Embed Size (px)
Citation preview

JOURNAL OF BACTERIOLOGY, Nov. 1981, p. 584-5930021-9193/81/110584-10$02.00/0
Vol. 148, No. 2
Biosynthetic and Bioenergetic Functions of Citric Acid CycleReactions in Rhodopseudomonas capsulata
J. THOMAS BEATTYt AND HOWARD GEST*Photosynthetic Bacteria Group, Department of Biology, Indiana University, Bloomington, Indiana 47405
Received 18 May 1981/Accepted 1 July 1981
Rhodopseudomonas capsulata can grow in a number of altemative modes,including (i) photosynthetic, defined here as anaerobic growth with light as theenergy source, and (ii) heterotrophic, referring to aerobic heterotrophic growth indarkness. The functions of citric acid cycle sequences in these growth modes wereinvestigated using wild-type and appropriate mutant strains. Results of growthtests and 02 utilization experiments showed that in the heterotrophic mode,energy conversion is dependent on operation of the classical citric acid cycle.Alpha-ketoglutarate dehydrogenase (KGD) activity in wild-type strain B10 issubstantially higher in cells grown heterotrophically than in cells grown photo-synthetically. Molecular oxygen, even at low concentration, appears to be impor-tant in regulation of KGD synthesis and, thus, in expression of citric acid cycleactivity. Extracts of (photosynthetically grown) mutant strain KGD11 lack de-monstrable KGD activity, and in contrast to the wild type, KGD11 is unable togrow heterotrophically on succinate, malate, or pyruvate owing to failure of theenergy conversion function of the citric acid cycle. KGD11, however, grows wellphotosynthetically on malate or on CO2 + H2. The KGD activity level requiredto support the bioenergetic function of the citric acid cycle is evidently muchhigher than that necessary to satisfy biosynthetic demands; thus, a very low rateof succinyl-coenzyme A formation (needed for biosynthesis) in the mutant wouldsuffice for growth under photosynthetic conditions. In wild-type R. capsulata,the a-ketoglutarate required for glutamate synthesis is ordinarily generated viacitric acid cycle reactions, which include the conversions catalyzed by citratesynthase and isocitrate dehydrogenase. Mutants blocked in the former or both ofthese enzymes can grow photosynthetically if provided with an exogenous sourceof a-ketoglutarate or glutamate, but grow very poorly (if at all) as heterotrophssince the energy supply under these conditions depends on operation of thecomplete citric acid cycle.
The citric acid cycle effects the catabolism ofacetyl-coenzyme A (CoA) to provide the reduc-ing power that drives the oxidative phosphoryl-ation machinery ofaerobic cells. Citric acid cyclereaction sequences, however, are also importantfor providing biosynthetic intermnediates in most,if not all, types of cells. Several cycle interme-diates (a-ketoglutarate [KG], oxaloacetate, andsuccinyl-CoA) are used for net synthesis of var-ious1essential cell constituents. In certain kindsof bacteria, segments of the cycle function onlyfor biosynthetic purposes, for example, in strictanaerobes such as Clostridium kluyveri (35) andin so-called "obligate" chemoautotrophs (32). Infacultative heterotrophic aerobes-anaerobes, therelative commitments of citric acid cycle se-quences to biosynthetic and bioenergetic func-
t Present address: Department of Genetics, Stanford Uni-versity School of Medicine, Stanford, CA 94305.
tions may be markedly affected by nutritionalcircumstances. Thus, when Escherichia coli isgrown fermentatively on sugars, the cells lackKG dehydrogenase (KGD), and under theseconditions, succinate required for fornation ofthe biosynthetic precursor succinyl-CoA is pro-duced reductively (2) by reversal of the se-quence:
-2H -2Hsuccinate -+ fumarate -. malate -+ oxaloacetateThis illustrates the special importance of KGDas a "regulatory catalyst" in overall operation ofthe citric acid cycle.
Little is known of the controls that govern"opening" (for biosynthesis) and "closing" (forenergy conversion) of the citric acid cycle, andthese are of particular significance for organisthat can grow either as anaerobes or aerobes.The photosynthetic bacterium Rhodopseudo-
584
on Septem
ber 8, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

CITRIC ACID CYCLE IN PHOTOSYNTHETIC BACTERIA 585
monas capsulata presents unusual opportu-nities for investigation of this and related prob-lems because it is capable of growing in fivedistinctly different ways (24): (i) anaerobicallyas a photoautotroph on H2 plus C02 with lightas the energy source, (ii) anaerobically as aphotoheterotroph on various organic carbonsources with light as the energy source, (iii) as afermentative anaerobe in darkness on sugars assole carbon and energy sources, (iv) aerobicallyas an ordinary chemoheterotroph in darkness,and (v) aerobically as a chemoautotroph in dark-ness with H2 as the source of energy and reduc-ing power. In this communication, we presentobservations on operation of the citric acid cycleand component sequences in R. capsulatamainly in connection with two particularlyprominent growth modes, namely, photoheter-otrophic (anaerobic) and aerobic (dark) chemo-heterotrophic.
MATERIALS AND METHODSBacterial strains. The general characteristics of
R. capsulata wild-type strain B10 are typical for thespecies (36). Mutant WE29 is a derivative of B10, andlacks both citrate synthase (CS) and isocitrate dehy-drogenase (ICD) activities (7). Although this strainwas originally thought to be a single-site mutant, moredetailed genetic analysis (J. T. Beatty, M.A. thesis,Indiana University, Bloomington, 1978) has shownthat the enzymatic deficiencies noted result from twoseparate mutations; similar mutants of E. coli havebeen described by others (19). Mutant T500, deficientin CS activity, was derived from B10 as detailed below.Strain KGD11 was also obtained from B10, after ethylmethane sulfonate mutagenesis, and is virtually devoidof KGD activity (6a)
Culture media and growth conditions. Unlessotherwise noted, the bacteria were grown in a minimalsynthetic medium, designated RCVB, which contains30mM DL-malate and 7.5mM (NH4)2S04 as the C andN sources, respectively (6a). When organic carbonsources other than malate were used, these and thepotassium phosphate component were added asepti-cally to the remainder of the sterile medium. In allinstances, organic carbon sources were added so as toprovide an initial C concentration equivalent to thatof 30 mM malate.
For photoautotrophic growth, RCVB medium wasmodified by omission of malate and increase of phos-phate buffer concentration from 10 mM to 20 mM(note that phosphate concentration was also doubledfor cultures grown photoheterotrophically on succi-nate and for cultures grown aerobically in darkness).Erlenmeyer flasks of 1-liter capacity, containing 50 mlof inoculated medium under a gas phase of 5% CO2 inH2, were sealed with rubber stoppers.
