Embed Size (px)
Citation preview

UNIVERSIDADE FEDERAL DO RIO GRANDE DO NORTE
CENTRO DE BIOCIÊNCIAS
PROGRAMA DE PÓS-GRADUAÇÃO EM CIÊNCIAS BIOLÓGICAS
MESTRADO EM CIÊNCIAS BIOLÓGICAS
BIOLOGIA ALIMENTAR E MORFOHISTOLOGIA DO TUBO
DIGESTÓRIO DO MUSSUM, Synbranchus marmoratus, BLOCH
(OSTEICHTHYES: SYNBRANCHIDAE) NO AÇUDE MARECHAL
DUTRA “GARGALHEIRAS”, LOCALIZADO NO SEMI-ÁRIDO
BRASILEIRO.
LUCIANA ARAÚJO MONTENEGRO
NATAL / RN
2009

II
LUCIANA ARAÚJO MONTENEGRO
BIOLOGIA ALIMENTAR E MORFOHISTOLOGIA DO TUBO
DIGESTÓRIO DO MUSSUM, Synbranchus marmoratus, BLOCH
(OSTEICHTHYES: SYNBRANCHIDAE) NO AÇUDE MARECHAL
DUTRA “GARGALHEIRAS”, LOCALIZADO NO SEMI-ÁRIDO
BRASILEIRO.
ORIENTADOR: Profª. Drª. SATHYABAMA CHELLAPPA
DOL / CB / UFRN
Versão da Dissertação de Mestrado apresentada ao
Programa de Pós-Graduação em Ciências Biológicas
do Centro de Biociências, da Universidade Federal
do Rio Grande do Norte, como um dos requisitos
para obtenção do título de Mestre em Ciências
Biológicas.
NATAL / RN
ABRIL, 2009


III
UNIVERSIDADE FEDERAL DO RIO GRANDE DO NORTE CENTRO DE BIOCIÊNCIAS
PROGRAMA DE PÓS-GRADUAÇÃO EM CIÊNCIAS BIOLÓGICAS MESTRADO EM CIÊNCIAS BIOLÓGICAS
BIOLOGIA ALIMENTAR E MORFOHISTOLOGIA DO TUBO DIGESTÓRIO
DO MUSSUM, Synbranchus marmoratus, BLOCH (OSTEICHTHYES:
SYNBRANCHIDAE) NO AÇUDE MARECHAL DUTRA “GARGALHEIRAS”,
LOCALIZADO NO SEMI-ÁRIDO BRASILEIRO.
LUCIANA ARAÚJO MONTENEGRO
Esta versão da Dissertação, apresentada pela aluna LUCIANA ARAÚJO
MONTENEGRO ao Programa de Pós-Graduação em Ciências Biológicas, do Centro
de Biociências, da Universidade Federal do Rio Grande do Norte, foi julgada adequada
e aprovada, pelos Membros da Banca Examinadora para conclusão do curso de
Mestrado em Ciências Biológicas.
MEMBROS DA BANCA EXAMINADORA:
_________________________________________________________
Profa. Dra. SATHYABAMA CHELLAPPA – DOL / CB / UFRN
_________________________________________________________
Prof. Dr. NAITHIRITHI T. CHELLAPPA – DOL / CB / UFRN
_________________________________________________________
Prof. Dr. BHASKARA CANAN – SENAI/RN
_________________________________________________________
Prof. Dr. JOSÉ ZANON DE OLIVEIRA PASSAVANTE – UFPE-PE
Natal/RN, 30 de Abril de 2009.

IV
AGRADECIMENTOS
A Universidade Federal do Rio Grande do Norte, pela oportunidade concedida
através do Programa de Pós-Graduação em Ciências Biológicas, em especial aos
professores do quadro de mestrado;
A professora Dra. SATHYABAMA CHELLAPPA, pela orientação e confiança
deposita em minha pessoa durante todo mestrado, a quem tanto admiro pelo seus
esforços e dedicação à pesquisa, sempre me auxiliando com suas valiosas sugestões
para o enriquecimento deste trabalho;
Ao professor Dr. NAITHIRITHI CHELLAPPA pela colaboração, permitindo
que as análises limnológicas fossem realizadas em seu laboratório, bem como à sua
equipe de laboratório;
A Empresa de Pesquisas Agropecuárias do Rio Grande do Norte (EMPARN),
pelos dados metereológicos;
Ao Sr. FERNANDO, pela colaboração, com as suas preciosas fotografias, que
foram tão importantes para o enriquecimento deste trabalho de pesquisa;
A professora RAIMUNDA GONÇALVES DE ALMEIDA, pelo incentivo,
apoio, colaboração e preciosas sugestões, a quem tanto admiro, com amizade e respeito
por sua pessoa;
Aos meus queridos pais, a quem tanto amo e admiro Sr. LUIZ MONTENEGRO
DA CUNHA e Sra. MARIA NAZARÉ ARAÚJO CUNHA, por me incentivarem e
apoiarem em todos os momentos de realização do meu mestrado, fazendo-me prosseguir
em frente com confiança e direcionamento;
Ao meu amigo e esposo, MARCOS ALEXANDRE, pela compreensão e
colaboração em todos os momentos do mestrado;
As minhas queridas filhas ADRIANNA e LYVIA, que em uma das fases mais
bonitas da sua infância, souberam da sua maneira, entender a minha ausência;
As colegas do LABICTIS pelas boas horas que passamos juntos, em especial a
DAISY pela preciosa contribuição nas coletas e apoio constante durante a realização das
biometrias;
A professora NAISSANDRA BEZERRA DA SILVA, pelo direcionamento,
apoio e colaboração na preparação e leituras das lâminas histológicas;

V
Ao DEPARTAMENTO DE MORFOLOGIA E HISTOLOGIA da UFRN, em
especial a MARIA DE LURDES pela paciência nata e colaboração durante a preparação
das lâminas histológicas;
A LILIANE GURGEL pelas preciosas sugestões e colaborações durante a
realização deste trabalho;
A ELIZETE CAVALCANTI, sugestões enriquecedoras e grandiosos momentos
de convívio;
A grande e querida amiga, ANA CARLA PETROVICH IORIO pelo apoio
constante, dedicação e amizade;
A todos que colaboraram de maneira a tornar possível a realização deste
trabalho;
E em especial, a nossa MÃE SANTÍSSIMA, que sem a sua permissão essa nova
conquista não teria sido possível!

VI
SUMÁRIO
Página
RESUMO VIII
ABSTRACT IX
CAPÍTULO 1
1. INTRODUÇÃO
1.1 Ictiofauna das bacias do Semi-Árido
2
2
1.2 Biologia alimentar dos peixes 4
1.3 Considerações sobre a espécie em estudo 5
2. OBJETIVOS DA PESQUISA 8
2.1 Objetivo geral 8
2.2 Objetivos específicos 8
3. MATERIAL E MÉTODOS 10
3.1 Área de estudo 10
3.2 Coleta e taxonomia da espécie em estudo 13
3.3 Parâmetros metereológicos 15
3.4 Caracterização morfométrica e biometria dos peixes
3.5 Aspectos alimentares
15
16
Grau de Repleção 16
Índice de repleção estomacal (IR): 17
3.6 Análise da dieta 17
3.7 Análise morfohistológica do tubo digestivo
3.8 Análise estatística
17
18
3.8 Normalização das referencias bibliográficas 18
REFERÊNCIAS BIBLIOGRÁFICAS (Capítulo 1) 19

VII
CAPÍTULO 2
Artigo Cientifico 1
REFERÊNCIAS
LISTA DE TABELAS
LEGENDA DE FIGURAS
Artigo Cientifico 2
REFERÊNCIAS
LISTA DE FIGURAS
28
39
42
43
48
54
57
CAPÍTULO 3
DISCUSSÃO GERAL
CONCLUSÃO
REFERÊNCIAS BIBLIOGRÁFICAS GERAIS
65
67
68

VIII
RESUMO
O regime alimentar e a morfohistologia do tubo digestório do mussum, Synbranchus marmoratus (Block, 1917) foram investigados. Os exemplares foram capturados mensalmente no período de agosto de 2007 a julho de 2008, no açude Marechal Dutra, Acari, Rio Grande do Norte. Foram verificados os parâmetros limnológicos, tais como, temperatura da água, oxigênio dissolvido e condutividade elétrica do açude. Os valores mensais de pluviosidade foram obtidos da EMPARN. Os peixes capturados foram medidos, pesados, eviscerados e registrado o peso de cada estômago. A análise do conteúdo estomacal foi realizada de acordo com o método de pontos e método de freqüência de ocorrência associado ao método de importância alimentar. Os estômagos tiveram o grau de repleção determinado e o índice de repleção relacionando a atividade alimentar com o período chuvoso e de estiagem. A precipitação pluviométrica variou de 0 a 335 mm. A média anual da pluviosidade foi de 71,62 mm ± SD 163,3. O maior valor de 335,5 mm ocorreu em março de 2008 e os menores valore ocorreram nos meses de agosto a dezembro de 2007. No período de seca, S. marmoratus apresentou valores mais altos no Índice médio de Repleção, com um pico no mês de setembro (4,54; ± SD = 0,56), sendo esse o maior valor médio de IR anual. O menor valor (3,99; ± SD = 0,25) ocorreu no mês de maio no período chuvoso. Os resultados obtidos indicam que S. marmoratus alimentou-se preferencialmente de material animal, sendo 78,22% de crustáceos, 13,5% de material orgânico semi digerido, 3,25% de moluscos, 2,85% de peixes e 1,4% de insetos. Portanto, a espécie em estudo pode ser caracterizada como carnívoro com preferência a carcinofagia. Os aspectos morfohistológicos do tubo digestório do S. marmoratus foram investigados e os resultados indicam que boca é terminal com grande abertura, lábios delgados com corpúsculos gustativos, os dentes são pequenos villiformes formando uma única série nas maxilas, com quatro pares de arcos branquiais com rastros branquiais curtos e espassados. O esôfago é curto e apresenta formato cilíndrico com pequeno diâmetro. A parede esofágica é espessa com a superfície interna formada por mucosa, contendo pregas paralelas. O estômago é do tipo retilíneo, formado pelas porções cárdica, cecal e pilórica. A região de maior comprimento é a região cecal, localizada entre a porção cárdica e pilórica. A região cárdica que é a primeira porção do estômago é curta e cilíndrica, formada por mucosa constituída por epitélio cilíndrico simples, com muitas células caliciformes. A região cecal é alongada e apresenta paredes delgadas, luz maior que a cárdica, com pregas de menor porte, formado por epitélio cilíndrico simples com glândulas gástricas. A porção pilórica não possui glândulas e apresenta pregas principais e secundárias em sua mucosa. Os aspectos morfohistológicos do trato digestório do S. marmoratus mostram que a espécie é adaptada para hábito alimentar carnívoro. Palavras chave: peixe, dieta alimentar, carnívoro, açude semi-árido.

IX
ABSTRACT Food habits and morpho-histology of the digestive tract of marbled swamp eel, Synbranchus marmoratus (Block, 1917) were investigated. The fish samples were captured during August, 2007 to July, 2008 in the Marechal Dutra reservoir, Acari, Rio Grande do Norte. The rain fall data was obtained from EMPARN. The fish captured, were measured, weighed, dissected, eviscerated and individual stomach weights were registered. The stomach contents analyses were carried out based on volumetric method, points, frequency of occurrence and applying the Index of Relative Importance. The degrees of repletion of the stomachs were determined besides the Index of Repletion relating to feeding activity variations and frequency of ingestion during the rainy and dry seasons. The rainfall varied from 0 mm a 335 mm with a mean value of 71.62 mm. Highest rainfall of 335.5 mm was registered in March, 2008 and August to December was the dry period. During the dry period the study species presented high degrees of repletion of the stomachs, with a peak value in the month of September (mean = 4.54; ± SD = 0.56). The minimum mean value of = 3.99 ± SD = 0.25 was registered in the month of May during the rainy period. The stomach contents of S. marmoratus registered show that this fish prefers animals, 78.22% of crustaceans 2.85% of mollusks, 3.25% of fish, 1.4% of insects and 13.5% of semi-digested organic matter, thus characterizing the study species as a carnivore with a preference for crustaceans. The morpho-histological aspects of the digestive tract of S. marmoratus indicate that the mouth is terminal adapted to open widely, thin lips with taste buds, small villiform teeth forming a single series on maxillas, four pairs of branchial arches with short and widely spaced branchial rays. The oesophagus is short and cylindrical with a small diameter. The oesophagus wall is thick with mucas surface and internal parallel folds. The stomach is retilinical in form, presenting cardiac, caecal and pyloric portions. The caecal portion is long and is intermediary in position between the cardiac and pyloric portions. The cardiac portion of the stomach is short and cylindrical formed of simple epithelial cylindrical mucus cells. The caecal portion is long with narrow walls, a big cavity and smaller folds which give rise to gastric glands. The phyloric portion has no glands and primary or secondary mucas folds. The morphohistological aspects of the digestive tract of S. marmoratus indicate its adaptation to a carnivorous feeding habit. Key Words: fish, food habits, carnivore, semi-arid reservoir.

1
CAPÍTULO 1
CAPÍTULO 1
INTRODUÇÃO GERAL

2
1. INTRODUÇÃO
1.1 - Ictiofauna das bacias do Semi-Árido
A ictiofauna dos ecossistemas dulciaquícolas da Caatinga inclui representantes de
diferentes grupos neotropicais e suas espécies distribuem-se em bacias interiores e costeiras do
nordeste brasileiro, que drenam parcialmente ou estão inteiramente localizadas na Caatinga.
Por isso, não há como caracterizar uma ictiofauna típica ou exclusiva deste ecossistema, uma
vez que a distribuição de muitas espécies nos rios (que cortam a caatinga) estendem-se para
além de seus limites, atingindo outros ecossistemas da região nordeste brasileiro e regiões
vizinhas (ROSA 2004; ROSA et al., 2005).
A ictiofauna dos açudes do semi-árido brasileiro representa o resultado de
processos ecológicos que determinaram a adaptação de espécies às condições climáticas e
ao regime hidrológico da região. A compilação taxonômica dos peixes que ocorrem no
bioma Caatinga revelou a presença de 240 espécies, distribuídas em 7 ordens. Ordem
Siluriformes apresentou a maior diversidade, com 101 espécies (Família Loricariidae com
34 espécies; Família Pimelodidae com 27 espécies e Família Callichthyidae com 15
espécies. A ordem Characiformes apresentou a segunda maior diversidade, com 89
espécies (Família Characidae com 50 espécies). Entre os Cyprinodontiformes, a família
Rivulidae foi a mais variada com 24 espécies. As demais ordens são representadas pelos
Gymnotiformes (6), Perciformes (17), Myliobatiformes (1), Clupeiformes (1) e
Symbranchiformes (1). Do total de espécies de peixes encontradas, 9 são introduzidas e
136 são consideradas endêmicas para Caatinga. As 9 espécies introduzidas são Poecilia
latipinna, Poecilia reticulata, Plagioscion squamosissimus, Astronotus ocellatus, Cichla
monoculus, Cichla ocellaris, Geophagus surinamensis, Oreochromis niloticus e Tilápia
rendalli (ROSA et al. 2005).
Estudos sobre peixes em açudes e lagos do Estado do Rio Grande do Norte foram
fundamentais para se ter mais informações e enriquecer o conhecimento sobre a ictiofauna
dulciaquícola da caatinga. Neste sentido, Os trabalhos realizados por Gurgel et al. (1995) e
Canan et al. (1997) relatam sobre as espécies de peixes mais frequentes nos açudes do
estado do Rio Grande do Norte: Cará, Cichlasoma orientale; tucunaré, Cichla ocellaris;
pescada branca, Plagiosciom squamosissimus; piaba, Astyanax bimaculatus; jacundá,
Crenicichla menezesi; cangati, Parauchenipterus galeatus; piranha, Serrasalmus brandtii.

3
A diversidade de espécies de peixes na Lagoa do Piató foi relatada no estudo de
Almeida et al. (1993). Levantamentos ictiofaunisticos realizados por Freire et al. (2000),
em cinco reservatórios da bacia do Piranhas Assu, demonstrou uma composição
ictiofaunística representada por algumas espécies nativas como curimatã, Prochilodus
brevis, piau, Leoporinus piau; pirambeba, Serrasalmus maculatus; sardinha, Triportheus
angulatus; traíra, Hoplias malabaricus, branquinha, Curimata ciliata e cascudo,
Hypostomus sp. E algumas espécies introduzidas como apaiari, Astronotus ocellatus,
pescada branca, Plasgioscion squamosisssimus, tilápia do Nilo, Oreochromis niloticus e
tucunaré, Cichla monoculus.
Estudos sobre a ecologia reprodutiva do tucunaré, Cichla monoculus foi realizado
no açude Campo Grande, São Paulo do Potengi, RN (CÂMARA, 1998; CÂMARA et al.,
2002; CHELLAPPA et al., 2003a), e os aspectos reprodutivos do cangati,
Parauchenipterus galeatus e pirambeba, Serrasalmus maculatus, foram realizados na
lagoa de Extremoz, RN (MEDEIROS, 2001; MEDEIROS et al., 2003; MEDEIROS et al.,
2004). No açude Marechal Dutra, RN onde foram realizadas pesquisas abordando aspectos
ictiofaunísticos, Silva (2000), concordou com a ocorrência de espécies de peixes relatadas
no trabalho de Freire et al. (2000). Menescal (2002) observou uma redução de espécies de
peixes nativos como uma consequência da introdução da tilápia do Nilo, Oreochromis
niloticus, no açude Marechal Dutra. Foram realizados estudos sobre a ictiofauna de valor
comercial (BUENO; CHELLAPPA, 2003) e sobre a biologia reprodutiva do cascudo,
Hypostomus pusarum no açude Marechal Dutra, Acari, RN (BUENO, 2004). Nascimento
(2006) estudou os aspectos reprodutivos do curimatã comum, Prochilodus brevis no
mesmo açude.
Os trabalhos realizados contribuíram significante em relação ao conhecimento dos
aspectos ecológicos dessas espécies nos ecossistemas aquáticos do semi-árido brasileiro.
Contudo, não há um consenso acerca do status taxonômico e os estudos alimentares de
muitas espécies de peixes do bioma Caatinga. Este fato reflete a carência de estudos sobre
a ecologia alimentar e sistemática dos peixes dos açudes da região.
Os ecossistemas do semi-árido brasileiro têm como características as variações de seus
recursos hídricos, os baixos índices pluviométricos, bem como a elevada taxa de evaporação,
onde as espécies presentes desenvolvem estratégias de sobrevivência, que acarretam em
competições intra e interespecífica. A barragem de Marechal Dutra é um ecossistema que
apresenta uma imensa importância para o abastecimento de água, recreação e sustento da
população local (COSTA, 2000; CHELLAPPA & COSTA, 2003).

