Embed Size (px)
Citation preview

Bioengineered 3D Brain Tumor Model To Elucidate the Effects ofMatrix Stiffness on Glioblastoma Cell Behavior Using PEG-BasedHydrogelsChristine Wang,† Xinming Tong,‡ and Fan Yang*,†,‡
†Department of Bioengineering, Stanford University, Stanford, California 94305, United States‡Department of Orthopaedic Surgery, Stanford University, Stanford, California 94305, United States
ABSTRACT: Glioblastoma (GBM) is the most common and aggressive form of primary brain tumor with a median survival of12−15 months, and the mechanisms underlying GBM tumor progression remain largely elusive. Given the importance of tumorniche signaling in driving GBM progression, there is a strong need to develop in vitro models to facilitate analysis of brain tumorcell-niche interactions in a physiologically relevant and controllable manner. Here we report the development of a bioengineered3D brain tumor model to help elucidate the effects of matrix stiffness on GBM cell fate using poly(ethylene-glycol) (PEG)-basedhydrogels with brain-mimicking biochemical and mechanical properties. We have chosen PEG given its bioinert nature andtunable physical property, and the resulting hydrogels allow tunable matrix stiffness without changing the biochemical contents.To facilitate cell proliferation and migration, CRGDS and a MMP-cleavable peptide were chemically incorporated. Hyaluronicacid (HA) was also incorporated to mimic the concentration in the brain extracellular matrix. Using U87 cells as a model GBMcell line, we demonstrate that such biomimetic hydrogels support U87 cell growth, spreading, and migration in 3D over thecourse of 3 weeks in culture. Gene expression analyses showed U87 cells actively deposited extracellular matrix and continued toupregulate matrix remodeling genes. To examine the effects of matrix stiffness on GBM cell fate in 3D, we encapsulated U87 cellsin soft (1 kPa) or stiff (26 kPa) hydrogels, which respectively mimics the matrix stiffness of normal brain or GBM tumor tissues.Our results suggest that changes in matrix stiffness induce differential GBM cell proliferation, morphology, and migration modesin 3D. Increasing matrix stiffness led to delayed U87 cell proliferation inside hydrogels, but cells formed denser spheroids withextended cell protrusions. Cells cultured in stiff hydrogels also showed upregulation of HA synthase 1 and matrixmetalloproteinase-1 (MMP-1), while simultaneously downregulating HA synthase 2 and MMP-9. This suggests that varyingmatrix stiffness can induce differential ECM deposition and remodeling by employing different HA synthases or MMPs.Furthermore, increasing matrix stiffness led to simultaneous upregulation of Hras, RhoA, and ROCK1, suggesting a potential linkbetween the mechanosensing pathways and the observed differential cell responses to changes in matrix stiffness. Thebioengineered 3D hydrogel platform reported here may provide a useful 3D in vitro brain tumor model for elucidating themechanisms underlying GBM progression, as well as for evaluating the efficacy of potential drug candidates for treating GBM.
KEYWORDS: tumor model, hydrogels, glioblastoma, tumor microenvironment, three-dimensional
■ INTRODUCTIONGlioblastoma (GBM) is the most common and aggressive formof primary brain tumor in adults, accounting for 60−70% ofmalignant glioma with an annual incidence of 14,000 cases peryear in the United States alone.1 Patients with GBM oftenundergo aggressive therapy with a median survival of 12 to 15months.1 Given the poor clinical outcomes of this devastatingdisease, there is a strong need to better understand the
Special Issue: Engineered Biomimetic Tissue Platforms for in VitroDrug Evaluation
Received: January 26, 2014Revised: April 4, 2014Accepted: April 8, 2014
Article
pubs.acs.org/molcularpharmaceutics
© XXXX American Chemical Society A dx.doi.org/10.1021/mp5000828 | Mol. Pharmaceutics XXXX, XXX, XXX−XXX

mechanisms underlying tumor progression. While previousresearch has largely focused on elucidating the intracellularprocesses that drive tumor growth, increasing evidence hashighlighted the tumor microenvironment as a significant playerin influencing GBM cell fate and tumor progression.2,3
The brain tumor microenvironment is a complex anddynamic system, consisting of various biochemical andmechanical cues that act in conjunction to fuel GBM cellgrowth and invasion. Extracellular matrix proteins (ECM), suchas collagen and hyaluronic acid (HA), provide structuralsupport, as well as influence intracellular signaling.3 Concen-trations of HA have been found to be elevated in GBMpatients, which can dictate glioma cell fate.4,5 Previous literaturealso suggests that HA can stimulate expression and secretion ofproteins, including matrix metalloproteases (MMP) orosteopontin, which can drive GBM cell migration andinvasion.6 In addition to biochemical cues, mechanical cues,such as matrix stiffness, can also significantly increase duringtumor growth, as GBM cells actively secrete fibrous ECMproteins, including laminin, collagen, and fibronectin.3 Thestiffness of GBM tissues has been reported to be 26 kPa, whichis much higher than the stiffness of normal brain tissue (0.1 to 1kPa).7,8
The current gold standard methods for studying cancerbiology include using 2D monolayer culture, organotypicculture, or animal models. Culturing cells in 2D monolayeroffers ease of use, but tissue culture plastic has a stiffness on theGPa scale, which is orders of magnitude higher than braintissues. Furthermore, 2D monolayer culture fails to mimic keyaspects of the tumor microenvironment, such as 3Darchitecture and presence of the ECM. Organotypic culture,on the other hand, is rich in ECM cues but has limited controlover varying the inputs and are difficult for mechanistic studies.Animal models provide the most sophisticated model for thetumor microenvironment, but are generally lengthy to produce,costly, and only suitable for low throughput studies.Furthermore, studying human cancer cell behavior in animalmodels requires the use of immunocompromised animals,which eliminates the potential roles of immune cells, such asmacrophages, in tumor progression. The limitations of currentplatforms for studying cancer biology call for the developmentof alternative in vitro models to facilitate analysis of tumor cell-niche interactions in a physiologically relevant, controllablemanner.
Biomaterials have been widely used as artificial niches tosupport culturing cells in 3D for tissue engineering. Hydrogelsare particularly attractive for studying brain cells due to theirhigh, tissue-mimicking water content, as well as tunablebiochemical and physical properties. Furthermore, manyhydrogel platforms can be formed using cytocompatibleconditions and permit transport of nutrients and waste. Recentstudies have employed hydrogels as in vitro models for studyingbrain tumor cell behavior in response to microenvironmentcues and showed stiffness-dependent GBM cell motility andproliferation.9,10 Previous work have primarily employednaturally derived materials, such as collagen or hyaluronicacid (HA), to study cancer cell behavior in 3D in vitrohydrogels.10,11 Despite their highly biomimetic nature, naturallyderived materials do not support independent tuning ofmicroenvironment cues and are subject to batch-to-batchvariability. In one study, Ananthanarayanan et al. used HA-based hydrogels to study GBM cell invasion as a function ofmatrix stiffness. To tune the matrix stiffness, HA concentrationwas varied, which introduced simultaneous changes in thebiochemical ligand concentration sensed by GBM cells. Thismakes it difficult to differentiate the respective contribution ofbiochemical or physical cues on observed cell fate changes. Inaddition, to achieve broad tunable matrix stiffness, theconcentration of HA used in the hydrogel exceeded thatfound in brain tissues.4,10 In another study, Pedron et al. usedgelatin-based hydrogels to study GBM cell behavior.12 Similarto HA, gelatin lacks decoupled tunability and may be lessphysiologically relevant, as concentrations of fibrous ECMproteins like collagen are lower in normal brain tissuecompared to connective tissue outside the central nervoussystem. To overcome the limitations of naturally derivedpolymers, recent research efforts have explored hydrogels basedon synthetic polymers, such as poly(ethylene-glycol) (PEG) orpolyacrylamide, as 3D in vitro cancer models that offer morecontrol and reliability.13−15 Unlike natural polymers, syntheticpolymers, such as PEG, are bioinert and present a blank slate.Biological epitopes can also be incorporated to mimic nativeECM and support desirable cell fate processes, such asadhesion, proliferation, or migration. Hydrogel platforms thatcan capitalize on the advantages of both naturally derived andsynthetic materials offer an attractive tool for engineering 3D invitro cancer niches with enhanced biomimicry and tunability.The goal of this study was to develop and characterize a
hydrogel platform with independently tunable biochemical and
Scheme 1. Glioma Cell Encapsulation in 3D Biomimetic Hydrogela
aHydrogel was formed from 8-arm PEG-norbornene, which was crosslinked with linear PEG-dithiol and MMP-cleavable sequence (mixed at 1:1ratio). RGD peptide (CRGDS) was covalently linked to the hydrogel network at a final concentration of 0.914 mM. Sodium hyaluronate (HA) wasadded to a final concentration of 0.266% (w/v) prior to crosslinking, allowing HA to be physically entrapped after gelation. Cells werehomogeneously mixed in the precursor solution at a final concentration of 0.5 M cells/mL. Hydrogel was crosslinked via UV photopolymerization(365 nm and 4 mW/cm2).
