Embed Size (px)
Citation preview

INFECriON AND IMMUNITY, Mar. 1994, p. 915-921 Vol. 62, No. 300 1 9-9567/94/$04.00+ 0Copyright (C 1994, American Society for Microbiology
Biofilm Formation by Candida Species on the Surface ofCatheter Materials In Vitro
STEPHEN P. HAWSER AND L. JULIA DOUGLAS*
Department of Microbiology, University of Glasgow, Glasgow G12 8QQ, United Kingdom
Received 18 October 1993/Returned for modification 10 November 1993/Accepted 28 December 1993
A model system for studying Candida biofilms growing on the surface of small discs of catheter material isdescribed. Biofilm formation was determined quantitatively by a colorimetric assay involving reduction of atetrazolium salt or by [3H]leucine incorporation; both methods gave excellent correlation with biofilm dryweight (r = 0.997 and 0.945, respectively). Growth of Candida albicans biofilms in medium containing 500 mMgalactose or 50 mM glucose reached a maximum after 48 h and then declined; however, the cell yield was lowerin low-glucose medium. Comparison of biofilm formation by 15 different isolates of C. albicans failed to revealany correlation with pathogenicity within this group, but there was some correlation with pathogenicity whendifferent Candida species were tested. Isolates of C. parapsilosis (Glasgow), C. pseudotropicalis, and C. glabrataall gave significantly less biofilm growth (P < 0.001) than the more pathogenic C. albicans. Evaluation ofvarious catheter materials showed that biofilm formation by C. albicans was slightly increased on latex orsilicone elastomer (P < 0.05), compared with polyvinyl chloride, but substantially decreased on polyurethaneor 100% silicone (P < 0.001). Scanning electron microscopy demonstrated that after 48 h, C. albicans biofilmsconsisted of a dense network of yeasts, germ tubes, pseudohyphae, and hyphae; extracellular polymericmaterial was visible on the surfaces of some of these morphological forms. Our model system is a simple andconvenient method for studying Candida biofilms and could be used for testing the efficacy of antifungal agentsagainst biofilm cells.
In recent years, the use of medical implants such as cathe-ters, pacemakers, prosthetic heart valves, and joint replace-ments has increased dramatically. These devices can becomecolonized by microorganisms which form a biofilm consistingof a mono- or multilayer of cells embedded within a matrix ofextracellular polymeric material (3, 4, 9, 11, 14, 15, 22).Release of microorganisms from the biofilm may initiate anacute disseminated infection. Implant-associated infections aredifficult to resolve, because biofilm microorganisms are resis-tant both to host defense mechanisms and antibiotic therapy(1, 4, 15). Frequently, the implant must be removed.Although the majority of implant infections are caused by
gram-positive bacteria, notably staphylococci, infections due togram-negative bacteria and fungi tend to be more serious (9).Fungal infections are most commonly caused by the patho-genic Candida species, particularly C. albicans, C. tropicalis,and C. parapsilosis (5). These organisms are regarded asincreasingly important nosocomial pathogens. A survey carriedout between 1983 and 1986 established that Candida blood-stream infections represented 10% of all nosocomial blood-stream infections recorded in a university hospital (31). Theattributable mortality rate was 38%. The majority of nosoco-mial septicemias caused by Candida species derive from intra-vascular catheters; more than 40% of all nosocomial septice-mias occurring in intensive care unit patients are infusionrelated (14, 18). Urinary catheters, prosthetic heart valves,cardiac pacemakers, silicone voice prostheses, endotrachealtubes, and cerebrospinal fluid shunts are also strongly associ-ated with Candida infections (25).
Bacterial biofilms have received increasing attention overthe last decade (3, 22), and a number of model systems have
* Corresponding author. Mailing address: Department of Microbi-ology, University of Glasgow, Glasgow G12 8QQ, United Kingdom.Phone: 041-339-8855 (x5842). Fax: 041-330-4600.
been devised for studying the colonization of various solidsurfaces by bacteria. These include the use of agar surfaces(24), submerged test pieces (8), the tubular Robbins device(21), and perfused biofilm fermenters (13). Investigations withpathogenic bacteria have shown that biofilm bacteria in vitrohave a substantially reduced sensitivity to clinically importantantibiotics compared with cells of the same organism in thedispersed form (4). With the aminoglycoside antibiotic tobra-mycin, for example, sensitivities of biofilm bacteria (strains ofPseudomonas aeruginosa) were decreased at least 1,000-fold(24). There is now direct evidence that bacterial growth rate,nutrient limitation, and reduced drug penetration into biofilmsall play a role in this resistance to antibiotics (1).
Formation of a biofilm on an implant surface dependsinitially on the ability of the microorganism to adhere. Adhe-sion of pathogenic Candida species to a variety of hostsurfaces, including inert materials, has received considerableattention (2, 10). However, there have been no reports to dateof studies with Candida biofilms. In this investigation, we havedevised a simple model system for Candida species by adaptingthe method described by Prosser et al. (27), previously used forEscherichia coli and other gram-negative bacteria. This in-volved forming biofilms on the surfaces of small discs cut fromdifferent types of catheters. Growth of the biofilms was mon-itored quantitatively by using dry weight, colorimetric, andradioisotope assays and was visualized by scanning electronmicroscopy (SEM).
