced chemiluminescence (ECL) using Immobilon
Western Chemiluminescence HRP substrate (Millipore, MA,USA). Band intensity was determined by Spot Denso (AlphaEaseFC software, Alpha Innotech). SNAP-23 membranelocalization is calculated as [{Band intensity of SNAP-23 inmembrane fraction/ (Band intensity of SNAP-23 in mem-brane fraction? Band intensity of SNAP-23 in cytosolicfraction)} 9 100]. That is, we took the percent of SNAP-23membrane association with respect to total SNAP-23 (Bandintensity of SNAP-23 in membrane fraction? Band intensityof SNAP-23 in cytosolic fraction).
For quantitative experiments, statistical analyses werecarried out by using a Student’s t test. Results were con-sidered significant when a p value of less than 0.05 wasobtained.
2.10 Immunoprecipitation
Immunoprecipitation from cell lysates was accomplished asdescribed previously (Hepp et al. 2005). Briefly, RBL cellswere lysed in ice-cold lysis buffer for 1 h. The lysis buffercomprises 10 mM Tris (pH 7.4), 150 mM NaCl containing1% Triton X-100 with protease inhibitors (50 mM phenyl-methylsulfonyl fluoride, 5 mM iodoacetamide, and 0.1 mMNa-p-tosyl-L-lysine chloromethyl ketone) and phosphataseinhibitors (5 mM EGTA, 5 mM EDTA, 10 mM Na4P2O7,50 mM NaF, and 1 mM Na 3VO4). Specific immunopre-cipitations were completed by incubating pre-cleared lysateswith specific antibodies bound to protein A-Sepharose beads(Sigma) for 2 h at 4�C. Proteins from the immunoprecipi-tates were separated on 12.5% SDS-PAGE gels.
2.11 Quantitation of membrane ruffling
Membrane ruffling was quantitated by 4 individuals, expertsin mast cell degranulation studies (were not involved with theexperiment) through blinded observation (theywere not awareof conditions under which the experiments were done). TheseDIC images of mast cells were captured (Confocal micro-scope, Olympus, 100X) after transfection with respectiveplasmid constructs. The membrane ruffles were scored withregards to the control/resting in WT showing 0 ruffling andcross-linked in WT being assigned a maximum score of 5.For each condition at least 10 fields from three independenttransfection experiments were examined.
2.12 Statistics
For all experiments statistical analysis has been done byStudent’s t-test in Microsoft excel. Here every data isexpressed as means ± SEM. The *, **, *** represent the p-values of B0.05, B0.005, B0.0005; respectively.
130 P Naskar, N Naqvi and N Puri

3. Results
3.1 Phosphorylation of SNAP-23 at Serine 120,during mast cell exocytosis is transient in nature
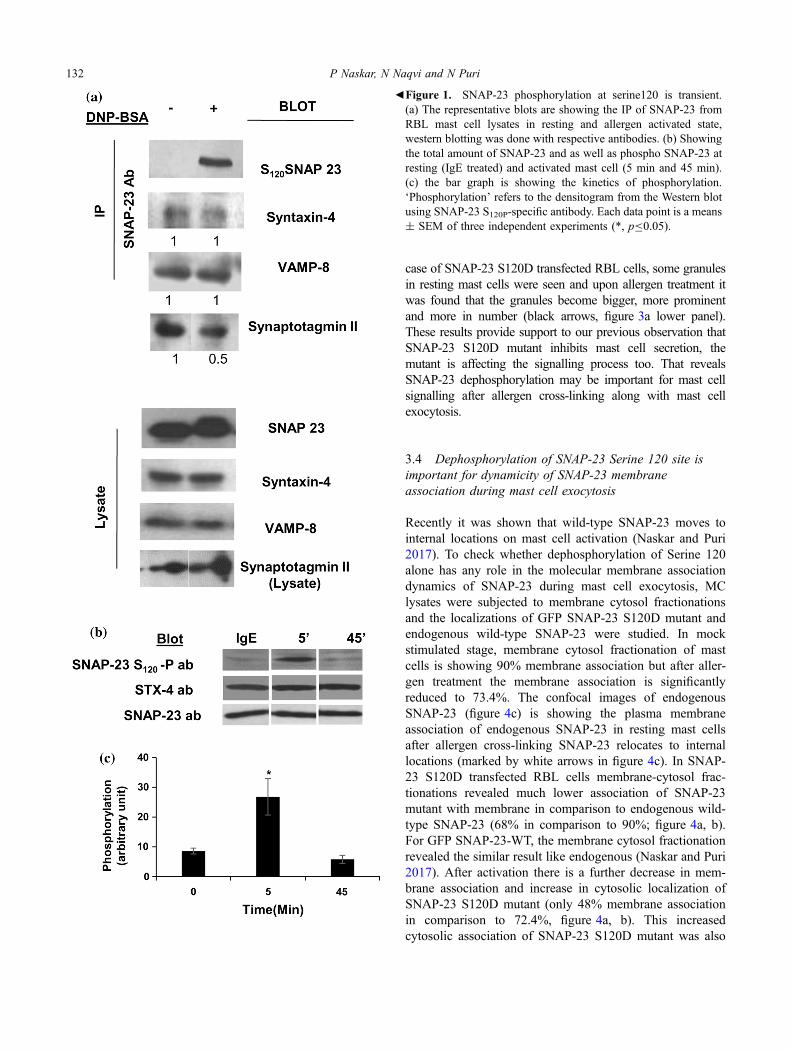
SNAP-23 has been shown to be important for mast cell exo-cytosis, the changes in phosphorylation of SNAP-23 and itsinteractions with other SNAREs, or interacting protein imme-diately after receptor cross-linking at 10 min have beeninvestigated. RBL mast cells were sensitized and receptorcross-linked with DNP-BSA for different time points as men-tioned (figure 1). As shown in the figure 1a, no band in the blotis seen corresponding to SNAP-23 S120-P ab in resting stage,but it appears 10 min after receptor cross-linking, while thetotal amount of SNAP-23, Syntaxin-4, VAMP-8 and, Synap-totagmin-II (Syt-II) remain the same (figure 1a). Co-immuno-precipitation (co-IP) studies by SNAP-23 specific antibodyindicate that Syntaxin-4 and VAMP-8 are bound to SNAP-23in resting and as well as activated mast cells and there is nochange in these interactions (figure 1a). Then