4
1.2 - Biologia alimentar dos peixes
Estudos acerca da biologia alimentar de espécies icticas vem recebendo atenção
especial principalmente devido à sua importância no que se refere à conservação de
ambientes aquáticos. Segundo Menezes (1994), sem estes estudos, não é possível
desenvolver planos de manejo sustentável visando à preservação das espécies de peixes,
bem como estabelecimento de parâmetros que devem nortear decisões sobre política de
preservação e programas de piscicultura.
Os ambientes de água doce oferecem poucas oportunidades para especialização de
peixes. Consequentemente, muitas espécies apresentam flexibilidade ampla em relação aos
diferentes tipos de hábito e habitat, compartilhando com outras espécies os recursos
disponíveis no ambiente (LARKIN, 1956). Os peixes diferem quanto ao tipo de alimento
consumido, mais do que qualquer outro grupo de vertebrados (NIKLOSKY, 1963)
indicando que embora haja diferenciação do hábito alimentar, exibem uma considerável
plasticidade que confere vantagens às espécies generalistas em relação às especialistas
(LOWE-MCCONNEL, 1999).
Em águas tropicais, apesar das espécies possuírem adaptações do trato digestório,
muitas delas mostram certa plasticidade em suas dietas, a qual pode estar relacionada a
diversos fatores, tais como, espaciais, sazonais, ontogenéticos e comportamentais
(AGOSTINHO e GOMES, 1997). O mesmo autor não recomenda fazer inferências a
respeito do hábito alimentar dos peixes baseando-se apenas em aspectos morfológicos.
Uma espécie pode mostrar caracteres morfológicos que indicam a exploração de um tipo
de alimento, mas ao se fazer à análise da dieta, esta pode apresentar hábito alimentar
diferente do esperado. Por exemplo, o ciclídeo Petrotilapia tridentiger, do Lago Malawi,
cuja dentição indica especialização para raspar algas de rochas, mas o estudo da dieta
alimentar mostra outros itens alimentares, diferentes do esperado pelos aspectos
morfológicos.
Segundo Zavala-Camin (1996), dados relacionados à alimentação fornecem
subsídios à compreensão dos seguintes temas: (1) nutrição da espécie, incluindo a
assimilação de alimentos; (2) os levantamentos faunísticos e florísticos do ambiente,
refletido no conteúdo estomacal; e (3) a transferência de energia nos ecossistemas. A
atividade alimentar gera elementos para se entender a relação entre os componentes da
ictiofauna e os demais organismos da comunidade aquática. Além disso, deve se levar em

5
conta que estudos sobre alimentação de peixes são indispensáveis para um efetivo manejo
da população, além de contribuir para o entendimento da biologia da espécie (HAHN,
1991; HAHN et al., 1997).
O conhecimento do regime alimentar das espécies nativas é de fundamental
importância para que possamos compreender toda a dinâmica do encadeamento biológico,
numa determinada coleção d’água. A primeira citação referente a alimentação de peixes
para o estado do Rio Grande do Norte é de Menezes & Menezes (1946), que estudaram
algumas espécies de peixes de ambiente dulciaquícolas do nordeste brasileiro, entre as
quais; cangati, Parauchenipterus galeatus.
Aspectos da alimentação de peixes para o Estado do Rio Grande do Norte foram
pesquisados por Magalhães et al. (1990) ao analisarem a dieta de Serrasalmus brandtii, do
rio Piranhas-Assu; Gurgel et al. (1994) estudando a dieta de Metynnis cf. roosevelti, da
Lagoa Redonda, Nísia Floresta. Chellappa et al. (1996), verificaram a dieta alimentar do
híbrido vermelho de tilapia, Oreochromis niloticus (Linnaeus) x Oreochromis
mossambicus (Peters) em viveiros dos peixes no RN; Canan et al. (1997) analisaram a
alimentação da comunidade de peixes da lagoa Boa Cicca, Gurgel et al. (1998) estudaram a
biologia alimentar da joaninha, Crenicchla lepidota da lagoa Redonda e Gurgel e Canan
(1999) que verificaram a dieta dos peixes da lagoa do Jiqui, RN. Mais recentemente, são
registrados os estudos de Raposo e Gurgel (2003), referindo-se a alimentação da piranha,
Serrasalmus spilopleura, em função do ciclo lunar e das estações do ano na lagoa de
Extremoz (Teixeira e Gurgel, 2004) e sobre a dinâmica da nutrição e do comportamento
alimentar de Steindachneria notonota, do açude Riacho da Cruz. Um estudo sobre a
ecologia alimentar e reprodutiva da piaba-do-rabo-amarelo, Astyanax cf. lacustris foi
realizado na lagoa do Piató, Assu, RN (SILVA, 2008).
1.3 - Considerações gerais sobre a espécie em estudo
O mussum, Synbranchus marmoratus, Bloch 1795, é um peixe teleósteo,
pertencente à família Synbranchidae, e ordem Synbranchiformes (LO NOSTRO, 2000).
Esses peixes habitam tocas nas lagoas e margens de rios. É um peixe de hábitos
crepusculares, noturnos e vive em covas que ele mesmo faz nas margens dos corpos d’água
de onde habita.
A ordem compreende três famílias, com 12 gêneros e cerca de 87 espécies. São
predominantemente dulciaquícolas, com apenas três espécies marinhas. Dos quatro

6
gêneros pertencentes à família Synbranchidae, Synbranchus spp, apresenta duas espécies,
S. marmoratus e S. madeira, ocorrem no continente americano, sendo encontrado desde o
sul do novo México estendendo-se pela América Central, até o sul da América do Sul.
Vivem principalmente em charcos, onde as águas são pobres em oxigênio e nos períodos
de seca, sobrevivem enterrados na lama (ROSEN e GREENWOOD, 1976; NELSON,
1994).
É comum encontrá-los nos ambientes lênticos e margens dos rios do nordeste do
Brasil, sendo mais facilmente capturados com covos. Zaret e Rand (1971) referenciam seu
comportamento como sendo carnívoro, predando peixes e sapos. S. marmoratus são
peixes de corpo alongado, sem nadadeiras pélvicas e abertura branquial localizada na
metade inferior do corpo (LAUDER e LIEM, 1983; BRITSKI, et al., 1999).
Os membros de Synbranchidae não possuem bexiga natatória, a pele é bastante
grossa e produz muco em quantidade que recobre todo seu corpo e a faz muito resistente a
infecções e a vida em cativeiro (GRAHAM et al., 1995).
S. marmoratus possui respiração aérea acessória, inspiram o ar através da câmara
branquial única que se abre na superfície ventral da cabeça quando a disponibilidade de
oxigênio diminui, apresentando assim, respiração aérea facultativa utilizando-se de sua
câmara buco-faríngea modificada, podendo viver tanto em águas pobres como ricas em
oxigênio No inicio da estação da seca, quando o nível da água declina constroem um
“túnel” no sedimento onde se enterram (ROBINS et al., 1991; KENNY, 1995; ESTEVES
1988).
Com base nessas informações e na necessidade imprescindível do conhecimento da
biologia de peixes do semi-árido do nordeste brasileiro, o presente trabalho teve como
objetivo complementar estudos já existentes sobre a ictiofauna em outras áreas do Estado
do Rio Grande do Norte, através da elucidação de aspectos alimentares de uma espécie de
peixe, nativa, encontrada no açude Marechal Dutra.

7
2. OBJETIVOS
CAPÍTULO 1

8
2. OBJETIVOS DA PESQUISA
2.1 - Objetivo Geral
Este trabalho teve como objetivo, verificar os aspectos da biologia alimentar e
morfohistologia do tubo digestório do mussum, Synbranchus marmoratus Bloch, 1795
(Osteichthyes: Synbranchidae), capturados no açude Marechal Dutra, RN do semi-árido
brasileiro a fim de fornecer subsídios para a conservação e uso sustentável desse recurso
pesqueiro da região.
2.2 - Objetivos específicos
Verificar as características morfométricas, a estrutura populacional em
comprimento e peso, o regime alimentar quanto ao grau de repleção (GR) e índice de
repleção estomacal (IR) da espécie em estudo, em relação aos períodos de estiagem e
chuvoso da região considerando os fatores ambientais.
(Artigo 1)
Caracterizar a morfohistologia do tubo digestório em relação ao regime
alimentar.
(Artigo 2)

9
CAPÍTULO 1
3. MATERIAIS E MÉTODOS

10
3. MATERIAL E MÉTODOS
3.1 - Área de estudo
O açude público Marechal Dutra, também denominado Gargalheiras, está
localizada nas coordenadas 6º 26’ 11’’ de Latitude Sul e 36º 38’ 28’’ de Longitude
Oeste, no distrito de Gargalheiras, município de Acari, RN, a 219 km de Natal (Figura
1). Compreende ao barramento do rio Acauã, formado pela confluência dos rios Currais
Novos (RN) e Picuí (PB), sendo um dos afluentes da bacia Piranhas-Assu. Sua
construção foi iniciada em 1909 e concluída em 1959. Com capacidade total de
acumulação de 40.000.000 m3 de água com uma profundidade máxima de 25 metros. A
profundidade média anual de 5,2 metros (Tabela 1 e Figura 2).
O açude Gargalheiras destaca-se na paisagem do semi-árido brasileiro como
núcleo de vida, sendo considerado a principal atração turística da cidade, sendo um
marco na história e na vida de Acari. Faz contraste com a paisagem seca e árida dos
sertões, proporcionando na época das chuvas, ao povo de Acari e aos turistas um grande
espetáculo na sua “sangria” sobre a parede da barragem. Teve sua construção iniciada
em 1909 e concluída em 1959. Na dependência dele vive hoje, uma população em torno
de 40 mil habitantes da cidade de Acari e Currais Novos.
A barragem Gargalheiras foi construída com a finalidade de armazenar água
para atender as necessidades da região, que pertence ao polígono das secas. Ela
apresenta o sétimo volume de água represada da bacia Piranhas-Açu. É um reservatório
de grande importância e tem como principal função o abastecimento público de água,
beneficiando a população tanto em termos armazenagem como para outros fins,
destacando-se a atividade pesqueira como principal fonte de renda da comunidade local,
seguida da exploração da agricultura de vazante.
A hidrologia do Município é caracterizada pelo Aqüífero Cristalino que, em
geral contém água de qualidade inferior para o consumo, em decorrência de seu teor
salino; e o Aquífero Aluvião, constituído pelo lençol freático depositado nos sedimentos
dos leitos e terraços de rios e riachos de grande porte, apresenta uma profundidade
média de 0,7m com água de boa qualidade. A vegetação local é de Caatinga
hiperxerófila, rala, de porte arbustivo e arbustivo-arbóreo, formado principalmente por
leguminosas e cactáceas.

11
O clima semi-árido, característico da região, apresenta um curto período
chuvoso e um longo período de estiagem, e ainda sendo marcado por constantes
irregularidades pluviométricas. O período chuvoso ocorre de janeiro a maio, mesmo nos
anos de menor pluviosidade. A precipitação anual média para a região é 430mm, a
máxima de 1.1337,6 mm e mínima de 18mm. A temperatura média mensal é de 27,4º
C, a média anual de umidade relativa do ar é de 65% e a insolação chega a 3.0000 h
/ano.
Tabela 1. Características hidrográficas e morfométricas do açude Gargalheiras, Acari,
RN.
Extensão da Bacia Hidrográfica 2.400 km2 Extensão da Bacia Hidráulica 780,00 há Capacidade máxima do açude 40.000.000mm3 Profundidade máxima do açude 25m Profundidade média do açude 5,2m
Fonte: SEMARH (Secretaria de Meio Ambiente e Recursos Hídricos do Rio Grande do Norte, 2007).
Figura 1. Área de estudo: Açude Marechal Dutra, Acari/RN, Brasil (6°24’06,87” Lat. S 36°35’07,23” Long. O) (Fonte: SECRETARIA DE RECURSOS HIDRICOS – SERHID, 2008).
Brasil
Rio Grande do Norte

12
Figura 2. Vista do açude Público Marechal Dutra: (a) período de estiagem, entre o agosto de 2008 a dezembro de 2008; (b) período chuvoso, entre janeiro de 2008 a julho de 2008.
a
b

13
3.2 - Coleta e taxonomia da espécie em estudo
Os membros de Synbranchidae são peixes serpentiformes, de corpo nu sem
escamas, não apresentam nadadeiras peitorais e pélvicas, as dorsais e anais são simples
pregas carnosas, sem raios e a caudal é pequena e vestigial ou ausente (NELSON 1994;
BRITSKI et al., 1999). Possui cabeça proeminente, com olhos pequenos e narinas
anteriores. A abertura branquial é única e situa-se sob a cabeça. A Figura 3 mostra a
distribuição geográfica e a espécie em estudo.
As coletas foram realizadas mensalmente por pescadores da região, no período
de agosto de 2007 a julho de 2008. Os exemplares de S. marmoratus foram capturados
de forma artesanal e também foram utilizados covos como apetrechos de pesca, que são
armadilhas confeccionadas em PVC, medindo cerca de 30cm de comprimento e 15cm
de diâmetro. Os covos são ligados uns aos outros por cordas (corda principal) e como é
projetado para não flutuar, descem ao substrato (Figura 4). Após as coletas, os peixes
foram acondicionados em caixas térmicas com gelo e transferidos ao Laboratório de
Ictiologia do Departamento de Oceanografia e Limnologia, UFRN, para a realização da
triagem e estudo.

14
Cla
sse:
Ost
eich
thye
s
Su
bcla
sse:
Act
inop
terig
ii
Div
isão
: Tel
eost
ei
Su
pero
rdem
: Aca
ntho
pthe
rygi
i
Ord
em: S
ynbr
anch
iform
es
Fa
míli
a: S
ynbr
anch
idae
Gên
ero:
Syn
bran
chus
Es
péci
e: S
ynbr
anch
us m
arm
orat
us
(FO
NTE
: NEL
SON
, 199
4)
Figu
ra 3
. Esp
écie
em
estu
do m
ussu
m, S
ynbr
anch
us m
arm
orat
us e
sua
dis
tribu
ição
geo
gráf
ica
(pon
tos v
erm
elho
s no
map
a).

15
Figura 4. Apetrechos de pesca: covos utilizados para captura do mussum, Synbranchus marmoratus no açude Marechal Dutra; a) covo aberto; b) covo fechado.
3.3 - Parâmetros Meteorológicos
Os dados de precipitação pluviométrica referentes ao período de estudo, foram
obtidos no Departamento de Metereologia e Recursos Hídricos da EMPARN (Empresa
de Pesquisas Agropecuárias do Rio Grande do Norte S/A), Natal/RN, para a
caracterização do regime pluviométrico da área de estudo.
3.4 - Caracterização morfométrica e biometria dos peixes
Os peixes foram numerados, pesados (g) e medidos (cm). A caracterização
morfométrica foi realizada baseando-se nas seguintes variáveis: Comprimento total
(LT), medição horizontal da ponta do focinho à maior extremidade da nadadeira caudal;
comprimento da cabeça (LC), distância da ponta do focinho à margem posterior do
opérculo; altura do corpo (AC), distância vertical, em ângulo reto com o eixo;
comprimento do focinho, ponta do focinho até o bordo posterior do opérculo;
a
b

16
comprimento do maxilar, a medida da porção inferior da boca; abertura da boca,
distância máxima entre as mandíbulas; comprimento pré-dorsal, comprimento desde o
extremo do focinho até nadadeira dorsal; comprimento pré-anal, comprimento desde o
extremo do focinho até o inicio da nadadeira anal e diâmetro do olho.
Após o registro desses dados, os peixes foram eviscerados com um
seccionamento na região abdominal a partir do poro urogenital para a retirada dos
órgãos foram verificados: peso corporal (Wt); peso corporal do peixe sem vísceras em
gramas (Wv) e peso do estômago (We). Os estômagos foram retirados na altura da
porção cárdica e pilorica, e obtido o peso em gramas. O grau de repleção do estômago
foi determinado de acordo com Santos (1978). Foram fixados em formol a 10% e
conservados em álcool a 70% para posterior leitura de itens alimentares.
Estrutura da população em peso e comprimento
A estrutura em comprimento e peso dos exemplares foram determinadas através
da distribuição das freqüências absolutas das classes de comprimento total e peso total
para sexo agrupado. A classe de intervalo foi de 5cm para o comprimento total, e
intervalo de 50g para o peso total dos espécimes.
3.5 - Aspectos alimentares
Grau de repleção:
A classificação do grau de enchimento do estômago (grau de repleção – GR) foi
estabelecida segundo a escala de Santos (1978), com adaptações de Gavilan-Leandro
(2003) variando de 0 a 3:
Onde:
0 = estômago vazio
1 = estômago parcialmente vazio (< 25% do volume ocupado)
2 = estômago parcialmente cheio (>25% e < 75% do volume ocupado)
3 = estômago totalmente cheio: repleto (> 75% e 100% do volume ocupado)

17
Índice de repleção estomacal (IR):
O Índice de Repleção estomacal foi calculado pela obtenção do peso do
estômago multiplicado por 100 e dividido pelo peso total do peixe (HAHN, 1991).
Variações nos valores de IR indicam uma maior ou menor atividade alimentar em
decorrência da disponibilidade ou escassez de recursos alimentares ao longo do ano. O
Índice de Repleção médio consiste na média ponderada dos valores numéricos do IR
considerado.
Onde:
We = Peso do estômago (g)
Wt = Massa corporal total do peixe (g)
3.6 - Análises da dieta
A identificação do conteúdo estomacal objetivou-se na descrição qualitativa e
quantitativa dos alimentos ingeridos pelo peixe. Alguns métodos foram combinados, para
que os erros fossem minimizados e o diagnóstico fosse o mais completo possível sobre a
dieta.
Após os procedimentos utilizados para investigação estomacal, comentados na
etapa de laboratório, os estômagos conservados foram retirados do álcool e preparados
para a análise. Os conteúdos estomacais foram examinados sob microscópio ótico e
estereomicroscópio. Procurou-se identificar até o menor nível taxonômico possível,
utilizando a bibliografia de apoio de Higuti e Franco (2001) e Wehr e Sheath (2003).
Para a investigação dos aspectos alimentares adotou-se o Método de Pontos
(SWYNNERTON e WORTHINGTON, 1940), Freqüência de Ocorrência
(HYNES,1950; HYSLOP, 1980), Volumétrico (HYSLOP, 1980), Índice de Importância
Alimentar estomacal (KAWAKAMI e VAZZOLER, 1980) e o hábito alimentar da
espécie (ROSECCHI e NOUAZE, 1987).
3.7 - Análise morfohistológica do tubo digestório
IR = We X 100 / Wt

18
Para a análise morfohistológica do tubo digestório, foi realizada a biometria e
dissecação dos animais, através de uma incisão no sentido longitudinal, partindo do poro
anal do animal até a cabeça. O tubo digestório foi retirado da cavidade celomática,
seguido de secções transversais do esôfago, estômago (porção cárdica, cecal e pilórica)
imediatamente fixados em Bouin a 10%, devidamente identificados.
Para análise anatômica dos órgãos digestórios foi utilizado um estereoscópio
(Olympus CH30). Dos órgãos digestórios foram retirados fragementos, que faziam parte
das regiões esofágica, cárdica, fúndica e pilórica estomacais e do intestino anterior,
médio e posterior. Em seguida efetuou-se a técnica histológica para microscopia óptica
desidratação, diafanização e inclusão em parafina, coradas com HE – Hematoxilina de
Harris, Eosina (MAIA, 1979) e PAS – e Ácido Periódico-Schiff (BEHMER et al., 1976)
com algumas modificações.
Para a descrição morfológica, especial atenção foi dada a caracterização do
formato geral do corpo, posição e tamanho da boca, posição e tamanho dos olhos, tipo
de dentição, tipo de forma das brânquias e rastros branquiais. Foram consideradas
também as estruturas da mucosa, quanto ao tipo de epitélio e os tipos celulares que o
compõem, assim como uma possível ocorrência de glândulas na lâmina própria e a
presença ou ausência de muscular da mucosa, a caracterização da submucosa e
distribuição de células granulares e, ou glândulas nesta túnica.
3.8 - Análise Estatística
Os resultados da análise de frequência de ocorrência dos itens da dieta de S.
marmoratus e o preenchimento estomacal foram comparados, aplicando-se o teste do 2
para se identificar as diferenças estatisticamente significativas.
3.9 - Normalização das referências
Para a normalização das referências bibliográficas da parte geral da dissertação
foi empregada a recomendação da ABNT – Associação Brasileira de Normas Técnicas
(MENEZES & CARVALHO, 2006) e para os Artigos foram utilizadas as normas das
revistas específicas (Anexo).