MATERIALS AND METHODS
Organisms. Fifteen isolates of C. albicans were used in thisstudy. The origin of six (GRI 681, GRI 682, GDH 2346, GDH2023, NCPF 3153, and Outbreak) is described elsewhere (29).Isolates GRI 272, GRI 922, and GRI 2773 were obtained fromwomen with vaginal candidiasis at Glasgow Royal Infirmary;isolates AAHB 59, AAHB 72, and AAHB 92 were obtained
915
on May 10, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

916 HAWSER AND DOUGLAS
from leukemic patients with line infections or patients withinfected aortic grafts at Crosshouse Hospital, Kilmarnock,Scotland. C. albicans 1001 (26) and two mutants derived fromit unable to form yeasts (strain 1001-FR) or hyphae (strain1001-92') were kindly supplied by C. Gil, Universidad Com-plutense, Madrid, Spain.
C. parapsilosis AAHB 4479, C. tropicalis AAHB 73, and C.glabrata AAHB 12 were isolated from patients with lineinfections at Crosshouse Hospital. C. tropicalis (Londonstrain), C. krusei (Glasgow strain), C. parapsilosis (Glasgowstrain), and C. pseudotropicalis (Glasgow strain) were fromclinical specimens and came from the Regional MycologyReference Laboratory, Glasgow, Scotland. All strains weremaintained on slopes of Sabouraud dextrose agar (Difco) andsubcultured monthly. Every 2 months, cultures were replacedby new ones freshly grown from freeze-dried stocks.Medium and culture conditions. All strains were grown in
yeast nitrogen base medium (Difco) containing either 50 mMglucose or 500 mM galactose as described previously (19).Batches of medium (20 ml, in 100-ml Erlenmeyer flasks) wereinoculated from fresh culture slopes and incubated at 37°C inan orbital shaker at 60 rpm. All strains and species, with theexception of the C. tropicalis (London) isolate, grew exclusivelyin the budding-yeast phase under these conditions. In contrast,the C. tropicalis (London) isolate grew predominantly in thepseudohyphal phase. Cells were harvested after 24 h andwashed twice in 0.15 M phosphate-buffered saline (PBS) pH7.2. Before use in biofilm experiments, all washed cell suspen-sions were adjusted to an optical density of 0.8 at 520 nm.
Biofilm formation. Discs of catheter material (surface area,0.5 cm2) were cut from catheters, sterilized with ethylene oxide(Caledonian Medical Ltd., Glasgow, United Kingdom) andplaced in wells of 24-well Nunclon tissue culture plates.Standardized cell suspension (80 ,ll) was applied to the surfaceof each disc, and the discs were incubated for 1 h at 37°C(adhesion period). Nonadherent organisms were removed bygentle washing with 0.15 M PBS (5 ml), and the discs were thenincubated for up to 72 h at 37°C, submerged in 1 ml of yeastnitrogen base medium containing either 50 mM glucose or 500mM galactose (biofilm formation). In control experiments,discs without cells were incubated in medium containing either50 mM glucose or 500 mM galactose. All biofilm and controlassays were carried out three times in triplicate.
Quantitative measurement of biofilm growth. (i) Determi-nation of dry weights. After biofilm formation, the discs wereremoved from the medium and gently washed with 0.15 M PBS(5 ml) to remove nonbiofilm cells. Biofilm organisms werescraped from the discs with a sterile scalpel and vortexed gentlyfor 3 min. The discs were washed in 0.15 M PBS (5 ml) toremove any remaining cells. All organisms were collected onpreweighed cellulose nitrate filters (0.45-,um pore size; 25-mmdiameter) and given three washes with water (5 ml). The filterswere dried to constant weight at 80°C, and the dry weights ofcells per disc were calculated. Dry-weights were determinedthree times in triplicate.
(ii) Incorporation of [3H]leucine. The method used formeasuring [3H]leucine incorporation was a modification of oneused for labelling biofilms of E. coli (8). After biofilm forma-tion, L-[4,5-3H]leucine in PBS (final concentration, 1 ,uCi/ml;120 Ci/mmol; Amersham) was added to each well, and theplates were incubated for a further 4 h at 37°C. Separateexperiments showed that the uptake of radioactive label bybiofilm cells was linear during this time. After labelling,organisms were removed from the discs by incubating the discsin wells containing 0.5 ml of 5% (wt/vol) sodium dodecylsulfate (SDS) for 3 h at 37°C. The SDS solution with the cells
was transferred to tubes containing 0.5 ml of 10% (wt/vol)trichloroacetic acid supplemented with 1% (wt/vol) CasaminoAcids (Difco). Each trichloroacetic acid precipitate was fil-tered through a prewetted cellulose nitrate filter (0.45-pLmpore size; 25-mm diameter). The filters were washed threetimes with 5 ml of 5% (wt/vol) trichloroacetic acid containing1% (wt/vol) Casamino Acids and were dried, and radioactivitywas counted in 4 ml of Opti-Fluor 0 scintillant (Packard).Filters without cells were used to determine backgroundcounts. All [3H]leucine incorporation assays were carried outthree times in triplicate.