co-immunopre-cipitation study was carried out between SNAP-23 and calciumsensor Syt-II to find out their direct interactions, as earlierreports showed that Syt-I interacts with SNAP-25 and playsimportant role in neurotransmitter release (Vogel et al. 2000).Our experiments revealed that in resting mast cells SNAP-23interacts with Syt-II. But this interaction is decreased in acti-vated stage by 50% (figure 1a). From this study we can con-clude that SNAP-23 phosphorylation is accompanied bydifferential interactions with other protein while their totalamounts remain the same during mast cell activation (Fig-ure 1a, lower panel). The extent of phosphorylation of SNAP-23 was also studied during early (5 min, highest kinetics ofmast cell secretion) and late (45 min, when the secretionreached a plateau) time points of mast cell exocytosis. RBLmast cells were sensitized and receptor cross-linked with DNP-BSA for different time points as mentioned. By using a SNAP-23 S120-Phospho specific antibody (SNAP-23 S120-P ab), itwas found that at the early time point, that is at 5 min, SNAP-23 got phosphorylated at S120, and after 45 min SNAP-23S120-Phospho band disappeared, while the total amount ofSNAP-23 and Syntaxin-4 remain the same (figure 1b and c).This confirms the transient nature of SNAP-23 phosphorylationat S120, which returns to basal non-phosphorylated state, afterinitial transient phosphorylation.
3.2 Blocking of dephosphorylation of SNAP-23at Serine 120 by S120D phosphomimetic mutationinhibits mast cell exocytosis
SNAP-23 transient phosphorylation at Serine 120 changeswith mast cell degranulation, from previous results. So, tocheck the effect of dephosphorylation on mast cell secretiondirectly, the serine (S) residue of SNAP-23 at 120 was
mutated to aspartate (D) by site directed mutagenesis, so thatthe new mutant mimics the phosphorylated phase of SNAP-23 at serine 120 site. The mutated SNAP-23 is cloned intoGFP vector as C-terminal protein for better visualization.This mutant was called the phosphomutant of SNAP-23 orSNAP-23 S120D, the sequencing was done to confirm thepoint mutation (figure 2a showing the sequence). Thismutant was transfected in RBL mast cells and after 24 h thecells were harvested by trypsinization and fixed with PFA.The transfection efficiency was measured by flow cytometry.Figure 2b is showing the transfection efficiency of SNAP-23S120D with respect to EGFP vector alone and GFP SNAP-23 wild-type. It was found that the transfection efficiency isreally less (only 16 ± 2%) in comparison to EGFP vectoralone (31.8 ± 3.9%) and wild-type SNAP-23 (33.3 ± 2%;figure 2b). The post transfection cell recovery was alsochecked that revealed, in case of GFP SNAP-23 S120D thecell recovery is 70% in comparison to wild-type transfectedcells (figure 2c). But the percent protein expression iscomparable to SNAP-23 WT (in comparison to endogenous,43% for SNAP-23 WT and for SNAP-23 S120D it is 37%;figure 2d and 2e). Then a secretion reporter assay was per-formed by using human Growth hormone (as described inmaterials and methods) to check the effect of this mutant onmast cell secretion. GFP SNAP-23 120D transfected mastcells showed a severe reduction in secretion (only 6.96 ±
4.3% in comparison to GFP SNAP-23 wild-type 33.37 ±
1.41%; figure 2f). So, it is clear that the constitutive phos-phorylation at SNAP-23 120 residue causes a massive, 79 %inhibition in mast cell exocytosis.