19
CAPÍTULO 1
REFERÊNCIAS BIBLIOGRÁFICAS
AGOSTINHO, A. A.; GOMES, L.C. 1997. Reservatório de Segredo: Bases ecológicas
para o Manejo. Maringá: Eduem, 387 p.
ALMEIDA, R.G.; SOARES, L.H. & EUFRÁSIO, M.M. 1993.Lagoa do Piató: Peixes e
Pesca. Coleção Vale do Açu, Coleção Humanas Letras. Natal, 84p.
BEHMER, A.O.; TOLOSA, E.M.C.; F. NETO, A.G. 1976 Manual de técnicas para
histologia normal e patológica. São Paulo: Editora da Universidade de São Paulo. p 98-101.
BRITSKI, H. A., SILIMOM, K. Z. S., LOPES, B. S. 1999. Peixes do pantanal. Manual
de identificação. H. A. Britski (ed.). Brasília: EMBRAPA, SPI, 184p.
BUENO, R. M. X. & CHELLAPPA, S. 2003. Composição ictiofaunística de valor
comercial do açude público Marechal Dutra, Acari, RN. Anais do Congresso
Nordestino de Ecologia. 10., Olinda, PE. (CD-ROM).
BUENO, R. M. X. 2004. Biologia Reprodutiva do cascudo, Hypostomus pusarum
(Starks, 1913) (Osteichthyes: Loricariidae) do açude Marechal Dutra, Acari, Rio Grande
do Norte. Dissertação (Mestrado em Bioecologia Aquática), Universidade Federal do
Rio grande do Norte, Natal, RN. 137p.
CÂMARA, M.R. Ecologia e Reprodução do Tucunaré comum, Cichla monoculus
(Osteichthyes: Cichlidae) da Barragem de Campo Grande, RN. 1998. 79f. Dissertação
(Mestrado em Bioecologia Aquática) Departamento de Oceanografia e Limnologia,
Universidade Federal do Rio Grande do Norte, Natal. 1998.
CÂMARA, M.R. VERANI, J.R.; CHELAPPA, S. Descrição morfométrica, mesrística e
produção de muco do acará disco, Symphysodon discus Heckel, 1840 (Osteichthyes:
Cichlidae) in: Congresso Brasileiro de Zoologia, 24.; 2002, Curitiba. Associação
Brasileira de Zoologia, 2002b.v.1.p.319.

20
CANAN, B., GURGEL, H. B.; NASCIMENTO, R. S. S., BORGES, S. A. G. V. &
BARBIERI, G.1997. Avaliação da comunidade de sete espécies de peixes da lagoa Boa
Cicca, Nísia Floresta – RN. Revista Ceres, Viçosa, 44 (256): 604-616.
CHELLAPPA, N. T. & A. M. COSTA. 2003. Dominant and co-existing species of
Cyanobacteria from a eutrophicated reservoir of Rio Grande do Norte State, Brazil.
Acta Oecologica – International Journal of Ecology. 24, 3-10.
CHELLAPPA, N.T.; CÂMARA, M.R. & CHELLAPPA, N.T. 2003a. Ecology of
Cichla monoculus (Osteichthyes: Cichlidae) from a reservoir in the semi-arid region of
Brazil. Hydrobiologia, 504. P. 267-273.
CHELLAPPA, S.; CHELLAPPA, N. T.; SILVA, E.A.; HUNTINGFORD, F. A. &
BEVERIDGE, M. C. M. 1996. The diet of hybrid red tilapia, Oreochromis niloticus
(Linnaeus) x Oreochromis mossambicus (Peters) reared in the freshwater ponds of
Northeastern Brazil. Aquaculture Research, 27 (12), 945-952.
COSTA, M. A. M. 2000. Ecologia da Comunidade Fitoplanctônica da Barragem
Gargalheiras em Acari, RN. 2000.. Dissertação ((Mestrado em Bioecologia Aquática),
Universidade Federal do Rio grande do Norte, Natal. 107p.
ESTEVES, F.A. 1988. Fundamentos de Limnologia. Rio de Janeiro.
Interciência.FINEP. 575pp
FREIRE, A.G.; ALMEIDA, R.G.; MENESCAL, R.; MEDEIROS, P.B.; SILVA, M. M.
2000. Caracterização ictiofaunistica dos grandes açudes do semi-árido do Rio Grande
do Norte. Coleção Mossoroense. Mossoró, RN.;série C, v. 1135, p. 269-275.
GAVILAN-LEANDRO. S. A. C. 2003. Variação temporal da freqüência de captura e
do comportamento alimentar de Astyanax bimaculatus Linnaeus, 1758 (Characidae,
Tetragonopterinae) do Rio Apodi-Mossoró, Rio Grande do Norte. Tese de Doutorado.
UFRN: Natal.

21
GRAHAM, J. B., LAI, N.C.,CHILLER, D. & ROBERTS J. L. 1995. The transition to
air breathing in fishes. Comparative aspects of cardio respiratory regulation in
Synbranchus marmoratus and Monopterus albus (Synbranchidae). J. Exp. Biol., v. 198,
pp. 1455 – 1467.
GURGEL, H. C. B. & CANAN, B. 1999. Feeding habits of seven fish species from the
semi-arid region of Rio Grande do Norte, Brasil. Acta Scientiarum 21 (2): 243-246.
GURGEL, H. C. B., B.W. F. MOLINA, L.J. SOARES & B. CANAN. 1995. Ictiofauna
de semi-árido do Rio Grande do Norte. P. 010-011. In: Resumos do XI Encontro
Brasileiro de Ictiologia. Campinas, Pontifícia Universidade Católica de Campinas/
Sociedade Brasileira de Ictiologia.
GURGEL, H. C. B.; ALMEIDA, R. G.; BARBIERI, G.;1994. Análise qualitativa da
alimentação e o coeficiente intestinal de Metynnis cf. Roosevelt Eigenmann, 1915
(Characidae, Myleinae), da lagoa Redonda, Nísia Floresta, Rio Grande do Norte, Brasil.
Revista Brasileira Zoologia 11(2):331-339.
GURGEL, H.C.B.; ALMEIDA, R.G.; BARBIERI, G.& VIEIRA, L. J. S. 1998. Dieta de
Crenicichla lepidota Heckel, 1840 (Perciformes, Cichlidae) da lagoa Redonda, Nísia
Floresta/RN. Acta Scientiarum 20 (2): 191-194.
HAHN, N. S. 1991. Alimentação e dinâmica da nutrição da curvina Plagioscion
squamosissimus (Heckel, 1840) (Pisces, Perciformes) e aspectos da estruturas trófica da
ictiofauna acompanhante no rio Paraná. Tese de Doutorado. Rio Claro: UNESP. 497p.
HAHN, S. N., FUGI, R., ALMEIDA, V. L. L., RUSSO, M. R. & LOUREIRO, V.E.
1997. Dieta e atividade alimentar de peixes do reservatório de Segredo, p.141-162.in:
EDUEM. Reservatório do Segredo: bases ecológicas para o manejo. Maringá.
Universidade Estadual de Maringá. 387p.
HIGUTI, J. & FRANCO, G. M. S. 2001. Identificação de invertebrados para análise de
conteúdo estomacal de peixes. Núcleo de Pesquisas em Limnologia, Ictiologia e
Aqüicultura. UEM: Paraná, 110 p.

22
HYNES, H. B. N. 1950. The food of freshwater Sticklebacks (Gasterosteus aculeatus
and Pygosteus pungitius), with a review of methods and used in studies of the food of
fishes. Journal of Animal Ecology, 19 (1): 36-58 p.
HYSLOP, E. J. 1980. Stomach contents analysis - a review of methods and their
application. Journal of Fish Biology, 17 (4): 411-429 p.
KAWAKAMI, E. & VAZZOLER, G. 1980. Método gráfico e estimativa de índice
alimentar aplicado no estudo de alimentação de peixes. Boletim do Instituto
Oceanográfico, 29 (2): 205-207 p.
KENNY, J. S., 1995. Views from the bridge: a memoir on the freshwater fishes of
Trinidad. St Joseph, Trinidad and Tobago, v. 3, 98p.
LARKIN, P. A 1956. Interspecific competition and population control in freshwater
fish. J. Fish. Res.Bd.Can.; 22 (6): 1357 – 1377.
LAUDER, G. V.; LIEM, K. F. 1983. The evolution and interrelationships of the
actinopterygian fishes. Bull. Mus. Comp. Zoo. 150: 95-197.
LO NOSTRO, F. L. 2000. Espermatogénesis, ciclo anual e inducción de la
espermiación en el pez protogínico diándrico, Synbranchus marmoratus Bloch 1795
(Teleostei, Synbranchidae) Buenos Aires, 138p. Tesis (Doctoral) – Depto de Cs
Biologicas, Facultad de Ciências Exactas y Naturales, Universidade de Buenos Aires.
LOWE-MCCONNELL, R.H. 1999. Estudos Ecológicos de Peixes Tropicais. São Paulo:
Editora da Universidade de São Paulo, 536p.
MAIA, V. 1979 Técnica Histológica. 2. ed. São Paulo: Atheneu. p.70-136.
MAGALHÃES, E. M.; ALMEIDA, R. G.; GURGEL, H. C. B. & BARBIERI, G. 1990.
Contribuição ao estudo da alimentação de Serraasalmus brandtii Reinhardt, 1874
(Characidae, Serrasalmidae) do rio Piranhas-açu, pendências, Rio Grande do Norte.
Revista Ceres 37(213):429-442.

23
MEDEIROS, A. P. T. 2001. Ecologia, diversidade e aspectos reprodutivos da ictiofauna
de valor comercial da lagoa de Extremoz, RN. Dissertação (Mestrado em Bioecologia
Aquática), Universidade Federal do Rio grande do Norte, Natal, RN. 75p.
MEDEIROS, A. P. T.; CHELLAPPA, N.T. & CHELLAPPA, S. 2003. Aspectos
reprodutivos do cangati, Parauchenipterus galeatus, Linnaeus, 1766 (Osteichthyes:
Auchenipteridae) da Lagoa de Extremoz, Rio Grande do Norte. Revista Brasileira de
Zoologia. 20 (4), 647-650.
MEDEIROS, A. P. T.; CHELLAPPA, S. & CHELLAPPA, N.T. 2004. Período
reprodutivo da pirambeba, Serrasalmus maculatus Kner, 1858 (Osteichthyes:
Characidae) na Lagoa de Extremoz, Rio Grando do Norte. p. 89 – 95. In: Ecologia
Aquática Tropical Ed. N. T. Chellappa , S. Chellappa & J.Z.O. Passavante. Editora:
Editora Serv-Graf, Natal / RN. 165p.
MENESCAL, R.A. 2002. Efeitos da Introdução da Tilápia do Nilo, Oreochromis
Niloticus, (Linnaeus, 1758) (Perciformes, Cichlidae) Sobre o Desembarque Pesqueiro
no Açude Marechal Dutra, Acari/RN. 2002. 49f. Dissertação (Mestrado em Bioecológia
Aquática) – Departamento de Oceanografia e Limnologia, Universidade Federal do Rio
Grande do Norte, Natal.
MENEZES, M.R.L. & CARVALHO, E.G. 2006. Referências Bibliográficas NBR 6023,
2. ed., Natal: EDUFRN, 60p.
MENEZES, N.A. 1994. Importância da Conservação da Ictiofauna dos Ecossistemas
aquáticos brasileiros in Seminário sobre fauna aquática e o setor elétrico brasileiro,
reuniões temáticas prepartórias: caderno 3 – conservação, Curitiba, 23-24 nov.; 1993/
Comitê Coordenador das Atividades do Meio Ambiente do Setor Elétrico – COMASE -
Rio de Janeiro: ELETROBRÁS, 40p.
MENEZES, R. S. & MENEZES, M. F. 1946. Notas sobre o regime alimentar de
algumas espécies ictiológicas de água doce do Nordeste. Revista Brasileira Biologia
6(4): 537-542.

24
NASCIMENTO, M. M. 2006. Biologia reprodutiva do curimatã comum, Prochilodus
brevis Steindachner, 1875 e limnologia do açude Marechal Dutra localizada na Caatinga
do Rio Grande do Norte. Dissertação (Mestrado em Ciências Biológicas), Universidade
Federal do Rio grande do Norte, Natal, RN. 85p.
NELSON, J. S. 1994. Fishes of the World. John Wiley & Sons,New York, 600p.
NIKOLSKY, G.V. 1963. The ecology of fishes. Londres: Academic Press.
RAPOSO, R. M. G. & GURGEL, H. C. B. 2003. Variação da alimentação natural de
Serrasalmus spilopleura Kner, 1860 (Pisces, Serrasalmidae) em função do ciclo lunar e
das estações do ano na lagoa de extremoz, Rio Grande do Norte, Brasil. Acta
Scientiarum. Animal Sciences 25(2): 267 – 272.
ROBINS C. R.; BAILEY R. M.; BOND C. E.; BROOKER J. R.; LACHNER E. A.;
LEA R.N.; SCOTT W. B. 1991. World fishes important to North Americans.
Exclusive of species from the continental waters of the United States and Canada. Am.
Fish. Soc. Spec.Publ., v 243, pp. 91.
ROSA, R. S., MENEZES, N.A., BRITSKI, H.A., COSTA, W.J.E.M. & GROTH, F.
2005. Diversidade, padrões de distribuição e conservação dos peixes da Caatinga. P.
135 – 180. In: I.R. Leal, M. Tabarelli & J.M.C. da Silva (Eds.) Ecologia e Conservação
da Caatinga. Editora Universitária da Universidade Federal de Pernambuco, Recife.
822p.
ROSA, R.S. 2004. Diversidade e conservação dos peixes da Caatinga. In: J.M.C. da
Silva, M. Tabarelli, M.T. Fonseca, L.V. Lins (Orgs.) Biodiversidade da Caatinga: Áreas
e Ações Prioritárias para a Conservação. Ministério do Meio Ambiente, Brasília-DF.
ROSECCHI, E. & NOUAZE, Y. 1987. Comparaison de cinq índices alimentaires
utilisés dans I’analyse des contenus stomacaus. Revista Trav. Int. de Peches Marit.,
Nantes. Vol. 49. n. 3-4, 111-123 p.

25
ROSEN, D. E.; GREENWOOD P. H. 1976. A fourth neotropical species of
Synbranchidae eel and phylogeny and systematics of fishes. Bull. Am. Mus. Nat. Hist.
157:1-69.
SANTOS, E. P. 1978. Dinâmica de populações aplicada à pesca e piscicultura. São
Paulo: Hucitec: EDUSP, 129 p.
SILVA, D. A. 2008. Ecologia alimentar e reprodutiva da piaba-do-rabo-amarelo,
Astyanax cf. lacustris, (Reinhardt, 1874) (Osteichthyes: Characidae) na Lagoa do Piató,
Assu, RN, Brasil. Dissertação (Mestrado em Bioecologia Aquática), Universidade
Federal do Rio grande do Norte, Natal, RN. 106p.
SILVA, M.M. 2000. Desembarque Pesqueiro das Principais Espécies da Ictiofauna da
Barragem Marechal Dutra, Acari/ RN. 2000. Dissertação (Mestrado em Bioecologia
Aquática), Departamento de Oceanografia e Limnologia, Universidade Federal do Rio
Grande do Norte, Natal. 120p.
SWYNNERTON, G. H.; WORTHINGTON, E. B. 1940. Notes on the food of fish in
Haweswater (Westmorland). Journal of Animal Ecology, 9: 183-187 p.
TEIXEIRA, J. L. A.; GURGEL, H. C .B. 2004. Dinâmica da nutrição e alimentação
natural de Steindacnerina notonota (Miranda-Ribeiro, 1937) (Pisces, Curimatidae),
Açude de Riacho da Cruz, Brasil, Brasil. Brasil. Ver. Bras. Zoociências. 6 (1): 19-28.
WEHR, J. D. & SHEATH, R. G. 2003. Freshwater Algae of North America: Ecology
and Classification. Elsevier Science: California, USA, 918 p.
ZARET, T.M. & RAND, A. S. 1971. Competition in tropical stream fishes. Ecology, 52
(2):336-342.
ZAVALA-CAMIN, L. 1996. A. Introdução aos estudos sobre alimentação natural em
peixes. Maringá, EDUEM.129 p.

26
CAPÍTULO 2
RESULTADOS Artigo Científico 1

27
4. RESULTADOS
4.1 Os resultados incluem dois artigos científicos em preparação a serem
submetidos à publicação.
ARTIGO 1 – ARTIGO EM PREPARAÇÃO
Luciana Araújo Montenegro; Daisy Nazareth Ferreira Damasceno & Sathyabama Chellappa
Biologia Alimentar do Mussum, Synbranchus marmoratus, Bloch
(Osteichthyes: Synbranchidae) em um Açude do Semi-Árido Brasileiro .
Freshwater Biology
ISSN: 0046-5070
Factor de impacto: 2,650
ARTIGO 2 – ARTIGO EM PREPARAÇÃO
Morfohistologia do tubo digestório do mussum, Synbranchus marmoratus,
Bloch (Osteichthyes: Synbranchidae) em um Açude do Semi-Árido Brasileiro
Luciana Araújo Montenegro; Naissandra Bezerra da Silva, Daisy Nazareth Ferreira Damasceno & Sathyabama Chellappa
Journal of Fish Biology
ISSN: 0022-1112 Factor de impacto: 1,404

28
BIOLOGIA ALIMENTAR DO MUSSUM, SYNBRANCHUS
MARMORATUS, BLOCH (OSTEICHTHYES: SYNBRANCHIDAE)
EM UM AÇUDE DO SEMI-ÁRIDO BRASILEIRO
Luciana Araújo Montenegro1; Daisy Nazareth Ferreira Damasceno2 & Sathyabama
Chellappa 1
1 UFRN, Programa de Pós-Graduação em Ciências Biológicas, Centro de Biociências,
Departamento de Oceanografia e Limnologia, Praia de Mãe Luíza, s/n, 59014-100,
Natal, Rio Grande do Norte, Brasil. E-mail: [email protected],
2 UFRN, Departamento de Oceanografia e Limnologia, Centro de Biociências, s/n,
59014-100, Natal, Rio Grande do Norte, Brasil. E-mail: [email protected]
RESUMO
O presente trabalho baseia-se na análise do conteúdo estomacal do mussum, Symbranchus marmoratus, capturados mensalmente utilizando covos no período de agosto de 2007 a julho de 2008, no açude Marechal Dutra, Acari, Rio Grande do Norte, Brasil. Foram obtidos de cada exemplar o comprimento total, massa corporal e peso do estômago. A análise do conteúdo estomacal dos peixes foi realizada de acordo com o método volumétrico, de pontos e o método de freqüência de ocorrência, aplicando-se sobre este o Índice de Importância Alimentar. As variações na dieta e a freqüência de ingestão alimentar foram relacionadas com a pluviosidade. Os itens alimentares identificados foram separados em grupos distintos. No período de seca a espécie apresentou valores mais altos no índice médio de repleção, com um pico no mês de setembro (4,54 ± SD = 0,56). O menor valor (3,99 ± SD = 0,25) ocorreu no mês de maio no período chuvoso. Os resultados obtidos indicam que S. marmoratus se alimentou preferencialmente de material animal, sendo 78,22% de crustáceos, 13,5% de material orgânico semi digerido, 3,25% de moluscos, 2,85% de peixes e 1,4% de insetos. Portanto, a espécie em estudo pode ser caracterizada como carnívoro com preferência a carcinofagia com a maior atividade alimentar durante a estiagem. Palavras Chave: dieta, peixe, açudes, carcinofagia, semi-árido.