(iii) Tetrazolium reduction assay. The tetrazolium salt3-[4,5-dimethylthiazol-2-yl]-2,5-diphenyltetrazolium bromide(MTT; Sigma) was used in an assay modified from thatdescribed by Mosmann (23). After biofilm formation, 50 ,ul ofMTT solution (a stock solution containing 5 mg ofMTT per mlof PBS, diluted 1:5 in prewarmed 0.15 M PBS prior toapplication) was added to each well. After incubation for 5 h at37°C, the medium plus MTT was removed, and the wells werewashed three times with 0.15 M PBS (2 ml) to remove all tracesof MTT. Dimethyl sulfoxide (1 ml) was then added to solubi-lize the MTT formazan product. MTT formazan formation wasmeasured at 540 nm by using a Shimadzu spectrophotometer.Control wells contained medium plus MTT to determinebackground formazan values. All assays were carried out threetimes in triplicate.SEM. Biofilms of C. albicans GDH 2346 formed on polyvinyl
chloride (PVC) catheter discs were fixed with 2.5% (vol/vol)glutaraldehyde in 0.15 M PBS for 1 h at room temperature.They were then treated with 1% (wt/vol) osmium tetroxide for1 h, washed three times with distilled water (3 ml), treated with1% (wt/vol) uranyl acetate for 1 h, and washed again withdistilled water (3 ml). The samples were dehydrated with aseries of ethanol solutions which ranged, in 10% increments,from 30% (vol/vol) ethanol in distilled water to dried absoluteethanol. All samples were dried to critical point by using aPolaron critical point drier, coated with gold with a Polaroncoater, and viewed under a Philips 500 scanning electronmicroscope.
Catheter materials. Different types of catheter were kindlydonated by a number of manufacturers. Latex urinary Foleycatheters and PVC central venous catheters were from RuschUK Ltd., High Wycombe, United Kingdom; silicone elas-tomer-coated latex urinary Foley catheters were provided bySimpla Plastics Ltd., Cardiff, United Kingdom; and siliconeurinary Foley catheters and polyurethane central venous cath-eters were from Vygon (UK) Ltd., Cirencester, UnitedKingdom.
RESULTS
Measurement of biofilm growth. Initially, biofilm growth wasmonitored by using a viable count procedure first described forbacterial biofilms (27). However, this method failed to givereproducible results when it was applied to C. albicans (datanot shown). Two alternative assays were therefore used; theyinvolved (i) measuring the uptake of [3H]leucine by biofilmcells, and (ii) determining the ability of the cells to reduce atetrazolium salt, MTT. Both assays appeared satisfactory, sincethere was a strong correlation between biofilm dry weight andeither [3H]leucine incorporation (r = 0.945) or MTT formazanformation (r = 0.997) (Fig. 1). By either assay method, growthof biofilms of C. albicans GDH 2346 in galactose-containingmedium reached a maximum after 48 h. Thereafter, cellnumbers appeared to decline (Fig. 1).Comparison of galactose-grown and glucose-grown biofilms.
INFECT. IMMUN.
on May 10, 2018 by guest
http://iai.asm.org/
Dow
nloaded from
CANDIDA BIOFILMS ON CATHETERS 917
2.0 to(a)
E 1.5
5 ~~~~~~~~~~~~~~61.0
4E
0.5
0.0 (I\ . I14{t L o
E
L.
E,_
._1
0 20 4 60
Time (h)
2.0
1.5
1.0-
0.5 ]
(b)
E
~1.2 E
tn
-0.8 C_
E
-0.4NI
E0.0oo
0 20 40 60 x()Time (h)
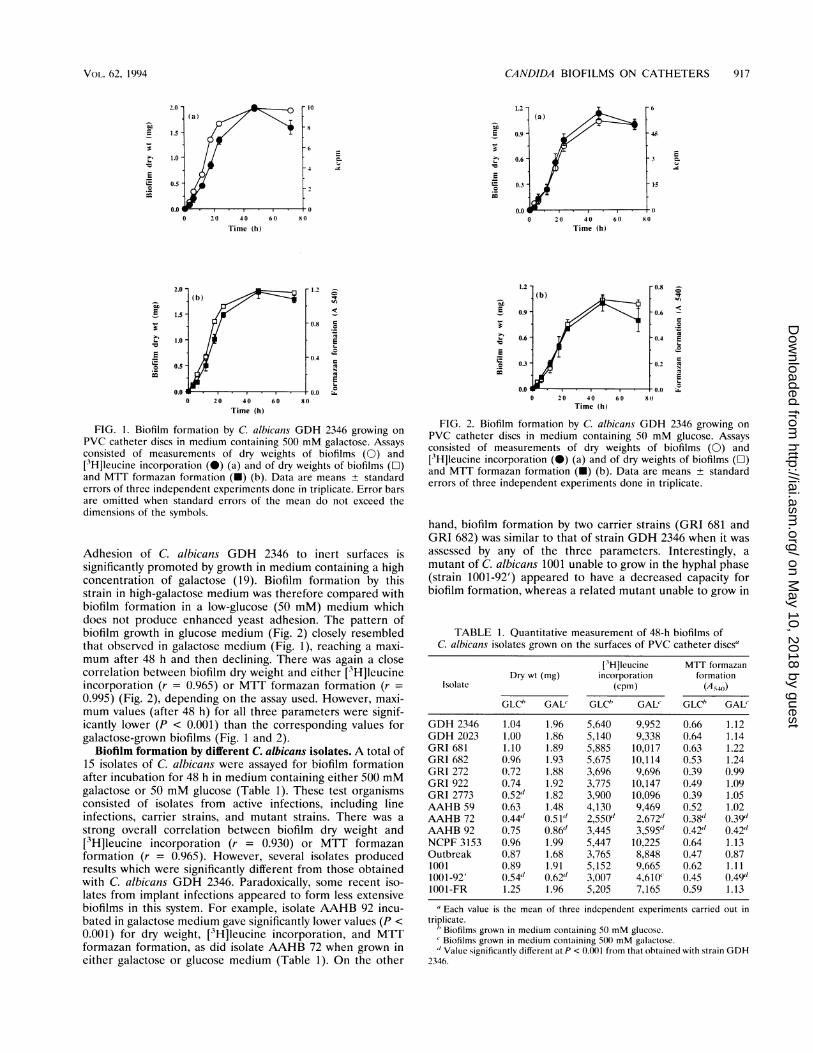
FIG. 1. Biofilm formation by C. albicans GDH 2346 growing on
PVC catheter discs in medium containing 500 mM galactose. Assaysconsisted of measurements of dry weights of biofilms (0) and[3H]Ieucine incorporation (0) (a) and of dry weights of biofilms (E)and MIT formazan formation (-) (b). Data are means ± standarderrors of three independent experiments done in triplicate. Error barsare omitted when standard errors of the mean do not exceed thedimensions of the symbols.