3.3 Transfection of SNAP-23 S120D mutant causesphenotypic changes in transfected mast cells indicativeof signalling defects
Recently studies have shown that secretion happens throughthe porosomal portal that is present between the ridges orruffles of the membrane and these ruffles are the hall mark forsecretion and further signalling (Jena 2009; Deng et al. 2010).Not only that, ruffling represents the cytoskeletal rearrangementthat also correlate with the downstream signalling after FceRIcross-linking with antigen (Deng et al. 2010; Draber et al.2012). To find out whether SNAP-23 120D has any effect inmast cell signalling after allergen cross-linking, the cellularmorphology was observed with respect to the prevalence ofruffles and granularity in SNAP-23 WT and SNAP-23 S120Dtransfected mast cells. The amount of ruffle formation wasanalysed (by blinded observer, n = 4) in GFP SNAP-23S120D transfected RBL, and was found significantly less incomparison to SNAP-23 wild-type transfected cells (2.8 incomparison to 4.16 respectively, figure 3a and b). In the wild-type SNAP-23 transfected cells no prominent granules wereseen (figure 3a) in resting and cross-linked mast cells. But in
SNAP-23 dephosphorylation in mast cell secretion 131
case of SNAP-23 S120D transfected RBL cells, some granulesin resting mast cells were seen and upon allergen treatment itwas found that the granules become bigger, more prominentand more in number (black arrows, figure 3a lower panel).These results provide support to our previous observation thatSNAP-23 S120D mutant inhibits mast cell secretion, themutant is affecting the signalling process too. That revealsSNAP-23 dephosphorylation may be important for mast cellsignalling after allergen cross-linking along with mast cellexocytosis.
3.4 Dephosphorylation of SNAP-23 Serine 120 site isimportant for dynamicity of SNAP-23 membraneassociation during mast cell exocytosis
Recently it was shown that wild-type SNAP-23 moves tointernal locations on mast cell activation (Naskar and Puri2017). To check whether dephosphorylation of Serine 120alone has any role in the molecular membrane associationdynamics of SNAP-23 during mast cell exocytosis, MClysates were subjected to membrane cytosol fractionationsand the localizations of GFP SNAP-23 S120D mutant andendogenous wild-type SNAP-23 were studied. In mockstimulated stage, membrane cytosol fractionation of mastcells is showing 90% membrane association but after aller-gen treatment the membrane association is significantlyreduced to 73.4%. The confocal images of endogenousSNAP-23 (figure 4c) is showing the plasma membraneassociation of endogenous SNAP-23 in resting mast cellsafter allergen cross-linking SNAP-23 relocates to internallocations (marked by white arrows in figure 4c). In SNAP-23 S120D transfected RBL cells membrane-cytosol frac-tionations revealed much lower association of SNAP-23mutant with membrane in comparison to endogenous wild-type SNAP-23 (68% in comparison to 90%; figure 4a, b).For GFP SNAP-23-WT, the membrane cytosol fractionationrevealed the similar result like endogenous (Naskar and Puri2017). After activation there is a further decrease in mem-brane association and increase in cytosolic localization ofSNAP-23 S120D mutant (only 48% membrane associationin comparison to 72.4%, figure 4a, b). This increasedcytosolic association of SNAP-23 S120D mutant was also
bFigure 1. SNAP-23 phosphorylation at serine120 is transient.(a) The representative blots are showing the IP of SNAP-23 fromRBL mast cell lysates in resting and allergen activated state,western blotting was done with respective antibodies. (b) Showingthe total amount of SNAP-23 and as well as phospho SNAP-23 atresting (IgE treated) and activated mast cell (5 min and 45 min).(c) the bar graph is showing the kinetics of phosphorylation.‘Phosphorylation’ refers to the densitogram from the Western blotusing SNAP-23 S120P-specific antibody. Each data point is a means± SEM of three independent experiments (*, pB0.05).