29
ABSTRACT
This work refers to stomach content analysis of the marbled swamp eel, Symbranchus marmoratus (Block, 1917), captured using traps on a monthly basis during the period of August, 2007 to July, 2008 in Marechal Dutra dam, Acari, Rio Grande do Norte State, Brazil. Each individual was measured and their total length, body mass and stomach weights were registered. The stomach content analysis was performed according to volumetric points, frequency of occurrence methods and the dietary importance index. The variations in diet and frequency of feeding were related to rainfall. The food items identified were separated in groups. During the dry season the study species showed higher values of fullness, with a peak value in September (4.54 + SD = 0.56). The lowest value (3,99 + SD = 0,25) was in May, during the rainy season. The stomach contents of S. marmoratus show that this fish prefers animals, 78.22% of crustaceans 2.85% of mollusks, 3.25% of fish, 1.4% of insects and 13.5% of semi-digested organic matter, thus characterizing the study species as a carnivore with a preference for crustaceans, with elevated feeding activity during the dry season. Key words: diet, fish, reservoir, carnivore, semi-arid.

30
INTRODUÇÃO
O conhecimento da alimentação de peixes em meio natural é indispensável á
compreensão de sua biologia e ecologia, permitindo explicar variações de crescimento,
certos aspectos da reprodução, migração e certos comportamentos de procura e captura
do item alimentar. As variações bióticas e abióticas podem levar a uma mudança nos
itens que compõem a dieta dos peixes, sugerindo que a maioria pode se utilizar uma
ampla gama de alimentos, e quando um destes itens encontra-se em proporções
diferentes do normal, em escassez ou em excesso, os peixes mudam seu comportamento
alimentar.
O mussum, Synbranchus marmoratus, Bloch 1795, é um peixe teleósteo,
pertencente a Família Synbranchidae, e ordem Synbranchiformes. Estudos ecológicos
sobre a alimentação de S.marmoratus fornecem informações relevantes sobre sua
estratégia de vida, interações com outros organismos do ecossistema e nos permitem
verificar variações na atividade alimentar em função de fatores ambientais. Além de
servir como suporte para a elaboração de um plano adequado de manejo, objetivando
um uso sustentável dos estoques pesqueiros e conservação das espécies nativas. Deve
ser enfatizado que a alimentação não é apenas um resultado das relações ecológicas dos
organismos num dado período de suas vidas, mas é também um reflexo de eventos
anteriores e preparação para a estação seguinte, se constituindo numa forma de
comportamento e que a força dos fatores ambientais e o significado biológico desses
mesmos fatores, alteram a velocidade das respostas comportamentais (MANTEIFEL et
al. 1978).
Embora a espécie S. marmoratus não seja uma espécie de grande interesse
comercial, a sua abundância em áreas dulciaquícolas do semi-árido nordestino, faz com
que seja utilizada pelas comunidades pesqueiras, como complemento alimentar. Nesse
sentido, levando-se em consideração a pequena quantidade de artigos publicados
referentes a alimentação do S. marmoratus a presente pesquisa visa contribuir para a
caracterização do padrão sazonal e temporal da dieta da espécie, atribuindo a
importância de cada item na alimentação da espécie, relacionando a atividade alimentar
com parâmetros meteorológicos e físico-químicos, tais como pluviosidade, temperatura,
condutividade elétrica e oxigênio dissolvido, verificando as variações na intensidade
alimentar em função das variáveis ambientais.

31
MATERIAIS E MÉTODOS
Área de estudo
A amostragem do estudo foi realizada no açude Marechal Dutra (6º 26’ 11’’ de
Lat. Sul e 36º 38’ 28’’ de Long. Oeste), situada no município de Acari, Estado do Rio
Grande do Norte. O açude está inserido no bioma Caatinga, possui uma capacidade de
acumulação de 40.000.000 m3, profundidade máxima de 25 metros e média anual de 5,2
metros localizada a 219km da capital do Estado. O açude Gargalheiras destaca-se na
paisagem do semi-árido brasileiro como núcleo de vida. Na dependência dele vive hoje,
uma população em torno de 40 mil habitantes da cidade de Acari e Currais Novos. Esse
reservatório vem sendo utilizado pela população para outros fins, destacando-se a
atividade pesqueira como principal fonte de renda da comunidade local, seguida da
exploração da agricultura de vazante.
Coleta do material
As coletas foram realizadas numa freqüência mensal do período de agosto de 2007 a
julho de 2008 utilizando-se covos como apetrechos de captura, fixados próximos à
vegetação marginal. Os exemplares capturados foram etiquetados e transportados ao
laboratório para o registro da biometria. Para cada exemplar foi obtida a massa corporal em
gramas e o peso do estômago, através do emprego de balança digital com precisão de
0,0001g. O comprimento total (uma medida entre a extremidade anterior da maxila e a
extremidade do lobo superior da nadadeira caudal, levemente distendida) foi expresso em
centímetros.
Pluviosidade
Os dados de precipitação pluviométrica referentes ao período de estudo foram
obtidos no Departamento de Metereologia e Recursos Hídricos da EMPARN (Empresa
de Pesquisas Agropecuárias do Rio Grande do Norte S/A) Natal/RN, para a
caracterização do regime pluviométrico da área de estudo.

32
Análise dos itens alimentares
Após as biometrias, os peixes foram eviscerados, com um seccionamento na
região abdominal a partir do poro urogenital para a retirada dos estômagos. Os
estômagos fixados no formol 10% e conservados no álcool 70% tiveram o conteúdo
examinado segundo o Método de Pontos (SWYNNERTON & WORTHINGTON,
1940), Freqüência de Ocorrência (HYNES, 1950; HYSLOP, 1980) e Volumétrico
(HYSLOP, 1980). O Índice de Repleção estomacal (IR) foi calculado com base na
massa corporal do peixe dividida pelo peso do estômago e multiplicada por cem. Os
valores médios mensais do IR foram comparados com fatores abióticos (pluviosidade) e
limnológicos (temperatura, condutividade elétrica e oxigênio dissolvido) relacionando-
os com as variações na atividade alimentar. Os itens alimentares identificados foram
separados em grupos distintos de acordo com a sua origem, visando à determinação do
hábito alimentar e aplicando o Índice de Importância Alimentar para cada item
consumido.
Análise estatística
Através do Software XL STAT foi realizada a Análise de Componentes
Principais – PCA, corroborada pelo teste de Spearman, com o intuito de verificar quais
das variáveis ambientais mais se relacionavam com a atividade alimentar.

33
RESULTADOS
A análise do conteúdo estomacal do S. marmoratus revelou que o peixe se
alimentou exclusivamente de itens de origem animal, sendo portanto considerado como
espécie carnívora. A dieta alimentar foi composta de itens separados em categorias
distintas. Os itens foram compostos de crustáceos, peixes, moluscos, nemátodos, restos
de inseto. Foram encontrados também escamas de peixe, exoesqueletos de crustáceos e
grande quantidade de material semi digerido.
→INSERIR Tab.1
O Índice de Importância alimentar (IAi) demonstrou que S. marmoratus se
alimentou preferencialmente de material animal, sendo (78,22%) de crustáceos, (13,5%)
de material orgânico semi digerido e (3,25%) de moluscos, (2,85%) de peixes, (1,4%)
de restos de insetos. Caracterizando a espécie em estudo como um organismo carnívoro
com tendência generalista (Fig. 1).
→INSERIR Fig. 1
A análise do Índice médio de Repleção estomacal (IR) demonstrou que os meses
de agosto a dezembro de 2007, correspondente ao período de estiagem, apresentaram os
valores mais altos para o período estudado. Houve uma diminuição acentuada do IR nos
meses de janeiro a junho, Houve maior atividade alimentar durante a estiagem e uma
diminuição no período chuvoso.
A precipitação pluviométrica variou de 0 mm a 335 mm. Durante o período
chuvoso a precipitação pluviométrica foi maior, com uma acentuada diminuição de
chuvas durante a estiagem. A média anual da pluviosidade foi de (71,62 mm ± SD
163,3). O maior valor ocorreu em março de 2008: 335,5 mm e os menores nos meses de
agosto a dezembro de 2007, nos quais não houve precipitação. A relação entre a
pluviosidade e o Índice médio de Repleção indica que nos meses de maior precipitação
os peixes ingeriram menos alimento, correspondente aos meses de janeiro a julho de
2008. No período em que há uma diminuição acentuada de chuvas, a espécie estudada

34
apresentou valores mais altos no Índice médio de Repleção, com um pico no mês de
setembro (média = 4,54; ± SD = 0,56), sendo esse o maior valor médio de IR anual. O
menor valor (média = 3,99; ± SD = 0,25) ocorreu no mês de maio, período chuvoso. A
atividade alimentar mostrou-se mais intensa durante os meses de baixa precipitação
pluviométrica (Fig. 2).
→INSERIR Fig. 2

35
DISCUSSÃO
Mudanças sazonais ocorridas no habitat afetam os peixes, principalmente através
de mudanças na quantidade e qualidade do alimento disponível. Em águas tropicais, a
despeito das especializações anatômicas para a alimentação, exibidas por muitas
espécies de peixes, a maioria delas mostra considerável plasticidade em suas dietas
(LOWE-MCCONNELL, 1987). A diversidade dos itens que constituem a dieta de S.
marmoratus nos permite inferir que esta espécie retrata bem o observado pela autora,
refletindo a característica oportunista da espécie.
No entanto, em S. marmoratus, ocorreu somente variação sazonal na quantidade
de alimento, estando a maioria dos exemplares com estômagos vazios no inverno e
alcançando o valor máximo de repleção no verão. Os crustáceos se mostram ser o
principal alimento desta espécie alcançou maior proporção em todas as estações. Esta
espécie foi carnívora em todas as estações, alimentando-se de crustáceos e peixes.
Contudo, não é o verão a estação em que os peixes estão mais bem nutridos. Aliás, estes
adquirem massa corporal no auge do inverno, precisamente quando se nota os valores
mais baixos do índice de repleção (IR).
Este fenômeno talvez possa ser explicado por alterações metabólicas, que
aumentam a conversão alimentar, relacionadas talvez com a reprodução.
Segundo Hyslop (1980), variações no peso médio dos conteúdos estomacais
durante o ano, indicam diferenças na intensidade da alimentação. A análise do conteúdo
estomacal de S. marmoratus evidenciou basicamente crustáceos ocorrendo em 80% dos
estômagos analisados durante o período de estiagem e de chuva, correspondendo a
42,5% do volume total de itens encontrados na análise da dieta alimentar.
Entretanto, diversos autores previnem o uso do método de freqüência de
ocorrência, com um índice isolado do “valor alimentar” (CRISP et al., 1978; HYNES,
1950; MANN, 1973; MANN & ORR, 1969). Estimadores numéricos superestimam a
importância de pequenas presas tomadas em grandes números. Nesta situação, a
percentagem de ocorrência encontrada em nossos estudos, foi associada a composição
volumétrica, no sentido de fornecer com certa confiança uma indicação da dieta
conforme sugerido por (CRISP et al., 1978; BONNEAU et al., 1972).
O índice de importância alimentar encontrado para os 42 estômagos analisados,
de S. marmoratus mostrou crustáceos (IAi=1,4), detritos(IAi = 0,34) Peixes (IAi

36
=0,09), restos de insetos (IAi=0,08), Moluscos (IAi=0,07), nemátoda (IAi =0,005). Os
resultados deste trabalho são semelhantes aos descritos por Soares et al. (1986) referente
a estrutura trófica do lago Camaleão no Amazonas onde os autores encontraram para
esta espécie, um espectro alimentar constituído basicamente de insetos, ácaros,
crustáceos e peixes, classificando-o como carnívoro. A dieta alimentar do mussum,
pode ser carnívora na fase adulta, ingerindo peixes, moluscos e crustáceos e
alimentando-se durante sua etapa juvenil, de zooplancton, pequenos crustáceos
(Cladócera e Ostacoda) e insetos associados à vegetação flutuante (LO NOSTRO,
1986).
Gerking (1994) relatou que um grande número de espécies de peixes tem
habilidade de mudar de dieta e de hábitos alimentares, respondendo a variações sazonais
e diárias, entre outras, na disponibilidade de alimento, o que cria dificuldades para
classificar os peixes em grupos tróficos. No entanto a esse despeito Caragitsou &
Papaconstatinou (1990) ressaltam que, de acordo com o alimento disponível no
ambiente, o peixe pode mudar tanto seu comportamento quanto seu padrão de migração.
Ainda conforme Larkin (1956), a competição caracteriza-se pela demanda de mais de
um tipo de organismo pelos mesmos recursos do meio ambiente para o suprimento de
suas necessidades imediatas, podendo ser causada por alimentação comum, por espaço,
ou por predação entre competidores.
A atividade alimentar possa ser influenciada pela variação do nível da água em
rios que possuem regime marcante de cheia e seco. O açude Marechal Dutra, durante o
período (agosto/2007 a julho 2008) apresentou variações marcantes no nível da água,
ocorrendo estiagem no período de agosto a dezembro que levou o nível do rio a 4
metros abaixo do considerado normal.
Alguns autores observaram variação sazonal na alimentação de peixes (Zaret &
Rand,1971; Guilen & Har, 1980; Laughlin & Werner, 1980; Wynes & Wissing, 1982),
sugerindo que isto pode mostra o ciclo de vida da presa ou o resultado de competição
pelo mesmo alimento. Nesse sentido, observou-se no conteúdo estomacal de S.
marmoratus uma maior incidência do item crustáceos durante os meses de estiagem,
onde a captura torna-se máxima, quando os camarões migram para as margens e mínima
durante o período de chuva, devido a velocidade muito alta da correnteza da água, que
diminui a capturabilidade das espécies.
O fato dos peixes diminuírem a alimentação em período de precipitação
pluviométrica pode não estar relacionado à disponibilidade de recursos e sim ao período

37
reprodutivo, que no nordeste brasileiro geralmente ocorre no período chuvoso. Dessa
forma eles teriam mais espaço para o desenvolvimento das gônadas. No açude Marechal
Dutra a análise efetuada por estação estiagem e chuvosa, mostra que os maiores valores
para o índice de repleção ocorreram durante o período de agosto de 2007 a dezembro de
2007. Essas variações sugerem que S.marmoratus possui maior atividade alimentar, nos
meses de menor precipitação pluviométrica, ou seja, na estação seca. Neste sentido,
Welcomme (1995), assinala que a intensidade alimentar em águas tropicais está sujeita
a poucas influências de temperatura, relacionando o regime das cheias ao suprimento
alimentar e a densidade populacional como determinantes da atividade alimentar, uma
vez que as chuvas ampliam os biótopos, favorecendo a atividade alimentar.
Para Goulding et al. (1984) a dieta de uma espécie de peixe, só pode ser
compreendida se o comportamento alimentar for estudado nos diversos tipos de água
que a espécie habita. Segundo estudos realizados por estes autores, o camarão é o
principal alimento do Sorubim lima em várzea de águas brancas, e sendo uma espécie
abundante em rios, com poucos exemplares capturados em águas pretas e claras.
Portanto, relacionaram a abundância de S.lima com a de camarões menor em rios de
águas pretas e claras do que em rios de água branca.
Em ambientes tropicais dulcícolas, as flutuações pluviométricas destacam-se
entre os fatores abióticos por apresentar um papel fundamental, pois provocam o
carreamento de nutrientes do solo, aumentando a disponibilidade de alimento e
enriquecendo os ecossistemas aquáticos, elevando as concentrações de oxigênio e
regulando as atividades especialmente reprodutivas, dos organismos, inclusive
fornecendo ambientes para nidificação e alimentação das larvas, em decorrência da
ampliação das margens. No açude Marechal Dutra os fatores abióticos exerceram
considerável influencia na alimentação dos peixes. De acordo com a análise dos
alimentos principais verificou-se que entre os fatores físico-químicos, a pluviosidade e o
oxigênio dissolvido foram os que mais interferiram. O oxigênio dissolvido apresenta
uma relação direta com a atividade alimentar tanto no período de chuva quanto no
período de estiagem, talvez por ser um fator limitante na distribuição dos organismos na
coluna d’água e na distribuição destes. A temperatura do açude não demonstrou grandes
variações ao longo do estudo e sua relação com a atividade alimentar foi constatada em
setembro e outubro, os meses de temperatura mais elevada durante a estiagem. De
acordo com Esteves (1996), em águas frias ou com a diminuição acentuada da
temperatura, pode haver diminuição no número de presas, o reservatório torna-se mais

38
abundante em ofertas de itens alimentares durante as estações quentes. Nesse sentido
pode-se inferir que a elevada atividade alimentar no açude Marechal Dutra, durante a
estação seca do que na chuvosa poderia ainda ser explicada pela disponibilidade dos
componentes na dieta, visto que apesar das chuvas aumentarem o aporte de material
trazido pelos rios e riachos que deságuam no açude Marechal Dutra, a elevada
movimentação da água dificulta sua disponibilidade na coluna d’água, não favorecendo
sua ingestão. A condutividade elétrica durante todo o período de estudo manteve-se alta
e apresentou influencia na atividade alimentar dos peixes durante o período de janeiro a
julho, correspondente ao período de estiagem. A alta condutividade elétrica é
característica dos reservatórios do Nordeste (CHELLAPPA et al., 2003).
Pelo exposto conclui-se que a espécie apresenta um maior índice de repleção
durante o período de estiagem. O padrão dos itens alimentares encontrados na dieta de
S. marmoratus, foi composto exclusivamente de itens de origem animal, principalmente
crustáceos, o que caracteriza a espécie como carnívora, com tendência a carcinofagia e
habito alimentar generalista.
Os autores agradecem ao CNPq pela concessão de suporte financeiro a pesquisa.