Adhesion of C. albicans GDH 2346 to inert surfaces issignificantly promoted by growth in medium containing a highconcentration of galactose (19). Biofilm formation by thisstrain in high-galactose medium was therefore compared withbiofilm formation in a low-glucose (50 mM) medium whichdoes not produce enhanced yeast adhesion. The pattern ofbiofilm growth in glucose medium (Fig. 2) closely resembledthat observed in galactose medium (Fig. 1), reaching a maxi-mum after 48 h and then declining. There was again a closecorrelation between biofilm dry weight and either [3H]leucineincorporation (r = 0.965) or MIT formazan formation (r =
0.995) (Fig. 2), depending on the assay used. However, maxi-mum values (after 48 h) for all three parameters were signif-icantly lower (P < 0.001) than the corresponding values forgalactose-grown biofilms (Fig. 1 and 2).
Biofilm formation by different C. albicans isolates. A total of15 isolates of C. albicans were assayed for biofilm formationafter incubation for 48 h in medium containing either 500 mMgalactose or 50 mM glucose (Table 1). These test organismsconsisted of isolates from active infections, including lineinfections, carrier strains, and mutant strains. There was a
strong overall correlation between biofilm dry weight and[3H]leucine incorporation (r = 0.930) or MIT formazanformation (r = 0.965). However, several isolates producedresults which were significantly different from those obtainedwith C. albicans GDH 2346. Paradoxically, some recent iso-lates from implant infections appeared to form less extensivebiofilms in this system. For example, isolate AAHB 92 incu-bated in galactose medium gave significantly lower values (P <0.001) for dry weight, [3H]leucine incorporation, and MTTformazan formation, as did isolate AAHB 72 when grown ineither galactose or glucose medium (Table 1). On the other
1.2-
E (a)
E 0.9
0.6-
0.3q-
0.0I
0 20 40 60Time (h)
E
E0
L
0 20 40 60Time (h)
-6
-4'
-3)
0800
aIT
E
E0
FIG. 2. Biofilm formation by C. albicans GDH 2346 growing onPVC catheter discs in medium containing 50 mM glucose. Assaysconsisted of measurements of dry weights of biofilms (0) and['3H]leucine incorporation (0) (a) and of dry weights of biofilms (1)and MIT formazan formation (U) (b). Data are means ± standarderrors of three independent experiments done in triplicate.
hand, biofilm formation by two carrier strains (GRI 681 andGRI 682) was similar to that of strain GDH 2346 when it wasassessed by any of the three parameters. Interestingly, amutant of C. albicans 1001 unable to grow in the hyphal phase(strain 1001-92') appeared to have a decreased capacity forbiofilm formation, whereas a related mutant unable to grow in
TABLE 1. Quantitative measurement of 48-h biofilms ofC. albicans isolates grown on the surfaces of PVC catheter discs'
['3Hileucine MTT formazanDry wt (mg) incorporation formation
Isolate (cpm) (A540)
GLCb GAL' GLCh GAL' GLCh GAL'
GDH 2346 1.04 1.96 5,640 9,952 0.66 1.12GDH 2023 1.(0 1.86 5,140 9,338 0.64 1.14GRI 681 1.10 1.89 5,885 10,017 0.63 1.22GRI 682 0.96 1.93 5,675 10,114 0.53 1.24GRI 272 0.72 1.88 3,696 9,696 0.39 0.99GRI 922 0.74 1.92 3,775 10,147 0.49 1.09GRI 2773 0.52" 1.82 3,900 10,096 0.39 1.05AAHB 59 0.63 1.48 4,130 9,469 0.52 1.02AAHB 72 0.44"d 0.51" 2,550" 2,672"d 0.38" 0.39"AAHB 92 0.75 0.86d 3,445 3,595"d 0.42"d 0.42"NCPF 3153 0.96 1.99 5,447 10,225 0.64 1.13Outbreak 0.87 1.68 3,765 8,848 0.47 0.871001 0.89 1.91 5,152 9,665 0.62 1.111001-92' 0.54"d 0.62" 3,007 4,610' 0.45 0.49"1001-FR 1.25 1.96 5,205 7,165 0.59 1.13
" Each value is the mean of three independent experiments carried out intriplicate.
" Biofilms grown in medium containing 5(1 mM glucose.Biofilms grown in medium containing 500 mM galactose.
d Value significantly different at P < 0.001 from that obtained with strain GDH2346.