Molecular Pharmaceutics Article
dx.doi.org/10.1021/mp5000828 | Mol. Pharmaceutics XXXX, XXX, XXX−XXXB

mechanical cues and employs both naturally derived andsynthetic materials as a 3D in vitro model for studying GBMcell-niche interactions (Scheme 1). To achieve independentlytunable biochemical and mechanical properties, we employed acombination of synthetic and naturally derived materials.Hydrogel stiffness was controlled by varying concentrationsof 8-arm PEG-norbornene (PEG-NB). To allow for cell-mediated degradation and migration, hydrogel network wascross-linked using both linear PEG-dithiol (PEG-SH) and aMMP-cleavable sequence (CGPQGIWGQC). To facilitate celladhesion in 3D, a cell adhesive peptide (CRGDS) wasconjugated to the hydrogel network. Lastly, to mimic thebrain ECM content, sodium hyaluronate (HA) (20−40 kDa)was mixed with the hydrogel precursor solution prior to cross-linking, allowing the HA to be physically entrapped aftergelation. We chose thiol-ene photopolymerization to cross-linkthe hydrogel, as this process is cytocompatible and permitsspatial and temporal control. The resulting hydrogels werecharacterized using mechanical testing and quantification ofequilibrium swelling ratio. Using this hydrogel platform, wethen studied the effect of varying matrix stiffness on modulatingthe cell fate of U87 cells, a commercially available GBM cellline, in 3D over 3 weeks. Cell fates in 3D hydrogels wereanalyzed by monitoring cell proliferation, morphology, andgene expression.
■ EXPERIMENTAL SECTIONMaterials. 8-Arm PEG (MW ≈ 40 kDa) was purchased
from JenKem Technology USA (Allen, TX, USA). Linear PEG(MW ≈ 1.5 kDa) was purchased from Sigma-Aldrich USA (St.Louis, MO). RGD peptide (CRGDS) was purchased from BioBasic, Inc. (Amherst, NY, USA). Sodium hyaluronate (HA)(MW ≈ 20−40 kDa) was purchased from Lifecore Biomedical(Cha s k a , MN , USA) . MMP- c l e a v a b l e p ep t i d e(CGPQGIWGQC) was synthesized by GenScript (Piscataway,NJ, USA). All other reagents and solvents were obtained fromFisher Scientific (Pittsburgh, PA, USA) unless otherwise noted.Hydrogel Formation. 8-Arm PEG-norbornene (PEG-NB)
and linear PEG-dithiol (PEG-SH) were synthesized aspreviously reported.16,17 To form the hydrogel network, 8-Arm 40 kDa PEG-NB was mixed with linear 1.5 kDa PEG-SHand an MMP-cleavable sequence at a molar ratio of 2:3:3 in thepresence of photoinitiator Igracure D2959 (0.05% w/v, CibaSpecialty Chemicals, Tarrytown, NY, USA). RGD peptide(CRGDS) was covalently linked to the hydrogel network at afinal concentration of 0.914 mM. Sodium hyaluronate (20−40kDa) was added at a final concentration of 0.266% (w/v),which was selected based on reported values of hyaluronic acidcontent in human brain tumor stroma.4 Chemical structures ofeach component are shown in Table 1. Each hydrogel samplecontained 75 μL of hydrogel precursor solution, which wasloaded in a cylindrical-shaped mold (3 mm in height, 5 mm indiameter). The hydrogel was exposed to UV light (365 nm and4 mW/cm2) for 5 min at RT to induce gelation.Mechanical Testing. Acellular hydrogels (n = 3) prepared
as above were allowed to equilibrate in PBS at roomtemperature overnight. To measure the stiffness of hydrogels,unconfined compressive tests were conducted using an Instron5944 materials testing system (Instron Corporation, Norwood,MA). The test setup consisted of custom-made aluminumcompression plates lined with PTFE to minimize friction. Alltests were conducted in PBS solution at RT. Hydrogel diameterand thickness were measured using digital calipers and the
material testing system’s position read-out, respectively. Beforeeach test, a preload of approximately 2 mN was applied. Theupper plate was then lowered at a rate of 1% strain/sec to amaximum strain of 30%. Load and displacement data wererecorded at 100 Hz. The modulus was determined for strainranges of 10−20% from linear curve fits of the stress vs straincurve in each strain range.
Swelling and Mesh Size Calculation. Acellular hydrogels(3% or 14% PEG, n = 3) prepared as above were allowed toequilibrate in PBS at room temperature overnight. Theequilibrium swelling ratio Q was calculated as the ratio of themass of the swollen hydrogel to the mass of the drycomponents after lyophilization. The theoretical hydrogelmesh size was calculated as done previously.18
Hydrogel Degradation. To verify that the hydrogelssynthesized are MMP-degradable, acellular hydrogels (14%PEG) were prepared as above and equilibrated in PBS at 37 °Covernight. On day 0, the wet weight of each hydrogel wasmeasured after equilibration. Additional hydrogels were used tomeasure the dry weight to calculate the average initial polymerconcentration in the hydrogels. On days 1 through 5, hydrogelswere placed in a fresh solution of collagenase (Type II, 5 U/mL, Worthington Biochemical Corp, Lakewood, NJ, USA) orPBS overnight at 37 °C. At each time point, the wet weight ofthe hydrogel was measured (n = 3). The equilibrium swellingratio Q was calculated at each time point using the initialpolymer concentration in the hydrogel and the hydrogel wetweight.
Cell Encapsulation in 3D Hydrogels. U87-mg/Luc+/GFP+ cells (U87-mg-df) were expanded in cell culturemedium consisting of Dulbecco’s minimal essential medium(DMEM, Life Technologies, Grand Island, NY), supplementedwith 10% (v/v) fetal bovine serum (FBS, Gibco, LifeTechnologies), 100 U/mL penicillin, and 100 μg/mLstreptomycin at 37 °C in 5% CO2.To mimic the mechanical stiffness found in normal brain and
GBM tissue, we chose 3% PEG (SOFT) and 14% PEG(STIFF) based on the mechanical testing results. U87-mg-dfcells were trypsinized, pelleted, and resuspended in thehydrogel precursor solution (3% or 14% PEG) at a finalconcentration of 0.5 million cells/mL. Then 75 μL of the cell-
Table 1. Chemical Structures of Hydrogel Components
Molecular Pharmaceutics Article
dx.doi.org/10.1021/mp5000828 | Mol. Pharmaceutics XXXX, XXX, XXX−XXXC

containing hydrogel solution was pipetted into a cylindrical-shaped mold and UV cross-linked as described above. Thesamples were then cultured in growth medium as describedabove for 21 days at 37 °C in 5% CO2 with medium changeevery other day.Cell Viability. U87-mg cells (GFP-/Luc-) were encapsu-
lated in soft and stiff hydrogels, and cell viability (n = 1) wasassessed 2 h after encapsulation using the Live/Dead CellViability Assay kit (Life Technologies). Briefly, the Live/Deadreagent was prepared per manufacturer’s instructions. Hydro-gels were immersed in Live/Dead reagent solution for 30−40min and imaged using a Zeiss fluorescence microscope.Cell Proliferation. Cell proliferation inside soft vs stiff
hydrogels was monitored over time using bright fieldmicroscopy. To quantify the cell proliferation, we measuredthe DNA content inside hydrogels at days 1 and 21 using theQuant-iT PicoGreen assay (Life Technologies). Briefly,lyophilized hydrogel samples (n = 3) were rehydrated anddigested using papain (Worthington Biochemical Corp) at 60°C for 16 h. After cooling to room temperature, samples werevortexed and centrifuged at 10,000 rpm for 5 min. Thesupernatant was used to measure DNA content using thePicoGreen assay per manufacturer’s instructions. Acellularhydrogels were used as control.Gene Expression. To examine the effects of varying matrix
stiffness on U87 cell fates, we analyzed gene expressions of U87cells after being cultured inside soft vs stiff hydrogels for 21days. A panel of 14 genes was examined including matrixmetalloproteinases (MMP1, MMP2, and MMP9), HAreceptors (CD44 and RHAMM), HA synthases (HAS1,HAS2, and HAS3), Ras proteins (Hras, Nras, and Kras), andmechanotransduction proteins RhoA and ROCK (ROCK1 andROCK2). Primer sequence of all tested genes are listed inTable 2. To measure the expression of target genes, total RNAwas extracted from the hydrogels (n = 3), and RT-PCR wasperformed. Briefly, hydrogel samples were homogenized inTRIzol (Life Technologies). RNA was extracted by the additionof chloroform and precipitated using RNeasy Mini Kit columns(Qiagen, Valencia, CA). cDNA was synthesized from extractedRNA using SuperScript III First-Strand Synthesis kit (LifeTechnologies) per manufacturer’s instructions. RT-PCR wasthen performed on an Applied Biosystems7900 Real-Time PCRsystem (Applied Biosystems, Life Technologies) using PowerSYBR Green PCR Master Mix (Applied Biosystems, Life
Technologies) per manufacturer’s instructions. Relative ex-pression levels of target genes was determined using thecomparative CT method. Target gene expression was firstnormalized to an endogenous gene GAPDH, followed by asecond normalization to the gene expression level in the controlgroup (3% PEG, day 1).