For routine maintenance, stock cultures were grownphotosynthetically in completely filled 17-ml screw-cap tubes: strain B10 in RCVB medium, mutantsWE29 and T500 in RCVB medium supplemented with10 mM L-glutamate, and strain KGD11 in RCVB
medium modified to contain succinate in place ofmalate. Media used for growth of mutants were sup-plemented with 0.05% L-ascorbate, freshly made andadded at the time of culture inoculation. Cultures forenzyme assays were grown photosynthetically in fullscrew-cap tubes or square "milk dilution" bottles of165-ml capacity.
Photosynthetically grown cultures were illuminated(at 32°C) with ca. 600 ft-c (ca. 6,400 lx) intensity ofincandescent light, provided by Lumiline lamps. Foraerobic dark growth, Erlenmeyer flasks containing10% of their nominal capacity of growth medium wereincubated (32°C) on a rotary shaker at 250 rpm.
Fluoroacetate selection for CS mutants. Theprocedure for selection of CS mutants (such as T500)was devised to obtain strains with the same phenotypeas WE29 and was based on the observation thatgrowth of the latter mutant is resistant to fluoroace-tate. After ethyl methane sulfonate mutagenesis (26)of a B10 culture, the cells were grown photosyntheti-cally (to the stationary phase) in RCVB medium sup-plemented with 10 mM L-glutamate and 100 uM fluo-roacetate. After three consecutive subcultures in thesame medium, plates of solid RCVB medium (1.5%[wt/vol] Difco agar) containing 65 JIM L-glutamatewere spread with culture dilutions. Potential mutantsof interest were identified as small colonies after in-cubation of plates in GasPak anaerobic jars (BBLMicrobiology Systems, Cockeysville, Md.) under pho-tosynthetic conditions for 3 days. Putative mutantswere picked and tested for photosynthetic growth onRCVB medium and on RCVB medium supplementedwith either 10mM L-glutamate or 10mM KG. Isolatesthat grew in the presence of L-glutamate or KG, butnot on unsupplemented DL-malate plates, were puri-fied by streaking on RCVB plates containing 10 mML-glutamate (plates were incubated photosyntheti-cally). A similar procedure was independently devisedand used for isolation of CS mutants of AcinetobacterIwoffi by Harford and Weitzman (14).
Photometric measurement of cell concentra-tions. Culture densities were measured with a Klett-Summerson photometer fitted with a no. 66 filter. Thephotometer was modified so as to accept 17-ml-capac-ity screw-cap culture tubes (completely filled for ex-periments in which anaerobic [photosynthetic] growthwas followed). For aerobic (dark) growth kinetics, theErlenmeyer flasks used were fitted with side-arm tubesto facilitate photometer measurements.
Preparation of cell extracts. Extracts were ordi-narily made of cells grown to the late logarithmicphase (in a few instances, stationary-phase cultures[density, >350 photometer units] were used).
For experiments concerned with assay of CS, acon-itate hydratase, and ICD, cells were harvested bycentrifugation, washed, and resuspended in 50 mMTris-hydrochloride buffer (pH 7.5) containing 2 mMMnCl2 and 0.1 mM sodium isocitrate. (The lattersupplements stabilize ICD activity.) Cell suspensionswere disrupted by passage through a French pressurecell (operated at 16,000 to 20,000 lb/in2). The crudeextracts were clarified by centrifugation at 140,000 xg for 90 min, and the supernatant fluids were retainedfor enzyme assays.When KGD activity or 02 uptake was to be mea-
VOL. 148, 1981
on Septem
ber 8, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

586 BEATTY AND GEST
sured, cells were washed and resuspended in 50 mMpotassium phosphate buffer (pH 7.0). Suspensions of5 ml or greater in volume were disrupted with a Frenchpressure cell as described above; with smaller volumes,extracts were made by sonication as detailed by Beattyand Gest (6a). The crude extracts were not centrifuged,and enzymatic activities were assayed immediately.Essentially the same activity levels were found withboth cell breakage procedures.Enzyme assays and measurement of 02 up-
take. CS (EC 4.1.3.7) activity was determined bycolorimetric measurement of oxaloacetate-dependentreduction of 5,5'-dithiobis-(2-nitrobenzoic acid) by re-
duced CoA (8). ICD (EC 1.1.1.42) was assayed bymeasurement of DL-isocitrate-dependent reduction ofNADP+ (7). The activity of KGD (EC 1.2.4.2) wasdetermined by measurement of KG-dependent reduc-tion ofNAD+ in the presence of 10-5 M rotenone (6a).Aconitate hydratase (EC 4.2.1.3) activity was esti-mated by measurement of increase in absorbance at250 nm due to formation of cis-aconitate from isocit-rate (31).
Uptake of 02 was measured polarographically witha Clark electrode. The latter was polarized at -0.8 Vand connected to a chart recorder on which 1.0 mVregistered as the maximum deflection. The glass re-
action chamber contained 3.0 ml of reaction mixtureand a small magnetic "flea" and was closed off fromthe atmosphere by means of a Teflon stopper whichhad a 0.75-mm hole drilled through it. The hole per-mitted additions to be made to the reaction mixturethrough a syringe needle, while the concentration of02 dissolved in the medium was continuously mom-
tored. The entire assembly was water-jacketed andmaintained at 30°C over a magnetic stirrer.
It was observed that intact cells showed little or nosubstrate-dependent 02 uptake when suspended inphosphate buffer. When cells were suspended inRCVB medium from which DL-malate had been omit-ted, however, significant respiratory activity was ob-served upon addition of substrates. Consequently, cells
were suspended in this way, and 02 utilization was
measured with 40 mM DL-malate as the substrate.The concentration of 02 dissolved in air-saturatedRCVB minus malate medium at 30°C was found to beca. 253 nmol ml' (estimated from oxidation of knownamount,s of NADH by cell extracts). This value wasused for all assays, regardless of atmospheric pressure.Oxygen uptake with 5 mM NADH as substrate(NADH oxidase activity) was measured using crudeunfractionated extracts.The quantities of extracts or cell suspensions used
for assays were in the range where activity was pro-
portional to amount of preparation used.Other procedures. Light intensity was measured
with a Weston illumination meter (model 756), cali-brated in foot-candles. Gases were mixed in volume-to-volume ratios with a Matheson rotameter, usingcommercially available (Matheson) calibration curves
of flow rate versus milliliters of gas per minute. Thebacteriochlorophyll (BChl) content of intact cells wasdetermined as described by Sojka et al. (33), andprotein was estimated by the metliod of Lowry et al.(22) with crystalline bovine serum albumin as thestandard.