VOL. 62? 1994
0
on May 10, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

918 HAWSER AND DOUGLAS
TABLE 2. Quantitative measurement of 48-h biofilms of Candidaspecies grown on the surfaces of PVC catheter discsa
[3H]leucine MaTDry wt (mg) incorporation formation
Isolate (cpm) (oratio)(A 540))
GLCb GAL' GLCh GALC GLCb GALC
C. albicans (GDH 2346) 1.04 1.96 5,640 9,952 0.67 1.12C. parapsilosis (AAHB 4479) 0.74 1.35 5,425 8,976 0.57 0.97C. parapsilosis (Glasgow) 0.41" 0.66" 3,865 4,495" 0.38 0.52C. tropicalis (AAHB 73) 1.10 2.05 6,990 12,260 0.70 1.23C. tropicalis (London) 0.85 1.74 4,790 9,649 0.48 0.55dC. pseudotropicalis (Glasgow) 0.45d 0.89" 3,453" 5,565" 0.46" 0.78C. krusei (Glasgow) 1.30 2.17 6,630 12,175 0.69 1.34C. glabrata (AAHB 12) 0.54d 0.35d 5,332 2,460" 0.39d 0.30"
a Each value is the mean of three independent experiments carried out intriplicate.
b Biofilms grown in medium containing 50 mM glucose.c Biofilms grown in medium containing 500 mM galactose.d Value significantly different at P < 0.001 from that obtained with strain GDH
2346.
the yeast phase (1001-FR) gave values for biofilm formationsimilar to those of the wild type (Table 1).For glucose-grown biofilms, the correlation between dry
weight and [3H]leucine incorporation (r = 0.825) or MTTformazan formation (r = 0.800) was less good than that forgalactose-grown biofilms. This corresponded with a greateramount of variation in biofilm formation within the 15 strainstested. Typically, glucose-grown biofilms gave values for dryweight, [3H]leucine incorporation, and MTT formazan forma-tion which were 35 to 55% lower (P < 0.001) than thoserecorded for galactose-grown biofilms (Table 1).
Biofilm formation by different Candida species. Biofilmformation by five other Candida species was compared withthat of C. albicans (Table 2). With galactose-grown cells, thecorrelation between biofilm dry weight and [3H]leucine incor-poration (r = 0.845) or MTT formazan formation (r = 0.815)was lower than that observed for galactose-grown strains of C.albicans. This finding presumably reflects the greater physio-logical differences between the species. Isolates of C. parapsi-losis (Glasgow), C. pseudotropicalis, and C. glabrata showedsignificantly less biofilm growth (P < 0.001) in medium con-taining either galactose or glucose than did C. albicans GDH2346 (Table 2). However, C. krusei which, like these species, isalso considered less pathogenic than C. albicans, exhibitedgood biofilm growth.For glucose-grown Candida species, the correlation between
biofilm dry weight and [3H]leucine incorporation (r = 0.920) orMTT formazan formation (r = 0.910) was greater than thatobtained with galactose-grown cells. Glucose-grown biofilms ofC. glabrata AAHB 12 gave increased values (P < 0.001) for dryweight, [3H]leucine incorporation, and MTT formazan forma-tion compared with galactose-grown biofilms of the sameorganism (Table 2).
Biofilm formation by C. albicans on different catheter mate-rials. Biofilm formation by C. albicans GDH 2346 on discsprepared from a range of different catheter materials wasexamined (Table 3). Almost identical results were obtainedwith PVC catheters supplied by two separate manufacturers.By comparison with biofilm formation on PVC, biofilm forma-tion on latex was slightly increased (P < 0.05), whereas that onpolyurethane was substantially decreased (P < 0.001). Twotypes of silicone catheter material produced different results;discs prepared from silicone elastomer catheters (which have a
TABLE 3. Effect of different catheter materials on the formation ofbiofilms by C. albicans GDH 2346a
[3tH]eucine formazanDry wt (mg) incorporation formatin
Catheter material (cpm) fAr4mt)
GLCb GAL' GLCb GAL' GLCb GALC
PVC (Riisch) 1.04 1.96 5,640 9,952 0.66 1.12PVC (Vygon) 1.06 2.05 4,995 9,946 0.65 1.22Latex (Rusch) 1.22d 2.36d 6,160" 10,405d 0.88" 1.52"Polyurethane (Vygon) 0.72' 1.53e 4,550' 9,525' 0.55' 1.18'Silicone elastomer (Simpla) 1.16d 2.14d 5,050" 9,005" 0.85" 1.46dSilicone (Vygon) 0.73" 1.52' 4,488' 7,789' 0.48' 1.02'
" Each value is the mean of three independent experiments carried out intriplicate.
b Biofilms grown in medium containing 50 mM glucose.cBiofilms grown in medium containing 500 mM galactose.dValue significantly different at P < 0.05 from that obtained with PVC
(Rusch) material.Value significantly different at P < 0.001 from that obtained with PVC
(Rusch) material.