Histology and Immunohistochemical Staining. Toprepare samples for histology, cell-laden or acellular hydrogels(n = 2) were fixed in 4% paraformaldehyde (Sigma) overnightat 4 °C, incubated in PBS containing 30% sucrose overnight at4 °C, and cyropreserved in optimum cutting temperature(OCT, Tissue-Tek) solution in liquid nitrogen. Samples werestored at −80 °C and sectioned at −20 °C. Sections werestained with H&E (Sigma) to visualize cell morphology. Forimmunostaining of the cell cytoskeleton, antigen retrieval wasperformed via incubation in 10 mM sodium citrate buffer(0.05% Tween 20, pH 6.0, Sigma) at 100 °C for 20 min. Cellswere then permeabilized in PBS containing 1% Triton X-100(Sigma) for 15 min at 37 °C. Nonspecific binding was blockedusing 1% BSA in PBS for 60 min at RT. To stain for actin,sections were then stained with phalloidin-rhodamine (50 μg/mL, Sigma) for 60 min at 37 °C. Cell nuclei werecounterstained with Hoechst dye 33342 (0.25 μg/mL, CellSignaling Technologies, Danvers, MA) for 60 min at RT.Sections were then mounted (Vectashield, Vector Laboratories,Burlingame, CA) and imaged using a Zeiss fluorescencemicroscope.
Statistical Analyses. GraphPad Prism (GraphPad software,San Diego, CA, USA) was used to perform statistical analysison cell proliferation and theoretical mesh size data. Unpairedstudent’s t tests (assuming Gaussian distribution) and two-wayanalysis of variance (ANOVA) with Tukey’s multiplecomparisons test were used to determine statistical significance(p < 0.05). Error was reported as standard deviation unlessotherwise noted.
■ RESULTS
Characterization of Hydrogels. To determine theoptimal hydrogel formulations that mimic the stiffness ofnormal brain and GBM tissues, we first varied theconcentration of PEG from 2% to 14% (w/v) and measuredthe resulting hydrogel stiffness using an unconfined compres-sion test. As expected, increasing PEG concentration led toincreased stiffness, ranging from 0.5 to 26 kPa (Figure 1A),
Table 2. Primer Sequences Used in RT-PCR
gene forward primer (5′ → 3′) reverse primer (5′ → 3′) ref
GAPDH ACAGTCAGCCCGCATCTTCTT CGACCAAATCCGTTGACTC 38CD44 GGTCCTATAAGGACACCCCAAAT AATCAAAGCCAAGGCCAAGA 39RHAMM GTTTCTGGAGCTGGCCGTC ACTGGTCCTTTCAATACTTCTAAAGT 40HAS1 GGTGGGGACGTGCGGATC ATGCAGGATACACAGTGGAAGTAG 41HAS2 GTGGATTATGTACAGGTTTGTGA TCCAACCATGGGATCTTCTT 42HAS3 CTCTACTCCCTCCTCTATATGTC AACTGCCACCCAGATGGA 41MMP1 AGCTAGCTCAGGATGACATTGATG GCCGATGGGCTGGACAG 43MMP2 CCACTGCCTTCGATACAC GAGCCACTCTCTGGAATCTTAAA 44MMP9 TGGGCTACGTGACCTATGACAT GCCCAGCCCACCTCCACTCCTC 45Hras ACATCCACCAGTACAGGGA TGCAGCCAGGTCACACTTGTTC 46Kras TTCCTACAGGAAGCAAGTAG CACAAAGAAAGCCCTCCCCA 47Nras GGTGAAACCTGTTTGTTGGA ATACACAGAGGAAGCCTTCG 47RhoA GGCTGGACTCGGATTCGTT CACAGGCTCCATCACCAACA 48ROCK1 GACCTGTAACCCAAGGAGAT GGAAAGTGGTAGAGTGTAGG 49ROCK2 ATGAAGATACAGCAAAACCAGTC CACCTTGAATAATGACTGCTTTC 50
Molecular Pharmaceutics Article
dx.doi.org/10.1021/mp5000828 | Mol. Pharmaceutics XXXX, XXX, XXX−XXXD
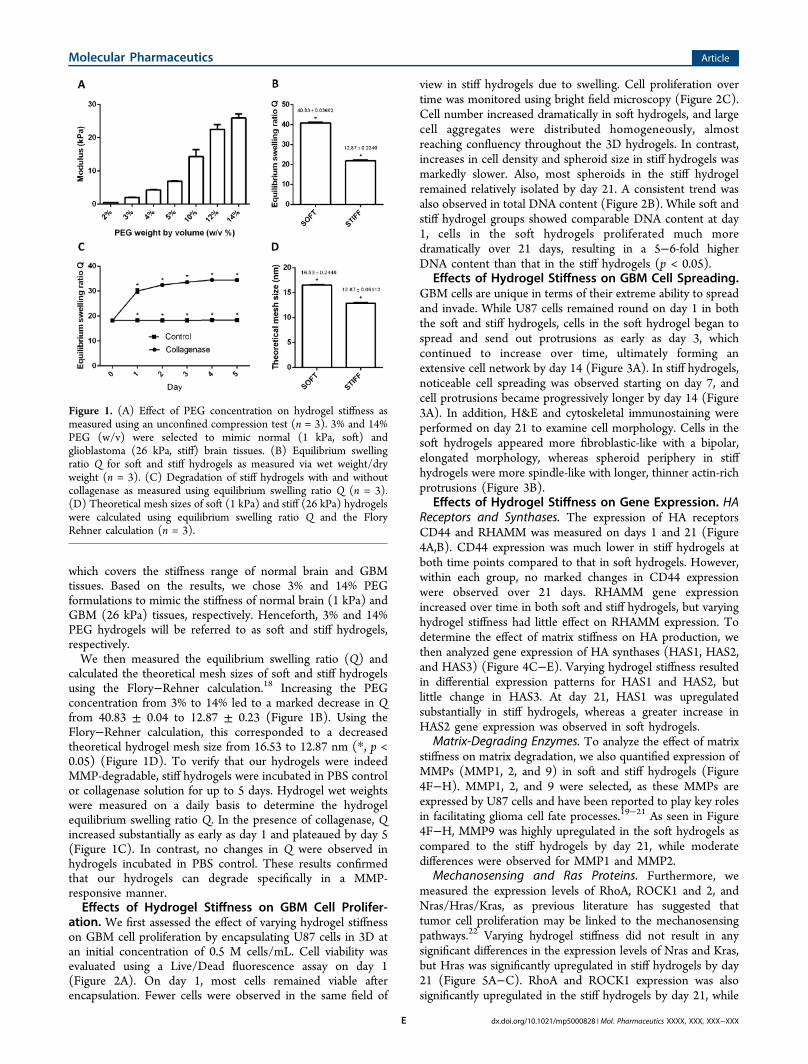
which covers the stiffness range of normal brain and GBMtissues. Based on the results, we chose 3% and 14% PEGformulations to mimic the stiffness of normal brain (1 kPa) andGBM (26 kPa) tissues, respectively. Henceforth, 3% and 14%PEG hydrogels will be referred to as soft and stiff hydrogels,respectively.We then measured the equilibrium swelling ratio (Q) and