39
REFERÊNCIAS
BONNEAU, D.L., McGUIRE, J.W.; TIEMEIER, O.W. & DEYONE, C.W. 1972. Food
habits and growth of channel catfish fry Ictularus punctatua. Trans.Am.Fish.Soc. 101, 613-
614.
CHELLAPPA, S.; CÂMARA, M. R. & CHELLAPPA, N. T. 2003. Ecology of Cichla
monoculus (Osteichthyes: Cichlidae) from a reservoir in the semi-arid region of Brazil.
Hydrobiologia, 504: 267-273 p.
CHELLAPPA, S.; CHELLAPPA, N. T.; SILVA, E.A.; HUNTINGFORD, F. A. &
BEVERIDGE, M. C. M. The diet of hybrid red tilapia, Oreochromis niloticus
(Linnaeus) x Oreochromis mossambicus (Peters) reared in the freshwater ponds of
Northeastern Brazil. Aquaculture Research, 27 (12), 945-952, 1996.
CARAGITSOU, E.; PAPACONSTATINOU. Food and feeding habits of large scale
gurnard, Lepidotrigla cavilloneI (Triglidae) in Grrek seas. Cybium, Paris, v.14, n.2,
p.95-100, 1990.
CRISP, D.T.; MANN, R.H.K. & McCORMACK, J.C. (1978). The effects of
impoundment and regulation upon the stomach contents of fish at Cow Green, Upper
Teesdale. J. Fish Biol. 12, 287-301.
ESTEVES, K. E. 1996. Feeding ecology of three Astyanax species (Characidae,
Tetragonopterinae) from a floodplain lake of Mogi-Guaçú River, Paraná River Basin,
Brazil. Environmental Biology of Fishes. 83-101 p.
GERKING, S.D. Feeding ecology of fishes. San Diego: Academic Press, 1994.
GOULDING, M.; CARVALHO, M. L.; FERREIRA, E. J. G. 1988. Rio Negro. Rich
life in poorwater. SPB Academic Publishing, The Hague. 200 p.

40
GUILLEN, A.L.& T.HART.1980. Feeding interrelationships between the sand shiner
and the striped shiner. Ohio j. Sci. 80 (2):71-76.
HYNES, H.B.N. 1950. The food of freshwater stickle back (Gasterosteus aculeatus and
Pungosteus pungitius) with a review of methods used in studies of the food fishes. Jour.
Anim.Ecol.19 (1):35-38.
HYSLOP. E.J. 1980. Stomach contents analysis – a review of methods and their
application. J. Fish Biol.;17 (4):411-429.
LAUGHLIN, D.R. & WERNER, E.E.1980. Resource partitioning in two coexisting
sunfish. J. Fish.Res.Bd.Canadá 13 (3):327-342.
LARKIN, P.A. Interespecific competition and population of water fish. J. Fish Res. Bd.;
Ottawa, v. 13, n.3, p. 327-342, 1956.
LO NOSTRO, F. L.; G. A. GUERRERO.1986. Presence of primary and secondary
males in a population of the protogynous Synbranchus marmoratus. Journal of Fish
Biology 49, 788–800.
LOWE-McCONNELL, R.H. 1987. Ecological studies in tropical fish communities.
Cambridge, Cambridge University Press. 382p.
MANTEIFEL, B.P.;I.I. GIRSA & D.S. PAVLOV.1978. On rhytms of fish behaviour, p.
215-224. In:J.E. THORPE (Ed). Rhytmic activity of fishes. London, Academic Press,
312p.
MANN, R.H.K. 1973.Observations on the age, growth, reproduction and food of the
roach, Rutilus rutilus (L.), in two rivers in Southern England. J. Fish Biol. 5, 707-736.
MANN, R.H.K. & ORR, D.R. 1969. A preliminary study of the feeding relationships
of fish in a hard-water and a soft-water stream in Southern England. J. Fish Biol. 1, 31-
44.

41
PILLAY, T.V.R. 1952. A critique of the methods of study of food of fish. Jour. Zool.
Soc. India, Orissa, 4:185-200.
SOARES,M.G.M.; ALMEIDA.; R.G, R.G.; JUNK, W. J.The trophic status of the fish
fauna in Lago Camaleão, a macrophyte dominated floodplain lake in the middle
Amazon. Amazoniana, v. IX n.4, p.511-526,1986.
SWYNNERTON. G.H.; WORTHINGTON.E.B. 1940. Notes on food of fish in
Haweswater (Westmorland). Journal of Animal Ecology. 9:183-187 p.
WELCOMME (1995), R.L. Fisheries ecology of floodplai Rivers. London: Longman,
1973.
WYNES, D.L. & WISSING, T.E.1982. Resource sharing among daters in an ohio
stream.Amer.Midl.Nat.107 (2):294-304.
ZARET, T.M.;RAND,A.S. 1972. Competition in tropical stream fishes: Support for the
competitive exclusion principle. Ecology, v.52, n.2, p.336-342.

42
LISTA DE TABELAS
Tabela 1. Espectro alimentar de Synbranchus marmoratus a)durante o período de
estiagem (agosto de 2007 a dezembro de 2007) b) Espectro alimentar de Synbranchus
marmoratus durante o período chuvoso (janeiro de 2008 a julho de 2008).
Tabela 1 (a). Espectro alimentar de Synbranchus marmoratus durante a) período de
estiagem (n=62 ) e (b) Período chuvoso (N=16) Fo = Freqüência de ocorrência, Fv =
Freqüência volumétrica% e IAi = Índice de Importância alimentar).
a)
Itens alimentares Fo Fv % IAi % Nematodos 3,57 2,77 0,16 Crustáceos 87,5 53,3 78,3 Moluscos 21,4 9,72 3,49 Peixes 19,6 12,5 4,1 Insetos 7,14 2,77 0,33 Detrito 42,8 18,8 13,5
b)
Itens alimentares Fo Fv % IAi % Nematodos - - - Crustáceos 54,5 73,5 90,6 Moluscos - - - Peixes - - - Insetos 27,2 7,35 4,52 Detrito 18,1 11,7 4,79

43
LEGENDA DE FIGURAS
Figura 1. Composição dos itens alimentares de Synbranchus marmoratus no período de
agosto de 2007 a julho de 2008 (N =78).
Figura 2. Índice de Importância alimentar de Synbranchus marmoratus no período de
agosto de 2007 a julho de 2008.
Figura 3. Relação entre a pluviosidade e o Índice médio de Repleção estomacal de
Synbranchus marmoratus no período de agosto de 2007 a julho de 2008.
Figura 4. Relação entre o peso e o comprimento de Synbranchus marmoratus no
período de agosto de 2007 a julho de 2008.
0 10 20 30 40 50 60 70 80 90 100
Nemátodos
Crustáceos
Moluscos
Peixes
Restos de Insetos
Detrito
Itens
alim
enta
res
Porcentagem (%)
Período de estiagem Período chuvoso
Figura 1. Composição dos itens alimentares de Synbranchus marmoratus no período de agosto de 2007 a julho de 2008 (N =78).
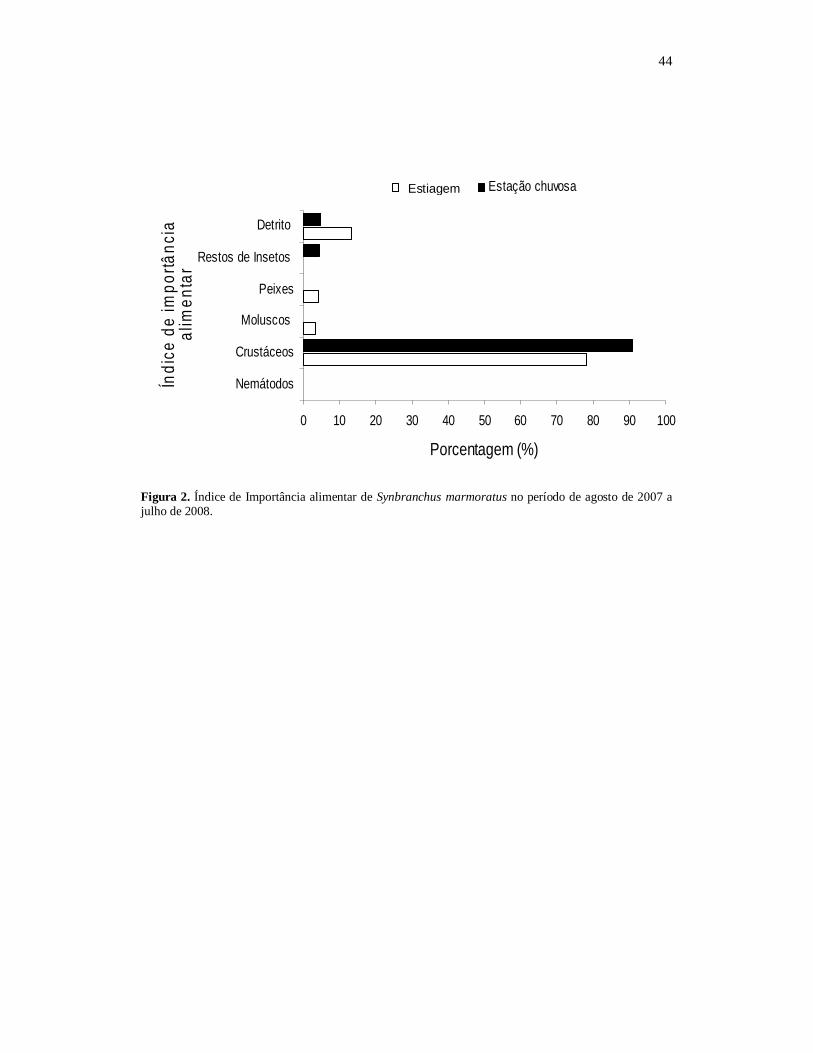
44
Figura 2. Índice de Importância alimentar de Synbranchus marmoratus no período de agosto de 2007 a julho de 2008.
0 10 20 30 40 50 60 70 80 90 100
Nemátodos
Crustáceos
Moluscos
Peixes
Restos de Insetos
Detrito
Índi
ce d
e im
portâ
ncia
al
imen
tar
Porcentagem (%)
Estação seca Estação chuvosaEstiagem
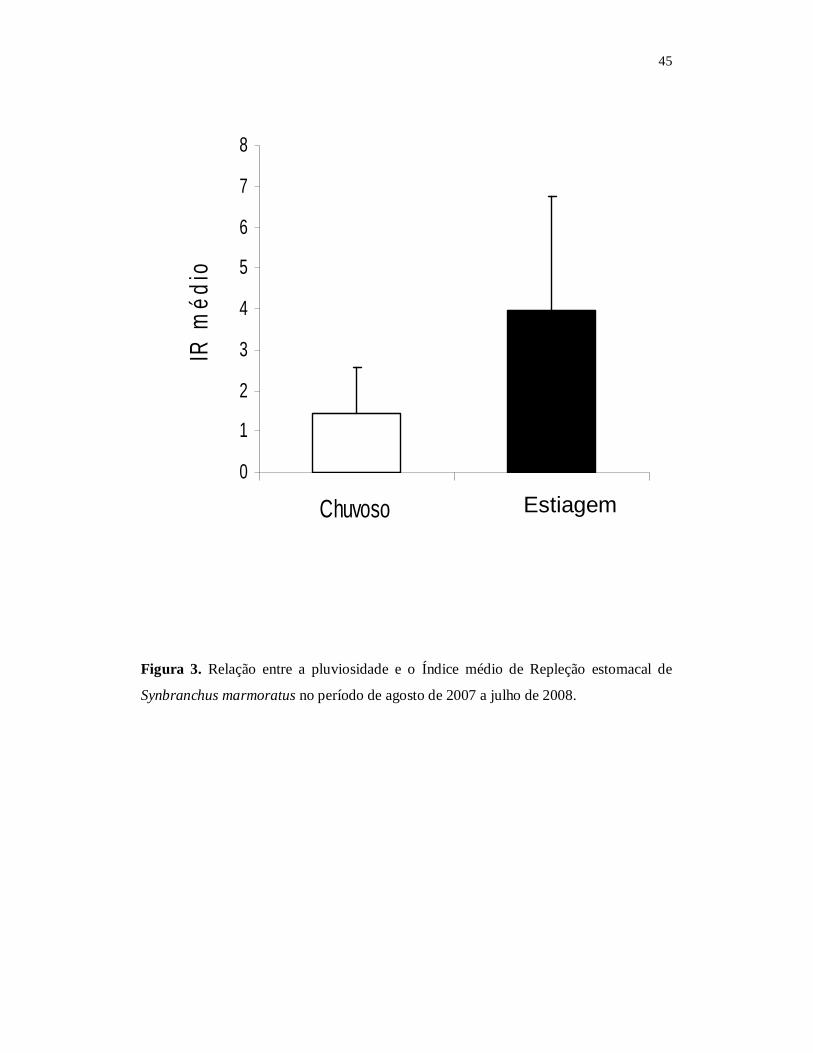
45
Figura 3. Relação entre a pluviosidade e o Índice médio de Repleção estomacal de
Synbranchus marmoratus no período de agosto de 2007 a julho de 2008.
0
1
2
3
4
5
6
7
8
Chuvoso Seco
IR m
édio
Estiagem

46
Figura 4. Relação entre o peso e o comprimento de Synbranchus marmoratus no
período de agosto de 2007 a julho de 2008.
9899
100101102103104105106107108
Chuvoso Seco
Peso
(g)
38
40
42
44
46
48
50
52
Com
prim
ento
(cm
)
Estiagem

47
CAPÍTULO 2
RESULTADOS Artigo Científico 2

48
Morfohistologia do tubo digestório do mussum, Synbranchus
marmoratus Bloch (Osteichthyes: Synbranchidae) em um açude do
semi-árido brasileiro
Luciana Araújo Montenegro1; Naissandra Bezerra da Silva 2 Daisy Nazareth Ferreira
Damasceno3 & Sathyabama Chellappa 1
1 UFRN, Programa de Pós-Graduação em Ciências Biológicas, Centro de Biociências,
Departamento de Oceanografia e Limnologia, Praia de Mãe Luíza, s/n, 59014-100,
Natal, Rio Grande do Norte, Brasil. E-mail: [email protected],
[email protected] 2 UFRN, Centro de Biociências, Departamento de Morfologia e Histologia, Natal, Rio
Grande do Norte, Brasil. E-mail: 3 UFRN, Departamento de Oceanografia e Limnologia, Centro de Biociências, s/n,
59014-100, Natal, Rio Grande do Norte, Brasil. E-mail: [email protected]
RESUMO
O presente estudo descreve e analisa morfohistologicamente o tubo digestório (esôfago, estômago e intestino) do mussum, Symbranchus marmoratus, capturados mensalmente no período de agosto de 2007 a julho de 2008, no açude Marechal Dutra, Acari, Rio Grande do Norte, Brasil. Foram coletados 78 exemplares de espécimes adultos e após s a biometria procedeu-se através de uma incisão ventral a retirada do tubo digestório. Foram feitas secções transversais do esôfago e estômago (porção cárdica, cecal e pilórica). Em seguida, efetuou-se a técnica histológica para obtenção de lâminas histológicas coradas e analisadas por microscopia. Os aspectos morfohistológicos do tubo digestório do S. marmoratus indicam que boca é terminal com grande abertura, lábios delgados, os dentes são pequenos villiformes formando uma única série nas maxilas, apresentam quatro pares de arcos branquiais com rastros branquiais curtos e espassados. O esôfago é curto e apresenta um formato cilíndrico com pequeno diâmetro. A parede esofágica é espessa com a superfície interna formado por mucosa, contendo pregas paralelas. O estômago é do tipo retilíneo, formado pelas porções cárdica, cecal e pilórica. A região de maior comprimento é a região cecal, localizada entre a porção cárdica e pilórica. A região cárdica que é a primeira porção do estômago é curta e cilíndrica, formada por mucosa constituída por epitélio cilíndrico simples, com muitas células caliciformes. A região cecal é alongada e apresenta paredes delgadas, luz maior que a cárdica, com pregas de menor porte, revestido por epitélio cilíndrico simples com glândulas gástricas na submucosa. A porção pilórica não possui glândulas e apresenta pregas principais e secundárias em sua mucosa. Os aspectos morfohistológicos do tubo digestório do S. marmoratus mostram que a espécie é adaptada para hábito alimentar carnívoro.
Palavras Chave: peixe, morfohistologia, tubo digestório, peixe carnívoro.

49
INTRODUÇÃO
Investigações do espectro alimentar, por meio de estudos de conteúdos
estomacais, aliadas a análises morfológicas de órgãos envolvidos na tomada do
alimentar ou que participam da digestão alimentar, têm auxiliado na interpretação da
dinâmica alimentar e ocupação de habitats por espécies icticas (Wootton,1990)
Nesse sentido Blazquez (1990) afirma que alem de processar os alimentos,
algumas partes do tubo digestório são especiaizadas, auxiliando a respiração, como
ocorre em Plecostomus comersonii (cascudo). Ribeiro e Fanta (2000), demonstraram,
através de estudos histológicos e histoquímicos, o envolvimento do epitélio do esôfago
de alguns peixes marinhos e continentais na função de osmorregulação.
Estudos histológicos e da morfologia, fornecem subsídios para a caracterização
dos órgãos do sistema digestório, de forma que possam ser melhor compreendidos a
fisiologia e o hábito alimentar da espécie a ser estudada possibilitam à obtenção de uma
descrição histológica padrão, além de poder auxiliar em avaliações de impacto
ambiental. (Al-Hussaini 1949) e (Morrison & Wright Jr. 1999).
Synbranchus marmoratus Bloch 1795, também conhecido como mussum é um
peixe teleósteo, pertencente a família Symbranchidae, ordem Synbranchiformes,
incluem peixes em forma de enguia (Lo Nostro, 1986).
Este trabalho visa contribuir com os estudos da morfologia e histologia dos
Teleósteos, descrevendo a histologia do tubo digestório, da espécie S. marmoratus, a
fim de facilitar o seu gerenciamento, colaborando na sua preservação, e abrindo novas
pespectivas para pesquisas futuras.
MATERIAL E MÉTODOS
A amostragem do estudo foi realizada no Açude Marechal Dutra (5°30’Lat. S e
37° Long. W), situada no município de Acari, Estado do Rio Grande do Norte. O Açude
Marechal Dutra está inserido no bioma Caatinga, possui uma capacidade de acumulação
de 40.000.000 m3, profundidade máxima de 25 metros e média anual de 5,2 metros. A
população de pescadores a utiliza como fonte alimentar e de renda, dependendo
exclusivamente da pesca para o seu sustento.