J. BACTERIOL.
RESULTS
Citric acid cycle activity in R. capsulataas measured by 02 consumption. The poten-tial capacity of R. capsulata cells to catalyzeaerobic (dark) citric acid cycle activity was as-
sessed from 02 utilization experimepts. Table 1shows the results of measurements using cells(or derived extracts) grown photosynthetically(anaerobically) or aerobically in darkness withmalate as the carbon source (RCVB medium).Cells grown aerobically, and extracts therefrom,showed activities ca. two- to threefold higherthan those grown photosynthetically. This dif-ference presumably indicates elevated synthesisof the respiratory system when growing cellsobtain energy by oxidative phosphorylation as-
sociated with dark aerobic electron flow.To validate the assumption that aerobic oxi-
dation of malate (and related substrates) by R.capsulata occurs via the citric acid cycle, exper-iments were performed with appropriate inhibi-tors and with two mutants blocked at differentsteps of the cycle. The inhibitors tested were
0uoroacetate, which gives rise to fluorocitrate,a potent inhibitor of aconitase (27), and arsenite,which inhibits ketoacid dehydrogenase com-
plexes through reaction with their lipoic acidmoieties (10). Both inhibitors caused strikinginhibition of malate-dependent 02 utilization byintact cells (Fig. 1). Depression of respiratoryrate to 50% of normal occurred with ca. 0.5 uMfluoroacetate or 0.5 mM arsenite. The lattercompound at 1 mM concentration completelyinhibits KGD activity in crude extracts of R.capsulata (6a). These results support the con-clusion that the citric acid cycle is the majormechanism used by R. capsulata for the aerobicoxidation of malate. Further eviconce was ob-tained by using mutants KGD11 (blocked inKGD activity; 6a) and T500 (blocked in CS).Cultures of the mutants were grown photosyn-thetically in malate medium supplemented witheither 5 mM succinate (KGD11) or 5 mM L-glutamate (T500). Cells of T500 and KGD1lwere found to consume 02 with malate as sub-
TABLE 1. UJtilization of O2 by intact cells andextracts ofR. capsulata B10
Rate of 02 coIsumptionaGrowth conditions (nmol.min-'.nmg protein-')
Intact cells Extracts
Aerobic/dark 104 ± 11 82 ± 25Anaerobic/light 38 ± 11 40 ± 12
a In darkness. Mean values and standard deviatio-nsare given for replicate experiments (four experimentswith intact cells and seven with extrabts). Substrateswere DL-malate for intact cells andNAPH for extracts.
on Septem
ber 8, 2018 by guesthttp://jb.asm
.org/D
ownloaded from
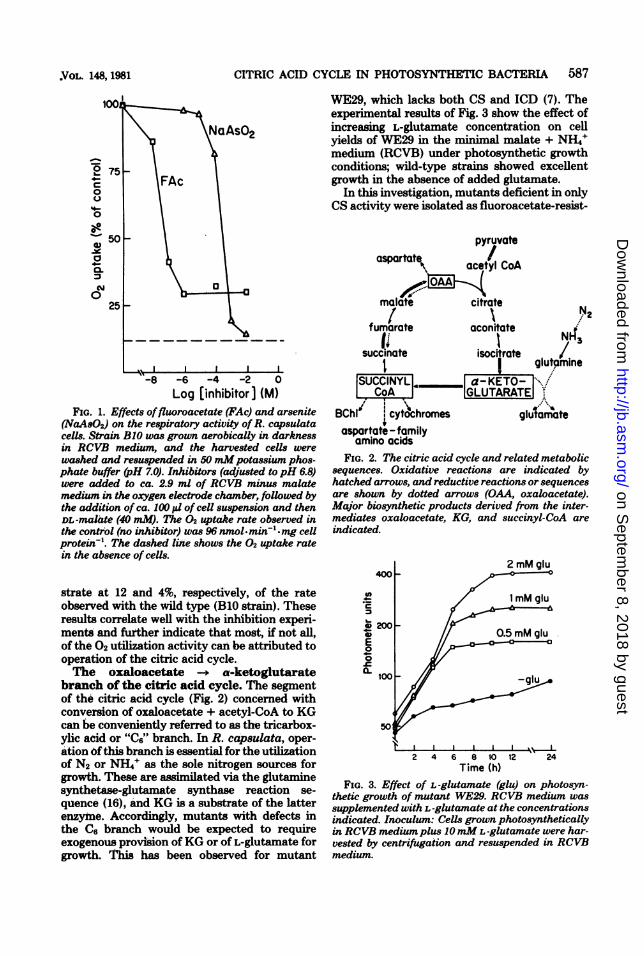
CITRIC ACID CYCLE IN PHOTOSYNTHETIC BACTERIA 587
° 75 -
C ~FAc0
0
850 l0I-
0
25 -
I I I.
-8 -6 -4 -2 0Log (inhibitor] (M)
FIG. 1. Effects offluoroacetate (FAc) and arsenite(NaAsO2) on the respiratory activity of R. capsulatacells. Strain BI1 was grown aerobically in darknessin RCVB medium, and the harvested cells were
washed and resuspended in 50 mMpotassium phos-phate buffer (pH 7.0). Inhibitors (adjusted to pH 6.8)were added to ca. 2.9 ml of RCVB minus malatemedium in the oxygen electrode chamber, foUowed bythe addition of ca. 100( p1 of cell suspension and thenDL-malate (40 mM). The 02 uptake rate observed inthe control (no inhibitor) was 96 nmol.min' *mg cellprotein-'. The dashed line shows the 02 uptake ratein the absence of cells.
strate at 12 and 4%, respectively, of the rateobserved with the wild type (BlO strain). Theseresults correlate well with the inhibition experi-ments and further indicate that most, if not all,of the 02 utilization activity can be attributed tooperation of the citric acid cycle.The oxaloacetate -- a-ketoglutarate
branch of the citric acid cycle. The segmentof the citric acid cycle (Fig. 2) concerned withconversion of oxaloacetate + acetyl-CoA to KGcan be conveniently referred to as the tricarbox-ylic acid or "C6" branch. In R. capsulata, oper-ation ofthis branch is essential for the utilizationof N2 or NH4+ as the sole nitrogen sources forgrowth. These are assimilated via the glutaminesynthetase-glutamate synthase reaction se-quence (16), and KG is a substrate of the latterenzyme. Accordingly, mutants with defects inthe C6 branch would be expected to requireexogenous provision ofKG or of L-glutamate forgrowth. This has been observed for mutant
WE29, which lacks both CS and ICD (7). Theexperimental results of Fig. 3 show the effect ofincreasing L-glutamate concentration on cellyields of WE29 in the minimal malate + NH4+medium (RCVB) under photosynthetic growthconditions; wild-type strains showed excellentgrowth in the absence of added glutamate.