calculated the theoretical mesh sizes of soft and stiff hydrogelsusing the Flory−Rehner calculation.18 Increasing the PEGconcentration from 3% to 14% led to a marked decrease in Qfrom 40.83 ± 0.04 to 12.87 ± 0.23 (Figure 1B). Using theFlory−Rehner calculation, this corresponded to a decreasedtheoretical hydrogel mesh size from 16.53 to 12.87 nm (*, p <0.05) (Figure 1D). To verify that our hydrogels were indeedMMP-degradable, stiff hydrogels were incubated in PBS controlor collagenase solution for up to 5 days. Hydrogel wet weightswere measured on a daily basis to determine the hydrogelequilibrium swelling ratio Q. In the presence of collagenase, Qincreased substantially as early as day 1 and plateaued by day 5(Figure 1C). In contrast, no changes in Q were observed inhydrogels incubated in PBS control. These results confirmedthat our hydrogels can degrade specifically in a MMP-responsive manner.Effects of Hydrogel Stiffness on GBM Cell Prolifer-
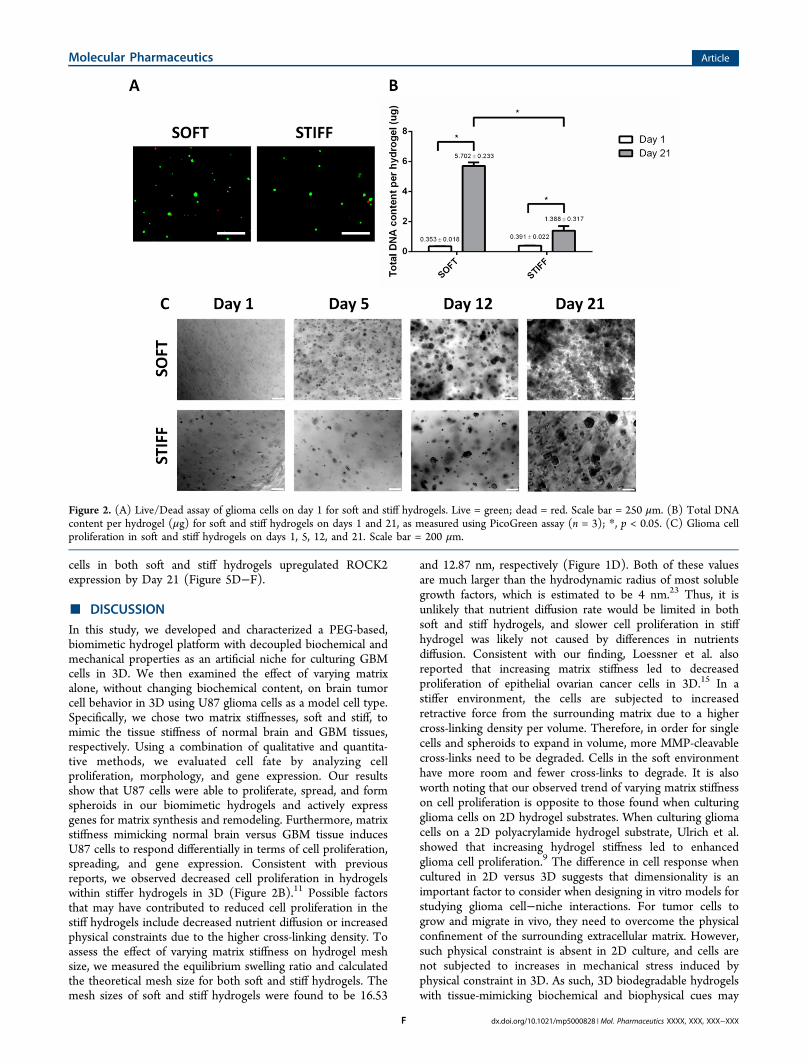
ation. We first assessed the effect of varying hydrogel stiffnesson GBM cell proliferation by encapsulating U87 cells in 3D atan initial concentration of 0.5 M cells/mL. Cell viability wasevaluated using a Live/Dead fluorescence assay on day 1(Figure 2A). On day 1, most cells remained viable afterencapsulation. Fewer cells were observed in the same field of
view in stiff hydrogels due to swelling. Cell proliferation overtime was monitored using bright field microscopy (Figure 2C).Cell number increased dramatically in soft hydrogels, and largecell aggregates were distributed homogeneously, almostreaching confluency throughout the 3D hydrogels. In contrast,increases in cell density and spheroid size in stiff hydrogels wasmarkedly slower. Also, most spheroids in the stiff hydrogelremained relatively isolated by day 21. A consistent trend wasalso observed in total DNA content (Figure 2B). While soft andstiff hydrogel groups showed comparable DNA content at day1, cells in the soft hydrogels proliferated much moredramatically over 21 days, resulting in a 5−6-fold higherDNA content than that in the stiff hydrogels (p < 0.05).
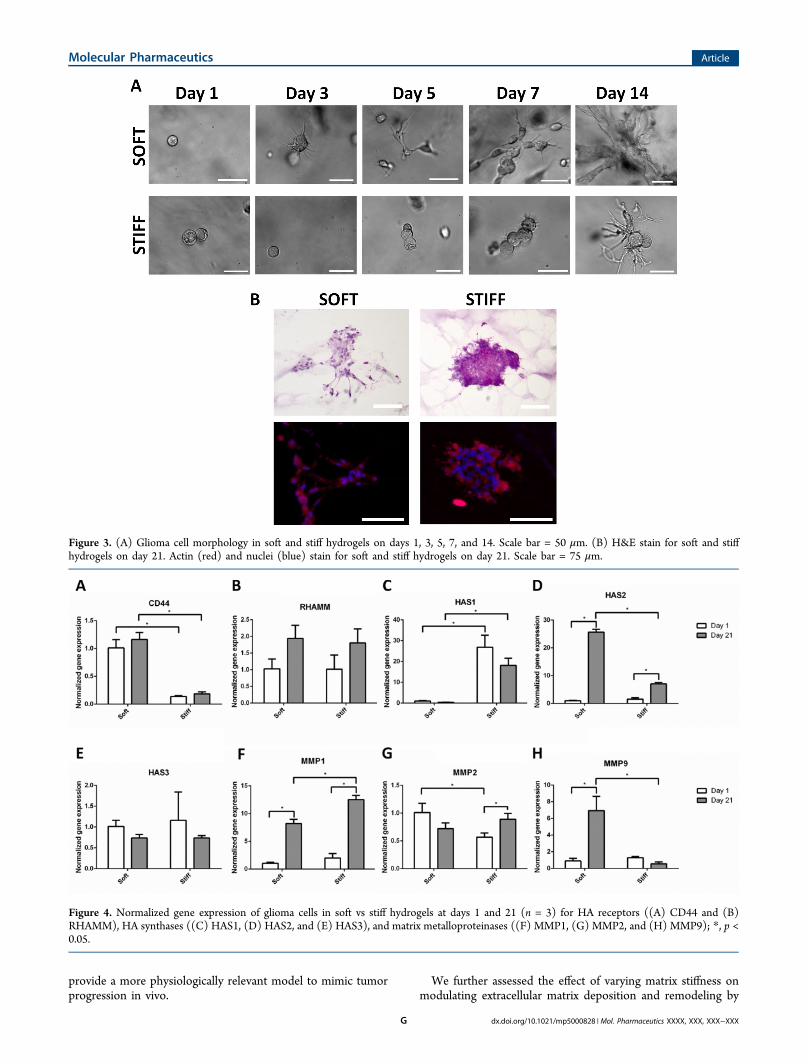
Effects of Hydrogel Stiffness on GBM Cell Spreading.GBM cells are unique in terms of their extreme ability to spreadand invade. While U87 cells remained round on day 1 in boththe soft and stiff hydrogels, cells in the soft hydrogel began tospread and send out protrusions as early as day 3, whichcontinued to increase over time, ultimately forming anextensive cell network by day 14 (Figure 3A). In stiff hydrogels,noticeable cell spreading was observed starting on day 7, andcell protrusions became progressively longer by day 14 (Figure3A). In addition, H&E and cytoskeletal immunostaining wereperformed on day 21 to examine cell morphology. Cells in thesoft hydrogels appeared more fibroblastic-like with a bipolar,elongated morphology, whereas spheroid periphery in stiffhydrogels were more spindle-like with longer, thinner actin-richprotrusions (Figure 3B).