50
As coletas foram realizadas numa freqüência mensal do período de agosto de
2007 a julho de 2008 utilizando-se covos fixados próximos à vegetação marginal. Neste
estudo foram utilizados 42 exemplares de Symbranchus marmoratus, com 103,8 ± 0,95
cm de comprimento padrão e 43,1 ± 8,6 g de peso corpóreo, coletados na região
marginal do açude Marechal Dutra, no distrito de Gargalheiras, município de Acari, RN,
a 219 km de Natal, o capital do Estado.
No Laboratório de Técnicas Histológicas (UFRN) Campus Universitário, foi
realizada a biometria e dissecação dos animais, através de uma incisão no sentido
longitudinal, partindo do poro anal do animal até a cabeça. O tubo digestório foi
retirado da cavidade celomática, seguido de seccionamentos transversais para separação
do esôfago, estômago (porção cárdica, cecal e pilórica) e intestino e logo após,
imediatamente fixados em Bouin a 10%, e devidamente identificados.
Para análise anatômica dos órgãos digestórios foi utilizado o microscópio
Olympus, modelo CH30 acoplado a câmara Olympus PM 10 AK. Dos órgãos
digestórios foram retirados fragementos, que faziam parte das regiões esofágica,
cárdica, fúndica e pilórica estomacais e do intestino anterior, médio e posterior. Em
seguida efetuou-se a técnicas histológicas de rotina para microscopia óptica
desidratação, diafanização e inclusão em parafina, coradas com Hematoxilina de Harris,
Eosina (Maia, 1979) e Ácido Periódico-Schiff (Behmer et al., 1976).
Para a descrição morfológica, especial atenção foi dada a caracterização do
formato geral do corpo, posição e tamanho da boca, posição e tamanho dos olhos, tipo
de dentição, tipo de forma das brânquias e rastros branquiais. Foram consideradas
também as estruturas da mucosa, quanto ao tipo de epitélio e os tipos celulares que o
compõem, assim como uma possível ocorrência de glândulas na lâmina própria e a
presença ou ausência de muscular da mucosa, a caracterização da submucosa e
distribuição de células granulares ou glândulas nesta túnica. Além disso, também foram
consideradas a organização da túnica muscular, da serosa ou da adventícia quando
presente.
RESULTADOS
A espécie S. marmoratus possui corpo tubular e comprido longitudinalmente,
com boca terminal e lábios delgados, aderidos a maxila com presença de corpúsculos
gustativos com função de localização, seleção e captura do alimento. Os dentes são

51
pequenos e viliformes, formando uma única série nas maxilas. Possui quatro pares de
arcos branquiais curtos e grossos, onde se apresentam dentes faringeanos com função de
apreensão do alimento.
→INSERIR Fig. 1
O esôfago do mussum, S. marmoratus apresenta-se de formato cilíndrico, de
pequeno diâmetro, curto. Sua parede se apresenta espessa, a superfície interna é
formado por mucosa, contendo pregas paralelas entre si. A mucosa apresenta epitélio
pseudo estratificado contendo células secretoras de muco, células basais e células de
revestimento apicais. Observa-se a presença de lâmina própria formada por tecido
conjuntivo denso e aglandular. Existem pregas na mucosa com feixes de musculatura
estriada chegando até a lâmina própria. A camada muscular é constituída por
musculatura lisa e estriada. O estômago do S. marmoratus delimita-se com o esôfago
pela presença de um estreitamento.
O esôfago é recoberto por uma camada adventícia ao se aproximar do estômago
surge a serosa. Na região de transição entre os dois órgãos o epitélio gradativamente
torna-se cilíndrico e aumenta o número de células secretoras de muco.
O estômago é do tipo retilíneo sendo a região de maior comprimento. A região
cecal foi intermediária com a porção cárdica e pilórica. A região cárdica, primeira
porção do estômago, é curta e cilíndrica, formada por mucosa constituída por epitélio
cilíndrico simples, com muitas células caliciformes. Apresenta paredes espessas e luz
reduzida pela grande quantidade de dobras que apresenta. Observa-se uma ausência de
glândulas nessa região e destaca-se nessa região o surgimento de uma serosa muito
vascular, o que pode estar relacionado com a respiração.
A região cecal é alongada para receber o alimento, e apresenta paredes mais
delgadas, luz maior, com pregas de menor porte, proporcionalmente ao tamanho,
formado por epitélio cilíndrico simples, que se invagina e origina glândulas gástricas
com função de secreção de ácido clorídrico e pepsinogênio.
As pregas estomacais são constituídas por mucosa e submucosa. A submucosa
não apresenta glândulas e é constituída por conjuntivo contendo vasos sanguíneos. A
camada muscular da porção cecal é dividida em uma camada circular e outra
longitudinal. A camada muscular é liso dividindo-se em duas camadas, a primeira
circular, desenvolvida na maior parte das regiões e a segunda uma camada longitudinal.
→INSERIR Fig. 2

52
A porção pilórica é aglandular, em sua mucosa observa-se a presença de pregas
principais que originam pequenas pregas secundárias. A mucosa encontra-se constituído
por epitélio cilíndrico simples muco secretor, seguido de lâmina própria contendo
apenas vasos sanguíneos. A camada muscular possui circular de diâmetro superior as
demais camadas musculares estomacais.
→INSERIR Fig. 3
Ao longo de toda a porção pilórica evidenciam-se dobras, em sentido
longitudinal de tamanhos variados. Ao longo das dobras observou-se invaginações
secundárias dando as primárias o aspecto denteado e fornecendo ao epitélio de
revestimento, uma superfície mais ampla de secreção.
Histologicamente foram observadas as seguintes camadas no estômago de S.
marmoratus: mucosa, submucosa, túnica muscular e serosa.
Levando-se em consideração a presença de glândulas estomacais foi observado
que ocorre variação na presença de glândulas de acordo com a região. A porção cárdica,
por exemplo, apresenta-se aglandular. Na porção cecal é observado um maior número
de glândulas gástricas, que se originam a partir de invaginações de pregas da mucosa,
contendo epitélio cilíndrico simples, estas persistem em grande quantidade na região
cecal, logo após, sofrendo diminuição no número até desaparecerem ao chegar na
porção pilórica.
Externamente o estômago é revestido por uma serosa, onde se evidenciam vasos
sanguíneos e nervos, As características histológicas do estômago, evidenciadas em S.
marmoratus, sugerem que o estômago dessa espécie é especializado na digestão
mecânica, sendo uma espécie carnívora.
A análise histológica do intestino demonstra a presença de vilosidades intestinais
constituídas de epitélio cilíndrico simples, que junto com a lâmina própria originam
vilosidades longas e ramificadas, revestidas por células secretoras de muco e células
absortivas. A mucosa intestinal é aglandular e não observa-se uma submucosa.
Posteriormente surge a muscular, constituída de túnica circular e longitudinal. A última
camada verificada é uma serosa que contém uma rica rede capilar.
DISCUSSÃO
Os peixes estão adaptados para se alimentar de um determinado item, ou
conjunto de itens. A plasticidade na dieta de peixes obedece a certo limite pré-

53
estabelecido pela forma do tubo digestivo, cujas estruturas podem refletir as tendências
alimentares da espécie. Conforme Pillay (1952), a natureza do alimento ingerido
depende primeiramente da morfologia e do comportamento alimentar do peixe, e
secundariamente da composição e quantidade do alimento disponível.
A posição, formato e tamanho da boca estão intimamente relacionados aos
hábitos alimentares e, em especial a forma de apreensão do alimento (Nikolsky, 1963).
Weatherley & Gil (1987) ressaltam que a posição da boca, formato e posição dos
dentes, morfologia dos rastros e presença de estruturas gustativas nos lábios estejam
diretamente relacionadas ao hábito alimentar do peixe. Russo et al. (2004), verificarm
que em adultos de Bryconamericus sp. do rio Iguaçu, que se alimentaram
principalmente da fauna bentônica, a boca terminal apresentou vantagens para a
sobrevivência do gênero, por facilitar a captura da presa. Assim, a posição terminal da
boca da espécie em estudo, S. marmoratus auxilia na apreensão de presas, favorecendo
a captura pelo alimento.
Em relação ao número de rastros branquiais observa-se uma relação inversa com
a presença de itens alimentares maiores e bentônicos na dieta (Watson & Balon, 1984).
O mussum, S. marmoratus, Possui quatro pares de arcos branquiais curtos e grossos,
onde se inserem dentes faringeanos com função de apreensão do alimento. De acordo
com MacNeill & Brandt (1990) a morfologia dos rastros branquiais determina a
eficiência de retenção de partículas de vários tamanhos.
O estômago do tipo retilíneo de S. marmoratus contrasta-se com a espécie
Prochilodus scrofa, que apresenta estômago sinfonal conforme verificado por
Hernandezblazquez et al. (1990), e com Hoplias malabaricus, que apresenta uma
forma intermediária entre os dois tipos conforme Braccini (2002).
A região cecal é alongada para receber o alimento, e apresenta paredes mais
delgadas, luz maior, com pregas de menor porte, proporcionalmente ao tamanho,
formado por epitélio cilíndrico simples, que se invagina e origina glândulas gástricas.
Semelhante ao observado por Gargiulo et al. (1998), estudando a mucosa gástrica da
tilápia, onde foi verificado que não existe diferenciação glandular, sendo o acído
clorídrico e o pepsinogênio secretado pela mesma célula.
Moraes & Barbola (1995) observam que o desenvolvimento da região cecal em
predadores, nadadores ativos, pode estar relacionado com a capacidade dos mesmos de
atacarem cardumes e consumirem grandes quantidades de presas em um curto espaço de
tempo.

54
A camada muscular é liso, dividindo-se em duas camadas, a primeira circular,
mais desenvolvida na maior parte das regiões, e externamente, uma camada
longitudinal.
Externamente o estômago de S. marmoratus é revestido por uma serosa, onde se
evidenciam vasos sanguíneos e nervos, também constatados por Alves & Tomé Kessler
et al. (1979) em Prochilodus sp. As características histológicas do estômago,
evidenciadas em S. marmoratus, sugerem que o estômago dessa espécie é especializado
na digestão mecânica, sendo considerado como uma espécie carnívora.
Dentre os vertebrados, são os teleósteos que apresentam uma das estruturas
histológicas intestinais mais simples (Al-Hussaini 1949; Khanna, 1971). e células
absortivas. Segundo Kappor et al (1975) o comprimento do intestino é uma variável que
responde sensivelmente a mudanças nas condições de alimentação. Zihler (1982)
destaca a dependência do comprimento do intestino ao valor nutricional do alimento
ingerido, peixes que se alimentam de itens resistentes a digestão, como detritos e
exoesqueletos calcários e quitinosos, apresentam o intestino mais longo.
O intestino propriamente dito (intestino médio e posterior na classificação
embriológica) dos peixes geralmente não apresenta particularidades anatômicas que
permitam dividi-lo em segmentos bem definidos. Assim, Para a distinção entre as
diferentes regiões do intestino de S. marmoratus, são ainda necessários estudos mais
elaborados, como os histofisiológicos e os histoquímicos, que podem permitir a
diferenciação, com total clareza, entre as áreas de absorção de gorduras (duodeno), de
absorção de proteínas (intestino grosso) e de absorção de água e íons (reto).
Os aspectos morfohistológicos do trato digestório do S. marmoratus mostram
que a espécie é adaptada para hábito alimentar carnívoro, pois o seu trato digestório
apresenta estruturas morfohistológicas semelhantes a outras espécies carnívoras, como
por exemplo, por exemplo estômago saculiforme e intestino longo.
REFERÊNCIAS
AL-HUSSAINI, A.H. 1949 On the functional morphology of the alimentary tract of
some fish in relation to differences in their feeding habits. II – Cytology and
Physiology. Quart. Jour. Microsc. Soc., London, 90: 323-354.

55
BEHMER, A.O.; TOLOSA, E.M.C.; F. NETO, A.G. 1976 Manual de técnicas para
histologia normal e patológica. São Paulo: Editora da Universidade de São Paulo. p 98-
101.
GARGIULO, A.M., CECCARELLI, P., DALLI AGLIO, & PEDINI V 1998. Histology
and ultrastructure of the gut of the tilapia (Tilápia spp), a hybrid teleost. Anatomy,
Histology and Embriology 27, 89-94.
HERNANDEZ-BLÁZQUEZ, F. J.; FERREIRA, N.; CARVALHO, C. A. F. de &
FERRI, S. Morphological aspects of the esophagus and stomach of a freshwater
iliophagus fish under scanning electron microscopy. Rev. Gengebaurs Morphol. Jahrb.,
136(1) 1990. p. 119-125.
KHANNA, S.S. 1964. The alimentary canal in some teleostean fishes. Jour. Zool.
Soc.India 13: 206-219.
KAPOOR, B.G.; H. SMITH & I.A. VEREGINA. 1975. The alimentary canal and
digestion in teleosts. Adv. Mar. Biol. 13:109-239.
LO NOSTRO, F. L.; G. A. GUERRERO. 1986. Presence of primary and secondary
males in a population of the protogynous Synbranchus marmoratus. Journal of Fish
Biology 49, 788–800.
MACNEILL, D.B. & S.B. BRANDT.1990. Ontogeneticshifts in gill-raker morphology
and predicted prey capture efficiency of the alewife Alosa Pseudoharengus . Copeia,
Carbondale, 1: 164-171.
MAIA, V. 1979 Técnica Histológica. 2. ed. São Paulo: Atheneu. p.70-136.
MORRISON, C.M. & WRIGHT Jr, J.R. 1999 A study of the histology of the digestive
tract of the Nile tilapia. J. Fish Biol., 54: 597-606.
NIKOLSKY, G.V. The ecology of fishes. London: Academic Press, 1963.

56
PILLAY, T.V.R.1952. A Critique of the methods of study of food of fishes. Jour. Zool.
Soc. India, Orissa, 4: 185-200.
RIBEIRO, C.A.O. & FANTA. E. 2000 Microscopic morphology and histochemistry of
the digestive system of a tropical freshwater fish Trichomycterus brasiliensis (Lütken)
Trichomycteridae). Revta bras. Zool., 17(4): 953-971.
ROSEN, D. E. & GREENWOOD, P. H. (1976). A fourth neotropical species of
synbranchid eel and the phylogeny and systematics of synbranchiform fishes. Bulletin
of American Museum of Natural History 257, 1–69.
RUSSO, M.R. et al. Feeding habits of two species of Bryconamericus Eigenmann,
1907 of the iguaçu river, Brazil. Acta Sci., Maringá, v. 26. N.4, p.431-426, 2004.
VEGAS-VELEZ, M. La structure histologique typique du tube digestif des poissons
télèosteens. Tethys, 4(1): 1972. p. 163- 174.
WATSON, D.J. & BALON, E.K. 1984. Ecomorphological analyses of fish taxocenes
in rainforest of northern Borneo, Jounal of Fish Biology 25, 371-384.
WEATHERLEY, A.H. & GILL, H.S. 1987. Feeding relations, correlated functional
Morphology, growth and size. London: Academic Press.
WOO, R.J., EVANS., G.W., MILLS, L.A (1978). Annual cycle in female three-spined
sticklebacks (Gasterosteus aculeatus L.) from an upland and lowland population. J. Fish
Biol., 12:331-343.
ZIHLER, F. 1982. Gross morphology and configuration of digestive tracts of Cichlidae
(Teleostei, perciformes): phylogenetic and functional significance. Neth. Jour. Zool.
32(4): 544-571.

57
LEGENDA DE FIGURAS
Figura 1. Espécie em estudo Synbranchus marmoratus
Figura 2. Tubo digestório de S. marmoratus com indicação das estruturas A) Faringe;
B) Esôfago; C1) Estômago região cárdica; C2) Estômago região cecal; C3)
Estômago, região pilórica; D) Intestino porção anterior e média.
Figura 3. Região esofágica de Synbranchus marmoratus, A a1) epitélio pseudo
estratificado; a2) células cubóides de revestimento (PAS); B b1) células
basais do esôfago; (b2) células mucosas (PAS); C c1) prega do esôfago; c2)
camada muscular (PAS); D d1) lâmina própria (PAS).
Figura 4. Região cárdica do estômago de Synbranchus marmoratus, A a1 )submucosa;
a2) vasos sanguíneos (PAS); B b1) pregas da mucosa; b2) luz da mucosa
(PAS); C c1) muscular; c2) serosa vascularizada (PAS).
Figura 5. Região cecal do estômago de Synbranchus marmoratus, A a1) camada
muscular da submucosa; a2) pregas da mucosa; (a2)luz contendo muco
(PAS); B b1) epitélio cilíndrico simples; b2) glândulas gástricas (PAS); C
(c1) serosa contendo vasos sangiíneos (PAS); D d1) vasos sanguíneos com
hemácias (PAS).
Figura 6. Região pilórica do estômago de Synbranchus marmoratus, A (a1) luz; a2)
epitélio cilíndrico simples (PAS); B b1) pregas da mucosa; b2) lâmina
própria (PAS); C (c1)camada muscular; (c2) vasos sanguíneos(HE).
Figura 7. Região intestinal de Synbranchus marmoratus, A a1)luz intestinal a2)pregas
intestinais(PAS); B b1)epitélio cilíndrico simples (PAS); C (c1) Célula
cilíndrica mucossecretora (PAS); D d1) camada muscular lisa; d2) serosa
contendo vasos sanguíneos (PAS).

58
Figura 1 - Espécie em estudo Synbranchus marmoratus.
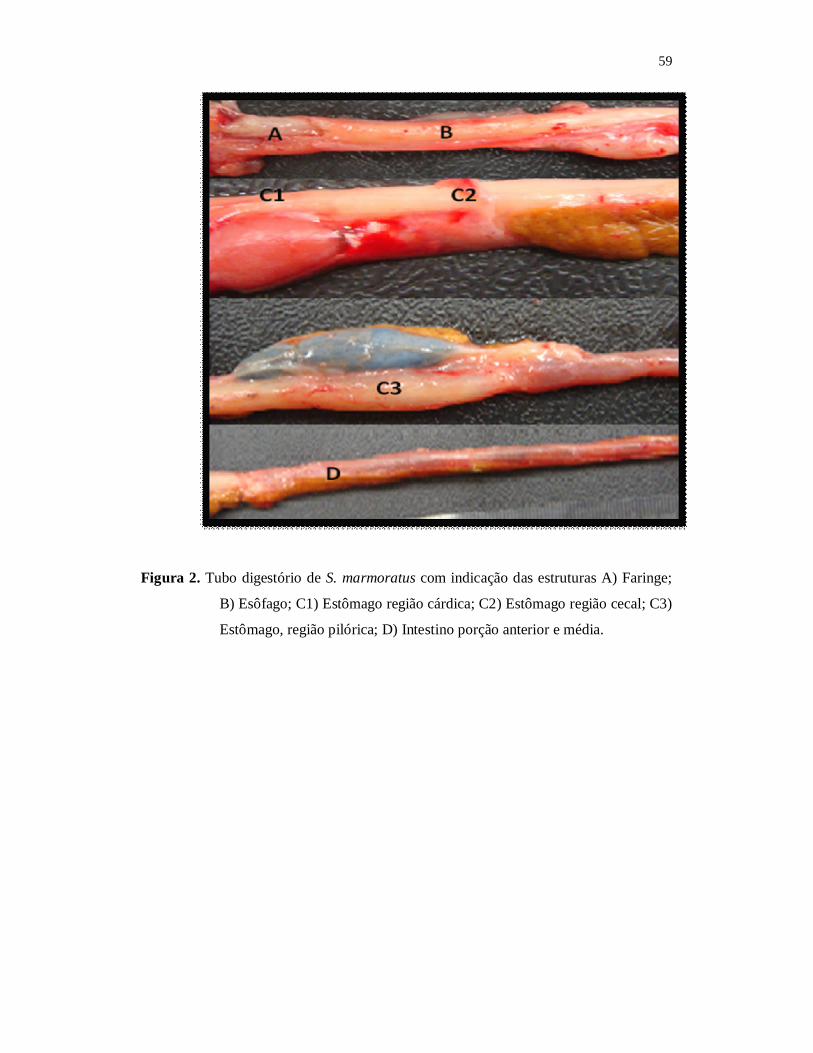
59
Figura 2. Tubo digestório de S. marmoratus com indicação das estruturas A) Faringe;
B) Esôfago; C1) Estômago região cárdica; C2) Estômago região cecal; C3)
Estômago, região pilórica; D) Intestino porção anterior e média.