In this investigation, mutants deficient in onlyCS activity were isolated as fluoroacetate-resist-
pyruvateasparte, Caeo acetyl CoA
malate citrate N2/fumarate aconitate
succinate isocitrate /tI glutomine
BChl cytochromes glulamateaspartate -fmily
amino acidsFIG. 2. The citric acid cycle and related metabolic
sequences. Oxidative reactions are indicated byhatched arrows, and reductive reactions or sequencesare shown by dotted arrows (OAA, oxaloacetate).Major biosynthetic products derived from the inter-mediates oxaloacetate, KG, and succinyl-CoA areindicated.
2 mM glu400
ImM glu
0.5 mM gluE00
100 ~~~~~-glu
50
2 4 6 8 10 12 24Time (h)
FIG. 3. Effect of L-glutamate (glu) on photosyn-thetic growth of mutant WE29. RCVB medium wassupplemented with L-glutamate at the concentrationsindicated. Inoculum: Cells grown photosyntheticallyin RCVB medium plus 1OmM L-glutamate were har-vested by centrifugation and resuspended in RCVBmedium.
.VOL. 148, 1981
on Septem
ber 8, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

588 BEATTY AND GEST
ant derivatives of the wild type (see Materialsand Methods). Such mutants, typified by strainT500, showed the same special growth require-ment as WE29, namely, an exogenous source ofKG such as L-glutamate. Levels of activities ofC6 branch enzymes in extracts of B10, WE29,and T500 cells (grown photosynthetically inRCVB medium supplemented with 10 mM L-glutamate) are shown in Table 2. The depend-ence of mutant WE29 and T500 on an exogenoussource of L-glutamate for photosynthetic growthdemonstrates the biosynthetic role of the C6branch of the citric acid cycle.Mutants of R. capsulata defective in the C6
branch would be expected to grow poorly, if atall, aerobically in darkness on malate even whensupplied with L-glutamate since in such circum-stances the energy supply depends on operationof the complete cycle. KGD- mutants, in fact,cannot grow aerobically in darkness on carbon/energy sources that must be catabolized via thecitric acid cycle (6a), and similar observationswere made with T500. In RCVB medium sup-plemented with 10 mM L-glutamate, the CS-negative mutant grew aerobically to a very lim-ited extent with a doubling time of about 24 h(the doubling time of the wild type in the samemedium is ca. 3 h).Biosynthesis of citric acid cycle enzymes
in R. capwulata: CS and ICD. As indicators ofC6 branch enzymes, levels of CS and ICD were
assayed in extracts of wild-type R. capsulatagrown under a wide variety of nutritional con-
ditions. These encompassed three modes of en-
ergy conversion, namely, photosynthetic, darkrespiratory, and dark fermentative. The growthconditions used and the enzyme-specific activi-ties observed are listed in Table 3, which alsogives the ratios of ICD to CS activities. In gen-eral, specific activities of both ICD and CS were
higher in extracts of aerobically grown cells thanin those from cells grown photosynthetically.The activities in "aerobic extracts," however,varied more widely, probably reflecting differ-ences among cultures in respect to medium pHand other conditions that change as cell densityincreases during growth. Despite the wide vari-ation in nutritional circumstances, the ratio ofICD to CS activities was relatively constant.This is also seen in the differential plot shown inFig. 4. An approximately linear relationship ofthe kind observed is interpreted as indicatingcoordinate expression of genes coding for thetwo enzymatic activities in question (3, 15). Thescatter of values from the line of the order seen
in Fig. 4 is commonly observed in other systemswhere enzymes are coordinately synthesized (3,12, 38).
TABLE 2. Activities of C6 branch citric acid cycleenzymes in extracts ofR. capsulata wild-type and
mutant strainsEnzyme activitya
Strain Aconi-CS tate hy- ICD
dratase
B10 (wild type) 142 67 627WE29 <1 85 <1T500 <1 37 541
a Values expressed in terms of nanomoles of sub-strate transformed per minute per milligram of pro-tein.
Factors affecting KGD synthesis in RCap8siat(L In R. capsulata, succinyl-CoA ap-pears to be produced from malate and fumarateonly via KGD activity (6a) and is required as abiosynthetic precursor under both respiratory(dark) and photosynthetic growth conditions. Itmight be supposed that with light as the energysource for growth, synthesis of KGD would beappreciably depressed as compared to cellsgrowing aerobically in darkness on typical or-ganic substrates, that is, under conditions inwhich KGD activity is also necessary for citricacid cycle-supported energy conversion. This no-tion was investigated by determination of KGDactivity levels in cells cultivated on various car-bon sources in the alternative growth modes (seeTable 4). Except for two special situations (withsuccinate or L-glutamate as carbon sources), ex-tracts of "aerobic dark cells" showed KGD ac-tivities three- to fivefold higher than extractsfrom cells grown photosynthetically. Thus, therelative KGD levels in aerobic and photosyn-thetic cells roughly parallel the relative respira-tory rates observed in cells grown in the twomodes (Table 1).
Exceptions to the general pattern of results inTable 4 were observed with cultures grown aer-obically on succinate and photosynthetically onL-glutamate as sole carbon sources. Succinate is,of course, derived from the succinyl-CoA gener-ated by KGD activity, and when the C4 acidserved as the sole carbon source aerobically, theenzyme level was significantly decreased. Glu-tamate as a sole carbon source under photosyn-thetic conditions, on the other hand, appearedto induce KGD synthesis.