latex core) showed slightly increased biofilm formation (P <0.05) compared with PVC discs, while discs from 100% siliconecatheters were more resistant (P < 0.001) to fungal coloniza-tion (Table 3). SEM has revealed that the surface topographiesof these two types of silicone catheter are quite distinct (resultsnot shown).SEM. Biofilm formation by C. albicans GDH 2346 on PVC
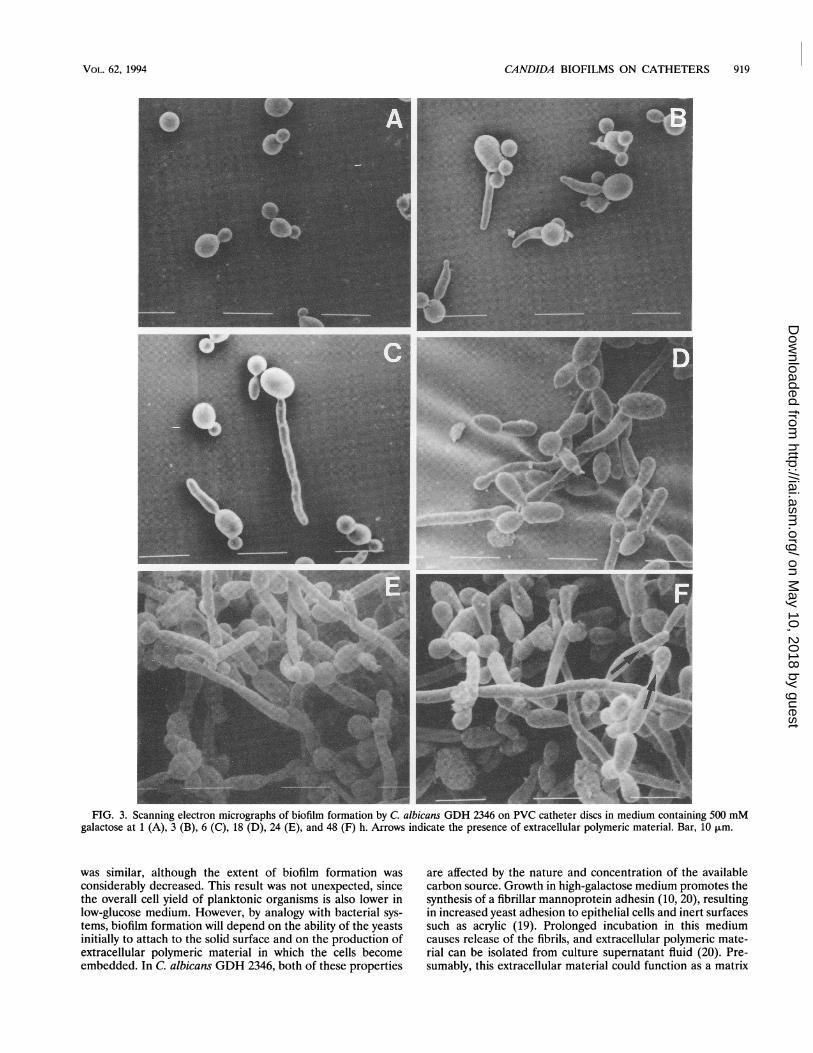
catheter discs was monitored by SEM. Replicate discs wereremoved from galactose medium for processing at intervals ofup to 48 h. After 1 h of incubation, the cell population attachedto the discs consisted exclusively of cells in the budding-yeastphase of growth (Fig. 3). However, after 3 to 6 h, some of thecells had produced germ tubes, and by 18 to 24 h, thepopulation had increased and consisted of a mixture of yeasts,germ tubes, and young hyphae (Fig. 3). The biofilms hadmatured by 48 h and comprised a dense network of yeasts,germ tubes, pseudohyphae, and hyphae. Extracellular poly-meric material (indicated by the arrows in Fig. 3) was visible onthe surfaces of some of these morphological forms.
DISCUSSION
Although a number of experimental model systems havebeen designed to study bacterial biofilms, this is the first reportof a biofilm system for pathogenic fungi. We have developedtwo assays for biofilm growth of Candida species on thesurfaces of small discs of catheter material. In the first assay,which was adapted from a procedure employed originally withE. coli (8), preformed Candida biofilms are labelled withradioactive leucine. The second assay depends on the conver-sion of the yellow tetrazolium salt MTT to a dark-blue product,MTT formazan, as a result of dehydrogenase activity by biofilmcells. This reaction has been used previously to measure theproliferation of various mammalian cell types (23) and todetermine the viability of some microorganisms, includingfungal species such as C. albicans (17). Both assay methodsshowed an excellent correlation between biofilm dry weightand the parameter being measured, and both are rapid andconvenient to carry out.Growth of biofilms of C. albicans GDH 2346 in high-
galactose medium reached a maximum after 48 h and thendeclined, possibly because of depletion of oxygen or nutrientswithin the biofilm. The growth pattern in low-glucose medium
INFEC-F. IMMUN.
on May 10, 2018 by guest
http://iai.asm.org/
Dow
nloaded from
CANDIDA BIOFILMS ON CATHETERS 919
FIG. 3. Scanning electron micrographs of biofilm formation by C. albicans GDH 2346 on PVC catheter discs in medium containing 500 mMgalactose at 1 (A), 3 (B), 6 (C), 18 (D), 24 (E), and 48 (F) h. Arrows indicate the presence of extracellular polymeric material. Bar, 10 jim.
was similar, although the extent of biofilm formation wasconsiderably decreased. This result was not unexpected, sincethe overall cell yield of planktonic organisms is also lower inlow-glucose medium. However, by analogy with bacterial sys-tems, biofilm formation will depend on the ability of the yeastsinitially to attach to the solid surface and on the production ofextracellular polymeric material in which the cells becomeembedded. In C. albicans GDH 2346, both of these properties
are affected by the nature and concentration of the availablecarbon source. Growth in high-galactose medium promotes thesynthesis of a fibrillar mannoprotein adhesin (10, 20), resultingin increased yeast adhesion to epithelial cells and inert surfacessuch as acrylic (19). Prolonged incubation in this mediumcauses release of the fibrils, and extracellular polymeric mate-rial can be isolated from culture supernatant fluid (20). Pre-sumably, this extracellular material could function as a matrix
VOL. 62, 1994
on May 10, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

920 HAWSER AND DOUGLAS
substance in biofilms. Our scanning electron micrographsprovide some evidence for such a matrix material, although itwas not detected in abundant quantities (Fig. 3).