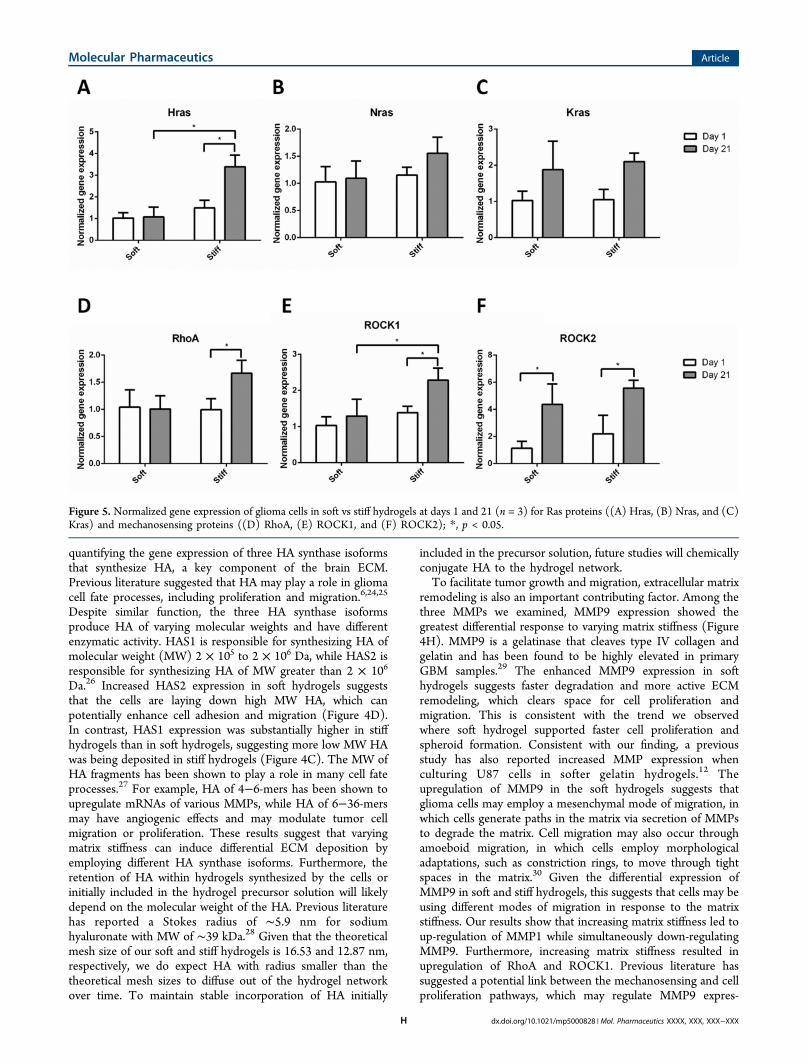
Effects of Hydrogel Stiffness on Gene Expression. HAReceptors and Synthases. The expression of HA receptorsCD44 and RHAMM was measured on days 1 and 21 (Figure4A,B). CD44 expression was much lower in stiff hydrogels atboth time points compared to that in soft hydrogels. However,within each group, no marked changes in CD44 expressionwere observed over 21 days. RHAMM gene expressionincreased over time in both soft and stiff hydrogels, but varyinghydrogel stiffness had little effect on RHAMM expression. Todetermine the effect of matrix stiffness on HA production, wethen analyzed gene expression of HA synthases (HAS1, HAS2,and HAS3) (Figure 4C−E). Varying hydrogel stiffness resultedin differential expression patterns for HAS1 and HAS2, butlittle change in HAS3. At day 21, HAS1 was upregulatedsubstantially in stiff hydrogels, whereas a greater increase inHAS2 gene expression was observed in soft hydrogels.
Matrix-Degrading Enzymes. To analyze the effect of matrixstiffness on matrix degradation, we also quantified expression ofMMPs (MMP1, 2, and 9) in soft and stiff hydrogels (Figure4F−H). MMP1, 2, and 9 were selected, as these MMPs areexpressed by U87 cells and have been reported to play key rolesin facilitating glioma cell fate processes.19−21 As seen in Figure4F−H, MMP9 was highly upregulated in the soft hydrogels ascompared to the stiff hydrogels by day 21, while moderatedifferences were observed for MMP1 and MMP2.
Mechanosensing and Ras Proteins. Furthermore, wemeasured the expression levels of RhoA, ROCK1 and 2, andNras/Hras/Kras, as previous literature has suggested thattumor cell proliferation may be linked to the mechanosensingpathways.22 Varying hydrogel stiffness did not result in anysignificant differences in the expression levels of Nras and Kras,but Hras was significantly upregulated in stiff hydrogels by day21 (Figure 5A−C). RhoA and ROCK1 expression was alsosignificantly upregulated in the stiff hydrogels by day 21, while
Figure 1. (A) Effect of PEG concentration on hydrogel stiffness asmeasured using an unconfined compression test (n = 3). 3% and 14%PEG (w/v) were selected to mimic normal (1 kPa, soft) andglioblastoma (26 kPa, stiff) brain tissues. (B) Equilibrium swellingratio Q for soft and stiff hydrogels as measured via wet weight/dryweight (n = 3). (C) Degradation of stiff hydrogels with and withoutcollagenase as measured using equilibrium swelling ratio Q (n = 3).(D) Theoretical mesh sizes of soft (1 kPa) and stiff (26 kPa) hydrogelswere calculated using equilibrium swelling ratio Q and the FloryRehner calculation (n = 3).
Molecular Pharmaceutics Article
dx.doi.org/10.1021/mp5000828 | Mol. Pharmaceutics XXXX, XXX, XXX−XXXE
cells in both soft and stiff hydrogels upregulated ROCK2expression by Day 21 (Figure 5D−F).
■ DISCUSSIONIn this study, we developed and characterized a PEG-based,biomimetic hydrogel platform with decoupled biochemical andmechanical properties as an artificial niche for culturing GBMcells in 3D. We then examined the effect of varying matrixalone, without changing biochemical content, on brain tumorcell behavior in 3D using U87 glioma cells as a model cell type.Specifically, we chose two matrix stiffnesses, soft and stiff, tomimic the tissue stiffness of normal brain and GBM tissues,respectively. Using a combination of qualitative and quantita-tive methods, we evaluated cell fate by analyzing cellproliferation, morphology, and gene expression. Our resultsshow that U87 cells were able to proliferate, spread, and formspheroids in our biomimetic hydrogels and actively expressgenes for matrix synthesis and remodeling. Furthermore, matrixstiffness mimicking normal brain versus GBM tissue inducesU87 cells to respond differentially in terms of cell proliferation,spreading, and gene expression. Consistent with previousreports, we observed decreased cell proliferation in hydrogelswithin stiffer hydrogels in 3D (Figure 2B).11 Possible factorsthat may have contributed to reduced cell proliferation in thestiff hydrogels include decreased nutrient diffusion or increasedphysical constraints due to the higher cross-linking density. Toassess the effect of varying matrix stiffness on hydrogel meshsize, we measured the equilibrium swelling ratio and calculatedthe theoretical mesh size for both soft and stiff hydrogels. Themesh sizes of soft and stiff hydrogels were found to be 16.53
and 12.87 nm, respectively (Figure 1D). Both of these valuesare much larger than the hydrodynamic radius of most solublegrowth factors, which is estimated to be 4 nm.23 Thus, it isunlikely that nutrient diffusion rate would be limited in bothsoft and stiff hydrogels, and slower cell proliferation in stiffhydrogel was likely not caused by differences in nutrientsdiffusion. Consistent with our finding, Loessner et al. alsoreported that increasing matrix stiffness led to decreasedproliferation of epithelial ovarian cancer cells in 3D.15 In astiffer environment, the cells are subjected to increasedretractive force from the surrounding matrix due to a highercross-linking density per volume. Therefore, in order for singlecells and spheroids to expand in volume, more MMP-cleavablecross-links need to be degraded. Cells in the soft environmenthave more room and fewer cross-links to degrade. It is alsoworth noting that our observed trend of varying matrix stiffnesson cell proliferation is opposite to those found when culturingglioma cells on 2D hydrogel substrates. When culturing gliomacells on a 2D polyacrylamide hydrogel substrate, Ulrich et al.showed that increasing hydrogel stiffness led to enhancedglioma cell proliferation.9 The difference in cell response whencultured in 2D versus 3D suggests that dimensionality is animportant factor to consider when designing in vitro models forstudying glioma cell−niche interactions. For tumor cells togrow and migrate in vivo, they need to overcome the physicalconfinement of the surrounding extracellular matrix. However,such physical constraint is absent in 2D culture, and cells arenot subjected to increases in mechanical stress induced byphysical constraint in 3D. As such, 3D biodegradable hydrogelswith tissue-mimicking biochemical and biophysical cues may
Figure 2. (A) Live/Dead assay of glioma cells on day 1 for soft and stiff hydrogels. Live = green; dead = red. Scale bar = 250 μm. (B) Total DNAcontent per hydrogel (μg) for soft and stiff hydrogels on days 1 and 21, as measured using PicoGreen assay (n = 3); *, p < 0.05. (C) Glioma cellproliferation in soft and stiff hydrogels on days 1, 5, 12, and 21. Scale bar = 200 μm.