Keevil et al. (18) have recently reported thateven trace concentrations of 02 (ca. 20 ppm) inthe gas phase can lead to detectably increasedKGD levels in Citrobacterfreundii cells growingin "anaerobic" cultures. This is also true for R.capsulata as evidenced from experiments whichwill be noted here in brief summary. R. capsu-
J. BACTERIOL.
on Septem
ber 8, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

CITRIC ACID CYCLE IN PHOTOSYNTHETIC BACTERIA 589
TABLE 3. CS and ICD activities in extracts ofR. capsulata cells grown under various conditionsd~~~~~~~~~~~~~~~~~~~~~
C/N sources for Growth condi- Symbol in Culture den- Activity'grourtha tions' Fig. 4 sityC CD S CD/CS
Malate/NH4+ An/L 1 300 389 80 4.8Malate/NH4+ An/L 2 256 463 90 5.2Malate/NH4+ An/L 3 310 634 80 8.0Malate/NH4+ An/L 4 138 640 142 4.5Malate/NH4+ An/L 5 315 405 65 6.3Lactate/NH4+ An/L 6 280 659 92 7.2Malate/glutamate An/L 7 294 683 95 7.2Malate/glutamate An/L 8 264 507 94 5.4Malate + KG/NH4+ An/L 9 305 487 98 5.0Glucose/NH4+ An/L 10 126 549 115 4.8Pyruvate/NH4+ An/L 11 192 369 92 4.0CO2/NH4+ e An/L 12 117 477 96 5.0Malate/N2 An/L 13 115 580 107 5.4Fructose/NH4+ f An/D 14 71 493 129 3.8Malate/NH4+ Aer/D a 175 989 185 5.3Malate/NH4+ Aer/D b 275 997 257 3.9Malate/NH4+ Aer/D c 153 497 103 4.8Malate/NH4+ Aer/D d 240 457 192 2.4Malate/NH4+ Aer/D e 300 471 165 2.9Malate/glutamate Aer/D f 320 1,080 284 3.8Malate/glutamate Aer/D g 385 1,010 281 3.6Lactate/NH4+ Aer/D h 455 1,630 415 4.0Lactate/NH4+ Aer/D i 300 914 287 3.2Pyruvate/NH4+ Aer/D j 190 809 143 5.7
a All media contained the mineral salts and vitamins specified for RCVB medium.b An/L, Anaerobic/light; An/D, anaerobic/dark; Aer/D, aerobic/dark.'Photometer units at time of cell harvest.d Activities are expressed in terms of nanomoles of substrate transformed per minute per milligram of protein.e Gas phase, 5% C02 + 95% H2.f Trimethylamine-N-oxide was used as the accessory oxidant for fermentative growth (23).
lata B10 was grown photosynthetically in con-tinuous culture essentially as described by Aik-ing and Sojka (1), using RCVB medium modifiedso as to contain only 6 mM DL-malate (thelimiting nutrient). Special precautions weretaken, however, to remove 02 from the gas phaseof ultrapure argon, and insofar as possible, allsupply lines and connections were constructedof glass tubing. At a dilution rate of 0.05 h-1(doubling time of 14 h), the KGD specific activ-ity in extracts of the cells from several experi-ments was 15 ± 4 nmol min-lmg protein-'.This activity corresponds to that observed inphotosynthetic batch-culture cells (Table 4).When the glass tubing that served as the me-dium supply line was replaced with latex rubbertubing, the KGD level in extracts of cells har-vested later increased to a specific activity of 25+ 0.6. The quantity of 02 that diffuses throughthe rubber tubing connection in such experi-ments is quite low and not easily measurable.The presence of a very low P02, however, isindicated by the fact that after replacement ofglass with rubber tubing, the (continuous) cul-ture color becomes more reddish owing to an O2-
dependent production of red carotenoids (H.Aiking, J. Cox, and G. Sojka, Abstr. Annu. Meet.Am. Soc. Microbiol. 1981, I123, p. 107). Molec-ular oxygen is clearly an important regulatorysignal for synthesis of KGD and, consequently,for expression of overall citric acid cycle activity.Photosynthetic growth of KGD- mu-
tants. KGD11 and similar mutants cannot growaerobically in darkness on succinate, malate, orpyruvate (6a). Experiments with KGD11 as thetest organism showed, however, that despite theapparent absence of KGD, this strain can growphotoheterotrophically on malate, fumarate, orsuccinate as sole carbon sources. Photosyntheticgrowth on succinate can be readily rationalizedsince the succinyl-CoA required for biosynthesiscan be generated directly from succinatethrough the action of succinyl-CoA synthetase.Since R. capsulata has no demonstrable fuma-rate reductase activity (6a), however, it is at firstglance difficult to see how a KGD- mutant cangrow on fumarate or malate. How is succinyl-CoA generated from fumarate or malate in amutant lacking both fumarate reductase andKGD activities? Isocitrate lyase activity was
VOL. 148, 1981
on Septem
ber 8, 2018 by guesthttp://jb.asm
.org/D
ownloaded from
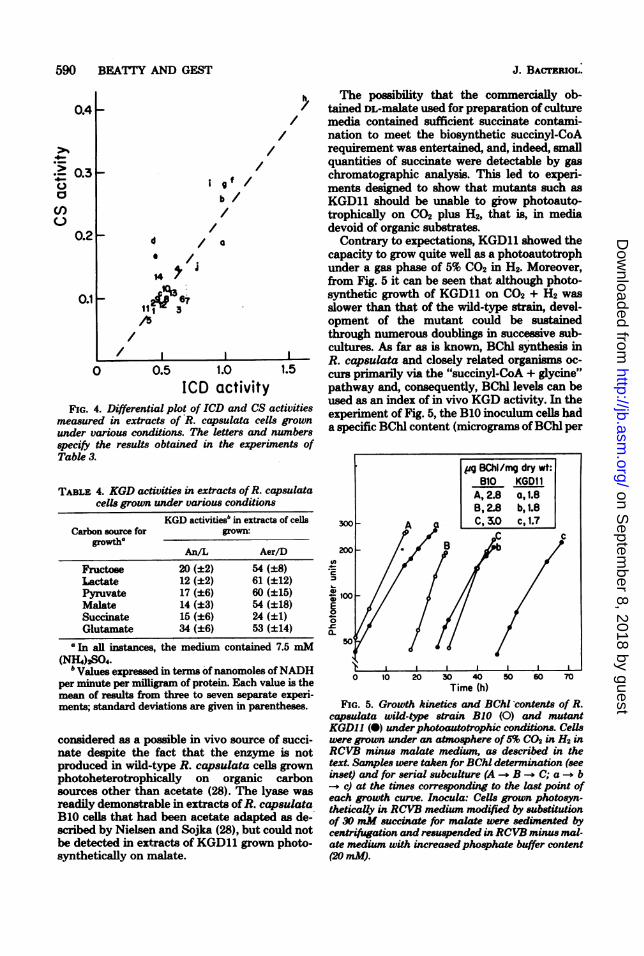
590 BEATTY AND GEST
0.4
0.3 _
0.2H_
0.1 _-
i gf /
d / a
/~~~
0
3
IA
0 0.5 1.0ICD activity
FIG. 4. Differential plot of ICD and CS activitiesmeasured in extracts of R. capsulata cells grownunder various conditions. The letters and numbersspecify the results obtained in the experiments ofTable 3.