Comparison of biofilm formation by 15 different isolates ofC. albicans failed to reveal any correlation with pathogenicitywithin this group. On the contrary, biofilm formation by twocarrier strains (GRI 681 and GRI 682) was similar to that ofstrains isolated originally from active infections, such as GDH2346 and GDH 2023. Moreover, two recent isolates fromimplant infections (AAHB 72 and AAHB 92) formed less-extensive biofilms than some of the other isolates tested here.This unexpected result may be explained by the incubationconditions used in the present assay. Preliminary experimentsindicate that gentle shaking to produce a flow of liquid over thecatheter disc increases the synthesis of extracellular matrixmaterial by biofilm cells. Such conditions may approximatethose found in vivo more than does static incubation and may,in consequence, favor biofilm formation by certain strains. Weare currently investigating this possibility.There was some correlation with pathogenicity when differ-
ent Candida species were tested for biofilm formation. Isolatesof C. parapsilosis (Glasgow), C. pseudotropicalis, and C. gla-brata all gave significantly less biofilm growth than the more-pathogenic C. albicans. However, a second isolate of C.parapsilosis (AAHB 4479), recently obtained from a lineinfection, showed a much greater propensity for biofilm for-mation than the first strain. Infections due to C. parapsilosis arefrequently associated with indwelling catheters or other pros-thetic devices (30). Both C. glabrata and C. krusei grew poorlyin shake cultures on high-galactose medium, reaching opticaldensities which were 26 and 35%, respectively, of that for C.albicans GDH 2346 (results not shown). A limited ability toassimilate galactose may also partly account for the relativelypoor biofilm formation by C. glabrata in this medium. Bycontrast, C. krusei gave better biofilm growth than C. albicansin both low-glucose and high-galactose medium, an anomalousresult that remains unexplained. Like C. parapsilosis, C. kruseiis now emerging as a significant nosocomial pathogen (5).There is increasing evidence that the chemical nature of
catheter material plays an important role in the pathogenesisof device-related infections. Central venous catheters are theintravascular devices most likely to cause iatrogenic septice-mia; most of these are made of PVC, although various studiesindicate that Teflon or polyurethane catheters are less likely tobecome colonized by bacteria (18). C. albicans is known toadhere to PVC more readily than to Teflon (28). Our resultsnow demonstrate that PVC catheters are more susceptible tobiofilm formation by this yeast than are polyurethane ones.The most extensive biofilm formation by C. albicans wasobserved on latex urinary catheter material. Most Candidaurinary tract infections are nosocomial and occur in patientswith urinary bladder catheters. Duration of catheter placementis directly related to the incidence of candiduria (12). A 100%silicone catheter produced significantly less biofilm formationthan either of the other urinary catheter materials. This may berelated to its surface topography, which appeared exceptionallysmooth when viewed by SEM. However, pure silicone cathe-ters are not ideally flexible and afford a lower degree of patientcomfort than silicone elastomer-coated latex catheters. In vivo,catheters and other prosthetic devices are rapidly coated withhost proteins such as albumin, fibrin, fibrinogen, and fibronec-tin. Some of these proteins are known to affect bacterialadhesion and biofilm formation (9). The biofilm system de-scribed here should facilitate analogous studies with Candidaspecies.A number of reports suggest that certain bacterial genes are
switched on in the presence of a surface. In a study byDagostino et al. (6), for example, transposon mutagenesis wasused to insert promoterless lacZ genes into appropriate recip-ient bacteria, giving some the ability to express ,3-galactosidaseat a polystyrene surface but not in liquid or on agar medium.Recently, Davies et al. (7) described activation of the algCpromoter, which is required for the production of the exo-polysaccharide alginate, by biofilms of P. aeruginosa. Activa-tion resulted from association of the bacteria with a Teflonsubstratum. In the present study of Candida biofilms, weobserved that biofilm cells growing on discs of PVC cathetermaterial exhibited mixed yeast and hyphal morphologies,whereas on an agar surface of the same medium (or in liquidculture), only the yeast form was found. The processes bywhich morphogenesis in C. albicans is regulated are poorlyunderstood at the molecular level; our SEM studies nowsuggest the possible involvement of some form of contact-induced gene expression.One of the most significant features of bacterial biofilms is
their resistance to antimicrobial agents (1, 4). Investigationsinto the mechanisms of resistance indicate that reduced drugpenetration into biofilms, nutrient limitation, and a low bacte-rial growth rate are all contributory factors (1). Preliminaryexperiments carried out in this laboratory have shown thatCandida biofilm cells are similarly less susceptible to a numberof antifungal agents than are planktonic yeasts (16). Animportant application of our model biofilm system will, there-fore, be to characterize the susceptibilities of different Candidabiofilms to the various antifungal agents in current clinical useand possibly also to others in development.
ACKNOWLEDGMENTS
This work was supported by a grant from The Sir Jules ThornCharitable Trust.We are indebted to M. D. Richardson at the Regional Mycology
Reference Laboratory, Glasgow, for identifying several of the isolatesused here and to L. Tetley and M. Mullin for assistance with the SEMstudies.
REFERENCES1. Brown, M. R. W., and P. Gilbert. 1993. Sensitivity of biofilms to
antimicrobial agents. J. Appl. Bacteriol. Symp. Suppl. 74:87S-97S.2. Calderone, R. A., and P. C. Braun. 1991. Adherence and receptor
relationships of Candida albicans. Microbiol. Rev. 55:1-20.3. Characklis, W. G., and P. A. Wilderer. 1989. Structure and
function of biofilms. Wiley, Chichester, United Kingdom.4. Costerton, J. W., K. J. Cheng, G. G. Geesey, T. I. Ladd, J. C.