132 P Naskar, N Naqvi and N Puri

observed by confocal fluorescence microscopy. When thetransfected RBL mast cells were co-stained with LAMP-3ab, a very high co-localization of SNAP-23 S120D mutantwith LAMP-3 was observed (P. coefficient 0.54 ± 0.01;
figure 4d and e), in IgE sensitized mast cell, a large portionof mutant SNAP-23 was already found associated withinternal lysosomal membranes rather than with plasmamembrane as occurs in case of wild-type endogenous
Figure 2. Transfection and expression of SNAP-23 S120D causes inhibition of MC exocytosis. (a) Showing the Sequence of mutantSNAP-23-S120D in comparison to SNAP-23 wild-type. (b) the transfection efficiency of the SNAP-23-S120D mutant in comparison toEGFP-SNAP-23WT and EGFP vector alone. (c) the graph is demonstrating the post transfection cell recovery of the mutant. (d) the percentexpression of the phosphomutant is shown here by western blotting and quantitation of blots shown by the bar graph in (e). (f) SNAP-23S120D transfected mast cells are showing inhibition in mast cell secretion. Here the hGh release was studied that was found reduced around80% with respect to SNAP-23 wild-type. Each data point is a means ± SEM of three independent experiments (*, pB0.05; **, pB0.005).
SNAP-23 dephosphorylation in mast cell secretion 133

SNAP-23 and these are present more in cytosol as shown inmembrane cytosol quantification (figure 4b). Even afterreceptor cross-linking, the SNAP-23 S120D mutant showsvery high co-localization with LAMP-3 (P. coefficient0.53 ± 0.06; figure 4d and e). These results indicate that thedephosphorylation following transient induced phosphoryla-tion of SNAP-23 at Ser120 plays a very important role andalone can mediate the movement of SNAP-23 to internallocations and then its association with lysosomal membranes.
4. Discussion
Protein phosphorylation and dephosphorylation are rever-sible posttranslational modifications that play key role inseveral physiological processes and are often deregulated inpathological conditions. The balance between activation anddeactivation of signalling pathways is a delicate one, which
is regulated not only through phosphorylation by kinases,but also through dephosphorylation events induced by adiverse range of phosphatases. Phosphorylation acts as amolecular switch for many regulatory events in signallingpathways that drive cell division, proliferation, differentia-tion, apoptosis, and exocytosis. For example, the phospho-rylation and dephosphorylation of t-SNAREs (Sso and Sec9)regulate their interactions and the exocytosis process in yeast(Marash and Gerst 2001).
It is possible that phosphorylated and non-phosphorylatedforms of SNAP-23 bind to different SNAREs or SNAREadapter proteins or any other proteins involved in exocytosis. Inthe current study as a model system for activation of mast cellswe have used cross-linking of DNP-specific IgE sensitizedmast cells with the multivalent antigen dinitrophenyl-bovineserum albumin (DNP-BSA), exactly as on mast cells of atopicindividuals during an allergen challenge (Gould et al. 2003).Here it was found that S120 dephosphorylation is importantand was shown to play an important role in SNAP-23 reloca-tion and dynamicity and subsequently in mast cell exocytosis.SNAP-23 phosphorylation is transient in nature because after 5min of allergen cross-linking phosphorylation is there but after45 min it disappeared. With phosphorylation the interaction ofSNAP-23 with syt II is decreasing, but STX-4, and VAMP-8interaction remains constant even after 10 min of allergencross-linking. There are reports that phosphorylation affects thebinding of affinity of different SNARE proteins like SNAP-25and Syntaxin-4 and even with Syt also. Phosphorylation of Sytregulates its interaction with t-SNAREs in neuronal cells(Verona et al. 2000). According to a model that explains themechanism of Syt functions in neurotransmitter release, SNAP-25, Syntaxin and VAMP form a loose tri molecular complexthat zips up half way, and is not enough for membrane fusion tohappen. This can convert to a tight SNARE-complex only afterSyt-I association upon Ca2? trigger. To explain the innermolecular mechanism one model suggests that without Ca2?,Syt prevents the vesicle fusion by binding with t-SNARE i.e.,blocking of the fusion pore formation. But upon Ca2? sig-nalling, Syt releases the t-SNAREs and subsequently they canmediate the fusion process (Chapman 2002; Snyder et al.2006). But for mast cells, the story is slightly different. Baramet al. (Baram et al. 1999) suggested that Syt-II inhibits granule-granule fusion during mast cell exocytosis upon calcium sig-nalling. On the other hand, it was found that syntaxin-4phosphorylation enhances its interaction with Syt I and phos-phorylation of Syntaxin-4 reduces its interaction with SNAP-25, during neurotransmitter release (Risinger and Bennett1999). Therefore, like these findings, it is possible that SNAP-23 phosphorylation may be down regulating its interaction withSyt-II to facilitate the granule-granule fusion during mast cellexocytosis.
A massive decrease was seen in mast cell exocytosis inSNAP-23 S120D transfected mast cells, which may be dueto the nature of this mutant. As this mutant is
Figure 3. SNAP-23 S120D affects the mast cell signallingfollowing the allergen cross-linking. (a) The DIC images fromSNAP-23WT and SNAP-23120D transfected mast cell (in cross-linked state) showing the difference in membrane ruffling andgranularity. Scale bar represents 10 lm. Membrane ruffling wasquantitated through blinded observers as explained in materials andmethods. For each condition at least 10 fields from threeindependent transfection experiments were examined. (b) Thetable shows reduction in membrane ruffles-formation in case ofSNAP-23 phosphomutant transfected mast cell in comparison towild-type upon allergen cross-linking. Each data point is a means ±SEM of three independent experiments (**, pB0.005).