Molecular Pharmaceutics Article
dx.doi.org/10.1021/mp5000828 | Mol. Pharmaceutics XXXX, XXX, XXX−XXXF
provide a more physiologically relevant model to mimic tumorprogression in vivo.
We further assessed the effect of varying matrix stiffness onmodulating extracellular matrix deposition and remodeling by
Figure 3. (A) Glioma cell morphology in soft and stiff hydrogels on days 1, 3, 5, 7, and 14. Scale bar = 50 μm. (B) H&E stain for soft and stiffhydrogels on day 21. Actin (red) and nuclei (blue) stain for soft and stiff hydrogels on day 21. Scale bar = 75 μm.
Figure 4. Normalized gene expression of glioma cells in soft vs stiff hydrogels at days 1 and 21 (n = 3) for HA receptors ((A) CD44 and (B)RHAMM), HA synthases ((C) HAS1, (D) HAS2, and (E) HAS3), and matrix metalloproteinases ((F) MMP1, (G) MMP2, and (H) MMP9); *, p <0.05.
Molecular Pharmaceutics Article
dx.doi.org/10.1021/mp5000828 | Mol. Pharmaceutics XXXX, XXX, XXX−XXXG
quantifying the gene expression of three HA synthase isoformsthat synthesize HA, a key component of the brain ECM.Previous literature suggested that HA may play a role in gliomacell fate processes, including proliferation and migration.6,24,25
Despite similar function, the three HA synthase isoformsproduce HA of varying molecular weights and have differentenzymatic activity. HAS1 is responsible for synthesizing HA ofmolecular weight (MW) 2 × 105 to 2 × 106 Da, while HAS2 isresponsible for synthesizing HA of MW greater than 2 × 106
Da.26 Increased HAS2 expression in soft hydrogels suggeststhat the cells are laying down high MW HA, which canpotentially enhance cell adhesion and migration (Figure 4D).In contrast, HAS1 expression was substantially higher in stiffhydrogels than in soft hydrogels, suggesting more low MW HAwas being deposited in stiff hydrogels (Figure 4C). The MW ofHA fragments has been shown to play a role in many cell fateprocesses.27 For example, HA of 4−6-mers has been shown toupregulate mRNAs of various MMPs, while HA of 6−36-mersmay have angiogenic effects and may modulate tumor cellmigration or proliferation. These results suggest that varyingmatrix stiffness can induce differential ECM deposition byemploying different HA synthase isoforms. Furthermore, theretention of HA within hydrogels synthesized by the cells orinitially included in the hydrogel precursor solution will likelydepend on the molecular weight of the HA. Previous literaturehas reported a Stokes radius of ∼5.9 nm for sodiumhyaluronate with MW of ∼39 kDa.28 Given that the theoreticalmesh size of our soft and stiff hydrogels is 16.53 and 12.87 nm,respectively, we do expect HA with radius smaller than thetheoretical mesh sizes to diffuse out of the hydrogel networkover time. To maintain stable incorporation of HA initially
included in the precursor solution, future studies will chemicallyconjugate HA to the hydrogel network.To facilitate tumor growth and migration, extracellular matrix
remodeling is also an important contributing factor. Among thethree MMPs we examined, MMP9 expression showed thegreatest differential response to varying matrix stiffness (Figure4H). MMP9 is a gelatinase that cleaves type IV collagen andgelatin and has been found to be highly elevated in primaryGBM samples.29 The enhanced MMP9 expression in softhydrogels suggests faster degradation and more active ECMremodeling, which clears space for cell proliferation andmigration. This is consistent with the trend we observedwhere soft hydrogel supported faster cell proliferation andspheroid formation. Consistent with our finding, a previousstudy has also reported increased MMP expression whenculturing U87 cells in softer gelatin hydrogels.12 Theupregulation of MMP9 in the soft hydrogels suggests thatglioma cells may employ a mesenchymal mode of migration, inwhich cells generate paths in the matrix via secretion of MMPsto degrade the matrix. Cell migration may also occur throughamoeboid migration, in which cells employ morphologicaladaptations, such as constriction rings, to move through tightspaces in the matrix.30 Given the differential expression ofMMP9 in soft and stiff hydrogels, this suggests that cells may beusing different modes of migration in response to the matrixstiffness. Our results show that increasing matrix stiffness led toup-regulation of MMP1 while simultaneously down-regulatingMMP9. Furthermore, increasing matrix stiffness resulted inupregulation of RhoA and ROCK1. Previous literature hassuggested a potential link between the mechanosensing and cellproliferation pathways, which may regulate MMP9 expres-
Figure 5. Normalized gene expression of glioma cells in soft vs stiff hydrogels at days 1 and 21 (n = 3) for Ras proteins ((A) Hras, (B) Nras, and (C)Kras) and mechanosensing proteins ((D) RhoA, (E) ROCK1, and (F) ROCK2); *, p < 0.05.
Molecular Pharmaceutics Article
dx.doi.org/10.1021/mp5000828 | Mol. Pharmaceutics XXXX, XXX, XXX−XXXH

sion.22,31 Specifically, MMP9 is a downstream effector of Ras,which may interact with the Rho/ROCK signaling axis.Our results demonstrate that U87 cell morphology and
spreading are differentially modulated in 3D hydrogels withvarying matrix stiffness. Cells in stiff hydrogels showed a morespindle-like morphology with longer and thinner actinprotrusions, whereas in soft hydrogels, cells displayed fewerand shorter protrusions (Figure 3B). These results may beexplained by differential expression of RhoA and ROCK1, twokey players in mechanotransduction pathways (Figure 5D−E).RhoA is a small GTPase that is known to regulate the actincytoskeleton and the formation of stress fibers via downstreameffectors. RhoA is regulated via the balance between GTP-aseactivating protein (GAP) and guanine nucleotide exchangefactor (GEF) proteins. ROCK1 and 2 are activated by GTP-bound Rho and share more than 30 intermediate downstreamsubstrates.32 Despite their similarities, ROCK1 and 2 may playdifferent roles in cell fate processes. For example, in mouseembryonic fibroblasts, ROCK1 deficiency inhibited actincytoskeleton remodeling, while ROCK2 downregulation ledto increased periphery membrane folding and altered celladhesion.32 In glioma cells, ROCK1 and 2 inhibition has beenreported to lead to differences in cell cycle progression.33 Also,Zohrabian et al. found that the addition of a ROCK inhibitor toglioblastoma cultures in 2D culture led to decreased radialmigration.34 Increased ROCK1 expression in the stiff hydrogelsmay explain the increase in the number of actin protrusions atthe periphery of the spheroid in stiff hydrogels. Although thematrix stiffness is significantly higher, the cells may activelyreorganize their actin cytoskeleton to probe and invade thematrix, a phenomenon that has been observed in breast cancercell lines when compressed in 2D.35
Previous literature has suggested that matrix stiffness sensedby Rho may also affect cell proliferation via the Ras pathway.22
Zohrabian et al. found that addition of ROCK inhibitors toglioblastoma cultures reduced the levels of phosphorylatedERK, a downstream effector of Ras, which may ultimately leadto reduced cell proliferation.34 Therefore, there may becooperation between the Rho/ROCK and Ras pathways toregulate cell motility and growth. In our studies, theupregulation of Rho/ROCK proteins may be correlated withthe significant upregulation in Hras expression. U87 cells havebeen shown to express the three isoforms of Ras, namely, Hras,Nras, and Kras.36 These three RAS genes encode for highlyhomologous Ras proteins, which have identical effector bindingdomains and can interact with the same set of downstreameffectors. However, each Ras protein exhibits different post-translational modifications, which results in different traffickingroutes and localization in domains of the plasma membrane.37
In our study, we observed upregulated Hras expression in stiffhydrogels (Figure 5A). However, despite increased expressionof Hras, cell proliferation rates seemed to be lower in the stiffhydrogels. This may be partially caused by other factors thatinfluence cell proliferation, such as the greater physicalconstraint that cells need to overcome in stiff hydrogels.In summary, here we report a PEG-based 3D hydrogel
platform as an artificial niche for studying the effect of matrixcues on GBM brain tumor cell fate in 3D. Using U87 cells as amodel GBM cell line, we demonstrate that such biomimetichydrogels support U87 cell growth, spreading, and migration in3D over the course of 3 weeks, and cells remain active indepositing new ECM and continue to upregulate matrixremodeling genes. Unlike previously reported hydrogels, our
platform allows varying matrix stiffness to mimic thephysiologically relevant matrix stiffness of normal brain andGBM tissues without changing biochemical content. Ourresults showed that soft and stiff hydrogels induced differentialtumor cell fates, such as proliferation, spreading, and geneexpressions, suggesting that changes in matrix stiffness intumors play an important role in modulating GBM progression.While the present study focuses on varying matrix stiffnessalone, our hydrogel platform allows decoupled tunability ofother niche cues, such as biochemical ligands or matrixdegradability, for examining complex interactions betweencancer cells and tumor niche. The platform reported here mayprovide a useful 3D in vitro model for elucidating themechanisms underlying GBM progression in a more physio-logically relevant and controlled manner, as well as forevaluating efficacy of potential drug candidates.