Nickel, M. Dasgupta, and T. J. Marrie. 1987. Bacterial biofilms innature and disease. Annu. Rev. Microbiol. 41:435-464.
5. Cox, G. M., and J. R. Perfect. 1993. Fungal infections. Curr. Opin.Infect. Dis. 6:422-426.
6. Dagostino, L., A. E. Goodman, and K. C. Marshall. 1991. Physi-ological responses induced in bacteria adhering to surfaces. Bio-fouling 4:113-119.
7. Davies, D. G., A. M. Chakrabarty, and G. G. Geesey. 1993.Exopolysaccharide production in biofilms: substratum activationof alginate gene expression by Pseudomonas aeruginosa. Appl.Environ. Microbiol. 59:1181-1186.
8. Dix, B. A., P. S. Cohen, D. C. Laux, and R. Cleeland. 1988.Radiochemical method for evaluating the effect of antibiotics onEscherichia coli biofilms. Antimicrob. Agents Chemother. 32:770-772.
9. Dougherty, S. H. 1988. Pathobiology of infection in prostheticdevices. Rev. Infect. Dis. 10:1102-1117.
10. Douglas, L. J. 1992. Mannoprotein adhesins of Candida albicans,p. 34-50. In J. E. Bennett, R. J. Hay, and P. K. Peterson (ed.), Newstrategies in fungal disease. Churchill Livingstone, Edinburgh.
11. Elliott, T. S. J. 1988. Intravascular-device infections. J. Med.Microbiol. 27:161-167.
INFECTF. IMMUN.
on May 10, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

CANDIDA BIOFILMS ON CATHETERS 921
12. Gentry, L. O., and M. F. Price. 1993. Urinary and peritonealCandida infections, p. 249-260. In G. P. Bodey (ed.), Candidiasis:pathogenesis, diagnosis and treatment. Raven Press, New York.
13. Gilbert, P., D. G. Allison, D. J. Evans, P. S. Handley, and M. R. W.Brown. 1989. Growth rate control of adherent bacterial popula-tions. Appl. Environ. Microbiol. 55:1308-1311.
14. Goldmann, D. A., and G. B. Pier. 1993. Pathogenesis of infectionsrelated to intravascular catheterization. Clin. Microbiol. Rev.6:176-192.
15. Gristina, A. G. 1987. Biomaterial-centered infection: microbialadhesion versus tissue integration. Science 237:1588-1595.
16. Hawser, S. P., and L. J. Douglas. Unpublished data.17. Levitz, S. M., and R. D. Diamond. 1985. A rapid colorimetric assay
of fungal viability with the tetrazolium salt MTT. J. Infect. Dis.152:938-945.
18. Maki, D. G. 1989. Pathogenesis, prevention and management ofinfections due to intravascular devices used for infusion therapy, p.161-177. In A. L. Bisno and F. A. Waldvogel (ed.), Infectionsassociated with indwelling medical devices. American Society forMicrobiology, Washington, D.C.
19. McCourtie, J., and L. J. Douglas. 1981. Relationship between cellsurface composition of Candida albicans and adherence to acrylicafter growth on different carbon sources. Infect. Immun. 32:1234-1241.
20. McCourtie, J., and L. J. Douglas. 1985. Extracellular polymer ofCandida albicans: isolation, analysis and role in adhesion. J. Gen.Microbiol. 131:495-503.
21. McCoy, W. F., J. D. Bryers, J. Robbins, and J. W. Costerton. 1981.Observations of fouling biofilm formation. Can. J. Microbiol.27:910-917.
22. Melo, L. F., T. R. Bott, M. Fletcher, and B. Capdeville. 1992.Biofilms-science and technology. Kluwer Academic Publishers,Dordrecht, The Netherlands.
23. Mosmann, T. 1983. Rapid colorimetric assay for cellular growthand survival: application to proliferation and cytotoxicity assays. J.Immunol. Methods 65:55-63.
24. Nichols, W. W., M. J. Evans, M. P. E. Slack, and H. L. Walmsley.1989. The penetration of antibiotics into aggregates of mucoid andnon-mucoid Pseudomonas aeruginosa. J. Gen. Microbiol. 135:1291-1303.
25. Odds, F. C. 1988. Candida and candidosis, 2nd ed. BailliereTindall, London.
26. Pomes, R., C. Gil, and C. Nombela. 1985. Genetic analysis ofCandida albicans morphological mutants. J. Gen. Microbiol. 131:2107-2113.
27. Prosser, B. T., D. Taylor, B. A. Dix, and R. Cleeland. 1987. Methodof evaluating effects of antibiotics on bacterial biofilm. Antimi-crob. Agents Chemother. 31:1502-1506.
28. Rotrosen, D., T. R. Gibson, and J. E. Edwards, Jr. 1983. Adher-ence of Candida species to intravenous catheters. J. Infect. Dis.147:594.
29. Sweet, S. P., and L. J. Douglas. 1991. Effect of iron deprivation onsurface composition and virulence determinants of Candida albi-cans. J. Gen. Microbiol. 137:859-865.
30. Weems, J. J. 1992. Candida parapsilosis: epidemiology, pathoge-nicity, clinical manifestations and antimicrobial susceptibility. Clin.Infect. Dis. 14:756-766.
31. Wey, S. B., M. Mori, and M. A. Pfaller. 1988. Hospital-acquiredcandidemia. The attributable mortality and excess length of stay.Arch. Intern. Med. 148:2642-2645.
VOL. 62, 1994
on May 10, 2018 by guest
http://iai.asm.org/
Dow
nloaded from





