TABLE 4. KGD activities in extracts ofR. capsulatacells grown under various conditions
KGD activitiesb in extracts of cellsCarbon source for grown:
growthaAn/L Aer/D
Fructose 20 (+2) 54 (+8)Lactate 12 (±2) 61 (±12)Pyruvate 17 (±6) 60 (±15)Malate 14 (±3) 54 (±18)Succinate 15 (+6) 24 (±1)Glutamate 34 (±6) 53 (±14)
a In al instances, the medium contained 7.5 mMNH4)SO4.
b Values expressed in terms of nanomoles ofNADHper minute per milligrm of protein. Each value is themean of results from three to seven separate experi-ments; standard deviations are given in parentheses.
considered as a possible in vivo source of succi-nate despite the fact that the enzyme is notproduced in wild-type R. capsulata cells grownphotoheterotrophically on organic carbonsources other than acetate (28). The lyase wasreadily demonstrable in extracts ofR. capsulataB10 cells that had been acetate adapted as de-scribed by Nielsen and Sojka (28), but could notbe detected in extracts of KGD11 grown photo-synthetically on malate.
The possibility that the commercially ob-tained DL-malate used for preparation of culturemedia contained sufficient succinate contami-nation to meet the biosynthetic succinyl-CoArequirement was entertained, and, indeed, smallquantities of succinate were detectable by gaschromatographic analysis. This led to experi-ments designed to show that mutants such asKGD11 should be unable to grow photoauto-trophically on C02 plus H2, that is, in mediadevoid of organic substrates.
Contrary to expectations, KGD11 showed thecapacity to grow quite well as a photoautotrophunder a gas phase of 5% C02 in H2. Moreover,from Fig. 5 it can be seen that although photo-synthetic growth of KGD11 on C02 + H2 wasslower than that of the wild-type strain, devel-opment of the mutant could be sustainedthrough numerous doublings in successive sub-cultures. As far as is known, BChl synthesis inR. capsulata and closely related organisms oc-curs primarily via the "succinyl-CoA + glycine"pathway and, consequently, BChl levels can beused as an index of in vivo KGD activity. In theexperiment of Fig. 5, the B10 inoculum cells hada specific BChl content (micrograms ofBChl per
.g BChl/rng dry wt:B10 KGDI1
A, 2.8 a, 1.8B, 2.8 b, 1.8
300 A C, 30 c, 1.7C c
0
a.50
0 10 20 30 40 50 60 70Time (h)
FIG. 5. Growth kinetics and BChl contents of R.capsulata wild-type strain B1O (0) and mutantKGD11 (0) under photoautotrophic conditions. Cellswere grown under an atmosphere of5% CO2 in H2 inRCVB minus malate medium, as described in thetext. Samples were taken for BChl determnination (seeinset) and for serial subculture (A -+ B -- C; a -- b
c) at the times corresponding to the last point ofeach growth curve. Inocula: Cells grown photosyn-theticaly in RCVB medium modified by substiutionof 30 mM succinate for malate were sedimented bycentriftigation and resuspended inRCVB minus mal-ate medium with increased phosphate buffer content(20mM).
4-
-.
0
C/)
J. BACTaOL.
on Septem
ber 8, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

CITRIC ACID CYCLE IN PHOTOSYNTHETIC BACTERIA 591
milligram [dry weight] of cells) of 2.7, whereasthe value for the KGD11 inoculum cells was 2.2.From the BChl values shown at the top of Fig.5, it is evident that the BChl content of B10 wasmaintained at the normal level during photoau-totrophic subculture. In KGD11, the specificBChl content diminished from the initial valueof 2.2 in the succinate-grown inoculum to 1.7 inthe third photoautotrophic subculture. The lat-ter content was obviously still sufficient to sup-port relatively good photoautotrophic growth,but since the BChl level was significantly lowerthan the wild-type value, it can be concludedthat the rate ofsuccinyl-CoA generation in grow-ing cells of the mutant was appreciably dimin-ished. In the experiment of Fig. 5, controls "wereincluded to determine the reversion frequenciesof KGD11 under the conditions used, and thesewere of the order of 8 x 10-6 or less; suchfrequencies cannot account for a significant frac-tion of the BChl observed in photoautotrophi-cally grown mutant populations.
DISCUSSION
Organic acids such as malate, lactate, andpyruvate are readily used by most representa-tives of the Rhodospirillaceae as carbon sourcesfor photoheterotrophic growth, or as carbon andenergy sources for aerobic dark growth. It seemsto be generally assumed that catabolism of theorganic acids occurs via the citric acid cycle, butthis view is largely an extrapolation from studieson the metabolism of Rhodospirillum rubrum(4, 29). The present investigation provides evi-dence for operation of the cycle as a majormechanism in the dark aerobic (heterotrophic)metabolism of a typical and widely investigatedspecies of the family Rhodospirillaceae. As inother kinds of cells, the citric acid cycle canfulfill several metabolic functions, primarily (i)generation of reduced pyridine nucleotide tosupport "electrophosphorylation" (13) and (ii)production of biosynthetic intermediates(mainly oxaloacetate, succinyl-CoA, and KG).The central importance of the cycle, or of com-ponent sequences, also is indicated by the factthat the activities of various enzymes are regu-lated by internediates of the cycle (5, 25).
Since it is a precursor of the glutamate neededfor protein synthesis, KG is required in substan-tial quantity for cell growth. Thus, photosyn-thetic growth is severely inhibited in mutantsthat have effective mutational blocks in the C6branch of the cycle, unless the cells are supple-mented with exogenous glutamate (or KG). Mu-tants of this kind are also unable to grow aero-bically in darkness on organic acids, even when
supplied with glutamate, due to failure of theenergy conversion function of citric acid cycleactivity.