134 P Naskar, N Naqvi and N Puri
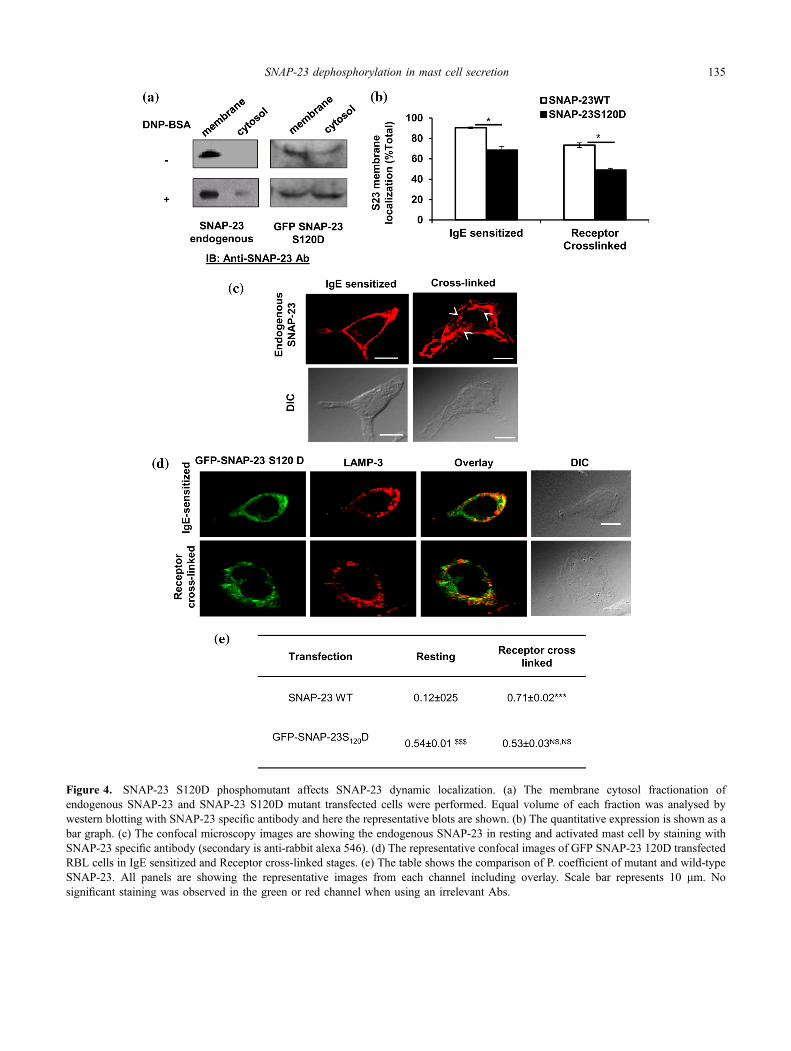
Figure 4. SNAP-23 S120D phosphomutant affects SNAP-23 dynamic localization. (a) The membrane cytosol fractionation ofendogenous SNAP-23 and SNAP-23 S120D mutant transfected cells were performed. Equal volume of each fraction was analysed bywestern blotting with SNAP-23 specific antibody and here the representative blots are shown. (b) The quantitative expression is shown as abar graph. (c) The confocal microscopy images are showing the endogenous SNAP-23 in resting and activated mast cell by staining withSNAP-23 specific antibody (secondary is anti-rabbit alexa 546). (d) The representative confocal images of GFP SNAP-23 120D transfectedRBL cells in IgE sensitized and Receptor cross-linked stages. (e) The table shows the comparison of P. coefficient of mutant and wild-typeSNAP-23. All panels are showing the representative images from each channel including overlay. Scale bar represents 10 lm. Nosignificant staining was observed in the green or red channel when using an irrelevant Abs.