60
Figura 3. Região esofágica de Synbranchus marmoratus, A a1) epitélio pseudo
estratificado; a2) células cubóides de revestimento (PAS); B b1) células
basais do esôfago; (b2) células mucosas (PAS); C c1) prega do esôfago; c2)
camada muscular (PAS); D d1) lâmina própria (PAS).
C
B
b1
A
B A
Cc
DD
b2
b1
d1
c2
C
c2
100µm
100µm
300µm
50µm
a1
c1
b c

61
Figura 4. Região cárdica do estômago de Synbranchus marmoratus, A a1 )submucosa;
a2) vasos sanguíneos (PAS); B b1) pregas da mucosa; b2) luz da mucosa
(PAS); C c1) muscular; c2) serosa vascularizada (PAS).
c2
b2
b1
a2
c1
B
A
Cc
DD
a1
100µm
50µm
50µm
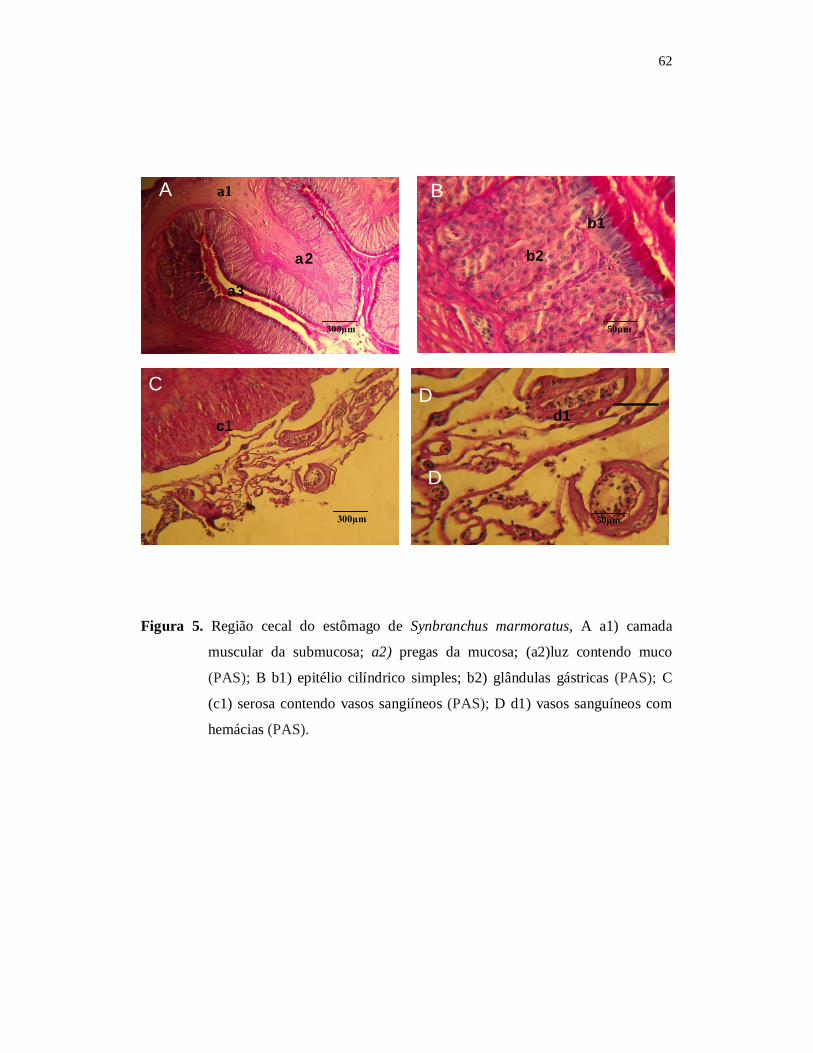
62
Figura 5. Região cecal do estômago de Synbranchus marmoratus, A a1) camada
muscular da submucosa; a2) pregas da mucosa; (a2)luz contendo muco
(PAS); B b1) epitélio cilíndrico simples; b2) glândulas gástricas (PAS); C
(c1) serosa contendo vasos sangiíneos (PAS); D d1) vasos sanguíneos com
hemácias (PAS).
a3
a2
b1
c1
A B
C D d1
a1
b2
300µm
50µm
300µm
50µm
D
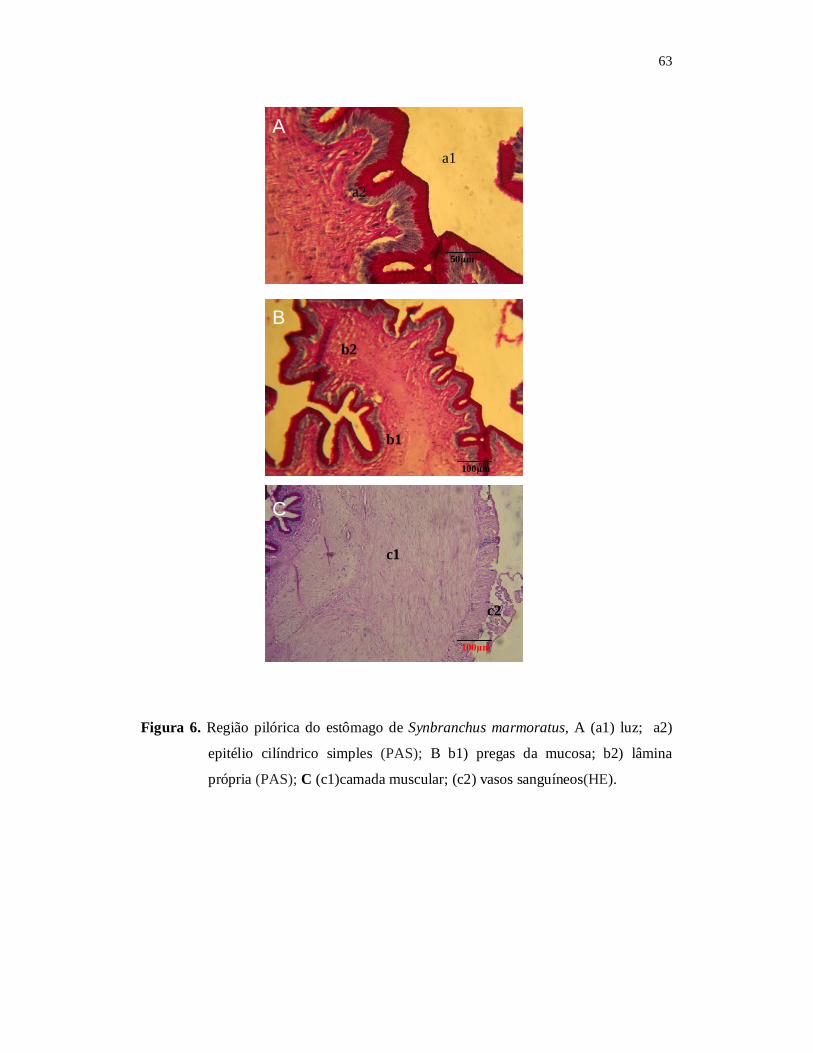
63
A
A D
B
Figura 6. Região pilórica do estômago de Synbranchus marmoratus, A (a1) luz; a2)
epitélio cilíndrico simples (PAS); B b1) pregas da mucosa; b2) lâmina
própria (PAS); C (c1)camada muscular; (c2) vasos sanguíneos(HE).
d1
A a1
A
B
C D
b1
50µm
100µm
a2
B b2
C
c1
100µm
c2
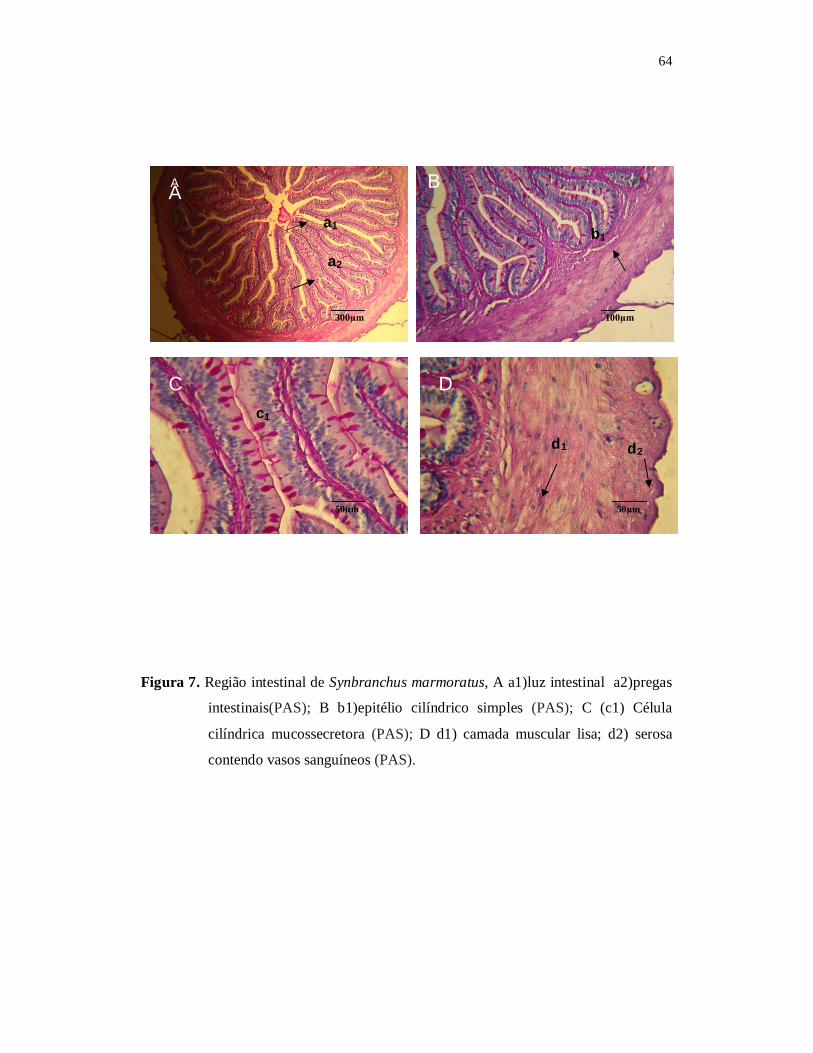
64
Figura 7. Região intestinal de Synbranchus marmoratus, A a1)luz intestinal a2)pregas
intestinais(PAS); B b1)epitélio cilíndrico simples (PAS); C (c1) Célula
cilíndrica mucossecretora (PAS); D d1) camada muscular lisa; d2) serosa
contendo vasos sanguíneos (PAS).
a1
a2
b1
c1
d1
A
d2
300µm
50µm
100µm
50µm
A
C
B
D
d d

65
5. DISCUSSÃO GERAL
O número de exemplares de S. marmoratus capturados variou sazonalmente
sendo maior na vazante, com 62 exemplares e menor no período chuvoso, com 16
exemplares. mas foi suficiente para analisar a dieta, durante o período de estudo. A
maioria dos peixes estudados foi maior que 43 cm, e a média do peso corporal foi de
103,9g.
Araujo-Lima et al. (1995) sugeriram que a oferta de alimento, a uma
comunidade de peixes, é dependente de variações ambientais, principalmente em
relação ao nível da água do rio. Durante a enchente e cheia o nível da água permite o
acesso dos peixes à região marginal alagada, criando um ambiente que favorece os
predadores que tocaiam suas presas.
Como o mussum. Durante a seca o espaço é reduzido e todas as espécies se
concentram no açude, o que favorece aos carnívoros (MERONA e BITTENCOURT,
1993).
A intensidade de alimentação ou oferta de alimento pode ser refletida pela
quantidade de estômagos vazios numa população. No presente trabalho a freqüência de
estômagos vazios do S. marmoratus foi em média 26,8% e esteve relacionada ao nível
do rio, sendo esta mais alta durante o período chuvoso. A quantidade de estômagos
vazios durante o período chuvoso, corrobora com a com a hipótese de aumento de
densidade de presas com a estiagem, que elevaria a oferta de alimento. Possivelmente o
jejum durante a reprodução e a ausência de floresta inundável exercem maior influência
sobre a intensidade de alimentação da espécie.
A grande ocorrência de camarões verificada na dieta de S. marmoratus durante
todo o período de estiagem pode ser explicado pelo fato que durante a estiagem, os
camarões ficam no fundo e são mais facilmente capturados pelo S. marmoratus, que é
um peixe bentônico. A grande freqüência de animais bentônicos na dieta quando o nível
da água está baixo, sugere uma tendência bentônica do mussum. Nesta época a
profundidade de água diminui (Odinetz-Collart & Moreira, 1993) e a luz penetra em
toda a coluna d’água atingindo partes mais profundas, permitindo ao mussum explorá-la
melhor.
O número mediano de presas no estômago foi igual a 3, apesar de ser encontrado
até dez indivíduos nos estômagos de alguns indivíduos, nestas situações as presas

66
estavam em estágios de digestão semelhantes. Provavelmente os indivíduos não
capturaram alimento continuamente, mas comeram durante períodos específicos do dia.
A presença ou ausência de um item alimentar no estômago de um peixe predador
pode estar relacionada à sua disponibilidade no ambiente (Nikolsky, 1978) embora
algumas vezes, indivíduos de uma mesma população, e amostrados na mesma época e
local, possam ter dietas diferentes. Na maioria dos casos o tamanho das presas tem
relação direta com o tamanho de peixes predadores (WOOTTON, 1990).
Russo et al. (2004) verificaram que em adultos de Bryconamericus sp. do rio
Iguaçu, que se alimentaram principalmente da fauna bentônica, a boca terminal
apresentou vantagens para a sobrevivência do gênero, por facilitar a captura da presa.
Assim, a posição terminal da boca da espécie em estudo, S. marmoratus auxilia na
apreensão de presas, favorecendo a captura pelo alimento.
Moraes e Barbola (1995), colocam que o desenvolvimento da região cecal em
predadores, nadadores ativos, pode estar relacionado com a capacidade dos mesmos de
atacarem cardumes e consumirem grandes quantidades de presas em um curto espaço de
tempo.
Os aspectos morfohistológicos do tubo digestório do S. marmoratus mostram
que a espécie é adaptada para hábito alimentar carnívoro, pois o seu trato digestório
apresenta estruturas morfohistológicas semelhantes a outras espécies carnívoras, como
por exemplo, estômago saculiforme e intestino longo.

67
6. CONCLUSÃO GERAL
Os estudos realizados para determinar a biologia alimentar e morfohistologia do
trato digestório do mussum, S. marmoratus (Osteichthyes: Synbranchidae), capturados
no açude Marechal Dutra, RN do semi-árido brasileiro, permitiram-nos chegar a
seguintes conclusões:
A espécie apresentou maior atividade alimentar durante os meses de estiagem.
A dieta foi composta de itens de origem animal. Dessa forma, o hábito alimentar
foi caracterizado como carnívoro com preferência a carcinofagia.
Os aspectos morfohistológicos do tubo digestório do S. marmoratus mostram
que a espécie é adaptada para hábito alimentar carnívoro.

68
REFERÊNCIAS BIBLIOGRÁFICAS GERAIS
AL-HUSSAINI, A.H. 1949 On the functional morphology of the alimentary tract of
some fish in relation to differences in their feeding habits. II – Cytology and
Physiology. Quart. Jour. Microsc. Soc., London, 90: 323-354.
ARAUJO-LIMA, C. A. R. M.; AGOSTINHO, A. A.; FABRÉ, N. N.; BICUDO, C. E.
M.;MATSUMURA TUNDISI, T. 1995. Trophic aspects of fish communities in
Brazilian rivers and reservoirs. In Tundisi, J. G.; Bicudo, C. E. M.; Matsumura Tundisi,
T. (Eds). Limnology in Brazil. ABC/SBL,Rio de Janeiro. p.105-136.
AGOSTINHO, A. A.; GOMES, L.C. Reservatório de Segredo: Bases ecológicas para o
Manejo. Maringá: Eduem, 1997. 387 p.
ALMEIDA, R.G.; SOARES, L.H. & EUFRÁSIO, M.M. 1993.Lagoa do Piató: Peixes e
Pesca. Coleção Vale do Açu, Coleção Humanas Letras. Natal, 84p.
BEHMER, A.O.; TOLOSA, E.M.C.; F. NETO, A.G. 1976 Manual de técnicas para
histologia normal e patológica. São Paulo: Editora da Universidade de São Paulo. p 98-101.
BRITSKY, H. A. (1970). Peixes de água doce do Estado de São Paulo. Sistemática.
Poluição e Piscicultura. Faculdade de Saúde Pública da USP e Instituto de Pesca –
C.P.R.N – S.A.
BRITSKI, H. A., SILIMOM, K. Z. S., LOPES, B. S. (1999). Peixes do pantanal.
Manual de identificação. H. A. Britski (ed.). Brasília: EMBRAPA – SPI, 184p.
BONNEAU, D.L., McGUIRE, J.W.; TIEMEIER, O.W. & DEYONE, C.W. 1972. Food
habits and growth of channel catfish fry Ictularus punctatua. Trans.Am.Fish.Soc. 101,
613-614.
BUENO, R. M. X. Biologia Reprodutiva do cascudo, Hypostomus pusarum (Starks,
1913) (Osteichthyes: Loricariidae) do açude Marechal Dutra, Acari, Rio Grande do

69
Norte. Dissertação (Mestrado em Bioecologia Aquática), Universidade Federal do Rio
grande do Norte, Natal, RN. 137p. 2004.
BUENO, R. M. X. & CHELLAPPA, S. Composição ictiofaunística de valor comercial
do açude público Marechal Dutra, Acari, RN. Anais do Congresso Nordestino de
Ecologia. 10., Olinda, PE. 2003. (CD-ROM).
CARAGITSOU, E.; PAPACONSTATINOU. Food and feeding habits of large scale
gurnard, Lepidotrigla cavilloneI (Triglidae) in Grrek seas. Cybium, Paris, v.14, n.2,
p.95-100, 1990.
CANAN, B., GURGEL, H. B.; NASCIMENTO, R. S. S., BORGES, S. A. G. V. &
BARBIERI, G.1997. Avaliação da comunidade de sete espécies de peixes da lagoa Boa
Cicca, Nísia Floresta – RN. Revista Ceres, Viçosa, 44 (256): 604-616.
CÂMARA, M.R. Ecologia e Reprodução do Tucunaré comum, Cichla monoculus
(Osteichthyes: Cichlidae) da Barragem de Campo Grande, RN. Dissertação (Mestrado
em Bioecologia Aquática) Universidade Federal do Rio Grande do Norte, Natal. 1998.
CÂMARA, M.R. VERANI, J.R.; CHELAPPA, S. Descrição morfométrica, mesrística e
produção de muco do acará disco, Symphysodon discus Heckel, 1840 (Osteichthyes:
Cichlidae) in: Congresso Brasileiro de Zoologia, 24.; 2002, Curitiba. Associação
Brasileira de Zoologia, 2002b.v.1.p.319.
CHELLAPPA, S.; CHELLAPPA, N. T.; SILVA, E.A.; HUNTINGFORD, F. A. &
BEVERIDGE, M. C. M. The diet of hybrid red tilapia, Oreochromis niloticus
(Linnaeus) x Oreochromis mossambicus (Peters) reared in the freshwater ponds of
Northeastern Brazil. Aquaculture Research, 27 (12), 945-952, 1996.
CHELLAPPA, N. T. & . A. M. Costa. Dominant and co-existing species of
Cyanobacteria from a eutrophicated reservoir of Rio Grande do Norte State, Brazil.
Acta Oecologica – International Journal of Ecology. 24, 3-10, 2003.