■ AUTHOR INFORMATIONCorresponding Author*(F.Y.) Tel: 650-725-7128. Fax: 650-723-9370. E-mail:[email protected] authors declare no competing financial interest.
■ ACKNOWLEDGMENTSThe authors would like to thank Stanford Child HealthResearch Institute (CHRI) Faculty Scholar Award for funding.C.W. would like to thank Stanford Graduate Fellowship forsupport. The authors also appreciate the technical assistancefrom Anthony Behn on mechanical testing.
■ REFERENCES(1) Wen, P. Y.; Kesari, S. Malignant gliomas in adults. N. Engl. J. Med.2008, 359 (5), 492−507.(2) Charles, N. A.; Holland, E. C.; Gilbertson, R.; Glass, R.;Kettenmann, H. The brain tumor microenvironment. Glia 2012, 60(3), 502−514.(3) Wiranowska, M.; Rojiani, M. V. Extracellular Matrix Micro-environment in Glioma Progression. In Glioma: Exploring Its Biologyand Practical Relevance; Ghosh, A., Ed.; Intech: Rijeka, Croatia, 2011;pp 257−284.(4) Delpech, B.; Maingonnat, C.; Girard, N.; Chauzy, C.; Maunoury,R.; Olivier, A.; Tayot, J.; Creissard, P. Hyaluronan and hyaluronectinin the extracellular matrix of human brain tumor stroma. Eur. J. Cancer1993, 29 (7), 1012−1017.(5) Pedron, S.; Becka, E.; Harley, B. A. Regulation of glioma cellphenotype in 3D matrices by hyaluronic acid. Biomaterials 2013, 34(30), 7408−17.(6) Park, J. B.; Kwak, H.-J.; Lee, S.-H. Role of hyaluronan in gliomainvasion. Cell Adhes. Migr. 2008, 2 (3), 202−207.(7) Polacheck, W. J.; Zervantonakis, I. K.; Kamm, R. D. Tumor cellmigration in complex microenvironments. Cell. Mol. Life Sci. 2012, 70(8), 1335−1356.(8) Engler, A. J.; Sen, S.; Sweeney, H. L.; Discher, D. E. Matrixelasticity directs stem cell lineage specification. Cell 2006, 126 (4),677−689.(9) Ulrich, T. A.; de Juan Pardo, E. M.; Kumar, S. The mechanicalrigidity of the extracellular matrix regulates the structure, motility, andproliferation of glioma cells. Cancer Res. 2009, 69 (10), 4167−4174.(10) Ananthanarayanan, B.; Kim, Y.; Kumar, S. Elucidating themechanobiology of malignant brain tumors using a brain matrix-mimetic hyaluronic acid hydrogel platform. Biomaterials 2011, 32 (31),7913−7923.(11) Kaufman, L. J.; Brangwynne, C. P.; Kasza, K. E.; Filippidi, E.;Gordon, V. D.; Deisboeck, T. S.; Weitz, D. A. Glioma expansion in
Molecular Pharmaceutics Article
dx.doi.org/10.1021/mp5000828 | Mol. Pharmaceutics XXXX, XXX, XXX−XXXI

collagen I matrices: analyzing collagen concentration-dependentgrowth and motility patterns. Biophys. J. 2005, 89 (1), 635−650.(12) Pedron, S.; Harley, B. A. C. Impact of the biophysical features ofa 3D gelatin microenvironment on glioblastoma malignancy. J. Biomed.Mater. Res., Part A 2013, 3404−3415.(13) Gill, B. J.; Gibbons, D. L.; Roudsari, L. C.; Saik, J. E.; Rizvi, Z.H.; Roybal, J. D.; Kurie, J. M.; West, J. L. A synthetic matrix withindependently tunable biochemistry and mechanical properties tostudy epithelial morphogenesis and EMT in a lung adenocarcinomamodel. Cancer Res. 2012, 72 (22), 6013−23.(14) Yang, X.; Sarvestani, S. K.; Moeinzadeh, S.; He, X.; Jabbari, E.Three-dimensional-engineered matrix to study cancer stem cells andtumorsphere formation: effect of matrix modulus. Tissue Eng., Part A2013, 19 (5−6), 669−84.(15) Loessner, D.; Stok, K. S.; Lutolf, M. P.; Hutmacher, D. W.;Clements, J. A.; Rizzi, S. C. Bioengineered 3D platform to explorecell−ECM interactions and drug resistance of epithelial ovarian cancercells. Biomaterials 2010, 31 (32), 8494−8506.(16) Fairbanks, B. D.; Schwartz, M. P.; Halevi, A. E.; Nuttelman, C.R.; Bowman, C. N.; Anseth, K. S. A versatile synthetic extracellularmatrix mimic via thiol-norbornene photopolymerization. Adv. Mater.2009, 21 (48), 5005−5010.(17) Anderson, S. B.; Lin, C.-C.; Kuntzler, D. V.; Anseth, K. S. Theperformance of human mesenchymal stem cells encapsulated in cell-degradable polymer−peptide hydrogels. Biomaterials 2011, 32 (14),3564−3574.(18) Zustiak, S. P.; Leach, J. B. Hydrolytically degradable poly-(ethylene glycol) hydrogel scaffolds with tunable degradation andmechanical properties. Biomacromolecules 2010, 11 (5), 1348−1357.(19) Rao, J. S. Molecular mechanisms of glioma invasiveness: the roleof proteases. Nat. Rev. Cancer 2003, 3 (7), 489−501.(20) Stojic, J.; Hagemann, C.; Haas, S.; Herbold, C.; Kuhnel, S.;Gerngras, S.; Roggendorf, W.; Roosen, K.; Vince, G. H. Expression ofmatrix metalloproteinases MMP-1, MMP-11 and MMP-19 iscorrelated with the WHO-grading of human malignant gliomas.Neurosci. Res. 2008, 60 (1), 40−49.(21) Hagemann, C.; Anacker, J.; Haas, S.; Riesner, D.; Schomig, B.;Ernestus, R.-I.; Vince, G. H. Comparative expression pattern of matrix-metalloproteinases in human glioblastoma cell-lines and primarycultures. BMC Res. Notes 2010, 3 (1), 293.(22) Jaalouk, D. E.; Lammerding, J. Mechanotransduction gone awry.Nat. Rev. Mol. Cell Biol. 2009, 10 (1), 63−73.(23) Berry, R.; Jowitt, T. A.; Ferrand, J.; Roessle, M.; Grossmann, J.G.; Canty-Laird, E. G.; Kammerer, R. A.; Kadler, K. E.; Baldock, C.Role of dimerization and substrate exclusion in the regulation of bonemorphogenetic protein-1 and mammalian tolloid. Proc. Natl. Acad. Sci.U.S.A. 2009, 106 (21), 8561−8566.(24) Toole, B. P. Hyaluronan promotes the malignant phenotype.Glycobiology 2002, 12 (3), 37R−42R.(25) Toole, B. P. Hyaluronan: from extracellular glue to pericellularcue. Nat. Rev. Cancer 2004, 4 (7), 528−539.(26) Itano, N.; Kimata, K. Mammalian Hyaluronan Synthases.IUBMB Life 2002, 54, 195−199.(27) Stern, R. Hyaluronan in Cancer Biology; Elsevier Inc: New York,2009.(28) Shimada, E.; Torii, N. K. Determination of Stokes radii andmolecular masses of sodium hyaluronates by sephacryl gelchromatography. J. Chromatogr. A 1994, 685, 172−177.(29) Choe, G.; Park, J. K.; Jouben-Steele, L.; Kremen, T. J.; Liau, L.M.; Vinters, H. V.; Cloughesy, T. F.; Mischel, P. S. Active matrixmetalloproteinase 9 expression is associated with primary glioblastomasubtype. Clin. Cancer Res. 2002, 8 (9), 2894−901.(30) Pathak, A.; Kumar, S. Biophysical regulation of tumor cellinvasion: moving beyond matrix stiffness. Integr. Biol. 2011, 3 (4), 267.(31) Iyer, V.; Pumiglia, K.; DiPersio, C. M. α3β1 integrin regulatesMMP-9 mRNA stability in immortalized keratinocytes: a novelmechanism of integrin-mediated MMP gene expression. J. Cell Sci.2005, 118 (6), 1185−1195.