SNAP-23 dephosphorylation in mast cell secretion 135
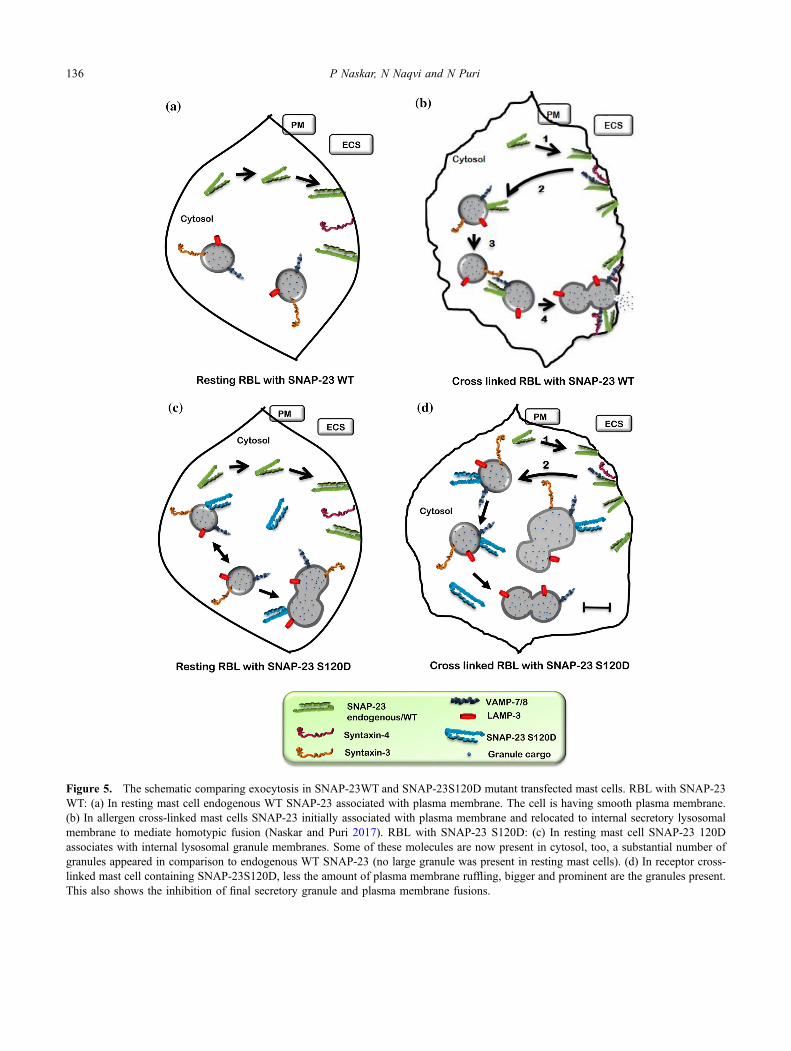
Figure 5. The schematic comparing exocytosis in SNAP-23WT and SNAP-23S120D mutant transfected mast cells. RBL with SNAP-23WT: (a) In resting mast cell endogenous WT SNAP-23 associated with plasma membrane. The cell is having smooth plasma membrane.(b) In allergen cross-linked mast cells SNAP-23 initially associated with plasma membrane and relocated to internal secretory lysosomalmembrane to mediate homotypic fusion (Naskar and Puri 2017). RBL with SNAP-23 S120D: (c) In resting mast cell SNAP-23 120Dassociates with internal lysosomal granule membranes. Some of these molecules are now present in cytosol, too, a substantial number ofgranules appeared in comparison to endogenous WT SNAP-23 (no large granule was present in resting mast cells). (d) In receptor cross-linked mast cell containing SNAP-23S120D, less the amount of plasma membrane ruffling, bigger and prominent are the granules present.This also shows the inhibition of final secretory granule and plasma membrane fusions.
136 P Naskar, N Naqvi and N Puri

phosphomimetic, and cannot be dephosphorylated so thisshows that the dephosphorylation is necessary for normalcellular functions to occur. There are reports that differentSNAREs including SNAP-23, Syntaxin-4 are associated inthe interaction with cytoskeleton during platelet secretion(Woronowicz et al. 2010). In mast cell it has been shownthat cytoskeleton rearrangement is associated with themembrane ruffling and other signalling processes (Denget al. 2010). From our data, we found reduced amount ofmembrane ruffling during mast cell secretion in SNAP-23S120D transfected cells. So, the freezing of phosphorylationat S120 site is inhibiting mast cell exocytosis not only byaffecting the membrane fusions but also the other signallingprocesses. As there is less exocytosis, there is no wonder wefound more granules in these cells that are big in size andmore prominent that may indicate that the granule–granulefusion is not affected but the final granule-plasma membranefusion is not occurring. So, it can be speculated that may bedephosphorylation of SNAP-23 is necessary for the finalplasma membrane fusion to occur.
Since the induced phosphorylation at S120 is in closeproximity to five cysteine residues in linker region in SNAP-23 involved in SNAP-23 palmitoylation (Vogel and Roche1999), phosphorylation may be important for determiningthe localization of SNAP-23 on different intracellularmembranes. To study the localization of SNAP-23 S120Dwe performed confocal study and membrane-cytosol frac-tionation. Following the defect in signalling we found themutant is accumulating more in cytosol in resting and cross-linked mast cells (figure 4 a and b). We could see that thereare membrane association of the mutant is only *40-60 %(from WB) and under confocal microscope we found veryhigh P. coefficient in resting and cross-linked mast cells.Although SNAP-23 S120D is now colocalized with LAMP-3 containing lysosomes but exocytosis is inhibited. Thatobservation can be explained along with our previousobservations including, SNAP-23 phosphorylation is tran-sient and freezing of phosphorylated state of SNAP-23 maystuck SNAP-23 with the SG membranes and prevent therecycling of SNAREs and SNARE priming (Puri et al.2003). So, in spite of showing high colocalization withinternal lysosomal SGs the SNAP-23 S120D mutant nega-tively regulates the mast cell exocytosis. Together phos-phorylation may be important for granule granule fusion incompound exocytosis but dephosphorylation is important forSNARE priming/recycling for granule plasma membranefusion to occur.