In cells growing photoheterotrophically withlight as the energy source, the KGD reaction issignificant only from a biosynthetic standpoint,that is, for generation of succinyl-CoA. The lat-ter is used primarily for synthesis of BChl andheme proteins, which in photosyntheticallygrown Rhodopseudomonas are found to the ex-tent of ca. 10 umol of BChl-mg protein-' (11, 34)and ca. 0.5 nmol of total heme.mg protein-' (30).From these values, the miniimum rate of succi-nyl-CoA synthesis necessary for maintaining the2-h doubling time characteristic of wild-type R.capsulata in RCVB medium (24) can be calcu-lated to be ca. 0.7 nmol.min-'.mg protein-' (atsaturating light intensity; at low light intensitywith a doubling time of 5 h, the calculated valueis ca. 0.6). The quantity ofsuccinyl-CoA requiredfor biosynthesis is clearly of a different order ofmagnitude than that of KG. In Rhodospirilla-ceae growing photosynthetically, the maximumpotential rate of KGD activity is much higherthan the miniimum needed to produce the re-quired succinyl-CoA (6a). Thus, typical specificactivities of KGD in extracts of R. capsulata areof the order of 15. Such values, of course, rep-resent maximal activities observed in vitro withall substrates and cofactors at saturating concen-trations. We assume that the in vivo rates mustbe considerably lower and subject to a variety ofregulatory controls.
Calculations similar to those noted above canbe made for mutant KGD11 growing photoau-totrophically (on CO2 + H2) with a doublingtime of 8 h and a somewhat diminished BChlcontent (Fig. 5). In this case, the required in vivorate ofsuccinyl-CoA production is ca. 0.03 nmol-min-'-mg protein-'. Such low rates cannot bedetected in cell extracts with available proce-dures, and this explains the seeming paradox ofthe capacity ofKGD11 to grow photoautotroph-ically despite the "absence" ofKGD activity andof fumarate reductase activity. Accordingly, inmutants of this kind, very low rates of KGDactivity could account for the succinyl-CoAneeded for biosyntheses. Even when there is asevere genetic block, "mutant enzymes" canhave low residual activity (leakiness), and it isalso possible that succinyl-CoA could be slowlyproduced by the pyruvate dehydrogenase com-plex or in some other way. It is relevant thatobservations similar to those under discussionhave been made with E. coli. Creaghan andGuest (9) constructed mutants of E. coli lackingKGD, fumarate reductase, succinate dehydro-genase, and isocitrate lyase activities, and these
VOL. 148, 1981
on Septem
ber 8, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

592 BEATTY AND GEST
still could grow slowly with glucose as the solecarbon and energy source. The cells showed areduced heme content as compared with wildtype, and it was concluded that leaks throughthe mutational blocks or an unknown metabolicpathway could supply succinate (or succinyl-CoA) at a low rate. The existence in green plantsof a mechanism for conversion of KG to theporphyrin precursor 8-aminolevulinic acid notinvolving succinyl-CoA has been established (6,17, 21), but the significance, if any, of this path-way in photosynthetic bacteria is still unclear(for recent studies in this connection, see refer-ence 8). Investigations on purple photosyntheticbacteria indicate that in these organisms, themajor (if not sole) source of 8-aminolevulinicacid is the succinyl-CoA + glycine condensationreaction (20).Although KGD- mutants of R. capsulata vir-
tually devoid of KGD activity apparently pro-duce sufficient succinyl-CoA for biosyntheticpurposes when light is the energy source, themetabolic lesion in such strains is lethal for theaerobic dark growth mode because the bioener-getic function of the citric acid cycle cannot bemaintained. The increase of KGD activity seenin extracts of cells grown aerobically as com-pared to extracts from "photosynthetic cells" isconsistent with the need for greatly increasedcarbon flow via the cycle to maintain an ade-quate supply of reducing equivalents for theenergy-converting respiratory apparatus.The various controls that affect formation of
citric acid cycle enzymes and their activities arestill not well understood (for a recent discussionofregulation of the cycle in mammalian systems,see reference 37). Deeper insights into this as-pect of cell metabolismn should be facilitated bymore detailed study of organisms like R. capsu-lata that have well-developed alternative mech-anisms of energy conversion. In this respect, theeffects of 02 on R. capsulata are of specialinterest. The observed increase in activity ofseveral citric acid cycle enzymes as well as of theNADH oxidase system in cells grown aerobicallysuggests that exposure to 02 induces a multicom-ponent metabolic system of considerable com-plexity. Molecular oxygen appears to exert reg-ulatory effects even when present at very lowconcentration, and this may well have meaningfor further analysis of the biochemical evolutionof aerobic systems from fermentative precursors.
ACKNOWLEDGMENTSThis,research was supported by National Science Foun-
dation Grant PCM-7910747. In addition, J.T.B. received sti-pend support from a National Institutes of Health traininggrant (Public Health Service; T32 CM-07227-04), and H.G.was the recipient of a fellowship from the John Simon Gug-genheim Memorial Foundation.
We thank Jeff Favinger and Marcia Dits for skilled tech-nical assistance, Michael Madigan for helpful discussions, andJohn Cox for collaboration in the continuous culture experi-ments.
LITERATURE CITED1. Aiking, H., and G. Sojka. 1979. Response ofRhodopseu-
domonas capsulata to illumination and growth rate ina light-limited continuous culture. J. Bacteriol. 139:530-536.
2. Amarasingham, C. R., and B. D. Davis. 1965. Regula-tion of a-ketoglutarate dehydrogenase formation inEscherichia coli. J. Biol. Chem. 240:3664-3668.
3. Ames, B. N., and B. Garry. 1959. Coordinate repressionof the synthesis of four histidine biosynthetic enzymesby histidine. Proc. Natl. Acad. Sci. U.S.A. 45:1453-1461.
4. Anderson, L., and R. C. Fuller. 1967. Photosynthesis inRhodospirillum rubrum. Ill. Metabolic control of re-ductive pentose phosphate and tricarboxylic acid en-zymes. Plant Physiol. 42:497-502.
5. Atkinson, D. E. 1968. Citrate and the citrate cycle in theregulation of energy metabolism, p. 23-40. In T. W.Goodwin (ed.), The metabolic roles of citrate. Biochem.Soc. Symposium no. 27. Academic Press, London.
6. Beale, S. I., S. P. Gough, and S. Granick. 1975. Biosyn-thesis of 8-aminolevulinic acid from the intact carbonof glutamic acid in greening barley. Proc. Natl. Acad.Sci. U.S.A. 72:2719-2723.
6a.Beatty, J. T., and H. Gest. 1981. Generation of succinyl-coenzyme A in photosynthetic bacteria. Arch. Micro-biol. 129:335-340.
7. Beatty, J. T., B. C. Johansson, J. D. Wall, and H.Gest. 1977. Nitrogen assimilation defects in a mutantof Rhodopseudomonas capsulata blocked in a-ketoglu-tarate generation. FEMS Microbiol. Lett. 2:267-270.
8. Chen, J., G. W. Miller, and J. Y. Takemoto. 1981.Biosynthesis of 8-amirolevulinic acid in Rhodopseu-domonas sphaeroides. Arch. Biochem. Biophys. 208:221-228.