70
CHELLAPPA, N.T.; CÂMARA, M.R.; CHELLAPPA, N.T. Ecology of Cichla
monoculus (Osteichthyes: Cichlidae) from a reservoir in the semi-arid region of Brazil.
Hydrobiologia, 504. P. 267-273. 2003a.
COSTA, M. A. M. Ecologia da Comunidade Fitoplanctônica da Barragem Gargalheiras
em Acari, RN. Dissertação ((Mestrado em Bioecologia Aquática), Universidade Federal
do Rio grande do Norte, Natal, 2000. Dissertação.
CRISP, D.T.; MANN, R.H.K. & McCORMACK, J.C. (1978). The effects of
impoundment and regulation upon the stomach contents of fish at Cow Green, Upper
Teesdale. J. Fish Biol. 12, 287-301.
ESTEVES, F.A. (1988). Fundamentos de Limnologia. Rio de Janeiro.
Interciência.FINEP. 575pp
ESTEVES, K. E. 1996. Feeding ecology of three Astyanax species (Characidae,
Tetragonopterinae) from a floodplain lake of Mogi-Guaçú River, Paraná River Basin,
Brazil. Environmental Biology of Fishes. 83-101 p
FREIRE, A.G.; ALMEIDA, R.G.; MENESCAL, R.; MEDEIROS, P.B.; SILVA, M. M.
Caracterização ictiofaunistica dos grandes açudes do semi-árido do Rio Grande do
Norte. Coleção Mossoroense. Mossoró, RN.;série C, v. 1135, p. 269-275, agosto, 2000.
FISHBASE: A global information of system on fishes. Word Wide Web Eletronic
Publication. Froesa, R. and D. Pauly. (Ed). Synbranchus marmoratus. Acesso em: 14 de
setembro/2008.
GARGIULO, A.M., CECCARELLI, P., DALLI AGLIO, & PEDINI V 1998. Histology
and ultrastructure of the gut of the tilapia (Tilápia ssp), a hybrid teleost. Anatomy,
Histology and Embriology 27, 89-94.
GERKING, S.D. Feeding ecology of fishes. San Diego: Academic Press, 1994.

71
GRAHAM, J. B., LAI, N.C.,CHILLER, D. & ROBERTS J. L. The transition to air breathing
in fishes. Comparative aspects of cardio respiratory regulation in Synbranchus marmoratus
and Monopterus albus (Synbranchidae). J. Exp. Biol., v. 198, pp. 1455 – 1467, 1995.
GOULDING, M.; CARVALHO, M. L.; FERREIRA, E. J. G. 1988. Rio Negro. Rich
life in poorwater. SPB Academic Publishing, The Hague. 200 p.
GURGEL, H. C. B.; ALMEIDA, R. G.; BARBIERI, G.;1994.Análise qualitativa da
alimentação e o coeficiente intestinal de Metynnis cf. Roosevelt Eigenmann, 1915
(Characidae, Myleinae), da lagoa Redonda, Nísia Floresta, Rio Grande do Norte, Brasil.
Revista Brasileira Zoologia 11(2):331-339.
GURGEL, H. C. B., B.W. F. MOLINA, L.J. SOARES & B. CANAN.1995. Ictiofauna
de semi-árido do Rio Grande do Norte. P. 010-011. XI Encontro Brasileiro de
Ictiologia. Campinas, Pontifícia Universidade Católica de Campinas/ Sociedade
Brasileira de Ictiologia.
GURGEL, H.C.B.; ALMEIDA, R.G.; BARBIERI, G.& VIEIRA, L. J. S. 1998. Dieta de
Crenicichla lepidota Heckel, 1840 (Perciformes, Cichlidae) da lagoa Redonda, Nísia
Floresta/RN. Acta Scientiarum 20 (2): 191-194.
GURGEL, H. C. B. & CANAN, B. 1999. Feeding of six species in Jiqui Lagoon,
eastern coast of Rio Grande do Norte, Brasil. Acta Scientiarum 21 (2): 243-246.
GUILLEN, A.L.& T.HART.1980. Feeding interrelationships between the sand shiner
and the striped shiner. Ohio j. Sci. 80 (2):71-76.
HYNES, H.B.N. 1950. The food of freshwater stickle back (Gasterosteus aculeatus and
Pungosteus pungitius) with a review of methods used in studies of the food fishes. Jour.
Anim.Ecol.19 (1):35-38.
HYSLOP. E.J. 1980. Stomach contents analysis – a review of methods and their
application. J. Fish Biol.;17 (4):411-429.

72
HERNANDEZ-BLÁZQUEZ, F. J.; FERREIRA, N.; CARVALHO, C. A. F. de &
FERRI, S. Morphological aspects of the esophagus and stomach of a freshwater
iliophagus fish under scanning electron microscopy. Rev. Gengebaurs Morphol. Jahrb.,
136(1) 1990. p. 119-125.
HAHN, N. S. (1991). Alimentação e dinâmica da nutrição da curvina Plagioscion
squamosissimus (Heckel, 1840) (Pisces, Perciformes) e aspectos da estruturas trófica da
ictiofauna acompanhante no rio Paraná. Tese de Doutorado. Rio Claro: UNESP. 497p.
HAHN, S. N., FUGI, R., ALMEIDA, V. L. L., RUSSO, M. R. & LOUREIRO, V.E.
1997. Dieta e atividade alimentar de peixes do reservatório de Segredo, p.141-162.in:
EDUEM. Reservatório do Segredo: bases ecológicas para o manejo. Maringá.
Universidade Estadual de Maringá. 387p.
HAHN, N. S. (1991). Alimentação e dinâmica da nutrição da curvina Plagioscion
squamosissimus (Heckel, 1840) (Pisces, Perciformes) e aspectos da estruturas trófica da
ictiofauna acompanhante no rio Paraná. Tese de Doutorado. Rio Claro: UNESP. 497p.
HAHN, S. N., FUGI, R., ALMEIDA, V. L. L., RUSSO, M. R. & LOUREIRO, V.E.
1997. Dieta e atividade alimentar de peixes do reservatório de Segredo, p.141-162.in:
EDUEM. Reservatório do Segredo: bases ecológicas para o manejo. Maringá.
Universidade Estadual de Maringá. 387p.
KENNY, J. S., Views from the bridge: a memoir on the freshwater fishes of Trinidad.
St Joseph, Trinidad and Tobago, v. 3, pp. 98, 1995
LAUDER, G. V., LIEM, K. F. (1983) The evolution and interrelationships of the
actinopterygian fishes. Bull. Mus. Comp. Zoo. 150: 95-197.
LAUGHLIN, D.R. & WERNER, E.E.1980. Resource partitioning in two coexisting
sunfish. J. Fish.Res.Bd.Canadá 13 (3):327-342.
LARKIN, P.A. Interespecific competition and population of water fish. J. Fish Res. Bd.;
Ottawa, v. 13, n.3, p. 327-342, 1956.

73
LOWE-McCONNELL, R.H. 1987. Ecological studies in tropical fish communities.
Cambridge, Cambridge University Press. 382p.
LOWE-MCCONNELL, R.H.1999. Estudos Ecológicos de Peixes Tropicais. São Paulo:
Editora da Universidade de São Paulo, 536p.
LO NOSTRO, F. L. (2000). Espermatogénesis, ciclo anual e inducción de la
espermiación en el pez protogínico diándrico, Synbranchus marmoratus Bloch 1795
(Teleostei, Synbranchidae) Buenos Aires, 138p. Tesis (Doctoral) – Depto de Cs
Biologicas, Facultad de Ciências Exactas y Naturales, Universidade de Buenos Aires.
LO NOSTRO, F. L.; G. A. GUERRERO.1986. Presence of primary and secondary
males in a population of the protogynous Synbranchus marmoratus. Journal of Fish
Biology 49, 788–800.
MACNEILL, D.B. & S.B. BRANDT.1990. Ontogeneticshifts in gill-raker morphology
and predicted prey capture efficiency of the alewife Alosa Pseudoharengus . Copeia,
Carbondale, 1: 164-171.
MAIA, V. 1979 Técnica Histológica. 2. ed. São Paulo: Atheneu. p.70-136.
MANTEIFEL, B.P.;I.I. GIRSA & D.S. PAVLOV.1978. On rhytms of fish behaviour, p.
215-224. In:J.E. THORPE (Ed). Rhytmic activity of fishes. London, Academic Press,
312p.
MANN, R.H.K. 1973.Observations on the age, growth, reproduction and food of the
roach, Rutilus rutilus (L.), in two rivers in Southern England. J. Fish Biol. 5, 707-736.
MANN, R.H.K. & ORR, D.R. 1969. A preliminary study of the feeding relationships of
fish in a hard-water and a soft-water stream in Southern England. J. Fish Biol. 1, 31-44.
MÉRONA, B. & BITTENCOURT, M.M. 1993. Fish communities of the lago do Rei, a
floodplain lake in the Central Amazon. A general description. Amazoniana, XII:415-
442.

74
MENEZES, N.A. Importância da Conservação da Ictiofauna dos Ecossistemas
aquáticos brasileiros in Seminário sobre fauna aquática e o setor elétrico brasileiro,
reuniões temáticas prepartórias: caderno 3 – conservação, Curitiba, 23-24 nov.; 1993/
Comitê Coordenador das Atividades do Meio Ambiente do Setor Elétrico – COMASE -
Rio de Janeiro: ELETROBRÁS, 40p.1994.
MEDEIROS, A. P. T. Ecologia, diversidade e aspectos reprodutivos da ictiofauna de
valor comercial da lagoa de Extremoz, RN. Dissertação (Mestrado em Bioecologia
Aquática), Universidade Federal do Rio grande do Norte, Natal, RN. 75p, 2001.
MEDEIROS, A. P. T.; CHELLAPPA, N.T. & CHELLAPPA, S. Aspectos reprodutivos do
cangati, Parauchenipterus galeatus, Linnaeus, 1766 (Osteichthyes: Auchenipteridae) da Lagoa
de Extremoz, Rio Grande do Norte. Revista Brasileira de Zoologia. 20 (4), 647-650, 2003.
MEDEIROS, A. P. T.; CHELLAPPA, S. & CHELLAPPA, N.T. Período reprodutivo da
pirambeba, Serrasalmus maculatus Kner, 1858 (Osteichthyes: Characidae) na Lagoa de
Extremoz, Rio Grando do Norte. p. 89 – 95. In: Ecologia Aquática Tropical Ed. N. T.
Chellappa S. Chellappa & J.Z.O. Passavante. Editora: Editora Serv-Graf, Natal / RN.
165p, 2004.
MENESCAL, R.A. Efeitos da Introdução da Tilápia do Nilo, Oreochromis Niloticus,
(Linnaeus, 1758) (Perciformes, Cichlidae) Sobre o Desembarque Pesqueiro no Açude
Marechal Dutra, Acari/RN. 49f. Dissertação (Mestrado em Bioecológia Aquática) –
Departamento de Oceanografia e Limnologia, Universidade Federal do Rio Grande do
Norte, Natal, 2002.49 f.
MENEZES, R. S. & MENEZES, M. F. 1946. Notas sobre o regime alimentar de algumas
espécies ictiológicas de água doce do Nordeste. Revista Brasileira Biologia 6(4): 537-542.
MORRISON, C.M. & WRIGHT Jr, J.R. 1999 A study of the histology of the digestive
tract of the Nile tilapia. J. Fish Biol., 54: 597-606.
MAGALHÃES, E. M.; ALMEIDA, R. G.; GURGEL, H. C. B. & BARBIERI, G. 1990.
Contribuição ao estudo da alimentação de Serraasalmus brandtii Reinhardt, 1874

75
(Characidae, Serrasalmidae) do rio Piranhas-açu, pendências, Rio Grande do Norte.
Revista Ceres 37(213):429-442.
NASCIMENTO, M. M. Biologia reprodutiva do curimatã comum, Prochilodus brevis
Steindachner, 1875 e limnologia do açude Marechal Dutra localizada na Caatinga do
Rio Grande do Norte. Dissertação (Mestrado em Ciências Biológicas), Universidade
Federal do Rio grande do Norte, Natal, RN. 85p. 2006.
NELSON, J. S. 1994. Fishes of the World. John Wiley & Sons,New York, 600 pp
NIKOLSKY, G.V. The ecology of fishes. Londres: Academic Press, 1963.
NIKOLSKY, G. V. 1978. The ecology of fishes, T.F.H. Publications, Neptune City. 352 p.
PILLAY, T.V.R. 1952. A critique of the methods of study of food of fish. Jour. Zool.
Soc. India, Orissa, 4:185-200.
RIBEIRO, C.A.O. & FANTA. E. 2000 Microscopic morphology and histochemistry of
the digestive system of a tropical freshwater fish Trichomycterus brasiliensis (Lütken
Trichomycteridae). Revta bras. Zool., 17(4): 953-971.
RAPOSO, R. M. G. & GURGEL, H. C. B. 2003. Variação da alimentação natural de
Serrasalmus spilopleura Kner, 1860 (Pisces, Serrasalmidae) em função do ciclo lunar e
das estações do ano na lagoa de extremoz, Rio Grande do Norte, Brasil. Acta
Scientiarum. Animal Sciences 25(2): 267 – 272.
ROSA, R. S., MENEZES, N.A., BRITSKI, H.A., COSTA, W.J.E.M. & GROTH, F.
Diversidade, padrões de distribuição e conservação dos peixes da Caatinga. P. 135 –
180. In: I.R. Leal, M. Tabarelli & J.M.C. da Silva (Eds.) Ecologia e Conservação da
Caatinga. Editora Universitária da Universidade Federal de Pernambuco, Recife . 2003.
ROSA, R.S. 2004. Diversidade e conservação dos peixes da Caatinga. In: J.M.C. da
Silva, M. Tabarelli, M.T. Fonseca, L.V. Lins (Orgs.) Biodiversidade da Caatinga: Áreas
e Ações Prioritárias para a Conservação. Ministério do Meio Ambiente, Brasília-DF.

76
ROBINS C. R.; BAILEY R. M.; BOND C. E.; BROOKER J. R.; LACHNER E. A.;
LEA R.N. & SCOTT W. B. World fishes important to North Americans. Exclusive of
species from the continental waters of the United States an Canada. Am. Fish. Soc.
Spec.Publ., v 243, pp. 91, 1991.
ROSEN, D. E., GREENWOOD P. H. (1976). A fourth neotropical species of Synbranchidae
eel and phylogeny and systematics fishes. Bull. Am. Mus. Nat. Hist. 157:1-69.
RUSSO, M.R. et al. Feeding habits of two species of Bryconamericus Eigenmann,
1907 of the iguaçu river, Brazil. Acta Sci., Maringá, v. 26. N.4, p.431-426, 2004.
SILVA, M.M. Desembarque Pesqueiro das Principais Espécies da Ictiofauna da
Barragem Marechal Dutra, Acari/ RN. 2000. 120f. Dissertação (Mestrado em
Bioecologia Aquática) – Departamento de Oceanografia e Limnologia, Universidade
Federal do Rio Grande do Norte, Natal, 2000.
SILVA, D. A. Ecologia alimentar e reprodutiva da piaba-do-rabo-amarelo, Astyanax cf.
lacustris, (Reinhardt, 1874) (Osteichthyes: Characidae) na Lagoa do Piató, Assu, RN,
Brasil. Dissertação (Mestrado em Bioecologia Aquática), Universidade Federal do Rio
grande do Norte, Natal, RN. 106p, 2008.
SOARES,M.G.M.; ALMEIDA.; R.G, R.G.; JUNK, W. J.The trophic status of the fish
fauna in Lago Camaleão, a macrophyte dominated floodplain lake in the middle
Amazon. Amazoniana, v. IX n.4, p.511-526,1986.
TEIXEIRA, J. L. A.; GURGEL, H. C .B. 2004. Dinâmica da nutrição e alimentação
natural de Steindacnerina notonota (Miranda-Ribeiro, 1937) (Pisces, Curimatidae),
Açude de Riacho da Cruz, Brasil, Brasil. Brasil. Ver. Bras. Zoociências. 6 (1): 19-28
VEGAS-VELEZ, M. La structure histologique typique du tube digestif des poissons
télèosteens. Tethys, 4(1): 1972. p. 163- 174.
WELCOMME (1995), R.L. Fisheries ecology of floodplai Rivers. London: Longman, 1973.

77
WYNES, D.L. & WISSING, T.E.1982. Resource sharing among daters in an ohio
stream.Amer.Midl.Nat.107 (2):294-304.
WATSON, D.J. & BALON, E.K. 1984. Ecomorphological analyses of fish taxocenes
in rainforest of northern Borneo, Jounal of Fish Biology 25, 371-384.
WEATHERLEY, A.H. & GILL, H.S. 1987. Feeding relations, correlated functional
Morphology, growth and size. London: Academic Press.
WOO, R.J., EVANS., G.W., MILLS, L.A (1978). Annual cycle in female three-spined
sticklebacks (Gasterosteus aculeatus L.) from an upland and lowland population. J. Fish
Biol., 12:331-343.
WATSON, D.J. & BALON, E.K. 1984. Ecomorphological analyses of fish taxocenes
in rainforest of northern Borneo, Jounal of Fish Biology 25, 371-384.
WEATHERLEY, A.H. & GILL, H.S. 1987. Feeding relations, correlated functional
Morphology, growth and size. London: Academic Press.
WOOTTON, R. J. 1990. Ecology of teleost fishes.Chapman and Hall, London. 404 p.
ZARET, T.M.;RAND,A.S. 1972. Competition in tropical stream fishes: Support for the
competitive exclusion principle. Ecology, v.52, n.2, p.336-342.
ZIHLER, F. 1982. Gross morphology and configuration of digestive tracts of Cichlidae
(Teleostei, perciformes): phylogenetic and functional significance. Neth. Jour. Zool.
32(4): 544-571.
ZAVALA-CAMIN, L. A. Introdução aos estudos sobre alimentação natural em peixes.
Maringá, EDUEM.129 p., 1996.






