We tried to make a schematic model (figure 5) showingthe effect of SNAP-23 120D dephosphorylation in restingstate and after allergen cross-linking (figure 5c and d). Thefirst panel of figure 5a shows SNAP-23 WT expressing mastcell at resting condition. Upon allergen cross-linking itshows membrane ruffling and SNAP-23 relocation to theinternal lysosomal membranes to mediate homotypic fusions
during compound exocytosis (figure 5b; Naskar and Puri2017). But in case of SNAP-23 S120D mutant transfections,mast cells in resting state seem more granular probably dueto promiscuous fusions of granules due to association ofSNAP-23 S120D with internal lysosomal membranes (fig-ure 5c). In allergen cross-linked state, SNAP-23 S120D stillassociates with internal lysosomal (LAMP-3 containing)granule membranes, and with cytosol as well (figure 5d). Asmutant is unable to dephosphorylate, so, it cannot dissociatefrom the internal membrane and the dynamicity of SNAP-23membrane association has been lost. Further there is severehindrance in mast cell mediator release in activated mast cell(as shown by our data, figure 2f) and as well as inhibition insignalling process also there, as the mutant transfected cellswere showing less amount of membrane ruffling as com-pared to endogenous control (figure 3a and b). So, here inthis model, figure 5d represents a SNAP-23 S120D trans-fected mast cell where fewer granule-plasma membranefusions are occurring, and the cell therefore contains biggergranules that may accumulate due to the inhibition of exo-cytosis. Further elucidation of this pathway of SNAP-23trafficking would help to generate a comprehensive view ofthe complex membrane fusion processes that occur in mastcells during exocytosis. Elucidating the role of SNAP-23dephosphorylation, in addition to studying the cell biologyof SNAREs, will generate a more comprehensive view of thecomplex trafficking processes that occur in mast cells.SNAREs themselves, or other components of membranefusion, might prove to be novel targets for therapeuticintervention in adverse inflammatory or allergic diseases.
Acknowledgements
We are grateful to Dr. Paul A Roche (ExperimentalImmunology Branch, National Cancer Institute, NationalInstitutes of Health, Bethesda, MD) for providing us withRBL cell line, SNAP-23 c-terminal antibody and pCMV-FLAG-RAT SNAP-23 plasmid and pCDNA3-hGH plasmid.We also thank confocal fluorescent Microscopy Facility,AIRF (Advanced instrument research facility), JNU for helpwith image analysis. This work was supported by researchgrants from Department of Science and Technology (DST)Govt. of India (SR/SO/HS-0122/2009; and DST-PURSE),and University Grants Commission (UGC), India (UPE-IIProject ID-54; and UGC-resource networking) to NP. PNand NN were supported by a grant from UGC, India.