(32) Shi, J.; Wu, X.; Surma, M.; Vemula, S.; Zhang, L.; Yang, Y.;Kapur, R.; Wei, L. Distinct roles for ROCK1 and ROCK2 in theregulation of cell detachment. Cell Death Dis. 2013, 4 (2), e483.(33) Inaba, N.; Ishizawa, S.; Kimura, M.; Fujioka, K.; Watanabe, M.;Shibasaki, T.; Manome, Y. Effect of inhibition of ROCK isoform onRT2 malignant glioma cells. Anticancer Res. 2010, 30 (9), 3509−3514.(34) Zohrabian, V. M.; Forzani, B.; Chau, Z.; Murali, R.; Jhanwar-Uniyal, M. Rho/ROCK and MAPK signaling pathways are involved inglioblastoma cell migration and proliferation. Anticancer Res. 2009, 29(1), 119−123.(35) Tse, J. M.; Cheng, G.; Tyrrell, J. A.; Wilcox-Adelman, S. A.;Boucher, Y.; Jain, R. K.; Munn, L. L. Mechanical compression drivescancer cells toward invasive phenotype. Proc. Natl. Acad. Sci. U.S.A.2012, 109 (3), 911−6.(36) Blum, R.; Jacob-Hirsch, J.; Amariglio, N.; Rechavi, G.; Kloog, Y.Ras inhibition in glioblastoma down-regulates hypoxia-induciblefactor-1a, causing glycolysis shutdown and cell death. Cancer Res.2005, 65, 999−1006.(37) Hancock, J. F. Ras proteins: different signals from differentlocations. Nat. Rev. Mol. Cell Biol. 2003, 4 (5), 373−385.(38) Ramasubramanian, A.; Shiigi, S.; Lee, G.; Yang, F. Non-viraldelivery of inductive and suppressive genes to adipose-derived stemcells for osteogenic differentiation. Pharm. Res. 2011, 28 (6), 1328−1337.(39) Kouvidi, K.; Berdiaki, A.; Nikitovic, D.; Katonis, P.; Afratis, N.;Hascall, V. C.; Karamanos, N. K.; Tzanakakis, G. N. Role of receptorfor hyaluronic acid-mediated motility (RHAMM) in low molecularweight hyaluronan (LMWHA)-mediated fibrosarcoma cell adhesion. J.Biol. Chem. 2011, 286 (44), 38509−38520.(40) Crainie, M.; Belch, A. R.; Mant, M. J.; Pilarski, L. M.Overexpression of the receptor for hyaluronan-mediated motility(RHAMM) characterizes the malignant clone in multiple myeloma:identification of three distinct RHAMM variants. Blood 1999, 93 (5),1684−1696.(41) Kramer, M. W.; Escudero, D. O.; Lokeshwar, S. D.; Golshani,R.; Ekwenna, O. O.; Acosta, K.; Merseburger, A. S.; Soloway, M.;Lokeshwar, V. B. Association of hyaluronic acid family members(HAS1, HAS2, and HYAL-1) with bladder cancer diagnosis andprognosis. Cancer 2011, 117 (6), 1197−1209.(42) Twarock, S.; Rock, K.; Sarbia, M.; Weber, A. A.; Janicke, R. U.;Fischer, J. W. Synthesis of hyaluronan in oesophageal cancer cells isuncoupled from the prostaglandin−cAMP pathway. Br. J. Pharmacol.2009, 157 (2), 234−243.(43) Huntington, J. T.; Shields, J. M.; Der, C. J.; Wyatt, C. A.;Benbow, U.; Slingluff, C. L.; Brinckerhoff, C. E. Overexpression ofcollagenase 1 (MMP-1) is mediated by the ERK pathway in invasivemelanoma cells: role of BRAF mutation and fibroblast growth factorsignaling. J. Biol. Chem. 2004, 279 (32), 33168−33176.(44) Lockwood, C. J.; Oner, C.; Uz, Y. H.; Kayisli, U. A.; Huang, S. J.;Buchwalder, L. F.; Murk, W.; Funai, E. F.; Schatz, F. Matrixmetalloproteinase 9 (MMP9) expression in preeclamptic deciduaand MMP9 induction by tumor necrosis factor alpha and interleukin 1beta in human first trimester decidual cells. Biol. Reprod. 2008, 78 (6),1064−1072.(45) McKenna, G. J.; Chen, Y.; Smith, R. M.; Meneghetti, A.; Ong,C.; McMaster, R.; Scudamore, C. H.; Chung, S. W. A role for matrixmetalloproteinases and tumor host interaction in hepatocellularcarcinomas. Ame. J. Surg. 2002, 183 (5), 588−594.(46) Barbier, J.; Dutertre, M.; Bittencourt, D.; Sanchez, G.; Gratadou,L.; de la Grange, P.; Auboeuf, D. Regulation of H-ras splice variantexpression by cross talk between the p53 and nonsense-mediatedmRNA decay pathways. Mol. Cell. Biol. 2007, 27 (20), 7315−7333.(47) Jin, Y.; Shima, Y.; Furu, M.; Aoyama, T.; Nakamata, T.;Nakayama, T.; Nakamura, T.; Toguchida, J. Absence of oncogenicmutations of RAS family genes in soft tissue sarcomas of 100 Japanesepatients. Anticancer Res. 2010, 30 (1), 245−251.(48) Pu, Y.-S.; Wang, C.-W.; Liu, G.-Y.; Kuo, Y.-Z.; Huang, C.-Y.;Kang, W.-Y.; Shun, C.-T.; Lin, C.-C.; Wu, W.-J.; Hour, T.-C. Down-
Molecular Pharmaceutics Article
dx.doi.org/10.1021/mp5000828 | Mol. Pharmaceutics XXXX, XXX, XXX−XXXJ

regulated expression of RhoA in human conventional renal cellcarcinoma. Anticancer Res. 2008, 28 (4B), 2039−2043.(49) Lefort, K.; Mandinova, A.; Ostano, P.; Kolev, V.; Calpini, V.;Kolfschoten, I.; Devgan, V.; Lieb, J.; Raffoul, W.; Hohl, D.; Neel, V.;Garlick, J.; Chiorino, G.; Dotto, G. P. Notch1 is a p53 target geneinvolved in human keratinocyte tumor suppression through negativeregulation of ROCK1/2 and MRCKα kinases. Genes Dev. 2007, 21(5), 562−577.(50) Zheng, F.; Liao, Y.-J.; Cai, M.-Y.; Liu, Y.-H.; Liu, T.-H.; Chen,S.-P.; Bian, X.-W.; Guan, X.-Y.; Lin, M. C.; Zeng, Y.-X.; Kung, H.-F.;Xie, D. The putative tumour suppressor microRNA-124 modulateshepatocellular carcinoma cell aggressiveness by repressing ROCK2 andEZH2. Gut 2012, 61 (2), 278−289.
Molecular Pharmaceutics Article
dx.doi.org/10.1021/mp5000828 | Mol. Pharmaceutics XXXX, XXX, XXX−XXXK