9. Creaghan, I. T., and J. R. Guest. 1978. Succinate de-hydrogenase-dependent nutritional requirement forsuccinate in mutants of Escherichia coli K12. J. Gen.Microbiol. 107:1-13.
10. Dawson, R. M. C., D. C. Elliott, W. H. Elliott, and K.M. Jones (ed.). 1969. Data for biochemical research,2nd ed. Oxford University Press, New York.
11. Dierstein, R., and G. Drews. 1974. Nitrogen-limitedcontinuous culture of Rhodopseudomonas capsulatagrowing photosynthetically or heterotrophically underlow oxygen tensions. Arch. Microbiol. 99:117-128.
12. Englesberg, E., J. Irr, J. Power, and N. Lee. 1965.Positive control of enzyme synthesis by gene C in theL-arabinose system. J. Bacteriol. 90:946-957.
13. Gest, H. 1980. The evolution of biological energy-trans-ducing systems. FEMS Microbiol. Lett. 7:73-77.
14. Harford, S., and P. D. J. Weitzman. 1980. Selection forcitrate synthase-deficient mutants with fluoroacetate.FEBS Lett. 114:339-341.
15. Jacob, F., and J. Monod. 1961. On the regulation of geneactivity. Cold Spring Harbor Symp. Quant. Biol. 26:193-211.
16. Johansson, B. C., and H. Gest. 1976. Inorganic nitrogenassimilation by the photosynthetic bacterium Rhodo-pseudomonas capsulata. J. Bacteriol. 128:683-688.
17. Jones, 0. T. G. 1978. Biosynthesis of porphyrins, hemes,and chlorophylls, p. 751-777. In R. K. Clayton and W.R. Sistrom (ed.), The photosynthetic bacteria. PlenumPress, New York.
18. Keevil, C. W., J. S. Hough, and J. A. Cole. 1979.Regulation of 2-oxoglutarate dehydrogenase synthesisin Citrobacter freundii by traces of oxygen in commer-cial nitrogen gas and by glutamate. J. Gen. Microbiol.
J. BACTERIOL.
on Septem
ber 8, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

CITRIC ACID CYCLE IN PHOTOSYNTHETIC BACTERIA 593
114:355-359.19. Lakshmi, T. M., and R. B. Helling. 1976. Selection for
citrate synthase deficiency in icd mutants of Esche-richia coli. J. Bacteriol. 127:76-83.
20. Lascelles, J. 1978. Regulation of pyrrole synthesis, p.795-808. In R. K. Clayton and W. R. Sistrom (ed.), Thephotosynthetic bacteria. Plenum Press, New York.
21. Lohr, J. B., and H. C. Friedman. 1976. New pathwayfor 8-aminolevulinic acid synthesis: forniation from a-
ketoglutaric acid by two partially purified plant en-
zymes. Biochem. Biophys. Res. Commun. 69:908-913.22. Lowry, 0. H., N. J. Rosebrough, A. L Farr, and R. J.
Randall. 1951. Protein measurement with the Folinphenol reagent. J. Biol. Chem. 193:265-275.
23. Madigan, M. T., J. C. Cox, and H. Gest. 1980. Physiol-ogy of dark fermentative growth ofRhodopseudomonascapsulata. J. Bacteriol. 142:908-915.
24. Madigan, M. T., and H. Gest 1979. Growth of thephotosynthetic bacterium Rhodopseudomonas capsu-lata chemoautotrophically in darkness with H2 as theenergy source. J. Bacteriol. 137:524-530.
25. Magasanik, B. 1977. Regulation of bacterial nitrogenfixation by glutamine synthetase. Trends Biochem. Sci.2:9-12.
26. Meynell, G. G., and E. Meynell. 1970. Theory andpractice in experimental bacteriology, 2nd ed., p. 257-259. Cambridge University Press, Cambridge, U.K..
27. Morrison, J. F., and R. A. Peters. 1954. Biochemistryof fluoroacetate poisoning: the effect of fluorocitrate onpurified aconitase. Biochem. J. 58:473-479.
28. Nielsen, A. M., and G. A. Sojka. 1979. Photohetero-trophic utilization of acetate by the wild type and an
acetate-adapted mutant of Rhodopseudomonas capsu-
lata. Arch. Microbiol. 120:39-42.29. Ormerod, J. G., and H. Gest. 1962. Symposium on
metabolism of inorganic compounds. IV. Hydrogen pho-tosynthesis and alternative metabolic pathways in pho-tosynthetic bacteria. Bacteriol. Rev. 26:51-66.
30. Porra, R. J., and J. Lascelles. 1965. Haemoproteins andhaem synthesis in facultative photosynthetic and deni-trifying bacteria. Biochem. J. 94:120-126.
31. Reeves, H. C., R. Rtabin, W. S. Wegener, and S. J. AJl.1971. Assays of enzymes of the tricarboxylic acid andglyoxylate cycles. Methods Microbiol. 6A:437-439.
32. Smith, A. J., J. London, and R. Y. Stanier. 1967.Biochemical basis of obligate autotrophy in blue-greenalgae and thiobacilli. J. Bacteriol. 94:972-983.
33. Sojka, G. A., H. H. Freeze, and H. Gest 1970. Quanti-tative estimation of bacteriochlorophyll in situ. Arch.Biochem. Biophys. 136:578-580.
34. Sojka, G. A., and H. Gest 1968. Integration of energyconversion and biosynthesis in the photosynthetic bac-terium Rhodopseudomonas capsulata. Proc. Natl.Acad. Sci. U.S.A. 61:1486-1493.
35. Stern, J. R., and G. Bambers. 1966. Glutamate biosyn-thesis in anaerobic bacteria. I. The citrate pathways ofglutamate synthesis in Clostridium kluyveri. Biochem-istry 5:1113-1118.
36. Weaver, P. F., J. D. Wall, and H. Gest. 1975. Charac-terization of Rhodopseudomonas capsulata. Arch. Mi-crobiol. 105:207-216.
37. Williamson, J. R., and R. H. Cooper. 1980. Regulationof the citric acid cycle in mammalian systems. FEBSLett. 117(Suppl.):K73-K85.
38. Zabin, I. 1963. Proteins of the lactose system. Cold SpringHarbor Symp. Quant. Biol. 28:431435.
VOL. 148, 1981
on Septem
ber 8, 2018 by guesthttp://jb.asm
.org/D
ownloaded from





