References
Baram D, Adachi R, Medalia O, Tuvim M, Dickey BF, Mekori YAand Sagi-Eisenberg R 1999 Synaptotagmin II negatively
SNAP-23 dephosphorylation in mast cell secretion 137

regulates Ca2?-triggered exocytosis of lysosomes in mast cells.J. Exp. Med. 189 1649–1658
Blank U, Cyprien B, Martin-Verdeaux S, Paumet F, Pombo I,Rivera J, Roa M, and Varin-Blank N 2002 SNAREs andassociated regulators in the control of exocytosis in the RBL-2H3 mast cell line. Mol. Immunol. 38 1341-1345
Chapman ER 2002 Synaptotagmin: a Ca(2?) sensor that triggersexocytosis? Nat. Rev. Mol. Cell Biol. 3 498–508
Deng Z, Lulevich V, Liu FT and Liu GY 2010 Applications ofatomic force microscopy in biophysical chemistry of cells. J.Phys. Chem. B. 114 5971–5982
Draber P, Sulimenko Vand Draberova E 2012 Cytoskeleton in mastcell signalling. Front. Immunol. 3 130
Galli S J, Grimbaldeston M and Tsai M 2008 Immunomodulatorymast cells: negative, as well as positive, regulators of immunity.Nat. Rev. Immunol. 8 478–486
Gould H J, Sutton BJ, Beavil AJ, Beavil RL, McCloskey N, CokerHA, Fear D and Smurthwaite L 2003 The biology of IGE andthe basis of allergic disease. Annu. Rev. Immunol. 21 579–628
Hepp R, Puri N, Hohenstein AC, Crawford GL, Whiteheart SW andRoche PA 2005 Phosphorylation of SNAP-23 regulates exocy-tosis from mast cells. J. Biol. Chem. 280 6610–6620
Huber M, Helgason CD, Damen JE, Liu L, Humphries RK andKrystal G 1998 The src homology 2-containing inositolphosphatase (SHIP) is the gatekeeper of mast cell degranulation.Proc. Natl. Acad. Sci. U S A 95 11330-11335
Jahn R and Sudhof TC 1999 Membrane fusion and exocytosis.Annu. Rev. Biochem. 68 863–911
Jena BP 2009 Porosome: the secretory portal in cells. Biochemistry.48 4009–18
Kalesnikoff J and Galli SJ 2008 New developments in mast cellbiology. Nat. Immunol. 9 1215–1223
Karim Z A, Zhang J, Banerjee M, Chicka MC, Al Hawas R,Hamilton TR, Roche PA and Whiteheart SW 2013 IkappaBkinase phosphorylation of SNAP-23 controls platelet secretion.Blood 121 4567–4574
Kraft S and Kinet JP 2007 New developments in FcepsilonRIregulation, function and inhibition. Nat. Rev. Immunol. 7365–378
Lin RC and Scheller RH 2000 Mechanisms of synaptic vesicleexocytosis. Annu. Rev. Cell Dev. Biol. 16 19–49
Marash M and Gerst JE 2001 t-SNARE dephosphorylationpromotes SNARE assembly and exocytosis in yeast. EMBO. J.20 411-421
Lorentz A, Baumann A, Vitte J and Blank U 2012 The SNAREmachinery in mast cell secretion. Front. Immunol. 3 143
Naskar P and Puri N 2017 Phosphorylation of SNAP-23 regulatesits dynamic membrane association during mast cell exocytosis.Biol. Open 6 1257–1269
Poirier MA, Xiao W, Macosko JC, Chan C, Shin YK and BennettMK 1998 The synaptic SNARE complex is a parallel four-stranded helical bundle. Nat. Struct. Biol. 5 765–769
Puri N, Kruhlak MJ, Whiteheart SW and Roche PA 2003 Mast celldegranulation requires N-ethylmaleimide-sensitive factor-medi-ated SNARE disassembly. J. Immunol. 171 5345–5352
Puri N and Roche PA 2008 Mast cells possess distinct secretorygranule subsets whose exocytosis is regulated by differentSNARE isoforms. Proc. Natl. Acad. Sci. USA 105 2580–2585
Risinger C and Bennett MK 1999 Differential phosphorylation ofsyntaxin and synaptosome-associated protein of 25 kDa (SNAP-25) isoforms. J. Neurochem. 72 614–624
Snyder DA, Kelly ML and Woodbury DJ 2006 SNARE complexregulation by phosphorylation. Cell. Biochem. Biophys. 45 111–123
Suzuki K and Verma IM 2008 Phosphorylation of SNAP-23 byIkappaB kinase 2 regulates mast cell degranulation. Cell 134485–495
Urb M and Sheppard DC 2012 The role of mast cells in the defenceagainst pathogens. PLoS Pathog. 8 e1002619
Vaidyanathan V V, Puri N and Roche PA 2001 The last exon ofSNAP-23 regulates granule exocytosis from mast cells. J. Biol.Chem. 276 25101–25106
Verona M, Zanotti S, Schafer T, Racagni G and Popoli M 2000Changes of synaptotagmin interaction with t-SNARE proteinsin vitro after calcium/calmodulin-dependent phosphorylation. J.Neurochem. 74 209–221
Vogel K, Cabaniols JP and Roche PA 2000 Targeting of SNAP-25to membranes is mediated by its association with the targetSNARE syntaxin. J. Biol. Chem. 275 2959–2965
Vogel K and Roche PA 1999 SNAP-23 and SNAP-25 arepalmitoylated in vivo. Biochem. Biophys. Res. Commun. 258407–410
Weber T, Zemelman BV, McNew JA, Westermann B, Gmachl M,Parlati F, Sollner TH and Rothman JE 1998 SNAREpins:minimal machinery for membrane fusion. Cell 92 759–772
Woronowicz K, Dilks JR, Rozenvayn N, Dowal L, Blair PS, PetersCG, Woronowicz L and Flaumenhaft R 2010 The platelet actincytoskeleton associates with SNAREs and participates in alpha-granule secretion. Biochemistry 49 4533–4542
Corresponding editor: RAJIV K SAXENA
138 P Naskar, N Naqvi and N Puri





























